cabai rawit 1
TRANSCRIPT

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 1/97
VOLUME 2
NOMOR 3
DESEMBER 2014

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 2/97

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 3/97

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 4/97
DAFTAR ISI
PENGARUH PEMBERIAN PUPUK HAYATI ( BIOFERTILIZER) DAN
MEDIA TANAM YANG BERBEDA PADA PERTUMBUHAN DAN
PRODUKTIVITAS TANAMAN CABAI RAWIT (Capsicum frutescens L.) DI
POLYBAG Agus supriyanto, Fita Khoirul Umah, Tini Surtiningsih
AKTIVITAS ANTIBAKTERI DAN ANTIFUNGI EKSTRAK PETROLEUM ETER
Dumortiera hirsuta
Junairiah, Hanik Faizah, dan Salamun
KEANEKARAGAMAN DAN POLA DISTRIBUSI LONGITUDINAL
SPESIES KERANG AIR TAWAR CORBICULIDAE DI SUNGAI BRANTAS PERIODE
JANUARI – FEBRUARI 2012
Ichsan Wardani, Drs. Moch. Affandi, M.Si., dan Dr. Bambang Irawan, M.Sc.
PENGARUH KOMBINASI ZAT PENGATUR TUMBUH KINETIN DAN IBA
TERHADAP KULTUR ANTERA CABAI RAWIT (Capsicum frutescens L.)
Irma Catur Prastyo W.(1), Dwi Kusuma Wahyuni (*)(1), dan Hery Purnobasuki (1)
DETEKSI ENZIM LIPASE DAN BIOSURFAKTAN PADA SUPERNATAN
KULTUR Bacillus sp. LII63B YANG DITUMBUHKAN PADA MINYAK
KELAPA
Ni’matuzahroh (*), Isnaini Septi Irmayanti, Tini Surtiningsih, Fatimah, Sri Sumarsih (**)
UJI ANTIBAKTERI EKSTRAK KULIT BUAH DAN BIJI MANGGIS (Garcinia
mangostana) PADA BAKTERI PENYEBAB JERAWAT (Staphylococcus epidermidis )
DENGAN MENGGUNAKAN SOLVEN ETANOL
Krisnina Maharani, Drs. Agus Supriyanto, M. Kes, dan Tri Nurhariyati, S. Si, M.
KAJIAN HISTOLOGI GONAD TERIPANG Phyllophorus sp. PADA BULANFEBRUARI, MARET DAN APRIL 2012Dwi Winarni(1), Moch. Affandi(1), Endang Dewi Masithah(2), Machmudhatun Nisa(1)
PENGARUH PEMBERIAN KONSORSIUM MIKROBA BIOFERTILIZER TERHADAP
PERTUMBUHAN DAN PRODUKSI JAMUR TIRAM PUTIH ( Pleorotus ostreatus)
Rizka Rakhmawati, Tini Surtiningsih, Tri Nurhariyati
Kajian Viabilitas dan Pola Pertumbuhan Lactobacillus plantarum pada Variasi Konsentrasi
Molase dan Waktu Inkubasi
Agus Supriyanto, Rochma Novirisandi, Ni’matuzahroh
PENGARUH KOMBINASI ZAT PENGATUR TUMBUH
BENZYLADENINE (BA) DAN INDOLE BUTIRIC ACID (IBA)
TERHADAP KULTUR ANTERA CABAI RAWIT (Capsicum frutescens L.)
Tining Sulistyowati1, Dwi Kusuma Wahyuni*1, dan Hery Purnobasuki1
1-11
12-22
23-30
31-38
39-47
48-57
58-67
68-76
77-86
87-93

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 5/97
PENGARUH PEMBERIAN PUPUK HAYATI ( BIOFERTILIZER) DAN
MEDIA TANAM YANG BERBEDA PADA PERTUMBUHAN DAN
PRODUKTIVITAS TANAMAN CABAI RAWIT (Capsicum frutescens L.) DI POLYBAG
Agus supriyanto, Fita Khoirul Umah, Tini SurtiningsihDepartemen Biologi, Fakultas Sains dan Teknologi
Universitas Airlangga Surabaya
Abstract
This research is aimed at knowing the effect of giving various doses ofbiofertilizer , using growing media, and combining biofertilizer and growing media on
growth and productivity of chili (Capsicum frutescens L.). This research is conductedat the Microbiology Laboratory Faculty of Science and Technology Airlangga
University and at the greenhouse of UPT Development of Food Crops and
Horticulture Agribusiness in Lebo, Sidoarjo, for six months from February 2012 until
July 2012. This research uses factorial design namely 2x5 pattern and with repetitionthree times. The first factor is the dose of fertilizer (D) consisting of D- : 0 ml dose of
biofertilizer / plant, D +: 10 g NPK fertilizer/ plant, D5: 5 ml dose of biofertilizer /
plant, D10: 10 ml dose of biofertilizer / plant , and D15: 15 ml dose of biofertilizer / plant. The second factor is the growing media (M) consisting of M1 (soil) and M2
(soil : compost with ratio 1:1). Growth parameter applied is the number of leaves
(sheet) and plant height (cm), while the productivity parameter used is the number offruits (fruit) and fruit weight (g). The data of observation were descriptively analyzed
and base on the value of its RAE( Relative Agronomic Effectiveness) . Based on result
of research, biofertilizer doses have effect on the growth and productivity of chili,
with the highest growth in doses D- and D15 (number of leaves), D+ (plant height),and the highest productivity in dose D10 (number of fruits and fruit weight). Growing
media have affects on the growth and productivity of chili, with the highest growth
in M1 growing media (number of leaves and plant height), and the highest productivity in M1 growing media (number of fruits and fruit weight).The
combination of biofertilizer and the media have effect on plant growth and
productivity of chili, with the highest growth in combination of biofertilizer and themedia M1D- (number of leaves), M1D15 (plant height), and the highest productivity
in combination of biofertilizer and the media M1D+ (number of fruits), M2D10
(fruit weight). Based on value of its RAE, the doses that effective to substitutechemistry fertilizer are D15 on M1 and D10 on M2.
Key words: Capsicum frutescens L., Biofertilizer , growing media, growth,
productivity

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 6/97
Pendahuluan
Cabai rawit (Capsicum frutescens L.) merupakan salah satu tanamanhortikultura dari jenis sayuran yang memiliki buah kecil dengan rasa yang pedas.
Produksi tanaman cabai rawit ini dari tahun ke tahun terus meningkat, tahun 2009
produksinya sebesar 591.294 ton, sedangkan pada tahun 2010 produksinya sebesar
521.704 ton. Setahun terahir ini produksi tanaman cabai rawit mengalami penurunan
sebanyak 69.590 ton (Deptan, 2011). Selain itu cabai rawit harganya di pasaran
seringkali lebih tinggi dari pada cabai jenis lainnya. Hal ini dikarenakan tidak sedikit
petani yang mengalami gagal panen. Terjadinya gagal panen diakibatkan karena
adanya beberapa kendala, terutama tingkat kesuburan tanah dan hama yang
berkembang di tengah udara lembab sehingga membuat bunga, daun dan tanaman
cabai rusak akhirnya mengakibatkan kegagalan panen (Anonimus, 2011).
Untuk meningkatkan hasil produksi cabai rawitnya, para petani berusaha
mengatasi kendala tersebut dengan melakukan pemupukan menggunakan pupuk
kimia. Akan tetapi pupuk kimia sering mengalami kelangkaan sehingga harganya
melonjak tinggi. Selain itu pemakaian pupuk ini dapat menyebabkan pencemaran
tanah, menurunkan pH tanah, cepat terserapnya zat hara dapat menjadikan tanah
menjadi miskin akan unsur hara, khususnya unsur hara mikro yang sangat diperlukan
oleh tanaman untuk meningkatkan hasil dan daya tahan tanaman terhadap serangan
hama dan penyakit (Syaifudin dkk., 2010).
Dengan adanya berbagai kendala tersebut, untuk meningkatkan produksi tanaman
cabai rawit dapat menggunakan alternatif pupuk hayati (biofertilizer ) yang
mempunyai keutungan ekologis maupun ekonomis, selain itu pupuk hayati yang
berbahan aktif organisme hidup ini dapat berfungsi sebagai penambat hara tertentu
atau memfasilitasi tersedianya hara dalam tanah bagi tanaman (Simanungkalit dkk.,
2006). Penggunaan pupuk hayati memerlukan takaran dosis pemupukan yang
disesuaikan dengan jenis media tanam yang dipakai agar hasilnya dapat sesuai
dengan harapan.

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 7/97
Sebagian besar unsur-unsur hara yang dibutuhkan tanaman disediakan melalui
media tanam, selanjutnya diserap oleh perakaran dan digunakan untuk proses
fisiologis tanaman (Ermina, 2010). Media tanam yang umum digunakan adalah tanah,
karena di dalam tanah tersedia faktor-faktor utama untuk pertumbuhan tanaman seperti
unsur hara, air, dan udara (Ningrum, 2010). Akan tetapi menurut Isroi (2009), kondisi
tanah sekarang semakin mengalami penurunan karena rendahnya bahan organik.
Sehingga para petani mengkombinasikan tanah dengan kompos.
Penanaman cabai membutuhkan lahan yang luas, tetapi lahan yang ada semakin
sempit, kondisi ini dapat diatasi dengan menanam cabai rawit di dalam polybag yang
juga dapat mempermudah pengamatan pertumbuhan dan produktivitasnya.
Berdasarkan latar belakang permasalahan tersebut, maka peneliti ingin melakukan
penelitian dengan judul “pengaruh pemberian pupuk hayati (biofertilizer ) dan media
tanam yang berbeda pada pertumbuhan dan produktivitas tanaman cabai rawit
(Capsicum Frutescens L.) di polybag ”.
Bahan dan Metode
Tahap Pembuatan Biofertilizer
Masing-masing inokulan yang terdiri atas : (1) Azotobacter sp. dan
Azospirillum sp. (bakteri fiksasi nitrogen non simbiotik); (2) Rhizobium sp. (bakteri
fiksasi nitrogen simbiotik); (3) Bacillus megaterium dan Bacillus subtilis (bakteri
pelarut fosfat); (4) Pseudomonas sp. (bakteri pelarut fosfat); (5) Cellulomonas sp.
(bakteri dekomposer); (6) bakteri dekomposer Lactobacillus sp.; dan (7)
Saccharomyces cereviceae ( yeast dekomposer) yang telah diremajakan, diambil satu
ose, kemudian ditanam pada tujuh botol kultur yang masing-masing botol berisi 100
ml media NB kemudian di homogenkan dengan shaker selama semalam. Setelah itu
biakan tersebut diinkubasi selama 24 jam pada suhu ruangan. Starter yang telah
diinkubasi kemudian dicampurkan dalam jerigen yang sudah berisi 6300 ml larutan
molase 2% dan dihomogenkan, maka pupuk hayati (biofertilizer ) sudah selesai

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 8/97
dibuat. Tetapi sebelum dipakai dalam proses pemupukan, pupuk tersebut harus
diinkubasi selama 24 jam terlebih dahulu untuk pengujian analisis kualitas pupuk .
Analisis kualitas pupuk hayati (biofertilizer )
Pada analisis kualitas pupuk ini dibutuhkan enam macam media untuk
menumbuhkan mikroba-mikroba yang ada dalam pupuk hayati. Keenam media
tersebut adalah: (1). Media semi solid NFB ( Nitrogen Fixing Bacteria) untuk
Azotobacter sp. dan Azospirillum sp. (2). Media Pikovskaya untuk Pseudomonas sp.,
Bacillus subtilis, dan Bacillus megaterium. (3). Media MSA ( Mannitol Salt Agar )
untuk Rhizobium sp.. (4). Media PDA ( Potato Dextose Agar ) untuk Saccharomyces
cereviceae. (5). Media MRSA ( Mannitol Rhogasa Sharpe Agar ) untuk actobacillus
sp.. (6). Media CMC Agar (Carboxyl Methil Cellulose) untuk Cellulomonas sp..
Tahap Penanaman Cabai Rawit
Proses penanaman cabai rawit terdiri atas beberapa tahap, yaitu : pembenihan,
penyiapan media tanam, penanaman, pemeliharaan, pemanenan, dan perawatan hasil
panen.
Prosedur Pengambilan Data
Data yang diambil untuk mengetahui proses pertumbuhan adalah tinggi
tanaman (cm) yang diukur dari permukaan tanah sampai ujung kuncup teratas dan
jumlah daun (helai) per tanaman. Sedangkan untuk proses produktivitas, data yang
diambil adalah jumlah buah (buah) per tanaman yang datanya didapat dari tiga kali
pengambilan dan berat buah (g/pohon cabai), buah yang dipanen adalah buah yang
sudah tua atau sudah matang.
Analisis Data
Data yang didapat dari penelitian ini dianalisis secara deskripsi dan dihitung
nilai RAE ( Relative Agronomic Effectiveness)nya.
Nilai RAE dihitung dengan persamaan :
RAE = hasil pada pupuk hayati yang diuji - hasil pada kontrol x 100%
hasil pada pupuk standar – hasil pada kontrol
(Machay et al . (1984) dalam Saraswati dkk. (2008).
7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 9/97
Hasil dan Pembahasan
a. Pengaruh dosis pupuk hayati (biofertilizer ) terhadap pertumbuhan dan
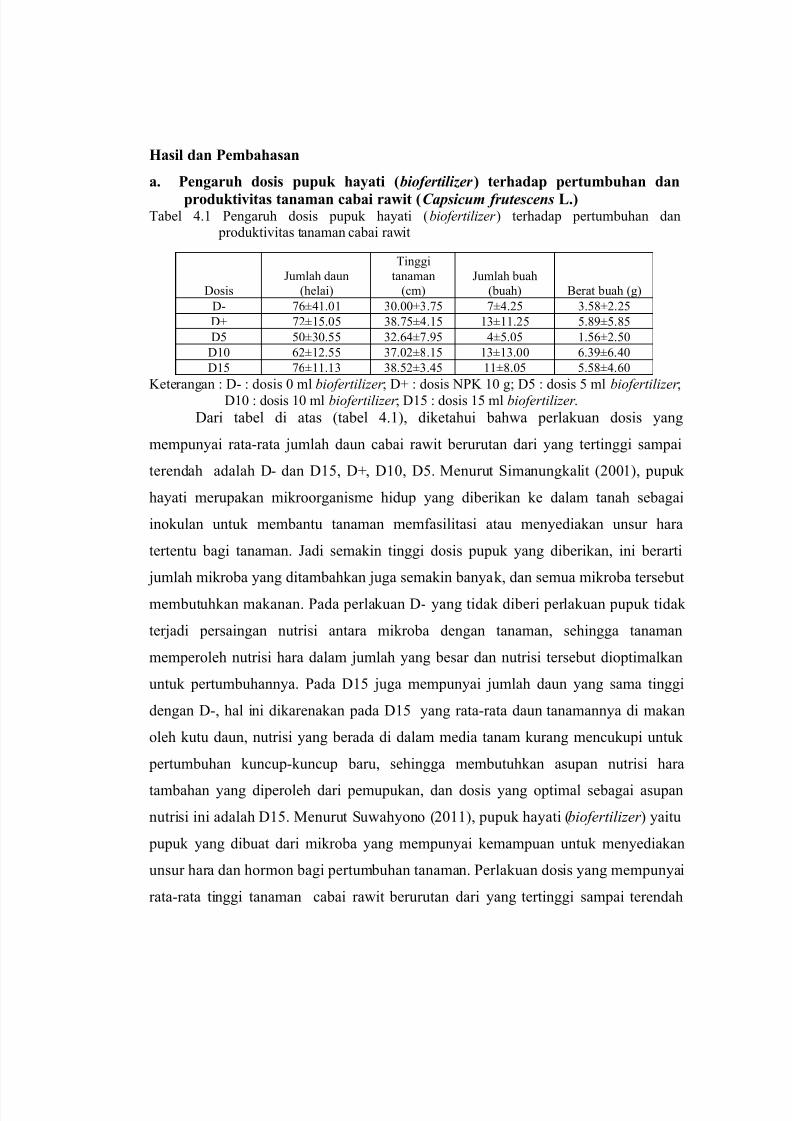
produktivitas tanaman cabai rawit (Capsicum frutescens L.)Tabel 4.1 Pengaruh dosis pupuk hayati (biofertilizer ) terhadap pertumbuhan dan
produktivitas tanaman cabai rawit
Dosis
Jumlah daun
(helai)
Tinggi
tanaman
(cm)
Jumlah buah
(buah) Berat buah (g)
D- 76±41.01 30.00±3.75 7±4.25 3.58±2.25
D+ 72±15.05 38.75±4.15 13±11.25 5.89±5.85
D5 50±30.55 32.64±7.95 4±5.05 1.56±2.50
D10 62±12.55 37.02±8.15 13±13.00 6.39±6.40
D15 76±11.13 38.52±3.45 11±8.05 5.58±4.60
Keterangan : D- : dosis 0 ml biofertilizer ; D+ : dosis NPK 10 g; D5 : dosis 5 ml biofertilizer ;
D10 : dosis 10 ml biofertilizer ; D15 : dosis 15 ml biofertilizer .
Dari tabel di atas (tabel 4.1), diketahui bahwa perlakuan dosis yang
mempunyai rata-rata jumlah daun cabai rawit berurutan dari yang tertinggi sampai
terendah adalah D- dan D15, D+, D10, D5. Menurut Simanungkalit (2001), pupuk
hayati merupakan mikroorganisme hidup yang diberikan ke dalam tanah sebagai
inokulan untuk membantu tanaman memfasilitasi atau menyediakan unsur hara
tertentu bagi tanaman. Jadi semakin tinggi dosis pupuk yang diberikan, ini berarti
jumlah mikroba yang ditambahkan juga semakin banyak, dan semua mikroba tersebut
membutuhkan makanan. Pada perlakuan D- yang tidak diberi perlakuan pupuk tidak
terjadi persaingan nutrisi antara mikroba dengan tanaman, sehingga tanaman
memperoleh nutrisi hara dalam jumlah yang besar dan nutrisi tersebut dioptimalkan
untuk pertumbuhannya. Pada D15 juga mempunyai jumlah daun yang sama tinggi
dengan D-, hal ini dikarenakan pada D15 yang rata-rata daun tanamannya di makan
oleh kutu daun, nutrisi yang berada di dalam media tanam kurang mencukupi untuk
pertumbuhan kuncup-kuncup baru, sehingga membutuhkan asupan nutrisi hara
tambahan yang diperoleh dari pemupukan, dan dosis yang optimal sebagai asupan
nutrisi ini adalah D15. Menurut Suwahyono (2011), pupuk hayati (biofertilizer ) yaitu
pupuk yang dibuat dari mikroba yang mempunyai kemampuan untuk menyediakan
unsur hara dan hormon bagi pertumbuhan tanaman. Perlakuan dosis yang mempunyai
rata-rata tinggi tanaman cabai rawit berurutan dari yang tertinggi sampai terendah

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 10/97
adalah D+, D15, D10, D5, D-. Perlakuan dengan D+ mempunyai nilai lebih tinggi
dari pada yang lainnya, hal ini menurut Gardner dkk. (1991), yang menyatakan
bahwa sepanjang masa pertumbuhan vegetatif, akar, daun, dan batang merupakan
bagian-bagian dari tanaman yang kompetitif dalam pemanfaatan hasil asimilasi. Jadi
kemungkinan proporsi energi yang dibutuhkan untuk proses pertumbuhan tinggi
tanaman lebih besar dari pada energi yang dibutuhkan untuk proses pertumbuhan
bagian tanaman lainnya.
Perlakuan dosis yang mempunyai rata-rata jumlah buah cabai rawit berurutan
dari yang tertinggi sampai terendah adalah D10, D+, D15, D-, D5. Meskipun D15
pada pengamatan jumlah daun yang memiliki jumlah tertinggi, tetapi D10 merupakan
dosis yang optimum untuk proses pembuahan, ini berarti dalam optimalisasi
produktivitas tanaman diperlukan pengurangan jumlah dosis pupuk, terutama pupuk
yang mengandung unsur N, karena jika tidak dihentikan maka pertumbuhan vegetatif
akan tumbuh terus menerus, sehingga akan menghambat pertumbuhan generatif, hal
ini diperkuat dengan pendapat Anonimus a (2008), yang menyatakan dosis pupuk
hayati yang dibutuhkan pada fase vegetatif berbeda dengan kebutuhan pada fase
generatif. Perlakuan dosis yang mempunyai rata-rata berat buah cabai rawit
berurutan dari yang tertinggi sampai terendah adalah D10, D+, D15, D-, D5.
Perlakuan D10 mempunyai nilai berat buah tertinggi, ini disebabkan selain dosis ini
optimum untuk peningkatan jumlah buah, dosis ini juga optimum untuk
meningkatkan berat buah cabai rawit. Sehingga ukuran buah pada perlakuan ini lebih
besar dari pada perlakuan yang lainnya, ini menyebabkan beratnya juga lebih tinggi.
b. Pengaruh media tanam terhadap pertumbuhan dan produktivitas tanaman
cabai rawit (Capsicum frutescens L.)
Tabel 4.2 Pengaruh media tanam terhadap jumlah daun (helai) cabai rawit
Media tanam
Jumlah daun
(helai)
Tinggi tanaman
(cm)
Jumlah buah
(buah)
Berat buah
(g)
M1 74±23.16 42.63±6.62 10±7.64 5.34±4.16
M2 60±20.92 28.13±4.36 8±9.00 3.86±4.48
Keterangan : M1: media tanam tanah; M2: media tanam tanah dan kompos.
7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 11/97
Secara deskriptif dari tabel 4.2, perlakuan media tanam yang mempunyai
rata-rata jumlah daun, tinggi tanaman, jumlah buah, dan berat buah cabai rawit
berurutan dari yang tertinggi sampai terendah adalah M1, M2. Pendapat
Hardjowigeno (2003) dalam Imanda dan Ketty (2012), yang mengatakan bahwa
tanah merupakan sumber utama zat hara untuk tanaman.Tanah mengandung unsur
hara esensial makro dan mikro. Unsur-unsur hara ini diserap akar tanaman dari dalam
tanah, hal ini yang menyebabkan perlakuan M1 yaitu media tanam tanah memiliki
nilai jumlah daun, tinggi tanaman, jumlah buah, dan berat buah lebih tinggi daripada
perlakuan M2. Selain itu mungkin karena tanah mempunyai sifat yang solid sehingga
hal ini menyebabkan penancapan perakaran tanaman lebih kuat, ini akan membantu
mengoptimalkan penyerapan hara dalam tanah.
c. Pengaruh dosis pupuk hayati (biofertilizer ) dan media tanam terhadap
pertumbuhan dan produktivitas tanaman cabai rawit (Capsicum frutescens
L.)
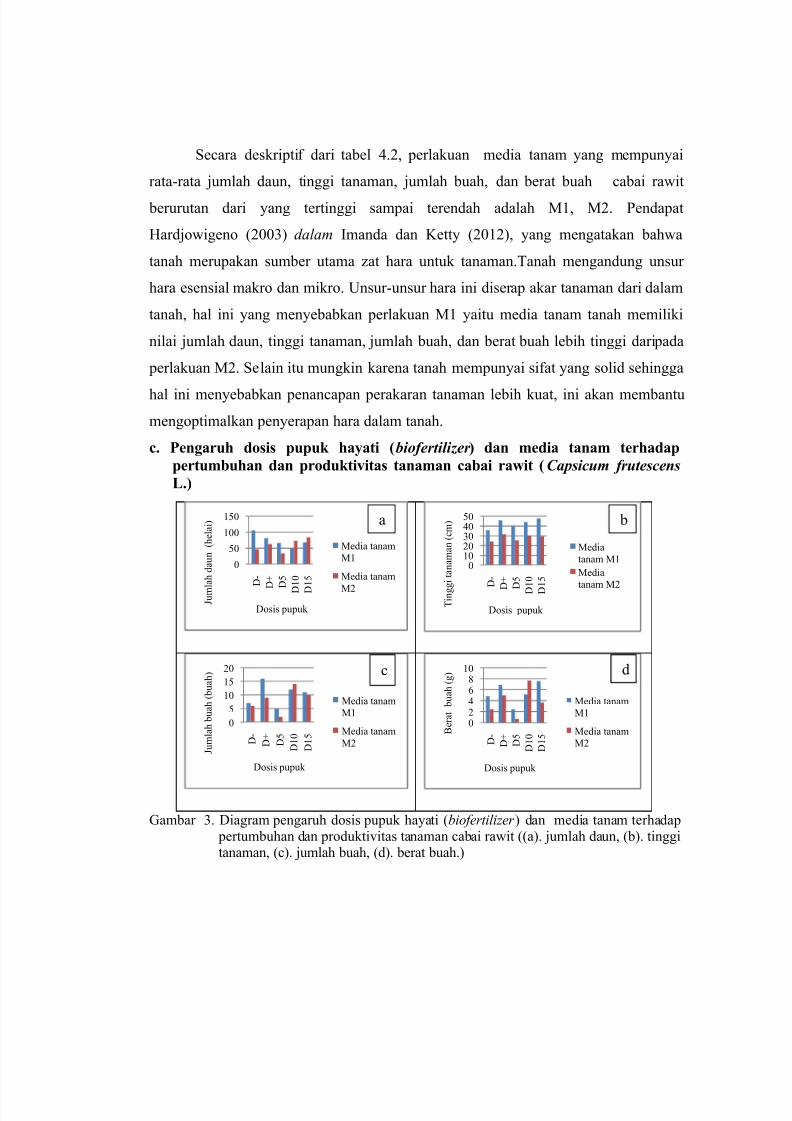
Gambar 3. Diagram pengaruh dosis pupuk hayati (biofertilizer ) dan media tanam terhadap pertumbuhan dan produktivitas tanaman cabai rawit ((a). jumlah daun, (b). tinggitanaman, (c). jumlah buah, (d). berat buah.)
0
50
100
150
D -
D +
D 5
D
1 0
D
1 5
J u m l a h d a u n
( h e l a i )
Dosis pupuk
Media tanam
M1
Media tanam
M2
01020304050
D -
D
+ D 5
D
1 0
D
1 5
T i n g g i t a n a m a n ( c m )
Dosis pupuk
Media
tanam M1
Media
tanam M2
0
5
10
15
20
D -
D +
D 5
D 1 0
D 1 5
J u m l a h b u a h ( b u a h )
Dosis pupuk
Media tanam
M1
Media tanam
M2
02468
10
D -
D +
D 5
D 1 0
D 1 5
B e r a t b u a h ( g )
Dosis pupuk
Media tanam
M1
Media tanam
M2
a b
c d

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 12/97
Keterangan : D- : dosis 0 ml biofertilizer ; D+ : dosis NPK 10 g; D5 : dosis 5 ml biofertilizer ;D10 : dosis 10 ml biofertilizer ; D15 : dosis 15 ml biofertilizer ; M1: media tanam
tanah; M2: media tanam tanah dan kompos.
Pada gambar 3 di atas, perlakuan dosis dan media tanam yang mempunyai
rata-rata jumlah daun cabai rawit berurutan dari yang tertinggi sampai terendah
adalah M1D-, M2D15, M1D+, M2D10, M1D15, M1D5, M2D+, M1D10, M2D-,
M2D5. Perlakuan M1D- mempunyai rata-rata jumlah daun tertinggi dibanding
dengan yang lainnya, hal ini mungkin disebabkan nutrisi yang ada dalam media M1
yaitu tanah sudah dapat mencukupi kebutuhan suplai nutrisi yang dibutuhkan untuk
proses pembentukan daun, sehingga dosis pupuk D- (tanpa diberi pupuk) lebih
optimal pembentukan jumlah daunnya dibandingkan dengan perlakuan yang lain.
Perlakuan dosis dan media tanam yang mempunyai rata-rata tinggi tanaman cabai
rawit berurutan dari yang tertinggi sampai terendah adalah M1D15, M1D+, M1D10,
M1D5, M1D-, M2D+, M2D10, M2D15, M2D5, M2D-. Perlakuan M1D15
mempunyai rata-rata tinggi tanaman paling tinggi dibanding dengan tanaman lainnya,
hal ini mungkin dikarenakan nutrisi yang dibutuhkan untuk proses pertumbuhan
tanaman sangat besar, sehingga nutrisi yang disediakan media tanam M1 masih
sangatlah kurang sehingga masih membutuhkan nutrisi tambahan dari proses
pemupukan, dan dosis yang paling optimal untuk ditambahkan ke media tanam M1
adalah D15.
Perlakuan dosis dan media tanam yang mempunyai rata-rata jumlah buah
cabai rawit berurutan dari yang tertinggi sampai terendah adalah M1D+, M2D10,
M1D10, M1D15, M2D15, M2D+, M1D-, M2D- M1D5, M2D5,. Perlakuan M1D+
menunjukan hasil rata-rata jumlah buah cabai rawit tertinggi. Antara jumlah daun dan
jumlah buah terdapat hubungan yang erat. Semakin banyak daun yang diproduksi,
secara normal hal ini akan berpengaruh secara langsung terhadap banyaknnya jumlah
produksi buah. Dengan adannya hubungan ini, maka pengamatan jumlah daun dan
jumlah buah yang mempunyai jumlah paling tinggi seharusnya terjadi pada tanaman
dengan perlakuan yang sama. Akan tetapi hal ini tidak terjadi pada penelitian ini,

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 13/97
karena pada pengamatan jumlah daun perlakuan yang mempunyai jumlah daun paling
banyak adalah M1D- (tanpa diberi pupuk), sedangkan pada pengamatan jumlah buah
perlakaun yang mempunyai jumlah buah paling banyak adalah tanaman dengan
perlakuan M1D+ (dengan pemupukan NPK), hal ini sangatlah berbeda, mungkin
dikarenakan nutrisi pada media tanam perlakuan M1D- jumlahnya masih kurang
untuk proses pembentukan buah, sehingga diperluakan nutrisi tambahan yang
didapatkan dari proses pemupukan. Hal ini didukung oleh pendapat Latifah (2008),
yang mengatakan bahwa pemberian pupuk melalui tanah dengan frekuensi yang
sangat jarang (sekaligus, dua atau tiga kali sepanjang siklus pertumbuhan)
membutuhkan jumlah pupuk yang sangat banyak karena adanya pencucian. Rosliani
dkk. (2001) dalam Latifah (2008), juga melaporkan bahwa dari pupuk N yang
diberikan kedalam tanah, hanya 30 -50 % yang diserap tanaman, sedangkan pupuk P
dan K lebih rendah lagi hanya sebesar 15 – 20 %, selebihnya menjadi residu dalam
larutan tanah dan tercuci.
Perlakuan dosis dan media tanam yang mempunyai rata-rata berat buah cabai
rawit berurutan dari yang tertinggi sampai terendah adalah M2D10, M1D15, M1D+,
M1D10, M2D+, M1D-, M2D15, M1D5, M2D- , M2D5. Secara deskriptif tanaman
yang mempunyai rata-rata berat buah yang paling tinggi adalah tanaman dengan
perlakuan M2D10. Meskipun pada pengamatan jumlah buah perlakuan M1D+
mempunyai nilai tertinggi, tetapi ini tidak bisa dijadikan patokan bahwa pada
perlakuan ini juga akan diperoleh berat buah tertinggi, jadi banyaknya jumlah buah
tidak bisa menentukan besarnya berat buah. Karena perlakuan M1D+ mempunyai
jumlah buah yang banyak, tetapi ukuran buahnya kecil-kecil, sehingga ini akan
mempengaruhi berat buahnya. Perlakuan M2D10 memiliki berat buah tertinggi
mungkin dikarenakan jumlah buah yang dimiliki memang tidak sebanyak M1D+,
tetapi ukuran buah yang dimiliki M2D10 lebih besar –besar, sehingga dapat
mempengaruhi berat buahnya.Dari hasil perhitungan RAE untuk setiap perlakuan,
diketahui bahwa D15 pada M1 dan D10 pada M2 memiliki nilai lebih besar dari
100%. Hal ini berarti D15 pada media tanam M1 (tanah) dan D10 pada media tanam

