p-issn 2354-8568benih-bogor.litbang.menlhk.go.id/assets/files/jpth... · pola faktorial 2 x 3 pada...
TRANSCRIPT


p-ISSN 2354-8568 e-ISSN 2527-6565
JURNAL PERBENIHAN TANAMAN HUTAN
Vol.7 No.2, Desember 2019
JURNAL PERBENIHAN TANAMAN HUTAN adalah publikasi ilmiah resmi dari Balai Penelitian dan Pengembangan Tanaman Hutan
(BP2TPTH). Jurnal ini mempublikasikan hasil-hasil penelitian dari berbagai aspek perbenihan tanaman hutan, meliputi pembangunan dan
pengelolaan sumber benih, biologi reproduksi, ekologi dan biologi benih, teknologi penanganan benih, teknologi perbanyakan vegetatif,
kesehatan benih, teknik persemaian, pengujian mutu benih dan bibit, sosial, ekonomi dan kebijakan perbenihan. Jurnal Perbenihan Tanaman
Hutan terbit dua kali setahun pada Bulan Agustus dan Desember, dan telah terakreditasi oleh Direktorat Jenderal Penguatan Riset dan
Pengembangan, Kementerian Riset, Teknologi dan Pendidikan Tinggi Republik Indonesia (Nomor: 30/E/KPT/2018) tanggal 24 Oktober
2018. Akreditasi berlaku dari Vol. 5 No. 2 Tahun 2017 sampai Vol. 10 No. 1 Tahun 2022.
Penanggung Jawab
Kepala Badan Penelitian, Pengembangan dan Inovasi
Wakil Penanggung Jawab
Kepala Balai Penelitian dan Pengembangan Teknologi Perbenihan Tanaman Hutan
Dewan Redaksi Ketua Merangkap Anggota
Dr. Ir. Yulianti Bramasto, M.Si (Silvikultur) Balai Penelitian dan Pengembangan Teknologi Perbenihan Tanaman Hutan, Kementerian Lingkungan Hidup dan Kehutanan, Indonesia
Anggota
Prof. Riset Dr. Dra. Dida Syamsuwida, M.Sc (Silvikultur / Produksi Benih) Balai Penelitian dan Pengembangan Teknologi Perbenihan Tanaman Hutan, Kementerian Lingkungan Hidup dan Kehutanan, Indonesia
Dr. Dede Jajat Sudrajat, S.Hut, MT (Silvikultur / Teknologi Benih) Balai Penelitian dan Pengembangan Teknologi Perbenihan Tanaman Hutan, Kementerian Lingkungan Hidup dan Kehutanan, Indonesia
Prof. Riset. Dr. Ir. Nina Mindawati, M.Si (Silvikultur) Pusat Penelitian dan Pengembangan Hutan, Badan Penelitian, Pengembangan dan Inovasi, Kementerian Lingkungan Hidup dan Kehutanan, Indonesia
Prof. Riset. Dr. Ir. Budi Leksono, MP (Pemuliaan) Balai Besar Penelitian dan Pengembangan Bioteknogi dan Pemuliaan Tanaman Hutan, Kementerian Lingkungan Hidup dan Kehutanan, Indonesia
Dr. Sri Utami,SP, M.Si (Hama Hutan, Perlindungan Hutan) Balai Penelitian dan Pengembangan Lingkungan Hidup dan Kehutanan Palembang, Kementerian Lingkungan Hidup dan Kehutanan, Indonesia
Mitra Bestari
Dato’ Dr. Marzalina Hj.Mansor (Genetik) Forest Research Institute Malaysia, Kepong, Malaysia Dr. Ir. Supriyanto (Fisiologi Pohon) Institut Pertanian Bogor, Indonesia
Prof. Dr. Ir. Iskandar Zulkarnaen Siregar, M.Sc.F.Trop (Genetik) Institut Pertanian Bogor, Indonesia Dr. Ir. Muhdin, M.Sc (Statistika) Institut Pertanian Bogor, Indonesia
Dr. Ir. Trimuji Ermayanti (Biotek) Lembaga Ilmu Pengetahuan Indonesia, Indonesia Prof. Dr. Ir. Sri Wilarso Budi R, MS (Silvikultur) Institut Pertanian Bogor, Indonesia
Dr. Darwo (Biometrika) Pusat Penelitian dan Pengembangan Hutan, Badan Penelitian, Pengembangan dan Inovasi,
Kementerian Lingkungan Hidup dan Kehutanan, Indonesia Dr.Ir. Nurul Khumaida, M.Si (Silvikultur) Institut Pertanian Bogor, Indonesia
Prof. Dr. Ir. Nurheni Wijayanto, MS (Ilmu Agroforestri) Fakultas Kehutanan, Institut Pertanian Bogor, Indonesia Dr.Ir. Arum Sekar Wulandari, MS (Mikrobiologi, Kultur Jaringan dan Bioteknologi Hutan) Fakultas Kehutanan, Institut Pertanian Bogor, Indonesia
Dr. Dede Rohadi (Ilmu Sosial) Center for International Forestry Research, Indonesia
Copyeditor
Ir. Danu, M.Si (Produksi Benih) Balai Penelitian dan Pengembangan Teknologi Perbenihan Tanaman Hutan, Kementerian Lingkungan Hidup dan Kehutanan, Indonesia
Ratna Uli Damayanti, S.Hut, M.Si (Kultur jaringan, Bioteknologi) Balai Penelitian dan Pengembangan Teknologi Perbenihan Tanaman Hutan, Kementerian
Lingkungan Hidup dan Kehutanan, Indonesia Fifi Gus Dwiyanti, S.Hut, M.Agr., Ph. D (Genetik) Institut Pertanian Bogor, Indonesia
Layout Editor
Tri Astuti Wisudayati, S.E, M.S.E
Sekretariat Dewan Redaksi
Redaksi Pelaksana Ketua Merangkap Anggota
Endah Nurhidajati, S.E., M.Si
(Kepala Seksi Data Informasi dan Sarana Penelitian)
Anggota
Wahyuni Munasri, A.Md
Diterbitkan oleh
Balai Penelitian dan Pengembangan Teknologi Perbenihan Tanaman Hutan, Badan Penelitian Pengembangan dan Inovasi, Kementerian Lingkungan Hidup dan Kehutanan
Terbit Pertama kali Agustus 1996 dengan judul Tekno Benih (ISSN 1410-1157), sejak Agustus 2003 berganti judul menjadi Info Benih (ISSN 1693-5314),
dan sejak Agustus 2013 berganti judul menjadi Jurnal Perbenihan Tanaman Hutan (ISSN 2354-8568)
Alamat
Balai Penelitian dan Pengembangan Teknologi Perbenihan Tanaman Hutan
Jl. Pakuan Ciheuleut P0 Box 105 Bogor, Telp./fax : (0251)8327768 Website : benih-bogor.litbang.menlhk.go.id

p-ISSN : 2354-8568
e-ISSN : 2527-6565
JURNAL PERBENIHAN TANAMAN HUTAN
Vol. 7 No. 2, Desember 2019
DAFTAR ISI
1. EFEKTIVITAS PENGGUNAAN MEDIA TANAM DAN PUPUK
TERHADAP KEMAMPUAN BERTUNAS TANAMAN PANGKAS KESS
(Lophostemon suaveolens (Sol.ex Gaertn.) Peter G.Wilson & J.T. Waterh)
(The Effectivity of Planting Media and Fertilizer in Sprouting Ability of Kess
(Lophostemon suaveolens (Sol.ex Gaertn.) Peter G.Wilson & J.T. Waterh) Stool
Plant)
Dwi Kartikaningtyas, Teguh Setyaji, dan/and Surip .............................................. 67-76
2. PENGARUH UMUR TUNAS PANGKASAN TERHADAP PERTUMBUHAN
SETEK Hopea odorata Roxb.
(The Influence of Pruning Shoot Age on the Growth of Hopea odorata Roxb.
Cutting)
Nurmawati Siregar dan/and Kurniawati Purwaka Putri ......................................... 77-85
3. KARAKTERISASI PERTUMBUHAN AWAL BEBERAPA FAMILI
MANGLID PADA UJI KETURUNAN DI SUKAMANTRI, KABUPATEN
CIAMIS
(Characterization of Early Growth of Several Families of Manglid on Progeny
Test at Sukamantri, Ciamis District)
Asep Rohandi dan/and Dede J. Sudrajat ..................................................................... 87-100
4. KETAHANAN BIBIT SENGON (Falcataria moluccana (Miq) Barneby & J.W.
Grimes) TERHADAP CENDAWAN Uromycladium falcatarium
BERDASARKAN ASAL BENIH DAN JENIS PENGENDALI
(The Endurance of Sengon (Falcataria moluccana (Miq) Barneby & J.W.
Grimes) Seedling on Uromycladium falcatarium Fungus Based on Seed Source
and Controller Agent)
Tati Suharti, Kurniawati Purwaka Putri, dan/and Yulianti Bramasto .................. 101-111
5. TEKNIK PENYIMPANAN BENIH MERANTI BALAU (Shorea seminis (de
Vriese) Sloot)
(The technique of storage of meranti balau (Shorea seminis (de Vriese) Sloot))
Muhammad Zanzibar, Naning Yuniarti, dan/and Ratna Uli Damayanti S ........... 113-125
6. KERAGAMAN PERTUMBUHAN BIBIT NYAMPLUNG (Calophyllum
inophyllum) ASAL SUMENEP, MADURA
(Growth Diversity of Nyamplung (Calophyllum inophyllum) Seedling from
Sumenep, Madura)
Aam Aminah, Danu dan/and Yulianti Bramasto ..................................................... 127-138

JURNAL PERBENIHAN TANAMAN HUTAN
p-ISSN 2354-8568 Vol.7 No.2, Desember 2019
e-ISSN 2527-6565
Kata kunci bersumber dari artikel. Lembar abstrak ini boleh di copy tanpa ijin dan biaya
UDC/ODC 630*245.13
Dwi Kartikaningtyas, Teguh Setyaji, dan/and Surip (Balai Besar Penelitian dan Pengembangan
Bioteknologi dan Pemuliaan Tanaman Hutan)
EFEKTIVITAS PENGGUNAAN MEDIA TANAM DAN PUPUK TERHADAP KEMAMPUAN
BERTUNAS TANAMAN PANGKAS KESS (Lophostemon suaveolens (Sol.ex Gaertn.) Peter G.Wilson &
J.T. Waterh))
J. Perbenihan Tanaman Hutan Vol.7 No.2 p. 67-76
Kebutuhan bibit dalam skala besar dibutuhkan untuk menunjang percepatan kegiatan rehabilitasi lahan
diantaranya pada lahan gambut. Untuk memenuhi kebutuhannya diperlukan teknik yang tepat, diantaranya
dengan perlakuan tanaman pangkas yang tepat sebagai penyedia materi genetik perbanyakan vegetatif.
Penelitian ini bertujuan untuk mengetahui adanya pengaruh penggunaan media tanam dan pemberian
berbagai jenis pupuk pada tanaman pangkas Kess terhadap pertumbuhan dan penambahan tunas baru.
Rancangan penelitian yang digunakan dalam penelitian ini adalah Rancangan Acak Lengkap (RAL) dengan
pola faktorial 2 x 3 pada 2 jenis media tanam (pasir, cocopeat) dan 3 jenis pupuk (pelepasan terkendali,
majemuk,daun). Hasil penelitian menunjukkan bahwa perlakuan media memberikan pengaruh terhadap
pembentukan tunas baru dan penambahan panjang tunas, namun demikian perlakuan pupuk hanya
berpengaruh terhadap pembentukan tunas. Penggunaan kombinasi media pasir dan pupuk pelepasan
terkendali lebih efektif dibandingkan kombinasi lainnya
Kata kunci : kemampuan bertunas, Lophostemon suaveolens, media tanam, pupuk
UDC/ODC 630*232.5 Nurmawati Siregar dan/and Kurniawati Purwaka Putri (Balai Penelitian dan Pengembangan Teknologi Perbenihan Tanaman Hutan) PENGARUH UMUR TUNAS PANGKASAN TERHADAP PERTUMBUHAN SETEK
Hopea odorata Roxb.
J. Perbenihan Tanaman Hutan Vol.7 No.2 p. 77-85
Alternatif perbanyakan Hopea odorata Roxb. diantaranya melalui perbanyakan vegetative dengan setek.
Bahan setek dengan tingkat juvenilitas tinggi diperoleh dengan cara pemangkasan stock plant. Informasi
umur tunas pangkasan terbaik sebagai bahan setek masih terbatas. Tujuan penelitian adalah mengetahui
pengaruh umur tunas pangkasan atau trubusan setelah pemangkasan terhadap keberhasilan perbanyakan
setek H. odorata. Rancangan penelitian adalah Rancangan Acak Kelompok dengan perlakuan umur tunas
(3, 4, 5 dan 6 bulan). Masing-masing perlakuan terdapat dalam 5 kelompok dan setiap perlakuan terdiri atas
20 setek. Respon pertumbuhan setek yang diamati adalah persentase setek tumbuh, tinggi tunas, panjang
akar, berat kering akar, berat kering tunas dan kandungan karbon dan nitrogen. Hasil penelitian
menunjukkan umur tunas pangkasan berpengaruh nyata terhadap semua respon pertumbuhan setek yang
diukur. Setek H. odorata dari tunas orthotrop umur 4 dan 5 bulan setelah pemangkasan menghasilkan
setek dengan persentase tumbuh, tinggi tunas, panjang akar, berat kering tunas dan akar yang lebih tinggi
dibandingkan tunas orthotrop umur 3 dan 6 bulan setelah pemangkasan. Tunas pangkasan umur 6 bulan
menghasilkan nilai rasio C/N (18,14) dan kandungan karbon tertinggi (47,48 persen), tetapi kandungan
nitrogen terendah (2,62 persen). Batang stek dari tunas orthotrop umur 4 dan 5 bulan setelah pemangkasan
hampir 50 persen sudah berkayu, sedangkan tunas umur 6 bulan setelah pemangkasan seluruh bagian
batangnya telah berkayu. Pengambilan materi perbanyakan setek H. odorata sebaiknya dilakukan 5 bulan
setelah pemangkasan karena persentase setek tumbuh yang tinggi (94,18 persen). Selain itu nilai C/N dari
tunas umur 5 bulan (15,11) relatif lebih besar dibanding tunas umur 4 bulan (13,14).
Kata kunci: setek, juvenilitas, Hopea, tunas pangkasan, pohon induk

JURNAL PERBENIHAN TANAMAN HUTAN
p-ISSN 2354-8568 Vol.7 No.2, Desember 2019
e-ISSN 2527-6565
Kata kunci bersumber dari artikel. Lembar abstrak ini boleh di copy tanpa ijin dan biaya
UDC/ODC 630*25
Asep Rohandi1) dan/and Dede J. Sudrajat2) (1)Balai Penelitian dan Pengembangan Teknologi Agroforestry, 2)Balai Penelitian dan Pengembangan Teknologi Perbenihan Tanaman Hutan)
KARAKTERISASI PERTUMBUHAN AWAL BEBERAPA FAMILI MANGLID PADA UJI
KETURUNAN DI SUKAMANTRI, KABUPATEN CIAMIS
J. Perbenihan Tanaman Hutan Vol.7 No.2 p. 87-100
Salah satu hambatan dalam pengembangan hutan rakyat manglid (Magnolia champaca (L.) Baill ex Pierre)
saat ini adalah besarnya keragaman pertumbuhan di lapangan. Oleh sebab itu, informasi karakteristik
morfologi sebagai penciri manglid berkualitas yang memiliki produktivitas tinggi sangat diperlukan.
Penelitian ini bertujuan untuk mengkarakterisasi pertumbuhan awal beberapa famili manglid pada uji
keturunan di Sukamantri, Kabupaten Ciamis. Rancangan percobaan yang digunakan adalah rancangan
acak lengkap berblok terdiri dari 42 famili yang berasal dari Ciamis, Tasikmalaya, Sumedang dan
Majalengka. Setiap famili terdiri dari 3 tanaman yang diulang sebanyak 7 ulangan (blok). Hasil penelitian
menunjukkan bahwa karakteristik pertumbuhan manglid pada umur 1 tahun memiliki keragaman yang
cukup besar (17,96 persen−24,70 persen). Karakter tinggi dan diameter batang manglid memiliki hubungan
yang positif secara kuat dengan lebar tajuk sehingga karakter tersebut dapat dijadikan sebagai kriteria
seleksi genotif manglid dengan produktivitas tinggi. Famili asal Ciamis (101 dan 104) memiliki
keunggulan dalam karakter tinggi total dan tinggi bebas cabang, sedangkan famili 218 (Tasikmalaya) dan
406 (Majalengka) memiliki keunggulan dalam karakter diameter batang. Famili-famili tersebut cukup
potensial sebagai sumber genetik untuk dikembangkan meskipun masih memerlukan evaluasi lebih lanjut.
Kata kunci : karakteristik pertumbuhan, keragaman, manglid, produktivitas
UDC/ODC 630*443.2
Tati Suharti, Kurniawati Purwaka Putri, dan/and Yulianti Bramasto
(Balai Penelitian dan Pengembangan Teknologi Perbenihan Tanaman Hutan)
KETAHANAN BIBIT SENGON (Falcataria moluccana (Miq) Barneby & J.W. Grimes) TERHADAP
CENDAWAN Uromycladium falcatarium BERDASARKAN ASAL BENIH DAN JENIS
PENGENDALI
J. Perbenihan Tanaman Hutan Vol.7 No.2 p. 101-111
Penyakit karat puru yang disebabkan cendawan Uromycladium falcatarium merupakan salah satu jenis
penyakit pada tanaman sengon (Falcataria moluccana) di persemaian dan di lapangan. Tujuan penelitian
ini adalah untuk mengetahui pengaruh asal benih dan jenis pengendali terhadap ketahanan bibit sengon
dari infeksi cendawan U. falcatarium di persemaian. Rancangan penelitian yang digunakan adalah
Rancangan Faktorial acak lengkap, terdiri dari 2 faktor, yaitu faktor Asal benih (A1 = Benih asal daerah
endemik karat puru/Kediri); A2 = benih asal bukan endemik karat puru/Cianjur) dan faktor jenis
pengendali (B1 = Kontrol; B2 = Pupuk hayati berupa PGPR (Plant Growth Promoting Rhizobacter) (5 g.l-
1); B3 = Biofungisida (5 g.l-1); B4 = Biofungisida berupa ekstrak daun sirsak (10 g.l-1); B5 = mankozeb (2
g.l-1). Setiap kombinasi perlakuan diulang 4 kali dan masing-masing ulangan terdiri dari 10 bibit. Variabel
pengamatan adalah insidensi, intensitas penyakit, jumlah teliospora dan ketebalan dinding sel epidermis.
Hasil peneltian menunjukkan bahwai interaksi asal benih dan jenis pengendali mempengaruhi jumlah
teliospora dan tebal epidermis. Jumlah teliospora (6,48 teliospora) terendah terdapat pada bibit asal benih
daerah bebas karat puru dengan jenis pengendali larutan ekstrak daun sirsak. Sel epidermis yang paling
tebal (5,43 μ – 5,84 μ) dihasilkan bibit asal benih daerah bukan endemik karat puru dengan jenis pengendali
larutan PGPR, ekstrak daun sirsak dan mankozeb. Intensitas penyakit pada bibit asal benih dari daerah
bukan endemik (3,5 persen) lebih rendah dibandingkan dari daerah endemik (5,2 persen). Larutan PGPR
dan mankozeb efektif mengurangi infeksi cendawan karena insidensi dan intensitas penyakit yang
ditimbulkannya relatif rendah setelah tahap pengendalian ke-3. Kata kunci : asal benih, bibit, epidermis, Falcataria moluccana, karat puru

JURNAL PERBENIHAN TANAMAN HUTAN
p-ISSN 2354-8568 Vol.7 No.2, Desember 2019
e-ISSN 2527-6565
Kata kunci bersumber dari artikel. Lembar abstrak ini boleh di copy tanpa ijin dan biaya
UDC/ODC 630*8232.315
Muhammad Zanzibar, Naning Yuniarti, dan/and Ratna Uli Damayanti S (Balai Penelitian dan
Pengembangan Teknologi Perbenihan Tanaman Hutan)
TEKNIK PENYIMPANAN BENIH MERANTI BALAU (Shorea seminis (de Vriese) Sloot)
J. Perbenihan Tanaman Hutan Vol.7 No.2 p. 113-125
Permasalahan yang dihadapi dari benih meranti balau (Shorea seminis (de Vriese) Sloot) adalah benihnya
berwatak rekalsitran sehingga viabilitasnya cepat menurun selama penyimpanan. Tujuan penelitian ini
adalah untuk mengetahui teknik penyimpanan terbaik benih meranti balau. Penelitian menggunakan
rancangan acak lengkap (RAL) pola faktorial dengan tiga faktor, yaitu faktor wadah simpan (kain belacu
dan kain belacu dalam kotak kayu), faktor ruang simpan (kamar dan AC), dan faktor periode simpan (0
hari, 3 hari, 6 hari, 9 hari, 12 hari, 15 hari dan 18 hari). Ulangan sebanyak 4 (empat) kali, @ 25 butir benih.
Parameter yang diamati adalah kadar air, daya berkecambah, dan nilai perkecambahan. Hasil penelitian
menunjukkan bahwa wadah, ruang, dan periode simpan berpengaruh terhadap nilai kadar air, daya
berkecambah dan nilai perkecambahan. Teknik penyimpanan terbaik untuk benih meranti balau adalah
menggunakan wadah kain blacu dalam kotak kayu, dimasukkan ke dalam ruang suhu kamar. Interaksi
penggunaan wadah kain belacu dalam kotak kayu dengan ruang suhu kamar dapat menekan laju penurunan
viabilitas benih meranti balau selama penyimpanan.
Kata kunci : Shorea seminis (de Vriese) Sloot, penyimpanan, viabilitas benih
UDC/ODC 630*24
Aam Aminah, Danu dan/and Yulianti Bramasto (Balai Penelitian dan Pengembangan Teknologi
Perbenihan Tanaman Hutan)
KERAGAMAN PERTUMBUHAN BIBIT NYAMPLUNG (Calophyllum inophyllum) ASAL SUMENEP,
MADURA
J. Perbenihan Tanaman Hutan Vol.7 No.2 p. 127-138
Salah satu sumber bahan baku biofuel yang berpotensi di Indonesia adalah nyamplung (Calophyllum
inophylum). Kelebihan nyamplung adalah memiliki rendemen minyak yang tinggi sebesar 40 persen―73
persen. Tujuan dari penelitian ini adalah untuk mengidentifikasi keragaman pertumbuhan dan nilai
heritabilitas untuk karakter tinggi dan diameter bibit nyamplung umur enam bulan dari 26 pohon induk
asal Sumenep, Madura. Penelitian menggunakan Rancangan Lengkap Berblok dengan 26 famili asal
Sumenep Madura sebagai perlakuan, setiap perlakuan terdiri dari 3 ulangan, setiap ulangan terdiri dari 8
bibit, sehingga jumlah bibit yang digunakan sebanyak 624 bibit. Hasil penelitian menunjukkan bahwa
terdapat keragaman pertumbuhan (tinggi dan diameter) pada tingkat bibit tanaman nyamplung asal
Sumenep, Madura. Pertumbuhan tinggi dan diameter bibit nyamplung dari 26 famili yang diuji bervariasi
antara 12,52 cm ―21,27 cm dan 3,04 mm ― 4,54 mm. Nilai heritabilitas individu pada tingkat bibit untuk
karakter tinggi dan diameter adalah 0,27 dan 0,16, sedangkan nilai heritabilitas famili adalah yaitu 0,65
untuk karakter tinggi dan 0,52 untuk diameter.
Kata kunci : famili, genetik, heritabilitas, Calophyllum inophyllum

