bab vi produksi lakase dari marasmius sp. dalam bioreaktor ... · pdf filekompresor sistem...
TRANSCRIPT

50
Bab VI Produksi Lakase dari Marasmius sp.
dalam Bioreaktor Imersi Berkala Termodifikasi
Abstrak
Lakase merupakan salah satu enzim yang dihasilkan oleh jamur pelapuk putih yang dapat digunakan pada degradasi lignin. Kemampuan ini dapat dimanfaatkan untuk proses pemutihan pulp. Penelitian produksi lakase telah banyak dilakukan pada berbagai spesies jamur, jenis reaktor, jenis kultivasi dan inducer yang digunakan. Salah satu faktor yang berpengaruh pada produksi lakase adalah tegangan geser (shear stress). Tegangan geser yang kuat pada kultur akan menghambat dan membuat lakase tidak aktif. Temporary immersion bioreactor merupakan bioreaktor yang dapat mengurangi tegangan geser terhadap kultur jamur. Pada penelitian ini dilakukan produksi lakase pada bioreaktor imersi berkala termodifikasi. Penelitian dilakukan untuk mengetahui pengaruh lamanya waktu imersi pada produksi lakase. Variasi waktu imersi yang dilakukan adalah waktu imersi 15 menit, 12 jam dan 24 jam. Hasil penelitian menunjukkan bahwa aktivitas maksimum lakase diperoleh pada kultur Marasmius sp dengan waktu imersi 12 jam, yaitu sebesar 457,6 U/l dan diikuti berturut-turut oleh kultur Marasmius sp. dengan waktu imersi 15 menit (348,4 U/l) dan 24 jam (281,9 U/l). Aktivitas lakase pada siklus kedua lebih rendah dibandingkan siklus pertama. Persentase penurunan tertinggi aktivitas lakase pada siklus kedua terjadi pada kultur dengan waktu imersi 24 jam (80,66%) kemudian diikuti oleh kultur dengan waktu imersi 12 jam (64,30%) dan 15 menit (3,83%). Kultur dengan waktu imersi 12 jam menghasilkan lakase lebih tinggi dibandingkan kultur dengan waktu imersi 15 menit dan 24 jam, namun produktivitas kultur tersebut tidak menunjukkan nilai yang paling tinggi. Hasil penelitian menunjukkan produktivitas kultur Marasmius sp. dengan waktu imersi 15 menit menghasilkan produktivitas yang paling tinggi yaitu 348,4 U/l/hari diikuti oleh kultur dengan waktu imersi 24 jam (281,9 U/l/hari) dan 12 jam (152,5 U/l/hari). Aktivitas lakase dapat juga dinyatakan sebagai Unit per mg total protein. Aktivitas lakase siklus pertama dan siklus kedua memiliki pola kecenderungan yang sama yaitu mula-mula aktivitasnya naik sampai aktivitas tertinggi kemudian turun aktivitasnya. Aktivitas lakase yang didapat pada siklus pertama untuk waktu imersi 12 jam sebesar 7,46 U/mg protein, berturut-turut kemudian adalah waktu imersi 15 menit (3,49 U/mg protein) dan waktu imersi 24 jam (2,57 U/mg protein). Aktivitas lakase pada siklus kedua menunjukkan pola yang sama dengan siklus pertama namun nilainya lebih rendah. Hal ini menunjukkan adanya protein (enzim ekstraseluler) lain selain lakase.
Kata kunci : lakase, Marasmius sp., bioreaktor imersi berkala termodifikasi, tegangan geser, waktu imersi, aktivitas lakase, produktivitas kultur.

51
VI.1 Pendahuluan
Lakase dikelompokkan ke dalam enzim oksidatif yang berperan sebagai biokatalis
proses degradasi lignin. Dengan kemampuannya ini, industri pulp, kertas dan
tekstil mulai menggunakan enzim ini pada proses produksinya untuk
mendapatkan proses produksi yang efisien dan ramah lingkungan karena dalam
kerjanya enzim ini hanya memerlukan oksigen dan menghasilkan air sebagai satu-
satunya produk samping (Riva, 2005). Namun demikian penggunaan enzim ini
pada industri tersebut di atas masih sangat rendah atau masih dalam tahap
pengembangan. Salah satu penyebabnya adalah harga lakase yang masih sangat
mahal. Jika industri tersebut dapat memproduksi enzim ini secara mandiri dan
langsung digunakan tanpa harus melalui proses pemurnian yang panjang, maka
diperkirakan biaya pengadaan enzim ini akan turun dan penggunaannya akan
meningkat.
Di alam terdapat tiga jenis jamur pengurai kayu yaitu pelapuk putih (white rot),
pelapuk coklat (brown rot), dan pelapuk lunak (soft rot, yang dapat mendegradasi
lignin. Jamur pelapuk putih merupakan kelompok jamur pengurai kayu yang
memiliki kemampuan mendegradasi lignin paling tinggi. Jamur ini melakukan
dekomposisi lignin sehingga dapat mencapai selulosa dan hemiselulosa (Pérez
dkk., 2005). Degradasi lignin dalam kayu menyebabkan terbentuknya kantung-
kantung yang berwarna putih, sehingga kelompok jamur ini disebut sebagai
jamur pelapuk putih (gambar VI.1). Beberapa contoh jamur pelapuk putih antara
lain Trametes versicolor, Irpex lacteus, P. chrysosporium, Heterobasidium
annosum, Ganoderma australe, Phlebia tremellosa, Pleurotus spp. dan Phellinus
pini yang merupakan kelompok Basidiomycetes, serta Xylaria hypoxylan yang
termasuk dalam kelompok Ascomycetes (Martinez dkk., 2005).

52
Gambar VI.1. Degradasi kayu pinus oleh jamur pelapuk putih oleh Phellinus
pini. Bagian berwarna putih merupakan daerah delignifikasi atau penyisihan lignin dan bukan merupakan degradasi selulosa (www.forestpathology.coafes.umn.edu).
Jamur pelapuk coklat mempunyai kemampuan degradasi lignin lebih rendah
daripada jamur pelapuk putih, yang lebih suka menguraikan selulosa dan tidak
mendegradasi lignin secara luas. Pertumbuhan jamur pelapuk coklat pada kayu
hanya akan mengakibatkan lignin termodifikasi secara terbatas agar dapat
mengakses karbohidrat dinding sel (Pérez dkk., 2005). Proses pelapukan oleh
jamur pelapuk coklat akan mengakibatkan kayu menjadi berwarna coklat, kering,
mudah patah dengan patahan seperti kubus. Seluruh jamur pelapuk coklat masuk
ke dalam kelompok Basidiomycetes seperti C. Puteana, Gleophyllum trabeum,
Laetiporus sulphureus, Piptoporus betulinus, Postia placenta dan Serpula
lacrimans (Martinez dkk., 2005).

53
Gambar VI.2.
Degradasi kayu oleh jamur pelapuk coklat. Perhatikan bentuk kotak-kotak pada kayu yang merupakan ciri khas pelapukan kayu oleh jamur pelapuk coklat. (www.forestpathology.coafes.umn.edu)
Jamur pelapuk lunak merupakan pendegradasi yang lambat dan kurang agresif
dibandingkan dengan pelapuk putih dan pelapuk coklat. Jamur ini memiliki
kemampuan yang paling rendah untuk mendegradasi lignin jika dibandingkan
dengan jamur pengurai kayu lainnya. Secara umum kelompok jamur ini
mendegradasi selulosa dan hemiselulosa dengan hanya sedikit memodifikasi
lignin. Hasil pelapukan kayu oleh jamur pelapuk lunak mirip dengan pelapukan
oleh jamur pelapuk coklat seperti disajikan pada gambar VI.3. Pada kelembaban
yang tinggi kayu yang diuraikan oleh jamur ini akan menjadi lunak sedangkan
pada kondisi yang kering kayu akan berwarna coklat dan mudah menjadi bubuk.
Beberapa contoh spesies jamur pelapuk lunak antara lain Chaetomium globusum,
Ustulina deusta yang termasuk dalam kelompok Ascomycetes serta Alternaria
alternata, Thielavia terrestris, Paecilomyces spp. yang termasuk dalam kelompok
Deuteromycetes.

