daya hambat bacillus thuringiensis kahn 15.39 terhadap ... · fakultas matematika dan ilmu...
TRANSCRIPT

DAYA HAMBAT Bacillus thuringiensis KAHN 15.39
TERHADAP CENDAWAN PATOGEN Curvularia affinis DAN
Colletotrichum gloeosporoides ASAL DAUN KELAPA SAWIT
SYIPA PAOZIAH
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
BOGOR
2014

2

PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa skripsi berjudul Daya Hambat Bakteri
Bacillus thuringiensis KAHN 15.39 terhadap Cendawan Patogen Curvularia
affinis dan Colletotrichum gloeosporoides Asal Daun Kelapa Sawit adalah benar
karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam
bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang
berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari
penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di
bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada
Institut Pertanian Bogor.
Bogor, September 2014
Syipa Paoziah
NIM G34100117

4
ABSTRAK
SYIPA PAOZIAH. Daya Hambat Bacillus thuringiensis KAHN 15.39 terhadap
Cendawan Patogen Curvularia affinis dan Colletotrichum gloeosporoides Asal
Daun Kelapa Sawit. Dibimbing oleh NISA RACHMANIA MUBARIK dan SRI
LISTIYOWATI.
Bibit yang berkualitas menjadi salah satu faktor yang memengaruhi
budidaya kelapa sawit di lapangan. Beberapa cendawan patogen dapat
menyebabkan bercak pada daun bibit kelapa sawit sehingga menurunkan kualitas
bibit. Penelitian ini bertujuan menguji daya hambat Bacillus thuringiensis KAHN
15.39 terhadap cendawan patogen Curvularia affinis dan Colletotrichum
gloeosporoides yang diisolasi dari bibit tanaman kelapa sawit melalui metode
Leaf-pieces assays. Peremajaan bakteri menggunakan medium Nutrient Agar
yang mengandung 1% Carboxy Methyl Celullose dan peremajaan cendawan
menggunakan medium Potato Dextrose Agar yang ditambahkan 500 mg/L
kloramfenikol. Bacillus thuringiensis KAHN15.39 dapat menghambat Curvularia
affinis dengan luas area terinfestasi sebesar 69 mm2 atau efektivitas biokontrolnya
sebesar 86%, dan menghambat Colletotrichum gloeosporoides dengan luas area
terinfestasi sebesar 152 mm2 atau efektivitas biokontrolnya sebesar 45%.
Penghambatan bakteri Bacillus thuringiensis KAHN15.39 terhadap Curvularia
affinis dan Colletotrichum gloeosporoides masing-masing bersifat kuratif.
Kata kunci: kelapa sawit, Bacillus thuringiensis KAHN15.39, Curvularia affinis,
Colletotrichum gloeosporoides, efektivitas biokontrol.

ABSTRACT
SYIPA PAOZIAH. Activity of Bacillus thuringiensis KAHN 15.39 as Biological
Control of Pathogenic Fungi Curvularia affinis and Colletotrichum
gloeosporoides from Leaves Seedling of Oil Palm
Some fungi caused disease on oil palm seedling so that it could be
decrease seedling quality. Therefore, the aim of this research was to investigate
activity of Bacillus thuringiensis KAHN 15.39 to inhibit pathogenic fungi
Curvularia affinis and Colletotrichum gloeosporoides which were isolated from
oil palm seedling. The bacteria ability to inhibit was examined with Leaf-pieces
assays method. Bacteria culture medium was Nutrient Agar which was
containing 1% Carboxy Methyl Celullose and medium for fungi used Potato
Dextrose Agar added with 500 mg/L chloramphenicol. The results showed B.
thuringiensis KAHN15.39 could inhibit Curvularia affinis which were the
infestation areas 69 mm2 or biocontrol effectiveness 86% either, while to inhibit
Colletotrichum gloeosporoides was the infestation areas 152 mm2 or biocontrol
effectiveness by 45%. Inhibition of Bacillus thuringiensis KAHN15.39 against
Curvularia affinis and Colletotrichum gloeosporoides have curative characteristic.
Key words: Oil palm, Bacillus thuringiensis KAHN15.39, Curvularia affinis,
Colletotrichum gloeosporoides, biocontrol effectiveness.

6

DAYA HAMBAT Bacillus thuringiensis KAHN 15.39
TERHADAP CENDAWAN PATOGEN Curvularia affinis DAN
Colletotrichum gloeosporoides ASAL DAUN KELAPA SAWIT
SYIPA PAOZIAH
Skripsi
sebagai salah satu syarat untuk memperoleh gelar
Sarjana Sains pada
Departemen Biologi
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
BOGOR
2014

8


10

PRAKATA
Alhamdulillahirabbil’alamin, puji dan syukur penulis panjatkan kepada
Allah Subhanahu wa Ta’ala atas segala karunia-Nya sehingga karya ilmiah ini
berhasil diselesaikan. Penelitian yang dilaksanakan sejak bulan Januari 2014 ini
berkaitan dengan mikrob sebagai biokontrol dengan judul Daya Hambat Bacillus
thuringiensis KAHN 15.39 terhadap Cendawan Patogen Curvularia affinis dan
Colletotrichum gloeosporoides Asal Daun Kelapa Sawit.
Terima kasih penulis ucapkan kepada Dr Nisa Rachmania Mubarik MSi dan
Dr Sri Listiyowati MSi selaku pembimbing. Terimakasih penulis ucapkan juga
kepada Dr Yohana Caecilia Sulistyaningsih Msi atas saran dan diskusi yang
diberikan sebagai dosen penguji skripsi. Penghargaan penulis tujukan kepada
Bapak, Ibu, Kakak-kakak (Teh Lilis, Teh Mumun, Teh Pipin, Teh Ifah, Teh Ade,
Teh Nunuy, Teh Ikah) serta segenap keluarga besar atas segala do’a, kasih sayang,
bantuan moral serta materil sehingga penulis dapat menyeleseikan studi di IPB
dengan lancar. Ungkapan terima kasih juga penulis sampaikan kepada Kak
Muhammad Asril atas diskusi dan bantuan yang diberikan selama penelitian di
laboratorium, sahabat sepenelitian (Suri Annisa), sahabat-sahabat seperjuangan
(Ismi, Tya, Ledy, Della, Yuli, Rahma), Bapak Jaka dan Mba Heni sebagai laboran
Mikrobiologi, rekan kerja di laboratorium Mikrobiologi yang telah banyak
membantu dan memberikan saran dalam proses penelitian, keluarga besar Pondok
Mahasiwa Al Ihya Dramaga yang telah memberikan dukungan dan do’anya serta
teman-teman Biologi47 atas dukungan, semangat dan kebersamaannya selama
kuliah di IPB. Terima kasih dan mohon maaf penulis sampaikan kepada pihak-
pihak terkait yang telah banyak membantu dalam penelitian dan penyusunan
skripsi yang tidak dapat penulis sebutkan satu persatu.
Semoga karya ilmiah ini bermanfaat.
Bogor, September 2014
Syipa Paoziah

12

DAFTAR ISI
DAFTAR TABEL xiv
DAFTAR GAMBAR xiv
PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
METODE 2
Waktu dan Tempat 2
Bahan 2
Peremajaan Isolat 2
Kemampuan Cendawan Uji Menginfestasi Daun Kelapa Sawit 3
Reisolasi Cendawan 3
Daya Hambat Bakteri terhadap Cendawan dengan 4
Metode Leaf-Pieces Assays 4
Pengamatan Struktur Anatomi Daun Terinfestasi Cendawan 5
HASIL 5
Peremajaan Isolat 5
Kemampuan Cendawan Uji Menginfestasi Daun Kelapa Sawit 7
Reisolasi Cendawan 8
Daya Hambat Bakteri terhadap Cendawan dengan 8
Metode Leaf-pieces assays 8
Pengamatan Struktur Anatomi Daun Terinfestasi Cendawan 11
PEMBAHASAN 12
SIMPULAN DAN SARAN 14
Simpulan 14
Saran 15
DAFTAR PUSTAKA 15
RIWAYAT HIDUP 17