EFEKTIVITAS PENGGUNAAN MEDIA TANAM DAN PUPUK TERHADAP KEMAMPUAN BERTUNAS
TANAMAN PANGKAS KESS (Lophostemon suaveolens) (Sol.ex Gaertn.) Peter G.Wilson & J.T. Waterh)
Dwi Kartikaningtyas, Teguh Setyaji dan Surip
Kontribusi penulis: Dwi Kartikaningtyas dan Teguh Setyaji sebagai kontributor utama
© 2019 BPTPTH All rights reserved. Open access under CC BY-NC-SA license.doi: //doi.org/10.20886/bptpth.2019.7.2.67-76 67
EFEKTIVITAS PENGGUNAAN MEDIA TANAM DAN PUPUK TERHADAP
KEMAMPUAN BERTUNAS TANAMAN PANGKAS KESS (Lophostemon suaveolens (Sol.ex
Gaertn.) Peter G.Wilson & J.T. Waterh)
(The Effectivity of Planting Media and Fertilizer in Sprouting Ability of Kess (Lophostemon
suaveolens (Sol.ex Gaertn.) Peter G.Wilson & J.T. Waterh) Stool Plant)
*Dwi Kartikaningtyas, *Teguh Setyaji, dan/and Surip
Balai Besar Penelitian dan Pengembangan Bioteknologi dan Pemuliaan Tanaman Hutan
Jl. Palagan Tentara Pelajar Km. 15, Purwobinangun, Pakem, Sleman, 55582, Yogyakarta, Indonesia
e-mail: [email protected]
Naskah masuk: 25 Juni 2018; Naskah direvisi:18 Maret 2019; Naskah diterima: 19 Agustus 2019
ABSTRACT
The need for large-scale seedlings is needed to support the speed-up of land rehabilitation among others on
peatlands. To reach the needs, appropriate techniques are needed, as well as the appropriate of stool plant
as a provider of vegetative propagation genetic material. The purpose of the study was to determine the
influence of the planting media and the application of various types of fertilizer on Kess (Lophostemon
suaveolens) stool plant in shoots growth and shoots length. The research was designed by using the
Completely Randomized Design (CRD) with 2 x 3 factorial design on two types of planting media (sand,
cocopeat) and three types of fertilizer (slow release , compound and leaf fertilizer). The results showed that
the planting media gave a real effect to the shoots growth and increasing of shoots length, however the
application of fertilizer only had an effect on shoots growth. The combination of sand medium and controlled
release fertilizer has more effective compared to other combinations. Keywords: fertilizer, Lophostemon suaveolens, planting media, sprouting ability
ABSTRAK
Kebutuhan bibit dalam skala besar dibutuhkan untuk menunjang percepatan kegiatan rehabilitasi lahan
diantaranya pada lahan gambut. Untuk memenuhi kebutuhannya diperlukan teknik yang tepat, diantaranya
dengan perlakuan tanaman pangkas yang tepat sebagai penyedia materi genetik perbanyakan vegetatif.
Penelitian ini bertujuan untuk mengetahui adanya pengaruh penggunaan media tanam dan pemberian
berbagai jenis pupuk pada tanaman pangkas Kess terhadap pertumbuhan dan penambahan tunas baru.
Rancangan penelitian yang digunakan dalam penelitian ini adalah Rancangan Acak Lengkap (RAL) dengan
pola faktorial 2 x 3 pada 2 jenis media tanam (pasir, cocopeat) dan 3 jenis pupuk (pelepasan terkendali,
majemuk,daun). Hasil penelitian menunjukkan bahwa perlakuan media memberikan pengaruh terhadap
pembentukan tunas baru dan penambahan panjang tunas, namun demikian perlakuan pupuk hanya
berpengaruh terhadap pembentukan tunas. Penggunaan kombinasi media pasir dan pupuk pelepasan
terkendali lebih efektif dibandingkan kombinasi lainnya. Kata kunci : kemampuan bertunas, Lophostemon suaveolens, media tanam, pupuk
I. PENDAHULUAN
Gambut merupakan sebuah ekosistem
rawa yang cukup dominan di beberapa wilayah
pengembangan Hutan Tanaman Industri (HTI)
khususnya Sumatera, Kalimantan dan Papua.
Keberadaan ekosistem hutan rawa gambut ini
terus mengalami degradasi dan kerusakan
akibat pola pemanfaatan yang tidak bijaksana
seperti terjadinya kebakaran hutan dan lahan
(Darmawan, Siregar, Sukendi, & Zahrah,
2016) maupun aktivitas penambangan
(Masganti, Wahyunto, Dariah, Nurhayati, &
Rachmiwati, 2014). Sebagai dampak dari
kerusakan tersebut, maka perlu adanya
kegiatan pemulihan ekosistem gambut.
Pemulihan fungsi ekosistem gambut dapat
dilakukan dengan berbagai cara, diantaranya
adalah suksesi alam, restorasi dan rehabilitasi

Jurnal PerbenihanTanaman Hutan Vol.7 No 2 Desember 2019:67-76
p-ISSN : 2354-8568
e-ISSN : 2527-6565
68
(KLHK, 2017). Dalam peraturan menteri
tersebut dijelaskan bahwa salah satu kriteria
tanaman untuk rehabilitasi ekositem gambut
adalah mengutamakan jenis asli dan
kesesuaian lahan. Salah satu tanaman yang
dapat ditemui pada ekosistem gambut adalah
Kess (Lophostemon suaveolens (Sol.ex
Gaertn.) Peter G.Wilson & J.T. Waterh).
Tanaman ini tumbuh di daerah pesisir dan
sering di dataran berawa (Wilson &
Waterhouse, 1982). Kess berbuah secara
periodik setiap tahunnya pada sekitar bulan
Nopember―Desember dan mempunyai ukuran
benih yang kecil serta mudah berkecambah
(https://tropical.theferns.info). Pada kegiatan
rehabilitasi sangat dibutuhkan bibit dalam
skala besar dengan waktu yang cepat, sehingga
musim berbuah seringkali menjadi
penghambat. Selain ketersediaan bibit dalam
skala besar, diharapkan bibit yang tersedia
mempunyai kualitas yang baik.
Salah satu cara yang dapat digunakan
dalam pemenuhan bibit dalam skala besar
adalah dengan perbanyakan vegetatif, dimana
menurut Goh & Monteuuis (2016) dengan
perbanyakan vegetatif dapat dihasilkan
tanaman yang lebih unggul dan seragam. Cara
perbanyakan vegetatif yang dapat dilakukan
diantaranya adalah dengan menggunakan stek
pucuk, dimana materi stek pucuk didapatkan
dari kebun pangkas dengan menggunakan
tanaman hasil seleksi (Adinugraha, Hasnah, &
Waris, 2017). Langkah yang penting dalam
pembangunan kebun pangkas adalah persiapan
indukan/tanaman pangkas (Kartikaningtyas,
2013). Tanaman pangkas diharapkan
mempunyai kemampuan bertunas (sprouting
ability) yang baik sehingga mampu mencukupi
kebutuhan sebagai bahan perbanyakan
vegetatif. Tanaman pangkas dari pohon induk
yang secara fenotipik bagus, pertumbuhan
yang cepat dan dengan heterosigositas tinggi
diharapkan menghasilkan bibit berkualitas
bagus (Mashudi & Susanto, 2013).
Sampai saat ini data dan informasi
mengenai perbanyakan vegetatif jenis Kess
khususnya penambahan tunas baru pada
tanaman pangkas masih sangat terbatas.
Dengan permasalahan tersebut maka penelitian
ini dilakukan dengan tujuan untuk mengetahui
adanya pengaruh penggunaan media tanam
dan pemberian berbagai jenis pupuk pada
tanaman pangkas tanaman Kess terhadap
pembentukan dan penambahan tunas baru.
Hasil penelitian ini diharapkan mampu
memberikan informasi tentang media tanam,
jenis pupuk maupun kombinasi keduanya yang
efektif terhadap pertumbuhan tanaman
pangkas Kess.
II. BAHAN DAN METODE
A. Bahan dan Alat
Penelitian dilakukan di rumah kaca/
greenhouse Balai Besar Penelitian dan
Pengembangan Bioteknologi dan Pemuliaan

EFEKTIVITAS PENGGUNAAN MEDIA TANAM DAN PUPUK TERHADAP KEMAMPUAN BERTUNAS
TANAMAN PANGKAS KESS (Lophostemon suaveolens) (Sol.ex Gaertn.) Peter G.Wilson & J.T. Waterh)
Dwi Kartikaningtyas, Teguh Setyaji dan Surip
69
Tanaman Hutan Yogyakarta pada bulan Juni
sampai dengan September 2017. Bahan yang
dipergunakan pada penelitian ini adalah semai
Kess dan media tanam pasir dan cocopeat,
pupuk pelepasan terkendali (slow release
fertilizer), pupuk majemuk dan pupuk daun.
Pupuk pelepasan terkendali merupakan pupuk
dengan pelepasan nitrogen secara terkendali
sesuai dengan waktu dan jumlah yang
dibutuhkan oleh tanaman, sehingga dapat
mengoptimalkan penyerapan nitrogen oleh
tanaman (Nainggolan, Suwardi, & Darmawan,
2009). Kandungan utama unsur hara yang
terdapat pada masing-masing pupuk adalah
nitrogen, fosfat dan kalium dengan
perbandingan sebagai berikut;17-8-9+3MgO
untuk pupuk pelepasan terkendali; 16-16-16
untuk pupuk majemuk dan 32-10-10 pada
pupuk daun. Peralatan yang dipergunakan
dalam penelitian ini diantaranya adalah
gunting, cutter, mistar, dan alat tulis lainnya.
B. Prosedur Penelitian
Penelitian ini dilakukan dengan tahapan
sebagai berikut:
1. Memilih bahan tanaman berupa semai
umur 3 bulan dengan ukuran seseragam
mungkin dengan diameter batang 2,0 mm
– 3,5 mm, yang kemudian dipergunakan
sebagai tanaman pangkas.
2. Semai yang dipergunakan sebagai tanaman
pangkas kemudian dipindah kedalam
polibag/plastik berukuran 18 cm x 18 cm
dengan menggunakan media pasir dan
cocopeat. Polibag/plastik yang
dipergunakan adalah polibag tanpa lubang,
dengan tujuan media cocopeat akan selalu
lembab dan mempunyai kondisi yang
hampir sama dengan gambut. Penyiraman
dilakukan menyesuaikan dengan kondisi
kelembaban media.
3. Perlakuan pupuk diberikan dengan
menggunakan 3 jenis pupuk, yaitu pupuk
pelepasan terkendali, pupuk majemuk dan
pupuk daun. Konsentrasi yang
dipergunakan untuk pupuk pelepasan
terkendali adalah 4,5 gram.liter-1 media,
pemberian pupuk adalah sekali pada awal
perlakuan dengan dicampur rata pada
media. Dosis ini mengacu pada dosis yang
dipakai oleh beberapa persemaian modern
yang dikembangkan oleh HTI (Setyaji,
komunikasi pribadi, 2017). Pupuk
majemuk menggunakan konsentrasi 5
gram.liter-1 dengan pemberian pupuk
seminggu sekali sebanyak 5ml/tanaman
pangkas. Hal ini mengacu pada dosis yang
biasa dipergunakan dalam pemeliharaan
tanaman pangkas dan penelitian
sebelumnya pada tanaman Eucalyptus
pellita oleh Kartikaningtyas (2018);
tanaman Meranti Tembaga oleh Mashudi
& Susanto (2013) dan pada tanaman Jati
(Suwandi & Adinugraha, 2016).
Sedangkan konsentrasi untuk pupuk daun
adalah 2 gram.liter-1 dengan pemberian
pupuk 2x seminggu sebanyak 5

Jurnal PerbenihanTanaman Hutan Vol.7 No 2 Desember 2019:67-76
p-ISSN : 2354-8568
e-ISSN : 2527-6565
70
ml/tanaman pangkas, sesuai dengan yang
tertera pada label aturan pakai.
4. Pemangkasan dilakukan setelah tanaman
pangkas tumbuh stabil.
5. Pengamatan dilakukan pada setiap
minggunya, dengan parameter yang
diamati adalah jumlah tunas dan panjang
tunas.
Rancangan penelitian yang digunakan
dalam penelitian ini adalah Rancangan Acak
Lengkap (RAL) faktorial 2 x 3 dengan 2 jenis
perlakuan yaitu jenis media tanam (2 taraf)
dan jenis pupuk (3 taraf) seperti tersaji pada
Tabel 1. Setiap kombinasi perlakuan diulang
sebanyak 3 kali/ulangan dengan jumlah semai
pada masing-masing ulangan adalah 5
tanaman pangkas, sehingga jumlah unit
pengamatan seluruhnya adalah sebanyak 90
tanaman pangkas.
C. Analisa Data
Data yang diperoleh dilakukan Analisis
Varians (ANOVA) dengan program SAS 9.0,
dan apabila terdapat perbedaan antar perlakuan
pada taraf kepercayaan 95% dilanjutkan
dengan menggunakan analisis DMRT
(Duncan’s Multiple Range Test). Tabel (Table) 1. Kombinasi perlakuan efektivitas media tanam dan pupuk terhadap pertumbuhan
tunas Kess(The combination of the effectivity of planting media and fertilizer on
the growth of Kess shoots)
Media(Media) Pupuk(Fertilizer)
Pelepasan terkendali (slow release) Majemuk(compound) Daun(leaf)
Pasir(sand) M1P1 M1P2 M1P3
Cocopeat(cocopeat) M2P1 M2P2 M2P3
Keterangan (Remark): M1P1: pasir +pupuk pelepasan terkendali; M2P1: cocopeat + pupuk pelepasan terkendali; M1P2
: pasir + pupuk majemuk; M2P2 : cocopeat + pupuk majemuk; M1P3 : pasir + pupuk daun;
M2P3 : cocopeat + pupuk daun(M1P1 : sand + slow release fertilizer ; M2P1 : cocopeat + slow
release fertilizer ; M1P2 : sand + compound fertilizer ; M21P2 : cocopeat + compound fertilizer
; M1P3 : sand + leaf fertilizer ; M2P3 : cocopeat + leaf fertilizer)
III. HASIL DAN PEMBAHASAN
A. Hasil
Efektivitas penggunaan media tanam dan
jenis pupuk yang berbeda terhadap
pertumbuhan tunas dilakukan pada tanaman
pangkas tanaman Kess. Untuk mengetahui
efektivitas penggunaan media tanam dan jenis
pupuk maka dilakukan analisis varians dengan
hasil sebagaimana tersaji pada Tabel 2.
Tabel (Table) 2. Hasil analisis varians efektivitas media tanam dan pupuk terhadap pertumbuhan
tunas Kess(Analysis of variance of the effectivity of planting media and fertilizer
on the growth of Kess shoots) Sumber Variasi
(Source of variation) db
Rerata kuadrat(Mean square)
Jumlah tunas (number of shoots) Panjang tunas (length of shoots)
Media(media) 1 15,769** 304,242***
Pupuk(fertilizer) 2 7,031** 4,005ns
Media*pupuk(mdia*fertilizer) 2 3,847ns 1,07ns
Keterangan (Remark) : ** : Berbeda nyata pada taraf kepercayaan 5%; *** : beda nyata pada taraf
kepercayaan 1%;ns : Tidak berbeda nyata(** : significant at the 5% level; *** :
significant at the 1% level; ns : not significant)
EFEKTIVITAS PENGGUNAAN MEDIA TANAM DAN PUPUK TERHADAP KEMAMPUAN BERTUNAS
TANAMAN PANGKAS KESS (Lophostemon suaveolens) (Sol.ex Gaertn.) Peter G.Wilson & J.T. Waterh)
Dwi Kartikaningtyas, Teguh Setyaji dan Surip
71
Dari hasil analisis varians terlihat bahwa
media tanam dan jenis pupuk berpengaruh
terhadap jumlah tunas yang tumbuh, namun
demikian tidak ditemukan adanya interaksi
antara media tanam dan jenis pupuk. Berbeda
halnya dengan panjang tunas, dimana panjang
tunas hanya dipengaruhi oleh media tanam
(Tabel 2.). Untuk mengetahui lebih lanjut
pengaruh media tanam dan jenis pupuk maka
dilakukan uji lanjut dengan hasil sebagaimana
tertera pada Tabel 3.
Tabel (Table) 3. Hasil uji Duncan efektivitas media tanam dan pupuk terhadap pertumbuhan tunas
Kess (Result of Duncan’s multiple range test of the effectivity of planting media
and fertilizer on the growth of Kess shoots)
Perlakuan(Treatment)
Rerata(Mean)
Jumlah tunas (number
of shoots)(tunas± SD)
Panjang tunas(length of
shoots) (cm± SD)
Media(media) Pasir(sand) 2,8907 ± 1,56A 5,1521 ± 2,77A
Cocopeat(cocopeat) 1,8386 ± 1,09B 0,5307 ± 0,54B
Pelepasan terkendali(slow release) 3,1453 ± 0,96A 3,4659 ± 2,73A
Pupuk(fertilizer) Majemuk(compound) 2,1505±1,61B 2,3980 ± 3,74A
Daun(leaf) 1,9415±1,49B 2,8695±2,81A
Keterangan (Remark): Huruf yang sama menunjukkan tidak terdapat beda nyata(Values followed by
the same letters are not significantly different)
Hasil uji lanjut menunjukkan bahwa
penggunaan media pasir lebih efektif dalam
menstimulasi pertumbuhan tunas Kess, baik
jumlah tunas maupun panjang tunas
dibandingkan dengan penggunaaan media
cocopeat. Sedangkan penggunaan pupuk
pelepasan terkendali menunjukkan hasil yang
lebih baik pada jumlah tunas maupun panjang
tunas dibandingkan dengan pupuk majemuk
maupun pupuk daun.
Kombinasi antara media tanam dan jenis
pupuk yang berbeda memberikan hasil yang
bervariasi terhadap pertumbuhan tunas.
Tabel (Table) 4. Rata-rata pertumbuhan tunas Kess pada masing-masing kombinasi(The growth
mean of Kess shoots on each combination)
Parameter(Trait) Media(Media)
Pupuk(Fertilizer)
Pelepasan terkendali
(slow release) Majemuk(compound) Daun(leaf)
Jumlah tunas (number of shoots)
(tunas± SD)
Pasir(sand) 3,666 ± 1,78 3,084 ± 1,56 2,000 ± 0,89
Cocopeat(cocopeat) 2,560 ± 1,25 1,883 ± 1,07 1,217 ± 0,55
Panjang tunas(length of shoots)
(cm± SD)
Pasir(sand) 5,892 ± 3,66 4,496 ± 2,61 5,142 ± 2,01
Cocopeat(cocopeat) 0,736 ± 0,79 0,597 ± 0,49 0,300 ± 0,24
B. Pembahasan
1. Pertumbuhan Indukan
Penelitian ini menggunakan semai umur 3
bulan yang diperlakukan sebagai
indukan/tanaman pangkas. Persentase hidup
tanaman pangkas pada akhir pengamatan
terendah adalah 80 persen dan tertinggi 100
persen. Hal ini menunjukkan bahwa perlakuan
media tanam dan pupuk yang diberikan pada
pelaksanaan penelitian sesuai dengan syarat
tumbuh tanaman pangkas tanaman Kess.
Selain adanya pertumbuhan yang baik pada

Jurnal PerbenihanTanaman Hutan Vol.7 No 2 Desember 2019:67-76
p-ISSN : 2354-8568
e-ISSN : 2527-6565
72
tanaman pangkas, selama pengamatan
berlangsung tidak ditemukan adanya serangan
hama maupun penyakit tanaman yang dapat
mengganggu pertumbuhan tanaman pangkas.
Pada penelitian ini tidak dilakukan kegiatan
pencegahan hama dan penyakit dengan
pemberian fungisida maupun insektisida di
awal penelitian. Dari hasil tersebut
menunjukkan bahwa tanaman pangkas
tanaman Kess mempunyai daya tahan terhadap
kondisi lingkungan dan hama penyakit
tanaman.
2. Jumlah Tunas
Efektivitas penggunaan media tanam dan
jenis pupuk yang dipergunakan pada penelitian
ini telah memberikan pengaruh terhadap
jumlah tunas yang terbentuk (Tabel 2). Jenis
media tanam maupun jenis pupuk yang
berbeda ternyata memberikan hasil yang
berbeda pula terhadap pembentukan tunas.
Pada penelitian ini pembentukan tunas pada
media pasir mempunyai rata-rata yang lebih
besar dibandingkan dengan media cocopeat
(Tabel 3). Demikian halnya dengan
penggunaan pupuk, dimana pupuk pelepasan
terkendali mempunyai kemampuan yang lebih
baik dalam pembentukan tunas dibandingkan
dengan pupuk majemuk dan pupuk daun
(Tabel 3). Pertumbuhan tunas dapat
dipengaruhi oleh faktor internal (genetik) dan
faktor eksternal (lingkungan). Adanya
perbedaan kondisi lingkungan, yaitu media
tanam, pada penelitian ini telah memberikan
hasil yang berbeda terhadap pembentukan
tunas baru pada stool plant. Selain itu adanya
kegiatan pemangkasan batang tanaman pada
bagian mampu meningkatkan jumlah tunas
yang tumbuh (Pramono & Danu, 2013).
Tekstur media tanam berpengaruh terhadap
kemampuan akar untuk menyerap unsur hara
yang tersedia. Sebagaimana diketahui bahwa
media pasir mempunyai porositas yang lebih
baik dibandingkan media cocopeat, sehingga
mempunyai drainase dan aerasi yang baik bagi
akar untuk menyerap unsur hara. Karakteristik
cocopeat sebagai media sapih/media tanam
adalah mampu menyimpan dan mengikat air
dengan kuat (Irawan & Hidayah, 2014). Media
sapih cocopeat memiliki kadar air dan daya
simpan air sebesar 119 persen dan 695,4
persen (Hasriani, Kalsim, & Sukendro, 2013)
dan memiliki pori mikro yang mampu
menghambat gerakan air lebih besar sehingga
menyebabkan ketersediaan air lebih tinggi
(Istomo & Valentino, 2012). Kemampuan
daya serap air yang tinggi akan menurunkan
kemampuan aerasi pada media tersebut,
sehingga akan menghambat akar untuk
menyerap oksigen. Selain itu kandungan
senyawa tanin yang terdapat pada cocopeat
menjadi penghalang mekanis dalam
penyerapan unsur hara (Irawan & Kafiar,
2015; Sukarman, Kainde, Rombang, &
Thomas, 2012).