54
Gambar VI.3. Degradasi kayu oleh jamur pelapuk lunak
(www.forestpathology.coafes.umn.edu)
Ketiga jamur tersebut mensekresikan enzim-enzim ekstraseluler untuk
menguraikan komponen kayu. Hal tersebut yang mendasari penelitian ini yaitu
memproduksi enzim ekstraseluler untuk mendegradasi lignin pada proses
biobleaching pulp. Lakase (EC 1.10.3.2, p-difenol oksidase) merupakan salah satu
enzim ekstraseluler yang berfungsi mengkatalisis proses pemecahan lignin,
sehingga dapat digunakan sebagai bahan aktif pada proses biobleaching pulp.
Enzim ini dihasilkan oleh spesies jamur yang masuk ke dalam kelompok
Ascomycetes dan Basidiomycetes.
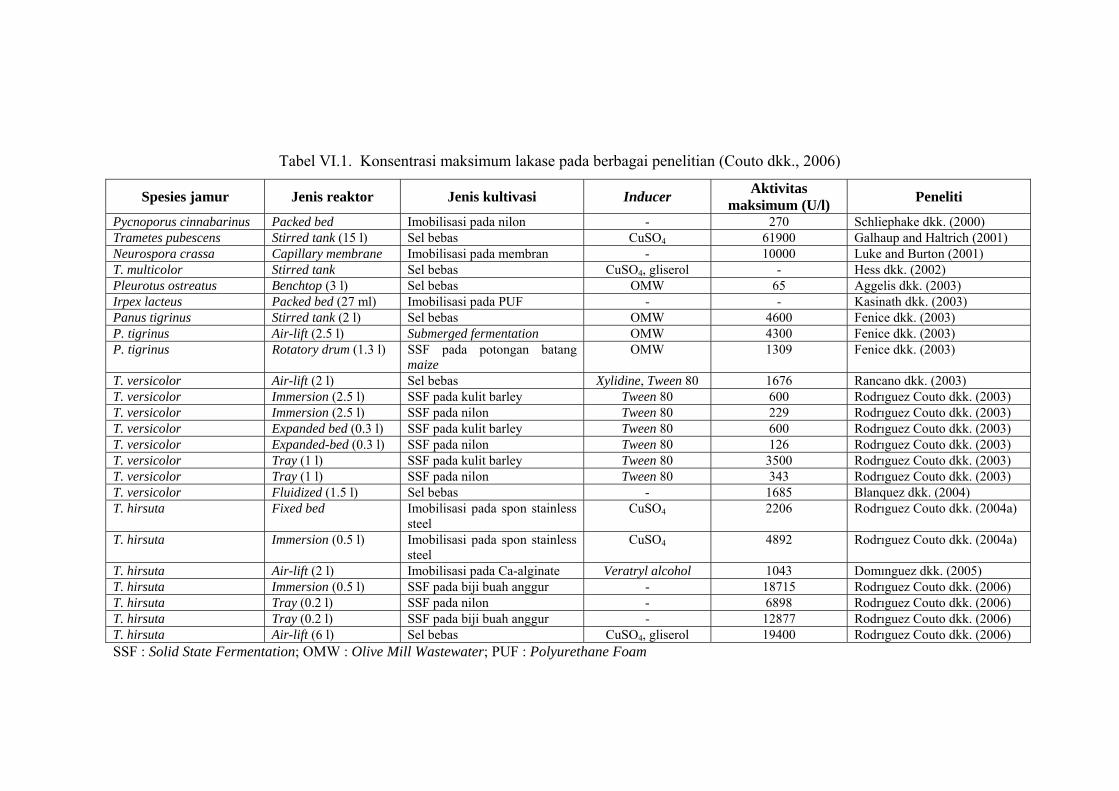
Penelitian produksi enzim lakase telah banyak dilakukan seperti disajikan pada
tabel VI.1. Penelitian tersebut dilakukan pada berbagai spesies jamur, jenis
reaktor, jenis kultivasi dan inducer yang digunakan (Couto dkk., 2006). Menurut
Van der Merwe (2002) produksi enzim ligninolitik jamur pelapuk putih
dipengaruhi oleh beberapa faktor antara lain spesies jamur, kandungan nitrogen,
temperatur, pH, inducer dan inhibitor. Sedangkan Prasad dkk. (2006)
mengungkapkan bahwa selain faktor tersebut transfer oksigen juga sangat penting.
Kultur yang teragitasi secara mekanis merupakan penghambat produksi lakase
karena adanya tegangan geser (shear stress) yang diterima oleh miselia jamur

55
dalam kultur rendam (Prasad dkk., 2005). Oleh karena itu pemilihan bioreaktor
sangat berpengaruh pada produksi enzim ini. Couto dkk. (2004) menggunakan
bioreaktor imersi dan peneliti yang sama pada tahun 2006 menggunakan
bioreaktor air-lift untuk memproduksi lakase. Pada kedua bioreaktor tersebut
tidak terdapat tegangan geser akibat agitasi mekanis, namun kebutuhan oksigen
tetap terpenuhi dengan baik.
Temporary immersion bioreactor telah digunakan dengan sukses untuk
menumbuhkan kultur Trametes versicolor dan Phanerochaete chrysosporium
yang digunakan pada proses penghilangan warna sintetis (Böhmer dkk., 2006).
Sistem kultivasi ini berhasil karena dapat mengurangi shear stress yang
menghambat produksi lakase.
Bioreaktor sistem RITA (Récipient à Immersion Temporaire Automatique)
(Artiles, 2003) terdiri dari dua kompartemen yaitu kompartemen untuk
pertumbuhan mikroorganisme dan kompartemen untuk medium cair. Kedua
kompartemen tersebut dihubungkan dengan selang silikon atau selang kaca. Udara
tekan steril dari kompresor dialirkan ke dalam kompartemen medium untuk
mendorong medium tersebut masuk ke dalam kompartemen tempat tumbuh
mikroorganisme. Keadaan ini akan menyebabkan mikroorganisme dapat
menggunakan nutrisi yang terdapat di dalam medium. Kondisi mikroorganisme
yang terendam dijaga beberapa saat kemudian udara tekan steril dialirkan ke
dalam kompartemen mikroorganisme untuk mendorong medium kembali ke
kompartemen medium. Keadaan ini akan memberikan kesempatan
mikroorganisme mendapatkan pasokan oksigen dari udara. Proses ini dilakukan
berulang-ulang (Artiles, 2003). Skema temporary immersion bioreactor disajikan
pada gambar VI.4.