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 14/97
M2 (tanah dan kompos) adalah efektif. Akan tetapi D10 pada M2 dengan nilai RAE
207,11% lebih efektif dari pada D15 pada M1 yang mempunyai nilai 133,49%.
Kesimpulan
Pemberian berbagai dosis pupuk hayati (biofertilizer ) secara deskriptif
berpengaruh pada pertumbuhan dan produktivitas tanaman cabai rawit (Capsicum
frutescens L.). Pertumbuhan tertinggi diperoleh dari perlakuan dosis D- dan D15
(jumlah daun) dan pada perlakuan D+ (tinggi tanaman). Produktivitas tertinggi
diperoleh dari perlakuan dosis D10 (jumlah buah dan berat buah). Media tanam
secara deskriptif berpengaruh pada pertumbuhan dan produktivitas tanaman cabai
rawit (Capsicum frutescens L.). Pertumbuhan tertinggi diperoleh dari perlakuan
media tanam M1 (jumlah daun dan tinggi tanaman ). Produktivitas tertinggi diperoleh
dari perlakuan media tanam M1 (jumlah buah dan berat buah). Pemberian dosis
pupuk hayati (biofertilizer ) dan media tanam secara deskriptif berpengaruh pada
pertumbuhan dan produktivitas tanaman cabai rawit (Capsicum frutescens L.).
Pertumbuhan tertinggi diperoleh dari perlakuan dosis dan media tanam M1D-
(jumlah daun) dan pada perlakuan dosis dan media tanam M1D15 (tinggi tanaman).
Produktivitas tertinggi diperoleh dari perlakuan dosis dan media tanam M1D+
(jumlah buah ) dan pada perlakuan dosis dan media tanam M2D10 (berat buah).
Daftar Pustaka
Anonimus, 2011, Laporan Survei Lapangan Produksi dan Pembentukan Harga Komoditas
Cabai di Kabupaten Magelang dan Wonosobo,http://www.bi.go.id/NR/rdonlyres/8AC2383D-37CE, diakses pada tanggal 02-01-2012
Anonimus a, 2008, Budidaya Tanaman Kedelai/ Kc. Hijau, Padi, Jagung/ Sorgum, Kc.Tanah, Cabe/ Tomat, Sawi/ Kubis, Rimpang/ Umbi, Jeruk/ Mangga Menggunakan
Pupuk Hayati Bio P 2000 Z National Patent: Id 0 000 438 S, Penyusun PT. AlamLestari Maju Indonesia (PT. ALAMI)
Deptan, 2011, Hasil Pencarian Berdasarkan Komoditi Hortikultura,http://cybex.deptan.go.id/lokalita/budidaya-cabe-rawit-0, diakses pada tanggal 7-10-2011
Ermina, Y., 2010, Media Tanaman Hidroponik dari Arang Sekam, Balai Besar PelatihanPertanian (BBPP Lembang)
Gardner, F. P., R. B. Pearce, dan R. L. Mitchell, 1991, Fisiologi Tanaman Budidaya,Universitas Indonesia Press, Jakarta

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 15/97
Imanda N. dan Ketty S., 2012, Pengaruh Jenis Media Tanam Terhadap Pertumbuhan BibitPepaya (Carica papaya L.) Genotipe IPB 3, IPB 4, dan IPB 9, Institut Pertanian
Bogor Isroi, 2009, Pupuk Organik Granul Sebuah Petunjuk Praktis, http://Isroi.wordpress.com ,
diakses pada tanggal 12-12-2011Latifah, 2008, Metode Penapisan dan Uji Ketahanan Cabai (Capsicum annuum L.) Terhadap
Chilli Veinal Mottle Virus dan Cucumber Mosaic Virus, Thesis, Institut Pertanian
Bogor Ningrum, F. G. K., 2010, Efektivitas Air Kelapa dan Ampas Teh Terhadap Pertumbuhan
Tanaman Mahkota Dewa ( Phaleria macrocarpa) Pada Media Tanam Yang Berbeda,Skripsi Program Studi Pendidikan Biologi Fakultas Keguruan dan Ilmu PendidikanUniversitas Muhammadiyah Surakarta
Saraswati, R., R.D.M. Simanungkalit, E. Husen, D. Santoso, D. Setyorini , dan A. Rachman,2008, Baku Mutu Pupuk Hayati, Balai Penelitian Tanah, Balai Besar LitbangSumberdaya Lahan Pertanian Badan Litbang Pertanian Departemen Pertanian
Simanungkalit, R. D. M., 2001, Aplikasi Pupuk Hayati dan Pupuk Kimia: Suatu PendekatanTerpadu, Buletin AgroBio 4(2) : 56-61.
Simanungkalit, R. D. M., D. A. Suriadikarta, R. Saraswati, D. Setyorini, dan W. Hartatik,2006, Pupuk Organik dan Pupuk Hayati, Balai Besar Penelitian dan PengembanganSumberdaya Lahan Pertanian, Jawa Barat
Suwahyono, U., 2011, Petunjuk Praktis Penggunaan Pupuk Organik Secara Efektif dan
Efisien, Penebar Swadaya, JakartaSyaifudin, A., L. Mulyani, M. Ariesta, 2010, Pupuk Kosarmas Sebagai Upaya Revitalisas
Lahan Kritis Guna Meningkatkan Kualitas dan Kuantitas Hasil Pertanian, Universitas Negeri Solo

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 16/97
1
AKTIVITAS ANTIBAKTERI DAN ANTIFUNGI EKSTRAK
PETROLEUM ETER Dumortiera hirsuta
Junairiah, Hanik Faizah, dan Salamun
Prodi S-1 Biologi, Depatermen Biologi, Fakultas Sains dan Teknologi,
Universitas Airlangga, Surabaya
ABSTRAK
The aims of this study were to investigate the secondary metabolite compoundand antimicrobial activity of petroleum eter extract of liverwort Dumortierahirsuta against Escherichia coli, Staphylococcus aureus and Candida albicans.
Secondary metabolite which contained in extract was tested with phytochemicalscreening method. Antimicrobial activity were investigated by disc diffusion
method to measure the inhibition zone diameter and tube dilution method to
determine minimal inhibitory concentration (MIC) and minimal bactericidal
concentration/minimal fungicidal concentration (MBC/MFC). Diameters of the
inhibition zone were analyzed using Kruskall-Wallis Test (α=0,05) followed up
with Mann-Whitney Test (α=0,05). Data of MIC, MBC/MFC and phytochemical
screening were descriptively analyzed. The result of phytochemical screening test
showed that there was steroid in petroleum eter extract of D. hirsuta. The result
showed that the diffferent concentration of petroleum eter extract of D. hirsuta
influenced diameter of the inhibition zone growth of E. coli, S. aureus and C.albicans. The kinds of microbe did not have effect on diameter of the inhibition
zone. MIC and MBC/MFC have not been able determined in this study.
Key words : Dumortiera hirsuta, antimicrobial, petroleum eter, Escherichia coli,
Staphylococcus aureus, Candida albicans
PENDAHULUAN
Penyakit infeksi merupakan salah satu masalah dalam bidang kesehatan
yang dari waktu ke waktu terus berkembang. Penyakit infeksi banyak disebabkan
oleh mikroorganisme flora normal, sebagai contoh, beberapa bakteri penting yang
dapat menyebabkan penyakit adalah Staphylococcus aureus dan Escherichia coli.
Infeksi juga dapat disebabkan oleh mikroorganisme lain seperti Candida albicans
yang juga merupakan anggota flora normal penyebab candidiasis (Jawetz et al.,
2005). Antibiotik memberikan dasar utama untuk terapi infeksi mikroba (bakteri

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 17/97
2
dan fungi). Namun, terlalu sering menggunakan antibiotik telah menjadi faktor
utama bagi munculnya dan penyebaran beberapa kelompok mikroorganisme yang
resisten terhadap antibiotik (Harbottle et al., 2006). Dalam beberapa tahun
terakhir, permasalahan resistensi bakteri pada penggunaan antibiotika merupakan
salah satu masalah yang berkembang di seluruh dunia (Bronzwaer et al., 2002).
Oleh karena itu diperlukan zat antibakteri baru dengan mekanisme aksi yang
berbeda (Tenover, 2006). Karena alasan ini, peneliti mengalihkan perhatiannya
untuk menemukan zat antimikroba dari sumber baru yang berasal dari tumbuhan
(Singh et al., 2010).
Zhu et al . (2006) mengungkapkan bahwa tumbuhan lumut adalah salah satu
sumber antibiotik yang paling signifikan dan menjanjikan di alam. Hasil
penelitian yang dilakukan oleh Veljic et al. (2010) mengungkapkan bahwa ekstrak
metanol lumut hati Ptilidium pulcherrimum memiliki aktivitas antibakteri dan
antifungi. Bodade et al. (2008) menjelaskan bahwa diantara kelompok lumut yang
diuji, lumut hati Plagiochasma appendiculatum adalah paling aktif menghambat
bakteri, lumut hati memiliki aktivitas antibiotik lebih baik (aktivitas 88%)
daripada lumut daun (aktivitas 33%), ini memperlihatkan bahwa senyawa
antibakteri terdapat pada sebagian besar takson dari lumut hati.
Pada beberapa penelitian sebelumnya telah dilaporkan bahwa lumut hati
Dumortiera hirsuta memiliki aktivitas antibakteri dan antifungi (Madsen dan
Pates, 1952 dalam Glime, 2007; Alam et al ., 2011). Escherichia coli,
Staphylococcus aureus dan Candida albicans merupakan flora normal manusia
akan tetapi akan berubah menjadi mikroba patogen dalam kondisi tertentu. Dalam
penelitian ini, untuk mengetahui aktivitas antimikroba ekstrak petroleum eter
lumut hati Dumortiera hirsuta terhadap penghambatan pertumbuhan bakteri gram
positif dan bakteri gram negatif serta penghambatan pertumbuhan fungi patogen
digunakan Escherichia coli, Staphylococcus aureus dan Candida albicans sebagai
mikroba uji. Petroleum eter digunakan untuk mengikat metabolit sekunder yang
terkandung dalam Dumortiera hirsuta yang diduga bersifat toksik bagi mikroba
uji. Penggunaan berbagai variasi konsentrasi ekstrak lumut tersebut diharapkan
dapat menunjukkan aktivitas antimikroba yang berbeda sehingga dapat diketahui

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 18/97
3
nilai MIC ( Minimal Inhibitory Concentration) dan MBC/MFC ( Minimal
Bactericidal Concentration/ Minimal Fungicidal Concentration)
BAHAN DAN METODE
Tempat dan Waktu Penelitian
Penelitian dilaksanakan selama 6 bulan pada bulan Pebruari 2012 sampai
dengan Juli 2012 di Laboratorium Biologi Dasar dan Laboratorium Mikrobiologi,
Departemen Biologi, Fakultas Sains dan Teknologi, Universitas Airlangga,
Surabaya.
Bahan Tumbuhan
Tumbuhan lumut Dumortiera hirsuta yang diperoleh dari Taman Hutan
Raya Raden Suryo, Cangar-Batu. Dumortiera hirsuta diidentifikasi dengan buku
Guide to the Liverworts of North Carolina (Hicks, 1992). Lumut diambil dari
habitatnya dan disimpan dalam kantong koleksi lumut. Sampel yang diambil
merupakan fase gametofit lengkap dan telah dewasa.
Mikroba Uji
Mikroba uji yang digunakan adalah Escherichia coli ATCC 25922,
Staphylococcus aureus ATCC 25923 dan Candida albicans ATCC 10231, yang
merupakan koleksi Laboratorium Mikrobiologi, Departemen Biologi, Fakultas
Sains dan Teknologi, Universitas Airlangga, Surabaya.
Ekstraksi Lumut D. hirsuta
Hasil koleksi tumbuhan lumut D. hirsuta yang diperoleh diekstraksi dengan
menggunakan pelarut petroleum eter. Sebanyak 10 gram lumut D. hirsuta kering
direndam secara keseluruhan dengan 300 mL petroleum eter. Larutan petroleum
eter dan tumbuhan lumut tersebut dimaserasi selama 3 hari. Ekstrak cair dari hasil
maserasi disaring menggunakan kertas saring kemudian disaring kembali dengan
menggunakan corong buchner. Filtrat yang dihasilkan kemudian ditampung dalam
gelas beaker dan ditutup dengan aluminium foil yang telah diberi lubang-lubang

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 19/97
4
kecil untuk mengguapkan pelarut petroleum eter. Kemudian 80 mg dari ekstrak
kering dilarutkan dalam 1 mL DMSO 20%.
Skrining Fitokimia
Ekstrak petroeum eter D. hirsuta diidentifikasi komponen fitokimianya
terhadap golongan senyawa triterpenoid dan steroid. Sampel dimasukkan ke
dalam spot plate dengan menggunakan tusuk gigi, kemudian ditambahkan larutan
Liebermann Burchard. Liebermann Burchard terdiri atas 3 tetes asam asetat
anhidrat dan 1 tetes asam sulfat pekat. Apabila terbentuk warna hijau maka
menunjukkan adanya senyawa steroid dan warna merah atau ungu menunjukkan
adanya triterpenoid (Harborne, 1987).
Penentuan Aktivitas Antimikroba
Metode cakram kertas
Metode cakram kertas menggunakan media Mueller-Hinton Agar (MHA)
steril sebagai media pertumbuhan dalam uji antimikroba. Metode ini meletakkan
kertas cakram (diameter 6 mm) yang ditetesi dengan 25 µL ekstrak peroleum eter D. hirsuta pada permukaan medium MHA (volume medium ± 15 mL) dalam
cawan petri yang sudah diinokulasi E. coli, S. aureus dan C. albicans secara
aseptik. Suspensi mikroba uji kurang lebih 1 mL dengan OD 0,1 pada λ 625 nm
untuk bakteri dan λ 600 nm untuk fungi. Kemudian medium yang telah diberi
perlakuan, yaitu dengan pemberian ekstrak petroleum eter D. hirsuta dengan
konsentrasi yang berbeda-beda (0 ppm, 2.500 ppm, 5.000 ppm, 10.000 ppm,
20.000 ppm, 40.000 ppm, 60.000 ppm, dan 80.000 ppm) diinkubasi selama 24
jam untuk bakteri dan 48 jam untuk fungi. Menurut Bailey dan Scott (1994)
terbentuknya daerah penghambatan (halo) di sekitar cakram uji menunjukkan
adanya aktivitas antimikroba. Diameter daerah penghambatan pertumbuhan
mikroba diukur dengan menggunakan jangka sorong.
Metode pengenceran dalam tabung
Metode pengenceren dalam tabung digunakan untuk memperoleh nilai MIC
dan MBC/MFC. Konsentrasi ekstrak petroleum eter D. hirsuta untuk metode

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 20/97
5
pengenceran ini adalah 2.500, 2.000, 1.500, 1.000, 750, 500, 250, dan 0 ppm.
Metode ini diawali dengan membuat suspensi mikroba uji pada media Muller-
Hinton Broth (MHB) dengan mengatur kekeruhan mikroba pada OD yang sesuai
dengan standar Mc. Farland 0,5 λ 625 nm=0,1 untuk bakteri dan 0,5 λ 600 nm=0,1
untuk fungi. Kemudian 1 mL larutan ekstrak untuk masing-masing konsentrasi
yang telah ditentukan dimasukkan kedalam tabung reaksi dan ditumbuhkan
dengan 1 mL mikroba uji di dalamnya. Kultur dihomogenkan dan diinkubasi
selama 24 jam untuk bakteri dan 48 jam untuk fungi. Aktivitas antimikroba dalam
kultur dapat diketahui jika terjadi penurunan densitas dalam kultur yang sudah
ditambah dengan ekstrak lumut. Dari kultur positif tersebut diambil sebanyak 0,1
mL untuk ditumbuhkan dalam media MHA dan diinkubasi selama 24 jam untuk
bakteri dan 48 jam untuk fungi. Setelah inkubasi, ketika terjadi pertumbuhan pada
media MHA, maka pada konsentrasi tersebut merupakan nilai MIC dan nilai
MBC/MFC dapat ditentukan jika dalam media tersebut tidak ditumbuhi oleh
mikroba, yang menunjukkan bahwa pada konsentrasi tersebut zat antimikroba
ekstrak lumut dapat membunuh mikroba uji (Bailey dan Scott, 1994).
Analisis Data
Data yang diperoleh, yaitu berupa diameter zona hambat pertumbuhan
bakteri E. coli, S. aureus, dan fungi C. albicans pada berbagai konsentrasi ekstrak
petroleum eter D. hirsuta. Data tersebut dianalisis secara statistik dengan
menggunakan program SPSS versi 16. Data diuji dengan Kruskall-Wallis Test
Kemudian dilakukan uji lanjutan menggunakan uji Mann-Whitney. Sedangkan
data jumlah sel mikroba uji, nilai MIC, nilai MBC/MFC dan skrining fitokimia
dianalisis secara deskriptif.
HASIL DAN PEMBAHASAN
Ekstraksi Lumut D. hirsuta
Hasil ekstraksi 10 gram serbuk lumut hati D.hirsuta dengan 1300 ml
petroleum eter diperoleh ekstrak padat berwarna kuning pekat sebanyak 0,22
gram. Ekstraksi ini dilakukan untuk mengambil komponen non polar dari

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 21/97
6
sampel lumut D. hirsuta. Ekstrak ini selanjutnya diidentifikasi komponen
fitokimianya dan dilakukan uji aktivitas antimikroba.
Skrining Fitokimia
Uji fitokimia kandungan metabolit sekunder ekstrak petroleum eter D. hirsuta
dilakukan terhadap uji triterpenoid dan steroid. Hasil pengujian skrining fitokimia
menunjukkan ekstrak mengandung senyawa steroid yang ditunjukkan dengan
terbentuknya warna hijau. Steroid bersifat nonpolar karena tersusun dari isopren-
isopren dari rantai panjang hidrokarbon (Robinson, 1995), sehingga steroid dapat
larut dalam petroleum eter.
Penentuan Aktivitas Antimikroba
Metode cakram kertas
Uji aktivitas antimikroba ekstrak petroleum eter D. hirsuta terhadap E. coli,
S. aureus, dan C. albicans dengan metode cakram kertas adalah positif yaitu
ditunjukkan dengan terbentuknya zona hambat (halo) di sekitar kertas cakram.
Dari hasil uji cakram kertas terlihat jelas ekstrak petroleum eter D. hirsuta
mempunyai aktivitas penghambatan terhadap mikroba uji. Diameter zona hambat
pertumbuhan bakteri E. coli, S. aureus, dan fungi C. albicans pada berbagai
konsentrasi ekstrak petroleum eter D. hirsuta disajikan pada gambar 1 berikut:
Gambar 1. Diagram batang diameter zona hambat pertumbuhan E. coli, S. aureus,
dan C. albicans pada berbagai konsentrasi ekstrak petroleum eter D.
hirsuta
0
5
10
15
20
25
D i a m e t e r Z o n a H a m b a t
( m
m )
Konsentrasi Ekstrak Petroleum Eter Dumortiera
hirsuta (ppm)
E.coliS.aureus
C.albicans

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 22/97
7
Pada gambar 1 terlihat bahwa konsentrasi ekstrak petroleum eter D. hirsuta
berpengaruh terhadap diameter zona hambat ketiga mikroba uji. Perlakuan
konsentrasi ekstrak petroleum eter D. hirsuta terhadap E. coli, S. aureus, dan C.
albicans mulai terlihat perbedaan yang nyata dengan kontrol pada konsentrasi
2.500 ppm, rerata diameter zona hambat E. coli sebesar (7,77 ± 0,17) mm, S.
aureus sebesar (8,64 ± 0,23) mm, dan C. albicans sebesar (11,16 ± 0,31) mm.
Rata-rata diameter zona hambat terbesar untuk perlakuan konsentrasi ekstrak
petroleum eter D. hirsuta terhadap E. coli dan S. aureus, yaitu pada konsentrasi
10.000 ppm, berturut-turut sebesar (9,22 ± 0,15) mm dan (9,68 ± 0,65) mm.
Sedangkan rata-rata diameter zona hambat terbesar untuk perlakuan konsentrasi
ekstrak petroleum eter D. hirsuta terhadap C. albicans sebesar (23,00 ± 2,52) mm
pada konsentrasi 60.000 ppm. Menurut Arora dan Bhardwaj (1997), aktivitas
antimikroba dikategorikan tingkat sensitifitas tinggi apabila diameter zona hambat
mencapai > 12 mm. Kategori tingkat sensitifitas sedang diberikan apabila ekstrak
mampu memberikan diameter zona hambat sekitar 9-12 mm. Kategori tingkat
sensitifitas rendah, apabila diameter berkisar antara 6-9 mm dan resisten apabila
<6 mm.
Pada penelitian ini, konsentrasi ekstrak petroleum eter D. hirsuta
berpengaruh terhadap diameter zona hambat akan tetapi jenis mikroba tidak
berpengaruh terhadap besarnya diameter zona hambat, hal ini dapat dilihat pada
gambar 1. Pada beberapa penelitian, telah dilaporkan bahwa bakteri gram positif
ditemukan lebih sensitif daripada bakteri gram negatif. Hal ini dapat terjadi karena
bakteri gram negatif memiliki membran luar sementara bakteri gram positif hanya
memiliki lapisan peptidoglikan. Membran luar ini bertanggung jawab untuk
melindungi bakteri dari antibiotik, deterjen, dan enzim yang biasanya merusak
membran dalam atau peptidoglikan (Lehninger et al ., 2005). Namun, hasil yang
diperoleh dalam penelitian ini adalah berbeda. Sensitivitas bakteri gram positif
atau bakteri gram negatif terhadap ekstrak petroleum eter D. hirsuta tidak
berbeda nyata. Hal ini menyimpulkan bahwa jenis mikroba uji, yaitu E. coli, S.
aureus, dan C. albicans tidak memiliki pengaruh terhadap besarnya diameter
zona hambat.
7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 23/97
8
Metode pengenceran dalam tabung
Metode pengenceran dalam tabung digunakan untuk mengetahui kadar
hambat terkecil (MIC) dan kadar bunuh (MBC/MFC) terkecil dari suatu bahan
antimikroba tertentu (Bailey dan Scott, 1994; Wistreich, 2003). Nilai MIC
ditentukan secara kasat yang didasarkan pada kekeruhan kultur mikroba yang
telah diinkubasi.
Pada penelitian ini, sulit dilakukan pengamatan terhadap tingkat kekeruhan
kultur pada berbagai konsentrasi, sehingga penentuan nilai MIC tidak didasarkan
pada kekeruhan kultur mikroba uji. Penentuan nilai MIC dilakukan dengan
pengamatan secara kuantitatif dengan cara menghitung jumlah koloni (nilai TPC
atau Total Plate Count ) dari masing-masing konsentrasi melalui metode
pencawanan. Hasil perhitungan nilai TPC tersebut dapat dilihat pada tabel 1
berikut:
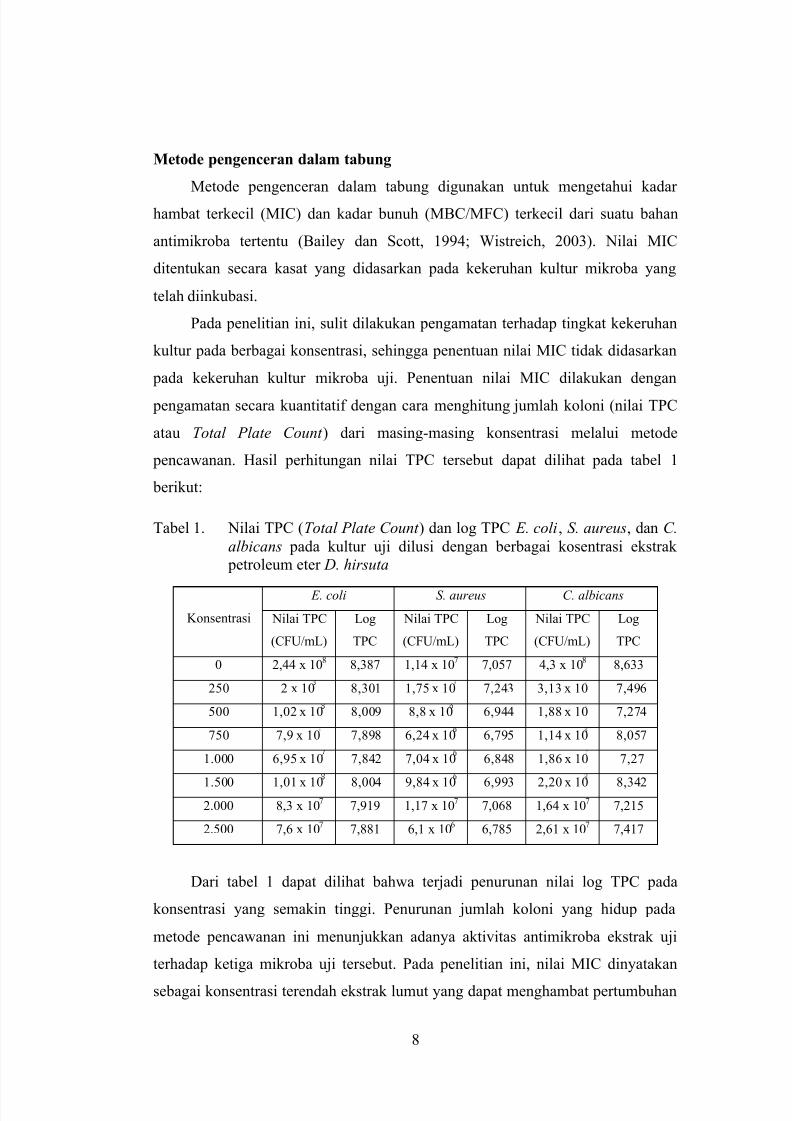
Tabel 1. Nilai TPC (Total Plate Count ) dan log TPC E. coli, S. aureus, dan C.
albicans pada kultur uji dilusi dengan berbagai kosentrasi ekstrak
petroleum eter D. hirsuta
Konsentrasi
E. coli S. aureus C. albicans
Nilai TPC
(CFU/mL)
Log
TPC
Nilai TPC
(CFU/mL)
Log
TPC
Nilai TPC
(CFU/mL)
Log
TPC
0 2,44 x 108 8,387 1,14 x 10
7 7,057 4,3 x 10
8 8,633
250 2 x 10 8,301 1,75 x 10 7,243 3,13 x 10 7,496
500 1,02 x 10 8,009 8,8 x 10 6,944 1,88 x 10 7,274
750 7,9 x 10 7,898 6,24 x 10 6,795 1,14 x 10 8,057
1.000 6,95 x 10 7,842 7,04 x 10 6,848 1,86 x 10 7,27
1.500 1,01 x 10 8,004 9,84 x 10 6,993 2,20 x 10 8,3422.000 8,3 x 10
7 7,919 1,17 x 10
7 7,068 1,64 x 10
7 7,215
2.500 7,6 x 107 7,881 6,1 x 106 6,785 2,61 x 107 7,417
Dari tabel 1 dapat dilihat bahwa terjadi penurunan nilai log TPC pada
konsentrasi yang semakin tinggi. Penurunan jumlah koloni yang hidup pada
metode pencawanan ini menunjukkan adanya aktivitas antimikroba ekstrak uji
terhadap ketiga mikroba uji tersebut. Pada penelitian ini, nilai MIC dinyatakan
sebagai konsentrasi terendah ekstrak lumut yang dapat menghambat pertumbuhan

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 24/97
9
mikroba sebanyak 90% koloni awal, sedangkan nilai MBC ditentukan dari cawan
dengan konsentrasi ekstrak terkecil yang tidak dapat ditumbuhi lagi oleh bakteri
atau mampu mereduksi 99,9% dari populasi bakteri awal selama inkubasi 24 jam
(Koketsu et al ., 1996). Dari definisi di atas, pada penelitian ini belum bisa
ditemukan nilai MIC karena penurunan jumlah koloni pada berbagai konsentrasi
ekstrak belum mencapai 90% baik pada E. coli, S. aureus maupun C. albicans.
Nilai MBC/MFC juga belum bisa ditemukan karena belum ada konsentrasi
ekstrak yang dapat mereduksi 99,9% koloni dari jumlah populasi awal baik pada
E. coli, S. aureus maupun C. albicans.
Dari hasil uji aktivitas antimikroba pada metode cakram kertas dan metode
pengenceran dalam tabung menunjukkan bahwa pada ekstrak petroleum eter D.
hirsuta terdapat zat aktif yang bersifat sebagai antimikroba. Pada beberapa
penelitian sebelumnya telah dilaporkan bahwa lumut hati D. hirsuta memiliki
aktivitas antibakteri dan antifungi (Madsen dan Pates, 1952 dalam Glime, 2007).
Ekstrak air lumut hati D. hirsuta ditemukan dapat menghambat sejumlah fungi
fitopatogen dengan mekanisme yang berbeda seperti penghambatan germinasi
spora, sintesis dinding sel sehingga dinding sel menjadi lembek atau tidak kaku,
sitoplasma hilang dari sel dan sitoplasma berbentuk granul atau butiran dan
lainnya (Alam et al ., 2011). Pada penelitian ini, ekstrak petroleum eter D. hirsuta
positif mengandung senyawa steroid. Senyawa ini diduga merupakan senyawa
aktif yang berperan sebagai antimikroba yang menghambat pertumbuhan E. coli,
S. aureus dan C. albicans. Hal ini didukung oleh penelitian sebelumnya bahwa
steroid dapat menghambat pertumbuhan mikroba. Menurut Zhu et al . (2000) dan
Varricchio et.al (1967) steroid dapat menghambat pertumbuhan bakteri gram
positif. Subhisha dan Subramoniam (2005) melaporkan bahwa ekstrak lumut hati
Pallavicinia lyellii mengandung senyawa steroid yang aktif menghambat
germinasi spora pertumbuhan C. albicans, mekanisme penghambatannya diduga
berkaitan dengan sistem intraseluler.