EFEKTIVITAS PENGGUNAAN MEDIA TANAM DAN PUPUK TERHADAP KEMAMPUAN BERTUNAS
TANAMAN PANGKAS KESS (Lophostemon suaveolens) (Sol.ex Gaertn.) Peter G.Wilson & J.T. Waterh)
Dwi Kartikaningtyas, Teguh Setyaji dan Surip
73
Penggunaan pupuk pelepasan terkendali
terbukti lebih efektif dalam pembentukan
tunas pada penelitian ini. Sebagaimana
diketahui bahwa tidak semua pupuk yang
diaplikasikan akan diserap oleh tanaman,
namun terdapat sebagian yang hanyut oleh air
maupun terdegradasi. Salah satu kendala
efisiensi penggunaan nitrogen adalah adanya
sifat nitrogen yang mudah menguap (volatile)
sehingga terjadi volatilisasi (hilangnya
nitrogen dalam tanah) (Nainggolan et al.,
2009). Hal ini yang sering terjadi pada
pemberian pupuk majemuk atau pupuk daun,
dimana penguapan nitrogen melalui proses
evaporasi maupun penguapan lebih cepat
dibandingkan daya serap tanaman terhadap
pupuk. Slow release fertilizer merupakan
salah satu modifikasi pupuk yang ditujukan
untuk meningkatkan efisiensi unsur-unsur
yang terdapat di dalam pupuk dengan
mengatur pelepasannya secara lambat atau
bertahap (Pratomo, Suwardi, & Darmawan,
2009). Pupuk dalam bentuk pelepasan
terkendali dapat mengoptimalkan penyerapan
nitrogen oleh tanaman, karena pupuk tersebut
dapat mengendalikan pelepasan unsur nitrogen
sesuai dengan jumlah dan waktu yang
dibutuhkan oleh tanaman (Nainggolan et al.,
2009). Dengan sifat pupuk pelepasan
terkendali yang telah disebutkan maka
penggunaan pupuk ini mampu mengurangi
volatilisasi sehingga lebih efektif apabila
dibandingkan dengan pupuk majemuk maupun
pupuk daun. Nasrullah & Tunggalini (2000)
juga menyatakan bahwa pupuk quick release
dianggap tidak efesien karena tercuci oleh
irigasi dan air hujan sehingga dengan
quickrelease, pemupukan tidak efesien atau
diperlukan dosis dan frekuensi yang lebih
tinggi untuk mendapatkan hasil yang baik.
Kombinasi perlakuan media tanam pasir
dengan pemberian pupuk pelepasan terkendali
pada penelitian ini telah memberikan dampak
yang lebih terhadap pembentukan tunas baru
pada tanaman pangkas Kess dibandingkan
dengan kombinasi lainnya.
3. Panjang Tunas
Pertambahan panjang tunas merupakan
pemanjangan ruas-ruas yang merentang
diantara buku-buku tempat melekatnya daun
(Hidayat, Hendalastuti, & Nurohman, 2007).
Adanya pemanjangan tunas atau batang
merupakan akibat adanya peningkatan jumlah
sel dan meluasnya sel (Gardner, Pearce, &
Roger, 1985) dan pemanjangan tunas lateral
akibat pemangkasan (Irawati & Setiari, 2009).
Adanya pengaruh media tanam terhadap
pertambahan panjang tunas disebabkan karena
pada media tanam terdapat unsur hara,
drainase dan aerasi maupun faktor lingkungan
lainnya yang dapat memicu adanya
pertumbuhan tunas. Hal ini senada dengan
penelitian Mahfudz, Isnaini, & Moko (2006)
yang menunjukkan bahwa media tanam
berpengaruh terhadap tinggi tunas pada stek
pucuk Merbau, maupun panjang tunas stek

Jurnal PerbenihanTanaman Hutan Vol.7 No 2 Desember 2019:67-76
p-ISSN : 2354-8568
e-ISSN : 2527-6565
74
tanaman Jarak Pagar (Hayati, Sabaruddin, &
Rahmawati, 2012) dan stek tanaman Bambang
Lanang (Danu & Putri, 2015). Pada
pengamatan ini, media pasir lebih efektif
dibandingkan media cocopeat terhadap adanya
pertambahan panjang tunas. Seperti telah
dijelaskan pada paragrap sebelumnya bahwa
media pasir mempunyai drainase dan aerasi
yang lebih baik dibandingkan media cocopeat,
sehingga pada media pasir kemampuan akar
tanaman untuk menyerap unsur hara maupun
oksigen akan lebih baik.
Penambahan panjang tunas pada
penelitian ini tidak dipengaruhi perlakuan
pemupukan. Namun demikian dari ketiga
pupuk yang diberikan, pupuk pelepasan
terkendali mempunyai rata-rata panjang tunas
terbesar (3,4659 cm) dibandingkan dengan
pupuk lainnya. Hal ini menunjukkan bahwa
pemberian pupuk pelepasan terkendali lebih
efektif dalam pertumbuhan panjang tunas
dibandingkan dengan pupuk majemuk dan
pupuk daun. Kombinasi media dan pupuk
yang paling efektif dalam penambahan
panjang tunas adalah kombinasi media pasir
dengan penambahan pupuk pelepasan
terkendali.
Pada penelitian Rugayah, Hermida,
Ginting, Agustian, & Agsya (2018), pemberian
pupuk pelepasan terkendali dan pupuk
Nitrogen lainnya dengan media tanah (sub
soil) tidak memberikan pengaruh yang nyata
terhadap pertumbuhan tanaman Kailan, tetapi
tidak demikian halnya dengan kombinasi
pupuk pelepasan terkendali dengan media
tanah yang telah diolah seperti yang dilakukan
oleh Gani (2009) pada tanaman padi dan
tanaman Krisan (Wasito & Komar, 2004).
Sedangkan penggunaan pupuk pelepasan
terkendali pada media pasir memberikan
pengaruh terhadap tinggi rumput bermuda dan
jumlah pucuk pada tanaman rumput
Cynodondactylon var. Tifdwarf (Nasrullah &
Tunggalini, 2000). Hal ini menunjukkan
bahwa media tanam memiliki peranan penting
dalam penyerapan unsur hara oleh akar
tanaman. Sebagaimana diketahui bahwa sub
soil merupakan lapisan tanah yang berada di
bawah lapisan top soil sehingga memiliki
struktur yang padat dan keras (Winarna &
Sutarta, 2003). Sehingga dibutuhkan media
tanam yang mempuyai aerasi dan drainase
yang baik untuk penyerapan unsur hara bagi
pertumbuhan tanaman seperti halnya media
pasir dengan penggunaan pupuk pelepasan
terkendali.
IV. KESIMPULAN
Media tanam dan pemupukan yang tepat
memberikan pengaruh terhadap pembentukan
dan pertumbuhan tunas pada tanaman pangkas.
Penggunaan media tanam pasir telah
mempengaruhi pembentukan tunas baru dan
pertambahan panjang tunas tanaman pangkas
Kess, begitu pula pemberian pupuk pelepasan

EFEKTIVITAS PENGGUNAAN MEDIA TANAM DAN PUPUK TERHADAP KEMAMPUAN BERTUNAS
TANAMAN PANGKAS KESS (Lophostemon suaveolens) (Sol.ex Gaertn.) Peter G.Wilson & J.T. Waterh)
Dwi Kartikaningtyas, Teguh Setyaji dan Surip
75
terkendali. Kombinasi antara media tanam
pasir dan pupuk pelepasan terkendali dengan
konsentrasi 4,5 gram.liter-1 media memberikan
pengaruh yang paling baik terhadap tunas
tanaman pangkas Kess dibandingkan
kombinasi lainnya.
Perlu dilakukan penelitian lebih lanjut
dengan melibatkan parameter lainnya yang
mampu menggambarkan pertumbuhan
tanaman lebih dalam, misalnya dengan
mengukur biomassa stool plant maupun
kemempuan berakar (rooting ability) stek hasil
pangkasan.
UCAPAN TERIMA KASIH
Ucapan terimakasih penulis sampaikan
kepada seluruh anggota Tim Peneliti Populasi
Pemuliaan untuk Unggulan Kayu Pulp di
BBPPBPTH atas bantuan dan kerjasamanya
dalam penelitian ini.
DAFTAR PUSTAKA
Adinugraha, H. A., Hasnah, T. M., & Waris.
(2017). Pertumbuhan tunas beberapa klon jati
terseleksi setelah pemangkasan di persemaian.
Jurnal Ilmu Kehutanan, 11(1), 109–117.
https://doi.org/10.22146/jik.24907
Danu, & Putri, K. P. (2015). Penggunaan media
dan hormon tumbuh dalam perbanyakan stek
Bambang Lanang (Michelia champaca L).
Jurnal Perbenihan Tanaman Hutan, 3(2), 61–
70.
Darmawan, B., Siregar, Y. I., Sukendi, & Zahrah,
S. (2016). Pengelolaan keberlanjutan
ekosistem hutan rawa gambut terhadap
kebakaran hutan dan lahan di Semenanjung
Kampar, Sumatera. Jurnal Manusia Dan
Lingkungan, 23(2), 195–205.
https://doi.org/10.22146/jml.18791
Gani, A. (2009). Keunggulan pupuk majemuk
NPK lambat urai untuk tanaman padi sawah.
Jurnal Penelitian Pertanian Tanaman
Pangan, 28(3), 148–157.
Gardner, F. P., Pearce, R. B., & Roger, L. M.
(1985). Physiologi of Crop Plants. USA: The
Iowa State University Press.
Goh, D., & Monteuuis, O. (2016). Teak. In Y.-S.
Park, J. Bonga, & K. M. Hyeun (Eds.),
Vegetative Propagation of Forest Trees
(Online Edi, pp. 425–440). Seoul, Korea:
National Institute Of Forest Science.
Hasriani, Kalsim, D. K., & Sukendro, A. (2013).
Kajian serbuk sabut kelapa (cocopeat) sebagai
media tanam.
Hayati, E., Sabaruddin, & Rahmawati. (2012).
Pengaruh jumlah mata tunas dan komposisi
media tanam terhadap pertumbuhan setek
tanaman Jarak Pagar (Jatropha curcas L.).
Jurnal Agrista, 16(3), 129–134.
Hidayat, A., Hendalastuti, H., & Nurohman, E.
(2007). Pengaruh ukuran diameter stek batang
Hopea odorata Rox b. dari kebun pangkas
terhadap kemampuan bertunas, berakar, dan
daya hidupnya. Jurnal Penelitian Hutan Dan
Konservasi Alam, 4(1).
Irawan, A., & Hidayah, H. N. (2014). Kesesuaian
penggunaan cocopeat sebagai media sapih
pada politube dalam pembibitan cempaka
(Magnolia elegans (Blume.) H.Keng). Jurnal
Wasian, 1(2), 73–76.
Irawan, A., & Kafiar, Y. (2015). Pemanfaatan
cocopeat dan arang sekam padi sebagai media
tanam bibit cempaka wasian (Elmerrilia
ovalis). In Prosiding Seminar Nasional
Masyarakat Biodiversitas Indonesia (pp. 805–
808).
https://doi.org/10.13057/psnmbi/m010423
Irawati, H., & Setiari, N. (2009). Pertumbuhan
tunas lateral tanaman nilam (Pogostemon
cablin Benth) setelah dilakukan pemangkasan
pucuk pada ruas yang berbeda. Buletin
Anatomi Dan Fisiologi, 17(2).
https://doi.org/10.14710/baf.v17i2.2558
Istomo, & Valentino, N. (2012). Pengaruh
Perlakuan Kombinasi Media terhadap
Pertumbuhan Anakan Tumih
(Combretocarpus rotundatus (Miq.) Danser).
Jurnal Silvikultur Tropika, 3(2), 81–84.

Jurnal PerbenihanTanaman Hutan Vol.7 No 2 Desember 2019:67-76
p-ISSN : 2354-8568
e-ISSN : 2527-6565
76
Kartikaningtyas, D. (2013). Pembangunan kebun
pangkas sebagai penyedia materi genetik
Eucalyptus pellita F. Muell. Informasi Teknis,
11(1), 25–30.
Kartikaningtyas, D. (2018). Respon tunas stool
plant Eucalyptus pellita terhadap variasi tinggi
pangkasan dan pemupukan pada media pasir.
Wana Benih, 19(2), 29–35.
KemenLHK. (2017). Peraturan Menteri
Lingkungan Hidup dan Kehutanan No.
P.16/Menlhk/Setjen/Kum.1/2/2017 tentang
Pedoman Teknis Pemulihan Fungsi Ekosistem
Gambut. Jakarta, Indonesia: Kementerian
Lingkungan Hidup dan Kehutanan.
Lophostemon suaveolens. (n.d.). Retrieved
February 15, 2019, from
http://tropical.theferns.info/viewtropical.php?i
d=Lophostemon+suaveolens
Mahfudz, Isnaini, & Moko, H. (2006). Pengaruh
zat pengatur tumbuh dan media tanam
terhadap pertumbuhan stek pucuk merbau.
Jurnal Penelitian Hutan Tanaman, 3:1(1),
25–34.
Masganti, Wahyunto, Dariah, A., Nurhayati, &
Rachmiwati, Y. (2014). Karakteristik dan
potensi pemanfaatan lahan gambut
terdegradasi di provinsi Riau. Jurnal
Sumberdaya Lahan, 8(1).
https://doi.org/10.2018/jsdl.v8i1.6444.g5747
Mashudi, & Susanto, M. (2013). Kemampuan
Bertunas Stool Plants Meranti Tembaga
(Shorea leprosula Miq.) dari Beberapa
Populasi di Kalimantan. Jurnal Pemuliaan
Tanaman Hutan, 7(2), 119–132.
https://doi.org/10.20886/jpth.2013.7.2.119-
132
Nainggolan, G. D., Suwardi, & Darmawan. (2009).
Pola pelepasan nitrogen dari pupuk tersedia
lambat (Slow Release Fertilizer) Urea-Zeolit-
Asam Humat. Jurnal Zeolit Indonesia, 8(2),
89–96.
Nasrullah, N., & Tunggalini, N. K. W. (2000).
Pengaruh pemupukan urea dan nitrogen slow
release terhadap pertumbuhan dan kualitas
rumput lapangan golf. Jurnal Agronomi
Indonesai, 28(2), 62–65.
Pramono, A. A., & Danu. (2013). Pengaruh
pemangkasan dan pelengkungan terhadap
produksi tunas pada pohon pangkas kayu
bawang (Azadirachta exelsa). Jurnal
Perbenihan Tanaman Hutan, 1(2), 93–101.
Pratomo, K. R., Suwardi, & Darmawan. (2009).
Pengaruh pupuk slow release urea-zeolit-asam
humat (UZA) terhadap produktivitas tanaman
padi var. Ciherang. Jurnal Zeolit Indonesia,
8(2), 83–88.
Rugayah, Hermida, L., Ginting, Y. C., Agustian, J.,
& Agsya, M. P. (2018). Uji aplikasi berbagai
jenis pupuk urea lepas lambat (slow release
urea) terhadap pertumbuhan tanaman kailan
(Brassica oleraceae L.). In Seminar Nasional
SINTA Fakultas Teknik : Riset PT-Eksplorasi
Hulu Demi Hilirisasi Produk (pp. 42–48).
Bandar Lampung.
Sukarman, Kainde, R., Rombang, J., & Thomas, A.
(2012). Pertumbuhan bibit sengon
(Paraserianthes falcataria) pada berbagai
media tumbuh. Eugenia, 18(3).
Suwandi, & Adinugraha, H. A. (2016). Pembuatan
model kebun pangkas Jati di persemaian.
Informasi Teknis, 14(1), 1–10.
Wasito, A., & Komar, D. (2004). Pengaruh jenis
pupuk terhadap pertumbuhan dan produksi
tanaman Krisan. Jurnal Hortikultura, 14(3),
1–5.
Wilson, P. G., & Waterhouse, J. T. (1982). A
review of the genus Tristania R. Br.
(Myrtaceae): a heterogeneous assemblage of
five genera. Australian Journal of Botany,
30(4), 413–446.
https://doi.org/https://doi.org/10.1071/BT9820
413
Winarna, & Sutarta, E. S. (2003). Pertumbuhan
dan serapan hara bibit kelapa sawit pada
medium tanam sub soil tanah Typic Paleudult,
Typic Tropopsamment, dan Typic Hapludult.
Warta PPKS, 11(1).

PENGARUH UMUR TUNAS PANGKASAN TERHADAP PERTUMBUHAN SETEK Hopea odorata Roxb.
Nurmawati Siregar dan Nurmawati Siregar
Kontribusi penulis: Nurmawati Siregar dan Kurniawati Purwaka Putri sebagai kontributor utama
© 2019 BPTPTH All rights reserved. Open access under CC BY-NC-SA license.doi: //doi.org/10.20886/bptpth.2019.7.2.77-85 77
PENGARUH UMUR TUNAS PANGKASAN TERHADAP PERTUMBUHAN SETEK
Hopea odorata Roxb.
(The Influence of Pruning Shoot Age on the Growth of Hopea odorata Roxb. Cutting)
*Nurmawati Siregar dan/and *Kurniawati Purwaka Putri
Balai Penelitian dan Pengembangan Teknologi Perbenihan Tanaman Hutan
Jl. Pakuan Ciheuleut PO.BOX 105 Telp/ Fax. 0251-8327768 Kode Pos 16001, Bogor, Jawa Barat, Indonesia
e-mail: [email protected]
Naskah masuk: 20 Juli 2019; Naskah direvisi: 10 September 2019; Naskah diterima: 21 Oktober 2019
ABSTRACT
Propagation of Hopea odorata Roxb. could be carried out through the vegetative methods by cuttings.
Material cuttings with a high juvenility are obtained by pruning a stock plant. Information on shoot age after
pruning as material cuttings were still limited. The purpose of this study was to determine the effect of shoot
age after pruning on the successful propagation of H. odorata cuttings. The study design was a randomized
block design with shoot age treatments of 3, 4, 5 and 6 months. Each treatment was placed in 5 blocks and
each treatment consisted of 20 cuttings. Growth response of cuttings observed was cutting survival
percentage, shoot height, root length, root dry weight, shoot dry weight, carbon, and nitrogen content. The
results showed that the age of pruning shoots had a significant effect on all measured growth response of
cuttings. H. odorata cuttings from orthotropic shoots aged 4 and 5 months after pruning showed cutting
survival percentage, shoot height, root length, shoot and root dry weight were higher than shoots aged 3 and
6 months after pruning. The age of 6 months pruning shoots showed the highest C/N ratio (18.14) and
carbon content (47.48 percent), but but the lowest nitrogen content (2.62 percent). The percentage of woody
on shoots aged 4 and 5 months after pruning was 50%, while almost all of the cuttings on shoots of the age
of 6 months were woody. The best propagation material for H. odorata cuttings was orthotropic shoots aged
5 months after pruning, because of the high cuttings survival percentage (94.18 percent). In addition C/N
ratio from shoot aged 5 months (15,11) was higher than from shoot aged 4 months (13,14). Keyword: cutting, juvenily, Hopea, shoot pruning, stock plant
ABSTRAK
Alternatif perbanyakan Hopea odorata Roxb. diantaranya melalui perbanyakan vegetative dengan setek.
Bahan setek dengan tingkat juvenilitas tinggi diperoleh dengan cara pemangkasan stock plant. Informasi
umur tunas pangkasan terbaik sebagai bahan setek masih terbatas. Tujuan penelitian adalah mengetahui
pengaruh umur tunas pangkasan atau trubusan setelah pemangkasan terhadap keberhasilan perbanyakan
setek H. odorata. Rancangan penelitian adalah Rancangan Acak Kelompok dengan perlakuan umur tunas (3,
4, 5 dan 6 bulan). Masing-masing perlakuan terdapat dalam 5 kelompok dan setiap perlakuan terdiri atas 20
setek. Respon pertumbuhan setek yang diamati adalah persentase setek tumbuh, tinggi tunas, panjang akar,
berat kering akar, berat kering tunas dan kandungan karbon dan nitrogen. Hasil penelitian menunjukkan
umur tunas pangkasan berpengaruh nyata terhadap semua respon pertumbuhan setek yang diukur. Setek H.
odorata dari tunas orthotrop umur 4 dan 5 bulan setelah pemangkasan menghasilkan setek dengan persentase
tumbuh, tinggi tunas, panjang akar, berat kering tunas dan akar yang lebih tinggi dibandingkan tunas
orthotrop umur 3 dan 6 bulan setelah pemangkasan. Tunas pangkasan umur 6 bulan menghasilkan nilai
rasio C/N (18,14) dan kandungan karbon tertinggi (47,48 persen), tetapi kandungan nitrogen terendah (2,62
persen). Batang stek dari tunas orthotrop umur 4 dan 5 bulan setelah pemangkasan hampir 50 persen sudah
berkayu, sedangkan tunas umur 6 bulan setelah pemangkasan seluruh bagian batangnya telah berkayu.
Pengambilan materi perbanyakan setek H. odorata sebaiknya dilakukan 5 bulan setelah pemangkasan karena
persentase setek tumbuh yang tinggi (94,18 persen). Selain itu nilai C/N dari tunas umur 5 bulan (15,11)
relatif lebih besar dibanding tunas umur 4 bulan (13,14). Kata kunci: setek, juvenilitas, Hopea, tunas pangkasan, pohon induk
I. PENDAHULUAN
Hopea odorata Roxb. merupakan salah
satu anggota famili Dipterocarpaceae yang
tersebar alami di beberapa negara di Asia
Tenggara dan Selatan (Dong, Beadle, Doyle,
& Worledge, 2014); (Sakai et al., 2009). Saat

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019:77-85
p-ISSN : 2354-8568
e-ISSN : 2527-6565
78
ini telah terjadi penurunan populasi H.
odorata yang sangat cepat di sebaran alaminya
akibat eksploitasi berlebihan yang terjadi
selama beberapa tahun (Dong et al., 2014).
Oleh karena itu dalam redlist IUCN
(International Union for Conservation of
Nature), H. odorata termasuk dalam kategori
spesies yang menghadapi resiko kepunahan di
alam dalam jangka waktu menengah
(vulnerable tree species) (Ly et al., 2017).
Untuk mempertahankan keberadaannya, maka
upaya konservasi harus dilakukan di antaranya
dengan penanaman baik in-situ maupun ek-
situ.
Ketersediaan bibit bermutu merupakan
salah satu faktor pendukung keberhasilan
konservasi H. odorata. Namun penyediaan
bibit melalui teknik perbanyakan generatif
terkendala karakter benihnya yang bersifat
rekalsitran, sehingga ketersediaan bibit
terbatas hanya sesaat setelah musim berbuah.
Perbanyakan vegetatif dengan menggunakan
setek menjadi salah satu alternatif dalam
pengadaan bibit H. odorata. Hal ini
disebabkan tingkat keberhasilan perbanyakan
setek H. odorata sangat tinggi dengan
persentase setek berakar lebih dari 90 persen
(Hidayat, Hendalastuti, & Nurohman, 2007).
Keberhasilan perbanyakan setek di
antaranya dipengaruhi tingkat juvenilitas
(kemudaan) pohon induk (stock plant) sebagai
sumber bahan setek. Bahan setek yang juvenil
(muda) secara kronologis dan fisiologis
menghasilkan persentase berakar yang lebih
tinggi dibandingkan bahan setek yang sudah
dewasa (Abdullateef & Osman, 2012; Danu &
Putri, 2017). (Hae & Funnah, 2011; Zhang et
al., 2013). Hal tersebut telah terbukti pada
beberapa jenis tanaman, antara lain: pinus
(Rasmussen & Hunt, 2010), Dalbergia
melanoxylon (Amri, Lyaruu, Nyomora, &
Kanyeka, 2010), Shorea leprosula (Danu,
Siregar, Wibowo, & Subiakto, 2010), Stevia
rebaudiana (Abdullateef & Osman, 2012),
Styrax benzoin (Putri & Danu, 2014),
Pongamia pinnata (Kurniaty, Putri, & Siregar,
2016) dan Voaconga africana (Kontoh, 2016).
Pemangkasan merupakan salah satu teknik
rejuvenasi stock plant untuk mendapatkan
bahan setek dengan tingkat juvenilitas tinggi
secara kronologis. Bagian tanaman yang dekat
dengan akar adalah bagian tanaman yang
tumbuh pada awal pertumbuhan, sehingga
secara kronologis bagian tanaman tersebut
paling muda (juvenil) (Danu & Putri, 2017).
Hidayat et al. (2007) melaporkan bahwa
rejuvenasi stock plant H. odorata umur 2
tahun dengan teknik pemangkasan setinggi 30
cm mampu menghasilkan tunas-tunas juvenil
dengan persentase setek berakar dengan
kisaran 92,31―95,59 persen. Namun,
informasi ilmiah mengenai umur bahan setek
setelah pemangkasan (rejuvenasi) yang terbaik
dengan tingkat juvenilitas bahan setek yang

PENGARUH UMUR TUNAS PANGKASAN TERHADAP PERTUMBUHAN SETEK Hopea odorata Roxb.
Nurmawati Siregar dan Nurmawati Siregar
79
tinggi khususnya jenis H. odorata masih
terbatas. Informasi ini sangat dibutuhkan
terutama dalam kaitannya dengan manajemen
pengadaan bibit di persemaian. Tujuan
penelitian adalah mengetahui pengaruh umur
tunas pangkasan atau trubusan setelah
pemangkasan terhadap keberhasilan
perbanyakan setek H. odorata.
II. BAHAN DAN METODE
A. Bahan dan Alat
Kegiatan penelitian perbanyakan H.
odorata secara vegetatif (setek) dilakukan di
Persemaian Balai Penelitian dan
Pengembangan Teknologi Perbenihan
Tanaman Hutan (BP2TPH) yang terletak di
Desa Nagrak, Cimahpar, Bogor. Waktu
pelaksanaan kegiatan dimulai dari bulan
Maret sampai dengan Nopember 2015.
Bahan penelitian yang digunakan adalah
tunas orthotrop yang diambil dari trubusan
pada pohon indukan (stock plant) H. odorata
umur 8 tahun yang ditanam di persemaian
BP2TPH. Bahan lainnya adalah tanah dan
pasir sebagai media pengakaran setek.
Peralatan yang digunakan antara lain: polybag,
Rumah Tumbuh Model Sungkup (Gambar 1),
shading net, sprayer, gunting setek, ember,
timbangan, oven dan alat ukur.
B. Prosedur Penelitian
Pohon induk H. odorata dipangkas
dengan tinggi pangkasan 20 cm dari
permukaan tanah. Setelah pemangkasan
dilakukan pemeliharan pohon seperti
penyiangan dan pemupukan untuk
mendapatkan tunas-tunas orthotrop yang sehat
untuk bahan setek. Umur trubusan yang
tumbuh setelah pemangkasan digunakan
sebagai materi setek dalam penelitian ini.
Waktu pengambilan trubusan disesuaikan
dengan masing-masing perlakuan yaitu 3, 4, 5
dan 6 bulan setelah pemangkasan.
Gambar (Figure) 1. Rumah tumbuh model
sungkup untuk
pengakaran setek Hopea
odorata (root chamber
of Hopea odorata
cutting)
Tunas-tunas orthotrop sebagai bahan
setek dipilih dengan kriteria tunas dorman dan
sehat. Tunas terpilih dipotong-potong dengan
ukuran panjang ± 10 cm, dan mempunyai 2
mata tunas (node). Bagian bawah dari tangkai
setek dipotong miring 45°. Selanjutnya tunas-
tunas tersebut dikurangi jumlah daunnya
hingga tersisa 2 helai. Daun tersebut
kemudian dipotong hingga tersisa 1/3 bagian
luasannya.
Setek pucuk diakarkan dalam sungkup
plastik yang disimpan di bawah naungan
(Gambar 1). Media setek adalah campuran
tanah dan pasir steril dengan perbandingan 1 :
1 (v/v) dengan wadah potray ukuran 4,5 cm X
4,5 cm X 12 cm.