56
Gambar VI.4. Skema temporary immersion bioreactor (www.bioplantas.cu)
Prinsip temporary immersion bioreactor tersebut telah digunakan oleh Couto dkk.
(2004) untuk memproduksi lakase menggunakan Trametes hirsuta yang
diimobilisasi dalam spon stainless steel. Perendaman kultur jamur terimobilisasi
dengan mencelupkannya ke dalam medium cair. Pada periode waktu tertentu
kultur diangkat dari medium cair (Couto dkk., 2004). Skema bioreaktor imersi
disajikan dalam gambar VI.5.
Gambar VI.5. Skema bioreaktor imersi produksi lakase
(Couto dkk., 2004)
kompresor sistem pneumatik pengambilan conto
medium cair
tempat kultur jamur
saringan steril
keluaran gas

57
Berdasarkan hal di atas maka pada penelitian ini digunakan prinsip dari temporary
immersion bioreactor yang dimodifikasi untuk memproduksi lakase yang
digunakan pada proses biobleaching pulp. Pemindahan medium cair dari satu
kompartemen ke kompartemen lainnya tidak menggunakan udara tekan melainkan
dengan memanfaatkan beda ketinggian kedua kompartemen. Pada penelitian ini
dilakukan kajian tentang pengaruh waktu imersi terhadap produksi lakase oleh
Marasmius sp.
58
Tabel VI.1. Konsentrasi maksimum lakase pada berbagai penelitian (Couto dkk., 2006)
Spesies jamur Jenis reaktor Jenis kultivasi Inducer Aktivitas maksimum (U/l) Peneliti
Pycnoporus cinnabarinus Packed bed Imobilisasi pada nilon - 270 Schliephake dkk. (2000) Trametes pubescens Stirred tank (15 l) Sel bebas CuSO4 61900 Galhaup and Haltrich (2001) Neurospora crassa Capillary membrane Imobilisasi pada membran - 10000 Luke and Burton (2001) T. multicolor Stirred tank Sel bebas CuSO4, gliserol - Hess dkk. (2002) Pleurotus ostreatus Benchtop (3 l) Sel bebas OMW 65 Aggelis dkk. (2003) Irpex lacteus Packed bed (27 ml) Imobilisasi pada PUF - - Kasinath dkk. (2003) Panus tigrinus Stirred tank (2 l) Sel bebas OMW 4600 Fenice dkk. (2003) P. tigrinus Air-lift (2.5 l) Submerged fermentation OMW 4300 Fenice dkk. (2003) P. tigrinus Rotatory drum (1.3 l) SSF pada potongan batang
maize OMW 1309 Fenice dkk. (2003)
T. versicolor Air-lift (2 l) Sel bebas Xylidine, Tween 80 1676 Rancano dkk. (2003) T. versicolor Immersion (2.5 l) SSF pada kulit barley Tween 80 600 Rodrıguez Couto dkk. (2003) T. versicolor Immersion (2.5 l) SSF pada nilon Tween 80 229 Rodrıguez Couto dkk. (2003) T. versicolor Expanded bed (0.3 l) SSF pada kulit barley Tween 80 600 Rodrıguez Couto dkk. (2003) T. versicolor Expanded-bed (0.3 l) SSF pada nilon Tween 80 126 Rodrıguez Couto dkk. (2003) T. versicolor Tray (1 l) SSF pada kulit barley Tween 80 3500 Rodrıguez Couto dkk. (2003) T. versicolor Tray (1 l) SSF pada nilon Tween 80 343 Rodrıguez Couto dkk. (2003) T. versicolor Fluidized (1.5 l) Sel bebas - 1685 Blanquez dkk. (2004) T. hirsuta Fixed bed Imobilisasi pada spon stainless
steel CuSO4 2206 Rodrıguez Couto dkk. (2004a)
T. hirsuta Immersion (0.5 l) Imobilisasi pada spon stainless steel
CuSO4 4892 Rodrıguez Couto dkk. (2004a)
T. hirsuta Air-lift (2 l) Imobilisasi pada Ca-alginate Veratryl alcohol 1043 Domınguez dkk. (2005) T. hirsuta Immersion (0.5 l) SSF pada biji buah anggur - 18715 Rodrıguez Couto dkk. (2006) T. hirsuta Tray (0.2 l) SSF pada nilon - 6898 Rodrıguez Couto dkk. (2006) T. hirsuta Tray (0.2 l) SSF pada biji buah anggur - 12877 Rodrıguez Couto dkk. (2006) T. hirsuta Air-lift (6 l) Sel bebas CuSO4, gliserol 19400 Rodrıguez Couto dkk. (2006) SSF : Solid State Fermentation; OMW : Olive Mill Wastewater; PUF : Polyurethane Foam

59
VI.2 Bahan dan Metode
VI.2.1 Mikroorganisme
Jamur yang digunakan adalah Marasmius sp. yang terpilih pada percobaan
sebelumnya. Sediaan Marasmius sp. yang digunakan sebagai inokulum,
ditumbuhkan pada medium Potato Dextrose Agar (PDA) dalam cawan petri
berumur 5 hari. Cara menumbuhkan jamur tersebut dapat dilihat pada subbab
III.2.1.a.
VI.2.2 Konfigurasi bioreaktor
Pada percobaan ini Marasmius sp. ditumbuhkan dengan metode temporary
immersion culture menggunakan bioreaktor sistem RITA (Récipient à Immersion
Temporaire Automatique) yang telah dimodifikasi. Satu set bioreaktor terdiri atas
dua kompartemen yang terbuat dari kaca dan terhubung dengan selang silikon.
Ukuran bioreaktor adalah tinggi 15 cm, panjang 10 cm dan lebar 10 cm. Tiap
bioreaktor diisi Marasmius sp. yang terimobilisasi sedemikian rupa sehingga
ketinggian kultur dalam bioreaktor adalah 8 cm. Salah satu kompartemen dapat
digerakkan naik dan turun dengan menggunakan mesin penggerak yang diatur
waktunya sesuai variasi waktu imersi yang digunakan. Variasi waktu imersi yang
digunakan pada penelitian ini adalah 15 menit, 12 jam dan 24 jam. Tiap variasi
waktu imersi dilakukan untuk dua siklus kultivasi dengan tiap siklus kultivasi
berjalan selama tiga hari. Satu siklus berarti kultur jamur dalam bioreaktor
menggunakan medium Kirk yang sama selama tiga hari sedangkan pada siklus
berikutnya medium Kirk seluruhnya diganti dengan medium Kirk yang baru.
Skema bioreaktor disajikan dalam gambar III.3 dan konfigurasi bioreaktor imersi
berkala termodifikasi disajikan dalam gambar VI.6

60
Gambar VI.6. Bioreaktor imersi berkala termodifikasi
Udara disuplai menggunakan aerator dan medium yang digunakan untuk produksi
lakase adalah sama dengan medium untuk merendam bulustru yaitu medium Kirk
(dengan kandungan lindi hitam 0,4%) sebanyak 700 ml tiap satu set bioreaktor.
Pengambilan conto dilakukan setiap 24 jam.
VI.2.3 Media imobilisasi
Media imobilisasi Marasmius sp. yang digunakan pada penelitian ini adalah
bulustru (Sunda)/gambas (Jawa). Bulustru merupakan serat buah oyong yang telah
dikeringkan. Sebelum digunakan, bulustru dengan luas kurang lebih 900 cm2
dipotong dengan ukuran sekitar 5 cm x 5 cm dan direndam dalam medium Kirk
selam 15 menit. Bulustru yang telah mengandung medium Kirk kemudian
ditempatkan dalam plastik tahan panas dan disterilisasi. Setelah dingin bulustru
tersebut selanjutnya diinokulasi dengan kultur Marasmius sp. yang telah tumbuh
dengan baik pada medium agar. Satu kantung plastik bulustru diinokulasi dengan
satu cawan petri kultur Marasmius sp. secara aseptis. Bulustru ini selanjutnya
diinkubasi pada temperatur ruang (± 28°C) selama kurang lebih 10 hari untuk
mendapatkan pertumbuhan Marasmius sp. yang baik.