14
DAFTAR TABEL
1 Daya hambat Bacillus thuringiensis KAHN 15.39 terhadap cendawan
Curvularia affinis dan Colletotrichum gloeosporoides melalui metode
uji tantang 7
2 Luas daun terinfestasi cendawan dan efektivitas biokontrol (BE) bakteri
selulolitik terhadap cendawan Curvularia affinis 9
3 Luas daun terinfestasi cendawan dan efektivitas biokontrol (BE) bakteri
selulolitik terhadap cendawan Colletotrichum gloeosporoides 10
DAFTAR GAMBAR
1 Bacillus thuringiensis KAHN15.39 5 2 Cendawan uji Curvularia affinis dan Colletotrichum gloeosporoides. 6 3 Penghambatan Bacillus thuringiensis KAHN 15.39 terhadap cendawan
patogen Curvularia affinis dan Colletotrichum gloeosporoides. 6 4 Penampakan hari kelima daun kelapa sawit yang diinokulasi cendawan. 7 5 Luas area terinfestasi cendawan Curvularia affinis pada berbagai
perlakuan bakteri 9
6 Luas area terinfestasi cendawan Colletotrichum gloeosporoides pada
berbagai perlakuan bakteri 10
7 Struktur anatomi daun kelapa sawit yang terinfestasi Curvularia affinis 11
8 Struktur anatomi daun kelapa sawit yang terinfestasi Colletotrichum
gloeosporoides 11

1
PENDAHULUAN
Latar Belakang
Kelapa sawit (Elaeis guineensis Jacq) merupakan tanaman produktif
daerah tropis yang cukup berkembang di Indonesia. Bahan mentah maupun
produk olahan dari kelapa sawit memiliki nilai ekonomi yang tinggi sebagai
penyumbang devisa negara. Berdasarkan data Kementerian Pertanian (2013), luas
lahan kelapa sawit di Indonesia diperkirakan 10 juta hektar dan produksi minyak
sawit sebanyak 27.7 juta ton dengan rata-rata produktivitas sawit 3.8 ton per
hektar per tahun. Sedangkan data Kementerian Perdagangan (2013) melaporkan
bahwa ekspor kelapa sawit berada pada kisaran 13 juta ton. Poduksi kelapa sawit
yang tinggi tersebut menempatkan Indonesia menjadi salah satu negara penghasil
kelapa sawit terbesar di dunia selain Malaysia dan Thailand.
Permintaan kelapa sawit yang cukup besar menyebabkan produksi dan
perluasan areal penanaman kelapa sawit semakin meningkat. Luas areal yang
meningkat berkaitan erat dengan pengadaan bibit berkualitas dalam jumlah besar
untuk produksi kelapa sawit yang optimal. Pembibitan di antaranya merupakan
faktor penentu budidaya kelapa sawit. Pembibitan menjadi langkah permulaan
untuk menentukan keberhasilan penanaman di lapangan. Beberapa faktor yang
memengaruhi pembibitan antara lain curah hujan, tanah, iklim, dan serangan
penyakit.
Keberadaan penyakit pada pembibitan kelapa sawit dapat menjadi faktor
pembatas pertumbuhan bibit. Penyakit yang sering ditemukan pada bibit kelapa
sawit yaitu bercak cokelat. Penyakit ini dapat merugikan pembibitan tanaman
kelapa sawit karena bibit menjadi kerdil, memperlama masa tanaman untuk mulai
berproduksi, meningkatkan kematian saat penanaman, dan menurunkan nilai jual
bibit. Beberapa cendawan patogen penyebab bercak daun pada kelapa sawit yaitu
Helminthosporium sp., Glomerella sp., Botridiplodia sp., Melacoiem elaedis, dan
Curvularia sp. (Ditjenbun 1993). Berdasarkan penelitian Solehudin et al. (2012)
mengenai status bercak cokelat di Kabupaten Sanggau, cendawan Curvularia
ditemukan pada bercak di seluruh lokasi pengamatan. Cendawan Curvularia dapat
bertahan hidup pada beberapa tumbuhan inang termasuk gulma di kebun kelapa
sawit. Pada bibit kelapa sawit cendawan Curvularia sp. menyerang bibit berumur
2 sampai 3 bulan. Cendawan Colletotrichum sp. merupakan cendawan umum
yang ditemukan dimana-mana. Penelitian cendawan ini pada tanaman kelapa
sawit belum banyak dilaporkan. Pada tanaman cabai cendawan Colletotrichum sp.
dapat menurunkan produksi dan kualitas cabai sebesar 45-60% (Wiratama et al.
2013).
Upaya pencegahan serangan penyakit pada kelapa sawit yang disebabkan
oleh cendawan patogen selama ini dilakukan dengan pemberian fungisida.
Pemanfaatan potensi mikrob seperti bakteri selulolitik yang mampu menghasilkan
enzim selulase sebagai pengendali biologi (biocontrol) berpotensi mengurangi
penggunaan fungisida sintetik. Selulosa merupakan salah satu komponen
penyusun dinding sel cendawan. Aktivitas enzim selulase dapat memotong ikatan
β-1,4-glikosidik pada selulosa sehingga bakteri selulolitik dapat digunakan untuk
menghambat pertumbuhan cendawan patogen yang menyerang kelapa sawit.

2
Penelitian yang telah dilakukan oleh Purnamasari (2013), menunjukkan
bahwa Bacillus thuringiensis KAHN 15.39 asal tanah hutan transformasi
(perkebunan) sekitar hutan Taman Nasional Bukit Dua Belas Jambi merupakan
bakteri selulolitik yang dapat menghambat cendawan patogen Curvularia affinis
dan Colletotrichum gloeosporoides. Penghambatannya dihitung melalui uji
tantang dan diperoleh sebesar 57.5% untuk Curvularia affinis dan 60% untuk
Colletotrichum gloeosporoides. Biakan cendawan yang digunakan merupakan
isolat yang diisolasi dari tanaman kelapa sawit di Balai Penelitian Bioteknologi
Perkebunan Indonesia, Bogor.
Tujuan Penelitian
Penelitian ini bertujuan menguji daya hambat Bacillus thuringiensis
KAHN15.39 terhadap cendawan patogen Curvularia affinis dan Colletotrichum
gloeosporoides, yang diisolasi dari daun bibit tanaman kelapa sawit, dengan
metode Leaf-pieces assays.
METODE
Waktu dan Tempat
Penelitian dilaksanakan bulan Januari 2014 - Juni 2014 yang bertempat di
Laboratorium Mikrobiologi dan Laboratorium Mikologi Departemen Biologi,
Fakultas MIPA, Institut Pertanian Bogor. Penyiapan preparat struktur anatomi
daun dilakukan di Laboratorium Zoologi LIPI Cibinong, Bogor.
Bahan
Bakteri yang digunakan ialah isolat Bacillus thuringiensis KAHN15.39
yang diisolasi oleh Purnamasari (2013). Cendawan yang digunakan ialah
Curvularia affinis dan Colletotrichum gloeosporoides yang diisolasi oleh
Purnamasari (2013) dan Haryanto (2013). Isolat bakteri dan cendawan yang
diujikan disimpan di IPB Culture Collection (IPBCC).
Daun kelapa sawit yang digunakan berasal dari bibit kelapa sawit yang
tumbuh di sekitar hutan Fakultas Perikanan IPB. Pemilihan daun didasarkan pada
kondisi daun yaitu sehat, utuh dan berasal dari bibit berumur 2 sampai 3 bulan
yang dicirikan dengan jumlah daun sekitar 3 sampai 4.
Peremajaan Isolat
Biakan bakteri diremajakan pada cawan yang berisi medium Nutrient Agar
(NA) yang mengandung 1% Carboxy Methyl Celullose (CMC) dan biakan
cendawan diremajakan pada cawan yang berisi medium Potato Dextrose Agar
(PDA) yang mengandung kloramfenikol 500 mg/L. Peremajaan bakteri dilakukan
dengan mengambil satu lup bakteri kemudian digoreskan dengan metode gores
kuadran pada medium NA yang mengandung 1% CMC. dan diinkubasi selama 24
jam. Selanjutnya biakan bakteri diamati melalui pewarnaan Gram. Peremajaan