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019:77-85
p-ISSN : 2354-8568
e-ISSN : 2527-6565
80
Pengamatan pertumbuhan dan
perkembangan setek dilakukan setelah 12
minggu dari waktu penanaman setek. Peubah
yang diamati meliputi persentase setek tumbuh
(persen), tinggi tunas (mm), panjang akar
(cm), berat kering akar (g) dan berat kering
tunas (g). Kriteria setek tumbuh adalah
apabila akar dan tunas sudah tumbuh, Selain itu
juga dilakukan analisis kandungan nutrisi
bahan setek yaitu karbon dan nitrogen.
Analisis dilakukan di laboratorium SEAMEO
BIOTROP.
Rancangan percobaan yang digunakan
adalah Rancangan Acak Kelompok, dengan
perlakuan umur bahan setek (A) sebanyak 4
taraf perlakuan, yaitu: A1 = 3 bulan, A2 = 4
bulan, A3 = 5 bulan, dan A4 = 6 bulan setelah
pemangkasan. Masing-masing perlakuan
terdapat dalam 5 kelompok dan setiap
perlakuan terdiri atas 20 setek.
Pengelompokkan berdasarkan sungkup plastik.
Model linier dari Rancangan acak kelompok
adalah sebagai berikut:
ij i j ijУ µ τ β= + + +ò ......................................(1)
dimana:
Уij = Respon atau nilai pengamatan dari
perlakuan ke i dan ulangan ke j;
µ = Nilai tengah umum;
τj = Pengaruh perlakuan ke-i;
βj = Pengaruh blok ke-j;
ϵij = Pengaruh galat dari perlakuan ke-i
dan ulangan ke-j.
C. Analisis Data
Data hasil pengamatan kemudian
dianalisis dengan analisis ragam. Apabila
hasil analisis uji-F menunjukkan perbedaan,
maka dilanjutkan dengan uji Tukey (Uji Beda
Nyata Jujur).
III. HASIL DAN PEMBAHASAN
A. Hasil
Hasil analisis ragam menunjukkan bahwa
umur tunas sebagai bahan setek berpengaruh
terhadap semua respon pertumbuhan setek
yang diukur (Tabel 1). Rata-rata persentase
tumbuh, tinggi tunas, panjang akar, berat
kering akar dan tunas serta kandungan unsur
karbon, nitrogen dan C/N tersaji pada Tabel 2.
Tabel (Table) 1. Nilai F hitung pengaruh umur bahan setek terhadap pertumbuhan setek (F value of
the influence of cutting age on the cutting growth).
Parameter (Parameters) F. hitung (F-calculation)
Persentase tumbuh (survival percentage) (%) 16,72 *
Tinggi tunas (shoots height) (cm) 18,15 *
Panjang akar (roots length) (cm) 28,01 *
Berat kering tunas (dry weight of shoots) (g) 13,43 *
Berat kering akar (dry weight of roots) (g) 14,49 *
Kandungan karbon (carbon content) (%)
Kandungan nitrogen (nitrogen content) (%)
C/N (ratio C/N) (%/%)
40,42 *
61,25 *
266,05 *
Keterangan (Remarks): tn = tidak nyata pada taraf uji 0,05 (not significantly at 0,05 level); * =
nyata pada taraf uji 0,05 (significantly at 0,05 level)
PENGARUH UMUR TUNAS PANGKASAN TERHADAP PERTUMBUHAN SETEK Hopea odorata Roxb.
Nurmawati Siregar dan Nurmawati Siregar
81
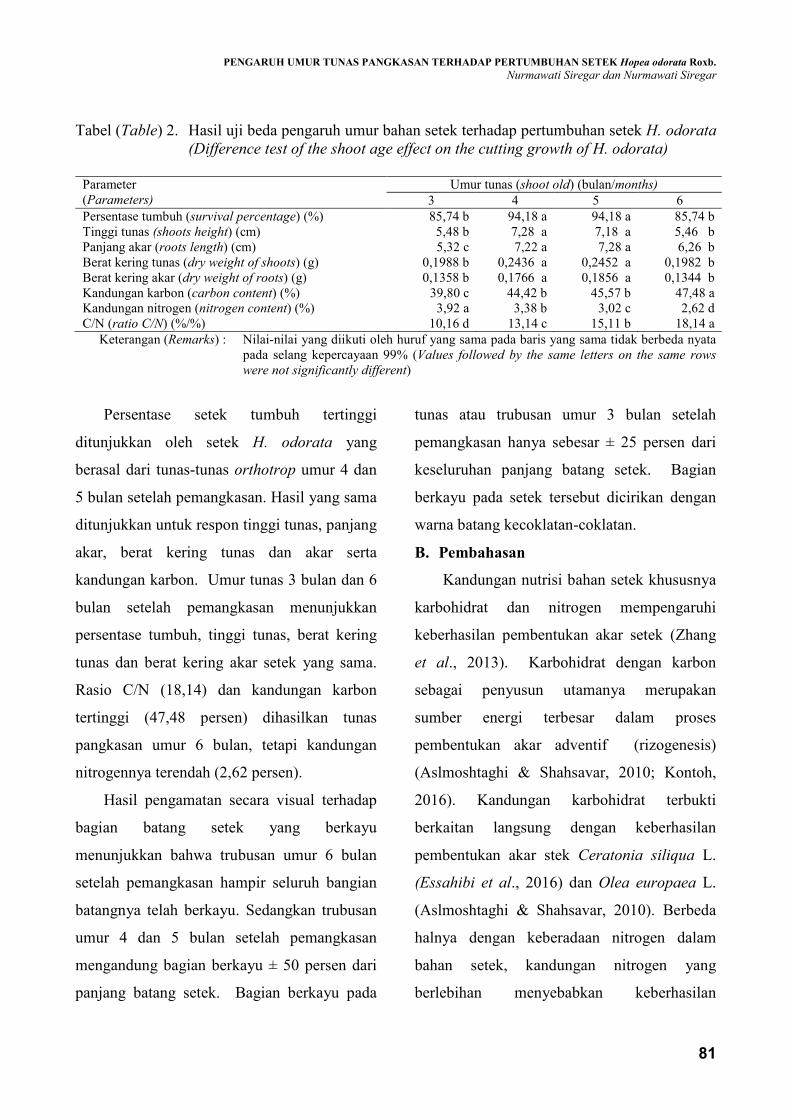
Tabel (Table) 2. Hasil uji beda pengaruh umur bahan setek terhadap pertumbuhan setek H. odorata
(Difference test of the shoot age effect on the cutting growth of H. odorata)
Parameter
(Parameters)
Umur tunas (shoot old) (bulan/months)
3 4 5 6
Persentase tumbuh (survival percentage) (%) 85,74 b 94,18 a 94,18 a 85,74 b
Tinggi tunas (shoots height) (cm) 5,48 b 7,28 a 7,18 a 5,46 b
Panjang akar (roots length) (cm) 5,32 c 7,22 a 7,28 a 6,26 b
Berat kering tunas (dry weight of shoots) (g) 0,1988 b 0,2436 a 0,2452 a 0,1982 b
Berat kering akar (dry weight of roots) (g) 0,1358 b 0,1766 a 0,1856 a 0,1344 b
Kandungan karbon (carbon content) (%) 39,80 c 44,42 b 45,57 b 47,48 a
Kandungan nitrogen (nitrogen content) (%) 3,92 a 3,38 b 3,02 c 2,62 d
C/N (ratio C/N) (%/%) 10,16 d 13,14 c 15,11 b 18,14 a
Keterangan (Remarks) : Nilai-nilai yang diikuti oleh huruf yang sama pada baris yang sama tidak berbeda nyata
pada selang kepercayaan 99% (Values followed by the same letters on the same rows
were not significantly different)
Persentase setek tumbuh tertinggi
ditunjukkan oleh setek H. odorata yang
berasal dari tunas-tunas orthotrop umur 4 dan
5 bulan setelah pemangkasan. Hasil yang sama
ditunjukkan untuk respon tinggi tunas, panjang
akar, berat kering tunas dan akar serta
kandungan karbon. Umur tunas 3 bulan dan 6
bulan setelah pemangkasan menunjukkan
persentase tumbuh, tinggi tunas, berat kering
tunas dan berat kering akar setek yang sama.
Rasio C/N (18,14) dan kandungan karbon
tertinggi (47,48 persen) dihasilkan tunas
pangkasan umur 6 bulan, tetapi kandungan
nitrogennya terendah (2,62 persen).
Hasil pengamatan secara visual terhadap
bagian batang setek yang berkayu
menunjukkan bahwa trubusan umur 6 bulan
setelah pemangkasan hampir seluruh bangian
batangnya telah berkayu. Sedangkan trubusan
umur 4 dan 5 bulan setelah pemangkasan
mengandung bagian berkayu ± 50 persen dari
panjang batang setek. Bagian berkayu pada
tunas atau trubusan umur 3 bulan setelah
pemangkasan hanya sebesar ± 25 persen dari
keseluruhan panjang batang setek. Bagian
berkayu pada setek tersebut dicirikan dengan
warna batang kecoklatan-coklatan.
B. Pembahasan
Kandungan nutrisi bahan setek khususnya
karbohidrat dan nitrogen mempengaruhi
keberhasilan pembentukan akar setek (Zhang
et al., 2013). Karbohidrat dengan karbon
sebagai penyusun utamanya merupakan
sumber energi terbesar dalam proses
pembentukan akar adventif (rizogenesis)
(Aslmoshtaghi & Shahsavar, 2010; Kontoh,
2016). Kandungan karbohidrat terbukti
berkaitan langsung dengan keberhasilan
pembentukan akar stek Ceratonia siliqua L.
(Essahibi et al., 2016) dan Olea europaea L.
(Aslmoshtaghi & Shahsavar, 2010). Berbeda
halnya dengan keberadaan nitrogen dalam
bahan setek, kandungan nitrogen yang
berlebihan menyebabkan keberhasilan

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019:77-85
p-ISSN : 2354-8568
e-ISSN : 2527-6565
82
perakaran setek menurun. Untuk itu
keseimbangan karbohidrat tinggi dan nitrogen
rendah sangat diperlukan untuk keberhasilan
perakaran setek yang sering dinyatakan
dengan perbandingan antara persediaan
karbohidrat dan nitrogen (C/N ratio).
Penelitian menunjukkan bahwa trubusan
H. odorata umur 4 dan 5 bulan setelah
pemangkasan menghasilkan nilai rasio C/N
yang lebih tinggi dibandingkan dengan
trubusan umur 3 bulan setelah pemangkasan.
Kondisi ini seiring dengan tingginya
persentase tumbuh, panjang tunas, panjang
akar dan bobot kering (biomassa) yang
dihasilkan trubusan H. odorata umur 4 dan 5
bulan setelah pemangkasan. Zhang et al.,
(2013) menyebutkan bahwa bahan setek
dengan rasio C/N yang tinggi cenderung akan
lebih mudah berakar. Pengaruh nilai rasio C/N
terhadap persentase keberhasilan berakar setek
juga dilaporkan untuk jenis S. leprosula (Danu
et al., 2010), Calophyllum inophyllum (Danu,
Subiakto, & Abidin, 2011), dan Robinia
pseudoacacia Linn (Ling & Zhong, 2012),
Sanseviera cylindrical (Rapilah, Rahayu, &
Rochman, 2016).
Namun tidak demikian halnya dengan
trubusan H. odorata umur 6 bulan setelah
pemangkasan, Rasio C/N yang dihasilkan
trubusan umur 6 bulan setelah pemangkasan
relatif tinggi tetapi persentase tumbuh setek
yang dihasilkannya relatif rendah. Persentase
tumbuh setek yang berasal trubusan umur 6
bulan setelah pemangkasan sebesar 85,74
persen sedangkan persentase tumbuh stek
setek yang berasal dari trubusan umur 4 dan 5
bulan setelah pemangkasan sebesar 94,18
persen. Hasil tersebut kemungkinan berkaitan
dengan kandungan nitrogen pada bahan setek.
Korelasi kandungan nitrogen dengan
perakaran setek seringkali tidak konsisten,
namun umumnya berkorelasi negatif (Ling &
Zhong, 2012), sehingga tidak dapat
diperkirakan secara pasti nilai C/N yang
terbaik untuk perakaran setek (Danu et al.,
2010). Walaupun demikian keberadaan
nitrogen sangat penting karena nitrogen
berperan dalam sintesis asam nukleat dan
protein, yang diperlukan untuk diferensiasi
akar.
Umur fisiologis bahan setek juga
ditunjukkan oleh kandungan lignin
(Nurhasybi, Danu, Sudrajat, & Dharmawati,
2003). Meskipun terjadi peningkatan kadar
karbohidrat tetapi apabila diikuti dengan
tingginya kandungan lignin maka proses
pembentukan tunas akan terhambat.
Kandungan lignin yang tinggi dicirikan dari
kondisi bahan setek yang sudah berkayu
(keras) dan berwarna coklat (Hidayat et al.,
2007; Santoso & Parwata, 2014). Dalam
penelitian ini trubusan umur 6 bulan
menunjukkan persentase bagian berkayu lebih
besar dibandingkan trubusan umur 4 dan 5

PENGARUH UMUR TUNAS PANGKASAN TERHADAP PERTUMBUHAN SETEK Hopea odorata Roxb.
Nurmawati Siregar dan Nurmawati Siregar
83
bulan. Selain itu produksi biomassa bahan
setek dari trubusan umur 6 bulan lebih rendah
dibandingkan trubusan umur 4 dan 5 bulan.
Produksi biomassa rendah dapat dikaitkan
bahwa setek lebih berkayu, karena sebagian
besar bahan makanan digunakan untuk proses
lignifikasi yang mengakibatkan batang lebih
tinggi mengandung liqnin sebagaimana yang
juga terjadi pada setek Jaropa curcas (Santoso
& Parwata, 2014). Perbedaan kandungan
lignin pada bahan setek inilah kemungkinan
yang menyebabkan terjadinya perbedaan
persentase tumbuh setek H. odorata.
Dipterocarpaceae termasuk kelompok
tanaman dengan tingkat juvenilitas rendah,
sehingga tidak mampu menghasilkan trubusan
atau tunas apabila sudah mencapai umur
kronologis dewasa (Danu & Putri, 2017).
Stock plant Dipterocarpaceae umur 3 tahun―7
tahun akan mulai mengalami penurunan
kualitas dan kuantitas tunas (Nurhasybi et al.,
2003). Akan tetapi dalam penelitian ini
ditunjukkan bahwa dengan teknik rejuvenasi
pemangkasan setinggi 20 cm, tanaman (stock
plant) H. odorata umur 8 tahun dapat
menghasilkan bahan perbanyakan setek yang
bersifat juvenile dengan rata-rata persentase
berakar setek secara keseluruhan mencapai
89,96 persen. Sebagai ilustrasi, dengan teknik
rejuvenilisasi yang sama, persentase setek
berakar yang dihasilkan dari stock plant H.
odorata umur 2 rata-rata mencapai lebih dari
90 persen (Hidayat et al., 2007) dan stock
plant H. odorata umur 5 tahun mencapai 76,6
persen (Nurhasybi et al., 2003).
Berkaitan dengan hasil penelitian ini
secara keseluruhan, maka pengambilan materi
perbanyakan setek H. odorata dapat dilakukan
pada waktu 5 bulan setelah pemangkasan.
Bertambahnya umur tunas pangkasan
cenderung akan mengurangi daya perakaran
setek H. odorata yang dihasilkan. Berbeda
halnya untuk jenis pulai, persentase setek
berakar dari umur pangkas 5 bulan lebih baik
dibandingkan dengan umur pangkas 3 bulan
(Mashudi, 2013). Sedangkan umur tunas
sungkai yang terbaik adalah 10 bulan setelah
pemangkasan dengan pertimbangan materi
setek relatif keras dan berkayu dengan
cadangan makanan relatif banyak (Sahwalita,
2017).
IV. KESIMPULAN
Tunas pangkasan Hopea odorata umur 5
bulan setelah pemangkasan menghasilkan
bahan setek juvenil yang terbaik dengan
persentase setek berakar sebesar 94,18 persen.
UCAPAN TERIMA KASIH
Penulis menyampaikan terima kasih yang
sebesar-besarnya kepada semua pihak yang
telah berpartisipasi dalam pelaksanaan
kegiatan penelitian ini, terutama untuk Bapak
H. Mufid Sanusi dan Bapak Ateng Rahmat
Hidayat yang telah membantu penyiapan
media dan bahan tanaman di persemaian serta
rekan-rekan yang telah memberikan masukan
dan saran dalam pelaksanaan dan penulisan.

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019:77-85
p-ISSN : 2354-8568
e-ISSN : 2527-6565
84
DAFTAR PUSTAKA
Abdullateef, R. A., & Osman, M. (2012). Effects
of stem cutting types, position and hormonal
factors on rooting in Stevia Rebaudiana
Bertoni. Journal of Agricultural Science, 4(1),
49–57. http://doi.org/10.5539/jas.v4n1p49
Amri, E., Lyaruu, H. V. M., Nyomora, A. S., &
Kanyeka, Z. L. (2010). Vegetative
propagation of African Blackwood (Dalbergia
melanoxylon Guill. & Perr.): Effects of age of
donor plant, IBA treatment and cutting
position on rooting ability of stem cuttings.
New Forests, 39, 183–194.
http://doi.org/10.1007/s11056-009-9163-6
Aslmoshtaghi, E., & Shahsavar, A. R. (2010).
Endogenous Soluble Sugars, Starch Contents
and Phenolic Compounds in Easy - and
Difficult – to – Root Olive Cuttings.
J.BioL.Environ.Sci., 4(11), 83–86.
Danu, D., Subiakto, A., & Abidin, A. Z. (2011).
Pengaruh umur pohon induk terhadap
perakaran stek nyamplung(Calophyllum
inophyllum L.). Jurnal Penelitian Hutan
Tanaman, 8(1), 41–49.
http://doi.org/10.20886/jpht.2011.8.1.41-49
Danu, & Putri, K. . (2017). Prinsip pembibitan
secara vegetatif. In I. Z. Siregar & N.
Mindawati (Eds.), Karakteristik dan prinsip
penanganan benih tanaman hutan berwatak
intermediet dan rekalsitran (pp. 187–197).
Bogor: IPB Press.
Danu, Siregar, I. ., Wibowo, C., & Subiakto, A.
(2010). Pengaruh umur bahan stek terhadap
keberhasilan stek pucuk meranti tembaga
(Shorea leprisula MIQ.). Jurnal Penelitian
Hutan Tanaman, 7(3), 1–14.
Dong, T. ., Beadle, C. ., Doyle, R., & Worledge, D.
(2014). Site conditions for regeneration of
hopea odorata in natural evergreen
dipterocarp forest in Southern Vietnam.
Journal of Tropical Forest Science, 26(4),
532–542.
Essahibi, A., Benhiba, L., Fouad, M. O., Babram,
M. A., Ghoulam, C., & Qaddoury, A. (2016).
Initial nutritional status and exogenous IBA
enhanced the rooting capacity of carob
(Ceratonia siliqua L .) cuttings under mist
system . J.Mater.Environ.Sci., 7(11), 4144–
4150.
Hae, M., & Funnah, S. M. (2011). The effect of
propagation media and growth regulars on
rooting potential of kei apple (dovyalis caffra)
stem cuttings at different physiological ages.
Life Science Journal, 8, 91–99.
Hidayat, A., Hendalastuti, H., & Nurohman, E.
(2007). Pengaruh ukuran diameter stek batang
Hopea odorata Roxb. dari kebun pangkas
terhadap kemampuan bertunas, berakar dan
daya hidupnya. Jurnal Penelitian Hutan
Tanaman, 4(1), 1–12.
Kontoh, I. H. (2016). Effect of growth regulators
and soil media on the propagation of
Voacanga africana stem cuttings.
Agroforestry Systems, 90, 479–488.
http://doi.org/10.1007/s10457-015-9870-2
Kurniaty, R., Putri, K. P., & Siregar, N. (2016).
Pengaruh bahan setek dan zat pengatur
tumbuh terhadap keberhasilan setek pucuk
malapari (Pongamia pinnata). Jurnal
Perbenihan Tanaman Hutan, 4(1), 1–8.
http://doi.org/10.20886/jpth.2016.4.1.1-10
Ling, W. X., & Zhong, Z. (2012). Seasonal
variation in rooting of the cuttings from
tetraploid locust in relation to nutrients and
endogenous plant hormones of the shoot.
Turkish Journal of Agriculture and Forestry,
36(2), 257–266. http://doi.org/10.3906/tar-
1011-1511
Ly, V., Newman, M., Khou, E., Barstow, M.,
Hoang, V. S., Nanthavong, K., & Pooma, R.
(2017). Hopea odorata. The IUCN red list of
threatened species 2017:e.T32305A2813234.
Mashudi. (2013). Pengaruh provenan dan
komposisi media terhadap keberhasilan teknik
penunasan pada stek pucuk pulai darat (
Alstonia angustiloba Miq.). Jurnal Penelitian
Hutan Tanaman, 10(1), 25–32.
Nurhasybi, Danu, Sudrajat, D., & Dharmawati.
(2003). Kajian Komprehensif benih tanaman
hutan jenis-jenis Dipterocarpaceae. (D.
Budiantho, Ed.). Bogor: Balai Penelitian dan
Pengembangan Teknologi Perbenihan.
Putri, K. P., & Danu. (2014). Pengaruh umur bahan
stek dan zat pengatur tumbuh terhadap
keberhasilan stek kemenyan ( Styrax benzoin
Dryand ). Jurnal Penelitian Hutan Tanaman,

PENGARUH UMUR TUNAS PANGKASAN TERHADAP PERTUMBUHAN SETEK Hopea odorata Roxb.
Nurmawati Siregar dan Nurmawati Siregar
85
11(3), 141–147.
Rapilah, Rahayu, A., & Rochman, N. (2016).
Pertumbuhan setek Sansvieria cylindrica
“Skyline” pada berbagai ukuran bahan
tanaman dan komposisi media tanam. Jurnal
Agronida, 2(1), 29–36.
Rasmussen, A., & Hunt, M. A. (2010). Ageing
delays the cellular stages of adventitious root
formation in pine. Australian Forestry, 73(1),
41–46.
http://doi.org/10.1080/00049158.2010.106763
08
Sahwalita. (2017). Pengaruh bahan setek terhadap
keberhasilan perbanyakan sungkai dan potensi
setek yang dihasilkan dari kebun pangkas.
Jurnal Perbenihan Tanaman Hutan, 5(1), 23–
34.
Sakai, A., Visaratana, T., Vacharangkura, T., Thai-
ngam, R., Tanaka, N., Ishizuka, M., &
Nakamura, S. (2009). Effect of species and
spacing of fast-growing nurse trees on growth
of an indigenous tree, Hopea odorata Roxb.,
in northeast Thailand. Forest Ecology and
Management, 257, 644–652.
http://doi.org/10.1016/j.foreco.2008.09.048
Santoso, B. B., & Parwata, I. A. (2014). Seedling
growth from stem cutting with different
physiological ages of Jatropha curcas L . of
West Nusa Tenggara genotypes The
University of Mataram. Int. Journal of App.
Scie and Tech., 4(6), 5–10.
Zhang, J., Chen, S., Liu, R., Jiang, J., Chen, F., &
Fang, W. (2013). Chrysanthemum cutting
productivity and rooting ability are improved
by grafting. The Scientific World Journal,
2013. http://doi.org/10.1155/2013/286328