61
Penumbuhan Marasmius sp. pada bulustru dimaksudkan agar kultur jamur tidak
terbawa aliran medium pada saat pengosongan bioreaktor, yang dikhawatirkan
akan menyumbat saluran. Imobilisasi Marasmius sp. ini memungkinkan luas
kontak kultur dengan oksigen akan meningkat dan menurunkan hambatan transfer
massanya. Pemindahan medium cair dengan memanfaatkan beda ketinggian
bioreaktor akan mengurangi tegangan geser yang dialami oleh kultur jamur. Hal
ini sesuai dengan yang dikemukakan oleh Prasad dkk. (2005) bahwa oksigen
merupakan salah satu faktor penting dalam produksi enzim ligninolitik oleh jamur
pelapuk putih dan agitasi mekanis yang menyebabkan tegangan geser pada kultur
merupakan penghambat produksi enzim tersebut.
VI.2.4 Medium pertumbuhan Marasmius sp.
Medium yang digunakan untuk memproduksi lakase pada percobaan ini adalah
medium Kirk. Komposisi dan cara pembuataan medium ini dapat dilihat pada
subbab III.2.1.c.
VI.2.5 Kultivasi Marasmius sp.
Kultivasi Marasmius sp. untuk memproduksi lakase dilakukan dengan
memasukkan satu kantung kultur Marasmius sp. terimobilisasi pada bulustru pada
masing-masing kompartemen bioreaktor. Kemudian ke dalam satu kompartemen
dimasukkan 700 ml medium Kirk steril yang mengandung lindi hitam dengan
persentase 0,4% (v/v). Kultivasi dilakukan pada suhu ruang (± 28°C) dengan
aerasi pada head space bioreaktor. Conto medium kultur diambil setiap 24 jam
untuk selanjutnya dianalisis aktivitas lakase dan konsentrasi proteinnya. Setelah
tiga hari kultivasi medium kultivasi dihentikan dengan cara mengeluarkan seluruh
medium kultur. Kemudian siklus kultivasi kedua langsung dilakukan dengan cara
menambahkan 700 ml medium baru.
VI.2.6 Uji aktivitas lakase
Uji aktivitas lakase dilakukan berdasarkan metode Bourbonnais dan Paice (1990).
Prinsip uji ini adalah sebagai berikut : pewarna non-phenol 2,2’-azinobis-di-(3-
ethylbenzthiazolinesulphonate) (ABTS) dioksidasi oleh lakase menjadi radikal

62
kation (ABTS+) yang lebih stabil (gambar VI.7). Konsentrasi radikal kation yang
berwarna biru kehijauan (dibaca pada panjang gelombang 420 nm) berkorelasi
dengan aktivitas lakase (Bar, 2001)
Gambar VI.7. Oksidasi lakase oleh ABTS menjadi radikal kation (ABTS+)
(Bar, 2001) Pengukuran aktivitas lakase pada conto kultur medium dilakukan dengan cara
sebagai berikut. Larutan 0.4 mM ABTS dalam buffer natrium asetat (pH 4.5)
sebanyak 1160 μl dimasukkan ke dalam kuvet 1,5 ml, selanjutnya ke dalam kuvet
dimasukkan 40 μl conto enzim dan dikocok agar tercampur homogen. Kemudian
absorbansi radikal kation diamati pada panjang gelombang 420 nm (εmM = 36 mM-
1cm-1) selama lima menit menggunakan spektrofotometer Genesis 10 UV-Visible.
Perubahan absorbansi radikal kation diamati setiap menit. Aktivitas lakase
dinyatakan sebagai International Unit (IU) per liter, dengan 1 IU didefinisikan
sebagai jumlah enzim yang dapat mengoksidasi 1 µmol ABTS tiap menit.
Aktivitas lakase ditentukan menggunakan persamaan VI.1.
( ) 1000min.2/ 1 xAxdxxv
VlIUaktivitas −Δ⎟⎟⎠
⎞⎜⎜⎝
⎛=
ε ........... (VI.1)