3
cendawan dilakukan dengan mengambil koloni cendawan beserta mediumnya
dengan ukuran diameter 1 cm, kemudian diletakkan pada medium baru dan
diinkubasi selama 3-5 hari.
Isolat bakteri yang telah diremajakan dilakukan uji tantang terhadap
cendawan uji menggunakan metode Fokkema (1973). Sebanyak 1 lup isolat
bakteri digoreskan sepanjang 2 cm dari tepi cawan petri berisi medium Potato
Dextrose Agar (PDA) yang berdiameter 9 cm, kemudian diletakkan cendawan uji
dengan jarak 3 cm dari bakteri. Pasangan kultur tersebut diinkubasi selama 7 hari
kemudian dihitung zona hambat bakteri terhadap cendawan dibandingkan dengan
kontrol.
Persentase penghambatan = (R1 – R2) /R1 x 100%.
Keterangan:
R1 = jari-jari cendawan ke arah tepi cawan
R2 = jari-jari cendawan ke arah bakteri
Kemampuan Cendawan Uji Menginfestasi Daun Kelapa Sawit
Cendawan yang telah menghasilkan konidium pada cawan petri berisi
medium PDA diberi aquades steril sebanyak 10 mL (yang mengandung 50 μL
tween 20%) pada permukaannya untuk memanen konidium. Kultur yang
digunakan umur 1 minggu untuk Curvularia affinis dan 10 hari untuk
Colletotrichum gloeosporoides. Selanjutnya untuk memisahkan konidium dari
miselium dilakukan penyaringan menggunakan kertas saring. Suspensi konidium
Curvularia affinis dan Colletotrichum gloeosporoides masing-masing dihitung
menggunakan hemasitometer sehingga diperoleh 106 konidium/ml. Biakan bakteri
selulolitik dibiakkan dalam medium Nutrient Broth yang mengandung 1% CMC.
Biakan bakteri yang digunakan sebanyak 108
sel/ml. Kultur bakteri yang diujikan
terhadap Curvularia affinis berumur 24 jam, dan berumur 36 jam untuk
Colletotrichum gloeosporoides sesuai data daya hambat bakteri terbesar dari
penelitian sebelumnya (Purnamasari 2013).
Daun kelapa sawit diambil dari bibit berumur sekitar tiga bulan,
didesinfeksi dengan cara dicuci bersih dan diseka menggunakan alkohol 70%.
Potongan daun berukuran 3 cm x 4 cm masing-masing diinokulasi dengan 100 μL
suspensi konidium cendawan Curvularia affinis (106
konidium/ml) dan
Colletotrichum gloeosporoides masing-masing sebanyak 106
konidium/ml lalu
diletakkan di cawan petri steril besar berdiameter 9 cm yang berisi tisu lembap.
Setiap cawan petri berisi lima potongan daun yang merupakan lima ulangan untuk
setiap perlakuan per cawan dan satu cawan petri berisi kontrol. Pengamatan hasil
inokulasi dilakukan pada 5-7 hari masa inkubasi.
Reisolasi Cendawan
Reisolasi dilakukan dengan memotong bagian daun yang mengalami
perubahan warna menjadi kecokelatan. Potongan daun didesinfeksi menggunakan
larutan 2% NaClO selama kurang lebih 10 detik, dicuci dengan aquades steril
sebanyak tiga kali, dikeringkan menggunakan kertas saring steril, lalu diletakkan
pada medium PDA. Isolat cendawan yang diperoleh kemudian dibandingkan
dengan isolat cendawan awal (sebelum reisolasi) melalui pengamatan
makroskopis berupa warna koloni, bentuk koloni dan bentuk mikroskopis meliputi

4
bentuk konidium dan warna konidium menggunakan preparat yang dibuat dengan
metode Riddle (Riddle 1950).
Daya Hambat Bakteri terhadap Cendawan dengan
Metode Leaf-Pieces Assays
Uji penghambatan bakteri terhadap cendawan dilakukan menggunakan
metode Leaf-piece assays (Hsieh dan Huang 2001). Metode ini menggunakan
potongan daun dengan ukuran tertentu yang diberi perlakuan untuk diamati. Daun
bibit kelapa sawit yang berumur tiga bulan dipotong ukuran 3 cm x 4 cm. Setiap
satu potong untuk satu kali ulangan. Potongan-potongan daun tersebut digunakan
untuk dua macam percobaan, masing-masing percobaan terdiri atas lima
perlakuan dan satu kontrol negatif. Percobaan kesatu yaitu: 1) daun sebagai
kontrol negatif (tanpa perlakuan), 2) daun diinokulasi dengan 100 μl cendawan
Curvularia affinis saja, 3) daun diinokulasi dengan 100μL bakteri B. thuringiensis
KAHN 15.39 dan dua jam kemudian diinokulasi 100 μl konidium Curvularia
affinis, 4) daun diinokulasi dengan 100 μL bakteri B. thuringiensis KAHN 15.39
terlebih dahulu dan tiga hari kemudian diinokulasi dengan 100 μl konidium
Curvularia affinis, 5) daun diinokulasi dengan 100 μL Curvularia affinis dan dua
jam kemudian diinokulasi 100 μL bakteri B. thuringiensis KAHN 15.39, 6) daun
diinokulasi dengan 100 μL cendawan Curvularia affinis terlebih dahulu dan tiga
hari kemudian diinokulasi dengan 100 μl bakteri B. thuringiensis KAHN 15.39.
Percobaan kedua dilakukan seperti percobaan kesatu dengan
menggunakan cendawan Colletotrichum gloeosporoides. Masing-masing
perlakuan sebanyak empat ulangan. Suspensi spora cendawan yang digunakan
masing-masing 106
konidium/ml dan bakteri yang digunakan 108
sel/ml. Potongan
daun yang telah diberi perlakuan diletakkan dalam cawan petri steril berisi tisu
steril yang lembap. Potongan daun tersebut diletakkan di atas alumunium foil
yang dibentuk huruf U sebagai alas sehingga tidak bersentuhan dengan tisu untuk
mencegah daun terlalu basah. Pengamatan dilakukan selama tujuh hari dengan
tiga hari pertama percobaan dilakukan dalam keadaan gelap. Kerusakan daun
diamati dengan menghitung luas daun yang terinfestasi cendawan yaitu dengan
mengukur panjang dan lebar daun yang rusak, sedangkan daya hambat bakteri
diamati dengan menghitung persentase penghambatan menggunakan persamaan:
BE = (dc-dt)/dc x 100%; (Chanchaichaovivat et al. 2007)
Keterangan:
BE = efektivitas biokontrol (%)
dc = luas area terinfestasi dari kontrol positif (hanya perlakuan cendawan saja)
(mm2)
dt = luas area terinfestasi dari daun yang diberi perlakuan bakteri dan cendawan
(mm2)
Berdasarkan ukuran potongan daun yang digunakan luas area terinfestasi
cendawan akan memiliki nilai paling rendah 0 mm2
dan paling tinggi 1200 mm2,
sedangkan nilai BE memiliki nilai paling tinggi 100%.