KARAKTERISASI PERTUMBUHAN AWAL BEBERAPA FAMILI
MANGLID PADA UJI KETURUNAN DI SUKAMANTRI, KABUPATEN CIAMIS
Asep Rohandi dan Dede J. Sudrajat
Kontribusi penulis: Asep Rohandi sebagai kontributor utama
© 2019 BPTPTH All rights reserved. Open access under CC BY-NC-SA license.doi: //doi.org/10.20886/bptpth.2019.7.2.87-100 87
KARAKTERISASI PERTUMBUHAN AWAL BEBERAPA FAMILI MANGLID PADA UJI
KETURUNAN DI SUKAMANTRI, KABUPATEN CIAMIS
(Characterization of Early Growth of Several Families of Manglid on Progeny Test at Sukamantri,
Ciamis District)
*Asep Rohandi dan/and Dede J. Sudrajat 1)Balai Penelitian dan Pengembangan Teknologi Agroforestry
Jalan Raya Ciamis - Banjar Km 4, Pamalayan, Ciamis, Jawa Barat, Indonesia 2)Balai Penelitian dan Pengembangan Teknologi Perbenihan Tanaman Hutan
Jl. Pakuan Ciheuleut PO.BOX 105 Telp/ Fax. 0251-8327768 Kode Pos 16001, Bogor, Jawa Barat, Indonesia
e-mail: [email protected]
Naskah masuk: 20 Februari 2019; Naskah direvisi: 14 Juni 2019; Naskah diterima: 6 November 2019
ABSTRACT
One of the obstacles in the development of private forest of manglid (Magnolia champaca (Linn.) Baill ex
Pierre) is the large diversity of growth in the field. Therefore, information on morphological characteristics
as a quality manglid character with high productivity is needed. This study aims to characterize the early
growth of several manglid families in progeny test at Sukamantri, Ciamis District. The experimental design
used was a completely randomized block design consisting of 42 families from Ciamis, Tasikmalaya,
Sumedang and Majalengka. Each family consists of 3 plants (3-tree plot) and 7 replications (blocks). The
results showed that the growth characteristics of manglid at 1 year old had a fairly large diversity (17.96
percent−24,70 percent). The characters of the height and stem diameter have the strong positive correlation
with the crown width so that the characters can be used as the criteria for selection of high productivity
manglid genotypes. Families from Ciamis (101 and 104) have superiority in the character of branch-free
height and total height, while families 218 (Tasikmalaya) and 406 (Majalengka) have advantages in stem
diameter character. These families are quite potential as genetic sources to be developed even though they
still need further evaluation. Keywords : diversity, growth characteristics, manglid, productivity
ABSTRAK
Salah satu hambatan dalam pengembangan hutan rakyat manglid (Magnolia champaca (L.) Baill ex Pierre)
saat ini adalah besarnya keragaman pertumbuhan di lapangan. Oleh sebab itu, informasi karakteristik
morfologi sebagai penciri manglid berkualitas yang memiliki produktivitas tinggi sangat diperlukan.
Penelitian ini bertujuan untuk mengkarakterisasi pertumbuhan awal beberapa famili manglid pada uji
keturunan di Sukamantri, Kabupaten Ciamis. Rancangan percobaan yang digunakan adalah rancangan acak
lengkap berblok terdiri dari 42 famili yang berasal dari Ciamis, Tasikmalaya, Sumedang dan Majalengka.
Setiap famili terdiri dari 3 tanaman yang diulang sebanyak 7 ulangan (blok). Hasil penelitian menunjukkan
bahwa karakteristik pertumbuhan manglid pada umur 1 tahun memiliki keragaman yang cukup besar (17,96
persen−24,70 persen). Karakter tinggi dan diameter batang manglid memiliki hubungan yang positif secara
kuat dengan lebar tajuk sehingga karakter tersebut dapat dijadikan sebagai kriteria seleksi genotif manglid
dengan produktivitas tinggi. Famili asal Ciamis (101 dan 104) memiliki keunggulan dalam karakter tinggi
total dan tinggi bebas cabang, sedangkan famili 218 (Tasikmalaya) dan 406 (Majalengka) memiliki
keunggulan dalam karakter diameter batang. Famili-famili tersebut cukup potensial sebagai sumber genetik
untuk dikembangkan meskipun masih memerlukan evaluasi lebih lanjut. Kata kunci : karakteristik pertumbuhan, keragaman, manglid, produktivitas
I. PENDAHULUAN
Manglid (Magnolia champaca (L.) Baill
ex Pierre) merupakan jenis asli Jawa Barat
yang mempunyai pertumbuhan relatif cepat
(Ahmad, 2016) dengan tinggi rata-rata pada
umur 8 tahun mencapai 13 m dan diameter 14
cm. Jenis ini merupakan kayu tropis penting
dengan nilai komersial cukup tinggi

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019:87-100
p-ISSN : 2354-8568
e-ISSN : 2527-6565
88
(Fernando, Jayasuriya, Walck, & Wijetunga,
2013). Manglid mempunyai nilai ekonomi
cukup baik bagi masyarakat karena pemasaran
kayunya mudah, relatif lebih tahan hama dan
penyakit serta kualitas kayunya relatif lebih
baik (kelas awet II, kelas kuat III-IV)
dibandingkan kayu-kayu yang sudah banyak
dibudidayakan di masyarakat, seperti sengon,
mahoni dan jabon (Sudomo & Mindawati,
2013). Kayu manglid banyak digunakan
sebagai bahan konstruksi ringan, kayu
pertukangan, kerajinan, perabot rumah tangga
dan potensial sebagai baku industri pulp dan
kertas (Siarudin, Sudomo, Indrjaya,
Puspitojati, & Mindawati, 2016). Hingga saat
ini, manglid menjadi jenis andalan setempat di
Jawa Barat dan menjadi salah satu unggulan
untuk hutan rakyat khususnya di wilayah Jawa
Barat Bagian Timur (Rohandi & Gunawan,
2014; Winara, Hani, & Pieter, 2016; Indrjaya,
2016).
Salah satu hambatan dalam
pengembangan hutan rakyat manglid saat ini
adalah besarnya keragaman pertumbuhan di
lapangan yang sangat berpengaruh terhadap
produktivitas dan pendapatan yang diperoleh
masyarakat. Berdasarkan perbedaan
morfologi, masyarakat menggolongkan
manglid dengan beberapa sebutan, seperti
campaka dan baros, yang memiliki
pertumbuhan relatif berbeda. Meskipun
demikian, Winara et al. (2016) melaporkan
bahwa semua manglid yang dikenal dan
dibudidayakan masyarakat hanya terdiri dari
satu jenis dengan nama latin Magnolia
champaca (L.) Baill ex Pierre dan satu variasi
manglid pada tingkat varietas yaitu Magnolia
champaca var. Pubinervia (Blume) Figlar &
Noot. Namun, keragaman antar sub-sub galur
manglid ini belum diketahui termasuk
karakteristik morfologinya sehingga banyak
petani yang menanam manglid mempunyai
pertumbuhan tidak sesuai dengan yang
diharapkan. Keragaman morfologi antar jenis
manglid yang dikenal di masyarakat ini terjadi
pada bagian batang, bunga, daun, tajuk dan
pola pertumbuhannya. Keragaman
pertumbuhan awal juga dilaporkan terjadi pada
beberapa famili Neolamarckia cadamba
(Sudrajat et al., 2016) dan Nuclea orientalis
(Riany, Siregar, & Sudrajat, 2018).
Keragaman tersebut menjadi dasar untuk
melakukan seleksi parameter-parameter yang
diinginkan.
Untuk memberikan informasi karakteristik
morfologi sebagai penciri manglid yang
memiliki pertumbuhan awal tinggi dan
sekaligus mengidentifikasi karakteristik
morfologi manglid untuk mempermudah
penjarangan seleksi pada uji keturunan
manglid di Kabupaten Ciamis, maka penelitian
karakterisasi morfologi manglid ini sangat
diperlukan. Penelitian ini bertujuan untuk
mengkarakterisasi pertumbuhan awal beberapa

KARAKTERISASI PERTUMBUHAN AWAL BEBERAPA FAMILI
MANGLID PADA UJI KETURUNAN DI SUKAMANTRI, KABUPATEN CIAMIS
Asep Rohandi dan Dede J. Sudrajat
89
famili manglid pada uji keturunan di
Sukamantri, Kabupaten Ciamis. Penelitian ini
diharapkan dapat mengidentifikasi famili-
famili manglid yang memiliki karakter
pertumbuhan unggul yang dapat dijadikan
dasar untuk penjarangan seleksi dan sebagai
sumber bahan tanaman untuk pengembangan
jenis ini dalam komersial dalam bentuk hutan
tanaman atau hutan rakyat.
II. BAHAN DAN METODE
A. Bahan dan Alat
Materi genetik manglid yang digunakan
terdiri dari 42 famili dan berasal dari 4 lokasi,
yaitu Ciamis, Tasikmalaya, Sumedang dan
Majalengka (Tabel 1). Sementara itu, alat yang
digunakan meliputi peta tanaman, alat ukur
tinggi, kaliper digitaldan lain-lain. Demplot uji
keturunan manglid berlokasi di Desa
Sukamantri, Kecamatan Sukamantri,
Kabupaten Ciamis. Lokasi tersebut terletak
antara 108°16’24,3” BT sampai 07°04’36,6”
LS dengan ketinggian 957 m dpl. Penelitian
dilaksanakan pada bulan Februari sampai
Maret 2016. Tanaman manglid ditanam pada
bulan Februari 2015 sehingga pada saat
pengamatan tanaman berumur 1 tahun.
Tabel (Table) 1. Lokasi pengumpulan materi genetik manglid untuk pembangunan uji keturunan
di Sukamantri, Kabupaten Ciamis (Locations of genetic material collection of
Magnolia champaca used for progeny test establishment in Sukamantri, Ciamis
District)
Asal pohon induk
(Mother tree origin)
Jumlah famili
(Number of family)
Posisi Koordinat
(Coordinat Position)
Elevasi
(Elevation) (m dpl)
Ciamis 18 7o07’0,09” LS - 108o14’25,3” BT 536
Tasikmalaya 11 7o30’26,6” LS - 108o05’14,3” BT 628
Sumedang 9 6o41’63,7” LS - 107o51’43,9” BT 553
Majalengka 4 6o55’38,3” LS - 108o20’59,5” BT 1157
Keterangan (Remark) : Pengumpulan materi genetik manglid diperoleh dari tegakan hutan tanaman (hutan rakyat)
(Genetic material collection of manglid is obtained from planting forest (cammunity forest).
B. Prosedur Penelitian
Rancangan percobaan yang digunakan
adalah rancangan acak lengkap berblok
(Randomized Complete Block Design).
Penanaman menggunakan 42 famili manglid
yang berasal dari pohon induk di Ciamis,
Tasikmalaya, Sumedang dan Majalengka.
Setiap famili terdiri dari 3 tanaman (3-tree
plot) yang diulang sebanyak 7 ulangan (blok)
dengan jarak tanam 2 m x 3 m. Pengamatan
karakter pertumbuhan manglid dilakukan pada
3 blok yaitu bok 1, 3 dan 5.
Pengamatan dilakukan terhadap beberapa
parameter pertumbuhan awal tegakan manglid
yang meliputi diameter pangkal batang, tinggi
total, tinggi bebas cabang dan lebar tajuk.
Pengukuran diameter batang dilakukan pada
pangkal batang dengan ketinggian 10 cm dari

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019:87-100
p-ISSN : 2354-8568
e-ISSN : 2527-6565
90
pangkal batang (Hadiyan, 2010), sedangkan
tinggi total diukur mulai pangkal batang
sampai pucuk serta tinggi bebas cabang diukur
mulai pangkal batang sampai cabang pertama.
Sementara itu, lebar tajuk merupakan rata-rata
hasil pengukuran tajuk terlebar dan terpendek
(Buba, 2013).
C. Analisis Data
Data pengamatan dianalisis dengan
menggunakan analisis ragam (ANOVA) untuk
mengetahui pengaruh perbedaan famili
terhadap pertumbuhan tanaman. Jika
perlakuan berpengaruh nyata dilanjutkan
dengan Duncan Multiple Range Test (DMRT)
pada α 0,05. Analisis klaster hierarki dan
analiasis komponen utama (Principle
Component Analysis) digunakan untuk
mengelompokkan seluruh famili berdasarkan
parameter yang diamati. Hubungan antar
karakteristik pertumbuhan dianalisis dengan
analisis korelasi sederhana (Pearson).
III. HASIL DAN PEMBAHASAN
A. Hasil
1. Keragaman karakteristik pertumbuhan
Karakterisasi sifat morfologis maupun
fisiologis pada tanaman manglid perlu
dilakukan agar dapat dideskripsikan dan
dimanfaatkan dalam kegiatan seleksi dan
selanjutnya dapat digunakan sebagai sumber
genetik dalam program pengembangan jenis
ini. Hasil analisis ragam menunjukkan bahwa
perbedaan famili berpengaruh sangat nyata
terhadap semua karakteristik pertumbuhan
manglid umur 1 tahun (Tabel 2). Hasil
penelitian terhadap 42 famili tanaman manglid
menunjukkan adanya keragaman karakteristik
morfofisiologi yang cukup besar. Nilai
koefisien keragaman tertinggi diperoleh
karakter tinggi bebas cabang (24,70),
sedangkan koefisien keragaman terendah
diperoleh pada karakter tinggi total (17,96
persen) (Tabel 2).
Tabel (Table) 2. Karakteristik morfologi manglid pada uji keturunan umur 1 tahun di Sukamantri,
Ciamis (Morphological characteristics of manglid on the progeny test at 1 year
age old at Sukamantri, Ciamis)
Parameter
(Parameters)
Maksimum
(Maximum)
Minimum
(Minimum)
Rataan
(Mean)
Standar deviasi
(Standard of
deviation)
Koefisien keragaman
(Coefficient of
variance)
F-Hitung
(F-test)
Diameter batang
(stem diameter) (mm)
45,50 21,00 32,26 5,10 21,02 2,95**
Tinggi bebas cabang
(clear bole height) (cm)
116,40 27,50 58,93 22,95 24,70 13,21**
Tinggi total
(total height) (cm)
226,00 107,00 152,15 22,83 17,96 3,78**
Lebar tajuk
(Crown width) (cm)
183,75 76,50 119,28 18,75 20,37 2,58**
Keterangan (Remark) : ** = berpengaruh sangat nyata pada p = 0.01, * = berpengaruh nyata pada p = 0.05, ns= tidak
berbeda nyata (* = significant at P < 0.05, ** = significant at P > 0.01, ns = no significant)
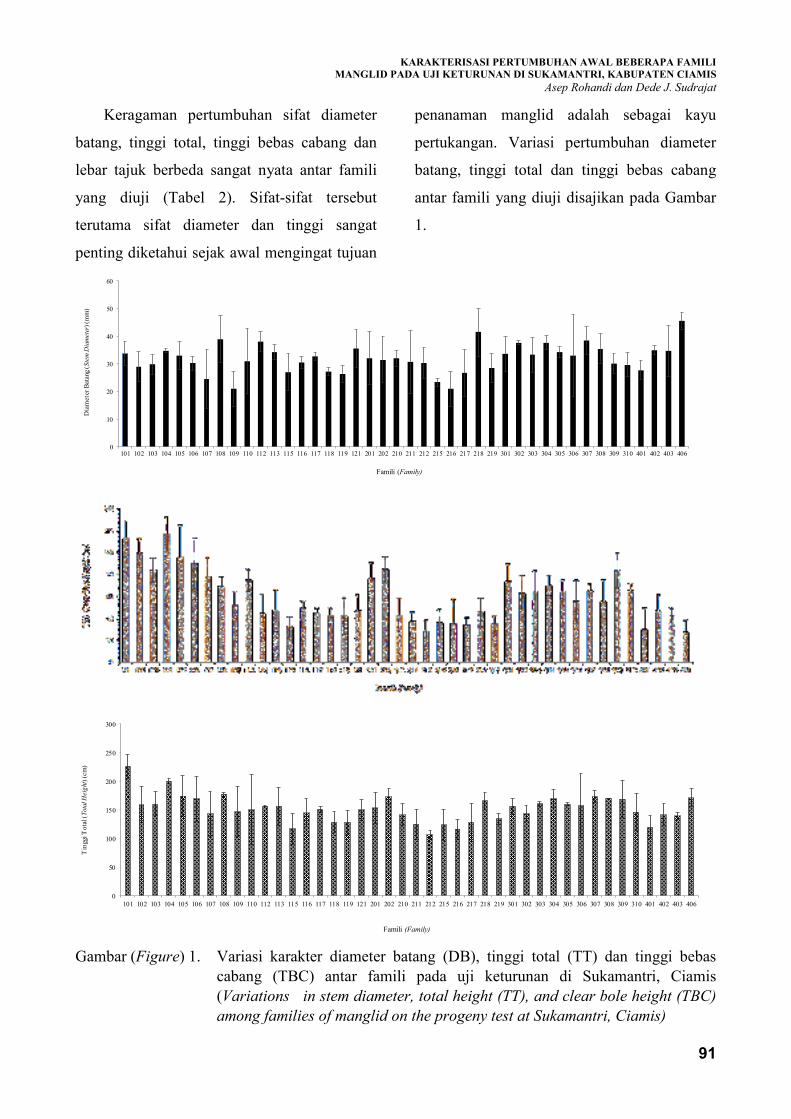
KARAKTERISASI PERTUMBUHAN AWAL BEBERAPA FAMILI
MANGLID PADA UJI KETURUNAN DI SUKAMANTRI, KABUPATEN CIAMIS
Asep Rohandi dan Dede J. Sudrajat
91
Keragaman pertumbuhan sifat diameter
batang, tinggi total, tinggi bebas cabang dan
lebar tajuk berbeda sangat nyata antar famili
yang diuji (Tabel 2). Sifat-sifat tersebut
terutama sifat diameter dan tinggi sangat
penting diketahui sejak awal mengingat tujuan
penanaman manglid adalah sebagai kayu
pertukangan. Variasi pertumbuhan diameter
batang, tinggi total dan tinggi bebas cabang
antar famili yang diuji disajikan pada Gambar
1.
0
10
20
30
40
50
60
101 102 103 104 105 106 107 108 109 110 112 113 115 116 117 118 119 121 201 202 210 211 212 215 216 217 218 219 301 302 303 304 305 306 307 308 309 310 401 402 403 406
Dia
met
er B
atan
g (
Ste
m D
iam
eter
) (m
m)
Famili (Family)
0
50
100
150
200
250
300
101 102 103 104 105 106 107 108 109 110 112 113 115 116 117 118 119 121 201 202 210 211 212 215 216 217 218 219 301 302 303 304 305 306 307 308 309 310 401 402 403 406
Tin
ggi
To
tal (
Tota
l Hei
ght)
(cm
)
Famili (Family)
Gambar (Figure) 1. Variasi karakter diameter batang (DB), tinggi total (TT) dan tinggi bebas
cabang (TBC) antar famili pada uji keturunan di Sukamantri, Ciamis
(Variations in stem diameter, total height (TT), and clear bole height (TBC)
among families of manglid on the progeny test at Sukamantri, Ciamis)

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019:87-100
p-ISSN : 2354-8568
e-ISSN : 2527-6565
92
Berdasarkan data pada Tabel 2 dan
Gambar 1 menunjukkan bahwa rata-rata
pertumbuhan diameter batang manglid umur 1
tahun sebesar 32,26 mm dengan nilai tertinggi
dicapai oleh famili 406 (Majalengka) sebesar
45,50 mm serta nilai terendah sebesar 21,00
mm dicapai famili 109 (Ciamis) yang tidak
berbeda nyata dengan famili 216
(Tasikmalaya). Rata-rata pertumbuhan tinggi
bebas cabang sebesar 27,50 cm dengan nilai
tertinggi dicapai famili 104 (Ciamis) sebesar
116,40 cm dan nilai terendah dicapai famili
212 (Tasikmalaya) sebesar 58,93 cm.
Sementara itu, rata-rata pertumbuhan tinggi
total sebesar 107,00 cm dengan nilai tertinggi
sebesar 226,00 cm dicapai famili 101 (Ciamis)
dan nilai terendah sebesar 152,15 cm dicapai
famili 212 (Tasikmalaya).
2. Hubungan antar karakteristik
pertumbuhan
Keeratan hubungan antar karakter
pertumbuhan dengan morfologi tanaman
manglid dilihat dengan menggunakan analisis
korelasi sederhana (Pearson). Analisis korelasi
digunakan untuk mengukur kekuatan
hubungan antara dua peubah melalui koefsien
korelasi. Hubungan antara karakteristik
pertumbuhan tanaman manglid umur 1 tahun
dapat dilihat pada Tabel 3.
Tabel (Table) 3. Korelasi antar karakteristik morfologi tanaman manglid umur 1 tahun
(Correlation between morphological characteristics of manglid at 1 year age old)
DB TBC TT TJ
DB 0,106 0,547** 0,721**
TBC 0,768** -0,249
TT 0,222
TJ
Keterangan (Remark ): DB = diameter batang, TBC = tinggi bebas cabang, TT = tinggi total, TJ = lebar tajuk, * =
nyata pada tingkat kepercayaan 95%, **=sangat nyata pada tingkat kepercayaan 99%
(DB=stem diameter, TBC=clear bole height, TT=total height, and TJ=crown width * =
Significant at P < 0.05, ** = Significant at P > 0.01)
Data pada Tabel 3 menunjukkan
terjadinya hubungan yang nyata antar sebagian
karakteristik pertumbuhan tanaman manglid
umur 1 tahun. Tinggi total memiliki hubungan
positif dengan diameter pangkal batang
(0,547) dan tinggi bebas cabang (0,768).
Selain berkorelasi positif dengan tinggi total,
diameter pangkal batang juga memiliki
hubungan positif dengan lebar tajuk (0,721).
3. Pengelompokkan famili berdasarkan
karakteristik pertumbuhan
Pengelompokkan famili manglid
menggunakan diagram biplot berdasarkan
analisis klaster hierarki (Gambar 2.a) dan
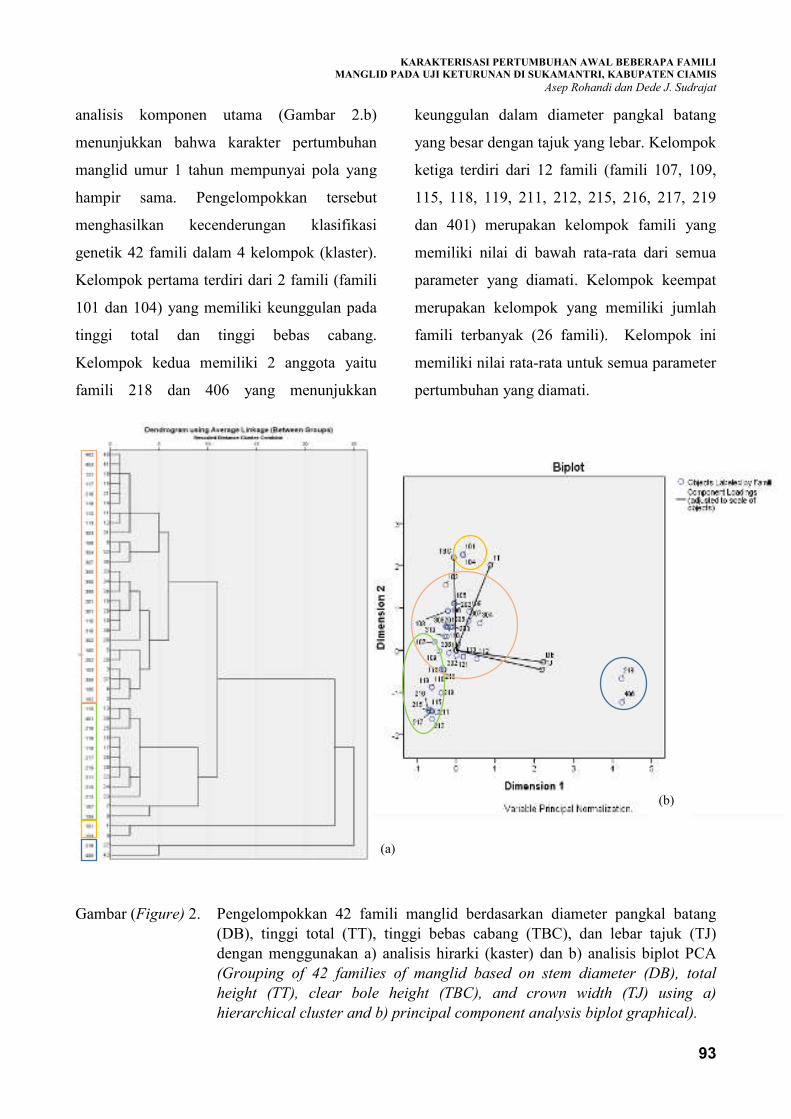
KARAKTERISASI PERTUMBUHAN AWAL BEBERAPA FAMILI
MANGLID PADA UJI KETURUNAN DI SUKAMANTRI, KABUPATEN CIAMIS
Asep Rohandi dan Dede J. Sudrajat
93
analisis komponen utama (Gambar 2.b)
menunjukkan bahwa karakter pertumbuhan
manglid umur 1 tahun mempunyai pola yang
hampir sama. Pengelompokkan tersebut
menghasilkan kecenderungan klasifikasi
genetik 42 famili dalam 4 kelompok (klaster).
Kelompok pertama terdiri dari 2 famili (famili
101 dan 104) yang memiliki keunggulan pada
tinggi total dan tinggi bebas cabang.
Kelompok kedua memiliki 2 anggota yaitu
famili 218 dan 406 yang menunjukkan
keunggulan dalam diameter pangkal batang
yang besar dengan tajuk yang lebar. Kelompok
ketiga terdiri dari 12 famili (famili 107, 109,
115, 118, 119, 211, 212, 215, 216, 217, 219
dan 401) merupakan kelompok famili yang
memiliki nilai di bawah rata-rata dari semua
parameter yang diamati. Kelompok keempat
merupakan kelompok yang memiliki jumlah
famili terbanyak (26 famili). Kelompok ini
memiliki nilai rata-rata untuk semua parameter
pertumbuhan yang diamati.
Gambar (Figure) 2. Pengelompokkan 42 famili manglid berdasarkan diameter pangkal batang
(DB), tinggi total (TT), tinggi bebas cabang (TBC), dan lebar tajuk (TJ)
dengan menggunakan a) analisis hirarki (kaster) dan b) analisis biplot PCA
(Grouping of 42 families of manglid based on stem diameter (DB), total
height (TT), clear bole height (TBC), and crown width (TJ) using a)
hierarchical cluster and b) principal component analysis biplot graphical).
(a)
(b)

