jurnal radioterapi
DESCRIPTION
radioterapiTRANSCRIPT

TinjauanPencegahan dan Terapi Kanker Melalui Modulasi dari Tumor MikroStephanie C. Caseya, Amedeo Amedeib, Katia Aquilanoc, Asfar S. Azmid, Fabian Benenciae, Dipita Bhaktaf, Alan E. Bilslandg, Chandra S. Boosanih, Sophie Cheni, Maria Rosa Cirioloc , Sarah Crawfordj, Hiromasa Fujiik, Alexandros G. Georgakilasl, Gunjan Guhaf, Dorota Halickam, William G. Helferichn, Petr Henebergo, Kanya Honokik, W. Nicol Keithg, Sid P. Kerkarp, Sulma I. Mohammedq, Elena Niccolair, Somaira Nowsheens, HP Vasantha Rupasinghet, Abbas Samadiu, Neetu Singhv, Wamidh H. Thalibw, Vasundara Venkateswaranx, Richard L. Whelany, Xujuan yangn, Dean W. Felshera, *
a Divisi Onkologi, Departemen Kedokteran dan Patologi, Stanford University School of Medicine, Stanford, CA, Amerika Serikat bDepartemen Eksperimental dan Clinical Medicine, University of Florence, Florence, Italia cDepartemen Biologi, University of Rome "Tor Vergata", Roma, Italia dDepartemen Onkologi, Wayne State University School of Medicine, Detroit , MI, Amerika Serikat eDepartemen Biomedical Sciences, Ohio University, Athens, OH, Amerika Serikat fSekolah Kimia dan Bioteknologi, SASTRA Universitas, Thanjavur 613401, Tamil Nadu, India g Institute of Cancer Sciences, University of Glasgow, Glasgow, Inggris Kerajaan h Departemen Ilmu Biomedis, School of Medicine, Universitas Creighton, Omaha, NE, Amerika Serikat I Ovarium dan prostat Laboratorium Penelitian Kanker, Guildford, Surrey, Inggris Raya jDepartemen Biologi, Southern Connecticut State University, New Haven, CT, Inggris negara kDepartemen Bedah ortopedi, Nara Medical University, Kashihara, Jepang lDepartemen Fisika, Fakultas Matematika Terapan dan Ilmu Pengetahuan Alam, Universitas Teknik Nasional Athena, Athena, Yunani m New York Medical College, Valhalla, NY, Amerika Serikat n Universitas Illinois di Urbana-Champaign, Champaign-Urbana, IL, Amerika Serikat o Charles University di Praha, Fakultas Ketiga Kedokteran, Praha, Republik Ceko p Pusat Penelitian Kanker, National Cancer Institute, National Institutes of Health, Bethesda, MD, Amerika Serikat q Departemen Perbandingan Pathobiology, Purdue University Center for Cancer Research, West Lafayette, IN, Amerika Serikat rUniversity of Florence, Florence, Italia s Program Pelatihan Scientist Medis, Mayo Graduate School, Mayo Medical School, Mayo Clinic, Rochester, MN, Amerika negaratDepartemen Ilmu Lingkungan, Fakultas Pertanian, Universitas Dalhousie, Nova Scotia, Kanada u Sanus Biosciences, San Diego, Amerika
Serikat v Lanjutan Molecular Science Research Centre (Pusat Advanced Research), University Medical Raja George, Lucknow, Uttar Pradesh, India w Departemen Farmasi Klinik dan Terapi, Universitas Sains Terapan, Amman, Jordan xDivisi Urologi, Sunnybrook Health Sciences Centre, Toronto, Ontario, Kanada yMount Sinai Hospital Roosevelt, Icahn Mount Sinai School of Medicine, New York City, NY, Amerika Serikat
A b s t r a kkanker muncul dalam konteks in vivo tumor lingkungan mikro. Lingkungan mikro ini adalah sebab dan akibat dari tumorigenesis. Tumor dan sel tubuh berko-evolusi secara dinamis melalui interaksi seluler langsung dan tidak langsung, memunculkan efek multiskala pada banyak program biologis, termasuk proliferasi seluler, pertumbuhan, dan metabolisme, serta angiogenesis dan hipoksia dan kekebalan bawaan dan adaptif. Di sini kita menyoroti proses biologis tertentu yang bisa dimanfaatkan sebagai target untuk pencegahan dan terapi kanker. Secara khusus, kami menjelaskan bagaimana penghambatan target seperti sintesis kolesterol dan metabolit, reakfif oksigen spesies dan hipoksia, aktivasi makrofag dan konversi, indoleamin regulasi 2,3-dioksigenase sel dendritik, faktor pertumbuhan endotel vaskular regulasi dari angiogenesis, fibrosis penghambatan, Endoglin, dan sinyal Janus kinase muncul sebagai contoh nexuses potensi penting dalam regulasi tumorigenesis dan lingkungan mikro tumor yang dapat ditargetkan. Kami juga telah mengidentifikasi agen terapi sebagai pendekatan, dalam produk alami tertentu seperti berberin, resveratrol, onionin A, epigallocatechin gallate, genistein, curcumin, naringenin, des oxyrhapontigenin, piperine, dan zerumbon, yang dapat menjamin penyelidikan lebih lanjut untuk menargetkan lingkungan mikro tumor . untuk pengobatan dan / atau pencegahan kanker.
Info Artikel
Pasal sejarah: Tersedia online 10 April 2015
Kata kunci: tumor mikro Kanker biologi Kanker terapi kanker pencegahan

1. Pendahuluan3.1 Tumor lingkungan mikro sebagai target terapi
Tumor lingkungan mikro sangat penting untuk inisiasi dan pemeliharaan tumorigenesis [1,2]. Tumor lingkungan mikro terdiri dari jaringan yang kompleks yang mencakup multipoten sel stroma / sel batang mesenchymal, fibroblas, pembuluh darah, prekursor sel endotel, sel-sel kekebalan tubuh, dan faktor-faktor yang disekresi seperti sitokin [2]. Selama perkembangan tumor, perubahan dalam lingkungan mikro terjadi melalui efek pada molekul serta tingkat sel dan melibatkan interaksi antara sel-sel kanker baru jadi dan sel tubuh struktural serta sel-sel kekebalan adaptif dan bawaan [3]. Banyak dari "keunggulan dari kanker" berhubungan dengan tumor lingkungan mikronya, termasuk kemampuan untuk menginduksi proliferasi dan menghambat apoptosis, untuk menginduksi angiogenesis dan menghindari hipoksia, menghambat sistem kekebalan tubuh dan menghindari deteksi kekebalan tubuh, dan mengaktifkan sel-sel kekebalan tubuh untuk dukungan invasi dan metastasis [4]. Jalur spesifik onkogenik dapat dikaitkan dengan perubahan dramatis dalam lingkungan mikro tumor [5-8]. Oleh karena itu, manipulasi tumor lingkungan mikro dapat digunakan sebagai pendekatan untuk mencegah serta mengobati kanker.
Identifikasi target terapi pada tumor lingkungan mikro dapat berguna dalam pengobatan dan pencegahan kanker. Pendekatan biologis yang khas telah menyelidiki mekanisme molekuler dan seluler tertentu dan kemudian untuk memeriksa apakah penghambatan atau aktivasi memiliki konsekuensi yang diharapkan untuk tumorigenesis. Namun, ada peringatan untuk pendekatan ini. Molekul yang sama dan sel efektor dapat memiliki peran baik pada pencegahan dan inisiasi tumorigenesis. Kanker yang berbeda dapat terjadi melalui mekanisme yang berbeda. Apa yang membatasi dalam beberapa konteks mungkin dalam keadaan lain tidak penting. Beberapa target mungkin memiliki efek pada beberapa jalur dan program yang dapat menyerang balik efektivitas mereka secara keseluruhan. Oleh karena itu, kemampuan untuk mengurangi reaksi target mikro dan mengidentifikasi terapi yang cocok masih dalam proses.
Dalam ulasan ini, kami telah mengambil pendekatan yang berbeda. Melalui sebuah inisiatif yang didukung oleh Proyek Halifax, sekelompok penyelidik bekerja sama sebagai sebuah tim untuk mengidentifikasi kedua target spesifik dan pendekatan baru untuk terapi menghambat aspek-aspek tertentu dari lingkungan mikro tumor. Melalui pendekatan integratif kami telah mengidentifikasi strategi untuk pengobatan dan pencegahan kanker. Kemudian, kami meneliti literatur dan mengidentifikasi kemungkinan agen, dalam produk alami tertentu, yang berpotensi menghambat sebagian atau beberapa dari target
tersebut. Tujuan kami adalah untuk mengidentifikasi agen yang dapat dimanfaatkan untuk pencegahan dan/atau pengobatan kanker. Akhirnya, tim memanfaatkan pendekatan validasi silang untuk memeriksa bagaimana target tersebut dan pendekatan, baik sendiri atau dalam kombinasi, dapatkah berguna untuk pencegahan dan/atau pengobatan kanker.
Kami mengidentifikasi 10 program yang bisa atau pasti muncul untuk menjadi target dan ada 10 agen alami yang dapat memediasi efek anti-kanker mereka melalui lingkungan mikro tumor (Gambar. 1 dan 2, Tabel 1 dan 2). Daftar kami bukan sebuah pemeriksaan lengkap dari semua target yang mungkin atau pendekatan terapi melainkan upaya untuk mengidentifikasi spektrum luas, toksisitas rendah terapi yang dapat dikombinasikan dengan terapi yang ada.
Target diidentifikasi meliputi program metabolik yang mungkin secara luas mempengaruhi banyak program biologi sel yang berdampak tumorigenesis dan lingkungan mikro tumor, termasuk (sintesis kolesterol dan metabolit, reaktif oksigen spesies (ROS) dan hipoksia, peradangan, program kekebalan terkait bawaan dan adaptif (konversi macrophage, sel dendritik (DC) aktivasi,sinyal kekebalan), sel tubuh lingkungan mikro yang terkait program seluler (fibrosis, angiogenesis), dan program sitokin yang dimediasi (interleukin (IL)-6,Endoglin,dan Janus terkait kinase (JAK)) (Gambar. 1, Tabel 1 dan 2).
Kami sangat fokus pada pendekatan identifikasi untuk menghambat target tersebut, termasuk produk-produk alami yang mungkin memiliki aktivitas antikanker yang signifikan. Beberapa molekul ini mungkin lebih umum mempengaruhi tumorigenesis dan lingkungan mikro (berberin), yang lain lebih khusus menargetkan ROS (resveratrol, desoxyrhapontigenin) konversi makrofag (onionin A), indoleamin 2,3-dioksigenase (IDO) regulasi sel dendritik (epigallocatechin-3-gallate (EGCG)), sintesis kolesterol (genis- Tein), fibrosis (naringenin), peradangan dan sinyal kekebalan (piperine), faktor pertumbuhan endotel vaskular (VEGF) inhibisi (kurkumin), dan JAK signaling (zerumbon). Pendekatan ini dapat menjamin penyelidikan lebih lanjut (Gambar. 1, Tabel 1 dan 2). Agen ini umumnya memiliki toksisitas rendah, menunjukkan bahwa mereka dapat dikombinasikan dengan terapi lain atau.