5
Pengamatan Struktur Anatomi Daun Terinfestasi Cendawan
Pengamatan struktur anatomi daun yang terinfestasi cendawan dilakukan
pada daun kontrol negatif, daun yang diinokulasi cendawan terlebih dahulu dan 3
hari kemudian diinokulasi bakteri, serta daun kontrol positif (daun yang
diinokulasi cendawan saja). Daun tersebut dipotong secara melintang
menggunakan mikrotom beku. Hasil sayatan diberi pewarna safranin dan diamati
pada mikroskop dengan perbesaran 400x. Bagian yang diamati ialah jaringan
epidermis, berkas pembuluh dan jaringan mesofil.
HASIL
Peremajaan Isolat
Bacillus thuringiensis KAHN 15.39 merupakan bakteri Gram positif yang
berbentuk batang (Gambar 1). Bakteri ini dapat menghasilkan enzim selulase
ekstraseluler yang dicirikan dengan terbentuknya zona bening di sekitar koloni
bakteri yang ditumbuhkan pada medium NA dengan CMC 1%.
Gambar 1 Bacillus thuringiensis KAHN15.39
Curvularia affinis memiliki warna koloni awal abu-abu dan setelah
dewasa berwarna hitam, bentuk konidiofor gelap, pada umumnya sederhana,
menghasilkan konidium pada ujung atau secara simpodial. Konidium Curvularia
affinis berwarna gelap, fusiform, terdiri atas 3-5 sel dengan sel yang ujung lebih
kecil dan lebih cerah dibanding sel yang di tengah sehingga bentuknya bengkok
(Gambar 2).
Colletotrichum gloeosporoides memiliki warna koloni awal putih,
kemudian menjadi abu-abu, karakteristik konidiofor sederhana yang memanjang,
konidia hialin, 1 sel, berbentuk ovoid (berbentuk telur dengan satu ujungnya
menyempit) atau oblong (Gambar 2).

6
Gambar 2 Cendawan uji Curvularia affinis dan Colletotrichum
gloeosporoides. (a) Warna koloni dan (b) bentuk
konidium Curvularia affinis, (c) warna koloni dan (d)
bentuk konidium Colletotrichum gloeosporoides
Uji tantang B. thuringiensis KAHN 15.39 terhadap cendawan patogen
Curvularia affinis dan Colletotrichum gloeosporoides dilakukan selama 7 hari
masa inkubasi (Gambar 3). Kontrol dengan perlakuan hanya menggunakan kedua
cendawan patogen tumbuh memenuhi seluruh permukaan agar-agar (Gambar 3c
dan 3d).
Gambar 3 Penghambatan Bacillus thuringiensis KAHN 15.39 terhadap
cendawan patogen Curvularia affinis dan Colletotrichum
gloeosporoides. (a) B. thuringiensis KAHN 15.39 + Curvularia
affinis, (b) B. thuringiensis KAHN 15.39 + Colletotrichum
gloeosporoides, (c) kontrol Curvularia affinis, dan (d) kontrol
Colletotrichum gloeosporoides

7
Hasil uji tantang B. thuringiensis KAHN 15.39 terhadap cendawan uji
menunjukkan daya hambat sebesar 27.48% untuk Curvularia affinis dan 34.21%
untuk Colletotrichum gloeosporoides (Tabel 1). Berdasarkan penelitian yang
dilakukan Purnamasari (2013), diperoleh nilai penghambatan sebesar 57.5% untuk
Curvularia affinis dan 60% untuk Colletotrichum gloeosporoides.
Tabel 1 Daya hambat Bacillus thuringiensis KAHN 15.39 terhadap cendawan
Curvularia affinis dan Colletotrichum gloeosporoides melalui metode uji
tantang
Hari ke-
Daya Hambat (%)
Curvularia affinis Colletotrichum gloeosporoides
1 24.24 23.33
2 31.50 36.48
3 30.76 38.46
4 30.00 38.75
5 27.50 37.50
6 27.50 32.50
7 27.50 32.50
Rata-rata 28.48 34.21
Kemampuan Cendawan Uji Menginfestasi Daun Kelapa Sawit
Daun yang diinokulasi dengan cendawan menunjukkan gejala kerusakan
pada hari kelima. Daun yang diinokulasi Curvularia affinis menunjukkan gejala
pada hari kedua dengan terbentuk area terinfestasi cendawan berwarna cokelat di
pinggiran daun, melebar pada hari ketiga dan keempat, dan berubah menjadi gelap
cenderung hitam pada hari kelima (Gambar 4a).
Daun yang diinokulasi Colletotrichum gloeosporoides menunjukkan gejala
pada hari ketiga dengan terbentuk area terinfestasi cendawan berwarna cokelat di
pinggirann daun, melebar pada hari keempat dan pada hari kelima berwarna
cokelat sedikit berbayang kuning (Gambar 4b). Daun kontrol yang tanpa inokulasi
cendawan hanya diberikan aquades steril pada hari kelima tidak menunjukkan
adanya perubahan warna (Gambar 4c).
Gambar 4 Penampakan hari kelima daun kelapa sawit yang diinokulasi
cendawan. (a) Curvularia affinis (b) Colletotrichum
gloeosporoides dan (c) daun kontrol

8
Reisolasi Cendawan
Daun yang menunjukkan gejala berupa terbentuknya area terinfestasi
cendawan pada uji infestasi cendawan digunakan untuk reisolasi cendawan. Daun
yang menunjukkan gejala dari masing-maisng cendawan dipotong berukuran 1 cm
untuk diletakkan di atas permukaan medium PDA dan diinkubasi selama 7 hari.
Hasil reisolasi menunjukkan morfologi konidium dan konidiofor yang sama
dengan inokulan awal yaitu cendawan Curvularia affinis dan Colletotrichum
gloeosporoides (Gambar 2).
Pengamatan hasil reisolasi secara makroskopis dan mikroskopis dilakukan
pada hari ketujuh. Cendawan Curvularia affinis memiliki warna koloni hitam
dengan konidium fusiform, terdiri atas 3-5 sel, dan sel yang ujung lebih kecil serta
lebih cerah dibanding sel yang di tengah sehingga bentuknya bengkok (Gambar
2b). Cendawan Colletotrichum gloeosporoides memiliki warna koloni putih
dengan konidium hialin, 1 sel, berbentuk ovoid (berbentuk telur dengan satu
ujungnya menyempit) atau oblong (Gambar 2d).
Daya Hambat Bakteri terhadap Cendawan dengan
Metode Leaf-pieces assays
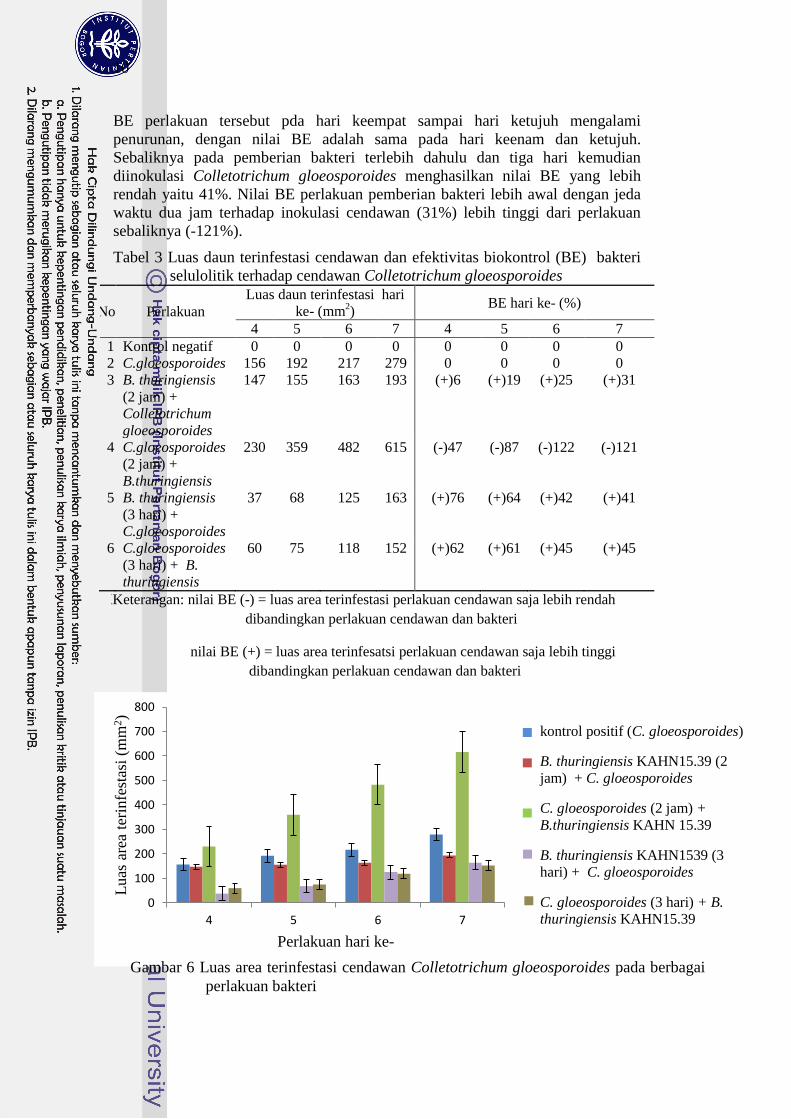
Luas daun terinfestasi cendawan pada hari ketujuh dengan perlakuan
inokulasi bakteri terlebih dahulu dan tiga hari kemudian diinokulasi Curvularia
affinis diperoleh sebesar 772 mm2. Nilai tersebut lebih besar dibandingkan dengan
perlakuan sebaliknya yaitu 69 mm2. Luas daun terinfestasi paling rendah
didapatkan pada perlakuan inokulasi cendawan terlebih dahulu dan tiga hari
kemudian diinokulasi bakteri dengan nilai sebesar 69 mm2 (Tabel 1).
Nilai BE paling tinggi sebesar 86% diperoleh pada perlakuan inokulasi
cendawan terlebih dahulu dan tiga hari kemudian diinokulasi bakteri. Nilai BE
perlakuan tersebut pada hari keempat sampai hari ketujuh mengalami penurunan,
nilai BE pada hari kelima dan keenam ialah sama, kemudian menurun pada hari
ketujuh. Sebaliknya pada pemberian bakteri terlebih dahulu dan tiga hari
kemudian diinokulasi Curvularia affinis tidak menghasilkan nilai BE (-50%).
Nilai BE perlakuan pemberian bakteri lebih awal dengan jeda waktu dua jam
terhadap inokulasi cendawan (7%) lebih tinggi dari perlakuan sebaliknya (-29%).