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 25/97
10
KESIMPULAN
Uji skrining fitokimia menunjukkan adanya senyawa steroid di dalam
ekstrak petroleum eter D. hirsuta. Pemberian ekstrak petroleum eter D. hirsuta
pada konsentrasi berbeda berpengaruh terhadap diameter zona hambat
pertumbuhan E. coli, S. aureus dan C. albicans. Jenis mikroba yang berbeda tidak
memiliki pengaruh terhadap besarnya zona hambat. Pada penelitian ini belum
dapat ditemukan nilai MIC (Minimum Inhibitor Concentration) dan MBC/MFC
(Minimum Bactericidal Concentration/Minimal Fungicidal Concentration) dari
ekstrak petroleum eter D. hirsuta terhadap E. coli, S. aureus dan C. albicans.
DAFTAR PUSTAKA
[1]
Alam, Afroz, T., Abhishek, V., Sharad, B.,K. Kumar,dan S.,Vinay, 2011,
In vitro antifungal efficacies of aqueous extract of D. hirsuta (Swaegr.)
Nees against sporulation and growth of postharvest phytopathogenic
fungi, Arc. Bryology, 103, 1-9
[2] Arora, D.S. dan Bhardwaj, 1997, Antibacterial Activity of Some
Medicinal Plants, Geo. Bios., 24, 127-131
[3]
Bailey, W.R. dan Scott, E.G, 1994, Diagnostic Microbiology, 4 edition,Saint Louis, The CV Mosby Company
[4] Bodade R.G., Borkar P.S., Md Saiful Arfeen, dan Khobragade C. N.,
2008, In vitro Screening of Bryophytes for Antimicrobial Activity, Journal of Medicinal Plans, 7 (4), 23-28
[5] Bronzwaer, SL., Cars, O., Buchhols, U., Molstad, S., dan Goettsch, W.,2002, A European Study on The Relationship between Antimicrobial Use
and Antimicrobial Resistance, Emerging Infectious Disease, 8, 278-282
[6] Glime, J.M, 2007, Bryophyte Ecology, http://www.bryoecol.mtu.edu
( diakses 6 Januari 2012)
[7] Harborne, JB, 1987, Metode Fitokimia, Edisi kedua, Bandung, Penerbit
ITB
[8] Harbottle H., Thakur, S., Zhao, S., dan White, D.G., 2006, Genetics of
Antimicrobial Resistance, Anim.Biotechnol , 17, 111-124[9]
Hicks, Marie L., 1992, Guide to the Liverworts of North Carolin, United
States of America, Duke University Press[10]
Jawetz E, Melnick GE, dan Adelberg CA, 2005, Mikrobiologi
kedokteran, Edisi II , Diterjemahkan oleh dr. Nani Widorini, Jakarta,
Salemba Medika
[11]
Koketsu, Mamoru, M., Kim, dan Yamamoto T., 1996, Antifungal activity
against foodborne fungi of Aspidistra elatior Blume, J of Agric Food
Chemistry 44, 301 – 303

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 26/97
11
[12] Lehninger,A., D. Nelson, M. Cox, dan Lehninger, 2005, Principles of
Biochemistry 4th Edition, New ork, W.H. Freeman New York
[13]
Robinson, T., 1995, Kandungan Senyawa Organik Tumbuhan Tinggi, Diterjemahkan oleh Prof. Dr. Kokasih Padmawinata, Bandung, ITB
[14] Singh Kaveri, Vandana Tiwar, dan Rajneesh Prajapat, 2010, Study of
Antimicrobial Activity of Medicinal Plants Against Various MultipleDrug Resistance Pathogens And Their Molecular Characterization And
it’s Bioinformatics Analysis Of Antibiotic Gene From Genomic Database
With Degenerate Primer Prediction, International Journal of Biological
Technology, 1(2), 15-19
[15] Subhisha, S. dan Subramoniam, A., 2005, Antifungal activities of a
steroid from Pallavicinia lyellii, a liverwort. Indian Journal of
Pharmacology, 37, 304-8.
[16]
Tenover, Fred C., 2006, Mechanisms of Antimicrobial Resistance inBacteria, The American Journal of Medicine, 119 (6A), S3 – S10
[17] Varricchio, F., Norman, J.D., dan Aundre, S., 1967, Effect of azasteroidson Gram-positive bacteria, Journal of Bacteriology, 93(2), 627-635
[18] Veljic, M., Ana Ciric, Marina Socovic, P. Janackovic, dan P. D. Marin,2010, Antibacterial and Antifungal Activity of The Liverwort ( Ptilidium
pulcherrimum) methanol extract. Arch. Biol. Sci., Belgrade, Serbia, 62
(2), 381-395
[19] Wistreich, G.A, 2003, Microbiology Laboratory Fundamentals and
Application, Los angeles, Pearson Education Inc.
[20]
Zhu Y., Zhu, Q.X., dan Jia, Z.J., 2000, Epoxide sesquiterpenes and
steroids from cremanthodium discoideum, Australian Journal of
Chemistry, 53(10), 831-834[21]
Zhu, R.L., Wang D., Xu L., Shi R. P, Wang J., dan Zheng M., 2006,
Antibacterial activity in extracts of some bryophytes from China andMongolia, Journal of Botanical Laboratory 100, 603-615

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 27/97
KEANEKARAGAMAN DAN POLA DISTRIBUSI LONGITUDINAL
SPESIES KERANG AIR TAWAR CORBICULIDAE DI SUNGAI BRANTAS
PERIODE JANUARI –
FEBRUARI 2012
Ichsan Wardani, Drs. Moch. Affandi, M.Si., dan Dr. Bambang Irawan, M.Sc.
Program Studi di S1 Biologi, Departemen Biologi, Fakultas Sains dan Teknologi,Universitas Airlangga, Surabaya.
Abstract
This study aims to reveal the diversity, abundance and dominance, as well as
longitudinal distribution patterns of species of freshwater mussels Corbiculidae in
Brantas river period of January-February 2012, and to obtain comparative results on
the same topic with a sampling period of April-May 2011. Samples of freshwatermussels Corbiculidae were taken at 15 stations in Brantas river using Ponar dredge ,
the two parts of the river and the middle section of the river, each as much as 5
Ponar dredge . Mussels samples are identified, characterized and analyzed to obtain
data on the identity of species, diversity, abundance, and distribution pattern of
longitudinal and comparative results on the same topic with the results of the study
period from April to May 2011. Species of freshwater mussels Corbiculidae obtained
in January-February 2012 there are two species, namely the abundance of Corbicula
lacunae range 3-38 individu/m2 , with total dominance index 78.13% and C. javanica
with the abundance range 3-18 individu/m2 , with total dominance index 21.88%.
Corbiculidae mussels species obtained in the April-May 2011 there was only one
species, namely C. lacunae in the abundance range 5-34 individu/m2. The presence
of species of freshwater mussels Corbiculidae well in the January-February 2012 and
April-May 2011 were both present only in the lower reaches of the river Brantas,
which is from Driyorejo until Kesamben in the January-February 2012 and from
Wringinanom until Ploso on the period of April to May 2011. It was concluded that
C. lacunae is the dominant species in both study periods, but the number of species
abundance in the January-February 2012 more. Longitudinal distribution pattern is
not continuous at any good research station in the January-February 2012 and April-
May 2011.
Key words : Freshwater mussels Corbiculidae, C. lacunae, C. javanica,
longitudinal distribution patterns, Brantas river.

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 28/97
Pendahuluan
Kerang air tawar memiliki arti penting dalam keseimbangan ekosistem di
lingkungannya, yaitu sebagai konsumen yang mengkonsumsi organisme-organisme berukuran lebih kecil dan komponen tersuspensi dalam air (filter feeder) dan juga
sebagai bioindikator (Grabarkiewicz dan Davis, 2008). Keberadaan kerang air tawar
saat ini mengalami penurunan. Master et al . (2000) dalam Grabarkiewicz dan Davis(2008) mengatakan bahwa saat ini 37 spesies kerang air tawar diduga mengalami
kepunahan. Hal tersebut dijelaskan Strayer et al . (2004) dalam Grabarkiewicz dan
Davis (2008) bahwa penurunan tajam jumlah spesies kerang air tawar disebabkan
oleh kerusakan habitat, penurunan kualitas air, introduksi spesies eksotis, dan perubahan hidrologi. Penelitian yang mengungkap keberadaan kerang air tawar
Corbiculidae di aliran sungai Brantas beberapa sudah pernah dilakukan. Affandi
(1990) dan Hidayati (1995) telah mendapati kerang air tawar Corbicula javanica dan
Corbicula lacunae di sepanjang sungai Kali Surabaya dan kanal Kali Wonokromoyang merupakan bagian hilir dari sungai Brantas. Penelitian lebih lanjut dilakukan
oleh Citriana (2002) yang juga mengambil sampel di sungai Kali Surabaya dan kanalkali Wonokromo dan hanya mendapatkan satu spesies kerang air tawar Corbiculidae,
yaitu Corbicula lacunae. Jutting (1953) menyebutkan bahwa di pulau Jawa termasuk
di sungai Brantas dijumpai tiga spesies kerang air tawar Corbiculidae dari genus
Corbicula, yaitu C. javanica, C. rivalis, dan C. pulchella. Penelitian ini merupakan bagian dari penelitian satu tahun untuk mendapatkan data terkini mengenai
keanekaragaman dan kelimpahan spesies kerang air tawar Corbiculidae yang
ditemukan di aliran sungai Brantas. Penelitian ini dirancang secara periodik pada periode waktu berbeda dengan kondisi lingkungan sungai yang bebeda pula. Kondisi
lingkungan sungai Brantas berfluktuasi secara periodik antara musim kemarau dan
musim penghujan, tingginya debit air dan kecepatan arus dapat menjadi kendala
dalam pengambilan sampel kerang air tawar Corbiculidae dan mempengaruhi tingkatakurasi hasil sampling, atau dengan kata lain bahwa akurasi hasil sampling kerang air
tawar Corbiculidae sangat dipengaruhi oleh kondisi lingkungan sungai pada waktu
pengambilan sampel dilakukan. Budiarto (2011) dan Ramadani (2011) telahmengawali penelitian ini, yaitu pada bulan April hingga Mei 2011. Dalam penelitian
terdahulu replikasi pengambilan sampel hanya sebanyak tiga kali dan pada penelitian
ini akan dilakukan replikasi pengambilan sampel sebanyak lima kali. Hal ini
dilakukan agar mendapat hasil yang lebih mewakili dan diharapkan dapat melengkapidata dari penelitian terdahulu. Penelitian ini diarahkan untuk mengungkap
keanekaragaman dan kelimpahan spesies kerang air tawar Corbiculidae yang
ditemukan di aliran sungai Brantas.
Bahan dan Metode Penelitian
Bahan yang digunakan adalah sampel kerang air tawar Corbiculidae dan
larutan fiksatif berupa larutan formalin 5 – 6%. Pengambilan sampel kerang air tawarCorbiculidae dilakukan pada 15 stasiun di sungai Brantas (Gambar 1). Penentuan

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 29/97
lokasi sampling pada sungai Brantas didasarkan pada penelitian terdahulu, yaitu pada
penelitian Budiarto (2011) atau Ramadani (2011). Hal ini dilakukan agar mendapat
data yang dapat dibandingkan karena penelitian ini merupakan penelitian lanjutan.Setiap stasiun penelitian dibagi menjadi tiga plot, yaitu dua plot pada bagian
tepi sungai dan satu pada bagian tengah sungai. Dalam tiap plot kemudian diambil
sampel sebanyak lima dregde dengan menggunakan Ponar dregde. Sampel yangdidapat kemudian disortir, yaitu dipisahkan dan diseleksi dari hewan lain atau
substrat secara selektif dan hati-hati. Selanjutnya sampel kerang air tawar
Corbiculidae dimasukkan dalam wadah atau botol kolektor dan diberi penambahan
larutan formalin 5 – 6% serta diberi label identitas sampel. Sampel air juga diambiluntuk keperluan analisis kandungan oksigen terlarut. Faktor fisik-kimia yang diamati
meliputi temperatur air, tingkat keasaman (pH), kecepatan arus, kadar oksigen
terlarut (DO), dan kedalaman.
Gambar 1. Peta Lokasi Pengambilan Sampel Kerang Air Tawar Corbiculidae
(Ramadani, 2011)
Hasil dan Pembahasan
Hasil pengambilan sampel di 15 stasiun penelitian diperoleh dua spesies kerang air
tawar Corbiculidae, yaitu Corbicula lacunae dan Corbicula javanica. SpesiesCorbicula lacunae didapati pada empat stasiun dengan urutan besar kelimpahan
berturut-turut dari yang terendah adalah stasiun 6 (3 individu/m2), 4 (9 individu/m
2),
7 (24 individu/m2), dan 9 (38 individu/m
2). Corbicula javanica hanya didapati pada
7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 30/97
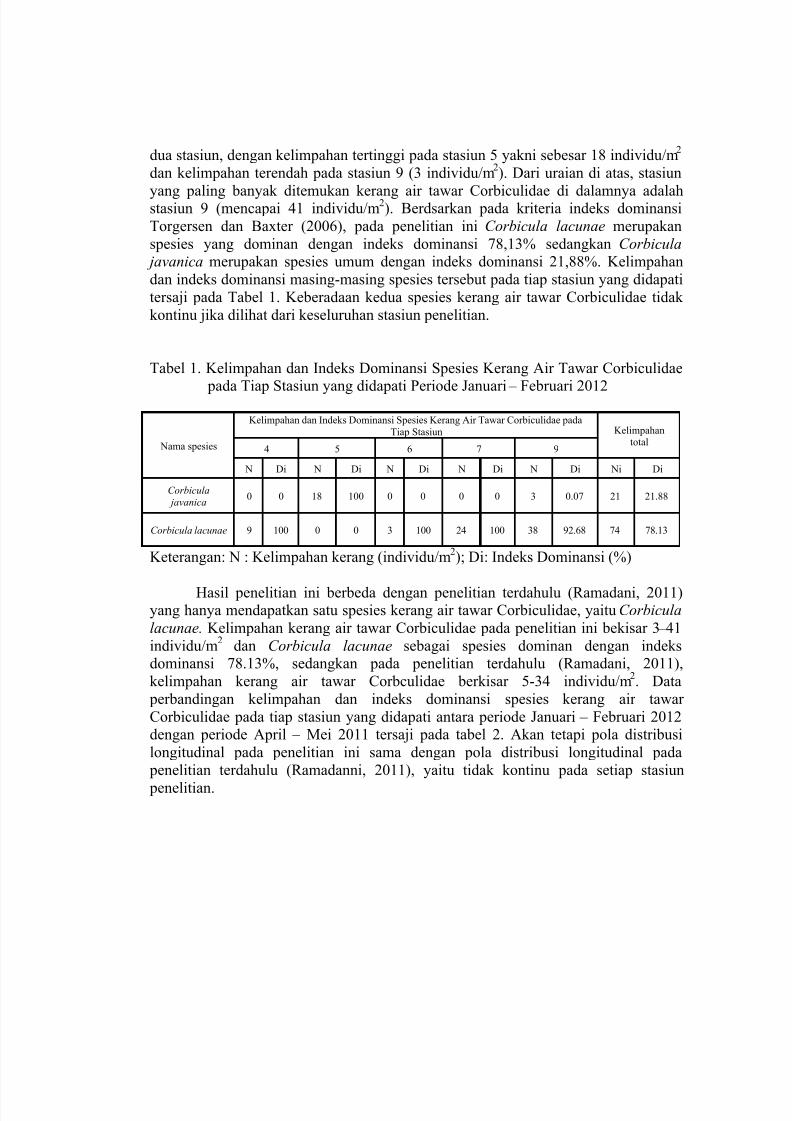
dua stasiun, dengan kelimpahan tertinggi pada stasiun 5 yakni sebesar 18 individu/m2
dan kelimpahan terendah pada stasiun 9 (3 individu/m2). Dari uraian di atas, stasiun
yang paling banyak ditemukan kerang air tawar Corbiculidae di dalamnya adalahstasiun 9 (mencapai 41 individu/m2). Berdsarkan pada kriteria indeks dominansi
Torgersen dan Baxter (2006), pada penelitian ini Corbicula lacunae merupakan
spesies yang dominan dengan indeks dominansi 78,13% sedangkan Corbicula javanica merupakan spesies umum dengan indeks dominansi 21,88%. Kelimpahan
dan indeks dominansi masing-masing spesies tersebut pada tiap stasiun yang didapati
tersaji pada Tabel 1. Keberadaan kedua spesies kerang air tawar Corbiculidae tidak
kontinu jika dilihat dari keseluruhan stasiun penelitian.
Tabel 1. Kelimpahan dan Indeks Dominansi Spesies Kerang Air Tawar Corbiculidae
pada Tiap Stasiun yang didapati Periode Januari –
Februari 2012
Nama spesies
Kelimpahan dan Indeks Dominansi Spesies Kerang Air Tawar Corbiculidae pada
Tiap Stasiun Kelimpahantotal
4 5 6 7 9
N Di N Di N Di N Di N Di Ni Di
Corbicula
javanica0 0 18 100 0 0 0 0 3 0.07 21 21.88
Corbicula lacunae 9 100 0 0 3 100 24 100 38 92.68 74 78.13
Keterangan: N : Kelimpahan kerang (individu/m2); Di: Indeks Dominansi (%)
Hasil penelitian ini berbeda dengan penelitian terdahulu (Ramadani, 2011)yang hanya mendapatkan satu spesies kerang air tawar Corbiculidae, yaitu Corbicula
lacunae. Kelimpahan kerang air tawar Corbiculidae pada penelitian ini bekisar 3 – 41
individu/m2
dan Corbicula lacunae sebagai spesies dominan dengan indeksdominansi 78.13%, sedangkan pada penelitian terdahulu (Ramadani, 2011),
kelimpahan kerang air tawar Corbculidae berkisar 5-34 individu/m2. Data
perbandingan kelimpahan dan indeks dominansi spesies kerang air tawar
Corbiculidae pada tiap stasiun yang didapati antara periode Januari – Februari 2012dengan periode April – Mei 2011 tersaji pada tabel 2. Akan tetapi pola distribusi
longitudinal pada penelitian ini sama dengan pola distribusi longitudinal pada
penelitian terdahulu (Ramadanni, 2011), yaitu tidak kontinu pada setiap stasiun
penelitian.

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 31/97
Tabel 2. Perbandingan Kelimpahan Dan Indeks Dominansi Spesies Kerang Air
Tawar Corbiculidae Pada Tiap Stasiun yang didapati antara Periode
Januari –
Februari 2012 Dengan Periode April –
Mei 2011
Keterangan: A : Periode Januari – Februari 2012; B : Periode April – Mei 2011
N : Kelimpahan kerang (individu/m2); Di: Indeks Dominansi (%)
Adanya perbedaan temuan spesies ini kemungkinan disebabkan oleh beberapa
hal, antara lain kondisi lingkungan sungai yang berbeda antara penelitian ini dengan
penelitian terdahulu. Perbedeaan kondisi lingkungan sungai tersebut meliputitemperatur air, tingkat keasaman (pH), kadar oksigen terlarut (DO), kecepatan arus,
dan kedalaman. Temperatur air dari 15 stasiun penelitian di perairan sungai Brantas
pada penelitian ini bervariasi bekisar antara 28 – 32⁰C, sedangkan temperatur air
pada penelitian terdahulu bervariasi bekisar antara 27 – 31⁰C. Hasil pengukuran
parameter pH (tingkat keasaman air) di seluruh stasiun penelitian ini juga bervariasi berkisar antara 5,6 – 6,3, yang menunjukkan bahwa air di seluruh stasiun penelitian di
sungai Brantas bersifat asam, sedangkan hasil pengukuran parameter pH pada penelitian terdahulu menunjukkan tingkat yang seragam yaitu bernilai 7, yang
menunjukkan bahwa air di seluruh stasiun penelitian di sungai Brantas pada penelitian terdahulu bersifat netral. Hasil pengukuran kadar oksigen terlarut dalam air
(DO = Dissolved oxygen) dari seluruh stasiun penelitian di perairan sungai Brantas
pada penelitian ini menunjukkan nilai yang beragam yaitu bekisar 5,88 – 10,61mgO2/liter, lain halnya dengan penelitian terdahulu yang memperoleh hasil
pengukuran kadar oksigen terlarut dalam air (DO = Dissolved oxygen) bekisar 4 – 8,3
mgO2/liter. Kecepatan arus di sungai Brantas pada penelitian ini berkisar antara
0,253 – 2,241 m/s dan kedalamannya berkisar 2 – 7 m, sedangkan kecepatan arus pada penelitian terdahulu berkisar 0.06 – 1,1 m/s dan kedalamannya berkisar 1,7 – 5,2 m.
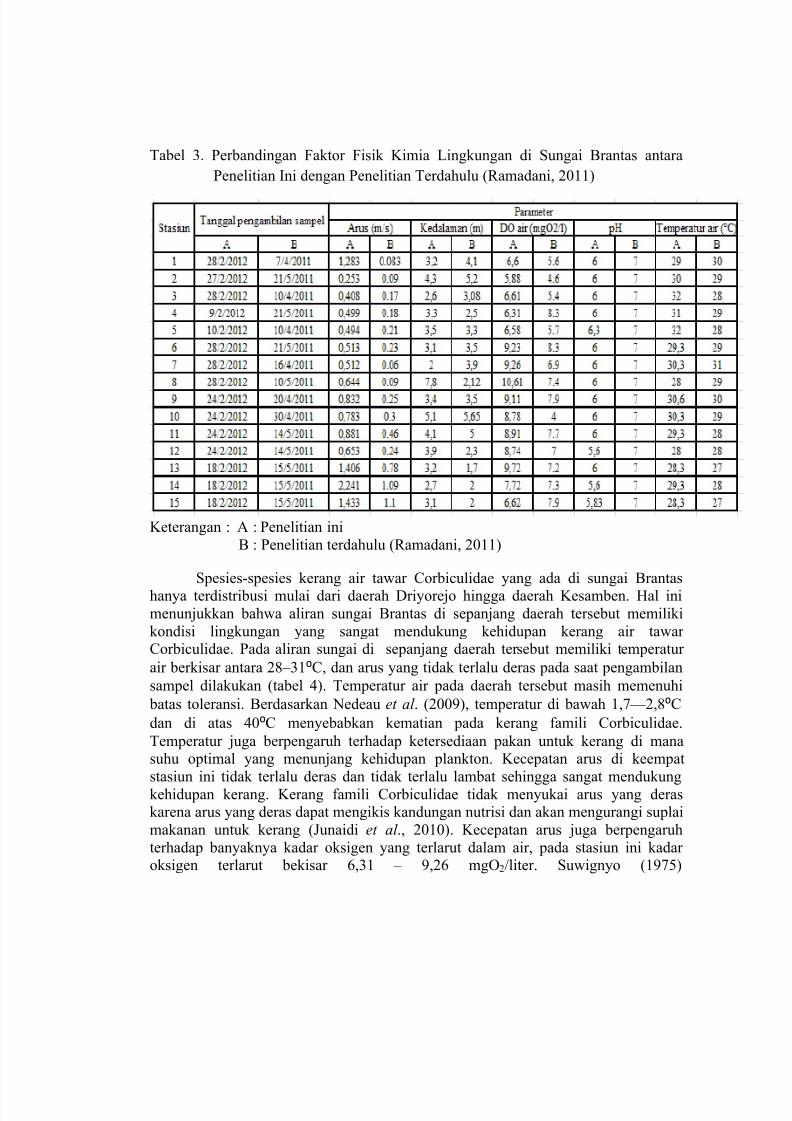
Data perbandingan hasil pengukuran parameter fisik kimia pada tiap stasiun dari penelitian ini dengan penelitian terdahulu (Ramadani, 2011) dapat dilihat pada tabel3.
7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 32/97
Tabel 3. Perbandingan Faktor Fisik Kimia Lingkungan di Sungai Brantas antara
Penelitian Ini dengan Penelitian Terdahulu (Ramadani, 2011)
Keterangan : A : Penelitian ini
B : Penelitian terdahulu (Ramadani, 2011)
Spesies-spesies kerang air tawar Corbiculidae yang ada di sungai Brantashanya terdistribusi mulai dari daerah Driyorejo hingga daerah Kesamben. Hal ini
menunjukkan bahwa aliran sungai Brantas di sepanjang daerah tersebut memiliki
kondisi lingkungan yang sangat mendukung kehidupan kerang air tawarCorbiculidae. Pada aliran sungai di sepanjang daerah tersebut memiliki temperatur
air berkisar antara 28 – 31⁰C, dan arus yang tidak terlalu deras pada saat pengambilan
sampel dilakukan (tabel 4). Temperatur air pada daerah tersebut masih memenuhi
batas toleransi. Berdasarkan Nedeau et al . (2009), temperatur di bawah 1,7 — 2,8⁰C
dan di atas 40⁰C menyebabkan kematian pada kerang famili Corbiculidae.
Temperatur juga berpengaruh terhadap ketersediaan pakan untuk kerang di manasuhu optimal yang menunjang kehidupan plankton. Kecepatan arus di keempat
stasiun ini tidak terlalu deras dan tidak terlalu lambat sehingga sangat mendukung
kehidupan kerang. Kerang famili Corbiculidae tidak menyukai arus yang deraskarena arus yang deras dapat mengikis kandungan nutrisi dan akan mengurangi suplai
makanan untuk kerang (Junaidi et al ., 2010). Kecepatan arus juga berpengaruh
terhadap banyaknya kadar oksigen yang terlarut dalam air, pada stasiun ini kadaroksigen terlarut bekisar 6,31 – 9,26 mgO2/liter. Suwignyo (1975)

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 33/97
dalam Prihatini (1999) menyatakan bahwa kerang menyukai lingkungan
dengan kandungan oksigen terlarut antara 3,8 — 12,5 mgO2/lt.
Kesimpulan dan Saran
Dari penelitian ini dapat disimpulkan bahwa Spesies kerang air tawar
Corbiculidae yang ada di perairan sungai Brantas adalah Corbicula javanica danCorbicula lacunae. Kelimpahan total masing-masing spesies kerang air tawar
Corbiculidae di seluruh stasiun penelitian adalah Corbicula javanica 21 individu/m2
dan Corbicula lacunae 74 individu/m2. Sedangkan nilai indeks dominansi spesies
kerang di seluruh stasiun penelitian Corbicula javanica 21,88% dan Corbiculalacunae 78,13%. Kedua spesies kerang air tawar Corbiculidae pada seluruh stasiun
penelitian di sungai Brantas memiliki pola keberadaan yang tidak kontinu.
Keberadaan yang kontinu hanya terjadi dalam daerah kisaran penyebaran di hilir
sungai Brantas. Pada penelitian ini dijumpai dua spesies kerang air tawarCorbiculidae, yaitu Corbicula lacunae dan Corbicula javanica, kelimpahan kedua
spesies tersebut berkisar antara 3 – 41 individu/m2
dengan C. lacunae sebagai spesies
yang dominan dengan indeks dominansi 78.13 %,
sedangkan pada penelitian
terdahulu (Ramadani, 2011) yang hanya mendapatkan satu spesies kerang air tawar
Corbiculidae, yaitu Corbicula lacunae dengan kelimpahan berkisar 5-34 individu/m2.
Pola distribusi longitudinal pada penelitian ini sama dengan pola distribusilongitudinal pada penelitian terdahulu (Ramadani, 2011), yaitu tidak kontinu pada
setiap stasiun penelitian.
Perlu adanya penelitian lebih lanjut mengenai keberadaan dan kelimpahankerang air tawar Corbiculidae pada waktu yang bebeda, pada waktu kecepatan arus
tidak terlalu deras agar tidak mengalami kesulitan pada saat pengambilan sampel,
serta metode sampling yang berbeda, misalnya pengambilan langsung dengan metode
plot agar mendapatkan hasil yang lebih mewakili mengenai keberadaan dankelimpahan kerang air tawar Corbiculidae di sungai Brantas.
Daftar Pustaka
Affandi, M. 1990. Pendugaan Tingkat Pencemaran Sungai Kali Surabaya dan Kanal
Kali Wonokromo dengan Menggunakan Indeks Diversitas Hewan Benthos
Makro. Skripsi. FMIPA. UNAIR.
Budiarto, N. 2011.Eksplorasi dan Visualisasi Morfologis Kerang Air Tawar
(Bivalvia: Corbiculidae) di Sungai BrantasJawaTimur. Skripsi. FSAINTEK.UNAIR.
Citriana, M.Y. 2002. Diversitas Dan Visualisasi Karakter Morfologi Invertebrata
Makro di Kali Surabaya.Skripsi. FMIPA. UNAIR.

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 34/97
Grabarkiewicz, J.D dan Davis, W.S. 2008. An Introduction to Freshwater mussel as
Biological indicator. U.S. Environmental Protection Agency:Washington DC.
Hidayati, U. 1995. Hewan Benthos Makro Sebagai Bioindikator di Perairan Sungai
Surabaya. Skripsi. FMIPA. UNAIR.
Junaidi, E., Effendi P. S. dan Joko. 2010. Kelimpahan Populasi dan Pola Distribusi
Remis (Corbicula sp) di Sungai Borang Kabupaten Banyuasin. FMIPA. Univ.
Sriwijaya, Jurnal Penelitian Sains. 13 (3D), 50-54.
Jutting, W.S.S. van B. 1953. Systematic Studies on the Non Marine Mollusc of the
Indo-Australia Archipelago : Revision of Freshwater Bivalvies.Vol 22. 19-
73.Zoological museum: Amsterdam.
Nedeau, E.J. A.K. Smith, J. Stone, dan S. Jepsen. 2009. Freshwater Mussels of the
Pacific Northwest 2nd
edition. The Xerces Society for Invertebrate
Conservation. Oregon.
Prihatini, W. 1999. Keragaman Jenis dan Ekobiologi Kerang Air Tawar FamiliUnionidae (Mollusca: Bivalvia) Beberapa Situ di Kabupaten dan Kotamadya
Bogor. Tesis. Prodi Biologi, Institut Pertanian Bogor: Bogor.
Ramadani, A.H. 2011. Keanekaaragaman dan Pola Distribusi Longitudinal KerangAir Tawar di Perairan Sungai Brantas. Skripsi. FSAINTEK. UNAIR.