3.2 Cross-validasi pendekatan dan targetKami mengidentifikasi pendekatan dan target
melalui analisis literatur ilmiah melalui tim peneliti dari banyak subspesialisasi. Kami membuat beberapa asumsi. Pertama, komplex biologi dan heterogenitas kanker menyarankan bahwa pendekatan terapi yang paling efektif mungkin memerlukan tindakan simultan pada mekanisme yang penting bagi banyak keunggulan dari kanker. Kedua, kami mengantisipasi bahwa sinergi akan dicapai dengan menggabungkan target spesifik dengan pendekatan tertentu. Ketiga, kita

menganggap bahwa kita bisa memvalidasi kedua target dan pendekatan melalui cross-validasi melalui analisis sastra. Akhirnya, kami menganggap itu penting untuk memeriksa hubungan sasaran diidentifikasi dan pendekatan dinominasikan di berbagai aspek biologi kanker.Khususnya, target dan pendekatan yang diidentifikasi untuk tumor lingkungan mikro telah terbukti relevan dengan keunggulan kanker lainnya. Ini dicatat sebagai efek “komplemen”, sementara mereka yang ditemukan memiliki tindakan pro-tumorigenic dicatat sebagai memiliki efek "bertentangan". Contoh di mana laporan tindakan yang relevan dalam aspek lain dari biologi kanker dicampur, di mana laporan yang menunjukkan
kedua potensi pro-kanker dan potensi anti-tumorigenic, kami telah menggunakan istilah "kontroversial." Akhirnya, dalam kasus di mana tidak ada dukungan literatur ditemukan dokumen yang berhubungan dengan situs target atau pendekatan dalam aspek tertentu biologi kanker, kita didokumentasikan ini sebagai hasil validasi ini ditunjukkan pada Tabel 1 dan 2.
Prioritas kami adalah untuk memilih target dan pendekatan setelah pertimbangan efek marker yang sesuai. Kami memeriksa untuk kemungkinan tindakan insidental dari intervensi terapeutik. Kami mengumpulkan pandangan lengkap dari literatur.
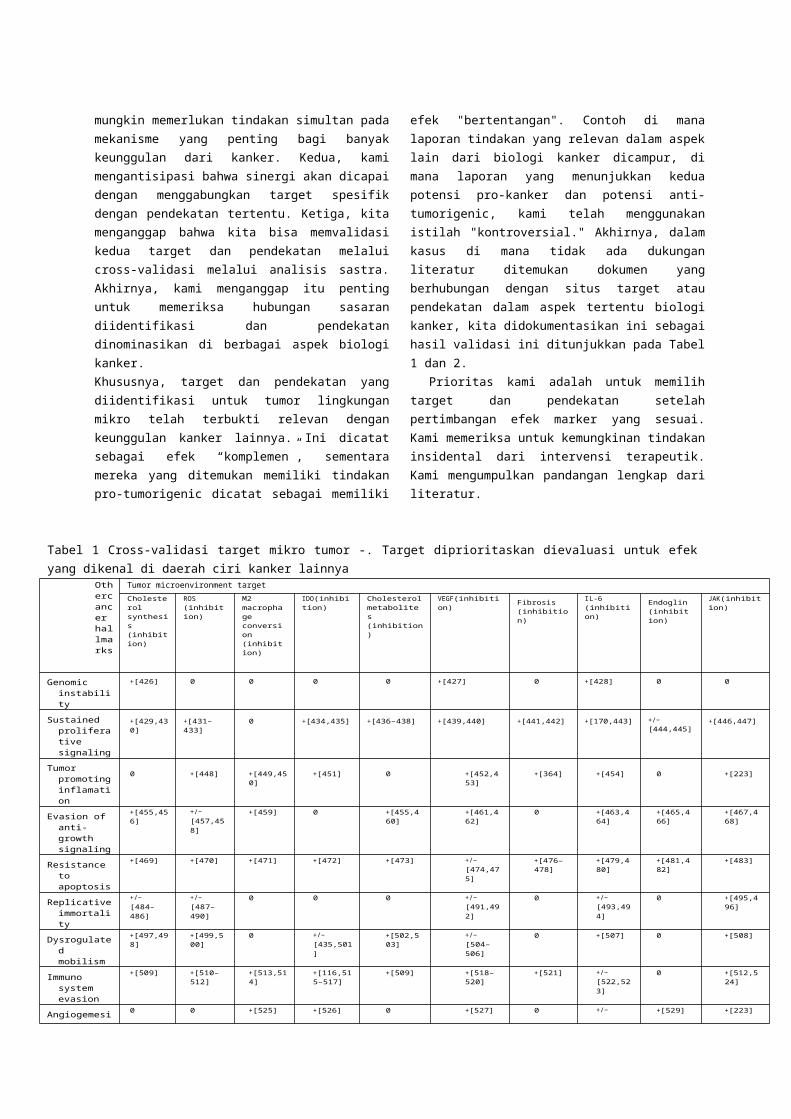
Tabel 1 Cross-validasi target mikro tumor -. Target diprioritaskan dievaluasi untuk efek yang dikenal di daerah ciri kanker lainnya
Othercancer hallmarks
Tumor microenvironment target
Cholesterol synthesis (inhibition)
ROS (inhibition)
M2 macrophage conversion (inhibition)
IDO(inhibition) Cholesterol metabolites (inhibition)
VEGF(inhibition) Fibrosis(inhibition)
IL-6 (inhibition) Endoglin
(inhibition)
JAK(inhibition)
Genomic instability
+[426] 0 0 0 0 +[427] 0 +[428] 0 0
Sustained proliferative signaling
+[429,430] +[431–433] 0 +[434,435] +[436–438] +[439,440] +[441,442] +[170,443] +/− [444,445] +[446,447]
Tumor promoting inflamation
0 +[448] +[449,450] +[451] 0 +[452,453] +[364] +[454] 0 +[223]
Evasion of anti-growth signaling
+[455,456] +/− [457,458]
+[459] 0 +[455,460] +[461,462] 0 +[463,464] +[465,466] +[467,468]
Resistance to apoptosis
+[469] +[470] +[471] +[472] +[473] +/− [474,475]
+[476–478]
+[479,480] +[481,482] +[483]
Replicative immortality
+/− [484–486]
+/− [487–490]
0 0 0 +/− [491,492]
0 +/− [493,494]
0 +[495,496]
Dysrogulated mobilism
+[497,498] +[499,500] 0 +/− [435,501]
+[502,503] +/− [504–506]
0 +[507] 0 +[508]
Immuno system evasion
+[509] +[510–512]
+[513,514] +[116,515–517]
+[509] +[518–520]
+[521] +/− [522,523]
0 +[512,524]
Angiogemesis 0 0 +[525] +[526] 0 +[527] 0 +/− [453,528]
+[529] +[223]
Tissue invasion and metastasis
+[530,531] +[532,533] +[513,534] +[535,536] +[537,538] +[539–541]
+[542,543] +[544] +[545,546] +[467,547]
Targetyang ditemukan memiliki komplementer, tindakan anti-karsinogenik dilaporkan di daerah ciri lain yang ditunjukkan dengan "+", sedangkan target yang ditemukan memiliki tindakan pro-karsinogenik di daerah ciri lain yang ditandai dengan"-".Dalam kasus di mana laporan tindakan yang relevan di daerah ciri lainnya dicampur (yaitu, laporan yang menunjukkan kedua potensi anti-karsinogenik dan potensi pro-karsinogenik), simbol "+ /-"digunakan. Akhirnya, dalam kasus di mana tidak ada dukungan literatur ditemukan untuk mendokumentasikan relevansi target dalam aspek tertentu biologi kanker, kami mendokumentasikan ini sebagai "0".
Othercancer hallmarks
Tumor microenvironment target
Cholesterol synthesis (inhibition)
ROS (inhibition)
M2 macrophage conversion (inhibition)
IDO(inhibition) Cholesterol metabolites (inhibition)
VEGF(inhibition) Fibrosis(inhibition)
IL-6 (inhibition) Endoglin
(inhibition)
JAK(inhibition)
Genomic instability
+[548,549] +[550,551] 0 +/− [552,553] +[554–556] +[557,558] +[559] 0 +[560,561] 0
Sustained proliferative signaling
+[562,563] +[284,564] 0 +[565,566] +/− [567–571] +[572,573] +[574] 0[387] +[391,575] +[413,576]
Tumor promoting inflamation
0+[577,578] 0 0 0 +[579] +[364] 0 0 0
Evasion of anti-growth signaling
+[580,581] +[582] 0 +[583] +[584] +[579,585] 0 0 +[586] 0
Resistance to apoptosis
+[587,588] +[589] 0 +[590] +[591] +[592] +/− [593,594] +[387] +[560] +[595]
Replicative immortality
+[581,596] +[597,598] 0 +[599,600] +[601,602] +[603–605] 0 0 0 0
Dysrogulated mobilism
+[606,607] +[608–610] 0 +[611–614] +[615,616] +[617–619] +[574,620] 0 +[391,621] +[415,622]
Immuno system evasion
−[623–625] +/− [626–631]
0 +[632–636] +/− [637,638] +/− [639–642] +/− [364,643,644] 0 −[645–648] +[418,649]
Angiogemesis +[650] +[651] 0 0 +[652] +[653] 0 0 0 0
Tissue invasion and metastasis
+[654,655] +[627,656] 0 +[657,658] +/− [659,660] +[661,662] +[364,366,663] 0 +[400,403] +[664,665]
Approaches yang ditemukan memiliki komplementer, tindakan anti-karsinogenik pada area ciri tertentu yang diindikasikan dengan "+", sedangkan pendekatan yang ditemukan memiliki tindakan pro-karsinogenik pada area ciri tertentu yang ditandai dengan"-".Dalam kasus di mana laporan tindakan yang relevan di keunggulan lainnya dicampur (yaitu, laporan yang menunjukkan potensi baik anti-karsinogenik dan pro-karsinogenik), simbol "+ /-"digunakan. Akhirnya, dalam kasus di mana tidak ada dukungan literatur ditemukan untuk mendokumentasikan relevansi pendekatan pada aspek tertentu biologi kanker, kami mendokumentasikan ini sebagai "0".