63
dengan :
V = volume total reaksi (ml)
v = volume enzim (ml)
ε = extinction coefficient ABTS pada 420 nm = 36 mM -1cm-1
d = Light path of cuvette (cm)
∆A.min-1 = perubahan absorbansi tiap menit pada 420 nm
VI.2.7 Pengukuran Konsentrasi Protein
Pengukuran konsentrasi protein dilakukan menggunakan metode Bradford
(Walker, 1996). Pengukuran konsentrasi protein metode Bradford didasarkan pada
pembentukan ikatan pewarna Coomasie Brilliant Blue G-250 dengan protein yang
dapat diamati absorbansinya pada panjang gelombang 595 nm. Penentuan
konsentrasi protein dilakukan dengan cara : ke dalam kuvet 1,5 ml dimasukkan
conto protein sebanyak 50 μl, selanjutnya ke dalam kuvet dimasukkan Reagen
Bradford sebanyak 1000 μl (Komposisi dan cara pembuatan reagen Bradford
dapat dilihat pada lampiran B). Campuran ini kemudian diinkubasi pada suhu
ruang ((± 28°C) selama 5 menit. Kemudian campuran ini diukur absorbansinya
pada panjang gelombang 595 nm. Konsentrasi protein ditentukan berdasarkan
kurva standar protein. Kurva standar dibuat dengan cara sebagai berikut :
pertama-tama dibuat larutan induk Buvine Serum Albumin (BSA) dengan
konsentrasi 2 mg/ml, larutan ini kemudian ini kemudian diencerkan agar memiliki
konsentrasi 0 – 1 mg/ml. Sediaan ini kemudian dinalisis dengan cara seperti yang
telah diuraikan di atas. Profil kurva standar larutan BSA ini dapat dilihat pada
lampiran B.
VI.3 Hasil dan Pembahasan
Percobaan produksi lakase dari Marasmius sp. ini selain untuk untuk mengetahui
pengaruh variasi waktu imersi terhadap produksi enzim, juga untuk mengetahui
pengaruh siklus kultivasi terhadap produksi enzim. Variasi waktu yang digunakan
adalah 15 menit, 12 jam dan 24 jam dan untuk setiap variasi waktu, kultivasi
dilakukan dua siklus.
64
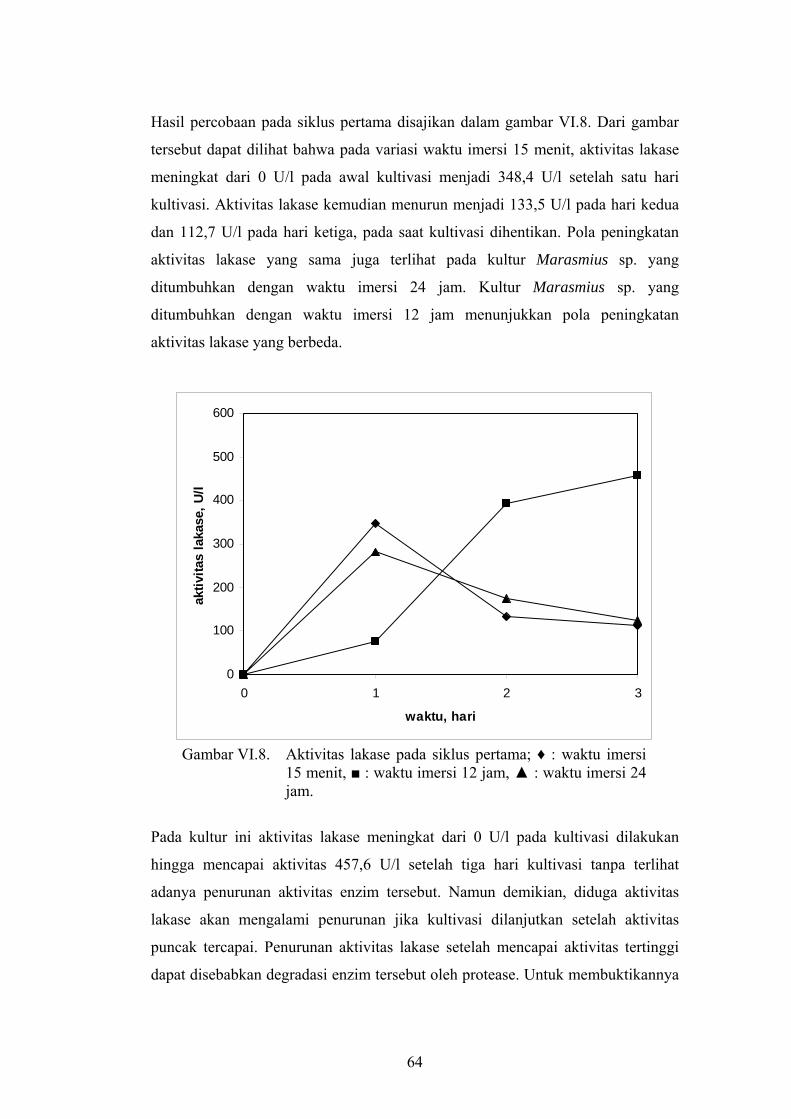
Hasil percobaan pada siklus pertama disajikan dalam gambar VI.8. Dari gambar
tersebut dapat dilihat bahwa pada variasi waktu imersi 15 menit, aktivitas lakase
meningkat dari 0 U/l pada awal kultivasi menjadi 348,4 U/l setelah satu hari
kultivasi. Aktivitas lakase kemudian menurun menjadi 133,5 U/l pada hari kedua
dan 112,7 U/l pada hari ketiga, pada saat kultivasi dihentikan. Pola peningkatan
aktivitas lakase yang sama juga terlihat pada kultur Marasmius sp. yang
ditumbuhkan dengan waktu imersi 24 jam. Kultur Marasmius sp. yang
ditumbuhkan dengan waktu imersi 12 jam menunjukkan pola peningkatan
aktivitas lakase yang berbeda.
0
100
200
300
400
500
600
0 1 2 3
waktu, hari
aktiv
itas
laka
se, U
/l
Gambar VI.8. Aktivitas lakase pada siklus pertama; ♦ : waktu imersi
15 menit, ■ : waktu imersi 12 jam, ▲ : waktu imersi 24 jam.
Pada kultur ini aktivitas lakase meningkat dari 0 U/l pada kultivasi dilakukan
hingga mencapai aktivitas 457,6 U/l setelah tiga hari kultivasi tanpa terlihat