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019:87-100
p-ISSN : 2354-8568
e-ISSN : 2527-6565
94
B. Pembahasan
Secara umum, koefisien keragaman
karakteristik pertumbuhan tanaman manglid
umur 1 tahun cukup besar (>20 persen). Hal
tersebut menggambarkan data yang diperoleh
cukup beragam (Widodo et al., 2004) kecuali
untuk karakter tinggi total. Berdasarkan
karakteristik pertumbuhan yang diamati
menunjukkan bahwa famili 406 memiliki nilai
tertinggi untuk dua karakteristik, yaitu
diameter pangkal batang dan lebar tajuk.
Famili 101 memiliki tinggi total tertinggi,
sedangkan famili 104 memiliki nilai tertinggi
untuk tinggi bebas cabang (Lampiran 1).
Hadiyan (2010) menjelaskan bahwa sumber
benih/provenans dan keragaman pada individu
famili menjadi salah satu faktor penting
timbulnya variasi sifat pertumbuhan.
Selanjutnya Setiadi (2011) melaporkan bahwa
adanya variasi sumber benih telah
berkontribusi terhadap pertumbuhan tinggi dan
diameter batang Araucaria cunninghamii umur
1 tahun. Variasi pertumbuhan sifat diameter
dan tinggi dari pohon induk yang berbeda juga
dilaporkan terjadi pada tanaman manglid umur
4 bulan (Pudjiono, 2018) dan tegakan hutan
rakyat manglid pada beberapa lokasi di
wilayah Priangan Timur (Rohandi, 2018).
Perbedaan tersebut dapat disebabkan oleh
perbedaan genetik antar pohon, perbedaan
lingkungan tempat tumbuh ataupun interaksi
antara keduanya (Nai’em, 2004). Perbedaan
morfologi antar populasi untuk jenis-jenis
tanaman hutan, seperti jabon putih (Sudrajat,
2016), suren (Djaman & Sudrajat, 2017) dan
pongamia (Supriyanto, Siregar, Suryani,
Aminah, & Sudrajat, 2017), lebih disebabkan
oleh besarnya kontribusi ragam genetik
dibandingkan ragam lingkungan.
Koefsien korelasi berperan penting dalam
kegiatan seleksi yang dapat digunakan untuk
mengestimasi secara tidak langsung karakter
yang lain. Semakin besar nilai koefisien
korelasi antar karakter maka hubungan antar
karakter tersebut semakin erat (Dermail et al.,
2016). Korelasi antara sifat dengan sifat
lainnya perlu dilakukan sehingga perbaikan
suatu sifat sudah mencakup perbaikan sifat-
sifat yang lain (Mahfudz, Na’iem, Sumardi, &
Hardiyanto, 2010; Sudrajat, Siregar,
Khumaida, Siregar, & Mansur, 2016).
Berdasarkan hubungan antar karakter
pertumbuhan menunjukkan bahwa terjadi
korelasi positif antara tinggi total dengan
diameter batang tanaman manglid umur 1
tahun. Tanaman manglid yang memiliki tinggi
total yang tinggi memiliki diameter batang
yang besar. Hasil yang sama dilaporkan
Hadiyan (2010) pada kebun benih semai
sengon (Falcataria moluccana) umur 4 bulan
yang menunjukkan adanya korelasi yang kuat
antara karakter tinggi dan diameter pangkal
batang sehingga perbaikan satu sifat tersebut
akan memperbaiki sifat lainnya. Hasil lainnya

KARAKTERISASI PERTUMBUHAN AWAL BEBERAPA FAMILI
MANGLID PADA UJI KETURUNAN DI SUKAMANTRI, KABUPATEN CIAMIS
Asep Rohandi dan Dede J. Sudrajat
95
dilaporkan Setiadi (2011) pada jenis A.
cunninghamii umur 1 tahun dan Halawane,
Kinho & Irawan (2015) pada jenis nyatoh
(Palaquium obtusifolium) umur 1,5 tahun.
Selanjutnya Handayani, Kartikaningtyas,
Setyaji, Sunarti, & Nirsatmanto (2018)
menjelaskan bahwa terdapat korelasi genetik
yang tinggi antara tinggi dengan diameter dan
bentuk batang jenis Acacia magium sehingga
seleksi berdasarkan tinggi mampu memberikan
tambahan perolehan genetik pada sifat
diameter dan bentuk batang. Korelasi positif
yang sangat nyata antara tinggi dan diameter
batang juga ditunjukkan pada jenis
Calycophyllum spruceanum (Weber, &
Montes, 2005) dan Neolamarckia cadamba
(Chaerani, Sudrajat, Siregar, & Siregar, 2019).
Hasil lain memperlihatkan bahwa
perbaikan pada karakter lebar tajuk akan
diikuti oleh peningkatan karakter tinggi dan
diameter pangkal batang tanaman manglid,
meskipun asumsi tersebut perlu diteliti lebih
lanjut. Hubungan positif antara lebar tajuk
dengan tinggi dan diameter batang juga
dilaporkan terjadi pada beberapa jenis pohon
di wilayah savana (Arzai, & Aliyu, 2010;
Buba, 2013). Selanjutnya Mahfudz et al.
(2010) menyebutkan bahwa variasi
pertumbuhan tinggi, diameter dan percabangan
merbau (Instia bijuga) pada tingkat individu
dan famili dipengaruhi kuat oleh faktor
genetik. Hal yang sama terjadi pada karakter
lebar tajuk yang juga lebih kuat dipengaruhi
oleh faktor genetik (Priyono, Sumirat, &
Crouzillat, 2011). Karakter-karakter tersebut
potensial digunakan sebagai kriteria seleksi
untuk mendapatkan tanaman manglid yang
adaptif dengan produktivitas tinggi.
Karakter yang digunakan sebagai kriteria
seleksi harus berkorelasi positif dengan
karakter terkait (Wirnas, Widodo, Sobir,
Trikoesoemaningtyas, & Sopandie, 2007).
Sementara itu, Lubis, Sutjahjo, Syukur, &
Trikoesoemaningtyas (2014) menjelaskan
bahwa karakter yang dapat digunakan sebagai
kriteria seleksi selain berkorelasi positif
dengan hasil juga harus memiliki nilai
koefisien keragaman genetik dan heritabilitas
tinggi, agar dapat diwariskan ke generasi
berikutnya. Seleksi tanaman manglid dari
berbagai lokasi di antaranya ditujukan untuk
mendapatkan tanaman manglid yang adaptif,
cepat tumbuh dan memiliki produktivitas
tinggi. Hal tersebut sangat penting untuk
pengembangan jenis ini dalam skala luas
(komersial).
Karakteristik morfologi yang
dikombinasikan dengan metode statistik
multivariate, seperti PCA, sangat umum
digunakan untuk memilih aksesi atau asal
tanaman terbaik pada suatu lokasi (Chalak et
al., 2007; Sorkheh et al., 2010). Hasil
pengelompokkan berdasarkan analisis klaster
hirarki (Gambar 2.a) dan analisis komponen
utama (Gambar 2.b), kelompok pertama
(famili 101 dan 104) dan kedua (famili 218

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019:87-100
p-ISSN : 2354-8568
e-ISSN : 2527-6565
96
dan 406) memiliki potensi yang cukup baik
sebagai kayu pertukangan karena memiliki
keunggulan pada karakter tinggi dan diameter.
Hadiyan (2010) menyebutkan bahwa
keragaman sifat diameter dan tinggi tanaman
sangat penting bagi plot pemuliaan tanaman
hutan untuk tujuan kayu pertukangan,
meskipun beberapa sifat lain seperti tinggi
bebas cabang, bentuk batang, sifat kayu dan
ketahanan serangan hama penyakit
kemungkinan besar akan menjadi variabel
signifikan untuk diintegrasikan dalam proses
seleksi. Hal tersebut didukung laporan
Rohandi (2018) yang menyebutkan bahwa
selain karakter tinggi dan diameter batang,
penampilan tanaman manglid di lapangan juga
memperlihatkan variasi pada karakter bentuk
dan ukuran (morfologi) daun, bentuk batang,
percabangan serta ketahanan terhadap hama
dan penyakit. Karakter-karakter kualitatif
seperti warna batang dan warna daun mungkin
perlu pertimbangkan dalam proses seleksi
sebagai penciri sehingga akan memudahkan
dalam pemilihan manglid berkualitas unggul.
Berdasarkan hasil pengamatan
pertumbuhan manglid umur 1 tahun belum
terlihat adanya famili yang memiliki
keunggulan pada karakter tinggi sekaligus
diameter batang. Hal tersebut diduga karena
potensi genetik dari famili-famili yang diuji
belum dapat diekspresikan dengan baik akibat
masih mudanya umur tanaman. Hadiyan
(2010) menyebutkan bahwa pengaruh faktor
genetik pada tanaman sengon muda (umur 4
bulan) masih belum terekspresi dengan baik.
Hasil penelitian Muslimin, Sofyan, & Islam,
(2013) mengindikasikan bahwa pengaruh
faktor genetik terhadap pertumbuhan klon jati
(Tectona grandis) umur 5,5 tahun relatif masih
sangat kecil. Sementara itu, Susanto &
Baskorowati (2018) melaporkan bahwa
pengaruh faktor genetik (aditif) maupun faktor
lingkungan terhadap keragaman pertumbuhan
sengon ras lahan Jawa berubah-ubah setiap
tahun. Selain itu, pada tanaman jabon putih,
tidak ada korelasi yang signifikan antara
pertumbuhan awal (< 1 tahun) dengan
pertumbuhan tinggi dan diameter tanaman
umur 4 tahun (Sudrajat, Yulianti, Danu,
Rustam, & Suwandhi, 2019). Meskipun
demikian, diketahuinya informasi sejak awal
adanya keragaman karakter tinggi dan
diameter sangat penting untuk mencari
individu-individu unggul jenis kayu
pertukangan (Hadiyan, 2010). Oleh sebab itu,
evaluasi secara periodik perlu terus dilakukan
sehingga dapat ditemukan famili-famili yang
memiliki karakter unggul sebagai sumber
genetik untuk dikembangkan secara luas
(komersial).
IV. KESIMPULAN
Pertumbuhan manglid pada umur 1 tahun
memiliki keragaman yang cukup besar.

KARAKTERISASI PERTUMBUHAN AWAL BEBERAPA FAMILI
MANGLID PADA UJI KETURUNAN DI SUKAMANTRI, KABUPATEN CIAMIS
Asep Rohandi dan Dede J. Sudrajat
97
Karakter pertumbuhan tinggi dan diameter
batang manglid memiliki hubungan positif
yang kuat dengan karakter lebar tajuk.
Terdapat variasi pada karakter diameter
batang, tinggi total dan tinggi bebas cabang
sehingga menjadi peluang untuk dilakukan
seleksi untuk menghasilkan individu-individu
unggul. Famili asal Ciamis (101 dan 104)
memiliki keunggulan dalam karakter tinggi
total dan tinggi bebas cabang, sedangkan
famili 218 (Tasikmalaya) dan 406
(Majalengka) memiliki keunggulan dalam
karakter diameter batang. Famili-famili
tersebut cukup potensial untuk dikembangkan
meskipun perlu dievaluasi lebih lanjut.
UCAPAN TERIMA KASIH
Ucapan terima kasih disampaikan
kepada Balai Penelitian dan Pengembangan
Teknologi Agroforestry, Ciamis yang telah
memberikan dana untuk pelaksanaan kegiatan
penelitian ini. Ucapan terima kasih juga
disampaikan kepada Budi Rahmawan,
Gunawan, Darsono Priono dan Kurniawan
serta semua pihak atas dedikasinya sehingga
penelitian ini dapat terlaksana dengan baik.
DAFTAR PUSTAKA
Ahmad, B. (2016). Perkembangan Tegakan
Manglid (Magnolia champaca) pada Hutan
Rakyat di Kabupaten Tasikmalaya. In Hutan
Rakyat Manglid (Status Riset dan
Pengembangan) (pp. 19–31). Bogor: FORDA
PRESS.
Arzai, A. H., & Aliyu, B. S. (2010). The
relationship between canopy width , height
and trunk size in some tree species growing
in the savana zone of Nigeria. Bayero
Journal of Pure and Applied Sciences, 3(1):
260-263.
Buba, T. (2013). Relationships between stem
diameter at breast height ( DBH ), tree height
, crown length , and crown ratio of Vitellaria
paradoxa C . F . Gaertn in the Nigerian
Guinea Savanna. African Journal of
Biotechnology, 12 (22) : 3441-3446.
https://doi.org/10.5897/AJB12.463
Chaerani, N., Sudrajat, D. J., Siregar, I. Z., &
Siregar, U. J. (2019). Growth performance
and wood quality of white jabon
(Neolamarckia cadamba) progeny testing at
Parung Panjang, Bogor, Indonesia.
Biodiversitas, 20(8), 2295–2301.
https://doi.org/10.13057/biodiv/d200826
Chalak, L, Chehade A., & Kadri, A. 2007.
Morphological characterization of cultivated
almonds in Lebanon, Fruits, 62 (3) : 177-186.
Dermail, A., Maryamah, U., Harahap, Y. P.,
Basrowi, H. A., Anggraeni, D. P., &
Suwarno, W. B. (2016). Seleksi Genotipe
Jagung Berkadar Amilopektin dan Padatan
Terlarut Total Tinggi untuk Mendukung
Diversifkasi Pangan. In Prosiding Seminar
Nasional dan Kongres Perhimpunan
Agronomi Indonesia 27 April 2016. Bogor.
Djaman, D. F., & Sudrajat, D. J. (2017).
Keragaman morfo-fisiologi benih suren
(Toona sinensis) dari berbagai tempat tumbuh
su Sumatera dan Jawa. Jurnal Pemuliaan
Tanaman Hutan, 11 (2), 139–150.
Fernando, M. T. R., Jayasuriya, K. M. G. G.,
Walck, J. L., & Wijetunga, A. S. T. B.
(2013). Identifying dormancy class and
storage behaviour of champak (Magnolia
champaca) seeds, an important tropical
timber tree. Journal of the National Science
Foundation of Sri Lanka, 41 (2) : 141-146.
https://doi.org/10.4038/jnsfsr.v41i2.5708
Hadiyan, Y. (2010). Evaluasi pertumbuhan awal
kebun benih semai uji keturunan sengon
(Falcataria moluccana sinonim :
Paraserianthes falcataria) umur 4 bulan di
Cikampek Jawa Barat. Jurnal Pemuliaan
Tanaman Hutan, 7 (2), 85–91.
Halawane, J., Kinho, J., & Irawan, A. (2015).
Variasi Genetik Pertumbuhan Tanaman Uji

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019:87-100
p-ISSN : 2354-8568
e-ISSN : 2527-6565
98
Keturunan Nyatoh (Palaquium obtusifolium)
umur 1,5 tahun di Hutan Penelitian
Batuangus, Sulawesi Utara. In Prosiding
Seminar Nasional Masyarakat Biodiversitas
Indonesia.
https://doi.org/10.13057/psnmbi/m010426
Handayani, B. R., Kartikaningtyas, D., Setyaji, T.,
Sunarti, S., & Nirsatmanto, A. (2018).
Keragaman Genetik Jenis Introduksi Acacia
auriculiformis Pada Uji Keturunan Generasi
Kedua di Gunungkidul. In Prosiding Seminar
Nasional Masyarakat Biodiversitas
Indonesia. Yogyakarta.
Indrjaya, Y. (2016). Pengaruh Jasa Lingkungan
Karbon Terhadap Daur Optimal Tegakan
Manglid dalam Proyek Aforestasi. In Hutan
Rakyat Manglid (Status Riset dan
Pengembangan) (pp. 115–127). Bogor:
FORDA PRESS.
Lubis, K., Sutjahjo, S. H., Syukur, M., &
Trikoesoemaningtyas. (2014). Pendugaan
parameter genetik dan seleksi karakter
morfofisiologi galur jagung introduksi di
lingkungan tanah masam. Penelitian
Pertanian Tanaman Pangan, 33 (2), 122–
128.
Mahfudz, Na’iem, M., Sumardi, & Hardiyanto, E.
B. (2010). Variasi pertumbuhan pada uji
keturunan merbau (Intsia bijuga O.Ktze) di
Sobang, Banten. Jurnal Pemuliaan Tanaman
Hutan, 4 (3), 157–165.
Muslimin, I., Sofyan, A., & Islam, S. (2013).
Parameter genetik pada uji klon jati (Tectona
grandis L. F) umur 5,5 tahun di Sumatera
Selatan. Pemuliaan Tanaman Hutan, 7 (2),
97–106.
Naiem, M. (2004). Keragaman Genetik, Pemuliaan
Pohon dan Peningkatan Produktivitas Hutan
di Indonesia. Pidato Pengukuhan Jabatan
Guru Besar pada Fakultas Kehutanan.
Universitas Gajah Mada, Yogyakarta.
Priyono, Sumirat, U., & Crouzillat, D. (2011).
Identification of quantitative trait loci
determining vegetative growth traits in
Coffea canephora. Pelita Perkebunan, 27(3),
150–167.
Pudjiono, S. (2018). Pertumbuhan Tanaman
Manglid ( Magnolia champaca ( L ) Baill Ex
Pierre ) Umur Empat Bulan Dari Beberapa
Pohon Induk di Trenggalek Jawa Timur. In
Prosiding Seminar Pendidikan dan Saintek
III. pp. 15–21.
Riany, F., Siregar, I. Z., & Sudrajat, D. J. (2018).
The growth and genetic potentials of gempol
(Nauclea orientalis L .) as shading trees in
urban landscapes. IOP Conf. Series: Earth
and Environmental Science 203 (2018)
012002, 1–14. https://doi.org/10.1088/1755-
1315/203/1/012002
Rohandi, A. (2018). Keragaman Pertumbuhan
Tegakan Manglid : Peluang dan Tantangan
Untuk Peningkatan Produktivitas Hutan
Rakyat di Jawa Barat. Jatinangor.
Rohandi, A., & Gunawan. (2014). Sebaran dan
Karakteristik Hutan Rakyat Manglid serta
Potensinya untuk Pengembangan Sumber
Benih di Wilayah Priangan Timur.
Yogyakarta.
Setiadi, D. (2011). Evaluasi awal kombinasi uji
provenans dan keturunan Araucaria
cunninghamii umur 12 bulan di Bondowoso,
Jawa Timur. Jurnal Ilmu Kehutanan, 5 (1) :
1-8.
Siarudin, M., Sudomo, A., Indrjaya, Y.,
Puspitojati, T., & Mindawati, N. (2016).
Hutan Rakyat Manglid (Status Riset dan
Pengembangan). Bogor : FORDA PRESS.
Sorkheh, K., Shiran, B., Khodambashi, M.,
Moradi, H., Gradziel, T. M., & Martínez-
gómez, P. (2010). Correlations between
quantitative tree and fruit almond traits and
their implications for breeding. Scientia
Horticulturae, 125, 323–331.
https://doi.org/10.1016/j.scienta.2010.04.014
Sudomo, A., & Mindawati, N. (2013). Penggunaan
zat pengatur tumbuh rootone-f pada stek
pucuk manglid (Manglieta glauca BI). Jurnal
Penelitian Hutan Tanaman, 10 (2), 57–63.
Sudrajat, D. J. (2016). Genetic variation of fruit,
seed, and seedling characteristics among 11
populations of white jabon in Indonesia.
Forest Science and Technology, 12 (1), 9–15.
https://doi.org/https://doi.org/10.1080/215801
03.2015.1007896
Sudrajat, D. J., Nurhasybi, N., Siregar, I. Z.,
Siregar, U. J., Mansur, I., & Khumaida, N.
(2016). Intraspecific variation on early

KARAKTERISASI PERTUMBUHAN AWAL BEBERAPA FAMILI
MANGLID PADA UJI KETURUNAN DI SUKAMANTRI, KABUPATEN CIAMIS
Asep Rohandi dan Dede J. Sudrajat
99
growth of Neolamarckia cadamba MIQ. In
provenance-progeny tests in West Java
Province, Indonesia. Biotropia, 23 (1).
https://doi.org/10.11598/btb.2016.23.1.439
Sudrajat, D. J., Siregar, I. Z., Khumaida, N.,
Siregar, U. J., & Mansur, I. (2016).
Keragaman antar populasi dan korelasi antar
karakter bibit jabon putih (Neolamarckia
cadamba (Roxb.) Bosser) pada cekaman
kekeringan dan genangan air. Jurnal
Penelitian Kehutanan Wallacea, 5(1), 13–24.
Sudrajat, D. J., Yulianti, Danu, Rustam, E., &
Suwandhi, I. (2019). Genetic diversity in the
growth of white jabon (Neolamarckia
cadamba) provenance-progeny test:
Comparing study in the nursery and field.
Biodiversitas, 20(5), 1325–1332.
Supriyanto, Siregar, I. Z., Suryani, A., Aminah, A.,
& Sudrajat, D. J. (2017). Keragaman
morfologi buah, benih dan bibit pongamia
(Pongamia pinnata (L) Pierre) di Pulau Jawa.
Jurnal Perbenihan Tanaman Hutan, 5(2),
103–114.
Susanto, M., & Baskorowati, L. (2018). Pengaruh
genetik dan lingkungan terhadap
pertumbuhan sengon (Falcataria Molucanna)
ras lahan Jawa. Jurnal Bioeksperimen, 4 (2),
35–41.
Weber, J. C., & Montes, C. S. (2005). Variation
and correlations among stem growth and
wood traits of variation and correlations
among stem growth and wood traits of
Calycophyllum spruceanum Benth . from the
Peruvian Amazon. Silvae Genetica, 5 (1) :
31-41. https://doi.org/10.1515/sg-2005-0005
Widodo, Chozin, M., & Mahmudin. (2004).
Hubungan pertumbuhan dan hasil beberapa
kultivar padi lokal pada tanah gambut dengan
pemberian dolomit. Jurnal Ilmu-Ilmu
Pertanian Indonesia, 6 (2), 75–82.
Winara, A., Hani, A., & Pieter, L. A. G. (2016).
Status Taksonomi dan Morfologi Manglid. In
Hutan Rakyat Manglid (Status Riset dan
Pengembangan) (pp. 11–18). Bogor: FORDA
PRESS.
Wirnas, D., Widodo, I., Sobir,
Trikoesoemaningtyas, & Sopandie, D.
(2007). Pemilihan karakter agronomi untuk
menyusun indeks seleksi pada 11 populasi
kedelai generasi F6. Buletin Agronomi, 34
(1), 19–24.