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 35/97
1
PENGARUH KOMBINASI ZAT PENGATUR TUMBUH KINETIN DAN IBA
TERHADAP KULTUR ANTERA CABAI RAWIT (Capsicum frutescens L.)
Irma Catur Prastyo W.(1)
, Dwi Kusuma Wahyuni(*)(1)
, dan Hery Purnobasuki(1)
(1) Departemen Biologi, Fakultas Sains dan Teknologi, Universitas Airlangga,
Surabaya.
*Email: [email protected]
ABSTRACT
The aims of this research were to know the effect of plant growth regulator
combination of kinetin and IBA at various concentration of kinetin (0 ppm and 0,5
ppm) and IBA (0 ppm, 0,5 ppm, 1 ppm, and 1,5 ppm) concentration for anther culture
of Capsicum frutescens L. This research used Completely Randomized Design. Data
analized with Kruskal-Walis Test. Anthers of C. frutescens were cultured in the
double layer Murashige & Skoog medium (liquid upper layer). Solid medium
contained 30g/L sucrose and agar 8 g/L. Liquid layer contained sucrose 60 g/L with
addition of glutamin (500 mg/L). The results showed that combination of kinetin and
IBA gave not signification effect for anther culture of C. frutescens.
Keyword: Anther culture, Capsicum frutescens L., Plant growth regulator.
Pendahuluan
Cabai mempunyai nilai ekonomis tinggi karena salah satu pemanfaatannya
adalah sebagai bahan baku industri (Santika, 1999). Peningkatan produksi cabai
terhambat oleh kendala dalam kegiatan budidaya. Kendala-kendala tersebut antara
lain produktifitas rendah, ukuran dan bentuknya tidak sesuai dengan yang diharapkan,
serta terbatasnya kultivar unggul yang berumur pendek (Harpenas dan Dermawan,
2011).
Kultur jaringan telah dikenal banyak orang sebagai usaha mendapatkan
varietas baru (unggul) dari suatu jenis tanaman dalam waktu yang relatif lebih singkat
daripada dengan cara pemuliaan tanaman yang harus dilakukan penanaman secara

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 36/97
2
berulang-ulang sampai beberapa generasi. Kultur antera merupakan salah satu metode
untuk mendapatkan tanaman haploid yang unggul yang dapat di pergunakan untuk
menghasilkan kultivar-kultivar baru atau hibrida F1 (Hendaryono & Wijayani, 1994).
Tanaman membutuhkan zat pengatur tumbuh alami (fitohormon) untuk proses
pertumbuhan, yaitu zat pengatur tumbuh auksin dan sitokinin. Zat pengatur tumbuh
berfungsi merangsang pertumbuhan, misalnya pertumbuhan akar, tunas,
perkecambahan dan sebagainya (Sandra, 2003). Selain itu, konsentrasi auksin dan
sitokinin dalam media kultur in vitro memainkan peranan penting dalam induksi dan
regenerasi kalus menjadi tunas (Desriatin, 2011).
Penggunaan media padat dengan kombinasi zat pengatur tumbuh auksin dan
sitokinin terhadap kultur antera Capsicum annum dengan masa kultur selama 70 hari
telah banyak dilakukan, tetapi hasil yang diperoleh sangat rendah dan kurang
memuaskan (Wang et al., 1973; George & Narayanaswamy, 1973). Dumas de Vaulx
et al. (1981) melakukan kultur antera Capsicum annum pada media semi solid dengan
penambahan zat pengatur tumbuh kinetin dan 2,4-D belum dapat meningkatkan
perkembangan embrio.
Untuk mendapatkan embrio dalam kultur antera, memerlukan tahap penelitian
yang panjang. Sebelum mendapatkan embrio, antera terlebih dahulu akan membesar
yang diakibatkan oleh pemanjangan dari 3 lapisan antera yaitu epidermis, lapisan
tengah, dan endotesium serta perkembangan mikrospora yang ada di dalam antera
(Anonim, 2011). Setelah terjadi proses pemanjangan, pada bagian stomium akan
membuka sehingga mikrospora yang ada di dalamnya akan keluar dari antera
(Sanders et al., 2000).
Berdasarkan uraian diatas, dapat diketahui bahwa perlu dilakukan penelitian
kultur antera C. frutescens dengan menggunakan jenis dan konsentrasi zat pengatur
tumbuh tertentu untuk menemukan formulasi yang tepat dalam meningkatkan hasil
kultur antera cabai rawit.

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 37/97
3
Bahan dan Metode
Penelitian dilaksanakan di Laboratorium Fisiologi Tumbuhan Departemen
Biologi, Fakultas Sains dan Teknologi, Universitas Airlangga Surabaya. Antera yang
digunakan adalah antera cabai rawit (Capsicum frutescens L.) dengan sedikit warna
ungu pada bagian ujung antera. Eksplan ditanam pada media Murashige & Skoog
double layer dengan penambahan zat pengatur tumbuh kinetin (0 ppm dan 0,5 ppm)
dan IBA (0 ppm, 0,5 ppm, 1 ppm, dan 1,5 ppm). Pada media padat (bagian atas)
mengandung sukrosa 3 g/L dan agar 8 g/L sedangkan pada media cair mengandung
sukrosa 60 g/L tanpa agar dengan penambahan glutamin 500 g/L.
Rancangan penelitian yang digunakan merupakan eksperimental laboratorium
dengan menggunakan rancangan acak lengkap. Kombinasi perlakuan berjumlah 7 dan
masing-masing perlakuan terdiri 3 kali ulangan. Data yang dikumpulkan adalah
persentase antera yang membesar dan antera yang pecah.
Hasil Penelitian
Pada penelitian ini digunakan antera Capsicum frutescens L. yang memiliki
warna putih kekuningan dengan ujung antera sedikit berwarna ungu (Gambar 1.A).
Pada penelitian ini dilakukan pengamatan antera C. frutescens yang mengalami
perubahan ukuran setelah ditanam pada media MS double layer yang membesar
setelah 10 minggu masa kultur (Gambar 1.B). Dalam hal ini jumlah antera yang
membesar (berubah ukuran dari kecil ke besar) menjadi objek pengamatan. Selain
perubahan ukuran antera, dalam penelitian ini juga dilakukan pengamatan antera yang
pecah (Gambar 1.C) pada setiap perlakuan.

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 38/97
4
Gambar 1. Struktur antera Capsicum frutescens L. A) Antera sebelum dikulturpada media, B) Antera yang membesar, C) Antera yang pecah (anak
panah) (Bar=1 mm).
Tabel 1. Pengaruh zat pengatur tumbuh auksin dan kinetin terhadap persentase anteracabai rawit (C. frutescens ) yang membesar.
PerlakuanPersentase Rata-rata
Antera Membesar (%)
K0B0,5 43,33±5,77
K0B1 66,67±11,55
K0B1,5 43,33±11,55
K0,5B0 33,33±11,55
K0,5B0,5 60,00±0,00
K0,5B1 60,00±10,00
K0,5B1,5 73,33±11,55
Tabel 2. Pengaruh zat pengatur tumbuh auksin dan kinetin terhadap persentase antera
cabai rawit (C. frutescens ) yang pecah.
PerlakuanPersentase Rata-rata
Antera Membesar (%)
K0B0,5 3,33±5,77
K0B1 0.00±0.00
K0B1,5 6,67±5,77
K0,5B0 6,67±5,77
K0,5B0,5 6,67±5,77
K0,5B1 0.00±0.00
K0,5B1,5 10,00±10,00
Pada setiap perlakuan, persentase antera yang membesar dan antera yang
pecah berbeda-beda (Tabel 1 dan 2). Namun, berdasarkan Uji Kruskal-Wallis
A B C

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 39/97
5
penambahan kombinasi zat pengatur tumbuh kinetin dan IBA tidak berpengaruh
terhadap persentase antera yang membesar sehingga tidak bisa dilakukan uji lanjutan
statistik.
PEMBAHASAN
Berdasarkan hasil penelitian tampak bahwa pemberian kombinasi zat pengatur
tumbuh kinetin dan IBA tidak berpengaruh terhadap persentase antera yang
membesar dan antera yang pecah. Hal ini berbeda dengan penelitian Supena et al.
(2004) yang menyatakan bahwa penambahan kombinasi zat pengatur tumbuh zaetin
2,5 µM dan IAA 5 µM ke dalam media pertumbuhan mampu menginduksi pecahnya
antera Capsicum annum serta menghasilkan embrio normal. Selain itu, menurut
Dumas de Vaulx et al. (1981) penambahan 0,01 mg/L kinetin dan 0,01 mg/L 2,4-D
merupakan kombinasi zat pengatur tumbuh yang tepat dalam menginduksi antera
Capsicum annum menjadi tanaman haploid.
Hal ini kemungkinan dikarenakan oleh browning. Browning merupakan
perubahan warna eksplan yang awalnya berwarna putih kekuningan dengan ujung
sedikit berwarna ungu menjadi berwana coklat. Browning kemungkinan terjadi
karena adanya akumulasi komponen fenolik pada eksplan dimana terjadi proses
perubahan adaptif bagian tanaman akibat adanya pengaruh fisik (memar, pengupasan,
pemotongan, atau kondisi yang tidak normal) dan biokimia, bisa juga merupakan
gejala ilmiah dari proses penuaan (Kartiningrum dkk., 2011).
Senyawa kuinon akan menghambat aktifitas enzim yang selanjutnya dapat
mematikan tanaman (Agustin, 2005). Berdasarkan hal tersebut, kemungkinan adanya
browning pada eksplan mempengaruhi jumlah antera yang pecah. Sehingga
pemberian kombinasi zat pengatur tumbuh auksin dan kinetin tidak memberikan
pengaruh terhadap jumlah antera cabai rawit (C. frutescens) yang pecah.
Menurut Sheeler & Bianchi (1987), bagian sel tanaman yaitu vakuola sebagai
tempat untuk menyimpan air dan produk-produk sel khususnya metabolit sekunder
termasuk fenol. Pada saat proses pemotongan jaringan, vakuola terpotong dan akan

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 40/97
6
mengeluarkan fenol yang bereaksi dengan enzim fenol oksidase di dalam sitosol,
sehingga terbentuk kuinon yang menyebabkan warna berubah menjadi coklat.
Untuk mencegah proses browning perlu dilakukan penambahan antioksidan
ke dalam media kultur dan perendaman eksplan dalam antioksidan sebelum
dikulturkan. Hal tersebut dilakukan untuk mencegah terbentuknya kuinon. Zat yang
biasa ditambahkan adalah polivinylpyrrolidone. Penambahan zat polivinylpyrrolidone
(PVP) cukup efektif untuk menyerap senyawa toksik (Chung et al., 1987). Namun
pada penelitian ini tidak dilakukan pemberian antibiotik sehingga browning tidak
dapat dikendalikan.
KESIMPULAN
Berdasarkan hasil penelitian diatas, maka dapat disimpulkan bahwa
pemberian kombinasi zat pengatur tumbuh kinetin dan IBA pada media pertumbuhan
kultur antera cabai rawit (Capsicum frutescens L.) tidak memberikan pengaruh
terhadap persentase jumlah antera yang membesar dan persentase jumlah antera yang
pecah.
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada Ibu Jemjem yang telah membantu
memelihara donor eksplan selama penelitian berlangsung.
DAFTAR PUSTAKA
Anonim, 2011, Sexual Reproduction in Flowering Plants, http://textbook.s-
anand.net/ncert/class-xii/biology/2-sexual-reproduction-in-flowering-plants, 9Oktober 2012.
Agustin, W., 2005, Pemuliaan Tanaman Pisang dengan Kultur Anther , MakalahIndividu Program S3.

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 41/97
7
Chung Hu, Shi-Chou, H., Ching-Po, H., Han-chih, L., Cheng-Chi, C., and Li-Ping, P.,
1987, On The Inductive Conditions of Rice Pollen Plantlets in Anther Culture,
Plant Tissue Culture.
Desriatin, N. L., 2011, Pengaruh kombinasi zat pengatur tumbuh IAA dan Kinetinterhadap morfogenesis pada kultur in vitro tanaman tembakau ( Nicotiana
tabacum L. var. Prancak-95), ITS-Undergraduate-15274-1506100021-Paper .
Hal 2.
Dumas de Vaulx, R., Chambonnent, D., and Pochard E., 1981, Culture In Vitro
d’antheres de piment (Capsicum annum L.): amelioration des taux d’obtention
de plantes chez differents genotypes par des traitements a +35oC, Journal of
Agronomie 1: 859-864.
George, L. and Narayanaswamy, S., 1973., Haploid Capsicum through experimentalandrogenesis, Protoplasma 78: 467-470.
Harpenas, A. dan Dermawan, R., 2011, Budi daya cabai unggul, Penebar Swadaya:Jakarta.
Hendaryono, D. P. dan Wijayani, A., 1994, Teknik kultur jaringan (pengenalan dan
petunjuk perbanyakan tanaman secara vegetatif-modern), Kanisius:Yogyakarta.
Kartakaningrum, S., Purwito, A., Wattimena, G. A., Marwoto, B., dan Sukma, D.,2011, Teknologi Haploid Anyelir: Studi Tahap Perkembangan Mikrospora dan
Seleksi Tanaman Donor Anyelir, Jurnal Hortikultura , 21(2): 108-109.
Sanders, P. M., Lee, P. Y., Biesgen, C., Boone, J. D., Beals, T. P., Weiler, E. W., and
Goldberg, R. B., 2010, The Arabidopsis Delayed Dehiscence1 Gene Encedes an
Enzyme in The Jasmonic Acid Synthesis Pathway, The Plant Cell, California.
Sandra, E., 2003, Kultur jaringan anggrek skala rumah tangga, Agromedia Pustaka.
Jakarta.
Santika, A., 1999, Agribisnis tanaman cabai, Penebar Swadaya: Jakarta.
Sheeler, P. and D. G. Bianchi, 1987, Cell and Molecular Biology, John Willey andSons, Inc Canada.
Supena, E. D. J., Suharsono, S., Jacobsen, E., and Custers, J. B. M., 2004, Succesfuldevelopment of shed-microspore culture protocol for double haploid production

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 42/97
8
in Indonesian hot pepper (Capsicum annum), Plant Research International:
Netherlands.
Wang, Y. Y., Sun, C. S., Wang, C. C., and Chien, N. F., 1973, The induction of the
pollen planlets of triticale and Capsicum annum from anther culture, Scientia
Sinica 16: 147-151.

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 43/97
DETEKSI ENZIM LIPASE DAN BIOSURFAKTAN PADA
SUPERNATAN KULTUR Bacil lus sp. LII63B YANG DITUMBUHKAN
PADA MINYAK KELAPA
Ni’matuzahroh (*), Isnaini Septi Irmayanti, Tini Surtiningsih, Fatimah, Sri Sumarsih (**)
(*) Laboratorium Mikrobiologi, Departemen Biologi, FSAINTEK, Universitas Airlangga
(**) Laboratorium Biokimia, Departemen Kimia, FSAINTEK, Universitas Airlangga
Kampus C Unair , Jln. Mulyorejo, Surabaya - Indonesia,
*)E-mail: [email protected]
ABSTRAKPenelitian ini bertujuan untuk mengetahui produksi enzim lipase dan biosurfaktan
pada supernatan kultur bakteri Bacillus sp. LII63B. Penelitian ini merupakan penelitian
eksperimental laboratoris. Bacillus sp. LII63B ditumbuhkan pada media yang berisi Nutrien
Broth dan 2 % minyak kelapa. Kultur diinkubasi selama 48 jam pada suhu 28oC.Pertumbuhan bakteri dievaluasi dengan metode turbidimetri. Supernatan kultur bakteri
Bacillus sp. LII63B diukur nilai tegangan permukaannya menggunakan tensiometer Du-
Nouy, nilai aktivitas emulsifikasi dengan minyak uji solar, serta nilai aktivitas enzimnya
menggunakan substrat p-nitrofenol palmitat. Hasil penelitian menunjukkan bahwa supernatan
Bacillus sp. LII63B memiliki nilai aktivitas lipolitik sebesar 11,54 U/mL, dapat menurunkan
nilai tegangan permukaan sebesar 13,92 dyne/cm, tetapi tidak dapat mengemulsi minyak uji
solar.
Kata kunci: supernatan Bacillus sp. LII63B, aktivitas enzim lipase, tegangan permukaan,
aktivitas emulsifikasi.
ABSTRACTThe aim of this study was to determine lipase enzyme and biosurfactant production
from Bacillus sp. LII63B culture supernatant. This study is an experimental research
laboratory. Bacillus sp. LII63B was grown on medium containing Nutrient Broth and 2%
coconut oil. Culture was incubated in 28oC for 48 hours. The bacterial growth was evaluated
by turbidimetry method. Supernatant of Bacillus sp. LII63B bacterial culture was detected its
surface tension with Du-Nouy tensiometer, emulsification activity with diesel oil, and lipolitic
activity using p-nitrofenol palmitat substrat. The result of this research showed that
supernatant of Bacillus sp. LII63B have a lipolitic activity up to 11,54 U/mL, have the ability
to lower the surface tension of 13.92 dyne/cm. However, it do not have the ability to emulsifydiesel oil.
Key words: supernatant of Bacillus sp. LII63B, lipase enzyme activity, surface tension,
emulsification activity.
PENGANTAR
Kontrol yang hampir tidak pernah dilakukan terhadap limbah industri telah
mengakibatkan terjadinya pencemaran yang sangat luas dan merupakan limbah yang
berbahaya, salah satunya adalah limbah oil sludge yang dihasilkan pertamina (Novianti,

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 44/97
2010). Oil sludge adalah limbah pengolahan minyak bumi yang terdiri atas minyak, air, dan
padatan mineral (Dibble dan Bartha, 1979). Selama ini, metode menggunakan enzim lipase
dan biosurfaktan mulai banyak dikembangkan dalam hal membantu remidiasi limbah minyak
atau oil sludge agar tidak mencemari lingkungan.
Enzim lipase merupakan enzim yang mengkatalisis proses hidrolisis dan sintesis dari
ester yang terbentuk dari gliserol dan asam lemak rantai panjang. Enzim ini digunakan dalam
proses perombakan lemak dan minyak, sintesis bahan kimia dan farmasitikal, pembuatan
kertas, produksi bahan kosmetik (Sharma et al., 2001), dan perlakuan terhadap air yang kaya
lipid. Surfaktan ( surface active agent ) adalah molekul amfifatik yang terdiri atas gugus
hidrofilik dan hidrofobik, sehingga dapat berada di antara cairan yang memiliki sifat polar dan
ikatan hidrogen yang berbeda, seperti minyak dan air (Desai dan Banat, 1997). Penggunaan
biosurfaktan lebih menguntungkan dibandingkan dengan surfaktan sintetik karena tidak
toksik, pengemulsi yang baik, dan lebih mudah didegradasi oleh mikroorganisme (Desai dan
Banat, 1997).
Penelitian sebelumnya oleh Fatimah dan Nurhariyati (2011) berhasil melakukan
skrining bakteri hidrokarbonoklastik dari limbah rumah potong hewan daerah Pegirian,
Surabaya. Hasil skrining menunjukkan bahwa kultur Bacillus sp. LII63B merupakan bakteri
yang memiliki potensi lipolitik. Sehingga, penelitian ini dilakukan untuk mengetahui aktivitas
enzim lipase dan kemampuannya sebagai biosurfaktan dari supernatan kultur Bacillus sp.
LII63B, berkaitan dengan potensinya yang dapat digunakan sebagai agen remidiasi dalam
membantu melarutkan minyak atau oil sludge.
BAHAN DAN METODE
Bahan
Isolat Bacillus sp. LII63B, media Nutrient Agar (NA), media Nutrient Broth (NB), minyak
goreng, media Bushnell Hash, solar, p-nitrofenol palmitat, serta beberapa bahan kimia yang
lazim digunakan dalam penelitian mikrobiologi yaitu alkohol dan spirtus.
Metode
1. Pembuatan stok bakteri uji
Biakan murni dari isolat Bacillus sp. LII63B diinokulasi pada media miring Nutrient
Agar (NA) steril dengan metode gores, kemudian diinkubasi pada suhu kamar (28o C) selama
24 jam.

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 45/97
2. Pembuatan kultur bakteri dalam media Nutir ent Broth
Sebanyak 1-2 ose biakan bakteri dari NA miring diinkubasikan ke dalam media Nutrient
Broth (NB) steril selama 24 jam. Penggunaannya dalam media produksi, kultur Bacillus sp.
LII63B nilai absorbansinya dihitung dengan metode turbidimetik hingga mendapatkan nilai
OD sebesar 0,5 pada λ 650 nm.
3. Pembuatan media Bushnell H aas+ 1 % minyak
Komposisi nutrien 100 ml media Bushnell Haas cair meliputi: K 2HPO4 0,5 gram,
KH2PO4 0,5 gram, NH4 NO3 0,5 gram, MgSO4.7H2O 0,05 gram, CaCl2.2H2O 0,02 gram, dan
FeCl3 0,01 gram. Medium 50 mL Bushnell Haas cair dan 1% minyak goreng, kedalamnya
dimasukkan 5% suspensi sel dengan A650 nm = 0,1 dari isolat. Campuran dikultivasi dengan
pengocokan 120 rpm pada suhu 37 ºC selama 48 jam. Selama proses kultivasi berlangsung,
setiap 4 jam sekali dilakukan penghitungan biomassa sel untuk membuat kurva pertumbuhan
mikroba dan pengambilan supernatan untuk menentukan waktu optimal aktivitas lipolitiknya.
4. Penentuan nilai aktivitas enzim lipase Bacil lus sp. LII63B
Aktivitas lipolitik dilakukan dengan metode Pereira-Meirelles et al . (1997) dalam
Sumarsih (2005) yang dimodifikasi. Aktivitas enzim ditentukan dengan metode
spektrofotometrik dengan substrat p-nitrofenil palmitat ( p-NPP). Kultur disentrifugasi pada
9.000 rpm dengan suhu 4º C selama 15 menit, sehingga terpisah antara supernatan dan
suspensi sel. Supernatan yang diperoleh mengandung enzim lipase ekstraseluler.
Disiapkan tabung Eppendorf yang diisi 700 µl larutan p-NPP 0,503 mM dalam buffer
fosfat pH 7.0. Kemudian ditambahkan 300 µl supernatan yang mengandung enzim lipase
ekstraseluler. Campuran diinkubasi dalam waterbath pada suhu 37º C selama 30 menit.
Tabung Eppendorf diangkat dari waterbath lalu ditambahkan 100 µl larutan Na2CO3 0,2 M.
p-nitrofenol yang terbentuk ditandai dengan warna kuning, kemudian diukur dengan
spektrofotometer UV-Vis pada λ 410 nm. Absorbansi yang diperoleh selanjutnya akan
digunakan untuk menetapkan nilai aktivitas enzim lipase berdasarkan pada kurva standart p-
nitofenol (U/mL).
5. Pembuatan media produksi NB + 2% minyak
Media produksi yang digunakan adalah media Nutrient Broth (13 g/L) yang ditambahkan
dengan substrat minyak goreng sebanyak 2% (v/v). Sebanyak 100 mL media dimasukkan ke
dalam masing-masing botol kultur ukuran 500 mL dan disterilisasi pada suhu 121o
C selama
15 menit. Sebanyak 5% (v/v) kultur Bacillus sp. LII63B dalam Nutrient Broth dengan nilai
absorbansi 0,5 pada λ 650 nm diinokulasikan ke dalam masing-masing botol kultur berukuran

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 46/97
500 mL yang telah berisi media produksi dengan volume 100 mL. Inkubasi dilakukan dalam
shaker incubator pada suhu ruang dengan agitasi 120 rpm selama waktu optimal yang
dimiliki oleh Bacillus sp. LII63B. Supernatan diperoleh dengan cara kultur disentrifugasi
pada kecepatan 9.000 rpm dalam suhu 4°C selama 15 menit.
6. Pengukuran tegangan permukaan supernatan kultur
Sebanyak 10 mL supernatan Bacillus sp. LII63B yang telah didapatkan pada waku
optimal 16 jam, kemudian dihitung nilai tegangan permukaannya dengan tensiometer Du-
Nouy. Kontrol yang digunakan adalah media produksi tanpa kultur. Satuan nilai tegangan
permukaan yang digunakan adalah dyne/cm.
7. Pengukuran aktivitas emulsifikasi
Aktivitas emulsifikasi diukur dengan mencampurkan 1 mL supernatan dengan 1 mL
minyak minyak uji solar di dalam tabung reaksi. Setelah divortex selama 2 menit, aktivitas
emulsifikasinya diamati pada waktu inkubasi 1 jam dan 24 jam.
HASIL DAN PEMBAHASAN
1. Kurva pertumbuhan dan waktu optimal aktivitas enzim Bacil lus sp. LII63B
Sebelum melakukan produksi untuk supernatan Bacillus sp LII63B, dilakukan
pengamatan pertumbuhan kultur bakteri dalam media Bushnell Hash + 1% minyak dengan
melihat nilai kekeruhan (OD) dari kultur di tiap jam pengamatan dengan menggunakan
spektrofotometer. Berikut gambar pertumbuhan kultur Bacillus sp. LII63B selama 48 jam
inkubasi.
Gambar 1. Kurva pertumbuhan bakteri Bacillus sp. LII63B selama 48 jam dalam media
pertumbuhan Bushnell Hash+1% minyak goreng

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 47/97
Dari gambar yang ditunjukkan di atas dapat diketahui bahwa dalam waktu 48 jam,
bakteri Bacillus sp.LII63B mengalami fasa log atau fasa eksponensial setelah mengalami fasa
adaptasi selama ± 4 jam pada jam awal. Disebut sebagai fasa eskponesial karena pembelahan
sel merupakan persamaan eksponensial. Pada fasa ini perbanyakan jumlah sel meningkat
sampai pada batas tertentu dan membutuhkan energi lebih banyak dibandingkan dengan fasa
yang lain. Selain itu pada fasa ini sel menjadi cenderung lebih sensitif terhadap
lingkungannya (Yuneta, 2010).
Adanya pertumbuhan dari bakteri dapat dilihat pada perubahan warna media Bushnell
Hash + 1% minyak goreng yang semula berwarna bening, seiring dengan berjalannya waktu
warna media berubah menjadi agak keruh dan semakin keruh di titik yang terakhir yaitu pada
titik jam ke-48. Selain media yang berwarna keruh, terjadi pula pembentukan globul minyak
yang terjadi akibat terhidrolisisnya hidrokarbon minyak goreng oleh enzim lipase. Hidrolisis
hidrokarbon minyak goreng terjadi pada ikatan ester trigliserida yang diputus oleh enzim
lipase, sehingga menjadi asam lemak dan gliserol. Produk yang terbentuk selanjutnya
digunakan oleh bakteri untuk kebutuhan metabolisme selnya (Renjana, 2011). Berikut
merupakan gambar perubahan kekeruhan dan globul minyak yang terbentuk setelah beberapa
jam inkubasi.
Gambar 2. Perbedaan media kultur Bushnell Hash + 1%minyak (A= Kontrol ; B= Setelah
ada pertumbuhan bakteri)
Sedangkan, waktu optimal untuk uji aktivitas lipolitik dari kultur Bacillus sp. LII63B
berdasarkan pada nilai absorbansinya disajikan sebagai berikut.
BA
Minyak
masih
mengumpul
globul minyakmulai
terbentuk
Media
Nampak
keruh seiring
pertumbuhan
bakteri
Media
masih
bening
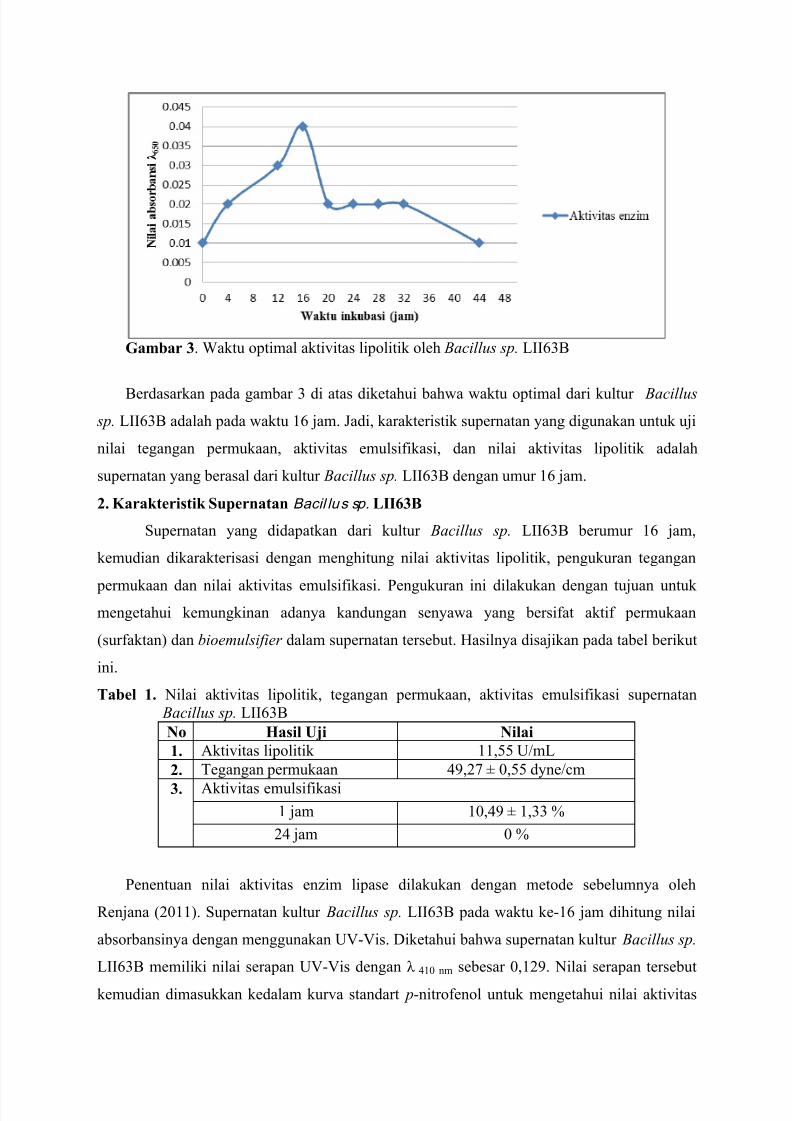
7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 48/97
Gambar 3. Waktu optimal aktivitas lipolitik oleh Bacillus sp. LII63B
Berdasarkan pada gambar 3 di atas diketahui bahwa waktu optimal dari kultur Bacillus
sp. LII63B adalah pada waktu 16 jam. Jadi, karakteristik supernatan yang digunakan untuk uji
nilai tegangan permukaan, aktivitas emulsifikasi, dan nilai aktivitas lipolitik adalah
supernatan yang berasal dari kultur Bacillus sp. LII63B dengan umur 16 jam.
2. Karakteristik Supernatan Bacil lus sp. LII63B
Supernatan yang didapatkan dari kultur Bacillus sp. LII63B berumur 16 jam,
kemudian dikarakterisasi dengan menghitung nilai aktivitas lipolitik, pengukuran tegangan
permukaan dan nilai aktivitas emulsifikasi. Pengukuran ini dilakukan dengan tujuan untukmengetahui kemungkinan adanya kandungan senyawa yang bersifat aktif permukaan
(surfaktan) dan bioemulsifier dalam supernatan tersebut. Hasilnya disajikan pada tabel berikut
ini.
Tabel 1. Nilai aktivitas lipolitik, tegangan permukaan, aktivitas emulsifikasi supernatan
Bacillus sp. LII63B
No Hasil Uji Nilai
1. Aktivitas lipolitik 11,55 U/mL
2. Tegangan permukaan 49,27 ± 0,55 dyne/cm
3. Aktivitas emulsifikasi
1 jam 10,49 ± 1,33 %
24 jam 0 %
Penentuan nilai aktivitas enzim lipase dilakukan dengan metode sebelumnya oleh
Renjana (2011). Supernatan kultur Bacillus sp. LII63B pada waktu ke-16 jam dihitung nilai
absorbansinya dengan menggunakan UV-Vis. Diketahui bahwa supernatan kultur Bacillus sp.
LII63B memiliki nilai serapan UV-Vis dengan λ 410 nm sebesar 0,129. Nilai serapan tersebut
kemudian dimasukkan kedalam kurva standart p-nitrofenol untuk mengetahui nilai aktivitas