9
Tabel 2 Luas daun terinfestasi cendawan dan efektivitas biokontrol (BE) bakteri
selulolitik terhadap cendawan Curvularia affinis
No Perlakuan
Luas daun terinfestasi hari
ke- (mm2)
BE hari ke- (%)
4 5 6 7 4 5 6 7
1 Kontrol negatif 0 0 0 0 0 0 0 0
2 Curvularia affinis 81 273 322 513 0 0 0 0
3 B. thuringiensis
KAHN15.39 (2 jam) +
Curvularia affinis
75 235 292 480 (+)8 (+)14 (+)9 (+)7
4 Curvularia affinis (2
jam) + B.thuringiensis
KAHN 15.39
223 330 471 662 (-)174 (-)21 (-)46 (-)29
5 B. thuringiensis
KAHN1539 (3 hari) +
Curvularia affinis
208 404 552 772 (-)156 (-)48 (-)72 (-)50
6 Curvularia affinis (3
hari) + B. thuringiensis
KAHN15.39
5 10 13 69 (+)94 (+)96 (+)96 (+)86
Keterangan: nilai BE (-) = luas area terinfestasi perlakuan cendawan saja lebih rendah dari
perlakuan cendawan dan bakteri
nilai BE (+) = luas area terinfestasi perlakuan cedawan saja lebih tinggi dari
perlakuan cendawan dan bakteri
Luas daun terinfestasi cendawan pada hari ketujuh dengan perlakuan
inokulasi bakteri terlebih dahulu dan tiga hari kemudian diinokulasi
Colletotrichum gloeosporoides diperoleh sebesar 163 mm2. Perlakuan sebaliknya
dengan luas daun terinfestasi cendawan sebesar 152 mm2
merupakan luas daun
terinfestasi paling rendah jika dibandingkan dengan perlakuan lainnya. Nilai BE
paling tinggi sebesar 45% diperoleh pada perlakuan cendawan terlebih dahulu dan
tiga hari kemudian diinokulasi Colletotrichum gloeosporoides (152 mm2). Nilai
-100
0
100
200
300
400
500
600
700
800
900
1000
4 5 6 7
Luas
are
a te
rinfe
stas
i (m
m2)
Perlakuan pada hari ke-
Gambar 5 Luas area terinfestasi cendawan Curvularia affinis pada berbagai
perlakuan bakteri
kontrol positif (C. affinis)
B. thuringiensis KAHN15.39
(2 jam) + C. affinis
C. affinis (2 jam) +
B.thuringiensis KAHN 15.39
B. thuringiensis KAHN1539
(3 hari) + C. affinis
C. affinis (3 hari) + B.
thuringiensis KAHN15.39
10
BE perlakuan tersebut pda hari keempat sampai hari ketujuh mengalami
penurunan, dengan nilai BE adalah sama pada hari keenam dan ketujuh.
Sebaliknya pada pemberian bakteri terlebih dahulu dan tiga hari kemudian
diinokulasi Colletotrichum gloeosporoides menghasilkan nilai BE yang lebih
rendah yaitu 41%. Nilai BE perlakuan pemberian bakteri lebih awal dengan jeda
waktu dua jam terhadap inokulasi cendawan (31%) lebih tinggi dari perlakuan
sebaliknya (-121%).
Tabel 3 Luas daun terinfestasi cendawan dan efektivitas biokontrol (BE) bakteri
selulolitik terhadap cendawan Colletotrichum gloeosporoides
No Perlakuan
Luas daun terinfestasi hari
ke- (mm2)
BE hari ke- (%)
4 5 6 7 4 5 6 7
1 Kontrol negatif 0 0 0 0 0 0 0 0
2 C.gloeosporoides 156 192 217 279 0 0 0 0
3 B. thuringiensis
(2 jam) +
Colletotrichum
gloeosporoides
147 155 163 193 (+)6 (+)19 (+)25 (+)31
4 C.gloeosporoides
(2 jam) +
B.thuringiensis
230 359 482 615 (-)47 (-)87 (-)122 (-)121
5 B. thuringiensis
(3 hari) +
C.gloeosporoides
37 68 125 163 (+)76 (+)64 (+)42 (+)41
6 C.gloeosporoides
(3 hari) + B.
thuringiensis
60 75 118 152 (+)62 (+)61 (+)45 (+)45
0
100
200
300
400
500
600
700
800
4 5 6 7
Luas
are
a te
rinfe
stas
i (m
m2)
Perlakuan hari ke-
KKeterangan: nilai BE (-) = luas area terinfestasi perlakuan cendawan saja lebih rendah
dibandingkan perlakuan cendawan dan bakteri
nilai BE (+) = luas area terinfesatsi perlakuan cendawan saja lebih tinggi
dibandingkan perlakuan cendawan dan bakteri
kontrol positif (C. gloeosporoides)
B. thuringiensis KAHN15.39 (2
jam) + C. gloeosporoides
C. gloeosporoides (2 jam) +
B.thuringiensis KAHN 15.39
B. thuringiensis KAHN1539 (3
hari) + C. gloeosporoides
C. gloeosporoides (3 hari) + B.
thuringiensis KAHN15.39
Gambar 6 Luas area terinfestasi cendawan Colletotrichum gloeosporoides pada berbagai
perlakuan bakteri
11
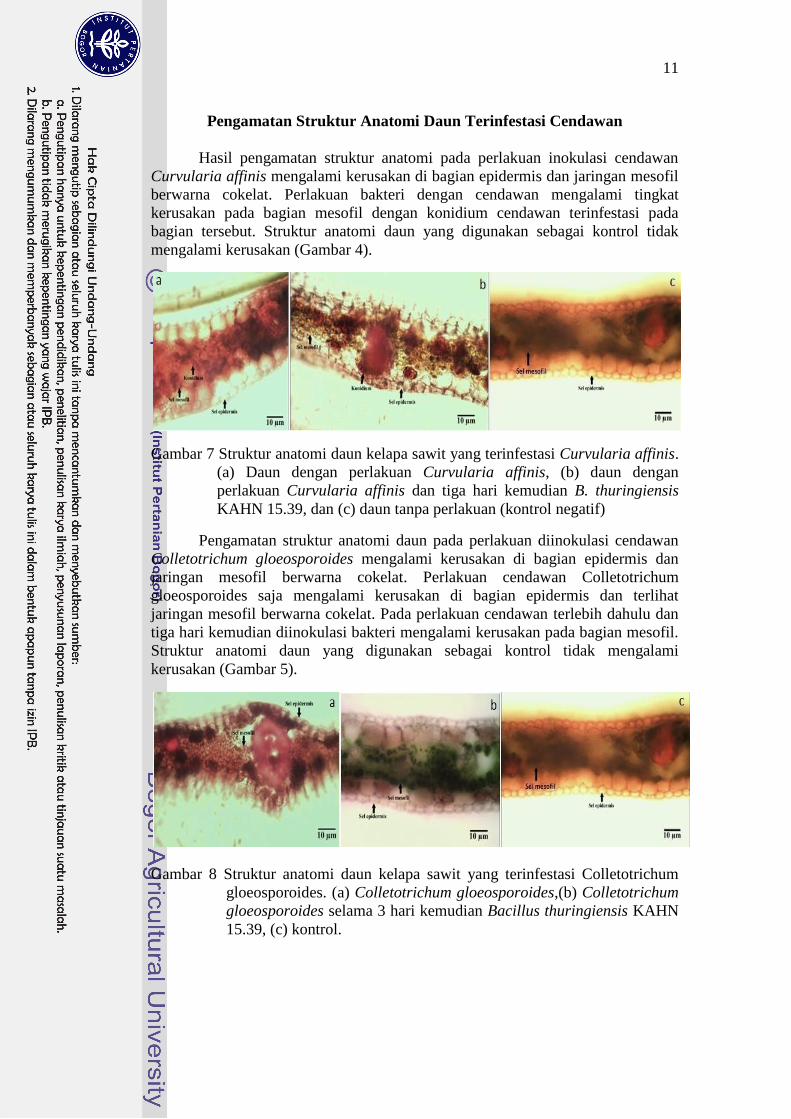
Pengamatan Struktur Anatomi Daun Terinfestasi Cendawan
Hasil pengamatan struktur anatomi pada perlakuan inokulasi cendawan
Curvularia affinis mengalami kerusakan di bagian epidermis dan jaringan mesofil
berwarna cokelat. Perlakuan bakteri dengan cendawan mengalami tingkat
kerusakan pada bagian mesofil dengan konidium cendawan terinfestasi pada
bagian tersebut. Struktur anatomi daun yang digunakan sebagai kontrol tidak
mengalami kerusakan (Gambar 4).
Gambar 7 Struktur anatomi daun kelapa sawit yang terinfestasi Curvularia affinis.
(a) Daun dengan perlakuan Curvularia affinis, (b) daun dengan
perlakuan Curvularia affinis dan tiga hari kemudian B. thuringiensis
KAHN 15.39, dan (c) daun tanpa perlakuan (kontrol negatif)
Pengamatan struktur anatomi daun pada perlakuan diinokulasi cendawan
Colletotrichum gloeosporoides mengalami kerusakan di bagian epidermis dan
jaringan mesofil berwarna cokelat. Perlakuan cendawan Colletotrichum
gloeosporoides saja mengalami kerusakan di bagian epidermis dan terlihat
jaringan mesofil berwarna cokelat. Pada perlakuan cendawan terlebih dahulu dan
tiga hari kemudian diinokulasi bakteri mengalami kerusakan pada bagian mesofil.
Struktur anatomi daun yang digunakan sebagai kontrol tidak mengalami
kerusakan (Gambar 5).
Gambar 8 Struktur anatomi daun kelapa sawit yang terinfestasi Colletotrichum
gloeosporoides. (a) Colletotrichum gloeosporoides,(b) Colletotrichum
gloeosporoides selama 3 hari kemudian Bacillus thuringiensis KAHN
15.39, (c) kontrol.