Gambar 1. Target dan pendekatan diidentifikasi yang dapat memodulasi mikro tumor untuk mencegah atau mengobati kanker . target terapi utama diidentifikasi termasuk penghambatan sintesis kolesterol dan metabolit, reakfif oksigen spesies dan
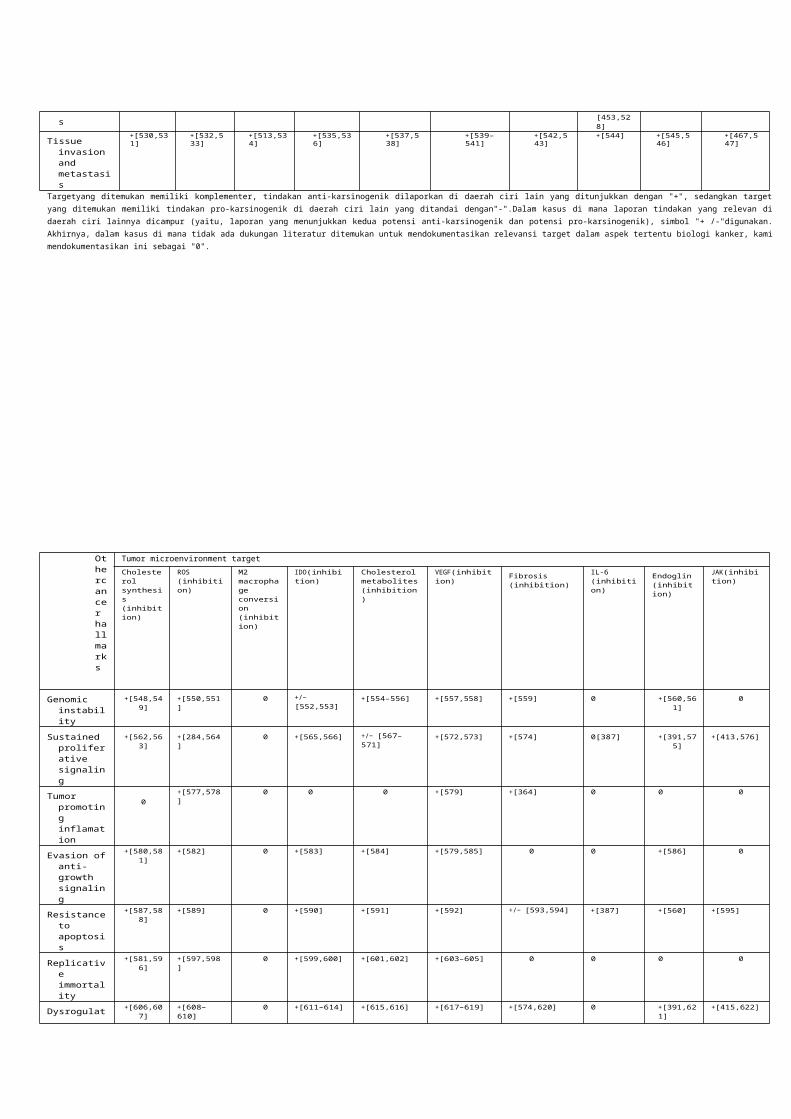
hipoksia, aktivasi makrofag dan konversi, regulasi IDO dari DC, regulasi VEGF
Namun,Kita mengakui bahwa hasil kami adalah titik
Dalam beberapa kasus, bukti diterbitkan marker yang sesuai hubungannya kuat. Dalam kasus lain, bukti yang mendasari lemah, yang terdiri dari hanya satu penelitian in vitro yang melibatkan jenis
2. Target2.1. Sintesis
Jalur kolesterol penting dalam patogenesis dari banyak penyakit
Sintesis kolesterol dan metabolit yang intim dengan patofisiologi karsinogenesis [11/09]. Kolesterol dan metabolitnya memiliki pengaruh pada banyak program biologis yang sangat penting untuk pertumbuhan dan sinyal sel. Kolesterol dan metabolitnya merupakan bagian integral
utusan dan mengatur sinyal seluler dan aktivasi onkogenik. Kolesterol sangat penting untuk sel tubuh fungsi
Kolesterol pertama kali diidentifikasi pada batu empedu [13]. Selanjutnya, kolesterol menjadi penting bagi banyak tujuan proses biologis, termasuk suhu tubuh inti, integritas struktural dan
Kolesterol biosintesis telah
didefinisikan dengan baik [16,17]. Kolestrol umumnya disintesis di hati dimulai dengan satu molekul masing-masing asetil CoA dan acetoacetyl CoA [18]. kolestrol diatur dalam retikulum endoplasma oleh protein sreb (SREBP) 1 dan 2 [19]. Sintesis kolesterol dikendalikan
Gambar 2. Struktur produk alami diidentifikasi yang dapat menargetkan mikro tumor untuk pengobatan atau pencegahan kanker. Molekul ditampilkan meliputi berberin, resveratrol, onionin A, EGCG, genistein, curcumin, naringenin, desoxyrhapontigenin, piperine, dan zerumbon .
Banyak studi menunjukkan bahwa kolestrol dan metabolitnya memiliki peran penting dalam tumorigenesis.
Pertama, studi model tikus menunjukkan bahwa
biosyntetis kolesterol adalah penyebab untuk tumorigenesis [21-23]. Demikian pula, pada model tikus transgenik diinduksi limfoma onkogen dan kanker hati, tumorigenesis dicegah ketika tikus diperlakukan dengan inhibitor HMG-CoA reductase [24,25], yang
ditemukan terkait dengan penghambatan RAS dan MYC onkogen, masing-masing.
Kedua, studi epidemiologi telah menunjukkan bahwa pasien yang menerima agen yang menghambat metabolisme kolesterol mengurangi risiko kanker [26]. Khususnya, kolesterol serum dan risiko kanker tampaknya tergantung pada situs kanker [27].
Ketiga, penelitian lain telah dilaporkan menunjukkan peningkatan kadar kolesterol dalam tumor
dibandingkan dengan jaringan biasa [28,29]. Keempat, kanker sering menunjukkan perubahan dalam program yang mengatur biosintesis kolesterol melalui peningkatan regulasi aktivitas HMG-CoA reductase [30,31], hilangnya umpan balik inhibisi [20], peningkatan penyerapan kolesterol ekstraseluler melalui reseptor LDL [32,33 ] dan penurunan ekspresi kolestrol eksportir ATP yang mengikat kaset transporter A1 (ABCA1) [33-35]. Akhirnya, obesitas dan kadar kolesterol tinggi dikaitkan dengan peningkatan risiko kanker payudara pada wanita pasca menopause [11,36].
Kolesterol metabolit memainkan peran kunci dalam regulasi seluler dan aktivasi nuklir onkogen. Kolesterol metabolit merupakan kunci untuk regulasi banyak onkogen melalui prenilasi termasuk onkogen RAS [25]. Pada gilirannya, ini mengarah pada peraturan bersama dari onkogen MYC [24,25]. Dengan demikian, metabolisme kolesterol kemungkinan memainkan peran dalam tumorigenesis. Kolesterol adalah komponen kunci membran sel, metabolit yang diperlukan untuk mengatur aktivasi onkogen, dan template untuk hormon penting. Pentingnya potensi biosintesis kolesterol pada kanker telah menyebabkan kepentingan yang signifikan dalam penggunaan inhibitor HMG-CoA, statin, untuk pengobatan atau pencegahan kanker manusia [37-41].
2.2. ROS
ROS mempengaruhi lingkungan mikro tumor melalui banyak mekanisme yang mungkin penting untuk pengobatan dan pencegahan yang kanker [42]. ROS dapat didefinisikan sebagai radikal oksigen dan zat pengoksidasi non-radikal yang dapat dengan mudah dikonversi ke radikal yang mengandung satu atau lebih elektron yang tidak berpasangan [43]. Enzim utama yang terlibat dalam generasi ROS yang nikotinamida adenin dinukleotida fosfat (NADPH), myeloperoxidase (MPO) dan xanthine oksidoreduktase (XOR).Reaksi non-enzimatik yang menghasilkan ROS adalah melalui mitokondria dan umumnya melibatkan penggunaan "catalytic" besi atau tembaga ion. ROS yang terlibat dalam berbagai proses metabolisme dan enzim reaksi dalam sel, dalam rantai transpor elektron di mitokondria, ekspresi gen, transduksi sinyal, aktivasi faktor transkripsi [44,45]. Kelebihan produksi ROS pada akhirnya dapat menyebabkan kerusakan jaringan [43].
Kontribusi ROS untuk tumorigenesis melalui mutagenesis serta efek pada lingkungan mikro tumor [46]. Tingkat ROS pada sel kanker lebih tinggi dibandingkan dengan mereka yang hadir dalam sel-sel normal [47,48], sel-sel kanker menghasilkan ROS melalui mitokondria [49]. Disregulasi antioksidan dari mesin matriks mitokondria telah terbukti memberikan
kontribusi yang signifikan selama transformasi seluler. Hal ini dicapai dengan peningkatan dalam tingkatan ROS dalam matriks [49] yang mungkin memainkan peran penting dalam regulasi ROS [50-52].
Regulasi ROS penting untuk tumorigenesis. Enzim mitokondria superoxide dismutase 2 (SOD2) [53,54] mengatur tumor hipoksia [55]. Onkogen dan gen supresor tumor diatur oleh ROS, termasuk fosfatase dan tensin homolog (PTEN) supresor tumor [56,57], kinase mitogen protein diaktifkan (MAPK), dan sinyal ekstraseluler diatur kinase (ERK) jalur [58-60 ]. Tingkat ROS telah terbukti mempengaruhi tumor angiogenesis [61] dan mengatur pembaruan tumor/berhubungan dengan sel induk kanker [62]. Akhirnya, sifat anti-neoplastik dari beberapa agen terapeutik dapat dimediasi oleh sifat antioksidan mereka [63], termasuk tamoxifen [63,64] dan sulphasalazine [65,66]. Manipulasi tingkatan ROS dapat dieksploitasi dimanfaatkan untuk pengobatan dan pencegahan kanker.
2.3. 2.3 Konversi makrofagFungsi dan regulasi makrofag berkontribusi dalam
tumorigenesis. Tumor terkait makrofag (TAM) dan sel-sel kekebalan tubuh bawaan lainnya telah ditemukan untuk mengatur tumor lingkungan mikro, termasuk promosi angiogenesis, inisiasi fibrosis, dan penekanan deteksi kekebalan [67]. Baru-baru ini, telah muncul bahwa tumor dapat mengeluarkan faktor yang mempromosikan konversi makrofag dari fenotip "M1" ke "M2" [68]. Fisiologis makrofag diaktifkan, atau jenis makrofag M1, menghasilkan sitokin seperti IL-1, IL-8, IL-12, IL-15, IL-18, IL-23 dan tumor necrosis factor (TNF) -α dalam menanggapi sinyal melalui reseptor yang serupa dipicu oleh kerusakan terkait oleh pola molekul bakteri, jamur, virus dan parasit [68]. Mediator inflamasi akut, khususnya, IL-12, mempromosikan pengembangan suatu respon imun Th1 untuk menghilangkan patogen asing dan sel-sel kanker [69]. Namun, makrofag dalam tumor tidak terkena sinyal bahaya dan menghasilkan lebih banyak IL-10, sitokin yang mengubah diferensiasi sel T jauh dari respon sitotoksik Th1 [70].M2 makrofag juga mengeluarkan lebih banyak transforming growth factor (TGF) -β, sitokin yang dapat meredam kemampuan sel T untuk

meningkatkan respon target dan dapat menyebabkan sel-sel kanker mencapai sel induk seperti fitur [71]. TGF-β juga menginduksi aktivasi fibroblas dan sel mesenchymal lainnya yang akhirnya mengarah ke fibrosis jaringan. Dengan demikian, tumor dihubungkan dengan makrofag diciptakan yang dapat mempromosikan karsinogenesis, angiogenesis dan menghilangkan kekebalan tubuh.
Makrofag merupakan Major histocompatibility complex (MHC) kelas utama I dan II. Dengan demikian, mereka dapat menyajikan antigen tumor melalui MHC II bagi sel-sel CD4 + T dan bisa menyeberang-sekarang MHC-I ke sel CD8 + T [72]. Setelah aktivasi oleh reseptor serupa seperti lipopolisakarida (LPS) atau interferon-gamma (IFN-Ɣ), Meningkatkan regulasi makrofag molekul kostimulatori seperti MHC- kelas I, CD80, CD83 dan CD86, yang memungkinkan sel T untuk sepenuhnya matang dan meningkatkan antigen respon imun spesifik [73]. Namun, dalam lingkungan mikro tumor, makrofag tidak mampu memunculkan antigen. Ini mungkin reversibel. Dengan demikian, IL-12 dapat mengkonversi M2 menjadi makrofag M1. Hal ini dapat memungkinkan presentasi antigen ke sel CD8 + T dan meningkatkan kekebalan anti-tumor [74].
TAM dapat berkontribusi untuk tumorigenesis dengan menginduksi hasil dari pemeriksaan kekebalan pada sel tumor. Sebagai contoh, TAM dapat menginduksi ekspresi program kematian-ligand 1 (PD-L1) [73]. Hal ini dapat melibatkan reseptor PD-1 pada sel T dan menghambat kemampuan mereka untuk menanggapi antigen tumor. Terapi yang memblokir PD-1 dan PD-L1 mungkin efektif untuk pengobatan berbagai jenis kanker. [75,76]. Secara umum, peningkatan TAM berkorelasi dengan prognosis buruk dari pasien [77,78]. Oleh karena itu, terapi yang menargetkan TAM atau mengubah fungsi mereka mungkin berguna untuk pengobatan kanker.