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 49/97
lipolitiknya. Dari hasil perhitungannya, nilai aktivitas lipolitik dari supernatan kultur Bacillus
sp. LII63B adalah 11,55 U/mL.
Tabel 1 menunjukkan bahwa supernatan kultur Bacillus sp. LII63B memiliki nilai
tegangan permukaan 49,27 ± 0.55 dyne/cm dengan nilai tegangan permukaan kontrol NB
adalah sebesar 63,19 ± 2,71 dyne/cm. Hal ini menunjukkan bahwa supernatan Bacillus sp.
LII63B memiliki kemampuan sebagai sufraktan atau memiliki surface active agent karena
dapat menurunkan tegangan permukaan sebesar sebesar 13,92 dyne/cm terhadap media
kontrolnya. Menurut Francy et al. (1991) mengatakan bahwa bakteri memiliki potensi
menghasilkan biosurfaktan jika dapat menurunkan nilai tegangan permukaan ≥ 10 dyne/cm.
Dari hasil penurunan yang dihasilkan oleh supernatan kultur Bacillus sp. LII63B
menunjukkan bahwa supernatan tersebut memilki potensi sebagai biosurfaktan.
Aktivitas emulsifikasi supernatan Bacillus sp. LII63B dilakukan pada minyak uji
solar. Hasil uji aktivitas emulsifikasi, dengan kontrol akuades memilki nilai persentase
aktivitas emulsifikasi sebesar 0 % pada solar. Sedangkan, supernatan kultur Bacillus sp.
LII63B dengan waktu inkubasi 1 jam memiliki nilai aktivitas emulsifikasi sebesar 10,49 ±
1,33 % dan pada waktu inkubasi 24 jam nilainya 0 %. Hal ini menunjukkan bahwa supernatan
kultur Bacillus sp. LII63B tidak memiliki kemampuan untuk mengemulsi minyak uji solar
dengan baik, ditunjukkan dengan hasil emulsi yang dihasilkan sangat rendah pada waktu
inkubasi 1 jam dan pada waktu inkubasi 24 jam emulsinya sudah tidak ada. Namun,
kemampuannya dalam mengemulsi pada waktu inkubasi 1 jam menunjukkan bahwa
supernatan kultur Bacillus sp. LII63B memiliki kemampuan sebagai emulsifier .
Aktivitas emulsifikasi dari supernatan kultur Bacillus sp. LII63B pada minyak uji
solar di tunjukkan pada gambar berikut,
Gambar 4. Aktivitas emulsifikasi crude enzim lipase Bacillus sp. LII63B dengan minyak uji
solarKeterangan: A = Sebelum di vortex
B = setelah divortex 24 jamC = emulsi yang terbentuk
1 2 31 2 3
BA
C

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 50/97
1: Supernatan 12: Supernatan 23: Supernatan 3
Dari penelitian ini didapatkan hasil bahwa supernatan kultur Bacillus sp. LII63B
merupkan bakteri hidrokarbonoklastik yang memiliki kemampuan untuk menghasilkan enzim
lipase dan memiliki kemampuan sebagai surface active agent dan emulsifier . Sehingga,
dengan adanya kemampuan tersebut, bakteri Bacillus sp. LII63B bisa dijadikan sebagai salah
satu bakteri yang dapat digunakan dalam proses biodegradasi sebagai agen remediasi dalam
membantu melarutkan limbah minyak atau oil sludge.
KESIMPULAN
Dari penelitian ini disimpulkan bahwa supernatan kultur Bacillus sp. LII63B yang
ditumbuhkan pada minyak kelapa memiliki aktivitas lipolitiknya sebesar 11,55 U/mL,
penurunan tegangan permukaan sebesar 13,92 dyne/cm, namun tidak memiliki aktivitas
emulsifikasi yang baik pada minyak uji solar.
KEPUSTAKAAN
Desai, J.D. dan Banat, I.M. 1997. Microbial Production of Surfactant and Commercial
Potential. Microbiol and Molecular Rev. 61: 47-64.
Dibble, J.T. dan R. Bartha. 1979. Effect of Environmental Parameter on the
Biodegradation of Oil Sludge. Applied Enviromental Microbiology 37: 729-739.
Fatimah dan Nurhariyati, T. 2011. Eksplorasi Bakteri Proteolitik dan Lipolitik dari
Limbah Rumah Potong Hewan. Laporan Penelitian Hibah Riset . Universitas
Airlangga: Surabaya.
Francy, D.S., Thomas, J.M., Raymond, R.I., dan Word, C.H. 1991. Emulsification of
Hydrocarbon by Subsurface Bacteria. J. Ind. Microbiol. Vol. 8. pp. 237-246.
Khumalo, L.W., L. Majoko, J. S. Read, dan I. Ncube. 2002. Characterisation of Some
Underutilised Vegetable Oils and Their Evalution as Starting Materials for
Lipase-Chatalysed Production Cocoa Butter Equivalens. Ind Crops Products, 16 :
237-244.
Novianti, S. 2010. Pembuatan dan Karakterisasi Bata Konstruksi dengan
Memanfaatkan Limbah Sludge Pertamina Pangkalan Susu. Tesis. Universitas
Sumatera Utara.
Renjana, Elga. 2011. Skrining dan Uji Aktivitas Lipolitik Mikroba Hidrokarbonoklastik .
Skripsi. Fakultas Sains dan Teknologi. Universitas Airlangga: Surabaya.

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 51/97
Sharma R., Chisti Y., dan Banerjee U.C. 2001. Production, Purification, Characterization,
and Applications of Lipases. Biotechnol Adv 19 : 627 – 662.
Sumarsih, S. 2005. Skrining Bakteri Penghasil Lipase Termostabil dari Reaktor pada
Pabrik Minyak Goreng. Laporan Penelitian Dipa Penerimaan Bukan Pajak Tahun
Anggaran 2005. FMIPA. Universitas Airlangga.
Yuneta, Rena. 2010. Pengaruh Suhu pada Lipase dari Bakteri Bacill us subtili s . Skripsi.
Jurusan Kimia FMIPA Institut Teknologi Sepuluh Nopember: Surabaya.

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 52/97
UJI ANTIBAKTERI EKSTRAK KULIT BUAH DAN BIJI MANGGIS (Garcinia
mangostana) PADA BAKTERI PENYEBAB JERAWAT (Staphylococcus
epidermidis ) DENGAN MENGGUNAKAN SOLVEN ETANOL
Krisnina Maharani, Drs. Agus Supriyanto, M. Kes, dan Tri Nurhariyati, S. Si, M.
Kes, Prodi S-1 Biologi, Departemen Biologi, Fakultas Sains dan Teknologi,
Universitas Airlangga, Surabaya.
ABSTRAK
Penelitian ini bertujuan untuk mengetahui pengaruh konsentrasi ekstrak
kulit buah dan biji manggis (Garcinia mangostana) sebagai antibakteri pada bakteri penyebab jerawat Staphylococcus epidermidis dengan menggunakan
solven etanol. Penelitian eksperimental ini menggunakan RAL (Rancangan Acak
Lengkap) yang terdiri atas uji difusi dan uji dilusi. Uji difusi terdiri atas 8
konsentrasi ekstrak, masing-masing terdiri atas (0, 12,5, 25, 50, 100, 200, 500, dan
1.000 ppm). Uji dilusi terdiri atas 12 konsentrasi ekstrak, masing-masing terdiri
atas (0, 12,5, 25, 50, 100, 200, 500, 1.000, 1.125, 1.250, 1.500 dan 2.000 ppm).
Parameter yang diukur untuk uji difusi adalah diameter daerah penghambatan
(mm), masing-masing perlakuan terdiri atas 3 ulangan. Uji dilusi digunakan untuk
mencari nilai MIC dan MBC. Data diameter daerah penghambatan dianalisis
secara statistika menggunakan uji Kruskal-Wallis dilanjutkan uji Mann-Whitney,
sedangkan data uji dilusi dianalisis secara deskriptif. Hasil penelitian uji difusimenunjukkan bahwa konsentrasi ekstrak kulit buah dan biji manggis (Garcinia
mangostana) berpengaruh menghambat pertumbuhan bakteri. Nilai tertinggi
untuk ekstrak kulit buah manggis pada konsentrasi 1000 ppm sebesar (0,8192 ±
0,05) mm, dan pada konsentrasi yang sama ekstrak biji sebesar (0,7208±0,05)
mm. Ekstrak kulit buah nilai MIC berada pada 1.000 ppm dan MBC pada 1.125
ppm, sedangkan pada ekstrak biji nilai MIC pada 2.000 ppm dan MBC belum
dapat ditemukan, namun memberikan pengaruh penghambatan pertumbuhan.
Kata kunci : Garcinia mangostana, Staphylococcus epidermidis, MIC, MBC

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 53/97
ABSTRACT
This research was aimed to know determine the effects of concentration of the
fruit skin and seed extracts of mangosteen (Garcinia mangostana) as an
antibacterial in acne-causing bacteria Staphylococcus epidermidis using ethanol
solvent. This experimental study using the CRD (Completely Randomized
Design) which consists of diffusion test and the test dilution. Diffusion test
consisted of 8 concentrations of extracts, each consisting of (0, 12,5, 25, 50, 100,
200, 500, dan 1.000 ppm). Dilution test consisted of 12 concentrations of extract,
each consisting of (0, 12,5, 25, 50, 100, 200, 500, 1.000, 1.125, 1.250, 1.500 dan
2.000 ppm). The parameters measured for the diffusion test is the diameter of the
inhibition area (mm), each treatment consisted of 3 replications. Dilution test is
used to find the value of MIC and MBC. Diameter of the inhibition data were
analyzed using a statistical test of Kruskal-Wallis test followed Mann-Whitney, thedilution of the test data were analyzed descriptively. The results of diffusion tests
showed that the concentration of skin extract of the fruit and seeds of mangosteen
(Garcinia mangostana) gives the effect of growth inhibition. The highest value for
the mangosteen rind extract at a concentration of 1000 ppm (0.8192 ± 0.05) mm,
and the mangosteen fruit seed extract at a concentration of 1000 ppm (0.7208 ±
0.05) mm. Fruit peel extract MIC values are at 1000 ppm and 1125 ppm MBC on,
while the MIC values of fruit seed extract at 2,000 ppm and MBC can not be
found, but it gives the effect of growth inhibition.
Key word : Garcinia mangostana, Staphylococcus epidermidis, MIC MBC
PENDAHULUAN
Jerawat merupakan kondisi abnormal kulit akibat gangguan berlebih
produksi kelenjar minyak ( sebaceous gland ) yang menyebabkan penyumbatan
folikel rambut dan pori-pori kulit sehingga terjadi peradangan pada kulit.
Keaktifan kelenjar minyak di bawah kulit dirangsang oleh hormone androgen
(hormone pertumbuhan). Pengentalan kelenjar minyak terjadi menutupi selubung
rambut, mendesak keluar dalam bentuk lemak kental, yang disebut jerawat
(Harmanto, 2006). Keberadaan mikroorganisme di tubuh manusia juga
mempengaruhi munculnya jerawat. Karena kebanyakan bakteri kulit dijumpai
pada epitelium (lapisan luar bersisik), membentuk koloni pada permukaan sel-sel
mati (aerobik) dan di dalam kelenjar lemak dijumpai bakteri-bakteri anaerob
lipolitik, seperti Staphylococccus epidermidis yang bersifat nonpatogen pada kulit
namun dapat menimbulkan penyakit, termasuk jerawat akibat lipase
Staphylococccus epidermidis melepaskan asam-asam lemak dari lipid dan

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 54/97
menyebabkan iritasi jaringan (Naturakos, 2009). Sejauh ini pengobatan jerawat
dilakukan dengan memberikan obat antibiotika, seperti doksisiklin untuk
menghambat perkembangan mikroba dan mengurangi jumlah asam lemak bebas
(Harmanto, 2006). Bahan antimikroba merupakan bahan yang dapat mengganggu
pertumbuhan dan metabolisme mikroba, yang menyatakan penghambatan
pertumbuhan. Antimikroba selain diperoleh dari bahan-bahan sintetik akhir-akhir
ini banyak ditemukan dari bahan alam seperti pada tanaman, rempah-rempah atau
dari mikroorganisme (Pelczar dan Chan, 1988). Melalui analisis kuantitatif
fitokimia biji manggis memiliki kemampuan sebagai antifungi dan antimikroba
yang masih memerlukan kajian dan penelitian lanjutan (Ajayi, 2011). Berdasarkan
penampisan fitokimianya diketahui biji manggis mengandung tanin, saponin,
terpenoid, flavonoid, dan alkanoid (Ajayi, 2011). Sementara kulit buah terluar
mengandung tanin, resin, alkaloid, saponin, triterpenoid, fenolik, flavonoid,
glikosida, steroid dan lateks kering manggis dengan sejumlah zat warna kuning
yang berasal dari dua metabolit sekunder, yaitu mangostin (C20H22O5) atau
mangosim (Nadkarni and Nadkarni, 1999). Senyawa aktif antibakteri dalam kulit
buah dan biji manggis adalah senyawa flavonoid, tannin dan saponin. Perlu
dilakukan uji pembanding keefektifan zat antibakteri kulit buah dan biji manggis,
mengingat perbedaan kosentrasi ekstrak akan menunjukkan perbedaan hasil uji
antibakteri (Mayachiew and Devahastin, 2008). Pohon manggis yang berumur
lebih dari 100 tahun dapat menghasilkan 50-80 kilogram (500-800buah/pohon)
(Paramawati, 2010). Efek antibakteri dari kulit buah dan biji manggis dapat
diketahui dengan melakukan uji yang dilakukan secara invitro dengan
menggunakan difusi cakram dan metode dilusi. Metode difusi cakram
diindikasikan dengan terbentuknya daerah hambatan pertumbuhan di sekitar
cakram uji. Metode dilusi dilakukan setelah sebelumnya melakukan uji dengan
metode difusi cakram sehingga didapatkan nilai MIC ( Minimal Inhibitory
Concentration), yaitu konsentrasi terendah suatu agen antimikroba yang
diperlukan untuk menghambat pertumbuhan mikroba serta MBC ( Minimal
Bactericidal Concentration), yaitu konsentrasi daya bunuh bakteri minimum.
Diduga komponen kimia ekstrak kulit buah dan biji manggis mempunyai

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 55/97
kemampuan sebagai antimikroba dapat mempengaruhi zona penghambatan
pertumbuhan pada bakteri
METODE PENELITIAN
Penelitian ini telah dilakukan di Laboratorium Mikrobiologi, Departemen
Biologi, Fakultas Sains dan Teknologi, Universitas Airlangga, Surabaya. Waktu
penelitian bulan Desember 2011 hingga Februari 2012. Penelitian ini merupakan
penelitian eksperimental menggunakan Rancangan Acak Lengkap. Kulit buah dan
biji manggis (Garcinia mangostana) diekstrak dengan pelarut etanol. Ekstrak kulit
buah dan biji dengan beberapa konsentrasi yang berbeda (0, 12,5, 25, 50, 100,
200, 500, dan 1.000 ppm,) yang dilakukan dengan tiga kali ulangan.
Prosedur penelitian terdiri dari beberapa tahapan, yaitu sebagai berikut :
1. Penyiapan Bahan Penelitian
Untuk membuat suatu ekstrak, terlebih dahulu dilakukan pengumpulan
bahan berupa kulit dan biji buah manggis. Agar homogen, warna dan usia kulit
buah manggis diusahakan seragam dari kulit buah manggis yang telah matang.
Tingkat kematangan buah manggis berdasarkan Indek atau tahapan, yaitu warna
kulit buah ungu kemerahan. Buah mulai masak dan siap dikonsumsi. Getah telah
hilang dan isi buah mudah dilepaskan. Tahapan ekstraksi adalah sebagai berikut:
(1)
Mencuci kulit buah dan biji manggis hingga bersih, memotong menjadi
potongan yang lebih kecil, kemudian mengering anginkan sampai kering.
Untuk mempermudah saat penggerusan. Kemudian menggerus dengan alat
penumbuk dan blender hingga halus dan ditimbang sebanyak 400 gram.
Merendam serbuk kulit buah manggis atau dimaserasi dengan pelarut
ethanol sebanyak 800 mL dalam tabung kaca sampai hasil tumbukan
terendam seluruhnya, diaduk-aduk. Filtrat yang didapatkan ditampung
kemudian menambahkan lagi pelarut ethanol hingga didapatkan filtrat yang
tidak berwarna.

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 56/97
(2)
Menyaring hasil rendaman dengan kertas saring, hingga diperoleh filtrat-
filtrat lalu dimasukkan ke dalam rotary vacuum evaporator sampai solven
ethanol habis menguap. Bahan kental serta pekat yang tertinggal disebut
ekstrak. Kemudian masing-masing ditimbang.
2. Peremajaan isolat murni bakteri
Kultur murni bakteri ditanam secara aseptik pada tabung reaksi yang berisi
media NA ( Nutrient Agar ) padat miring dengan menggunakan jarum ose
digoreskan ( streak ), kemudian diinkubasi pada inkubator selama 24 jam dengan
suhu kamar.3. Uji Aktivitas Antimikroba
Metode Kertas Cakram (disc dif fusion method )
Metode kertas cakram dilakukan dengan menyiapkan media Mueller Hinton Broth
(MHB) steril sebagai media pertumbuhan khusus untuk uji antimikroba. Suspensi
mikroba uji dibuat dengan mengatur kekeruhan mikroba sampai didapat nilai
rapat optis (Optical Density) setara dengan standar McFarland 0,5 (108 CFU/ml)
Sebanyak 1 mL suspensi mikroba uji dimasukkan ke dalam cawan petri steril,
kemudian ditambahkan 15 mL media Mueller Hinton Agar (MHA) ke dalam
cawan tersebut lalu dihomogenkan dengan cara menggerakkan cawan seperti
angka delapan, setelah itu didiamkan sampai memadat. Selanjutnya sebanyak 3
lembar kertas cakram steril dengan diameter 6 mm dan ketebalan yang sama
diletakkan di permukaan agar dengan jarak yang diupayakan sama satu sama lain
membentuk segitiga. Sebelum diletakkan di permukaan agar, pada kertas tersebut
diinjeksikan masing-masing sebanyak 15µl ekstrak kulit buah dan biji manggis
dari masing-masing konsentrasi. Masing-masing mempunyai 3 ulangan.
Kemudian diinkubasi pada suhu 37oC selama 48 jam. Adanya aktivitas
antimikroba ditunjukkan terbentuknya daerah penghambatan (halo) disekitar
cakram uji (Bailey dan Scott, 2004). Diameter daerah hambatan diukur dengan
menggunakan jangka sorong (LC-0,05mm).
Metode Pengenceran dalam Tabung (tube dilution method
Metode pengenceran dalam tabung dibuat dengan membuat suspensi
mikroba uji pada media Mueller-Hinton Broth (MHB) dengan mengatur

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 57/97
kekeruhan mikroba sampai didapat nilai rapat optis (Optical Density) setara
dengan standar McFarland 0,5 (108 CFU/ml). Masing-masing sebanyak 1 mL
ekstrak kulit dan biji buah manggis dengan beberapa konsentrasi (12,5 ppm, 25
ppm, 50 ppm, 100 ppm, dan 200 ppm) dimasukkan dalam tabung reaksi yang
sebelumnya telah diisi dengan 1 mL suspensi mikroba uji. Kultur dihomogenkan
dan diinkubasi selama 24 jam. Apabila terdapat aktivitas antimikroba, maka tidak
ada kekeruhan (larutan jernih) dalam kultur tersebut. Dari seluruh kultur diambil
sebanyak 1 mL untuk ditumbuhkan pada media MHA dan diinkubasi selama 24
jam. Ketika terjadi pertumbuhan pada media MHA, maka pada konsentrasi
tersebut merupakan nilai MIC ekstrak kulit buah dan biji manggis dan apabila
tidak terjadi pertumbuhan pada media MHA tersebut, maka pada konsentrasi
tersebut didapatkan nilai MBC ekstrak kulit buah manggis dan nilai MBC ekstrak
biji manggis (Bailey dan Scott, 2004).
4. Analisis Data
Data yang diperoleh dari penelitian ini adalah diameter daerah
penghambatan ekstrak buah dan biji manggis terhadap pertumbuhan
Staphylococcus epidermidis, nilai MIC, nilai MBC dan jumlah sel bakteri
(CFU/mL). Data TPC, data nilai MIC dan MBC ekstak kulit buah dan biji
manggis terhadap pertumbuhan Staphylococcus epidermidis dianalisis secara
deskriptif. Data diameter daerah penghambatan ekstrak kulit buah dan biji
manggis terhadap pertumbuhan Staphylococcus epidermidis dianalisis secara
statistik. Uji ANAVA dilakukan atas dasar asumsi bahwa data berdistribusi
normal dan variansi data homogen. Jika p < 0,05 (tidak ada beda nyata) pada
Homogencity of Variance variansi data tidak homogen, maka analisis dilanjutkan
dengan uji non-parametrik K-Independent sampel yaitu Kruskal-Wallis. Jika p <
0,05 (ada beda nyata ) pada uji Kruskal-Walls maka analisis dilanjutkan dengan
uji Mann-Whitney.
HASIL DAN PEMBAHASAN
Hasil ekstraksi
7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 58/97
Hasil ekstraksi kulit dan biji buah manggis (Garcinia mangostana) didapatkan
suspensi gel berwarna coklat untuk kulit buah dan berwana kuning kecoklatan
untuk biji manggis (Garcinia mangostana). Sebanyak 0,4 g ekstrak kulit dan biji
masing-masing dilarutkan dalam 5 mL etanol dan 95 mL aquades steril untuk
dibuat variasi konsentrasi tiap ekstrak, selanjutnya dilakukan uji aktivitas
antimikroba pada Staphylococcus epidermidis.
Pengaruh konsentrasi ekstrak pada proses antibakteri
1. Metode difusi kertas cakram (disc dif fusion method )
Data diameter penghambatan Pertumbuhan Staphylococcus epidermidis pada
variasi konsentrasi ekstrak kulit dan biji buah manggis (Garcinia mangostana)
berdistribusi normal dan tidak homogen dari hasil analisis uji Kolmogorov-
Smirnov dan Homogeneity of variances sehingga dilanjutkan dengan uji non
parametrik alternatif yaitu uji Kruskal-Wallis. Hasil uji Kruskal-Wallis
menunjukkan bahwa, didapatkan nilai signifikansi < 0,05 (lampiran 3),
konsentrasi ekstrak kulit buah manggis dan biji memiliki pengaruh terhadapdiameter daerah penghambatan pertumbuhan (=0,013, pada kulit buah manggis
dan = 0,051, pada biji) sehingga Ho ditolak dan H1 diterima serta dilanjutkan
dengan uji Mann-Whitney untuk mengetahui beda pengaruh antara perlakuan.
Gambar 1. Grafik diameter daerah penghambatan pertumbuhan Staphylococcus
epidermidis pada setiap konsentrasi ekstrak.

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 59/97
Data daerah penghambatan secara keseluruhan dari ekstrak kulit buah
manggis, data yang terbesar adalah (0,8192 ± 0,05) mm pada konsentrasi 1.000
ppm, sedangkan rata-rata daerah penghambatan terkecil adalah (0,453±0,05) mm
pada konsentrasi 12,5 ppm. Sementara untuk data daerah penghambatan biji buah
manggis secara keseluruhan, data terbesar adalah (0,7208±0,05) mm dan data
terkecil adalah (0,4367±0,05) mm pada konsentrasi 12,5 ppm. Senyawa
antibakteri ekstrak kulit dan biji manggis mulai menunjukkan penghambatan
pertumbuhan Staphylococcus epidermidis pada konsentrasi 12,5 ppm yang
ditunjukkan adanya daerah penghambatan (hallo) dengan diameter sebesar
(0,453±0,05) mm pada kulit buah manggis dan (0,4367±0,05) mm pada biji buah
manggis.
2. Metode pengenceran dalam tabung (tube dilution method )
Tahapan penentuan konsentrasi ekstrak yang diencerkan dalam tabung
didasarkan atas hasil uji difusi cakram kertas yang sebelumnya telah dilakukan,
dimana pada konsentrasi ekstrak 12,5 ppm sudah terbentuk zona (hallo).
Berdasarkan hasil uji dilusi diketahui bahwa pada konsentrasi 12,5 ppm, 25 ppm,50 ppm, 100 ppm, 200 ppm, 500 ppm, dan 1.000 ppm ekstrak kulit buah manggis
dan biji manggis menunjukkan penghambatan pertumbuhan bakteri
Staphylococcus epidermidis dengan ditunjukkannya perubahan kekeruhan larutan
suspensi setelah diinkubasi selama 24 jam, namun mengingat suspensi manggis
merupakan larutan yang berwarna membuat hasil dari uji menjadi bias, karena
kekeruhan akibat adanya ekstrak dan kekeruhan bakteri hampir tidak bisa
dibedakan. Sehingga visualisasi kultur tidak dapat digunakan untuk menentukan
nilai MIC dan MBC.
Perlakuan dilanjutkan dengan metode TPC yang merupakan salah satu teknik
penghitungan koloni mikroba yang paling sederhana. Jumlah koloni bakteri atau
TPC CFU/mL (10log) dianalisis secara deskriptif untuk mengetahui pertumbuhan
bakteri Staphylococcus epidermidis
yang telah diberikan perlakuan ekstrak kulit
buah manggis ataupun biji buah manggis manggis pada beberapa konsentrasi pada
media MHA ( Muller Hinton Agar ).

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 60/97
KESIMPULAN
Konsentrasi ekstrak kulit buah manggis (Garcinia mangostana)
mempengaruhi diameter daerah penghambatan pertumbuhan Staphylococcus
epidermidis. Rerata diameter daerah penghambatan pertumbuhan terbesar pada
konsentrasi 1.000 ppm sebesar 0,8192±0,05 mm.. Konsentrasi ekstrak biji buah
manggis (Garcinia mangostana) mempengaruhi diameter daerah penghambatan
pertumbuhan Staphylococcus epidermidis. Rerata diameter daerah
penghambatan pertumbuhan terbesar pada konsentrasi 1.000 ppm sebesar
0,733±0,05 mm.. Konsentrasi minimum ekstrak kulit buah (Garcinia
mangostana) nilai MIC ( Minimal Inhibitory Concentration) pada konsentrasi
1.000 ppm dan nilai MBC ( Minimal Bactericidal Concentration)
Staphylococcus epidermidis pada konsentrasi 1.125 ppm, sementara pada biji
buah manggis (Garcinia mangostana) nilai MIC ( Minimal Inhibitory
Concentration) pada konsentrasi 2.000 ppm, namun belum ditemukan nilai
MBC ( Minimal Bactericidal Concentration) Staphylococcus epidermidis.
DAFTAR PUSTAKA
Ajayi, I. A., Adebowale, K. O., Dawodu, F. O., and Oderinde, R. A. 2011.
Chemical analysis and preliminary toxicological evaluation of Garcinia
mangostana seeds and seed oil. 999-1004. Diakses 28 November 2011.
Akiyama, H., Fujii, K., Yamasaki, O., Oono, T., Iwatsuki, T., 2001. Antibacterial
Action of Several Tannins Agains Staphylococcus aureus, Journal of Anti- microbial Chemotherapy . Vol. 48 : 487-91.
Bailey, W. R., and Scott, E. G. 2004. Diagnostic Mikr obiologi . Elevent Edition.The CV Mosby Company. Saint Louis. 168-187.
Burkill, H.M. 1994. The useful plants of West Tropical Africa. Edition 2. Vol.
2. Families E-I. Royal Botanic Gardens Kew. ISBN No. 0-947643-56-7.
Bernasconi, G., Gerster, H., Hauser, H., Stauble, H., and Schneiter, E. 1995.
Teknologi Kimia Bagian 2 (edisi terjemahan ). PT Pranadya Paramita.
Jakarta.
Cappuccino, J.G. and N. Sherman, 2005. Microbiology-A laboratory manual.
7th Edn., Pearson Int., Ontario, Canada.