12
PEMBAHASAN
Daun kelapa sawit yang berumur 3 sampai 4 bulan dapat diinfeksi oleh
cendawan Curvularia affinis dan Colletotrichum gloeosporoides. Menurut Lubis
(2008), terdapat sejumlah patogen penyebab bercak cokelat pada pembibitan
kelapa sawit yaitu Botryodiplodia sp., Glomerella sp., Melanconium sp.,
Curvularia sp., Cochliobolus sp.. Semangun (2000) menyatakan bahwa jenis-jenis
patogen penyebab bercak cokelat di Indonesia belum banyak dilaporkan sehingga
dalam penelitian ini tidak dicantumkan pustaka mengenai cendawan
Colletotrichum gloeosporoides yang menyerang bibit kelapa sawit. Penelitian
yang telah dilakukan pada cendawan Colletotrichum gloeosporoides melaporkan
bahwa cendawan tersebut dapat menyebabkan penyakit antraknosa pada pepaya
(Hamdayanty et al. 2012)
Nilai BE pada perlakuan cendawan terlebih dahulu, baik Curvularia affinis
maupun Colletotrichum gloeosporoidedes dengan jeda waktu 2 jam diinokulasi
bakteri menunjukkan hasil yang negatif. Hal ini menunjukkan luas area
terinfestasi yang mendapat perlakuan bakteri lebih besar daripada perlakuan
cendawan saja (Gambar 7 dan 8). Bakteri pada perlakuan ini tidak memiliki daya
hambat terhadap pertumbuhan cendawan yang menyebabkan kerusakan pada
jaringan daun. Hal ini kemungkinan cendawan mampu menghasilkan suatu
senyawa untuk menginfestasi jaringan dalam waktu dua jam. Senyawa tersebut
kemungkinan mampu menghambat pertumbuhan bakteri pada saat diinokulasikan,
sehingga pertumbuhan bakteri tidak maksimum.
Perlakuan yang sebaliknya yaitu inokulasi bakteri terlebih dahulu dengan
jeda waktu dua jam diinokulasi cendawan menghasilkan nilai BE positif. Nilai BE
positif menunjukkan luas area terinfestasi cendawan yang mendapat perlakuan
bakteri lebih kecil daripada luas area terinfestasi cendawan tanpa mendapat
perlakuan bakteri. Hal tersebut kemungkinan dalam waktu dua jam bakteri telah
menginfestasi dan menghasilkan senyawa antagonis pada waktu terjadi interaksi
dengan cendawan. Pada perlakuan ini interaksi antagonis yang terjadi antara
bakteri dan cendawan berupa kompetisi ruang (Baker dan Cook 1982). Bacillus
thuringiensis KAHN 15.39 yang digunakan dalam penelitian ini berumur 24 jam
untuk Curvularia affinis dan 36 jam untuk Colletotrichum gloeosporoides. Bakteri
tersebut telah memasuki fase stasioner berdasarkan hasil penelitian Purnamasari
(2013).
Nilai BE menunjukkan negatif jika bakteri diinokulasi terlebih dahulu
dengan jeda waktu 3 hari terhadap inokulasi cendawan Curvularia affinis, namun
nilai BE positif terhadap inokulasi cendawan Colletotrichum gloeosporoides.
Berdasarkan hasil penelitian sebelumnya melalui uji tantang daya hambat bakteri
terhadap Curvularia affinis lebih rendah daripada terhadap Colletotrichum
gloeosporoides (Purnamasari 2013), sehingga bakteri pada waktu diujikan ke
substrat yang berbeda yaitu berupa jaringan daun tidak memberikan efek
penghambatan terhadap cendawan Curvularia affinis. Hal tersebut kemungkinan
pertumbuhan bakteri tidak optimum seperti pada uji tantang (ditumbuhkan pada
medium kaya nutrisi). Sedangkan daya hambat bakteri terhadap Colletotrichum
gloeosporoides yang lebih tinggi melalui uji tantang masih memberikan efek
penghambatan pada penelitian ini.