adanya penurunan aktivitas enzim tersebut. Namun demikian, diduga aktivitas
lakase akan mengalami penurunan jika kultivasi dilanjutkan setelah aktivitas
puncak tercapai. Penurunan aktivitas lakase setelah mencapai aktivitas tertinggi
dapat disebabkan degradasi enzim tersebut oleh protease. Untuk membuktikannya
65
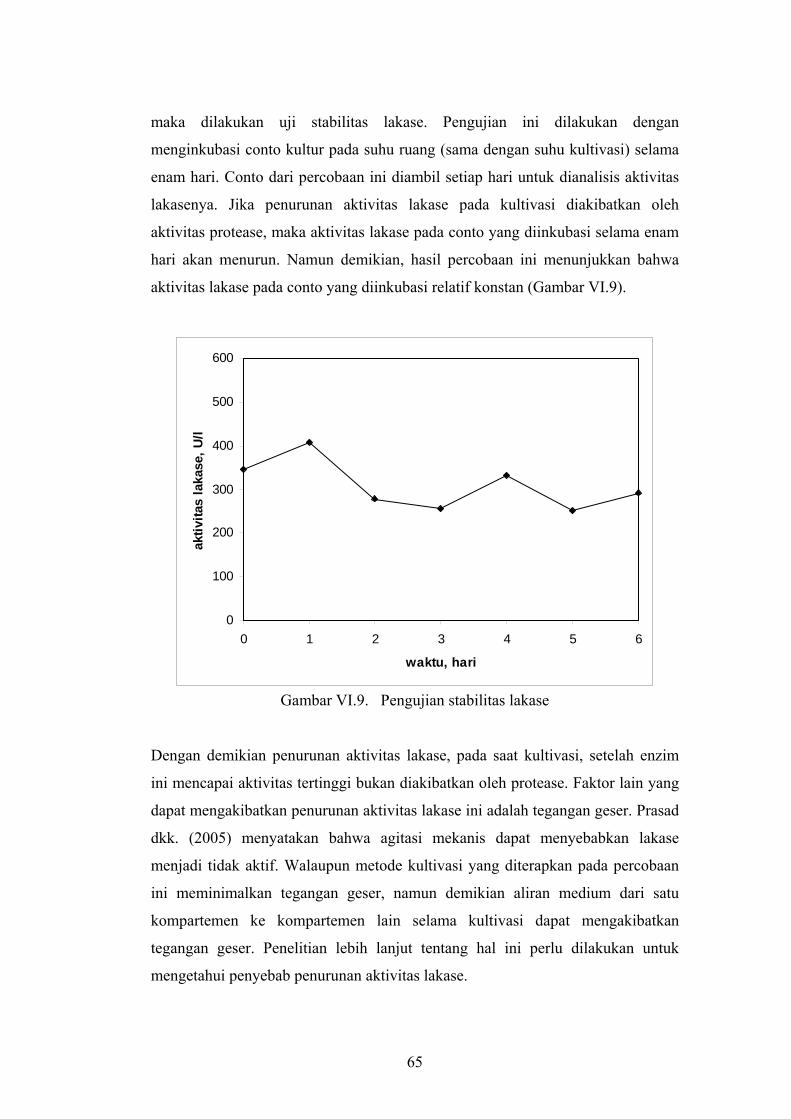
maka dilakukan uji stabilitas lakase. Pengujian ini dilakukan dengan
menginkubasi conto kultur pada suhu ruang (sama dengan suhu kultivasi) selama
enam hari. Conto dari percobaan ini diambil setiap hari untuk dianalisis aktivitas
lakasenya. Jika penurunan aktivitas lakase pada kultivasi diakibatkan oleh
aktivitas protease, maka aktivitas lakase pada conto yang diinkubasi selama enam
hari akan menurun. Namun demikian, hasil percobaan ini menunjukkan bahwa
aktivitas lakase pada conto yang diinkubasi relatif konstan (Gambar VI.9).
0
100
200
300
400
500
600
0 1 2 3 4 5 6
waktu, hari
aktiv
itas
laka
se, U
/l
Gambar VI.9. Pengujian stabilitas lakase
Dengan demikian penurunan aktivitas lakase, pada saat kultivasi, setelah enzim
ini mencapai aktivitas tertinggi bukan diakibatkan oleh protease. Faktor lain yang
dapat mengakibatkan penurunan aktivitas lakase ini adalah tegangan geser. Prasad
dkk. (2005) menyatakan bahwa agitasi mekanis dapat menyebabkan lakase
menjadi tidak aktif. Walaupun metode kultivasi yang diterapkan pada percobaan
ini meminimalkan tegangan geser, namun demikian aliran medium dari satu
kompartemen ke kompartemen lain selama kultivasi dapat mengakibatkan
tegangan geser. Penelitian lebih lanjut tentang hal ini perlu dilakukan untuk
mengetahui penyebab penurunan aktivitas lakase.
66
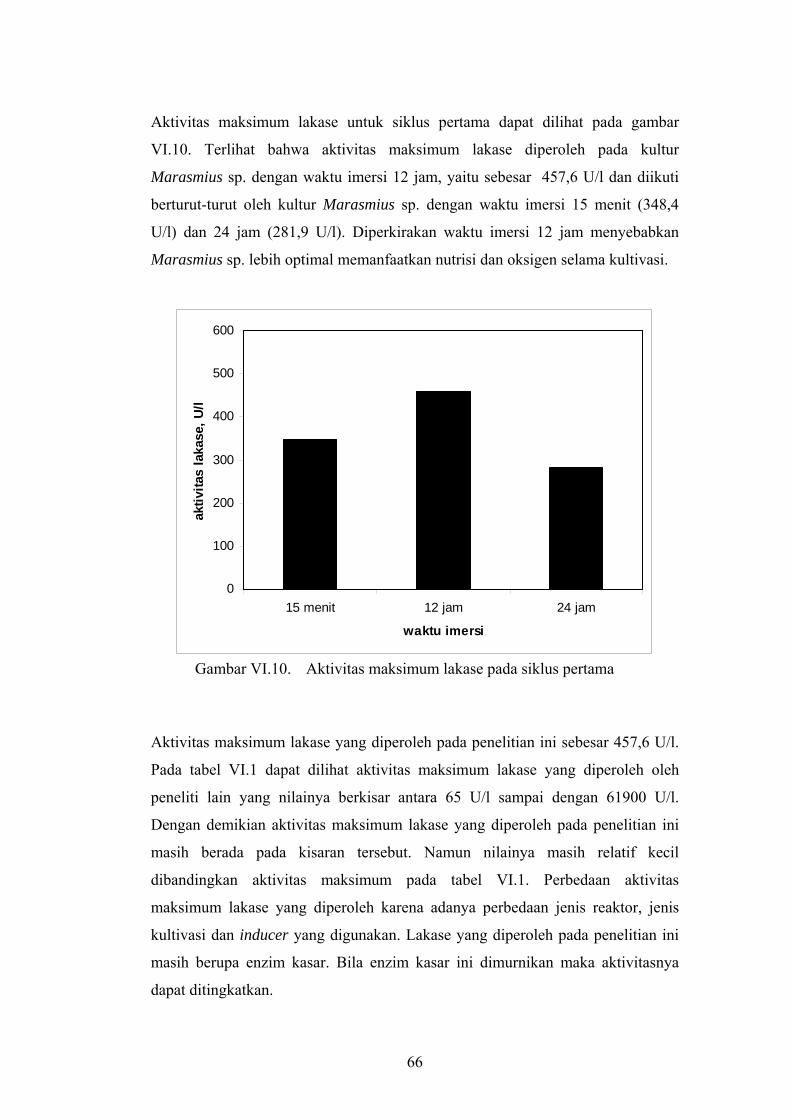
Aktivitas maksimum lakase untuk siklus pertama dapat dilihat pada gambar
VI.10. Terlihat bahwa aktivitas maksimum lakase diperoleh pada kultur
Marasmius sp. dengan waktu imersi 12 jam, yaitu sebesar 457,6 U/l dan diikuti
berturut-turut oleh kultur Marasmius sp. dengan waktu imersi 15 menit (348,4
U/l) dan 24 jam (281,9 U/l). Diperkirakan waktu imersi 12 jam menyebabkan
Marasmius sp. lebih optimal memanfaatkan nutrisi dan oksigen selama kultivasi.
0
100
200
300
400
500
600
15 menit 12 jam 24 jam
waktu imersi
aktiv
itas
laka
se, U
/l
Gambar VI.10. Aktivitas maksimum lakase pada siklus pertama
Aktivitas maksimum lakase yang diperoleh pada penelitian ini sebesar 457,6 U/l.
Pada tabel VI.1 dapat dilihat aktivitas maksimum lakase yang diperoleh oleh
peneliti lain yang nilainya berkisar antara 65 U/l sampai dengan 61900 U/l.
Dengan demikian aktivitas maksimum lakase yang diperoleh pada penelitian ini
masih berada pada kisaran tersebut. Namun nilainya masih relatif kecil
dibandingkan aktivitas maksimum pada tabel VI.1. Perbedaan aktivitas
maksimum lakase yang diperoleh karena adanya perbedaan jenis reaktor, jenis