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 61/97
Chen, S. X., Wan, M., and Loh, B. N. 1996. Active constituents against HIV-1
protease from Garcinia mangostana, Planta Med . 62(4):381-2.
Fardiaz, S. 1998. Mikrobiologi Pangan. Dirjen Pendidikan Tinggi. Depdikbud.
PAU Bogor.
Harborne J.B. 1987. Metode Fitokimia Penuntun Cara Modern Menganalisis
Tumbuhan. Bandung :Penerbit ITB.
Harmanto, N. 2006. Ibu Sehat dan Cantik dengan Herba. PT. Elex Media
Komputindo, Jakarta.
Jinsart, W., Ternai, B., Buddhasukh, D., and Polya GM. 1992. Inhibition of
wheat embryo calcium-dependent protein kinase and other kinases bymangostin and gammamangostin. Phytochemistry. 31(11):3711-3713.
Kanisius. 2003. Bibit Manggis. Penerbit Kanisius, Yogyakarta.
Kastaman, R. 2007. Prospective Analysis on Development of Mangosteen
(Garcinia mangostana ) Processing Product in order to Improve Farmers
Income (Case Study in Kecamatan Puspahiang Kabupaten Tasikmalaya).
Jurnal Agrikultural. Vol 18. No. 15.
Kristanti, A. N., Aminah, N. S., Tanjung, M., dan Kurniadi, B. 2008.Buku Ajar
Fitokimia. Airlangga University Press, Surabaya.
Mayachiew, P., Pasuk, S. and Ritthiruangdej, P. 2008. Relationship Between
antioxidant activity of Indian gooseberry and galangan extracts. LWT –
Food Science and technology 41 : 1153 – 1159.
Nadkarni, K. M., and Nadkarni, A. K. 1999. Indian Materia Medica - with
Ayurvedic, Unani-Tibbi, Siddha, Allopathic, Homeopathic, Naturopathicand Home remedies. Popular Prakashan Private Ltd., Bombay, India. ISBN
No. 81-7154-142-9. Vol.1.
Naturakos. 2009. Bahan-bahan kosmetik sebagai antiacne . Juli, Vol. IV/ no. 10,
Hal 1-3.
Paramawati, R.. 2010. Dasyatnya manggis untuk menumpas penyakit.
Agromedia Pustaka. Jakarta.
Pelczar, Jr., M.J., dan Chan, E.C.S. 1988. Dasar-dasar Mikrobiologi, Jilid 1 &
2. Penerbit Universitas Indonesia, Jakarta.
Purnomo, M. 2001. Isolasi Flavonoid dari Daun Beluntas (Pluchea indica
Less) yang mempunyai Aktivitas Antimikroba Terhadap Penyebab Bau

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 62/97
KAJIAN HISTOLOGI GONAD TERIPANG Phyll ophorus sp. PADABULAN FEBRUARI, MARET DAN APRIL 2012
Dwi Winarni(1), Moch. Affandi(1), Endang Dewi Masithah(2), Machmudhatun Nisa(1)
(1) Departemen Biologi Fakultas Sains dan Teknologi Universitas
Airlangga,(2)
Departemen Budidaya Perairan Fakultas Perikanan dan Kelautan Universitas
Airlangga,
Email :[email protected]
ABSTRACT
The aim of this study was to know the histological structure of ball sea cucumbers
( Phyll ophorus sp .) gonad in February, March and April 2012. Those ball sea
cucumbers were taken from East Surabaya Coastal, 25 animals per month. Five
tubules from each gonad were fixed in neutral buffered formalin, processed by
paraffin method, and stained with hematoxylin-eosin. Histological analysis was
done in six serial cross section (4 m thick, at 40 m from one to another section) for each tubule.
The result showed that Phyll ophorus sp. has separate sexes and asynchronousreproductive pattern. There were various stages of oocytes in the female gonads
which were classified to previtellogenic oocytes (it has diameter < 150 µm),
vitellogenic oocytes (it has diameter150-350 µm) and post-vitellogenic oocytes (it
has diameter> 350 µm). While in the male gonads showed spermatogenic cells,
there were spermatocytes, spermatid and spermatozoa. Typically, various stages
of gametogenic cells could occupy in the same section of tubule, both in male and
female gonad. The thickness of gonadal wall was tend to be affected by lumen
diameter and it will shrink when the lumen was dominated by post-vittelogenic
oocytes in female gonads and spermatozoa in male gonads.
Key words: Phyllophorus sp. , ball sea cucumber, gonad, oocytes, spermatozoa
PENGANTAR
Teripang atau mentimun laut yang termasuk dalam kelas Holothuroidea
merupakan hasil laut yang mempunyai nilai penting dan memiliki potensi yang
cukup besar karena Indonesia memiliki perairan pantai dengan habitat teripang
yang cukup luas (Dewi et al., 2010). Teripang juga merupakan salah satu produk
perikanan yang telah lama dikenal dan dikonsumsi oleh masyarakat pesisir di

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 63/97
Indonesia, dan juga sangat dikenal di negara-negara Benua Eropa, Jepang, dan
Amerika Serikat. Akan tetapi, berdasarkan hasil pengamatan di beberapa lokasi
penangkapan teripang, mulai dirasakan bahwa peningkatan produksi dengan cara
penangkapan dari alam tidak dapat lagi dipertahankan, karena dikhawatirkan akan
merusak sumberdaya hayati (Rustam, 2006).
Menurut Anonimus (2009), pengelolaan sumberdaya teripang
memerlukan jumlah yang cukup dari pemijah (hewan dewasa yang reproduksinya
sudah matang) dan lingkungan yang sesuai agar setiap tahap dalam daur hidupnya
dapat dilalui dengan baik. Hal tersebut dapat dicapai dengan pengaturan untuk
mengendalikan mortalitas yang terjadi akibat penangkapan dan melalui budidaya.
Untuk kepentingan tersebut, diperlukan informasi mengenai siklus dan pola
reproduksi, serta faktor-faktor lingkungan yang mempengaruhi keduanya.
Pola reproduksi merupakan gambaran tahapan reproduksi pada waktu
tertentu sedangkan siklus reproduksi merupakan pola reproduksi yang terjadi
secara teratur dan berulang seiring dengan putaran waktu. Pola reproduksi dan
siklus reproduksi ditentukan berdasar kajian tahap-tahap kematangan gonad
selama kurun waktu tertentu. Setiap spesies teripang memiliki kekhasan tersendiri
dalam setiap tahap kematangan gonad, baik dalam struktur histologi jantan
maupun betina. Misalnya pada Psolus fabricii yang memiliki ciri khas berupa
oosit mature dengan variasi diameter mulai dari 600 — 800 µm pada tahap
maturationdan tebal dinding gonad yang dipengaruhi oleh diameter lumen, yakni
semakin lebar diameter lumen maka dinding gonad akan semakin tipis (Hamel et
al., 1993). Ciri khas Psolus fabricii ini berbeda dengan Holothuria scabra yang
memiliki dinding gonad yang relatif tipis dengan diameter oosit mature sekitar
110 — 140 µm (Rasolofonirina et al., 2005).
Teripang Phyllophorus sp. atau yang lebih dikenal dengan sebutan
terung merupakan spesies yang dapat ditemukan di Pantai Timur Surabaya dan
memiliki tingkat distribusi tinggi (1,9062) dengan kelimpahan relatif 44,44%.
Selama ini terung hanya dimanfaatkan untuk dibuat makanan ringan berupa
keripik terung, meskipun ada informasi dari beberapa pengepul yang mengatakan
bahwa terung kering merupakan salah satu jenis teripang yang diekspor ke

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 64/97
Taiwan dan Hongkong. Dari pengamatan di lapangan, diketahui bahwa
pengambilan terung oleh nelayan dilakukan seiring dengan penangkapan ikan dan
bahwa pengambilan tidak lagi dilakukan di pinggir pantai, melainkan lebih ke
tengah, kearah laut lepas.
Belum ada informasi mengenai manfaat lain terung, sampai
dilakukannya penelitian oleh Winarni et al. (2010), meskipun sudah banyak
dilakukan penelitian-penelitian teripang spesies lain dengan bioaktivitas sebagai
anti jamur, anti mikroba, sitotoksik dan imunomodulasi (Dang et al., 2007). Di
samping itu sudah banyak pula beredar produk suplemen kesehatan impor
berbahan dasar teripang dari spesies lain. Penelitian Winarni et al.(2010) dengan
menggunakan hewan coba mencit, menunjukkan bahwa terung berpotensi sebagai
imunomodulator yang meningkatkan respons imun tubuh terhadap
Mycobacterium tuberculosis.
Dengan diketahui besarnya potensi terung dan keberadaan di habitat
alaminya, diperlukan informasi tentang pola dan siklus reproduksi untuk
kepentingan pengendalian populasi di alam dan upaya budidaya di waktu yang
akan datang. Penelitian ini dilakukan untuk memberikan informasi tentang
struktur histologi gonad yang dilakukan pada bulan Pebruari, Maret dan April
2012. Dari hasil penelitian ini, paling tidak, dapat diketahui pola reproduksi
terung.
BAHAN DAN CARA KERJA
Bahan
Sampel teripang spesies Phyllophorus sp.(Gambar 1) diperoleh dari
habitat teripang di Pantai Timur Surabaya yang berada pada daerah dengan
kisaran geografis di antara 07º12’48,71” hingga 07º15’54,53” lintang Selatan, dan
di antara 112º48’16,21’ hingga 112º52’34,97’ bujur Timur . Pengambilan sampel
teripang ini dilakukan pada lokasi yang sama pada awal bulan Februari, Maret,
dan April 2012. Identifikasi spesies dilakukan di Laboratorium Ekologi
Departemen Biologi Fakultas Sains dan Teknologi.Pembuatan sediaan histologi
dilakukan di Laboratorium Histologi Hewan, Departemen Biologi, Fakultas Sains

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 65/97
dan Teknologi, Universitas Airlangga. Semua bahan kimia yang digunakan (xylol,
etanol, formaldehid, disodium hidrogen fosfat dan sodium dihidrogen fosfat)
merupakan bahan pro analisa.
Gambar 1. Phyllophorus sp. (terung) di Pantai Timur Surabaya (Winarni et al., 2010)
Cara kerja
Pengambilan sampel teripang dilakukan dengan menggunakan tenaga
bantuan nelayan. Setiap bulan diambil 25 individu, sehingga mendapatkan 75
sampel dalam waktu tiga bulan. Sampel segar yang didapatkan dari nelayan
dibersihkan dari kotoran. Kemudian gonad dikeluarkan dari coeloem, bentuk dan
warna gonad dicatat. Berikutnya, sebanyak 5 tubulus dipotong dari gonad tersebut
dan kemudian difiksasi dalam fiksatif neutral buffered formalin selama sekurang-
kurangnya 24 jam.
Potongan tubulus kemudian diproses menjadi sediaan histologi dengan
metode parafin, dan diwarnai dengan pewarna hematoksilin eosin. Setiap satu
tubulus dibuat 6 potongan seri melintang tubulus dari arah distal, denganketebalan masing-masing irisan 4 m dan jarak antar irisan 40 m. Pengamatan
histologi dilakukan di bawah mikroskop dengan perbesaran 100 kali. Data yang
diamati adalah ukuran dan frekuensi jenis-jenis oosit ( previtellogenic,
vitellogenic, dan post-vitellogenic), diameter lumen tubulus dan tebal dinding
tubulus. Penentuan jenis oosit berdasarkan Purwati (2009). Hubungan antara
diameter lumen dan tebal dinding gonad Phyllophorus sp., dianalisis
menggunakan uji korelasi Pearson pada α = 0,05.

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 66/97
HASIL
Hasil pengamatan morfologi gonad dan histologi gonad menunjukkan
bahwa gonad betina (Gambar 2B) umumnya berwarna hijau lumut dengan bentuk
tubulus yang lebih menggembung dibandingkan dengan gonad jantan (Gambar
2A) yang umumnya berwarna cokelat muda.
Gambar 2. Morfologi gonad Phyllophorus sp. (A) Jantan ; (B) Betina
Pada lumen tubulus gonad betina teripang Phyllophorus sp. ditemukan berbagai
jenis oosit. Berdasarkan Purwati (2009), oosit tersebut dikategorikan menjadi 3, yakni oosit
pre-vitellogenic (diameter < 150 µm), oosit vitellogenic (diameter 150 — 350 µm), dan oosit
post-vitellogenic (diameter > 350 µm) (Gambar 3D,3E. dan 3F). Oosit pre-vitellogenic
terletak melekat di dinding, sangat eosinofilik, tidak ditemukan sel-sel folikuler dan zona
radiata di sekelilingnya, belum memiliki germinal vesicle, nukleus bulat dan berada di
tengah. Sedangkan oosit vitellogenic dengan ukuran yang lebih besar, ada yang terletak di
dekat dinding, tetapi ada juga yang terletak hampir ke tengah lumen, eosinofilik, tidak
ditemukan sel-sel folikuler dan zona radiata di sekelilingnya, belum memiliki germinal
vesicle, nukleus bulat dan berada di tengah.
7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 67/97
Gambar 3.Histologi tubulus gonad jantan (A, B, C) dan betina (D, E, F)
Phyllophorus sp. dg: dinding gonad ; L: lumen ; rs: relict spermatozoa ; sc: spermatosit ; st: spermatid ; sz: spermatozoa ; lf:
longitudinal fold ; ro: oosit relict ; prvo: oosit pre-vitellogenic ; vo:
oosit vitellogenic ; ptvo: oosit post-vitellogenic ; zr: zona radiata ; gv:
germinal vesicle ; n: nukleus ; f: fagosit (Skala : 200 µm).
Sedangkan oosit post-vitellogenic dengan diameter > 350 µm, memiliki
ciri eosinofilik lemah, terletak di tengah lumen, dikelilingi oleh sel folikular dan
zona radiata, terdapat germinal vesicle, nukleus seringkali terletak di tepi
germinal vesicle dan bentuknya memanjang. Berbagai jenis oosit dengan
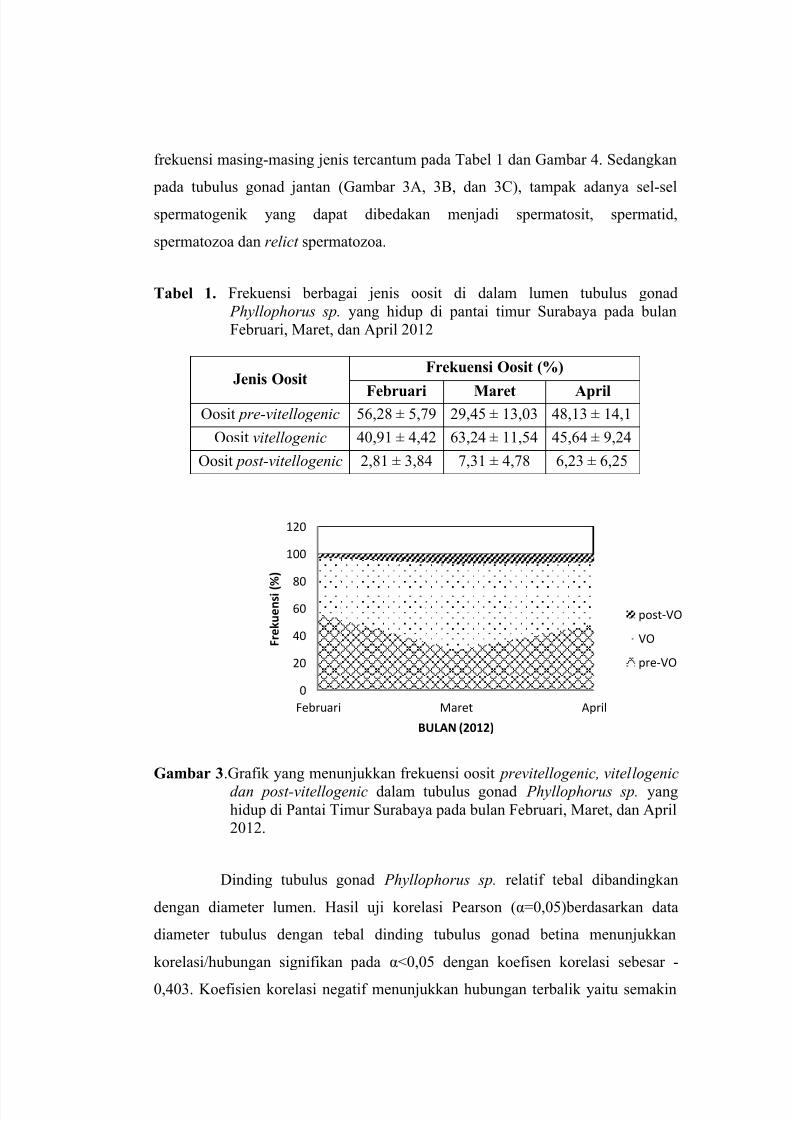
7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 68/97
frekuensi masing-masing jenis tercantum pada Tabel 1 dan Gambar 4. Sedangkan
pada tubulus gonad jantan (Gambar 3A, 3B, dan 3C), tampak adanya sel-sel
spermatogenik yang dapat dibedakan menjadi spermatosit, spermatid,
spermatozoa dan relict spermatozoa.
Tabel 1. Frekuensi berbagai jenis oosit di dalam lumen tubulus gonad
Phyllophorus sp. yang hidup di pantai timur Surabaya pada bulan
Februari, Maret, dan April 2012
Jenis OositFrekuensi Oosit (%)
Februari Maret AprilOosit pre-vitellogenic 56,28 ± 5,79 29,45 ± 13,03 48,13 ± 14,1
Oosit vitellogenic 40,91 ± 4,42 63,24 ± 11,54 45,64 ± 9,24
Oosit post-vitellogenic 2,81 ± 3,84 7,31 ± 4,78 6,23 ± 6,25
Gambar 3.Grafik yang menunjukkan frekuensi oosit previtellogenic, vitellogenic
dan post-vitellogenic dalam tubulus gonad Phyllophorus sp. yanghidup di Pantai Timur Surabaya pada bulan Februari, Maret, dan April
2012.
Dinding tubulus gonad Phyllophorus sp. relatif tebal dibandingkan
dengan diameter lumen. Hasil uji korelasi Pearson (α=0,05)berdasarkan data
diameter tubulus dengan tebal dinding tubulus gonad betina menunjukkan
korelasi/hubungan signifikan pada α<0,05 dengan koefisen korelasi sebesar -
0,403. Koefisien korelasi negatif menunjukkan hubungan terbalik yaitu semakin
0
20
40
60
80
100
120
Februari Maret April
F r e k u e n s i (
% )
BULAN (2012)
post-VO
VO
pre-VO

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 69/97
besar diameter lumen maka semakin tipis dinding tubulus, demikian sebaliknya,
semakin sempit lumen, semakin tebal dinding tubulus.
PEMBAHASAN
Hasil pengamatan menunjukkan bahwa Phyllophorus sp. merupakan
teripang dengan jenis kelamin terpisah (dioecious). Individu jantan dan betina
tidak dapat dibedakan berdasarkan morfologi luar, namun dapat dibedakan
melalui pengamatan bentuk dan warna gonad.
Variasi struktur histologi yang identik dengan variasi tahap kematangan
gonad antar individu, menunjukkan bahwa populasi Phyllophorus sp. memiliki
pola reproduksi asinkron. Populasi yang memiliki pola reproduksi asinkron dapat
memijah sepanjang tahun, dengan puncak pemijahan pada waktu-waktu tertentu
dan dapat bervariasi tergantung pada variasi perubahan lingkungan (Conand,
1993).
Pada pengamatan gonad jantan dan betina Phyllophorus sp.,dinding
tubulus gonad yang relatif tebal dibanding diameter lumen tubulus berbeda
dengan teripang tropis pada umumnya. Pada individu betina ketebalan dinding
gonad bergantung secara signifikan pada jumlah dan jenis oosit yang terdapat
pada lumen dengan kata lain bergantung pada tahap kematangan gonad. Kondisi
ini mirip dengan gonad teripang subtropis Psolus fabricii (Hamel et al ., 1993).
Relict spermatozoa atau spermatozoa sisa, yaitu spermatozoa yang tidak
tersalurkan ke gonoduct, ditemukan pada tubulus gonad jantan (Gambar 3A).
Berdasarkan pengamatan Hamel et al . (1993) pada teripang Psolus fabricii, relict
spermatozoa merupakan ciri bahwa gonad mengalami fase post-spawning dan
umumnya terdapat sel-sel fagosit yang siap mengabsorbsi spermatozoa yang
tersisa tersebut tanpa adanya sel-sel spermatogenik dalam tahap perkembangan
lebih awal seperti spermatosit. Namun pada gonad Phyllophorus sp., keberadaan
relict spermatozoa pada lumen justru disertai dengan kehadiran spermatosit yang
masih melekat pada dinding gonad. Demikian juga pada tubulus lainnya dimana
terdapat spermatozoa yang hampir memenuhi lumen dan secara bersamaan
terdapat spermatid yang masih melekat di dinding gonad. Berdasarkan Hamel et

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 70/97
al . (1993), keberadaan spermatid dan spermatozoa merupakan ciri bahwa gonad
Psolus fabricii sedang mengalami fase maturation. Umumnya dinding tubulus
gonad sudah mengalami penipisan akibat lumen berdilatasi karena jumlah
spermatozoa terus melimpah. Namun pada gonad Phyllophorus sp. dinding gonad
masih cukup tebal meskipun terdapat spermatid dan spermatozoa didalamnya.
Dinding tubulus gonad Phyllophorus sp. baru akan benar-benar menipis dan
lumen mengalami dilatasi saat lumen hanya dipenuhi oleh spermatozoa. Namun,
tidak pernah benar-benar dijumpai spermatozoa memenuhi seluruh isi tubulus
dalam satu gonad jantan karena pasti sudah dijumpai keberadaan spermatid.
Seperti halnya relict spermatozoa pada gonad jantan Phyllophorus sp.,
pada gonad betina juga terdapat oosit relict , yakni merupakan oosit post-
vitellogenic yang tidak dilepaskan ke gonoduct. Berdasarkan Hamel et al . (1993),
relict oosit merupakan ciri bahwa gonad Psolus fabricii mengalami fase post-
spawning dan umumnya terdapat sel-sel fagosit yang siap mengabsorbsi oosit
yang tersisa tersebut. Namun pada gonad Phyllophorus sp., keberadaan relict
oosit selain disertai dengan keberadaan fagosit, juga disertai dengan keberadaan
oosit pre-vitellogenic. Berdasarkan Hamel et al . (1993), mengacu pada gonad
Psolus fabricii,oosit pre-vitellogenic merupakan ciri bahwa gonad sedang
mengalami fase recovery, oosit vitellogenic merupakan ciri bahwa gonad sedang
dalam fase advanced growth, dan oosit post-vitellogenic merupakan ciri gonad
sudah dalam fase maturation.Akan tetapi, pada gonad betina Phyllophorus sp.
tidak pernah benar-benar ditemukan hanya satu jenis oosit yang memenuhi lumen
karena ketiga jenis oosit tersebut sering terlihat bersamaan dalam satu tubulus.
Nampaknya, tahap-tahap kematangan gonad yang umum ditemukan
pada teripang spesies lain, pada Phyllophorus sp. ini saling tumpang-tindih. Hal
ini memungkinkan Phyllophorus sp. yang memiliki pola reproduksi asinkron
dapat memijah sepanjang tahun dengan waktu antar terjadinya puncak pemijahan
lebih singkat dari spesies lain. Jika benar demikian, dimungkinkan pula bahwa
keberadaan teripang di pantai timur Surabaya dengan kelimpahan relatif tertinggi
dibanding spesies teripang lain (Winarni et al ., 2010), disebabkan oleh faktor
tersebut, meskipun untuk sampai pada kesimpulan tersebut, memerlukan data

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 71/97
yang diperoleh dari bulan-bulan berbeda dengan lama waktu paling tidak selama 1
tahun. Kajian histologi pada irisan memanjang tubulus juga diperlukan untuk
mengetahui distribusi berbagai jenis sel-sel gametogenik dalam tubulus.
KEPUSTAKAAN
Anonimus, 2009. Laporan Ilmiah Institut Pertanian Bogor .
http://repository.ipb.ac.id/bitstream/handle/123456789/5912/DaftarPustak
a_2009yal-7.pdf?sequence=12. Diunduh pada 22 November 2011.
Conand, C., 1993. Reproductive Biology Of The Holothurians From The Major
Communities Of The New Caledonian Lagoon. Journal of Marine
Biology. Springer Verlag. 116: 439-450.
Dang, N.H., N.V. Thanh, P.V. Kiem, L.M. Huong, C.V. Minh and Y.H.Kim.2007. Two New Triterpene Glycosides friom Vietnamese Sea
Cucumber Holothuria scabra. Archives of Pharmacal Research. Vol.30.
no.11. p. 1387-1391.
Dewi, K. H., Silsia, D., Susanti, L., Markom, M., dan Mendra, H. 2010.Ekstraksi
Teripang Pasir ( Holothuria scabra) sebagai Sumber Testosteron pada
Berbagai Kecepatan dan Lama Pengadukan. Prosiding Seminar Nasional
Teknik Kimia “Kejuangan” Pengembangan Teknologi Kimia untuk
Pengolahan Sumber Daya Alam Indonesia. ISSN 1693 – 4393.
Hamel, J. F., Himmelman, J. H., dan Dufresne, L. 1993. Gametogenesis and
Spawning of the Sea Cucumber Psolus fabricii (Duben and Koren). Biol
Bull. 184:125-143.
Purwati, P. 2009. Oocytes of Holothuria leucospilota (Echinodermata :
Holothuroidea) : An Ultra Structural Study. Journal of Oceanology. Mar.
Res. Indonesia 34 : 17-26.
Rasolofonirina, R., Devarajen V., Igor E., dan Michel J. 2005. Reproductive
Cycle of Edible Echinoderms from the South-Western Indian Ocean.
Journal Western Indian Ocean. Mar. Sci. 4 (1):61-75.
Rustam.2006. Pelatihan Budidaya Laut (Coremap Fase II Kabupaten Selayar),
Budidaya Teripang. Pelatihan Budidaya Laut Coremap Tahap II
Kabupaten Selayar, Yayasan Mattirotasi.
Winarni, D., Affandi, M., Masithoh, E. D. dan Kristanti, A. N. 2010. PotensiTeripang Pantai Timur Surabaya Sebagai Modulator Imunitas Alami
Terhadap Mycobacterium tuberculosis. Laporan Akhir Penelitian Strategis
Nasional Batch II . Universitas Airlangga : Surabaya.

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 72/97
PENGARUH PEMBERIAN KONSORSIUM MIKROBA BIOFERTILIZER
TERHADAP PERTUMBUHAN DAN PRODUKSI JAMUR TIRAM PUTIH
(Pleorotus ostreatus )
Rizka Rakhmawati, Tini Surtiningsih, Tri Nurhariyati
Program Studi S1 Biologi, Fakultas Sains dan Teknologi, Universitas Airlangga
ABSTRACT
This research was aimed to know the effect of the use consortium of
biofertilizer microbes with different dosage to growth and production of white
oyster
mushroom (Pleorotus ostreatus). The growth parameter is the number of
pileus and the production parameter is the weight of white oyster
mushroom. This
study was conducted in completely randomized design with six treatments, D0-
(without biofertilizer as a control (-)), D0+ (with NPK as a control (+)), D2 (with 2
ml biofertilizer in each baglog), D4 (with 4 ml biofertilizer in each baglog), D6
(with 6 ml biofertilizer in each baglog), dan D8 (with 8 ml biofertilizer in each
baglog), each treatment have seven replications. Data from harvest time was
analized using ANOVA and continued by Duncan test using α = 0,05. The result
of this study showed that the gift of consortium of biofertilizer microbes affect the production but it didn’t affect the growth. The best result to the weight is D6
(128,43 ± 37,66 gram).
Key words : white oyster mushroom, biofertilizer, dosage, growth, production
PENGANTAR
Indonesia merupakan negara agraris dan sektor pertanian menjadi salah
satu sektor penting dalam mendukung perekonomian, sehingga bidang pertanian
harus terus dipacu untuk dapat meningkatkan hasil. Salah satu jenis pertanian
yang menarik untuk dikembangkan di kalangan masyarakat saat ini adalah budi
daya jamur. Kandungan gizi yang cukup baik dari jamur dan kebutuhan konsumen
yang cenderung meningkat menyebabkan usaha budi daya jamur memiliki
prospek yang bagus untuk dikembangkan. Salah satu jenis jamur yang banyak
dikembangkan di Indonesia adalah jamur tiram putih.
Jamur tiram putih mempunyai manfaat bagi kesehatan manusia, protein
nabati yang dikandungnya tidak mengandung kolesterol sehingga dapat mencegah

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 73/97
timbulnya penyakit darah tinggi dan jantung serta untuk mengurangi berat badan
dan diabetes. Kandungan asam folat (vitamin B komplek) yang tinggi dapat
menyembuhkan anemia dan obat antitumor. Jamur tiram putih dapat digunakan
untuk mencegah dari kekurangan gizi dan pengobatan kekurangan zat besi
(Pasaribu, 2002).
Seiring dengan semakin populer dan bertambahnya pengetahuan
masyarakat mengenai jamur tiram putih sebagai bahan makanan yang lezat dan
bergizi, maka permintaan konsumen dan permintaan pasar terhadap jamur tiram
putih terus meningkat. Namun kenaikan permintaan konsumen tersebut tidak
diikuti dengan peningkatan produksi jamur tiram putih. Menurut Wardani (2010),
budi daya jamur konsumsi masih jarang ditemui di sebagian besar wilayah
Indonesia sehingga kebutuhan konsumen tidak terpenuhi. Pada beberapa tempat
budi daya jamur tiram putih yang ada di wilayah Indonesia, petani jamur tiram
putih masih menggunakan pupuk kimia untuk memacu pertumbuhan dan
meningkatkan produksi.
Alasan masyarakat memilih jamur tiram putih untuk dikonsumsi ialah
karena ingin hidup sehat tanpa bahan kimia. Jadi jika diketahui bahwa dalam
pengelolahan jamur tiram putih masih digunakan bahan kimia, maka itu dapat
menjadi faktor pengurang minat bagi masyarakat untuk mengonsumsi jamur tiram
putih. Sosialisasi untuk mengganti penggunaan pupuk kimia dengan beralih pada
pupuk organik dan biofertilizer pun mulai digencarkan.
Sel-sel jamur tiram memerlukan karbon berantai enam (C6) untuk
pertumbuhan. Kebutuhan karbon dapat terpenuhi dengan mengurai media tumbuh
jamur (baglog). Media tumbuh jamur tiram putih yang berupa baglog memiliki
penyusun utama yaitu serbuk gergaji kayu, bekatul, dan tepung jagung. Secara
umum, kayu mengandung selulosa, hemiselulosa, lignin, pentosan dan
sebagainya. Unsur-unsur tersebut terdapat pada dinding sel kayu. Bagian yang
terbesar adalah selulosa (Anonimus, 2011). Selulosa merupakan komponen utama
penyusun dinding sel tanaman dan hampir tidak pernah ditemui dalam keadaan
murni di alam, melainkan berikatan dengan bahan lain, yaitu lignin dan
hemiselulosa membentuk suatu lignoselulosa. Mikroba memproduksi enzim

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 74/97
ekstraselular untuk depolimerasi senyawa berukuran besar menjadi berukuran
kecil dan larut dalam air (Saraswati dkk.,2011) sehingga kebutuhan nutrisi jamur
tiram putih dapat terpenuhi dengan baik.
BAHAN DAN METODE
Bahan yang digunakan dalam penelitian ini antara lain bakteri
Cellulomonas sp., Bacillus megaterium, Azospirillum sp., dan yeast
Saccharomyces cereviceae serta media pertumbuhan mikroba berupa NA, PDA,
NB, SDB, NB + glukosa 1% dan molase 2% serta media selektif yang merupakan
koleksi dari Laboratorium Mikrobiologi, Fakultas Sains dan Teknologi,
Universitas Airlangga. Media tanam baglog dan bibit jamur tiram putih var.
Oystern yang diperoleh dari Desa Pepe, Sidoarjo.
Penelitian ini merupakan penelitian eksperimental dengan rancangan acak
lengkap yang terdiri dari enam perlakuan (D0-, D0
+, D2, D4, D6 dan D8) dan setiap
perlakuan di ulang sebanyak tujuh kali. D0- merupakan kontrol negatif (tanpa
konsorsium mikroba biofertilizer ), D0+ merupakan kontrol positif (tanpa
konsorsium mikroba biofertilizer , namun dengan NPK 2 gram/baglog), D2 (biofertilizer 2 ml/baglog), D4 (biofertilizer 4 ml/baglog), D6 (biofertilizer 6
ml/baglog), dan D8 (biofertilizer 8 ml/baglog). Variabel terikat pada penelitian ini
yaitu pertumbuhan yang dapat dilihat dari jumlah tudung (buah) serta produksi
yang dilihat dari total berat basah (gram) jamur tiram putih saat panen.
Pembuatan konsorsium mikroba biofert il izer
Membuat media pertumbuhan mikroba, yaitu Nutrien Agar (NA) untuk
bakteri dan Potato Dextrose Agar (PDA) untuk yeast, kemudian membiakkan
mikroba dalam media tersebut. Setelah masa inkubasi dalam media padat,
mikroba dibiakkan dalam media NB (untuk bakteri) dan media SDB (untuk
yeast). Kemudian dibuat stok mikroba dengan cara mengambil 10 ml mikroba
dalam media Broth yang dimasukkan ke dalam media NB + glukosa 1% dan
dilakukan inkubasi selama dua hari. Melakukan penghitungan jumlah koloni
mikroba pada media NB + glukosa 1% dengan metode TPC untuk mengetahui
jumlah sel apakah sudah memenuhi baku mutu biofertilizer (total koloni 106 untuk