13
Perlakuan inokulasi cendawan terlebih dahulu, baik Curvularia affinis
maupun Colletotrichum gloeosporoides dan 3 hari kemudian diinokulasi bakteri
menunjukkan hasil BE positif. Nilai tersebut menunjukkan luas area terinfestasi
cendawan pada perlakuan dengan bakteri lebih kecil daripada perelakuan
cendawan saja (Gambar 7 dan 8). Hal ini mungkin disebabkan bakteri mampu
tumbuh dengan baik menggunakan nutrisi hasil degradasi jaringan daun oleh
cendawan. Bakteri menghasilkan senyawa tertentu pada saat terjadi interaksi
dengan cendawan seperti yang dilaporkan Hwang et al. (2001).
Perlakuan dengan nilai BE paling tinggi dan area terinfestasi cendawan
paling rendah pada penghambatan Curvularia affinis dan Colletotrichum
gloeosporoides didapatkan pada perlakuan dengan inokulasi cendawan terlebih
dahulu kemudian inokulasi bakteri setelah tiga hari. Hal ini mengindikasikan
bahwa bakteri Bacillus thuringiensis KAHN15.39 bersifat kuratif
(penanggulangan dilakukan setelah terinfeksi penyakit) dalam penerapan di daun.
Kerusakan yang terjadi pada struktur anatomi baik pada perlakuan
Curvularia affinis maupun Colletotrichum gloeosporoides (perlakuan cendawan
saja) menunjukkan kerusakan pada bagian mesofil. Cendawan dapat merusak sel
atau jaringan dari tumbuhan dan menyebabkan penurunan fungsi fisiologis sel
atau jaringan. Jenis sel dan jaringan yang terinfeksi akan menentukan fungsi
fisiologis yang dipengaruhinya. Infeksi pada daun akan mengganggu fotosintesis
(Agrios 1996) seperti yang dilaporkan oleh Dimaro et al. (2009) yang melakukan
penelitian mengenai histopatologi pada daun Raphia hookeri yang diserang
Glomerella cingulata. Jaringan yang terinfeksi menunjukkan perubahan warna
menjadi gelap dan gangguan transfusi pada jaringan. Setelah terjadi penetrasi pada
epidermis daun, patogen akan menyerang hipodermis dan menginfeksi jaringan
mesofil atau palisade. Cendawan dapat menyebabkan penyakit pada tumbuhan
melalui mekanisme seperti melemahkan inang dengan menyerap makanan terus-
menerus, menghasilkan atau mengganggu metabolisme sel inang dengan toksin,
enzim dan zat pengatur tumbuh yang disekresikannya, menghambat transportasi
makanan, hara mineral dan air melalui jaringan pengangkut, serta mengkonsumsi
kandungan sel inang setelah terjadi kontak (Agrios 1996)
Bakteri selulolitik sebagai biokontrol alami diharapkan mampu menghambat
pertumbuhan cendawan yang diduga patogen seperti yang telah dilaporkan Kucuk
dan Kivanc (2004), bahwa aktivitas β-1,3 glukanase yang dihasilkan oleh Bacillus
sp. dapat mendegradasi dinding sel cendawan Trichoderma harzianum. Senyawa
bioaktif yang dihasilkan oleh B. subtilis diketahui juga memiliki aktivitas terhadap
cendawan Rhizoctonia solani (Kondoh et al. 2001). Menurut Kim et al. (2004),
B. thuringiensis dapat menghasilkan lipopeptida dari kelompok fengycin yang
memiliki aktivitas antagonisme terhadap fitopatogen. Terdapat tiga kelompok
lipopeptida yang dihasilkan Bacillus sp. dalam peranannya sebagai agens
biokontrol yaitu surfactin, iturins, dan fengycins (Ongena dan Jacques 2007).
Pengamatan yang dilakukan pada penelitian penyakit tanaman umumnya
dengan menghitung derajat keparahan. Tingkat keparahan dipengaruhi beberapa
faktor yaitu kondisi bibit, umur inang, resistensi inang dan sifat genetik inang
(Sinaga 2009). Dalam penelitian ini tidak digunakan tanaman utuh sehingga
penghitungan bercak tidak dapat dilakukan. Bercak cokelat Curvularia paling
berpotensi menyerang bibit pada umur tiga bulan, sedangkan bibit umur empat
bulan sudah lebih tahan terhadap penularan bercak (Solehudin et al. 2012).

14
Penelitian terhadap cendawan patogen Curvularia yang selama ini dilakukan di
antaranya terhadap padi untuk menguji ketahahanannya (Taufik et al. 2012) dan
jagung untuk pengujian efektivitas antifungi (Ankinbode 2010).
Selama ini penelitian serangan bercak Colletotrichum lebih banyak
dilakukan pada cabai (Herwidyarti et al. 2013; Wiratama 2013), gulma dan
pepaya (Hamdayanty et al. 2012). Tingkat area terinfeksi dipengaruhi oleh masa
inkubasi cendawan pada inang. Menurut Herwidyarti et al. (2013), masa inkubasi
Colletotrichum cukup lama (3-27 hari) jika dibandingkan Curvularia. Hal
tersebut sejalan dengan hasil penelitian ini yaitu luas area terinfestasi cendawan
Colletotrichum gloeosporoides (279 mm2) lebih kecil dibandingkan Curvularia
affinis (513 mm2) (Tabel 1 dan 2). Daun setelah diinokulasi cendawan disimpan
dalam ruang tertutup dengan kondisi lembap selama 48 jam. Menurut
Sugiprihatini (2013), kelembapan selama 48 jam secara terus menerus sangat
rentan bagi tanaman untuk terserang penyakit.
Metode Leaf-pieces assays sering digunakan dalam meneliti ketahanan
inang terhadap cendawan dan efektivitas fungisida (Zadoks dan Schein 1979).
Beberapa penelitian yang menggunakan metode tersebut, bagian daun yang
dipotong ditutup agar atau medium untuk mencegah bakteri atau cendawan masuk
dari bagian tersebut (Boydom et al. 2013; Jackson et al. 2008), sehingga tingkat
keparahannya dapat dibandingkan dengan penelitian menggunakan tanaman utuh.
Kelebihan metode tersebut ialah reproduksibilitas lebih tinggi karena ukuran dan
umur daun sama, meningkatkan perbanyakan, lebih konsisten, kondisi inkubasi
seragam, peletakan inokulasi pada bagian yang spesifik di daun, kuantifikasi
penyakit lebih akurat dan mudah dilakukan (Parke et al. 2005). Kelemahan dari
metode Leaf-pieces assays ialah tidak dapat menjelaskan respon inang terhadap
patogen. Selain itu, pada penelitian ini gejala yang muncul pada daun tidak sesuai
dengan gejala bercak daun dan lebih mengarah pada gejala akibat saprofit bukan
parasit, sehingga kemungkinan metode Leaf-pieces assays tidak cocok digunakan
untuk semua jenis tanaman.
SIMPULAN DAN SARAN
Simpulan
Isolat Bacillus thuringiensis KAHN15.39 dapat menghambat cendawan
Curvularia affinis dengan tingkat efektivitas biokontrol sebesar 86% dan
cendawan Colletotrichum gloeosporoides dengan tingkat efektivitas biokontrol
sebesar 45% melalui metode Leaf-pieces assays. Penghambatan bakteri Bacillus
thuringiensis KAHN15.39 terhadap Curvularia affinis dan Colletotrichum
gloeosporoides bersifat kuratif yaitu penanggulangan dilakukan setelah terinfeksi
penyakit. Kerusakan struktur anatomi daun terjadi pada bagian epidermis dan
jaringan mesofil dengan tingkat kerusakan perlakuan bakteri dan cendawan lebih
rendah dibandingkan perlakuan cendawan saja.