2.4. IDO
DC adalah sel antigen-digambarkan sebagai sel yang menghubungkan kekebalan bawaan dan adaptif dan telah terlibat dalam regulasi kekebalan kanker [79,80]. DC adalah pemain kunci dalam merangsang respon kekebalan anti-tumor. DC terpapar antigen dalam ketiadaan kostimulasi yang benar dapat menginduksi toleransi [81]. Fungsi tolerogenic dari DC telah dikaitkan dengan rendahnya tingkat molekul spesifik termasuk anggota keluarga B7 dan PD-L1, B7-H2, B7-H3, B7-H4 dan BTLA [82-89].
Toleransi kekebalan yang dimediasi oleh DC muncul untuk dimediasi oleh enzim yang tidak meregulasi fungsi dari efektor limfosit dalam antigen-independen. Ini termasuk nitrat oksida sintase yang diinduksi, yang menghasilkan oksida nitrat, arginase-1, yang menguras lingkungan arginin, dan IDO, yang mendegradasi triptofan asam amino esensial (TRP)
dan mengkatalisis generasi kynurenine (KYN) [90-95] .Sistem kekebalan tubuh dapat berfungsi sebagai
pelindung terhadap perkembangan tumor [96-100]. DC menyimpan IDO aktif yang telah terdeteksi dalam lingkungan mikro tumor atau menguras kelenjar getah bening [101-103]. Sel-sel ini dapat menekan fungsi sel T melalui IDO,mengaktivasi elevasi oleh dua mekanisme. Dalam kasus KYN, pada interaksi dengan reseptor aril hidrokarbon, molekul ini telah terbukti menghambat proliferasi sel T dan sel NK, mempromosikan regulasi sel T (Treg) diferensiasi, dan penghambatan DC imunogenisitas [90,91,104]. Selain itu, deplesi TRP cepat dari lingkungan mikronya mengirimkan sinyal stres pada sel-sel T, menginduksi anergi sel CD8 sitotoksik T dan mempromosikan diferensiasi CD4 terhadap Tregs [90,91,105].
Sel-sel tumor dapat mensintesis IDO. Tapi, itu tidak jelas apakah kontributor utama KYN generasi dan penipisan TRP dalam lingkungan mikro tumor adalah sel tumor atau infiltrasi leukosit, di DC atau TAM tertentu [90,91,106]. Terlepas dari sumber, aktivasi IDO dapat menginduksi imunosupresi. Tingginya kadar IDO berkorelasi dengan prognosis buruk [107-112]. Penghambatan IDO dapat menekan pertumbuhan tumor pada model tikus [113-117]. Oleh karena itu, IDO inhibitor mungkin berguna untuk menargetkan mikro tumor untuk pengobatan kanker.
2.5. 2.5 VEGFVEGF adalah regulator penting dari angiogenesis
tumor. Mereka meliputi keluarga (VEGF-A, -B, -C, -D, -E dan pertumbuhan plasenta faktor [PGF]) dari faktor pertumbuhan yang menunjukkan pola yang dilestarikan delapan cysteine residu [118-120]. Secara khusus, VEGF-A (molekul paradigmatik dari keluarga ini dan biasanya disebut sebagai VEGF) memiliki kemampuan untuk bertindak baik sebagai mitogen yang, merangsang proliferasi sel endotel, dan juga faktor kemotaktik dengan kemampuan untuk menarik monosit [118 -120].
Manusia dengan VEGF-A memiliki empat isoform berbeda (VEGF 121, 165, 189 dan 206), konsekuensi dari alternatif ekson splicing [118]. Sifat VEGF asli paling dekat sesuai dengan VEGF-165, yang merupakan VEGF-A isoform dominan[118]. VEGF berpartisipasi dalam proses fisiologis yang berbeda seperti angiogenesis, penyembuhan luka, dan embriogenesis [118-120]. VEGF telah ditunjukkan untuk berpartisipasi dalam proses patologis seperti diabetic retinopathy dan onkogenesis [118]. Tumor memerlukan faktor-faktor angiogenik untuk menginduksi pembentukan neovessels [121-123]. VEGF saja dapat memulai kaskade angiogenik [124]. VEGF disekresikan oleh sebagian besar kanker pada manusia [124], dan ekspresi VEGF dapat dikorelasikan dengan prognosis buruk pada ovarium [100] dan jenis-jenis tumor kanker [125-128].
VEGF berinteraksi dengan reseptor umum

(VEGFR-1, VEGFR- 2, VEGFR-3 dan neuropilin-1) [129]. Mereka terdiri dari keluarga reseptor tirosin kinase(RTKs) menunjukkan beberapa domain imunoglobulin-seperti dalam domain ekstraseluler, sebuah daerah transmembran tunggal dan konsensus tirosin urutan yang terganggu oleh domain kinase-insert [118-120]. VEGF-A juga berinteraksi dengan keluarga neuropilins dari co-reseptor.
VEGFR-1 memiliki afinitas yang sangat tinggi untuk VEGF-A [130]. VEGF-A mencegah mengikat VEGFR-2 [118]. VEGFR-1 mampu menginduksi mitokondria sinyal genic dan pro-survival di beberapa sel [131]. VEGFR-1 juga telah dikaitkan dengan induksi molekul angiogenik seperti metalloproteases matriks (MMPs) dan faktor pertumbuhan hepatosit (HGF) [132133]. VEGFR-1 juga dapat berpartisipasi dalam hematopoiesis, perekrutan progenitor endotel, dan migrasi monosit. Akhirnya, VEGFR-1 dapat heterodimerize dengan VEGFR-2, yang mengarah ke transactivation molekul ini [134].
VEGFR-2 menengahi mitogenesis dan angiogenesis [118-120]. Setelah ligan mengikat, VEGFR-2 dimerizes dan autophosphorylates di beberapa residu tirosin. Ligasi VEGFR-2 dengan hasil VEGF dalam fosforilasi protein yang berbeda seperti PI-3-kinase, RAS GTPase-activating protein, keluarga protein SRC, dan protein dari RAF-MEK-ERK jalur [119120135]. VEGFR- 2 signaling dapat mempromosikan kelangsungan hidup sel endotel, proliferasi dan angiogenesis. Dengan demikian, VEGF dan reseptornya dianggap molekul kunci dalam proses neovaskularisasi dan pertumbuhan akibat dari banyak tumor.
VEGF telah menjadi target untuk terapi antitumor [136]. Sebuah antibodi monoklonal manusiawi yang menargetkan VEGF (beva- cizumab / Avastin) telah disetujui untuk pengobatan kanker kolorektal yang berbeda, kanker ginjal, kanker paru-paru atau glioblastoma [137]. Beberapa studi menyoroti kemanjurannya sebagai bagian dari therapi kombinasi [138-140]. Aflibercept / VEGF-trap dapat bertindak sebagai reseptor umpan untuk VEGF. Senyawa ini memiliki khasiat antitumor [137,141-144]. Akhirnya, RTK inhibitor seperti sunitinib dan sorafenib memiliki aktivitas pada kanker lambung, kanker ginjal, tumor pankreas atau kanker hati [137.145].
2.6. FibrosisJaringan fibrosis umumnya diamati dalam tumor
lingkungan mikronya terkait dengan proliferasi cepat dari fibroblas [67]. Fibroblas juga dapat mengeluarkan berbagai sitokin dan kemokin seperti TGF-, IL-1, IL-6, IL-8, CXCR4, CXCL12, dan monosit-chemotactic protein 1 (MCP-1) [146] dan faktor platelet pertumbuhan yang berasal ( PDGF), HGF, stroma sel-diturunkan faktor 1 (SDF1), VEGF, dan faktor pertumbuhan fibroblast dasar (bFGF) [147](CAF).
Kanker terkait fibroblast sering dikaitkan dengan biologi tumor lebih agresif karena sekresi MMPs yang meningkatkan pemecahan ECM dan membantu dalam sel-sel kanker menuju ke pembuluh darah dan metastasis ke tempat yang jauh [148]. MMPs juga terlibat dalam menginduksi epitel untuk mesenchymal transision (EMT), sebuah proses yang memicu diferensiasi sel-sel kanker asal epitel ke dalam sel mesenchymal dengan sifat stemness. EMT mungkin biomarker dari prognosis yang buruk [149].Fibroblast dapat dikaitkan dengan hasil klinis yang lebih buruk pada pasien dengan berbagai jenis kanker [150-152]. Dengan demikian, penghambatan ditargetkan fibroblas mungkin berguna untuk mengobati kanker.
Namun, beberapa studi menunjukkan peran yang lebih kompleks untuk fibroblast di tumorigenesis. Menargetkan fibroblast mengaktifkan protein (FAP) tidak menghasilkan regresi tumor tetapi dikaitkan dengan toksisitas sumsum tulang [153]. Penghapusan ditargetkan halus aktin otot myofibroblasts positif khususnya terkait dengan tahap pra-ganas karsinoma pankreas (pankreas intraep- neoplasia ithelial) [154]. Hal ini menyebabkan fenotipe tumor lebih buruk dibedakan dan agresif. Oleh karena itu, fibroblas dan myofibroblasts tampaknya memainkan peran penting dalam pembentukan matriks ekstra-selular dan merangsang fibrosis dalam tumbuh tumor.
2.7. 2.7 IL-6IL-6 merupakan sitokin inflamasi yang terkait
dengan respon imun bawaan dan pertahanan terhadap infeksi, tetapi baru-baru ini ditemukan untuk memainkan peran dalam lingkungan mikro tumor. Makrofag, monosit dan sel T dapat menghasilkan IL-1 dan TNF [155]. Disregulasi IL-6 dikaitkan dengan penyakit inflamasi, seperti rheumatoid arthritis, resistensi insulin, sepsis dan kanker [155.156].
Sinyal dari IL-6 terjadi melalui kolaborasi dari reseptor membran-terikat (IL-6R / gp80) dan sinyal transduser glikoprotein (gp130), reseptor untuk sitokin seperti IL-11 dan IL-27 [155-158]. Ekspresi permukaan IL-6R terbatas sebagian besar sel-sel kekebalan tubuh dan hepatosit. Namun, gp130 dinyatakan sebagai banyak jenis sel, termasuk endotel dan tumor sel [157,159-161]. Bentuk larut dari IL-6R (sIL-6R) mampu berinteraksi dengan IL-6 dalam larutan dan kemudian menghubungi membran sel untuk menginduksi sinyal melalui gp130. Dengan demikian, sel-sel lacking pada membran terikat IL-6R masih dapat dipengaruhi oleh IL-6 dihasilkan di lingkungan mikro [157,159-162]. IL-6 mengaktifkan JAK dan transduser sinyal dan aktivator transkripsi (JAK / STAT) mengaktifkan STAT3 [155163164]. STAT3 menyebabkan kelangsungan hidup sel kanker,proliferasi, dan metastasis; itu juga mempromosikan angiogenesis dan ekspresi faktor penekan kekebalan dalam lingkungan mikro tumor [165]. IL-6 dapat meningkatkan pertumbuhan kanker

payudara [166], glioma [167], limfoma [168], multiple myeloma [169], kanker ovarium [170], dan kanker prostat [171]. Tingginya kadar larut IL-6 atau tinggi tingkat pewarnaan IL-6 dalam sampel tumor berkorelasi dengan hasil yang buruk [172-181]. Akhirnya, IL-6 dapat menginduksi produksi VEGF pada sel kanker atau sel tumor terkait [182-184]. Dengan demikian, IL-6 dapat mempromosikan tumorigenesis melalui banyak mekanisme.