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 75/97
bakteri dan 104 untuk yeast). Stok mikroba yang telah diketahui jumlah sel
dimasukkan dalam larutan molase 2%. Setelah proses inkubasi dalam larutan
molase 2% selama dua hari, dilakukan uji keberadaan mikroba pada media
selektif.
Pembudidayaan jamur tiram putih
Pembudidayaan jamur tiram putih yang dilakukan dalam penelitian ini
antara lain persiapan media baglog, inokulasi bibit serta perawatan pada masa
inkubasi dan masa penumbuhan hingga panen. Bibit diinokulasikan ke dalam
baglog dengan kondisi yang steril pada suhu 230C - 25
0C
Selain itu perlu dilakukan Proses perawatan dan pemeliharaan jamur
tiram putih, antara lain melepas cincin baglog agar tubuh buah jamur tumbuh
semakin mudah dan cepat karena luasnya area tumbuh dan melakukan pengabutan
atau penyiraman (kelembaban kumbung harus dijaga minimal 80%).
HASIL DAN PEMBAHASAN
Pengaruh pemberian konsorsium mikroba biofertilizer terhadap
pertumbuhan jamur tiram putih
Proses pertumbuhan jamur tiram putih diawali dengan pembentukan
miselium oleh spora setelah inokulasi bibit jamur (F2) pada media tanam serbuk
gergaji kayu (baglog). Setelah miselium memenuhi seluruh pemukaan baglog,
akan muncul pin head yang kemudian menjadi badan buah jamur siap panen. Hal
yang harus diperhatikan pada saat proses pemanenan adalah pengambilan seluruh
bagian tubuh buah jamur tiram putih beserta bagian stalk yang menembus baglog
media tanam (Elsera,2009). Pada penelitian ini, proses pertumbuhan miselium
memerlukan waktu ± 30 hari.
Dari hasil uji ANOVA dengan derajad signifikan 0,05 (α = 0,05) untuk
pertumbuhan jamur tiram putih diketahui nilai signifikan pada perlakuan
konsorsium mikroba biofertilizer terhadap pertumbuhan jamur tiram putih ≥ 0,05
yaitu sebesar 0,370. Karena nilai signifikan yang diperoleh lebih besar dari
derajad signifikan yang ditetapkan (α = 0,05), maka keputusan yang diambil
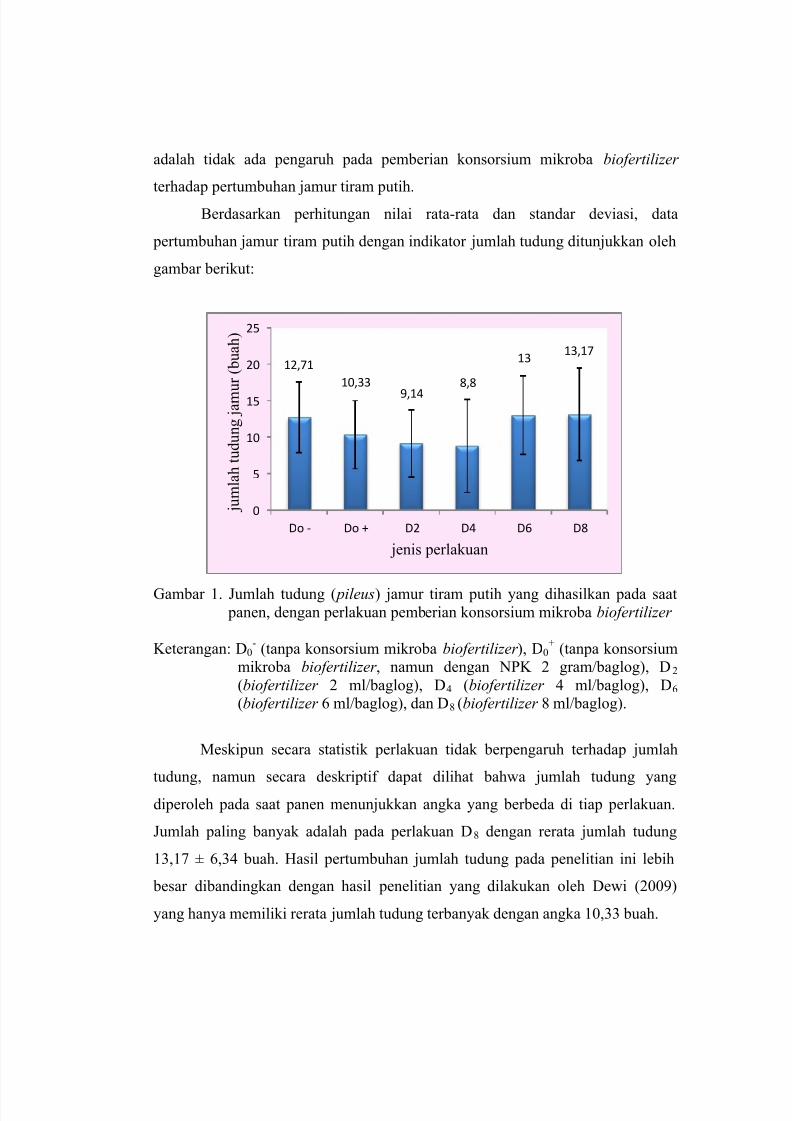
7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 76/97
adalah tidak ada pengaruh pada pemberian konsorsium mikroba biofertilizer
terhadap pertumbuhan jamur tiram putih.
Berdasarkan perhitungan nilai rata-rata dan standar deviasi, data
pertumbuhan jamur tiram putih dengan indikator jumlah tudung ditunjukkan oleh
gambar berikut:
Gambar 1. Jumlah tudung ( pileus) jamur tiram putih yang dihasilkan pada saat panen, dengan perlakuan pemberian konsorsium mikroba biofertilizer
Keterangan: D0- (tanpa konsorsium mikroba biofertilizer ), D0
+ (tanpa konsorsium
mikroba biofertilizer , namun dengan NPK 2 gram/baglog), D2
(biofertilizer 2 ml/baglog), D4 (biofertilizer 4 ml/baglog), D6
(biofertilizer 6 ml/baglog), dan D8 (biofertilizer 8 ml/baglog).
Meskipun secara statistik perlakuan tidak berpengaruh terhadap jumlah
tudung, namun secara deskriptif dapat dilihat bahwa jumlah tudung yang
diperoleh pada saat panen menunjukkan angka yang berbeda di tiap perlakuan.
Jumlah paling banyak adalah pada perlakuan D8 dengan rerata jumlah tudung
13,17 ± 6,34 buah. Hasil pertumbuhan jumlah tudung pada penelitian ini lebih
besar dibandingkan dengan hasil penelitian yang dilakukan oleh Dewi (2009)
yang hanya memiliki rerata jumlah tudung terbanyak dengan angka 10,33 buah.
12,71
10,33
9,14
8,8
1313,17
0
5
10
15
20
25
Do - Do + D2 D4 D6 D8
j u m l a h t u d u n g j a m u r ( b u a h )
jenis perlakuan

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 77/97
Pengaruh pemberian konsorsium mikroba biofertilizer terhadap produksi
jamur tiram putih
Pada penelitian ini, produksi jamur tiram putih yang diukur adalah berat
basah tubuh buah yang dihasilkan ketika proses pemanenan. Dari hasil ANOVA
dengan derajad signifikan 0,05 (α = 0,05) untuk produksi jamur tiram putih
diketahui nilai signifikan pada perlakuan konsorsium mikroba biofertilizer
terhadap produksi jamur tiram putih ≤ 0,05 yaitu sebesar 0,013. Karena nilai
signifikan yang diperoleh lebih kecil dari derajad signifikan yang ditetapkan (α =
0,05), maka keputusan yang diambil adalah ada pengaruh pada pemberian
konsorsium mikroba biofertilizer terhadap produksi jamur tiram sehingga dapat
dilanjutkan dengan uji Duncan dengan derajad signifikan 0,05 (α = 0,05).
Tabel 1. Pengaruh perlakuan konsorsium mikroba biofertilizer dengan dosis yang
berbeda terhadap produksi jamur tiram putih pasca panen
No. Jenis perlakuan
(dosis)
berat basah
(gram)
1 D0- 71,86 ± 41,20 a
2 D0+ 66,67 ± 25,65 a3 D2 60,57 ± 22,86 a
4 D4 103,4 ± 50,18 ab
5 D6 128,43 ± 37,66 b
6 D8 99,0 ± 56,46 ab
Keterangan: Huruf yang berbeda di belakang angka menunjukkan perbedaan
signifikan berdasarkan hasil uji Duncan
Tabel 1. di atas menunjukkan bahwa perlakuan D6 memberi beda nyata
dengan perlakuan D2 maupun kontrol dan menghasilkan berat yang terbesar
dengan nilai 128,43 ± 37,66 gram. Data produksi jamur tiram putih disajikan oleh
gambar berikut:
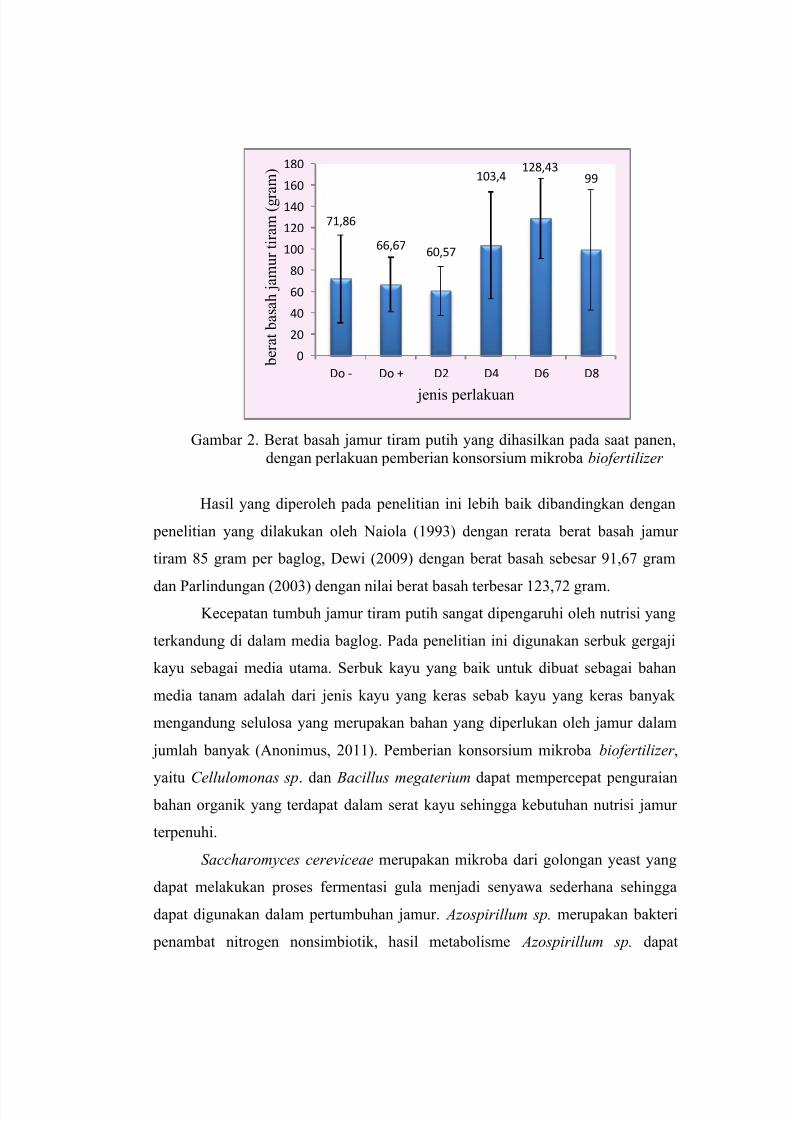
7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 78/97
Gambar 2. Berat basah jamur tiram putih yang dihasilkan pada saat panen,
dengan perlakuan pemberian konsorsium mikroba biofertilizer
Hasil yang diperoleh pada penelitian ini lebih baik dibandingkan dengan
penelitian yang dilakukan oleh Naiola (1993) dengan rerata berat basah jamur
tiram 85 gram per baglog, Dewi (2009) dengan berat basah sebesar 91,67 gram
dan Parlindungan (2003) dengan nilai berat basah terbesar 123,72 gram.Kecepatan tumbuh jamur tiram putih sangat dipengaruhi oleh nutrisi yang
terkandung di dalam media baglog. Pada penelitian ini digunakan serbuk gergaji
kayu sebagai media utama. Serbuk kayu yang baik untuk dibuat sebagai bahan
media tanam adalah dari jenis kayu yang keras sebab kayu yang keras banyak
mengandung selulosa yang merupakan bahan yang diperlukan oleh jamur dalam
jumlah banyak (Anonimus, 2011). Pemberian konsorsium mikroba biofertilizer ,
yaitu Cellulomonas sp. dan Bacillus megaterium dapat mempercepat penguraian bahan organik yang terdapat dalam serat kayu sehingga kebutuhan nutrisi jamur
terpenuhi.
Saccharomyces cereviceae merupakan mikroba dari golongan yeast yang
dapat melakukan proses fermentasi gula menjadi senyawa sederhana sehingga
dapat digunakan dalam pertumbuhan jamur. Azospirillum sp. merupakan bakteri
penambat nitrogen nonsimbiotik, hasil metabolisme Azospirillum sp. dapat
71,86
66,6760,57
103,4128,43
99
0
20
40
60
80
100
120
140
160
180
Do - Do + D2 D4 D6 D8 b e r a t b a s a h j a m u r t i r a m ( g r
a m )
jenis perlakuan

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 79/97
menyediakan unsur nitrogen yang dibutuhkan dalam pertumbuhan dan produksi
jamur tiram putih.
KESIMPULAN
Pemberian konsorsium mikroba biofertilizer dengan dosis yang berbeda
tidak berpengaruh terhadap pertumbuhan (jumlah tudung) tetapi berpengaruh
terhadap produksi (berat basah) jamur tiram putih ( Pleoratus ostreatus).
Perlakuan memberikan hasil paling baik terhadap berat basah pada D6 dengan
128,43 ± 37,66 gram.
DAFTAR PUSTAKA
Anonimus, 2011, Budi Daya Jamur Tiram dan Jamur Kuping ,
bisnisjamurtiram.blogspot.com,usahajamur.co.cc, Diakses pada hari
Kamis, 8 Desember 2011.
Dewi, I. K., 2009, Efektifitas Pemberian Blotong Kering terhadap Pertumbuhan
Jamur Tiram Putih (Pleorotus ostreatus) pada Media Serbuk Gergaji Kayu, Skripsi, Fakultas Keguruan dan Ilmu Pendidikan Universitas
Muhammadiyah Surakarta.
Elsera, L., 2009, Risiko Produksi Jamur Tiram Putih pada Usaha Cempaka Baru
di Kecamatan Cisarua Kabupaten Bogor, Skripsi, Departemen Agribisnis
Institut Pertanian Bogor.
Naiola, E., 1993, Budi Daya Jamur Merang dan Jamur Tiram Putih pada
Pekarangan di Daerah DAS Cisadane, Balitbang Mikrobiologi, Puslitbang
Biologi, LIPI.
Parlindungan, A. K., 2003, Karakteristik Pertumbuhan dan Produksi Jamur
Tiram Putih (Pleorotus ostreatus) dan Jamur Tiram Kelabu (Pleorotus
sajor Caju) pada baglog alang-alang , Jurnal Natur Indonesia 5(2): 152-
156 (2003).

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 80/97
Pasaribu, T., 2002, Aneka Jamur Unggulan yang Menembus Pasar , PT Gramedia,
Jakarta.
Saraswati, R., Edi Santoso, Erny Yuniarti, 2011, Organisme Perombak Bahan
Organik . Pupuk organik dan pupuk hayati, 211-230.
Wardani, Isnaeni, 2010, Budi Daya Jamur Konsumsi, Penerbit ANDI,
Yogyakarta.

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 81/97
1
Kajian Viabilitas dan Pola Pertumbuhan Lactobacil lus plantarum pada Variasi
Konsentrasi Molase dan Waktu Inkubasi
Agus Supriyanto, Rochma Novirisandi, Ni’matuzahroh
Program Studi S1 Biologi, Departemen BiologiFakultas Sains dan Teknologi, Universitas Airlangga
ABSTRACT
This study was aim to determine the growth pattern of Lactobacillus plantarum on the variation concentration of molasses and the incubation time, the
influence combination of molasses concentration and incubation time on total of
Lactobacillus plantarum cells and to know viability of Lactobacillus plantarum inlast incubation on variation concentration of molasses. This study used a factorial
design 4x13 with 3 replications. Growth medium consisted of molasses of different
concentrations (0%, 1%, 2%, 3%) with 2% starter bacteria Lactobacillus plantarum and was incubated for 12 weeks. Observational data in the form of total bacteria
(CFU/mL) were then analyzed in a descriptively and statistically using Brown
Forsithe Test , followed by Gomes-Howell Test . The descriptive results were analyzed
showed that Lactobacillus plantarum had no lag phase in growth patterns and growth
patterns of Lactobacillus plantarum on molasses concentration 1%, 2% and 3% havethe same phase. The log phase of growth pattern was toward the stationary phase
followed by a deceleration phase, while the statistic results were analyzed showedthat a combination of molasses concentration and incubation time gave an effect on
the total of Lactobacillus plantarum cells. The combination of molasses
concentration at 2% for 12 weeks incubation period was a highest combinationamong the other combination. The total of Lactobacillus plantarum cells on molasses
concentration 1%, 2% and 3% at the end of incubation period was 107 (CFU/mL) so
the viability of Lactobacillus plantarum is still good.
Keyword :, Incubation, Lactobacillus plantarum, molasses concentration, total of
Lactobacillus plantarum cells

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 82/97
2
PENDAHULUAN
Masalah pada lingkungan perairan terutama adanya akumulasi zat-zat organik
yang kadarnya berlebih dan melampaui kadar yang seharusnya ditetapkan untuk
kategori perairan dapat mencemari perairan. Sebagai akibatnya ikan yang
dibudidayakan pada perairan akan tercemar bahan organik. Adanya bahan organik
yang ada pada tubuh ikan akan memicu tumbuhnya bakteri patogen di dalam tubuh
ikan pada bagian sistem pencernaannya (Soeharsono, 2010).
Semua permasalahan ini dapat diatasi dengan teknik akuakultur yang akan
memanfaatkan kerja bakteri probiotik untuk menghambat bahkan membunuh bakteri
patogen. Salah satu bakteri probiotik yang berperan dalam menghambat dan
membunuh pertumbuhan bakteri patogen pada sistem pencernaan ikan adalah
Lactobacillus plantarum (Surono, 2004).
Adanya bakteri probiotik pada sistem akuakultur akan menjadi penetralisir
bagi sistem pencernaan pada tubuh ikan sehingga bakteri patogen yang ada pada
sistem pencernaan ikan dapat dihambat pertumbuhannya. Hal ini dapat mendukung
pertumbuhan ikan yang cepat pada suatu budidaya perairan sehingga dapat menekan
pengeluaran untuk pakan, mempercepat masa panen dan ikan bisa dipanen dalam
ukuran yang seimbang karena terjaganya struktur organoleptik pada ikan (Emanuel,
et al ., 2005).
Untuk mendukung pertumbuhan bakteri probiotik ini diperlukan suatu
substrat yang dapat menstimulasi bakteri probiotik untuk menghasilkan suatu
metabolit yang nantinya akan berperan sebagai penghambat dan pembunuh bakteri
patogen. Mekanisme pencampuran bakteri probiotik pada sistem akuakultur
membutuhkan suatu media yang dapat mendukung metabolisme bakteri probiotik .
Salah satu media yang biasanya digunakan yaitu molase. Molase merupakan
media pembawa yang selama ini sudah dikenal sebagai substrat pertumbuhan bakteri.
Molase dipilih karena efektifitas dan efisiensinya sebagai bahan pembawa untuk

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 83/97
3
mengontrol pertumbuhan bakteri sudah diketahui tingkat keberhasilannya terhadap
hasil penelitian sebelumnya cukup signifikan (Murdiyatmo, 2003).
Dalam penelitian ini akan digunakan variasi konsentrasi molase yang berbeda
dengan kadar di bawah 5% yaitu 1, 2, dan 3% sebagai formulasi perlakuan.
Penggunaan kadar molase yang relatif rendah memiliki alasan tertentu yaitu karena
bentuk molase yang kental akan meningkatkan tekanan osmose pada substrat
sehingga kurang efisien untuk pertumbuhan bakteri.
Berdasarkan latar belakang diatas perlu diadakan penelitian terhadap pola
pertumbuhan dan viabilitas bakteri Lactobacillus plantarum pada beberapa
konsentrasi molase serta lamanya waktu inkubasi sehingga penggunaan bakteri
probiotik ini diharapkan mampu mengatasi permasalahan rusaknya struktur
organoleptik pada ikan sehingga akan menghambat proses pembusukan pada ikan
dikarenakan pertumbuhan bakteri patogen pada usus ikan serta mampu meningkatkan
produktivitas tambak dalam rangka mewujudkan target pemerintah dalam
meningkatkan produksi produk perikanan.
BAHAN DAN METODE
Penelitian dilakukan di Laboratorium Mikrobiologi Umum, Departemen
Biologi, Fakultas Sains dan Teknologi, Universitas Airlangga, Surabaya. Waktu
pelaksanaan penelitian 4 bulan (Februari-Mei 2012).
Isolat Lactobacillus plantarum dan media molase berasal dari hasil koleksi
Laboratorium Mikrobiologi Umum, Departemen Biologi, Fakultas Sains dan
Teknologi, Universitas Airlangga, Surabaya. Sedangkan bahan kimia yang digunakan
dalam penelitian adalah media MRS broth, MRS agar, aquades, alkohol dan molase.
Isolat Lactobacillus plantarum diperbanyak dengan cara metode streak
kemudian diinokulasikan pada media MRS broth dan diinkubasi selama 24 jam.
Setelah 24 jam masa inkubasi biakan kultur Lactobacillus plantarum pada MRS broth

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 84/97
4
diukur nilai kekeruhannya untuk menentukan starter bakteri yang digunakan untuk
penelitian ini. Jika nilai kekeruhan menunjukkan angka 0,5 maka kultur Lactobacillus
plantarum siap untuk dijadikan kultur perlakuan.
Penelitian disusun berdasarkan Rancangan Faktorial (RF) yang terdiri dari 4
perlakuan (tanpa molase, molase 1%, molase 2%, molase 3%) dengan 3 kali
pengulangan untuk masing-masing perlakuan. Pengambilan data dilakukan selama 12
minggu lalu dianalisis secara deskriptif dan statistik. Analisis data secara statistik
menggunakan uji Kolmogorov-sminorv dan homogenitas datanya diuji dengan uji
Levane. Kemudian menggunakan uji lanjutan Brown-Forsithe untuk mengetahui
apakah ada pengaruh antar perlakuan dengan menggunakan derajat signifikansi 5%.
Setelah itu dilanjutkan dengan uji Gomes-Howell untuk mengetahui beda nyata
terkecil antar perlakuan.
Parameter yang diamati dalam penelitian ini adalah jumlah sel Lactobacillus
plantarum. Penghitungan jumlah sel Lactobacillus plantarum dilakukan dengan
metode cawan tuang dalam CFU/mL.
HASIL DAN PEMBAHASAN
Dari hasil penelitian, pengamatan terhadap jumlah sel Lactobacillus
plantarum dengan perbedaaan periode kultur dan komposisi media dimaksudkan
untuk mengetahui pengaruhnya terhadap jumlah sel Lactobacillus plantarum. Setelah
itu dibuat kurva pertumbuhan bakteri serta dianalisis fase pertumbuhannya,
kombinasi antar perlakuan terhadap viabilitas bakteri ( Lactobacillus plantarum) dan
viabilitas sel Lactobacillus plantarum di akhir inkubasi.
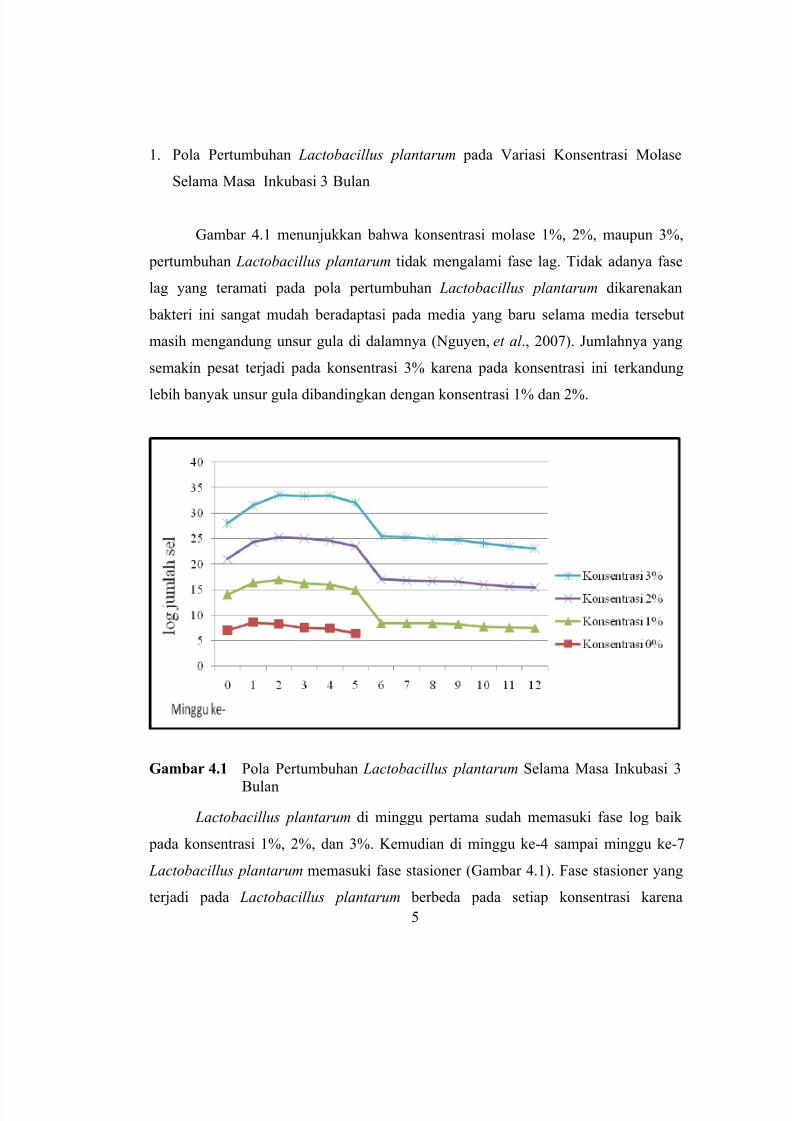
7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 85/97
5
1. Pola Pertumbuhan Lactobacillus plantarum pada Variasi Konsentrasi Molase
Selama Masa Inkubasi 3 Bulan
Gambar 4.1 menunjukkan bahwa konsentrasi molase 1%, 2%, maupun 3%,
pertumbuhan Lactobacillus plantarum tidak mengalami fase lag. Tidak adanya fase
lag yang teramati pada pola pertumbuhan Lactobacillus plantarum dikarenakan
bakteri ini sangat mudah beradaptasi pada media yang baru selama media tersebut
masih mengandung unsur gula di dalamnya (Nguyen, et al ., 2007). Jumlahnya yang
semakin pesat terjadi pada konsentrasi 3% karena pada konsentrasi ini terkandung
lebih banyak unsur gula dibandingkan dengan konsentrasi 1% dan 2%.
Gambar 4.1 Pola Pertumbuhan Lactobacillus plantarum Selama Masa Inkubasi 3
Bulan
Lactobacillus plantarum di minggu pertama sudah memasuki fase log baik
pada konsentrasi 1%, 2%, dan 3%. Kemudian di minggu ke-4 sampai minggu ke-7
Lactobacillus plantarum memasuki fase stasioner (Gambar 4.1). Fase stasioner yang
terjadi pada Lactobacillus plantarum berbeda pada setiap konsentrasi karena