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019:87-100
p-ISSN : 2354-8568
e-ISSN : 2527-6565
100
Lampiran (Appendix) 1. Karakteristik morfologi tanaman manglid pada umur 1 tahun pada
demplot uji keturunan di Sukamantri, Ciamis (Morphological
characteristics of manglid on the progeny test at 1 year age old at
Sukamantri, Ciamis)
Famili (Lokasi)
Families (Location)
DB
(mm)
TBC
(cm)
TT
(cm)
TJ
(cm)
101 (Ciamis) 33.67 b-h 112.33 a-b 226.00 a 114.00 c-i
102 (Ciamis) 29.00 c-i 99.00 a-c 159.33 b-h 91.17 ij
103 (Ciamis) 29.75 c-i 83.75 c-f 160.50 b-g 102.00 d-j
104 (Ciamis) 34.80 b-g 116.40 a 200.60 ab 123.50 b-i
105 (Ciamis) 33.00 b-h 95.50 b-d 174.33 b-d 117.89 c-i
106 (Ciamis) 30.22 c-i 89.67 c-e 169.67 b-e 112.28 c-j
107 (Ciamis) 24.43 g-i 78.43 c-f 143.43 c-j 95.29 g-j
108 (Ciamis) 38.86 a-c 69.43 e-i 177.14 bc 131.00 b-h
109 (Ciamis) 21.00 i 52.00 h-m 147.00 c-j 76.50 j
110 (Ciamis) 31.00 c-i 74.44 e-g 150.33 c-i 115.61 c-i
112 (Ciamis) 38.14 a-d 44.14 j-n 157.29 c-i 138.43 b-d
113 (Ciamis) 34.33 b-g 47.00 j-n 156.67 c-i 141.92 bc
115 (Ciamis) 27.00 e-i 32.50 mn 117.25 h-j 99.00 e-j
116 (Ciamis) 30.40 c-i 49.20 i-n 145.00 c-j 133.00 b-g
117 (Ciamis) 32.75 b-h 45.00 j-n 150.88 c-i 132.38 b-h
118 (Ciamis) 27.13 e-i 42.13 k-n 128.63 e-j 121.25 b-i
119 (Ciamis) 26.25 f-i 42.25 k-n 128.50 e-j 118.75 c-i
121 (Ciamis) 35.56 b-f 46.67 j-n 149.89 c-i 134.50 b-f
201 (Tasikmalaya) 32.14 b-h 76.00 d-g 153.64 c-i 106.14 c-j
202 (Tasikmalaya) 31.50 b-h 84.13 c-f 173.00 b-d 130.44 b-h
210 (Tasikmalaya) 32.00 b-h 43.22 j-n 142.00 c-j 126.33 b-i
211 (Tasikmalaya) 30.75 c-i 37.00 l-n 124.88 f-j 116.25 c-i
212 (Tasikmalaya) 30.25 c-i 29.00 n 107.00 j 119.50 b-i
215 (Tasikmalaya) 23.50 hi 36.00 l-n 124.42 f-j 115.38 c-i
216 (Tasikmalaya) 21.00 i 35.20 l-n 116.40 ij 97.10 f-j
217 (Tasikmalaya) 26.67 f-i 34.11 l-n 128.50 e-j 119.72 b-i
218 (Tasikmalaya) 41.56 ab 45.89 j-n 166.44 b-f 156.33 ab
219 (Tasikmalaya) 28.43 c-i 35.14 l-n 134.71 c-j 114.21 c-i
301 (Tasikmalaya) 33.57 b-h 73.14 e-h 156.57 c-i 98.00 e-j
302 (Sumedang) 37.57 a-e 62.71 f-k 143.79 c-j 115.71 c-i
303 (Sumedang) 33.33 b-h 64.17 f-k 161.00 b-g 130.75 b-h
304 (Sumedang) 37.67 a-e 70.00 e-i 169.67 b-e 135.58 b-e
305 (Sumedang) 34.20 b-g 64.00 f-k 160.60 b-g 105.80 c-j
306 (Sumedang) 33.00 b-h 55.20 g-l 158.20 c-i 105.00 c-j
307 (Sumedang) 38.50 a-c 64.63 f-j 173.50 b-d 113.63 c-i
308 (Sumedang) 35.44 b-f 55.11 g-l 169.78 b-e 111.17 c-j
309 (Sumedang) 30.00 c-i 83.60 c-f 168.80 b-e 102.20 d-j
310 (Sumedang) 29.63 c-i 65.00 f-j 146.38 c-j 107.38 c-j
401 (Majalengka) 27.67 d-i 29.67 n 120.33 g-j 94.50 h-j
402 (Majalengka) 34.86 b-g 46.43 j-n 142.00 c-j 128.93 b-i
403 (Majalengka) 34.67 b-g 42.50 k-n 139.50 c-j 129.92 b-h
406 (Majalengka) 45.50 a 27.500 n 171.50 b-d 183.75 a
Rata-rata (Average) 32,26 58,93 152,15 119,28
F-hit : Blok (Block) 6,30* 6,07* 12,18** 0,68ns
Famili (Family) 2,78** 13,55** 3,37** 2,67**
CV (%) 21,02 24,70 17,96 20,37
Keterangan (Remark ): DB = diameter batang, TBC = tinggi bebas cabang, TT = tinggi total, TJ = lebar tajuk, * =
nyata pada tingkat kepercayaan 95%, **=sangat nyata pada tingkat kepercayaan 99%
(DB=stem diameter, TBC = clear bole height, TT = total height, and TJ = crown width, * =
Significant at P < 0.05, ** = Significant at P > 0.01)

KETAHANAN BIBIT SENGON (Falcataria moluccana (Miq) Barneby & J.W. Grimes)
TERHADAP CENDAWAN Uromycladium falcatarium BERDASARKAN ASAL BENIH DAN JENIS PENGENDALI
Tati Suharti, Kurniawati Purwaka Putri, dan Yulianti Bramasto
Kontribusi penulis: Tati Suharti, Kurniawati Purwaka Putri, dan Yulianti Bramasto sebagai kontributor utama
© 2019 BPTPTH All rights reserved. Open access under CC BY-NC-SA license.doi: //doi.org/10.20886/bptpth.2017.5.2.101-111 101
KETAHANAN BIBIT SENGON (Falcataria moluccana (Miq) Barneby & J.W. Grimes)
TERHADAP CENDAWAN Uromycladium falcatarium BERDASARKAN ASAL BENIH
DAN JENIS PENGENDALI
(The Endurance of Sengon (Falcataria moluccana (Miq) Barneby & J.W. Grimes) Seedling against
Uromycladium falcatarium Fungus Based on Seed Source and Controller Agent)
Tati Suharti, Kurniawati Purwaka Putri, dan/and Yulianti Bramasto
Balai Penelitian dan Pengembangan Teknologi Perbenihan Tanaman Hutan
Jl. Pakuan Ciheuleut PO Box 105; Telp. (0251) 8327768, Bogor, Jawa Barat, Indonesia
e-mail: [email protected]
Naskah masuk: 2 September 2019; Naskah direvisi: 16 Oktober 2019; Naskah diterima: 18 November 2019
ABSTRACT
Gall rust disease caused by Uromycladium falcatarium fungus is one of sengon (Falcataria moluccana)
diseases in the nursery and field. The purpose of this study was determining the effect of seed sources and
controller type against the resistance of sengon seedlings from infection of Uromycladium falcatarium in the
nursery. The research design used a factorial completely randomized design with two 2 factors i.e sources of
the seed (A1 = seed from the endemic area of gall rust/Kediri; A2 = seed from non-endemic of gall
rust/Cianjur) and the type of cotrollers (B1 = no treatment l; B2 = biological fertilizer of Plant Growth
Promoting Rhizobacteria (PGPR) (5 g.l-1); B3 = Biofungicide (5 g.l-1); B4 = biofungicide of soursop leaf
extract (10 g.l-1); B5 =mancozeb (2 g.l-1). Each treatment combination consisted of 10 seedlings repeated 4
times. The observation variables were the disease incidence and intensity, the number of teliospores and
thickness of the epidermal cell wall. The results of the study showed that the interaction of seed sources and
type of controller affected the number of teliospores and thickness of the epidermis. The lowest number of
teliospores (6.48 teliospores) was found in seedlings from non endemic gall rust areas with the controller of
soursop leaf extract. The thickest epidermal cells (5.43 μ - 5.84 μ) were produced from seedlings from gall
rust-free area with PGPR solution controller, soursop leaf extract and mancozeb. The disease intensity in
seedlings originating from gall-free areas (3.5 percent) is lower than endemic areas (5.2 percent). PGPR
and mancozeb were effective to the infection of fungus because the disease incidence and intensity were low
after the 3rd control stage. Keywords: epidermis, Falcataria moluccana, gall rust, seedling, seed source
ABSTRAK
Penyakit karat puru yang disebabkan cendawan Uromycladium falcatarium merupakan salah satu jenis
penyakit pada tanaman sengon (Falcataria moluccana) di persemaian dan di lapangan. Tujuan penelitian ini
adalah untuk mengetahui pengaruh asal benih dan jenis pengendali terhadap ketahanan bibit sengon dari
infeksi cendawan U. falcatarium di persemaian. Rancangan penelitian yang digunakan adalah Rancangan
Faktorial acak lengkap, terdiri dari 2 faktor, yaitu faktor Asal benih (A1 = Benih asal daerah endemik karat
puru/Kediri); A2 = benih asal bukan endemik karat puru/Cianjur) dan faktor jenis pengendali (B1 = Kontrol;
B2 = Pupuk hayati berupa PGPR (Plant Growth Promoting Rhizobacter) (5 g.l-1); B3 = Biofungisida (5 g.l-
1); B4 = Biofungisida berupa ekstrak daun sirsak (10 g.l-1); B5 = mankozeb (2 g.l-1). Setiap kombinasi
perlakuan diulang 4 kali dan masing-masing ulangan terdiri dari 10 bibit. Variabel pengamatan adalah
insidensi, intensitas penyakit, jumlah teliospora dan ketebalan dinding sel epidermis. Hasil peneltian
menunjukkan bahwai interaksi asal benih dan jenis pengendali mempengaruhi jumlah teliospora dan tebal
epidermis. Jumlah teliospora (6,48 teliospora) terendah terdapat pada bibit asal benih daerah bebas karat puru
dengan jenis pengendali larutan ekstrak daun sirsak. Sel epidermis yang paling tebal (5,43 μ – 5,84 μ)
dihasilkan bibit asal benih daerah bukan endemik karat puru dengan jenis pengendali larutan PGPR, ekstrak
daun sirsak dan mankozeb. Intensitas penyakit pada bibit asal benih dari daerah bukan endemik (3,5 persen)
lebih rendah dibandingkan dari daerah endemik (5,2 persen). Larutan PGPR dan mankozeb efektif
mengurangi infeksi cendawan karena insidensi dan intensitas penyakit yang ditimbulkannya relatif rendah
setelah tahap pengendalian ke-3. Kata kunci : asal benih, bibit, epidermis, Falcataria moluccana, karat puru

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019: 101-111
p-ISSN : 2354-8568
e-ISSN : 2527-6565
102
I. PENDAHULUAN
Sengon (Falcataria moluccana (Miq)
Barneby & J.W. Grimes) dari family Fabaceae
merupakan salah satu jenis tanaman di hutan
rakyat bernilai ekonomis. Hal tersebut
didasarkan dari manfaat kayu sengon yang
beragam antara lain untuk konstruksi ringan
,bahan kemasan ringan, korek api, bahan baku
triplex, kayu lapis, papan partikel dan papan
blok (Krisnawati, Varis, Kallio, & Kanninen,
2011).
Tanaman sengon merupakan jenis cepat
tumbuh dan mempunyai banyak manfaat
sehingga pengusahaan tanaman sengon
meningkat dari tahun ke tahun terbukti dengan
perluasan hutan tanaman sengon di hutan
rakyat sudah ke luar Jawa (Anggraeni, 2018).
Namun saat ini serangan berbagai jenis hama
dan penyakit pada berbagai fase pertumbuhan
tanaman menjadi salah satu faktor pembatas
produktivitas tanaman sengon. Penanaman
sengon umumnya dilakukan secara
monokultur sehingga kondisi ini akan
menyebabnya cepatnya perkembangan hama
atau penyakit termasuk karat puru.
Tahap pertumbuhan tanaman yang sangat
penting adalah saat tanaman ditumbuhkan di
persemaian. Pada tahap tersebut, salah satu
jenis penyakit yang sering ditemukan yaitu
penyakit karat puru yang disebabkan
cendawan Uromycladium tepperianum
(Rahayu, Nor, See, & Saleh, 2009). Penyakit
karat puru dapat menghambat pertumbuhan
bibit dan mematikan bibit (Putri, 2018) bahkan
dapat mematikan pohon sengon (Nurrohmah
& Baskorowati, 2011). Selain pada bibit,
penyakit karat puru ini juga terdapat pada
tanaman muda hingga tegakan di lapangan
yang menyebabkan batang menjadi cacat
sehingga volume dan kualitas kayu berkurang,
bahkan dapat menyebabkan kematian
tanaman hingga 90 persen (Corryanti &
Novitasari, 2015). Oleh karena itu perlu
adanya pengelolaan penyakit salah satunya di
tingkat benih dan bibit untuk mencegah atau
mengurangi perkembangan karat puru.
Penggunaan fungisida kimia efektif
mengurangi keparahan karat sengon di tingkat
bibit namun tidak efektif untuk pengendalian
karat sengon di tegakan (Wiryadiputra, 2007).
Penggunaan fungisida kimia dapat berbahaya
bagi kesehatan manusia dan lingkungan
sehingga perlu mencari alternatif pengendalian
karat puru di tingkat bibit yang ramah
lingkungan.
Alternatif pengendalian karat puru di bibit
sengon sebaiknya menggunakan fungisida
nabati seperti penggunaan daun sirsak.
Ekstrak daun sirsak disamping potensinya
yang besar sebagai obat untuk penyakit pada
manusia, juga dilaporkan dapat mengatasi
serangan hama dan penyakit pada tanaman
(Coria-Téllez, Montalvo-Gónzalez, Yahia, &
Obledo-Vázquez, 2018). Daun sirsak (Annona

KETAHANAN BIBIT SENGON (Falcataria moluccana (Miq) Barneby & J.W. Grimes)
TERHADAP CENDAWAN Uromycladium falcatarium BERDASARKAN ASAL BENIH DAN JENIS PENGENDALI
Tati Suharti, Kurniawati Purwaka Putri, dan Yulianti Bramasto
103
muricata L.) mengandung senyawa aktif
acetogenin yang dapat menghambat dan
membunuh sel kanker (Moghadamtousi,
Fadaeinasab, Nikzad, Mohan, Ali, & Kadir,
2015). Acetogenin yang terkandung dalam
sirsak dapat juga digunakan sebagai pestisida
(Shibula & Velavan, 2015). Bahan-bahan
pengendali lainnya yang ramah lingkungan
seperti Plant Growth Promoting Rhizobacter
(PGPR), dan biofungisida baik yang berasal
dari tanaman maupun mikroorganisme. Hasil
penelitian sebelumnya menunjukkan bahwa
PGPR dan biofungisida mampu menghambat
perkembangan spora U. tepperianum (Putri &
Bramasto, 2017). Pengendalian penyakit karat
puru dapat juga melalui pemilihan asal benih
yang sehat yaitu tegakan sengon yang tidak
terkena karat puru. Tujuan penelitian ini
adalah untuk mengetahui pengaruh asal benih
dan jenis pengendali terhadap ketahanan bibit
sengon dari serangan cendawan
Uromycladium falcatarium di persemaian.
II. BAHAN DAN METODE
A. Bahan dan Alat
Bahan yang digunakan adalah benih
sengon asal dari Kabupaten Kediri (Jawa
Timur) yang merupakan sumber benih
endemik karat puru dan asal Kecamatan
Sindangbarang Kabupaten Cianjur (Jawa
Barat) yang diambil dari tegakan yang sehat
dan bebas karat puru. Bahan lainnya adalah
biofungisida yang mengandung Cryptococcus
terreus, C. Albidus dan Candida edax; PGPR
yang mengandung Rhizobium sp. Bacillus
polymixa dan Pseudomonas flourescens;
fungisida kimia (mankozeb); larutan ekstrak
daun sirsak; haemacytometer untuk
menghitung teliospora; larutan tween;
suspensi teliospora karat puru untuk inokulasi
ke bibit serta tanah, pasir, sekam padi, arang
sekam padi dan kompos sebagai bahan media
tabur dan sapih. Peralatan yang digunakan
diantaranya mikroskop, laminar airflow,
autoclave, oven, caliper, polybag, kamera.
B. Prosedur Penelitian
Benih sengon direndam terlebih dahulu
dalam air mendidih dan dibiarkan sampai
dingin selama 24 jam sebelum ditabur di atas
media tabur (Nusantara, 2002). Media tabur
yang digunakan adalah campuran tanah dan
pasir (1:1,v/v) steril. Bibit siap sapih setelah
± 3- 4 minggu dari penaburan atau setelah
muncul sepasang daun (kecambah normal).
Media sapih yang digunakan adalah campuran
tanah dengan sekam padi, arang sekam padi
dan kompos (3:1:1:1,v/v).
Bibit sengon yang segar dan sehat
diinfeksi teliospora karat puru (U. falcatarium)
pada umur ± 8 minggu dari penyapihan. Proses
inokulasi teliospora dengan cara
menyemprotkan suspensi teliospora sebanyak
± 1 ml per bibit diatas permukaan daun secara
merata dan dibiarkan selama 1 minggu.
Suspensi teliospora cendawan U. falcatarium
diperoleh dengan cara mencampurkan

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019: 101-111
p-ISSN : 2354-8568
e-ISSN : 2527-6565
104
teliospora ke dalam 200 ml aquadest yang
ditambahkan dengan 4 ml larutan tween 20
(konsentrasi 2 persen). Dalam ± 1 ml suspensi
U. falcatarium mengandung 1,18 x 107
teliospora. Menurut (Putri, Baskorowati,
Nurhidayati, Herawan & Nurrohmah, 2016),
inokulasi spora karat puru pada bibit sengon
menggunakan konsentrasi minimal 1x105
teliospora.
Pemberian perlakuan pengendali penyakit
setelah 1 minggu dari infeksi teliospora
cendawan dengan cara penyiraman yang
dilakukan setiap 2 minggu yaitu pada saat bibit
berumur 9, 11, 13, 15 dan 17 minggu. Larutan
pengendali penyakit karat puru yang
digunakan adalah larutan PGPR (5 g.l-1);
biofungisida (5 g.l-1); ekstrak daun sirsak (10
g.l-1) dan mankozeb (2 g.l-1). Konsentrasi yang
digunakan untuk PGPR, bofungsida dan
mankozeb adalah sesuai dengan petunjuk
penggunaan yang terdapat di kemasan
formulasi sedangkan untuk konsentrasi sirsak
berdasarkan Zulkipli, Marsuni dan Rosa
(2018).
Rancangan penelitian yang digunakan
adalah Rancangan Faktorial dalam acak
lengkap, terdiri dari 2 faktor, yaitu faktor asal
benih dan faktor jenis pengendali. Faktor asal
benih terdiri dari 2 taraf, yaitu:
A1 = Benih asal daerah endemik karat
puru/Kediri
A2 = Benih asal daerah bukan endemik karat
puru/Cianjur
Faktor aplikasi pengendali terdiri dari 5 taraf
yaitu: B1 = Kontrol
B2 = Pupuk hayati berupa PGPR (5 g.l-1)
B3 = Biofungisida (5 g.l-1)
B4 = Biofungisida nabati berupa ekstrak
daun sirsak (10 g.l-1)
B5 = Mankozeb (2 g.l-1)
Setiap kombinasi perlakuan terdiri dari 10
bibit diulang 4 kali.
Variabel pengamatan adalah insidensi dan
intensitas penyakit serta jumlah teliospora
yang diamati secara periodik setiap 1 minggu
setelah dilakukan penyemprotan larutan
pengendali yaitu pada saat bibit berumur
10,12, 14, 16 dan 18 minggu. Selain itu pada
akhir penelitian diamati ketebalan dinding sel
epidermis. Pengamatan intensitas penyakit
dengan melakukan skoring/penilaian (Tabel
1).
Tabel (Table) 1. Klasifikasi tingkat keparahan penyakit (Classification level of disease severity)
Tingkat keparahan (severity level) Intensitas penyakit (disease intensity) Nilai
(score)
Sehat (healthy) Tidak ada penyakit (0%) 0
Ringan (light) Intensitas penyakit (≤ 10 %) 1
Agak berat (bit heavy) Intensitas penyakit 10% < X ≤ 25% 2
Berat (heavy) Intensitas penyakit 25% < X ≤ 45% 3
Sangat berat (very heavy) Intensitas penyakit >45% < X ≤ 75% 4
Gagal (failed) Intensitas penyakit X > 75 % 5
Sumber (Source) : Asmaliyah, Lukman, & Mindawati (2016)

KETAHANAN BIBIT SENGON (Falcataria moluccana (Miq) Barneby & J.W. Grimes)
TERHADAP CENDAWAN Uromycladium falcatarium BERDASARKAN ASAL BENIH DAN JENIS PENGENDALI
Tati Suharti, Kurniawati Purwaka Putri, dan Yulianti Bramasto
105
Berdasarkan hasil pengamatan dan
skoring penyakit pada bibit sengon, dihitung
insidensi penyakit dan intensitas penyakit
dengan menggunakan rumus berikut ini
(Lestari, Rahayu, Wdiyatno, 2013) :
Jumlah tanaman yang terkena penyakitInsidensi penyakit= X 100%
jumlah keseluruhan tanaman
...(1)
( )Intensitas penyakit= %
X 100
Ni x Vj
Z x N
∑ ……......... (2)
Keterangan:
Ni = Jumlah tanaman yang terkena
penyakit dengan klasifikasi tertentu
Vj = Nilai untuk klasifikasi tertentu
Z = Nilai tertinggi dalam klasifikasi
N = Jumlah bibit seluruhnya dalam petak
contoh
C. Analisis Data
Data yang diperoleh dianalisis dengan
analisis sidik ragam kemudian apabila
signifikan maka akan dilanjutkan dengan
dengan uji lanjut Duncan.
III. HASIL DAN PEMBAHASAN
A. Hasil
Hasil analisis ragam menunjukkan bahwa
terjadi interaksi nyata perlakuan asal sumber
benih dan jenis larutan pengendali terhadap
jumlah teliospora setelah pengendalian ke-1
dan tebal epidermis (Tabel 2). Secara umum
rata-rata jumlah teliospora yang ditemukan
pada bibit yang benihnya asal Cianjur relatif
lebih sedikit dibanding benih asal Kediri. Bibit
yang benihnya asal Cianjur memiliki
epidermis yang lebih tebal dibandingkan bibit
yang benihnya asal Kediri.
Hasil analisis ragam menunjukkan bahwa
tidak terjadi interaksi nyata antara asal benih
dan jenis larutan pengendali terhadap insidensi
dan intensitas penyakit (Tabel 3).
Tabel (Table) 2. Rata-rata jumlah teliospora dan tebal epidermis sebagai pengaruh dari interaksi
asal benih dan jenis pengendali (The average number of teliospores and epidermis
thicknesses as an effect of seed sources and controller type treatments).
Interaksi
Jumlah teliospora setelah pengendalian ke- (number of teliospores after controlling at weeks to-)
Tebal epidermis
(epidermis thickness) 1 2 3
Kediri – Kontrol
(Kediri-control)
17,90 a 8,34 a 1,67 a 2,17 f
Kediri- PGPR
(Kediri-PGPR)
9,73 bcd 5,09 a 0,5 a 2,72 ed
Kediri- Biofungisida
(Kediri- biofungicide)
10,13 bc 5,11 a 0,5 a 2,42 ef
Kediri- Ekstrak daun sirsak
(Kediri-soursop leaf extract)
7,58 cde 4,15 a 0 a 2,74 ed
Kediri –Mankozeb
(Kediri-Mankozeb)
8,76 bcde 5,32 a 0,88 a 2,92 d
Cianjur-PGPR
(Cianjur-PGPR)
9,55 bcd 5,45 a 0,07 a 5,43 a
Cianjur- Biofungisida
(Cianjur- biofungicide)
7,29 cde 4,32 a 0 a 4,84 b
Cianjur-Ekstrak daun sirsak
(Cianjur-soursop leaf extract)
6,48 e 3,56 a 0 a 5,47 a
Cianjur-Mankozeb (Cianjur-Mankozeb)
7,15 de 4,32 a 0 a 5,84 a
Nilai F hitung (F-test) 3,85 * 2,14 ns 2,3 ns 1,80*
Keterangan (Remarks): Nilai-nilai yang diikuti oleh huruf yang sama pada kolom yang sama tidak berbeda nyata pada taraf uji 5% (Values followed
by the same letters on the same colom are not significantly different at the test level 5%); * = berbeda nyata pada taraf uji 5% (significantly different at the test level 5%); ns = tidak berbeda nyata pada taraf uji 5% (different not significant
at test level 5%)