IL-6 telah menjadi terapi target. Anti-IL6 antibodi (siltux- imab / CNTO 328) meningkatkan efek sitotoksik obat kemoterapi seperti paclitaxel atau melphalan [185-187] dan penurunan pertumbuhan tumor, infiltrasi makrofag dan angiogenesis [185]. Siltuximab sendiri atau dalam kombinasi dengan obat sitotoksik telah dipelajari pada pasien manusia [188-190]. Beberapa efek diamati bila digunakan dalam terapi kombinasi [191.192]. Demikian pula, antibodi manusiawi anti-IL-6R, tocilizumab, telah terbukti menghambat IL-6 sinyal dalam sel-sel kanker dalam studi praklinis [193-195]. antibodi ini telah digunakan untuk pengobatan peradangan [196] dan cachexia [197].
2.8.2.9.
2.8.Endoglin
Endoglin (CD105) adalah faktor pertumbuhan glikoprotein co-reseptor homodynamic untuk TGF- dalam jaringan endotel yang memainkan peran penting dalam angiogenesis dan remodeling vaskular [198.199]. Endoglin memodulasi Smad fosforilasi dan dapat mengontrol adhesi sel dan migrasi dengan mengatur komposisi kompleks adhesi fokus dan dapat mengatur angiogenesis. Penyimpangan dari fungsi coreseptor yang sangat penting untuk banyak proses sel yang terlibat dalam kanker [200]. Peradangan dan angiogenesis tumor mungkin terkait akibat dari disregulasi fungsi Endoglin co-reseptor [201]. Ekspresi Endoglin diamati di neo-angiogenesis, perkembangan tumor dan metastasis [202]. Cara menghambat jalur Endoglin mungkin berguna untuk pengobatan kanker [203.204].Antibodi TRC105 memiliki aviditas tinggi untuk Endoglin-mengikat dan mungkin memiliki aktivitas sebagai agen tunggal maupun dikombinasikan kemoterapi dengan bevacizumab [205] dan mungkin mengatasi resistensi terapi untuk bevacizumab [206].
2.9 JAKKeluarga JAK termasuk reseptor terkait tirosin
kinase JAK1, JAK2, JAK3 dan Tyk2 yang regulator penting dari banyak proses signaling normal yang telah terlibat kausal dalam tumorigenesis [207.208]. JAK jalur umumnya penting untuk sinyal sel normal [209.210]. Di antara contoh klasik adalah JAK-dimediasi STAT3 tirosin fosforilasi dalam menanggapi
sitokin IL-6 keluarga (termasuk IL-11) sinyal melalui gp130 [211].
Mutasi pada JAK2 dan lebih umum di SOCS1 telah terlibat dalam tumorigenesis [212]. Jalur JAK / STAT3 adalah konstitutif aktif dalam sel-sel tumor [213214] dan dalam tumor terkait sel stroma [163.164.184.215.216]. IL-6 -. Dimediasi JAK/STAT sinyal parakrin umumnya diamati pada kanker [217-219]
JAK sinyal penting untuk interaksi tumor-sel tubuh di lingkungan mikro. Dalam kanker kepala dan leher, IL-6 menengahi EMT dan meningkatkan potensi metastasis sel berubah [220]. Dalam Waldenstrom macroglobulinemia, ekspresi CCL5 dysregulated memodulasi IL-6 sekresi pada sel stroma, mengakibatkan peningkatan sekresi IgM oleh sel-sel ganas melalui JAK / STAT jalur [221]. Pankreas sel stellata terkait kanker mensekresi IL-6 dan faktor larut lain yang mempromosikan akumulasi sel penekan myeloid yang diturunkan melalui JAK / STAT3 tergantung mekanisme [222]. Karsinogenesis paru-paru, AZD1480 menghambat aktivasi STAT3 di tumor- sel myeloid terkait, mengurangi jumlah sel, menghambat metastasis tumor, dan myeloid angiogenesis sel-dimediasi .AZD1480 memblok angiogenesis, infiltrasi paru-paru sel myeloid dan pembentukan metastasis paru pada tikus syngeneic eksperimental dan spontan model metastasis serta dalam xenograft manusia. Aktivasi STAT3 dalam sel kanker sudah cukup untuk mengatasi mikro-dimediasi dan AZD1480-menghambat perkembangan kanker paru-paru [223].
Efek terapi dari TLR4 dan TLR9 kompleks agonis terhadap melanoma metastasis bergantung pada penggunaan simultan inhibitor dari JAK / STAT jalur (seperti AG490 onist antag-). Terapi gabungan seperti mengaktifkan kematian autophagy terkait sel melanoma via IFN-Ɣ/aktivasi STAT1 dan penghambatan diciptakan tumor metastasis [224]. Dalam karsinogenesis Barrett, IL-6 memblokir antibodi dan AG490 dan JAK inhibitor blok STAT3 fosforilasi menurunkan resistensi terhadap apoptosis [225]. JAK / akumulasi STAT3 tergantung dari sel Treg pada tumor tergantung pada peningkatan protein S1PR1 dalam sel CD4 + T, sedangkan jalur penghambatan JAK / STAT3 dalam sel T berkurang karena akumulasi dari sel-sel Treg pada tumor dan pertumbuhan tumor. Migrasi Treg menuju tumor hampir sepenuhnya diblokir oleh AZD1480 [226]. Sinyal gp130-IL6ST / JAK1 menghasilkan actomyosin contractility melalui Rho-kinase tergantung sinyal di kedua sel-sel tumor dan sel-sel stroma. Oleh karena itu, penghambatan jalur JAK dapat berguna untuk memodulasi tumorigenesis melalui banyak mekanisme, termasuk menargetkan lingkungan mikro tumor.
3. Pendekatan

3.1 3.1 BerberineBerberin (Gambar. 2) adalah amonium kuaterner
garam dari kelompok protoberberine alkaloid isoquinoline dengan sifat umum anti neoplastik [227.228]. Berberin memiliki bioavailabilitas rendah dengan kurang dari 5% dari dosis yang tertelan menemukan jalan ke dalam sirkulasi sistemik [229230]; pada tikus nilai jauh lebih rendah (0,68%) [231]. Pada manusia, dosis 1000-1500 mg per hari telah terbukti efektif dalam hal dampak berberin pada profil glucidic dan lipid pada pasien dengan hypercholesteremia dan diabetes tipe 2. Produk dimetabolisme dari berberin juga bertindak sebagai senyawa asli tapi dengan potensi kurang [232.233]. Bentuk umum dari produk ekskresi kemih dari berberin diyakini jatrorrhizine [234.235].
Sebagai obat tradisional atau suplemen makanan, berberin telah menunjukkan aktivitas terhadap jamur, Candida albicans, parasit, dan infeksi virus / bakteri [236]. Utilitas klinis telah dinilai untuk banyak penyakit dan kondisi termasuk hyperlipidemia, diabetes, obesitas dan penyakit hati berlemak. Saat ini ada 17 yang selesai dan studi klinis yang sedang berlangsung terdaftar mengenai berberin (www.clinicaltrials.org).
Interaksi Berberin dengan berbagai jalur metabolik telah diteliti secara luas. Adenosine mono-fosfat kinase (AMPK) adalah protein sensor nutrisi; berberin mengaktifkan AMPK dalam dosis dan waktu tergantung caranya [237.238]. Data menunjukkan bahwa berberin-diinduksi AMPK menghambat kompleks I dari mitokondria rantai transpor elektron [239]. Efek ini juga diamati dengan metformin obat anti-diabetes dan rosiglitazone. Metabolisme lipid, efek penurun lipid dari berberin diyakini terkait dengan stabilisasi hati LDL-C reseptor (LDLR) oleh jalur ERK-dependent dan juga oleh peningkatan nasional aktivitas transkripsi dari LDLR promotor oleh c-Jun N kinase terminal (JNK) [240241]. Dalam 3T3L1 sel, berberin telah terbukti mengurangi enzim adipogenic kunci dalam vitro seperti sintase asam lemak, acetyl koenzim A (asetil-CoA) karboksilase, asil-CoA sintase, dan lipoprotein lipase [242]. Selanjutnya, berberin telah ditunjukkan untuk menghambat kolesterol dan sintesis trigliserida dalam sel hati melalui aktivasi AMP kinase [243].
Efek anti-neoplastik Berberin terpilih telah dicatat [244.245]. Berberin muncul untuk menekan peradangan dalam menanggapi rangsangan pro-inflamasi [246]. Berberin pada konsentrasi 10-20 mcg / mL, in vitro, telah terbukti sedikit meningkatkan proliferasi sel T dalam menanggapi antigen, sedangkan konsentrasi di atas bahwa hasil tingkat di imunosupresi tergantung dosis [247]. Penghambatan selektif JAK3 oleh berberin juga dapat memediasi imunosupresi [248]. Berberin memberikan efek anti-tumor yang melalui berbagai mekanisme yang meliputi penghambatan proliferasi sel, induksi apoptosis, dan penindasan angiogenesis dan metastasis tumor. Efek
imunomodulator berberin ini, melalui JAK3 inhibisi, mungkin juga berdampak pada pertumbuhan kanker [248]. Berberin telah ditemukan untuk meningkatkan sitotoksisitas doxorubicin, yang menunjukkan bahwa agen ini mungkin memiliki potensi sebagai tambahan untuk beberapa agen kemoterapi tradisional [249]. Efek sitotoksik berberin telah ditunjukkan untuk berbagai tumor termasuk paru-paru, payudara, prostat, neuroblastoma kolorektal, limfoma, koma osteosar- dan leukemia [228].
Berberin memiliki efek apoptosis caspase-independen pada IMCE usus sel kanker tetapi tidak sel normal colonocyte, YAMC [250]. Berberin juga menginduksi sitotoksisitas melalui G1-fase siklus sel penangkapan dan caspase-3-dependent apoptosis pada glioblastoma, epidermoid dan prostat sel karsinoma [251-253]. Efek apoptosis berberin dikaitkan dengan peningkatan regulasi gen apoptosis pro Fas, FasL, p53, dan Bax [254-256]. Berberin memiliki efek anti-angiogenik terkait dengan penurunan ekspresi MMP-1, MMP-2 dan MMP-9 [257-259].
Sebagai surfaktan, kelarutan berberin adalah rendah. Penggunaan Berberin terhambat oleh bioavailabilitas rendah yang terkait dengan biotransformasi cepat selama periode panjang yang tetap dalam usus. Berbagai sistem pengiriman nano-partikel telah digunakan untuk meningkatkan penyerapan berberine, termasuk metode rotary-evaporated film ultrasonication [260], nanoemulsification dengan isopropil miristat / gliserin [261], dan penggabungan lipisomal [262]. Berberin diproduksi dengan sistem pengiriman nanoparticulate menunjukkan peningkatan bioavailabilitas dan optimalisasi anti-inflamasi, anti angiogenik [263], dan efek antikanker [264]. Berberin memiliki potensi sebagai agen anti-kanker [250]. Dasar molekul efek neoplastik, bagaimanapun, perlu diteliti lebih lanjut.Resveratrol
Resveratrol (3,4, 5-trihidroksi-trans-stilbene) adalah senyawa polifenol yang fungsional milik phytoalexins dengan aktivitas anti-ROS. Resveratrol dihasilkan melalui stilbene synthase [265] dalam menanggapi patogen infeksi [266] atau stres kondisi [267] menggunakan malonyl-koenzim A (CoA) dan p-coumaroyl CoA sebagai sors precur-. Senyawa ini mungkin memiliki potensi untuk pencegahan dan pengobatan kanker[268].