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 86/97
6
kandungan nutrisi yang tersedia pada masing-masing konsentrasi juga berbeda
(Schlegel dan Schmidt, 1994). Pada umumnya fase stasioner ini terjadi karena
penumpukan racun akibat metabolisme sel dan kandungan nutrisi mulai habis
sehingga terjadi kompetisi akibatnya beberapa sel akan mati dan jumlah sel bakteri
cenderung konstan. Hal ini sesuai dengan pernyataan Suriawiria (1995).
Kemudian dari minggu ke-7 sampai minggu ke-12 merupakan fase
perlambatan (kematian) dari Lactobacillus plantarum (Gambar 4.1). Fase
perlambatan (kematian) terjadi karena penumpukan racun yang semakin banyak
sedangkan nutrisi yang tersedia semakin habis menyebabkan jumlah sel bakteri yang
mati semakin banyak dibandingkan dengan jumlah sel bakteri yang hidup sehingga
jumlah sel bakteri menurun secara eksponensial (Waluyo, 2004).
Jika dikaitkan dengan pengukuran pH, maka dapat diketahui bahwa pada saat
awal inkubasi pH yang berkisar antara 5-6 berubah menjadi kisaran pH 2,5-4 dalam 2
minggu inkubasi. Ini berarti bahwa Lactobacillus plantarum mengeluarkan metabolit
sekunder yaitu asam laktat yang merubah pH media menjadi asam sehingga
menghambat pertumbuhan bakteri patogen, hal ini sesuai dengan penelitian Jeni dan
Rini (1995). Kisaran pH ini tidak berubah sampai akhir inkubasi. Penurunan nilai pH
tidak berpengaruh pada penelitian ini karena nilai pH terendah tidak dikuti dengan
jumlah sel terbanyak begitu juga sebaliknya.
2. Pengaruh Kombinasi Variasi Konsentrasi Molase dan Waktu Inkubasi terhadap
Jumlah Sel Lactobacillus plantarum
Untuk mengetahui ada/tidaknya pengaruh pada kombinasi variasi konsentrasi
molase dan waktu inkubasi terhadap jumlah sel Lactobacillus plantarum dapatdianalisis secara statistik menggunakan uji normalitas dan homogenitas data. Uji
normalitas data dengan uji KS menunjukkan bahwa data berdistribusi normal karena
nilai > 0,05. sedangkan uji homogenitas data mengggunakan uji levane
menunjukkan data tidak homogen. Karena data normal dan tidak homogen maka
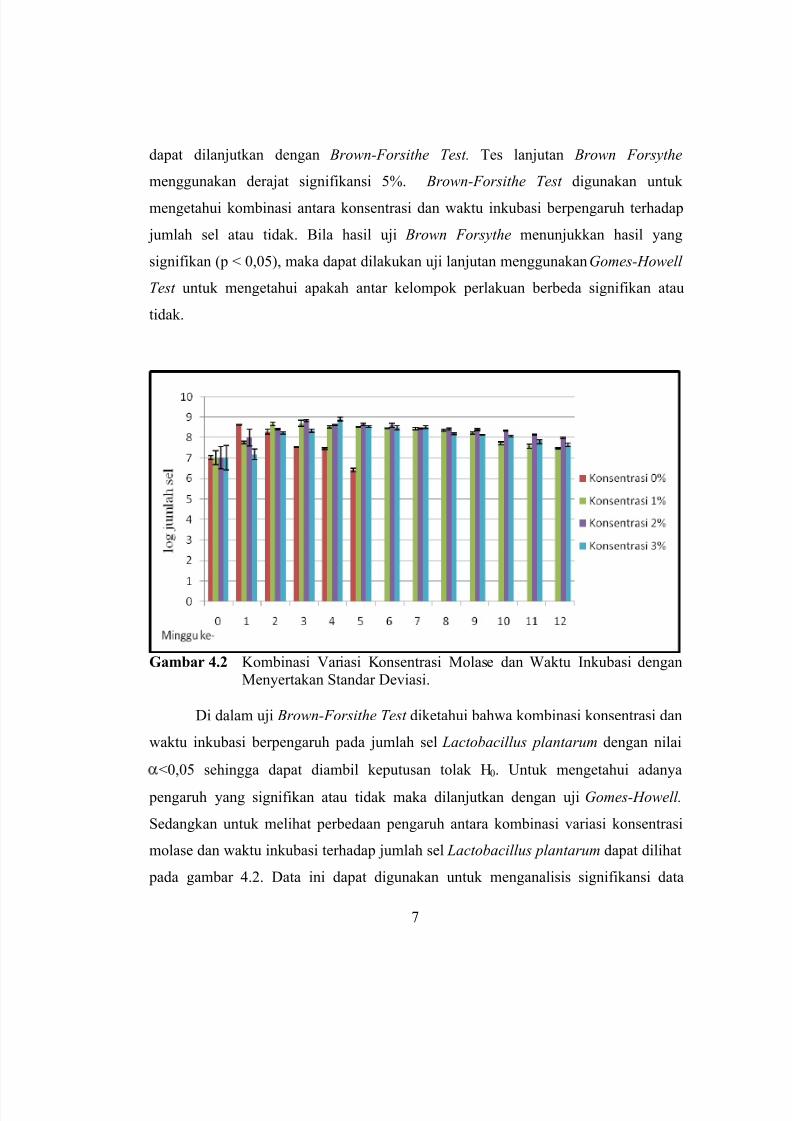
7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 87/97
7
dapat dilanjutkan dengan Brown-Forsithe Test. Tes lanjutan Brown Forsythe
menggunakan derajat signifikansi 5%. Brown-Forsithe Test digunakan untuk
mengetahui kombinasi antara konsentrasi dan waktu inkubasi berpengaruh terhadap
jumlah sel atau tidak. Bila hasil uji Brown Forsythe menunjukkan hasil yang
signifikan (p < 0,05), maka dapat dilakukan uji lanjutan menggunakan Gomes-Howell
Test untuk mengetahui apakah antar kelompok perlakuan berbeda signifikan atau
tidak.
Gambar 4.2 Kombinasi Variasi Konsentrasi Molase dan Waktu Inkubasi denganMenyertakan Standar Deviasi.
Di dalam uji Brown-Forsithe Test diketahui bahwa kombinasi konsentrasi dan
waktu inkubasi berpengaruh pada jumlah sel Lactobacillus plantarum dengan nilai
<0,05 sehingga dapat diambil keputusan tolak H0. Untuk mengetahui adanya
pengaruh yang signifikan atau tidak maka dilanjutkan dengan uji Gomes-Howell.
Sedangkan untuk melihat perbedaan pengaruh antara kombinasi variasi konsentrasi
molase dan waktu inkubasi terhadap jumlah sel Lactobacillus plantarum dapat dilihat
pada gambar 4.2. Data ini dapat digunakan untuk menganalisis signifikansi data
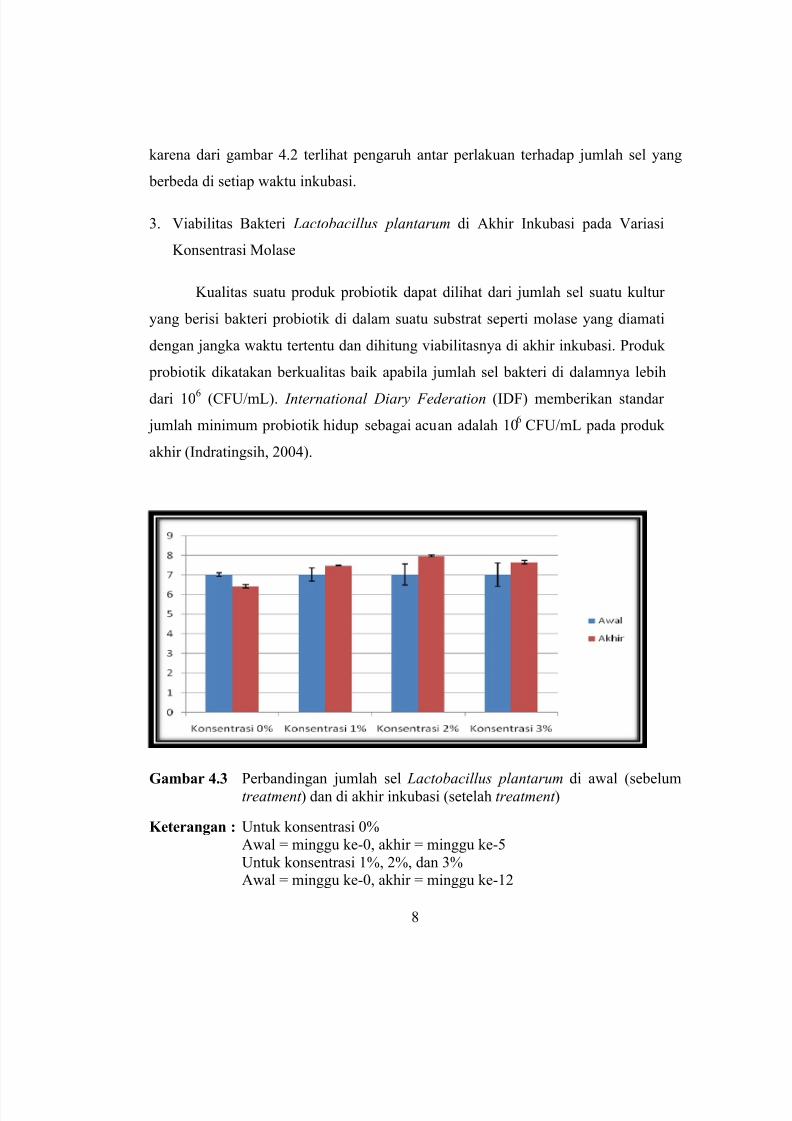
7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 88/97
8
karena dari gambar 4.2 terlihat pengaruh antar perlakuan terhadap jumlah sel yang
berbeda di setiap waktu inkubasi.
3. Viabilitas Bakteri Lactobacillus plantarum di Akhir Inkubasi pada Variasi
Konsentrasi Molase
Kualitas suatu produk probiotik dapat dilihat dari jumlah sel suatu kultur
yang berisi bakteri probiotik di dalam suatu substrat seperti molase yang diamati
dengan jangka waktu tertentu dan dihitung viabilitasnya di akhir inkubasi. Produk
probiotik dikatakan berkualitas baik apabila jumlah sel bakteri di dalamnya lebih
dari 106 (CFU/mL). International Diary Federation (IDF) memberikan standar
jumlah minimum probiotik hidup sebagai acuan adalah 106
CFU/mL pada produk
akhir (Indratingsih, 2004).
Gambar 4.3 Perbandingan jumlah sel Lactobacillus plantarum di awal (sebelumtreatment ) dan di akhir inkubasi (setelah treatment )
Keterangan : Untuk konsentrasi 0%
Awal = minggu ke-0, akhir = minggu ke-5
Untuk konsentrasi 1%, 2%, dan 3%Awal = minggu ke-0, akhir = minggu ke-12

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 89/97
9
Konsentrasi molase 1%, 2%, dan 3% di akhir inkubasi masih mencapai 107
dan kualitasnya sebagai produk probiotik masih baik. Ini dikarenakan jumlah sel
Lactobacillus plantarum pada ketiga konsentrasi tersebut masih di atas 106 (CFU/mL)
di akhir inkubasi (Gambar 4.3). Kultur Lactobacillus plantarum pada konsentrasi
molase 2% adalah produk probiotik yang memiliki jumlah sel paling banyak di akhir
inkubasi dibandingkan dengan konsentrasi yang lain (Gambar 4.3).
KESIMPULAN
Pola pertumbuhan bakteri Lactobacillus plantarum pada konsentrasi molase
1%, 2%, dan 3% memiliki persamaan fase pertumbuhan yaitu fase log, stasioner dan
perlambatan. Kombinasi antara konsentrasi molase dan waktu inkubasi berpengaruh
terhadap pertumbuhan Lactobacillus plantarum. Kombinasi konsentrasi molase 2%
selama 12 minggu inkubasi merupakan kombinasi yang paling baik terhadap jumlah
sel Lactobacillus plantarum. Dan viabilitas bakteri Lactobacillus plantarum pada
variasi konsentrasi molase 1%, 2%, dan 3% di akhir masa inkubasi masih dapat
digunakan sebagai produk probiotik.
DAFTAR PUSTAKA
Dwidjoseputro, D., 2003, Dasar-Dasar Mikrobiologi, Edisi 14, Djambatan, Jakarta.
Emanuel, Vamanu, Vamanu Adrian, Popa Ovidiu, Campeanu Gheorghe, (2005),
Isolation Of a Lactobacillus plantarum strain used for obtaining a product for
the preservation of fodders, African Journal of Biotechnology, Vol. 4 (5), Hal:
403-408.
Indratingsih,W. S., Salasia, S. dan Wahyuni, E., 2004, Produksi Yoghurt Shiitake
(Yohsitake) Sebagai Pangan Kesehatan Berbasis Susu, Jurnal Teknologi dan
Industri Pangan Vol. XV (1), No. 54-60.

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 90/97
10
Murdiyatmo, U., 2003, Prospek Industri Ethanol dari Molase di Indonesia, PT
Perkebunan Nusantara XI, Surabaya.
Nguyen, T.D.T. et al., Characterization of Lactobacillus plantarum PH04, a
potential probiotic bacterium with cholesterol-lowering effects. International
Journal of Food Microbiology. Volume 113, Issue 3, 15 February 2007, Pages
358-361.
Schlegel,H.G. dan Schmidt, K, 1994, Mikrobiologi Umum, Gadjah Mada University
Press, Yogyakarta.
Suriawiria, Unus, 1995, Pengantar Mikrobiologi Umum, Angkasa, Bandung.
Surono, I, 2004, Probiotik Susu Fermentasi dan Kesehatan, PT.Zitri Cipta Karya:
Jakarta.
Waluyo, Lud, Drs. M.Kes, 2004, Mikrobiologi Umum, Universitas Muhammadiyah
Press, Malang.

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 91/97
1
PENGARUH KOMBINASI ZAT PENGATUR TUMBUH
BENZYLADENINE (BA) DAN INDOLE BUTIRIC ACID (IBA)
TERHADAP KULTUR ANTERA CABAI RAWIT (Capsicum frutescens L.)
Tining Sulistyowati1, Dwi Kusuma Wahyuni
*1, dan Hery Purnobasuki
1
1Departemen Biologi, Fakultas Sains dan Teknologi
Universitas Airlangga, Surabaya
*Email: [email protected]
AbstractThe study was aimed to know the influence of indole butiric acid (IBA)
and benzyladenine (BA) combination on the growth of anther culture of Capsicum
frutescens L. This research was designed with Completely Randomized Design
(CRD), used indole butiric acid (IBA) in various consentration of 0, 0.5, 1, and
1.5 ppm and benzyladenine (BA) in various consentration of 0 and 0.5 ppm.
Anthers were collected from buds when the length of petal is slightly longer than
sepal (ratio petal:sepal approximately 3:2). Buds were given a one day of 4oC
pretreatment, followed by culture of the anthers in double layer medium (solid
medium and liquid upper layer). The medium contained MS component solidified
with 8 gr/L agar in solid medium and 6% sucrose and 500 mg/L glutamin in the
liquid upper layer. Data of enlarged anther were analyzed with ANOVA followed
by Duncan, meanwhile data of dehisced anther were analyzed with Kruskal-
Wallis Test followed by Mann-Whitney Test. The result showed that the BA-IBA
combination influenced the growth of anther culture. Combination of 0,5 ppm BA
1 ppm IBA was the best treatment to influence anther culture of Capsicum
frutescens L. Combination of 0,5 ppm BA 0,5 ppm IBA gave result 70±0% of
enlarged anther and 10±0% on percentage of dehisced anther.
Keywords: Anther culture, benzyladenine, indole butiric acid, Capsicum
frutescens L.
Pendahuluan
Cabai rawit (Capsicum frutescens L.) memiliki nilai tinggi untuk industri
makanan dan farmasi. Kebutuhan cabai di Indonesia terus meningkat seiring
dengan meningkatnya jumlah penduduk dan industri makanan. Namun, budidaya
cabai di Indonesia pada umumnya masih belum dilakukan secara intensif,
sehingga produktivitasnya masih sangat rendah (Anonimus, 2005). Pemuliaan

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 92/97
2
tanaman dengan menggunakan galur murni sangat diperlukan untuk tanaman itu.
Salah satu metode pemuliaan tanaman yaitu menggunakan kultur haploid.
Keberhasilan aplikasi teknik hapkultur antera dalam mendapatkan tanaman
haploid ganda pada tanaman cabai masih sedikit yang dilaporkan (Barany et al.,
2005). Menurut Datta (2005), keberhasilan kultur haploid dipengaruhi oleh
beberapa faktor antara lain genotip tanaman donor, kondisi tanaman donor, tahap
perkembangan mikrospora, perlakuan pra kultur, dan media kultur.
Wang et al. (1973) berhasil mengkultur dan meregenerasi tanaman haploid
dari antera cabai (Capsicum annuum) dengan menggunakan zat pengatur tumbuh
kinetin dan NAA, serta kinetin dan 2,4-D. Demikian juga yang telah dilakukan
Dumas de Vaulx et al. (1981) dan Gyulai et al. (2000). Supena (2004)
mengembangkan protokol kultur sebar mikrospora pada C. annuum menggunakan
zeatin dan IAA.
Karena itulah perlu dilakukan penelitian mengenai kombinasi zat pengatur
tumbuh indole butiric acid (IBA) dan benzyladenine (BA) untuk meningkatkan
keberhasilan kultur antera cabai rawit. Pecahnya antera merupakan jalan bagi
mikrospora di dalam antera untuk keluar dan menyebar pada media yang telah
disediakan.
Bahan dan Metode
Penelitian ini dilaksanakan pada bulan Januari sampai Juni 2012 di
Laboratorium Fisiologi Tumbuhan, Departemen Biologi, Fakultas Sains dan
Teknologi, Universitas Airlangga Surabaya.
Tanaman cabai rawit (Capsicum frutescens L.) varietas Dewata F1.
Kuncup bunga cabai rawit dengan ukuran yang sudah ditentukan (perbandingan
petal:sepal=3:2) dipanen pada pagi hari. Bahan-bahan kimia yang digunakan
untuk kultur antera cabai rawit adalah bahan penyusun media MS ( Murashige and
Skoog), zat pengatur tumbuh (IBA, dan BA), akuades steril, clorox, alkohol 70%,
dan spiritus.
Penelitian menggunakan rancangan acak lengkap dengan 7 perlakuan
kombinasi IBA dan BA. Tiap perlakuan menggunakan 3 ulangan. Pengamatan
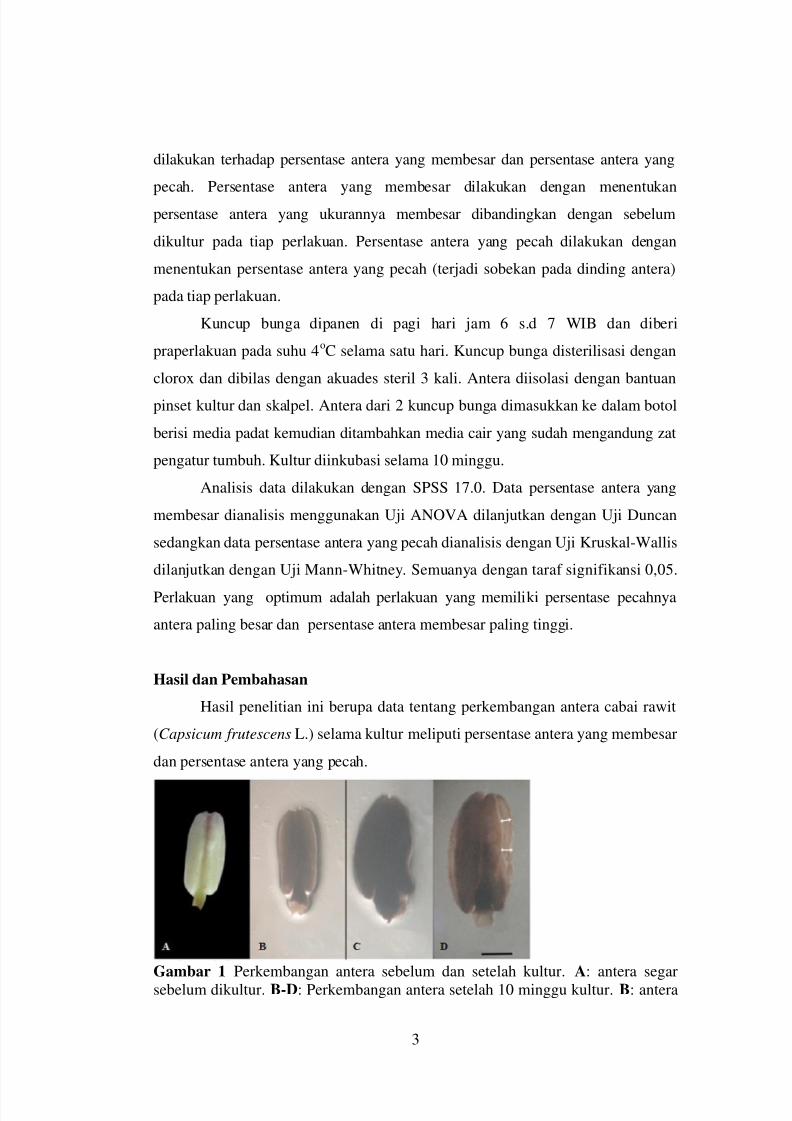
7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 93/97
3
dilakukan terhadap persentase antera yang membesar dan persentase antera yang
pecah. Persentase antera yang membesar dilakukan dengan menentukan
persentase antera yang ukurannya membesar dibandingkan dengan sebelum
dikultur pada tiap perlakuan. Persentase antera yang pecah dilakukan dengan
menentukan persentase antera yang pecah (terjadi sobekan pada dinding antera)
pada tiap perlakuan.
Kuncup bunga dipanen di pagi hari jam 6 s.d 7 WIB dan diberi
praperlakuan pada suhu 4oC selama satu hari. Kuncup bunga disterilisasi dengan
clorox dan dibilas dengan akuades steril 3 kali. Antera diisolasi dengan bantuan
pinset kultur dan skalpel. Antera dari 2 kuncup bunga dimasukkan ke dalam botol
berisi media padat kemudian ditambahkan media cair yang sudah mengandung zat
pengatur tumbuh. Kultur diinkubasi selama 10 minggu.
Analisis data dilakukan dengan SPSS 17.0. Data persentase antera yang
membesar dianalisis menggunakan Uji ANOVA dilanjutkan dengan Uji Duncan
sedangkan data persentase antera yang pecah dianalisis dengan Uji Kruskal-Wallis
dilanjutkan dengan Uji Mann-Whitney. Semuanya dengan taraf signifikansi 0,05.
Perlakuan yang optimum adalah perlakuan yang memiliki persentase pecahnya
antera paling besar dan persentase antera membesar paling tinggi.
Hasil dan Pembahasan
Hasil penelitian ini berupa data tentang perkembangan antera cabai rawit
(Capsicum frutescens L.) selama kultur meliputi persentase antera yang membesar
dan persentase antera yang pecah.
Gambar 1 Perkembangan antera sebelum dan setelah kultur. A: antera segar
sebelum dikultur. B-D: Perkembangan antera setelah 10 minggu kultur. B: antera

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 94/97
4
yang tidak membesar dan tidak pecah. C: antera yang membesar. D: antera yang
pecah; panah putih merupakan sobekan pada dinding antera. bar = 0,1 cm untuk
A-D.
Pengamatan terhadap perkembangan antera selama kultur menunjukkan
antera yang semula berwarna kekuningan dengan ukuran sekitar 0,1x0,2 cm
(Gambar 1A), beberapa hari kemudian antera mulai mengalami pembesaran
ukuran dimana pada minggu kesepuluh ukurannya dapat mencapai 0,15x0,27 cm
(Gambar 1C) dan pada minggu ketiga mulai terdapat antera yang pecah (Gambar
1D). Pecahnya antera terjadi pada sisi lateral pada dinding antera (panah putih).
Namun ada juga antera yang tidak mengalami perubahan ukuran dan juga tidak
pecah (Gambar 1B).
Tabel 1 menunjukkan rerata persentase antera yang membesar dan
persentase antera yang pecah pada berbagai kombinasi perlakuan benzyladenine
(BA) dan indole butiric acid (IBA). Analisis statistik pada kedua parameter
tersebut menunjukkan bahwa pemberian kombinasi BA dan IBA memberikan
pengaruh terhadap persentase antera yang membesar dan persentase antera yang
pecah.
Tabel 1 Rerata persentase antera yang membesar dan persentase antera yang
pecah pada kultur antera cabai rawit (Capsicum frutescens L.) pada berbagai
kombinasi benzyladenine (BA) dan indole butiric acid (IBA). Data merupakan
rerata dari 3 replikasi pada pengamatan minggu kesepuluh.
PerlakuanRerata persentase antera
yang membesar (%)
Rerata persentase antera
yang pecah (%)
BA0IBA0,5 50,00±10a
0,00±0ac
BA0IBA1 53,33±11,55a 3,33±5,77a c
BA0IBA1,5 53,33±5,77a
0,00±0ac
BA0,5IBA0 60,00±0a
0,00±0ac
BA0,5IBA0,5 66,67±15,28a
10,00±0
BA0,5IBA1 70,00±0 10,00±0
BA0,5IBA1,5 60,00±10a
6,67±5,77a c
Keterangan: Angka-angka yang diikuti oleh huruf yang sama berarti berbeda tidak
nyata (0,05) dengan Uji Duncan (persentase antera yang membesar)
dan Uji Mann-Whitney (persentase antera yang pecah).

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 95/97
5
Pada pengamatan persentase antera yang membesar, hasil terendah sebesar
50% ditunjukkan oleh perlakuan BA 0 ppm IBA 0,5 ppm sedangkan hasil
tertinggi (70%) ditunjukkan oleh perlakuan BA 0,5 ppm IBA 1 ppm. Pada
pengamatan persentase antera yang pecah, hasil terendah sebesar 0% ditunjukkan
oleh perlakuan BA 0 ppm IBA 0,5 ppm, BA 0 ppm IBA 1,5 ppm, dan BA 0,5
ppm IBA 0 ppm sedangkan hasil tertinggi (10%) ditunjukkan oleh perlakuan BA
0,5 ppm IBA 0,5 ppm dan BA 0,5 ppm dan IBA 1 ppm.
Dengan melihat hasil pada kedua parameter tersebut, dapat disimpulkan
bahwa perlakuan yang paling baik untuk kultur antera C. frutescens adalah
perlakuan BA 0,5 ppm dan IBA 1 ppm. Hal ini memperlihatkan bahwa pemberian
kombinasi auksin lebih tinggi daripada sitokinin memberikan hasil yang lebih
baik dibandingkan perlakuan lainnya.
Hasil penelitian menunjukkan bahwa keberhasilan kombinasi auksin yang
lebih rendah daripada sitokinin ini sama dengan hasil penelitian Wang et al.,
(1973) yang melakukan kultur antera dan regenerasi C.annuum dimana kombinasi
yang paling baik adalah kombinasi 4,65µM kinetin dan 5,37 µM NAA dan
4,65µM kinetin dan 4,52 µM 2,4-D. Adanya perbedaan konsentrasi antara hasil
penelitian dengan penelitian sebelumnya terutama dikarenakan adanya perbedaan
kebutuhan akan zat pengatur tumbuh eksogen. Kebutuhan akan zat pengatur
eksogen sangat dipengaruhi oleh genotip tanaman. Selain itu Reinert dan Bajaj
(1977) melaporkan bahwa kebutuhan akan auksin dan sitokinin eksogen
tergantung pada kadar auksin dan sitokinin endogen di dalam antera.
Perlakuan dengan penambahan zat pengatur tumbuh auksin saja atau
sitokinin saja menyebabkan persentase antera yang membesar dan pecah menjadi
kecil. Hal ini disebabkan karena pembelahan sel diatur oleh auksin dan sitokinin,
keduanya mempengaruhi fase berbeda dalam siklus sel. Auksin mempengaruhi
replikasi DNA sedangkan sitokinin mengendalikan peristiwa menuju mitosis
(Jouanneau dalam George et al., 2008). Adanya auksin menyebabkan sel
mikrospora di dalam antera berkembang, namun jika tidak diikuti dengan
pembelahan sel maka ukuran antera tidak dapat bertambah secara signifikan.

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 96/97
6
Pada pengamatan dijumpai antera yang tidak membesar atau tidak pecah
meskipun telah diinkubasi selama sepuluh minggu, hal itu mungkin disebabkan
eksplan telah mati karena sterilisasi yang terlalu kuat atau karena adanya variasi
individu yang menyebabkan adanya respon yang berbeda terhadap perlakuan yang
diberikan. Selain itu ada juga antera yang membesar namun tidak pecah, hal ini
disebabkan perkembangan mikrospora di dalam antera tidak cukup kuat untuk
mendorong dinding antera sehingga tidak mampu membuat dinding antera pecah.
Pada penelitian ini antera dikultur pada media dua lapis. Mikrospora di
dalam antera mengalami perkembangan namun pada beberapa antera terjadi
perubahan warna menjadi kecoklatan (browning) yang menunjukkan kematian sel
dinding antera. Hal ini sejalan dengan teori apotopsis dimana akan terjadi
kematian sel untuk menyokong sel lain yang sedang berkembang. Selain itu
browning dapat terjadi karena dihasilkan senyawa fenol yang akan menghambat
aktivitas enzim yang selanjutnya dapat mematikan tanaman (Hu and Wang, 1983)
Kesimpulan
Berdasarkan hasil penelitian ini, dapat disimpulkan bahwa:
1. Pemberian kombinasi benzyladenine (BA) dan indole butiric acid
(IBA)berpengaruh secara nyata terhadap pertumbuhan kultur antera cabai
rawit (Capsicum frutescens L.), yaitu pada persentase antera yang
membesar dan persentase antera yang pecah.
2. Perlakuan BA0,5IBA1 (0,5 ppm BA dan 1 ppm IBA) merupakan
kombinasi yang terbaik untuk kultur antera C. frutescens yang
menghasilkan persentase antera yang membesar dan pecah paling tinggi
dengan nilai persentase antera yang membesar 70±0% dan persentase
antera yang pecah 10±0%..
Ucapan Terima Kasih
Terima kasih kepada Bapak Tunut dan Ibu Saijah yang telah memelihara
donor eksplan untuk penelitian ini.

7/26/2019 cabai rawit 1
http://slidepdf.com/reader/full/cabai-rawit-1 97/97
Daftar Pustaka
Anonimus, 2005, Statistik Pertanian 2005, Departemen Pertanian, Jakarta.
Barany, I., Melendi, P.G., Fadon, B., Mitykot, J., Risueno, M.C., and Testillano,
P.S., 2005, Microspore-derived embryogenesis in pepper (Capsicum
annuum L.): subcellular rearrangements through development, Biol.
Cell, 97 (9): 709 – 722.
Datta, S. K., 2005, Androgenic haploids: factors controlling development and its
application in crop improvement, Cureent Science, 89 (11): 1870-1878.
Dumas de Vaulx R., Chambonnet, D., and Pochard, E., 1981, Culture in vitro
d'antheres de piment (Capsicum annuum): amelioration des taux
d'obtention de plantes chez differents genotypes par traitments a+35 °C.
Agronomie, Vol 1: 859 – 64.
George, E.F., Hall, M.E., and Klerk, G.D., 2008, Plant propagation by tissueculture, third edition, Vol 1. The Background, Springer, Netherlands.
Gyulai, G., Gemesne, J.A., Sagi, Z.S., Venczel, G., Pinter, P., and Kyristof, Z.,
2000, Doubled haploid development and PCR analysis of F1-hydrid
derived DH-R2 paprika (Capsicum annuum L.) lines. J Plant Physiol,
156: 168 – 74.
Hu, C., and Wang, J. P., 1983, Meristem shoot tip and bud culture. In: Handbook
of Plant Cell, MacMillan Publ, New York.
Reinert, J. and Bajaj, Y. P. S., 1977, Anther culture: haploid production and its
significance. In ‘Applied and fundamental Aspects of Plant Cell, Tissue
and Organ Cult., Spinger-Verlag, Berlin.
Supena, E.D.J., 2004, Innovations in microspore embryogenesis in Indonesian hot
pepper (Capsicum annuum L.) and Brassica napus L., Thesis,
Wageningan University, Netherlands.
Wang, Y.Y., Sun, C.S., Wang, C.C., and Chien, N.F., 1973. The induction of
pollen plantlets of Triticale and Capsicum annuum from anther culture,
Sci sin, 16: 147-151.