kultivasi dan inducer yang digunakan. Lakase yang diperoleh pada penelitian ini
masih berupa enzim kasar. Bila enzim kasar ini dimurnikan maka aktivitasnya
dapat ditingkatkan.
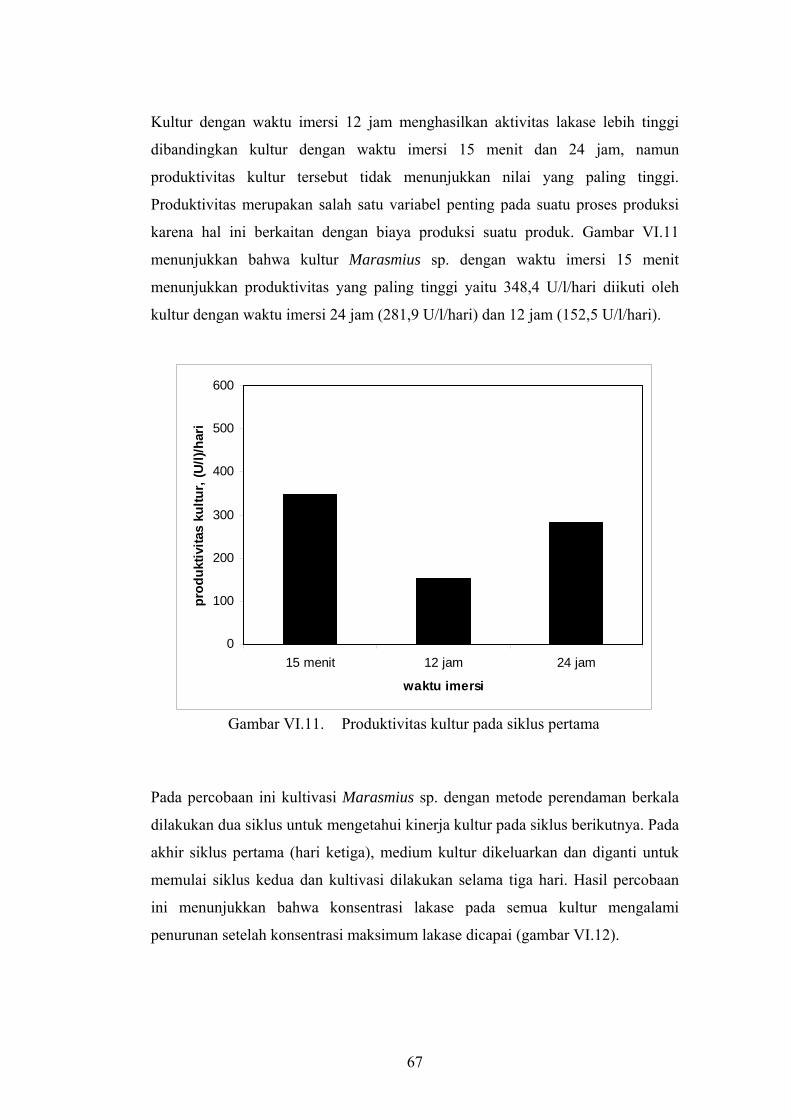
67
Kultur dengan waktu imersi 12 jam menghasilkan aktivitas lakase lebih tinggi
dibandingkan kultur dengan waktu imersi 15 menit dan 24 jam, namun
produktivitas kultur tersebut tidak menunjukkan nilai yang paling tinggi.
Produktivitas merupakan salah satu variabel penting pada suatu proses produksi
karena hal ini berkaitan dengan biaya produksi suatu produk. Gambar VI.11
menunjukkan bahwa kultur Marasmius sp. dengan waktu imersi 15 menit
menunjukkan produktivitas yang paling tinggi yaitu 348,4 U/l/hari diikuti oleh
kultur dengan waktu imersi 24 jam (281,9 U/l/hari) dan 12 jam (152,5 U/l/hari).
0
100
200
300
400
500
600
15 menit 12 jam 24 jam
waktu imersi
prod
uktiv
itas
kultu
r, (U
/l)/h
ari
Gambar VI.11. Produktivitas kultur pada siklus pertama
Pada percobaan ini kultivasi Marasmius sp. dengan metode perendaman berkala
dilakukan dua siklus untuk mengetahui kinerja kultur pada siklus berikutnya. Pada
akhir siklus pertama (hari ketiga), medium kultur dikeluarkan dan diganti untuk
memulai siklus kedua dan kultivasi dilakukan selama tiga hari. Hasil percobaan
ini menunjukkan bahwa konsentrasi lakase pada semua kultur mengalami
penurunan setelah konsentrasi maksimum lakase dicapai (gambar VI.12).
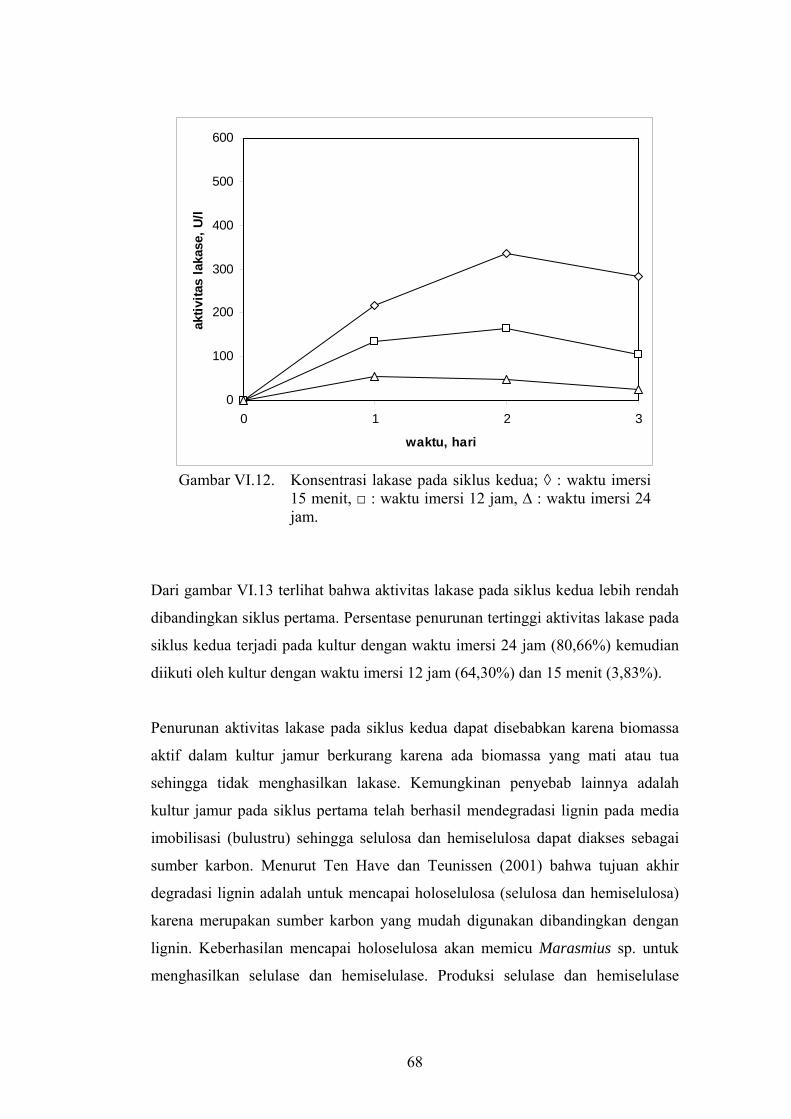
68
0
100
200
300
400
500
600
0 1 2 3
waktu, hari
aktiv
itas
laka
se, U
/l
Gambar VI.12. Konsentrasi lakase pada siklus kedua; ◊ : waktu imersi
15 menit, □ : waktu imersi 12 jam, ∆ : waktu imersi 24 jam.
Dari gambar VI.13 terlihat bahwa aktivitas lakase pada siklus kedua lebih rendah
dibandingkan siklus pertama. Persentase penurunan tertinggi aktivitas lakase pada
siklus kedua terjadi pada kultur dengan waktu imersi 24 jam (80,66%) kemudian
diikuti oleh kultur dengan waktu imersi 12 jam (64,30%) dan 15 menit (3,83%).
Penurunan aktivitas lakase pada siklus kedua dapat disebabkan karena biomassa
aktif dalam kultur jamur berkurang karena ada biomassa yang mati atau tua
sehingga tidak menghasilkan lakase. Kemungkinan penyebab lainnya adalah
kultur jamur pada siklus pertama telah berhasil mendegradasi lignin pada media
imobilisasi (bulustru) sehingga selulosa dan hemiselulosa dapat diakses sebagai
sumber karbon. Menurut Ten Have dan Teunissen (2001) bahwa tujuan akhir
degradasi lignin adalah untuk mencapai holoselulosa (selulosa dan hemiselulosa)
karena merupakan sumber karbon yang mudah digunakan dibandingkan dengan
lignin. Keberhasilan mencapai holoselulosa akan memicu Marasmius sp. untuk
menghasilkan selulase dan hemiselulase. Produksi selulase dan hemiselulase