Resveratrol terjadi secara alami di lebih dari 70 spesies tanaman termasuk kacang, blueberry, raspberry, mulberry, pinus, dan anggur [269]. Tingkat yang relatif tinggi resveratrol hadir dalam kulit anggur segar, yang menjelaskan konsentrasi tinggi dalam anggur merah dan jus anggur [270]. Bentuk terkonjugasi yang berbeda dari resveratrol yang terdeteksi pada tanaman. Trans-resveratrol ada dalam bentuk glikosilasi dan memiliki isomer cis dan trans. Konjugasi lainnya, termasuk kelompok 1-2 metil,

kelompok sulfat, dan asam lemak, juga diamati [270]. Glikosilasi meningkatkan stabilitas, kelarutan dan penyerapan di saluran pencernaan manusia. Selain itu, melindungi resveratrol dari degradasi oksidatif [271].
Resveratrol metabolisme dalam tubuh manusia termasuk konversi untuk larut dalam air trans-resveratrol-3-O-glukuronida dan trans-resveratrol-3-O-sulfat dengan fase-2 hati enzim obat-metabolisme [272]. Produk-produk metabolik memiliki paruh plasma sekitar 9,2 jam, yang secara signifikan lebih tinggi daripada paruh resveratrol (8-14 menit) [272]. Konsentrasi antara 32 nM dan 100 M yang digunakan untuk studi in vitro yang berbeda, sedangkan konsentrasi 100 ng sampai 1500 mg / kg (berat badan) yang digunakan dalam studi hewan [273]. Resveratrol dan produk metabolismenya terdeteksi dalam hati, lambung, ginjal, empedu dan urin setelah administrasi oral tunggal 14C-trans-resveratrol di Balb / c tikus [274], sedangkan 24,6% dari resveratrol dan metabolitnya terdeteksi di urine manusia setelah pemberian oral [275].
Aktivitas biologis dikaitkan hanya dengan bentuk trans, yang merupakan pengambil radikal bebas [276]. Yang normal seluler respirasi, tekanan lingkungan, dan radiasi UV adalah pemicu utama produksi ROS. Ketidakseimbangan dalam rasio antara oksidatif pasangan redoks dized dan mengurangi seperti glutathione (GSH / GSSG) atau NADPH / NADP + penyebab ROS akumulasi [277]. Tingginya kadar ROS bereaksi dengan komponen seluler termasuk DNA, protein, dan lipid yang menyebabkan kerusakan sel dan jaringan [278]. Resveratrol dan lainnya diet dengan stilbenes mengurangi stres oksidatif dengan bertindak baik sebagai penangkap langsung dari ROS [279] atau sebagai inhibitor ekspresi NADPH oksidase dan aktivitas xantin oksidase [280]. Resveratrol memiliki toksisitas rendah [281]. Berbagai penelitian melaporkan efek anti-kanker [282-284], termasuk penindasan metastasis [285] dan induksi penangkapan pro liferative [286].
Sel-sel normal memiliki enzim antioksidan dan molekul yang menjaga ROS di bawah tingkat fisiologis normal [287]. Dalam sel-sel kanker,sinyal onkogenik merangsang metabolisme sel aktif, yang meningkatkan produksi ROS dan menyebabkan stres oksidatif permanen [288]. Selain itu, tumor terkait kerusakan mitokondria menyebabkan peningkatan besar dalam produksi ROS [289]. Resveratrol menghambat ROS dan mengurangi stres oksidatif [290]. Ini mengurangi produksi intraseluler ROS dan stres oksidatif oleh mekanisme yang melibatkan degradasi dari Keap 1 protein, yang merupakan penekan Nrf2 [291]. Dalam model tikus hepatocarcinogenesis, resveratrol ditemukan upregu- akhir hati Nrf2 [292]. Dalam studi lain, total kadar oksidan dalam plasma, hati dan otak yang menurun dan jumlah antioksidan di organ-organ ini meningkat pada tikus diobati dengan resveratrol
[293]. Selain itu, resveratrol mengurangi stres oksidatif dan mempertahankan fungsi mitokondria terkandung melalui kemampuannya untuk mengaktifkan sirtuin 1 (SIRT1), yang memiliki banyak peran dalam mengurangi stres oksidatif dan mempromosikan fungsi mitokondria [294]. Selain itu, penurunan serum dan stres oksidatif hati dalam diet tinggi lemak [295] dan tikus diabetic [296]. Resveratrol merupakan kandidat untuk pengobatan dan pencegahan kanker yang berbeda dengan penghambatan ROS.
Onionin A
Onionin A adalah produk alami dalam sayuran Allium yang baru-baru ini telah diidentifikasi sebagai agen potensial untuk mengatur aktivitas makrofag yang bisa memiliki aktivitas anti-neoplastik. Konsumsi Allium sayuran dikaitkan dengan penurunan risiko beberapa kanker. Sebuah studi epidemiologi Eropa melaporkan penurunan 55-80% dari odds rasio hampir semua kanker utama, termasuk mulut, esofagus, laring, kolorektal, prostat, payudara dan kanker ovarium, pada populasi yang sering mengkonsumsi dalam jumlah yang cukup besar dari bawang atau bawang putih dalam makanan mereka [297].Sayuran termasuk bawang, bawang putih, kebocoran, daun bawang dan bawang milik keluarga Allium. Sebelumnya diidentifikasi senyawa bioaktif dalam bawang (Allium cepa) adalah flavonoid dan fenol [298]. Flavonoid merupakan keluarga terbesar dari senyawa polifenol dan dengan nama "polifenol" dan "flavonoid", lama kelamaan, digunakan secara bergantian. Senyawa ini dipercaya untuk membatasi dan mencegah perkembangan kanker dari sel-sel yang rusak melalui efek antiinflamasi mereka [299-302].
Efek sitotoksik ekstrak polifenol bawang yang diturunkan baru-baru ini diselidiki. Ekstrak polifenol dari A.cepa dapat menginduksi caspase apoptosis tergantung dari sel-sel kanker lambung manusia melalui jalur mitokondria oleh peningkatan regulasi p53 dan protein Bax serta dengan memodulasi Bcl-2 protein. Selanjutnya, onion berasal ekstrak polifenol diinduksi apoptosis caspase-tergantung dari beberapa baris sel leukemia manusia in vitro telah dikaitkan, setidaknya sebagian, penghambatan sinyal jalur PI3K / AKT [303]. Sifat antioksidan dan antimutagenik ekstrak bawang terhadap mutagen terkait dengan polifenol dan flavonoid mereka [304]. Lipid senyawa organosulfur yang larut hadir dalam ekstrak bawang menghambat proliferasi berbudaya usus manusia, kulit dan tumor sel paru-paru [305]. Salah satu mekanisme yang mungkin untuk menghambat aktivasi karsinogen oleh ekstrak derivatif bawang mungkin penghambatan sitokrom P450 2E1, yang diaktifkan oleh sejumlah zat xenobiotik [306].
Bawang juga kaya akan senyawa organosulfur. Hal ini phytochemical, termasuk diallyl disulfide, S-allylcysteine dan ajoene, melindungi terhadap kanker secara kimiawi pada model binatang dengan mengubah metabolisme karsinogen [307-310]. Baru-baru ini, onionin A dimurnikan [311] dan diidentifikasi

sebagai 3,4-dimetil-5-(1E-propenil) - tetrahydrothiophen-2-sulfoxide-S-oksida. Onionin A dapat menghambat TAM [68.312]. Efek toksik dari onionin A pada IL-10 yang diinduksi elevasi mengaktivasi M2 makrofag dengan menilai ekspresi
unik M2 penanda CD163. Onionin A secara signifikan menekan ekspresi dari CD163 pada konsentrasi 10 dan 30 µM. Hasil ini menunjukkan bahwa onionin A dapat menekan proliferasi sel tumor. Agen ini mungkin berguna sebagai agen anti-kanker.
3.4 EGCG
EGCG menghambat ekspresi IDO pada sel kanker manusia. Konsumsi teh hijau, yang dihasilkan dari daun tanaman Camellia sinensis, telah dikaitkan dengan insidensi yang lebih rendah pada kanker manusia [313]. Teh hijau mengandung banyak polifenol, di EGCG ini secara khusus, yang telah terbukti untuk menekan pembentukan tumor dan perkembangan pada hewan model [314]. The kemopreventif dan terapi efek dari EGCG yang dikaitkan dengan spektrum luas kemampuan anti-kanker polifenol ini, termasuk penghambatan proliferasi, peradangan, apoptosis, dan angiogenesis [314.315].
EGCG juga telah ditemukan untuk menghambat ekspresi IDO , yang merupakan enzim kunci dalam menekan sel T dan merangsang toleransi kekebalan sel tumor melalui menipisnya tryptophan. Banyak jalur sitokin-dependent dan signaling independen yang terlibat dalam ekspresi IDO. Interferon-dirangsang IDO elevasi mengaktivasi adalah, bagaimanapun, dimediasi oleh JAK / STAT signaling jalur [316]. Ada sejumlah bukti yang menunjukkan bahwa Feres EGCG antar- dengan JAK / STAT-diatur aktivasi IDO, mengakibatkan penindasan IDO dan IDO terkait ekspresi gen hilir dalam sel kanker manusia.
EGCG telah terbukti untuk menekan ekspresi IDO melalui menghambat IFN -Ɣ diinduksi di lini sel kanker mulut manusia [317]. Translokasi STAT1 ke dalam inti, yang akibatnya menghambat aktivasi transkripsional IDO, diblokir oleh EGCG. Chen et al. [317] juga menunjukkan bahwa EGCG secara signifikan menekan fosforilasi protein kinase C (PKC-) dan Jaks, mengakibatkan penghambatan IFN-Ɣ-distimulasi STAT1 fosforilasi. Serupa, kelompok lain menunjukkan bahwa EGCG blok Ido ekspresi pada kanker kolorektal manusia pada tingkat transkripsi melalui penghambatan STAT1 fosforilasi, yang akibatnya mendukung penekanan aktivitas STAT1 yang diaktifkan elemen urutan promotor IDO, IFN-dirangsang elemen respon ( ISRE) dan IFN-Ɣ-. urutan aktivasi (GAS) [318]
EGCG diteliti menunjukkan aktivitas anti-IDO dalam sumsum yang diturunkan sel dendritik tulang murine (BMDCs) [319]. EGCG diblokir oleh pengikatan STAT1 terfosforilasi untuk INF regulasi faktor-1 (IRF-1) promotor, dalam menanggapi rangsangan IFN- Ɣ. Hasil dari prostaglandin E2 (PGE2) dan cyclooxygenase (COX-2) juga secara signifikan dihambat EGCG-diobati BMDCs murine. Lebih dari ekspresi PGE2, lipid bioaktif, dan COX-2, enzim kunci dalam biosintesis prostaglandin, sering dikaitkan dengan pengawasan kekebalan dan kanker [320]. Efek penghambatan EGCG pada COX- 2 ekspresi juga telah terlihat di lini sel kanker lainnya seperti kanker prostat manusia dan karsinoma kolon [321.322]. Dalam sebuah studi in vivo, Ogawa et al. [323] menunjukkan efek EGCG pada azoxymethane (AOM) -diinduksi lesi preneoplastic di F344 tikus melalui penekanan ekspresi IDO. EGCG-tikus diperlakukan dibiarkan secara signifikan mengurangi tingkat menyimpang crypt fokus, yang berlebih dari IDO. Ekspresi mRNA COX-2 di AOM- diperlakukan pada tikus juga dihambat oleh pengobatan EGCG [323].