15
Saran
Penggunaan metode Leaf-pieces assays sebaiknya disertai dengan
penelitian in planta untuk melihat respon inang terhadap patogen maupun agens
biokontrol.
DAFTAR PUSTAKA
Agrios GN. 1996. Ilmu Penyakit Tumbuhan. Busnia M, penerjemah. Yogyakarta
(ID): Gadjah Mada Pr. Terjemahan dari: Plant Pathology.
Akinbode, OA. 2010. Evaluation of antifungal efficacy of some plant extracts on
Curvularia lunata the causal organism of maize leaf spot. J Environ Sci
Technol. 4(11):797-800.
Baker KF, Cook RJ. 1982. Biological Control of Plant Pathogen. San Francisco
(US): American Phytopathological Society.
Boydom A, Dawit W, Getaneh W. 2013. Evaluation of detached leaf assay for
assessing leaf rust (Puccinia triticana Eriks) resistance in wheat. J Plant
Pathol Microb. 4(5):1-4
Chanchaichaovivat A, Ruenwongsa P, Panjipan B. 2007. Screening and
identification of yeast strains from fruits snd vegetables: potenstial for
biological control of postharvest chilli anthracnose (Colletotrichum capsici).
J Biol Control. 42:326-325.
[Ditjenbun] Direktorat Jendral Perkebunan (ID). 1993. Vademecum Budidaya
Kelapa Sawit (Elaeis guineensis). Serpong (ID): PT Perkebunan XI.
Hamdayanty, Yunita R, Amin NN, Damayanti TI. 2012. Pemanfaatan kitosan
untuk mengendalikan antraknosa pada pepaya (Colletotrichum
gloeosporoides) dan meningkatkan daya simpan buah. J Fitopatol Indones.
8(4):97-102.
Haryanto A. 2013. Isolation of chitinolytic bacteria used as biological control of
suspected pathogen fungi on oil palm seedlings [skripsi]. Bogor (ID): IPB
Univ Pr.
Herwidyarti KH, Ratih S, Sembodo DRJ. 2013. Keparahan penyakit antraknosa
pada cabai (Capsicum annum L) dan berbagai jenis gulma. J Agrotek Trop.
1(1):102-106.
Hsieh TF, Huang JW. 2001. Leaf-disk method assessment of disease severity of
lily leaf blight caused by Botrytis elliptica. Plant Pathol Bul. 10:37-44.
Hwang BK, Lee JY, Kim BS, Lim SW, Moon SS. 2001. Isolation and in vivo and
in vitro antifungal activity of phenylacetic acid and sodium phenylacetate
from Streptomyces humidus. J Appl Environ Microbiol. 67(1):3739-3745.
Jackson EW, Obert DE, Chong J, Avant JB, Bonman JM. 2008. Detached-leaf
method for propagating Puccinia coronata and assesing crown resistence in
oat. J Plant Dis. 92(10):1400-1406.
[Kementan] Kementerian Pertanian (ID). 2013. Statistik Pertanian 2013. Jakarta
(ID): Kementerian Pertanian.
Kim PI, Bai H, Bai D, Chae H, Chung S, Kim Y, Park R, Chi YT. 2004.
Purification and characterization of a lipopeptide produced by Bacillus
thuringiensis CMB26. J Appl Microbiol. 97(5):942-949.

16
Kondoh M, Hirai M, Shoda M. 2001. Integrated biological and chemical control
of damping-off caused by Rhizoctonia solani using Bacillus subtilis RB14-C
and flutolanil. J Biosci Bioeng. 1(2):173-177.
Kucuk C, Kivanc M. 2004. In vitro antifungal activity of strain of Trichoderma
harzianum. J Biol. 25(1):111-115.
Lubis AU. 2008. Kelapa sawit (Elaeis guinensis Jacq) di Indonesia. Pematang
Siantar (ID): Puslitbun Marihat.
Ongena M, Jacques P. 2007. Bacillus lipopeptides: versatile weapons for plant
disease biocontrol. J Trend Microbiol. 16(3):115-125.
Parke JL, Roth M, Choquette C. 2005. Detached Leaf Assays with Phytophthora
ramorum: Are They Valid? Sudden Oak Death Science Symposium II;18-21
January 2005; Monterey, California. Monterey (CA): Oregon State Univ Pr.
Purnamasari D. 2013. Isolasi bakteri selulolitik penghambat pertumbuhan
cendawan pada kelapa sawit dari tanah hutan di Jambi [skripsi]. Bogor (ID):
IPB Univ Pr.
Riddle RW. 1950. Permanent stained mycological preparation obtained by slide
culture. Mycologia. 42: 265-270.
Semangun H. 2000. Penyakit-penyakit Bibit Perkebunan di Indonesia.
Yogyakarta(ID): UGM Univ Pr.
Sinaga MS. 2003. Dasar-dasar Penyakit Tumbuhan. Depok (ID): Penebar
Swadaya.
Solehudin D, Suswanto I, Supriyanto. 2012. Status penyakit bercak coklat pada
pembibitan kelapa sawit di Kabupaten Sanggau. J Perkeb Lahan Trop. 2(1):
1-4.
Sugiprihatini D, Wiyono S, Widodo. 2011. Selection of yeasts antagonists as
biocontrol agent of mango fruit rot caused by Botryodiplodia theobromae. J
Microbiol Indones. 5(4): 155.
Taufik M, Asniah, Syair. 2012. Ketahanan lapangan padi gogo terhadap infeksi
Curvularia oryzae. J Fitopatol Indones. 8(2): 50-53.
Wiratama ID, Sudiarta IP, Sukewijaya IM, Sumiartha K, Utama MP. 2013. Kajian
ketahanan beberapa galur dan varietas cabai terhadap serangan antraknosa
di Desa Abang Songan Kecamatan Kintamani Kabupaten Bangli. J
Agroekotek Trop. 2(2):71-81.
Zadoks JC, Schein RD. 1979. Epidemiology and Plant Disease Management.
New York (US): Oxford Univ Pr.

17
RIWAYAT HIDUP
Penulis dilahirkan di Kuningan pada tanggal 11 Maret 1991 dari ayah
Mohammad Jahuri dan Almh. ibu Siti Khodijah. Penulis merupakan anak
kedelapan dari delapan bersaudara. Tahun 2010 penulis lulus dari MAN Insan
Cendekia Gorontalo. Pada tahun yang sama, penulis lulus seleksi masuk IPB
melalui jalur Seleksi Nasional Masuk Perguruan Tinggi Negeri (SNMPTN) di
Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam. Penulis
mendapatkan beasiswa dari BCA Finance tahun 2011-2013 dan beasiswa dari
Women International Club (WIC) tahun 2013-2014.
Penulis aktif mengikuti beberapa organisasi dalam masa studi seperti
Himpunan Mahasiswa Biologi (HIMABIO) periode 2011-2012 dan 2012-2013
pada divisi Bioworld, Ikatan Alumni Insan Cendekia Gorontalo (IAICG) periode
2011-sekarang dan Ikatan Santri Mahasiswa Al Ihya (ISMA) periode 2012-2014.
Penulis juga aktif mengikuti kegiatan seperti menjadi peserta kompetisi olimpiade
biologi pada OSN Pertamina (2011-2012) dan peserta pelatihan Leadership and
Enterpreunership School (LES) IPB (2011). Penulis juga aktif mengikuti lomba
Karya Tulis Ilmiah seperti pada The 2nd
Airlangga Ideas Competition (AIC),
Bisnis Challenge UI Green Festival, Karya Tulis Ilmiah Al Quran (LKTIA) IPB
dan Pekan Kreativitas Mahasiswa Penelitian (PKMP) yang didanai oleh DIKTI.
Penulis juga mengikuti beberapa kegiatan kepanitiaan seperti pada Lomba Karate
se Jawa-Bali (2010), Explo Science (2012), Fun with Bioworld (2012), Biology
on Science and Application (2012/2013).
Selama mengikuti perkuliahan, penulis pernah menjadi asisten praktikum
Biologi Dasar (2013). Tanggal 3-5 Juli 2012 penulis melaksanakan Studi Lapang
di Taman Nasional Gunung Gede-Pangrango (TNGGP) dengan judul Struktur
Sekretori Tanaman Obat Anggota Suku Asteraceae di Taman Nasional Gunung
Gede Pangrango, Jawa Barat. Setelah itu, pada bulan Juli hingga Agustus 2013,
penulis melaksanakan Praktik Lapangan dengan topik Perbanyakan Bibit dan
Perawatan Tanaman Anggrek Dendrobium sp. di Taman Anggrek Indonesia
Permai (TAIP) dan Pondok Bunga Nursery.