69
mengakibatkan terjadinya kompetisi sistem produksi enzim di dalam sel
Marasmius sp. sehingga produksi lakase menurun.
0
100
200
300
400
500
600
15 menit 12 jam 24 jam
waktu imersi
aktiv
itas
laka
se, U
/l
Gambar VI.13. Aktivitas maksimum lakase; ■ : siklus pertama dan □ :
siklus kedua
Komponen utama penyusun enzim adalah protein (Shuler dan Kargi, 1992).
Dengan demikian aktivitas enzim juga dapat dinyatakan dalam Unit per mg total
protein (U/mg protein protein). Dari gambar VI.14 dapat diketahui bahwa
aktivitas lakase siklus pertama dan siklus kedua memiliki pola kecenderungan
yang sama yaitu mula-mula aktivitasnya naik sampai aktivitas tertinggi kemudian
turun aktivitasnya. Aktivitas lakase didapat pada siklus pertama untuk waktu
imersi 12 jam sebesar 7,46 U/mg protein, berturut-turut kemudian adalah waktu
imersi 15 menit (3,49 U/mg protein) dan waktu imersi 24 jam (2,57 U/mg
protein).
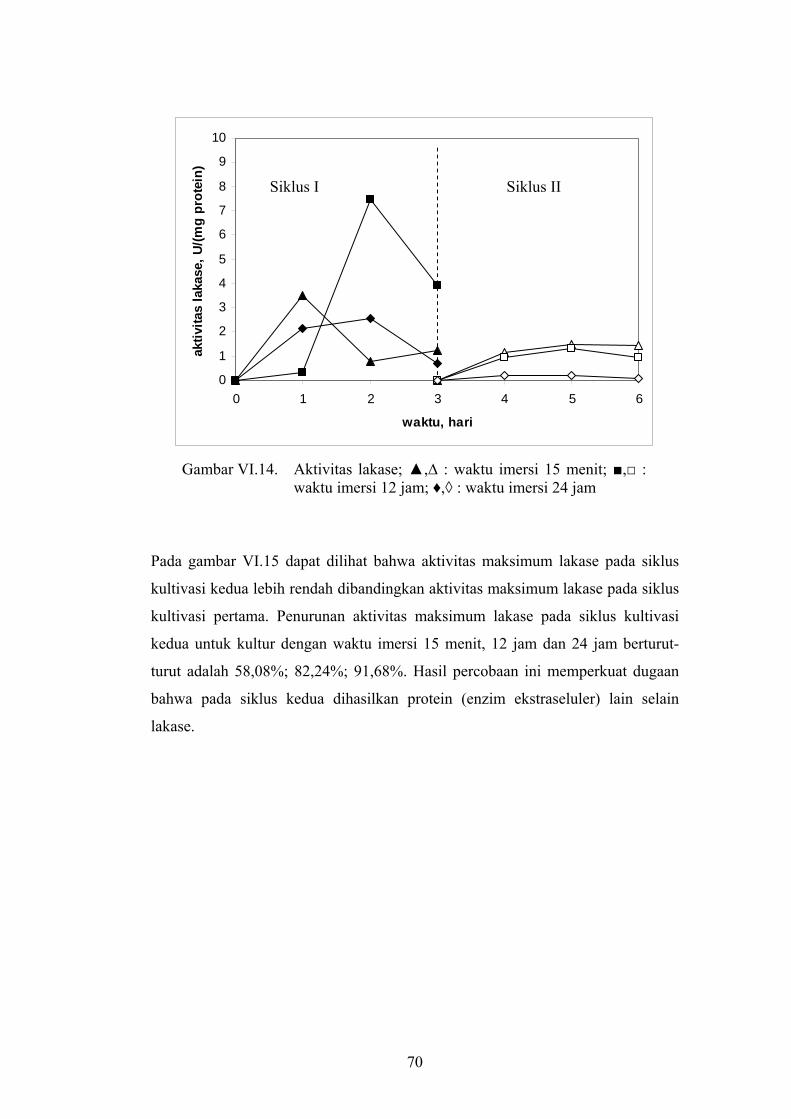
70
0
1
2
3
4
5
6
7
8
9
10
0 1 2 3 4 5 6
waktu, hari
aktiv
itas
laka
se, U
/(mg
prot
ein)
Gambar VI.14. Aktivitas lakase; ▲,∆ : waktu imersi 15 menit; ■,□ :
waktu imersi 12 jam; ♦,◊ : waktu imersi 24 jam
Pada gambar VI.15 dapat dilihat bahwa aktivitas maksimum lakase pada siklus
kultivasi kedua lebih rendah dibandingkan aktivitas maksimum lakase pada siklus
kultivasi pertama. Penurunan aktivitas maksimum lakase pada siklus kultivasi
kedua untuk kultur dengan waktu imersi 15 menit, 12 jam dan 24 jam berturut-
turut adalah 58,08%; 82,24%; 91,68%. Hasil percobaan ini memperkuat dugaan
bahwa pada siklus kedua dihasilkan protein (enzim ekstraseluler) lain selain
lakase.
Siklus I Siklus II
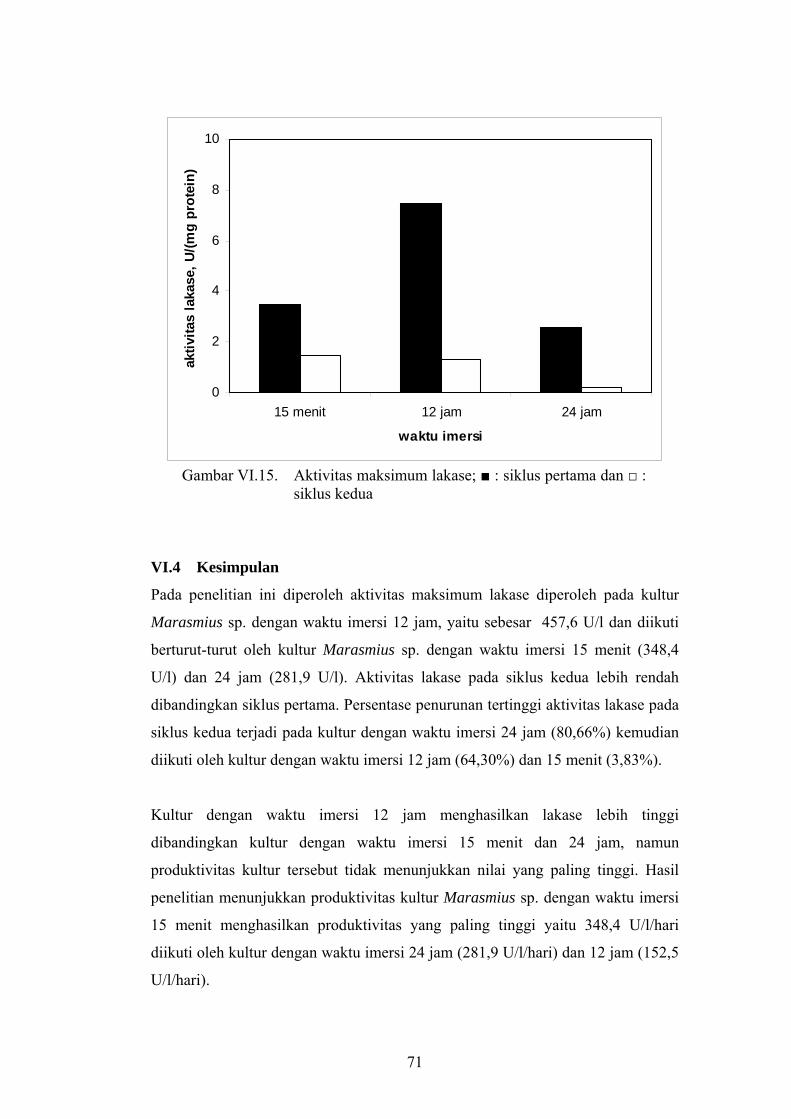
71
0
2
4
6
8
10
15 menit 12 jam 24 jam
waktu imersi
aktiv
itas
laka
se, U
/(mg
prot
ein)
Gambar VI.15. Aktivitas maksimum lakase; ■ : siklus pertama dan □ :
siklus kedua
VI.4 Kesimpulan
Pada penelitian ini diperoleh aktivitas maksimum lakase diperoleh pada kultur
Marasmius sp. dengan waktu imersi 12 jam, yaitu sebesar 457,6 U/l dan diikuti
berturut-turut oleh kultur Marasmius sp. dengan waktu imersi 15 menit (348,4
U/l) dan 24 jam (281,9 U/l). Aktivitas lakase pada siklus kedua lebih rendah
dibandingkan siklus pertama. Persentase penurunan tertinggi aktivitas lakase pada
siklus kedua terjadi pada kultur dengan waktu imersi 24 jam (80,66%) kemudian
diikuti oleh kultur dengan waktu imersi 12 jam (64,30%) dan 15 menit (3,83%).
Kultur dengan waktu imersi 12 jam menghasilkan lakase lebih tinggi
dibandingkan kultur dengan waktu imersi 15 menit dan 24 jam, namun
produktivitas kultur tersebut tidak menunjukkan nilai yang paling tinggi. Hasil
penelitian menunjukkan produktivitas kultur Marasmius sp. dengan waktu imersi
15 menit menghasilkan produktivitas yang paling tinggi yaitu 348,4 U/l/hari
diikuti oleh kultur dengan waktu imersi 24 jam (281,9 U/l/hari) dan 12 jam (152,5
U/l/hari).

72
Aktivitas lakase berdasarkan total protein dalam enzim kasar menunjukkan bahwa
pada siklus pertama dan siklus kedua memiliki pola kecenderungan yang sama
yaitu mula-mula aktivitasnya naik sampai aktivitas tertinggi kemudian turun
aktivitasnya. Aktivitas lakase yang didapat pada siklus pertama untuk waktu
imersi 12 jam sebesar 7,46 U/mg protein, berturut-turut kemudian adalah waktu
imersi 15 menit (3,49 U/mg protein) dan waktu imersi 24 jam (2,57 U/mg
protein). Aktivitas lakase pada siklus kedua menunjukkan pola yang sama dengan
siklus pertama namun nilainya lebih rendah. Hal ini menunjukkan adanya protein
(enzim ekstraseluler) lain selain lakase.