EGCG menghambat jalur sinyal JAK / STAT. Pra-pengobatan dengan EGCG menyebabkan penekanan STAT1 fosforilasi dan IRF-1 ekspresi baris sel kanker yang berbeda seperti mammae karsinoma, karsinoma serviks, dan hepatocarcinoma [324.325]. STAT3 dikaitkan dengan ekspresi IDO konstitutif pada sel kanker manusia [326]. EGCG menghambat fosforilasi dan ekspresi dari kedua JAK3 dan STAT3 protein dalam sel-sel kanker pankreas [327]. EGCG mengurangi kadar protein terfosforilasi-STAT3 dirangsang oleh faktor insulin-like growth (IGFs) dalam sel karsinoma hepatoseluler, mungkin melalui menghambat bioavailabilitas IGFs [328]. EGCG menghambat STAT3 di kepala dan leher kanker [329] dan bisa- payudara cer [330]. Penghambatan JAK / STAT jalur melalui EGCG mungkin berguna untuk mengatur IDO.
EGCG memiliki sifat anti-kanker. Kemampuan EGCG bertindak sebagai agen multi-target dalam

mengatur sinyal JAK / STAT dan JAK / STAT-dimediasi IDO luar biasa. Khasiat mengkombinasikan EGCG dengan sejumlah obat kemoterapi seperti tamoxIfen dan paclitaxel telah menunjukkan efek sinergis [331], EGCG merupakan kandidat sebagai terapi kanker dengan menargetkan IDO.
3.4 3.5 Genisteingenistein (4, 5,7-trihidroksiisoflavon), sebuah
isoflavon polifenol, ditemukan dalam produk kedelai dan muncul untuk memodulasi metabolisme kolesterol. Ini memiliki bioavailabilitas rendah karena kelarutan miskin air, metabolisme ekstensif usus tahap pertama-pass II, dan ekskresi berikutnya metabolit terkonjugasi mereka [332]. Studi epidemiologil menunjukkan bahwa diet asupan kaya kedelai dapat menurunkan kejadian kanker payudara dan prostat di negara-negara Asia [333.334]. Genistein dapat memediasi efek anti-kanker melalui faktor inti (NF) -K B modulasi, pengurangan tingkat protein AKT, penurunan regulasi karsinogenesis androgen-dimediasi, dan / atau efek antioxidation lebih umum [334.335]. Genistein berpotensi untuk aktivitas anti-kanker terhadap prostat [336337], ovarium [338], payudara [339], paru-paru [340], dan kanker pankreas [341].
Peningkatan aktivitas HMGR, mevalonate, dan prenilasi protein dikaitkan dengan tumorigenesis [342]. Genistein telah ditunjukkan untuk menekan HMGR dan menghambat pertumbuhan tumor [343.344]. Genistein juga dapat meningkatkan reseptor LDL dan menurunkan ekspresi HMGR dalam sel kanker usus besar [345]. Genistein memiliki banyak efek lain pada metabolisme lipid yang dapat berkontribusi terhadap sifat anti-neoplastik [346]. Namun, penting untuk dicatat bahwa genistein mungkin memiliki efek pro-proliferasi dalam beberapa konteks [347-350].
3.5 3.6 Curcumin
curcumin (diferuloylmethane), bahan aktif dari rempah-rempah kunyit dari tanaman Curcuma longa, milik kelompok senyawa herbal polifenol dan memiliki beberapa efek yang menguntungkan termasuk tindakan anti-tumorigenic yang tampaknya berkaitan dengan penghambatan VEGF. Bubuk kunyit banyak digunakan dalam pengobatan Ayurveda, Unani, dan Siddha sebagai obat rumah untuk berbagai penyakit [351]. Selain kurkumin, kunyit mengandung fraksi ringan seperti demethoxycurcumin, bisdemethoxycurcumin, dan cyclocurcumin [352]. Curcumin telah terlibat sebagai penekan inisiasi tumor dan promosi, angiogenesis, dan metastasis [353.354].
Kurkumin menurunkan regulasi ekspresi VEGF pada sel kanker prostat secara dosis-dan tergantung waktu [355]. Osteo Pontin / integrin avb3 sinyal melalui aktivasi MMP9 meningkatkan VEGF dan ekspresi Angiostatin pada sel kanker prostat dan sebaliknya
kurkumin mengurangi ekspresi VEGF [355], menekan aktivitas MMP9 di sel-sel kanker prostat. Kurkumin yang diturunkan secara analogi CDF menghambat VEGF serta IL-6 dan gen kanker stem sel signature genes Nanog, Oct4, dan EZH2 in vitro dan in vivo [356]. Demikian pula, CDF berkurang VEGF dan IL-6 ekspresi dalam sel-sel kanker prostat [357]. Curcumin menghambat migrasi dan invasi sel kanker paru-paru manusia melalui penghambatan MMP-2 dan MMP-9 dan penekanan ekspresi VEGF [358]. Paparan jangka panjang curcumin diselidiki dalam hati dari tikus bearing yang limfoma. Pengobatan Curcumin menginduksi aktifasi gen aktif stres HIF- 1a, MYC, dan aktivitas LDH ke tingkat normal. Selain itu, menyebabkan penghambatan angiogenesis yang dibuktikan dengan berkurangnya MMP-2, MMP-9, PKC-dan kadar VEGF [359].
Mirip dengan IL-6, IL-1 sinyal sangat penting untuk inflamasi dan keganasan proses. IL-1 diinduksi IKB alpha fosforilasi dan penghambatan hilir NF B, yang mengarah ke ekspresi gen beberapa wakil yang berkaitan dengan proliferasi sel, invasi dan angiogenesis [360]. Pengobatan kurkumin diblokir IL-1 dan VEGF ekspresi dalam sel chondrosarcoma. Selanjutnya, kurkumin menghambat IL-1 beta-induced angiogenesis dan gen NF B terkait expression sion [361]. Kurkumin merupakan salah satu unsur utama bumbu kunyit, yang telah digunakan selama berabad-abad. Fase 1 percobaan manusia dengan 25 mata pelajaran menggunakan hingga 8000 mg kurkumin per hari selama 3 bulan tidak menemukan toksisitas, dan secara keseluruhan telah dianggap aman dalam enam percobaan manusia [362]. Curcumin memiliki sangat rendah bioavailabilitas sistemik, karena kelarutan air rendah dan stabilitas miskin.
NaringeninNaringenin memiliki prospek yang baik
sebagai agen terapi yang ideal vis à-vis pengaruh pada metastasis dan efek tertentu pada fibrosis [363]. Naringenin secara signifikan mengurangi metastasis paru-paru pada tikus (C57BL / 6 dan BALB / c) dengan fibrosis paru dan meningkatkan kelangsungan hidup mereka dengan meningkatkan lingkungan fibrotik-imunosupresif dan mengurangi sel-sel T regulator [364]. Dalam sel HSC-T6, naringenin diberikannya efek antifibrogenic dengan langsung atau tidak langsung mengatur turunnya ekspresi protein Smad3 dan fosforilasi melalui sinyal TGF-beta [365] dan menurunkan regulasi dari vimentin, cadherin N-, MMP2 dan MMP9 [366]. Naringenin menghambat pembentukan virus dan dalam jangka panjang produksi partikel menular hepatitis C [367]. Ini memiliki agonistik [368] dan

kegiatan antagonistik terhadap estrogen [369]. Ini juga mungkin memiliki nilai pelindung terhadap kontaminan makanan bisphenol A [370]. Oleh karena itu, naringenin diberikannya efek penghambatan pada pertumbuhan sel kanker, migrasi dan invasi, dan juga memiliki properti kemopreventif.
Naringenin memiliki bioavailabilitas yang rendah (2,8% dari asupan) karena ekskresi tinggi dan rendah C max dalam plasma. Prinsip metabolisme, naringenin-7-0-glucoronide dan naringenin-4’-0-glucoronide puncaknya pada 6 jam sesudah intake. Distribusi dari metabolis juga bervariasi karena mengikat serum albumin. Modulasi tersebut mempengaruhi half-life dari plasma dan distribusi jaringan. Sebuah NRG glucoronide afinitas untuk serum albumin sama seperti naringenin. Upaya untuk meningkatkan bioavabilitas melaui berbagai teknik seperti menggabungkan NRG dengan β-cyclodextrin melalui dispersi padat dan teknik Self Nano Emulsifying Drug (SNEDDS) [373]. Dari campuran dan nanopartikel ditingkatkan secara signifikan terjadi peningkatan dalam perbandingan absorbsi NRG untuk sebuah NRG. Bagian bawah dari waktu konsentrasi obat (AUC) 0-24 jam yang secara signifikan meningkat untuk SNEDDS yang di gabungkan dengan obat murni. Bahkan nanopartikel dimuat dalam NRG menunjukkan peningkatan peroksidatif anti lipid yang menimbulkan efek antiproliferatif dan potensial antioksidan untuk meningkatkan afinitas kemopreventif kombinasi untuk free naringenin dalam 7,12-dimethylbenz(a)anthtacene (DMBA)- menginduce oral karsinogenesi[374]. Naringenin flavonoid mempresentasikan GJF dalam berbagai interaksi dengan obat untuk mengurangi efek tarapetik atau meningkatkan efek samping yang kompetitif atau mekanisme inhibisi dari isoenzim dinding usus CYP3A4, P-glukoprotein yang banyak resisten terhadap protein obat, dan anion organik mentrasnfer inhibisi polipeptida. Sangat aman bila dievaluasi secara in vitro dan in vivo [375,376].
3.8 DesoxyrhapontigeninDesoxyrhapontigenin (3,5-dihidroksi-4* O-
methylresveratrol) adalah antioksidan [377]. Desoxyrhapontigenin mungkin berguna dalam modulasi lingkungan mikro tumor. Menghambat enzim sitokrom P450 [378,379], peradangan, ROS, dan jaringan terkait [380-385].
Desoxyrhapontigenin mempengaruhi ROS dan peradangan. Hal ini terjadi melalui peningkatan ekspresi enzim antioksidan [383] dan penghambatan NF B dan AP-1, mengurangi produksi COX-2, TNF, dan IL-6, dan mengurangi peradangan dalam hewan yang mal karagenan-induced peradangan Model [385].Desoxyrhapontigenin dihasilkan oleh tanaman [386] dan cyototoxic dan efek anti-proliferasi yang tergantung dosis [387-390]. Dengan demikian, desoxyrhapontigenin mungkin berguna sebagai agen terapi untuk cance melalui dampaknya pada ROS dan peradangan.
3.9 PiperinPiperin (1-piperoyl piperidin) adalah alkaloid utama
dalam hitam (Piper nigrum) dan paprika panjang (Piper longum) dan memiliki potensi beberapa kegiatan anti-kanker. Hal ini banyak digunakan baik sebagai bumbu makanan serta dalam pengobatan tradisional [391].
Piperine memiliki toksisitas rendah [392].Selain itu, in vitro penyerapan tingkat tion dari piperin relatif tinggi dibandingkan dengan produk alami lainnya seperti curcumin tanpa metabolik modifikasi dari piperin selama proses penyerapan [393]. Piperin banyak digunakan sebagai penambah bioavailabilitas untuk berbagai kelompok agen terapetik termasuk agen antimikroba rifampisin [394], nevirapine yang merupakan inhibitor poten dari immunode manusia virus defisiensi (HIV) -1 reverse transcriptase [395] dan curcumin yang memiliki sifat antikanker [396]. Piperin memiliki berbagai efek biologis termasuk aktivitas antioksidan [397] serta anti-inflamasi dengan menghambat PMA-diinduksi COX-2 [398].
Piperin memiliki efek signifikan sebagai anti-kanker [399]. Mekanisme yang berbeda telah diusulkan, termasuk apoptosis dan pression dukungan dari metastasis [400], penghambatan angiogenesis [401],dan menghambat invasi oleh downregulation dari MMPs [402.403]. Piperin mengeluarkan aktivitas kemopreventif terhadap beberapa karsinogen termasuk benzo (a) pyrene-diinduksi paru karsinogenesis [404] dan DMBA-diinduksi karsinogenesis kulit [405].Piperin, dengan toksisitas rendah dan aktivitas anti-angiogenik kuat, dapat dianggap sebagai agen terapi mungkin dalam pencegahan dan pengobatan kanker.
3.10 ZerumbonZerumbon, sesquiterpene, diberikannya efek
antikanker melalui modulasi jalur dari JAK / STAT. Di dalam sel karsinoma sel ginjal dan model xenograft mouse, ketergantungan zerumbon dosis dan waktu, menghambat aktivitas STAT-3 melalui dukungan menekan kinase c-SRC, JAK-1 dan JAK-2. Pervanadate, adalah protein tirosin fosfatase (PTP) sebagai pengobatan inhibitor terbalik yang diinduksi downregulation zerumbon STAT3, menunjukkan keterlibatan PTP a. SHP-1 tirosin fosfatase berinteraksi

dengan STAT3 dan diinduksi oleh zerumbon. Setelah knockdown dari SHP-1 oleh siRNA, kemampuan zerumbon untuk menghambat aktivasi STAT3 dimediasi apoptosis ditekan, menunjukkan keterlibatan SHP-1 dalam aksinya. Zerumbon tidak hanya menekan STAT3 tetapi juga mengurangi ekspresi regulasi STAT3-diatur produk gen yang terlibat dalam perkembangan, kelangsungan hidup, dan angiogenesis [406]. Oleh karena itu, zerumbon menghambat aktivasi STAT3, menyebabkan penekanan pertumbuhan dan sensitisasi sel kanker.
Zerumbon adalah inhibitor konstitutif JAK / STAT serta IL-6 dengan merangsang jalur, sehingga menghalangi aktivitas IL-6 pada karsinoma pankreas [407]. Sebuah efek sinergis dari zerumbon dengan paclitaxel dalam sel kanker prostat dimediasi melalui aktifasi jalur JAK-2 / STAT-3 [407.408]. Zerumbon dengan cisplatin menunjukkan efek antikanker sinergis pada neoplasia intraepithelial serviks pada perempuan BALB / c tikus melalui serum IL-6 [409]. Dalam beberapa kasus, penghambatan JAK-2 / STAT-3-dimediasi jalur sinyal diinduksi sitotoksisitas melalui PARP di garis sel kanker prostat manusia (DU 145) [407]. Namun, mekanisme apoptosis lainnya juga telah dilaporkan melalui induksi G2 / M penangkapan dan penurunan cyclin B1 / tingkat protein Cdk1 di HL-60 sel [410]. G2 / M menahan dan Fas- dan mitokondria dimediasi-apoptosis telah diamati pada sel T-akut lymphoblastic leukemia [411] dan sel-sel leukemia [412],dan Bax- dan Bak-dimediasi apoptosis telah diamati pada sel kanker payudara manusia dan othotopic xenograft [413]. Ini mungkin memodulasi Bax rasio / Bcl-2 pada sel kanker hati independen dari p53 fungsional [414],TRAIL-menginduksi kematian reseptor pada kanker usus besar manusia [415], dan Gli-1 / Bcl-2 jalur dimediasi apoptosis pada karsinoma ginjal manusia [416].
Zerumbon dihambat CXC kemokin reseptor-4 ekspresi dengan penghambatan berikutnya CXCL-12 menginduksi invasi dari payudara dan sel-sel pankreas tumor [417] dan sel skuamosa karsinoma lidah manusia [417]. Zerumbon menghambat IL-6 dan menginduksi apoptosis di sel-sel kanker ovarium dan rahim [418]. Hal ini juga menurunkan kadar nitrit dan prostaglandin (E2) dengan tidak berubah COX-1 di LPA dan gamma radiasi meningkatkan sintesa NO dan COX-2 serta pelepasan TNF di RAW 264.7 makrofag tikus [419-421]. Zerumbon menekan aktivasi TPA-diinduksi dari ekspresi EBV, LOX-1 mRNA [422], O2 –anion menjeneralkan melalui NADPH oksidase dalam DMSO dibedakan sel leukemia promyelocytic (HL-60) manusia dan melalui xanthine oksidatif Dase di AS52 sel ovarium hamster Cina [423].
Keamanan zerumbon telah dibuktikan dalam sel manusia yang normal [424]. Konsumsi oral dari zerumbon (20/40 mg / kg / hari) selama 8 minggu mengurangi hiperglikemia yang diinduksi p38 dimediasi
respon inflamasi (infiltrasi makrofag dan peningkatan kadar IL-1, -6 dan TNF) dan juga mengurangi ekspresi antar adhesi molekul-1, MCP-1, TGF- 1 dan fibronectin pada ginjal tikus Terbukti secar in vitro dan in vivo efisiensi farmakologis dari zerumbon memberikan dasar untuk menjelaskan bioaktivitas antikanker. Namun untuk meningkatkan bioavailabilitas, struktur nano lipid zerumbon juga telah disiapkan dan ditandai untuk efek mereka antileukemic [410].
4. Diskusi: mengevaluasi target dan pendekatan
Kami telah mengidentifikasi 10 pendekatan potensi untuk menghambat 10 identifikasi target untuk mengobati dan mencegah kanker dengan menargetkan aspek yang spesifik dari lingkungan mikro tumor (Gambar1 dan 2.Tabel 1 dan 2) Daftar kami adalah tidak lengkap tetapi lebih ilustratif. Beberapa tema muncul.
Pertama, pertimbangan lingkungan mikro tumor sebagai target untuk pencegahan dan pengobatan kanker menyediakan perspektif yang unik pada kedua tumorigenesis dan terapi kanker. Mayoritas terapi yang ada telah difokuskan pada efek pada sel kanker baru jadi. Namun, penghambatan program biologis yang berhubungan dengan lingkungan mikro tumor mungkin penting untuk pencegahan dan pengobatan kanker.
Kedua, ada banyak ada produk alami yang telah dilaporkan memiliki sifat anti-neoplastik dan / atau pencegahan tumor potensial. Dalam banyak kasus, agen ini tampaknya memiliki efek diskrit dan mudah terukur pada lingkungan mikro tumor. Namun, itu akan menjadi penting untuk mendefinisikan bioavailabilitas serta kinetika eliminasi senyawa ini bila digunakan sendiri atau dalam kombinasi [425].
Ketiga, pengaruh dari obat yang menargetkan mikro membutuhkan pengembangan tes yang unik. Kebanyakan terapi bertarget dapat dievaluasi melalui pemeriksaan ekspresi dan / atau aktivitas molekul yang menjadi target. Pertimbangan lingkungan mikro tumor membutuhkan pengukuran seluler, humoral, dan program mediasi sitokin dan ini memerlukan analisis in situ. Bagi banyak studi klinis, ini adalah masalah karena tidak mungkin untuk mendapatkan bahan klinis yang sesuai untuk evaluasi ini.
Oleh karena itu, pengembangan terapi yang menargetkan microenvironment membutuhkan pendekatan baru untuk membuat pengukuran ini baik melalui teknik yang lebih sensitif yang tidak memerlukan pemeriksaan langsung bahan tumor atau melalui pengukuran bahan yang lebih mudah didapat seperti serum darah.
Keempat, lingkungan mikro tumor dan sel-sel neoplastik baru jadi coevolve temporal selama tumorigenesis. Oleh karena itu, dalam sasaran dari lingkungan mikro, kita harus mengakui bahwa sangat penting untuk dipertimbangkan ketika memperkenalkan

terapi dan untuk mengevaluasi efisiensinya pada waktu yang tepat. Dengan demikian, agen yang mengubah aktifasi kekebalan mungkin penting dalam mencegah tumorigenesis. Oleh karena itu, maka diperlukan pemeriksaan efek yang sangat dini selama inisiasi tumor sebagai lawan dalam model praklinis atau pasien dengan kanker stadium lanjut.
Kelima, evaluasi agen yang menargetkan tumor microenvironment harus dipertimbangkan dalam konteks pendekatan terapi yang ada untuk pencegahan tumor. Dengan demikian, bagi banyak kanker ada diterima pendekatan untuk mengobati tumor primer atau metastasis, atau untuk mengurangi kemungkinan lesi kanker awal untuk maju ke kanker lebih maju. Pertimbangan dari target atau pendekatan kami sarankan harus mempertimbangkan standar saat perawatan.
Keenam, pengukuran perubahan dalam tumor microenvironment mungkin endpoint penting untuk mengevaluasi praklinis dan efisiensi klinis terapi. Meneliti bagaimana individu atau sendirian spesifik pendekatan dapat memengaruhi spesifik target bisa memberikan pengukuran menengah mensugestif efisiensi terapetik.
Akhirnya, kami menyadari bahwa pendekatan multi-disiplin yang luas itu penting untuk mengidentifikasi kedua pendekatan dan target. Tumor mikro sifatnya terjadi dalam program biologis yang berbeda di beberapa skala (molekul, sel, organ, host) selama periode waktu yang lama. Peneliti dengan keahlian di berbagai disiplin ilmu yang lain diperlukan untuk mempertimbangkan kompleksitas ini.
Conflik kepentinganSemua penulis mengungkapkan tidak ada
kepentingan keuangan konflik.
Ucapan Terima KasihRLW sedang melakukan ujicoba Tahap I untuk EGCG.
Para penulis mengucapkan terima kasih kepada
Kementerian Italia Universitas dan Penelitian
(2009FZZ4XM 002 EN, AA), Program Penelitian
Intramural dari NIH, National Cancer Institute, Center
for Cancer Research (SPK), National Institutes of
Health (R01CA128704 ke DH), National Institutes of
Health F32CA177139, R01CA170378, U54CA149145,
P50CA114747, R01CA184384, dan R21CA169964
(SCC, DWF), sebuah NIH R15 CA137499-01 (FB),
dana startup dari Ohio Uni- hayati, hibah RSAC
(RP1206) dari Heritage College of Osteopathic
Medicine di OU (FB), Prostat dan Kanker ovarium
Research Trust di Surrey, UK (S. Chen), sebuah
Connecticut State, Universitas sity Penelitian hibah (S.
Crawford), sebuah Reintegrasi hibah MC- Uni Eropa
Marie Curie CIG-303514, dana Nasional Yunani
melalui Program nasional Opera- 'Pendidikan dan
Lifelong Learning Kerangka Acuan Strategis Nasional
(NSRF) -Penelitian Program Pendanaan: THALES
(nomor Hibah MIS 379.346) dan BIAYA
ActionCM1201'Biomimetic Radical Chemistry '(untuk
AGG), Universitas Charles di Praha proyek UNCE
204.015 dan PRVOUK P31 / 2012, dengan P301
proyek Republik Science Foundation / 12/1686, dan
oleh Inter nal Hibah Badan Departemen Kesehatan
proyek Republik Ceko NT13663-3 / 2012 (untuk PH),
Departemen Pendidikan, kultur, Olahraga, Sains dan
Teknologi, Jepang (No. 24590493) (KH), dan swasta
sumbangan (RLW). WNK dan AEB acknowl- tepi
Universitas Glasgow, Beatson Onkologi Pusat Fund,
CRU (www.cancerresearchuk.org/) hibah C301 /
A14762. Sponsor studi tidak terlibat dalam penelitian
ini. Para penulis juga mengakui upaya Leroy Lowe dan
Kanker tim Mengenal.