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019: 101-111
p-ISSN : 2354-8568
e-ISSN : 2527-6565
106
Tabel (Table) 3. Rata-rata insidensi dan intensitas penyakit karat puru (The average of disease
incidence and intensity).
Interaksi (Interaction)
Insidensi penyakit setelah pengendalian pada
minggu ke- (Disease incidence after
controlling time at weeks to-)
Intensitas penyakit setelah pengendalian pada
minggu ke-(Disease intensity after contrrolling
time at weeks to-)
10 12 14 1 2 3
Kediri – Kontrol
(Kediri-control)
45,0 a 62,5 a 77,5 a 4,4 a 8,55 a 8,38 a
Kediri- PGPR (Kediri-PGPR)
42,5 a 42,5 a 40,0 a 4,13 a 3,13 a 3,63 a
Kediri- Biofungisida
(Kediri- biofungicide)
40,0 a 60,0 a 37,5 a 3,13 a 5,13 a 3,25 a
Kediri- Ekstrak daun sirsak (Kediri-soursop leaf extract)
42,5 a 42,5 a 72,5 a 3,75 a 5,0 a 7,87 a
Kediri –Mankozeb
(Kediri-Mankozeb)
20,0 a 55,0 a 32,5 a 2,5 a 4,13 a 2,75 a
Cianjur-Kontrol (Cianjur-control)
27,5 a 30,0 a 45,0 a 3,13 a 4,13 a 5,5 a
Cianjur-PGPR
(Cianjur-PGPR)
30,0 a 35,0 a 27,5 a 2,88 a 2,13 a 2,5 a
Cianjur- Biofungisida (Cianjur- biofungicide)
47,5 a 47,5 a 45,0 a 3,5 a 3,88 a 3,63 a
Cianjur-Ekstrak daun sirsak
(Cianjur-soursop leaf extract)
57,5 a 52,5 a 47,5 a 4,4 a 3,38 a 3,0 a
Cianjur-Mankozeb (Cianjur-Mankozeb)
37,5 a 37,5 a 40,0 a 2,25 a 2,0 a 2,88 a
Rata-rata (Average) 39,0 46,5 46,5 3,4 4,1 4,3
Nilai F hitung (F test) 1,55 tn 1,23 tn 2,18 tn 0,30 tn 1,20 tn 2,15 tn
Keterangan (Remarks): Nilai-nilai yang diikuti oleh huruf yang sama pada kolom yang sama tidak berbeda nyata pada taraf uji 5% (Values
followed by the same letters on the same colom are not significantly different at the test level 5%); * = berbeda nyata pada taraf uji 5% (significantly different at the test level 5%); ns = tidak berbeda nyata pada
taraf uji 5% (no significant different at test level 5%)
Hasil analisis ragam menunjukkan bahwa
asal benih berpengaruh nyata terhadap
intensitas penyakit pada pengendalian ke-2
dan ke-3, jumlah teliospora pada ketiga waktu
pengendalian dan tebal epidermis. Asal benih
tidak berpengaruh nyata terhadap insidensi
penyakit untuk ketiga waktu pengendalian dan
intensitas penyakit pada pengendalian minggu
ke-1 (Tabel 4 ).
Tabel (Table) 4. Rataan insidensi penyakit, intensitas penyakit, jumlah teliospora dan tebal sel
epidermis bibit sengon berdasarkan asal benih (The average of disease incidence
and intensity, number of teliospores and thickness of epidermis based on seeds
sources)
Respon (Response) Pengendaliaan ke-
(control to-)
Asal benih (seed sources) F-hit
(F-test) Kediri Cianjur
Insidensi penyakit
(disease incidence) (%)
1 38,0 a 40,0 a 0,12 tn
2 52,5 a 40,0 a 3,71 tn 3 52,0 a 41,0 a 3,92 tn
Intensitas penyakit
(disease intensity) (%)
1 3,6 a 3,2 a 0,23 tn
2 6,8 a 3,1 b 4,60 * 3 5,2 a 3,5 b 6,21 *
Jumlah teliospora (number of teliospores)
1 10,8 a 8,4 b 19,08 **
2 5,5 a 4,5 b 4,55 * 3 0,7 a 0,3 b 9,39 **
Tebal epidermis (thickness of epidermis) - 2,59 b 5,186 a 687,75 **
Keterangan (Remarks): Nilai-nilai yang diikuti oleh huruf yang sama pada baris yang sama tidak berbeda nyata pada selang kepercayaan 99%
(Values followed by the same letters on the same row are not significantly different); ** = berbeda nyata pada taraf uji 1%; * = berbeda nyata pada taraf uji 5 %; tn = tidak berbeda nyata pada taraf uji 5% (** = significantly different at the test
level of 1%; * = significantly different at the test level of 5%; ns= no significant different at test level of 5%)

KETAHANAN BIBIT SENGON (Falcataria moluccana (Miq) Barneby & J.W. Grimes)
TERHADAP CENDAWAN Uromycladium falcatarium BERDASARKAN ASAL BENIH DAN JENIS PENGENDALI
Tati Suharti, Kurniawati Purwaka Putri, dan Yulianti Bramasto
107
Jenis pengendali berpengaruh nyata
terhadap insidensi dan intensitas penyakit pada
waktu pengendalian ke-3 serta berpengaruh
sangat nyata terhadap jumlah teliospora pada
semua waktu pengendalian dan tebal
epidermis (Tabel 5). Hasil uji beda
menunjukkan bahwa pada pengendalian ke-3,
pemberian PGPR, biofungisida dan mankozeb
menurunkan insidensi penyakit. Sedangkan
ekstrak daun sirsak (60 persen) tidak berbeda
nyata dengan kontrol (61,3 persen).
Pengendalian pada minggu ke-14, perlakuan
PGPR, biofungisida, ekstrak daun sirsak dan
mankozeb dapat mengurangi insidensi dan
intensitas penyakit, serta meningkatkan tebal
epidermis bibit sengon. Secara umum keempat
jenis pengendali yang digunakan akan
menurunkan jumlah teliospora yang terdapat
pada bibit sengon.
Tabel (Table) 5. Rataan insidensi dan intensitas penyakit, jumlah teliospora dan tebal sel epidermis
bibit sengon berdasarkan jenis pengendali (The average of incidence and disease
intensity, number of teliospores and thickness of epidermis based on controller
types)
Respon (Response)
Pengendaliaan ke-
(Controlling to-) Jenis pengendali (Thypes of controller)
F-hit
(F-test)
1 2 3 4 5
Insidensi penyakit
(disease incidence) (%)
1 36,2 a 36,2 a 43,7 a 50,0 a 28,7 a 1,59 tn
2 46,2 a 38,7 a 53,7 a 47,5 a 46,2 a 0,59 tn
3 61,3 a 33,7 b 41,2 b 60,0 a 36,2 b 4,51*
Intensitas penyakit
(disease intensity) (%)
1 3,7 a 3,5 a 3,1 a 4,1 a 2,4 a 0,62 tn
2 10,1 a 2,6 a 4,5 a 4,2 a 3,1 a 2,38 tn
3 6,94 a 3,1 c 3,4 bc 5,4 ab 2,8 c 5,64*
Jumlah teliospora
(number of teliospores)
1 14,7 a 9,6 b 8,7 bc 7,0 c 7,9 bc 22,90 **
2 6,7 a 5,0 b 4,7 b 3,9 b 4,7 b 4,23 **
3 1,7 a 0,2 bc 0,2 bc 0,0 c 0,4 b 26,84 **
Tebal epidermis
(thickness of epidermis) - 3,25 c 4,07 a 3,63 b 4,10 a 4,38 a 16,18 **
Keterangan (Remarks): (1) kontrol; 2 PGPR; 3 Biofungisida; 4 ekstrak daun sirsak; 5 mankozeb
Jenis pengendali hanya berpengaruh nyata
terhadap insidensi dan intensitas penyakit pada
pengendalian ke-3. Setelah pengendalian ke-3,
insidensi penyakit pada bibit kontrol dan
perlakuan sirsak berbeda nyata dengan
perlakuan lainnya. Insidensi penyakit yang
paling tinggi yaitu pada kontrol (61,26 persen).
Begitu pula dengan intensitas penyakit setelah
pengendalian ke-3, bibit kontrol mempunyai
intensitas penyakit yang paling tinggi (6,94
persen). Intensitas penyakit kontrol dan
perlakuan sirsak berbeda nyata dengan
perlakuan PGPR dan mankozeb.
B. Pembahasan
Asal benih (endemik, bukan endemik) dan
jenis larutan pengendali terbukti secara
sinergis menurunkan jumlah teliospora U.
falcatarium setelah satu kali dilakukan aplikasi
pengendalian. Secara umum bibit asal benih
Cianjur yang diberi larutan pengendali
cenderung menghasilkan jumlah spora
(teliospora) yang lebih rendah dibandingkan

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019: 101-111
p-ISSN : 2354-8568
e-ISSN : 2527-6565
108
kombinasi bibit asal benih Kediri dengan
larutan pengendali. Dalam hal ini asal sumber
benih berperan dalam menghambat proses
perkembangan teliospora, sehingga menjadi
faktor penting dalam kaitannya dengan
kemampuan menahan infeksi patogen.
Sebagaimana diketahui bahwa kemampuan
tanaman untuk bertahan terhadap penyakit
akan bervariasi diantara berbagai asal sumber
benih (Rahayu et al., 2009; Nurrohmah &
Baskorowati, 2011; Baskorowati, Rohandi &
Gunawan, 2012).
Dalam penelitian ini intensitas penyakit
karat puru pada bibit sengon dari kedua lokasi
masih di bawah ≤ 10 persen, sehingga tingkat
keparahan masih ringan (Asmaliyah et al.,
2016). Namun, secara umum intensitas
penyakit pada bibit asal Kediri lebih tinggi 1,7
persen pada pengendalian ke-3 dibanding bibit
asal benih Cianjur. Perbedaan intensitas
penyakit tersebut menunjukkan bahwa kedua
asal benih mempunyai daya tahan berbeda.
Bibit dengan benih asal Kediri diduga
memiliki tingkat ketahanan terhadap infeksi
cendawan U. falcatarium yang lebih rendah
dibanding benih asal Cianjur. Nurrohmah &
Baskorowati (2011) menyatakan bahwa
perbedaan ketahanan penyakit disebabkan
adanya keragaman genetik pada tanaman
sengon yang dibawa oleh benih. Benih asal
Kediri dikumpulkan dari wilayah yang
merupakan daerah endemik karat puru,
sedangkan benih asal sumber benih Cianjur
dikumpulkan dari tegakan sengon yang berada
< 5 km dari pinggir pantai serta kondisi
tegakan sehat dan bebas dari serangan
penyakit terutama cendawan karat puru. Hasil
yang sama ditunjukkan pada bibit yang berasal
dari Wamena lebih kecil intensitas
penyakitnya dibanding asal Kediri (Rahayu et
al., 2009). Oleh karena itu salah satu
pengendalian hama dan penyakit dapat
dilakukan antara lain dengan menggunakan
benih tanaman yang tidak hanya memiliki
pertumbuhan bagus tetapi juga tahan terhadap
serangan hama dan penyakit (Nurrohmah &
Baskorowati, 2011).
Interaksi asal sumber benih dan jenis
larutan pengendali juga mempengaruhi
ketebalan sel epidermis bibit sengon.
Ketebalan dinding sel epidermis tanaman
inang menyebabkan hifa cendawan kesulitan
pada saat penetrasi sehingga menjadi salah
satu faktor teknis yang dapat mempertahankan
tanaman dari serangan cendawan. (Gil &
Chang-Duck, 2015) melaporkan bahwa
epidermis yang tebal dapat meningkatkan
ketahanan tumbuhan terhadap infeksi patogen
Uromyces truncicola pada pohon Sophora
japonica. Tanaman dapat ditingkatkan
ketahanannya antara lain melalui ketahanan
struktural yaitu penghalang yang bersifat fisik
seperti penebalan dinding sel dengan
pemberian elisitor/inducer biotik atau abiotik

KETAHANAN BIBIT SENGON (Falcataria moluccana (Miq) Barneby & J.W. Grimes)
TERHADAP CENDAWAN Uromycladium falcatarium BERDASARKAN ASAL BENIH DAN JENIS PENGENDALI
Tati Suharti, Kurniawati Purwaka Putri, dan Yulianti Bramasto
109
seperti mikroorganisme, bahan tanaman atau
senyawa kimia (Inayati, 2016). Hasil analisis
mikroskopis menunjukkan bahwa sel
epidermis pada bibit sengon Cianjur dan
Kediri yang tidak diberi larutan pengendali
relatif lebih tipis dibanding bibit yang diberi
larutan pengendali. Larutan PGPR, mankozeb,
dan ekstrak daun sirsak dapat meningkatkan
tebal sel epidermis. Shanmugaiah, Ramesh,
Jayaprakashvel, & Mathivanan (2006)
melaporkan bahwa PGPR yang berasal dari
Pseudomonas sp. dapat menginduksi
ketahanan tanaman terhadap patogen salah
satunya menyebabkan modifikasi struktur dan
biokimia/fisiologi dinding sel.
Larutan PGPR dan mankozeb dalam
penelitian ini diketahui sebagai pengendali
yang efektif mengurangi insidensi dan
intensitas penyakit karat puru pada bibit
sengon. PGPR yang digunakan mengandung
bakteri yang bersifat antagonis seperti B.
polymixa dan P. flourescens. Untuk itu PGPR
dapat menjadi salah satu alternatif
pengendalian U. falcatarium yang ramah
lingkungan. Sejalan dengan penelitian
(Pracoyo, 2013) yang melaporkan bahwa
PGPR yang mengandung B. polymixa dan P.
flourescens dapat mengurangi insidensi dan
intensitas karat puru pada tegakan sengon.
Kavitha, Senthilkumar, Gnanamanickam,
Inayathullah, & Jayakumar (2005) melaporkan
B. polymixa dapat menyebabkan perubahan
bentuk sel sehingga hifa menjadi membengkak
dan membulat. Kemampuan P. flourescens
dalam menghambat pertumbuhan cendawan
salah satunya karena menghasilkan antibiotik
yaitu phenazine 1- carboxylic acid, 2, 4-
diacetylphloroglucinol, pyoluteorin and
pyrrolnitrin (Prabhukarthikeyan &
Raguchander, 2016).
Aplikasi mankozeb, P. fluorescens dapat
meningkatkan aktivitas enzim-enzim yang
terlibat dalam ketahanan tanaman seperti
enzymes peroxidase (PO) dan polyphenol
oxidase (PPO) (Anand, Chandrasekaran,
Kuttalam, Raguchander, Prakasam, &
Samiyappan, 2007). Menurut Ghosh (2015),
peroxidase (PO) mengoksidasi fenol menjadi
quinone dan menghasilkan peroksida (H2O2)
yang berperan sebagai antimikroba,
selanjutnya terjadi polimerasi fenol menjadi
senyawa lignin yang akan tersimpan di
dinding sel dan selanjutnya berperan dalam
menghambat perkembangan patogen. Aktivitas
PPO yang meningkat juga akan menghasilkan
senyawa-senyawa beracun sehingga
ketahanan inang terhadap infeksi patogen
meningkat.
Perkembangan jumlah teliospora
cenderung menurun dengan semakin seringnya
pemberian larutan pengendali. Namun
efektivitasnya terhadap insidensi dan intensitas
penyakitnya baru menampakkan perbedaan
dengan kontrol setelah pengendalian ke-3.
Menurut Widyastuti, Harjono, & Surya (2013),
kegagalan teliospora dalam proses infeksi

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019: 101-111
p-ISSN : 2354-8568
e-ISSN : 2527-6565
110
disebabkan oleh adanya senyawa penghambat
salah satunya senyawa fenol.
IV. KESIMPULAN
Insidensi dan intensitas penyakit karat
puru lebih tinggi pada bibit asal Kediri
dibandingkan asal Cianjur berkaitan dengan
kondisi sel epidermis bibit Cianjur yang lebih
tebal dibandingkan Kediri. Pengendalian
penyakit karat puru pada tingkat bibit dapat
dilakukan dengan pemberian larutan PGPR (5
g.l-1), mankozeb (2 g.l-1), atau biofungisida (5
g.l-1). Pengendalian terbaik yang mampu
menurunkan insidensi, intensitas karat puru
dan menghambat perkembangan teliospora
yang ramah lingkungan adalah pemberian
larutan PGPR.
UCAPAN TERIMA KASIH
Penulis mengucapkan terimakasih yang
sebesar-besarnya kepada Dr. Ratih Damayanti,
S.Hut., M.Si peneliti bidang anatomi kayu di
Puslitbang Hasil Hutan atas bimbingan dan
arahan yang diberikan sehingga pengamatan
sel epidermis bibit dapat terlaksana. Ucapan
terima kasih juga disampaikan kepada Ibu
Dina Agustina, S.Si, Bapak Emuy, Bapak
Suherman dan Ibu Wildani Asfari Hanifah atas
kerjasamanya sehingga penelitian ini dapat
dilaksanakan dan diselesaikan.
DAFTAR PUSTAKA
Anand, T., Chandrasekaran, A., Kuttalam, S.,
Raguchander, T., Prakasam, V., &
Samiyappan, R. (2007). Association of some
plant defense enzyme activities with systemic
resistance to early leaf blight and leaf spot
induced in tomato plants by azoxystrobin and
Pseudomonas fluorescens. Journal of Plant
Interactions, 2(4), 233–244.
Anggraeni, I. (2018). Penyakit karat tumor pada
sengon (Paraserianthes falcataria (L)
Nielsen) DI perkebunan Glenmore
Banyuwangi, Jawa Timur. Jurnal Penelitian
Hutan Tanaman, 6(5), 311–321.
Asmaliyah, A., Lukman, A. H., & Mindawati, N.
(2016). Perkembangan serangan hama dan
penyakit pada tanaman bambang lanang pada
berbagai sistem persiapan lahan. Jurnal
Penelitian Hutan Tanaman, 13(2), 139–155.
Baskorowati, L., Rohandi, A., & Gunawan.
(2012). Proceeding of International
Conference on The Impact of Climate
Change to Forest Pests and Diseases in The
Tropics. In Response of young Falcataria
moluccana to gall rust. (pp. 162-168).
Coria-Téllez, A. V., Montalvo-Gónzalez, E.,
Yahia, E. M., & Obledo-Vazquez, E. N.
(2018). Annona muricata: A comprehensive
review on its traditional medicinal uses,
phytochemicals, pharmacological activities,
mechanisms of action and toxicity. Arabian
Journal of Chemistry, 11(5), 662–691.
Corryanti, & Novitasari. (2015). Sengon dan
Penyakit Karat Tumor. Puslitbang Perum
Perhutani, Cepu. 29 p.
Ghosh, R. ( 2015). Enzymatic responses of ginger
plants to Pythium infection after sar
induction. Journal of Plant Pathology and
Microbology, 6(7).
Gill, Hee-Young, & Chang-Duck K. (2015).
Morphological characteristic of the rust fungi,
Uromyces truncicola, and histological
changes in the infected host tree, Sophora
japonica. Journal of Korean Forest Society,
99(3), 277-284.
Inayati, A. (2016). Ketahanan terimbas tanaman
kacang-kacangan. Iptek Tanaman Pangan
11(2), 175–186.
Kavitha, S., Senthilkumar, S., Gnanamanickam,
S., Inayathullah, M., & Jayakumar, R. (2005).
Isolation and partial characterization of

KETAHANAN BIBIT SENGON (Falcataria moluccana (Miq) Barneby & J.W. Grimes)
TERHADAP CENDAWAN Uromycladium falcatarium BERDASARKAN ASAL BENIH DAN JENIS PENGENDALI
Tati Suharti, Kurniawati Purwaka Putri, dan Yulianti Bramasto
111
antifungal protein from Bacillus polymyxa
strain VLB16. Process Biochemistry, 40(10),
3236-3246.
Krisnawati, H., Varis, E., Kallio, M. dan
Kanninen, M. (2011). Paraserienthes
falcataria (L.) Nielsen: Ekologi, Silvikultur
\dan Produktivitas. CIFOR, Bogor,
Indonesia.
Lestari, P., Rahayu, S., & Widiyatno. (2013).
Dynamics of gall rust disease on sengon
(Falcataria moluccana) in various
agroforestry patterns. Procedia
Environmental Sciences, 17,167 – 171.
Moghadamtousi, S. Z., Fadaeinasab, M., Nikzad,
S., Mohan, G., Ali, H. M., & Kadir, H. A.
(2015). Annona muricata (Annonaceae): A
review of its traditional uses, isolated
acetogenins and biological activities.
International Journal of Molecular Sciences,
16(7), 15625–15658.
Nurrohmah, S. H., & Baskorowati, L. (2011).
Prosiding Seminar Nasional Pendidikan
Biologi dan Saintek II 49. Serangan awal
penyakit karat tumor pada tanaman sengon
di plot uji provenan sengon Candiroto, Jawa
Tengah. (pp 48-61).
Nusantara, A. D. (2002). Tanggap semai sengon
(Paraserianthes falcataria L. Nielsen)
terhadap inokulasi ganda cendawan mikoriza
arbuskular dan Rhizobium sp. Jipi, 4(2), 62–
70.
Prabhukarthikeyan S.R., & Raguchander, T.
(2016). Antifungal metabolites of
Pseudomonas fluorescens against Pythium
aphanidermatum. Journal of Pure and
Applied Microbiology, 10(1), 579-584.
Pracoyo, A. (2013). Pengaruh plant growth
promoting rhizobacter (PGPR) dan pupuk
mikro terhadap penyakt karat puru dan
pertumbuhan tanaman sengon
(Paraserianthes falcataria) di lapangan.
Skripsi. Departemen Proteksi Tanaman.
Fakultas Pertanian. Bogor: Institut Pertanian
Bogor.
Putri, A.I., Baskorowati, L., Nurhidayati,
Herawan, T., & Nurrohmah, S. T. (2016).
Proceedings of the International Conference
of Indonesia Forestry Researchers III. Effect
filter cover of seedling in direct inoculation
screening of Uromycladium tepperianum for
Falcataria moluccana disease tolerant. (pp
11-18).
Putri, A.I. (2018). Microscopic callus selection of
sengon tree (Falcataria moluccana) putative
tolerant to Uromycladium falcatarium). IOP
Conf. Ser.: Earth Environ. Sci. 183 012006. 6
p.
Putri, K. P., & Bramasto, Y. (2017). Pengendalian
cendawan Uromycladium tepperianum pada
bibit sengon (Falcataria moluccana (Miq.)
Barneby & J. W. Grimes ). JPTH, 5(1):13-22.
Rahayu, S., Nor, N. A., See, L. S., & Saleh, G.
(2009). Responses of Falcataria moluccana
seedlings of different seed sources to
inoculation with Uromycladium tepperianum.
Silvae Genetica, 58(1–2), 62–68.
Shanmugaiah, V., Ramesh, S., Jayaprakashvel,
M., & Mathivanan, N. (2006). Proceedings of
the 1st International Symposium on
Biological Control of Bacterial Plant
Diseases. In Biocontrol and plant growth
promoting potential of Pseudomonas sp.
MML2212 from the rice rhizosphere.
(pp.320-324).
Shibula, K., & Velavan, S. (2015). Determination
of phytocomponents in methanolic extract of
Annona muricata leaf using GC-MS
technique. International Journal of
Pharmacognosy and Phytochemical
Research, 7(6), 1251–1255.
Wiryadiputra, S. (2007). Epidemi penyakit tumor
pada sengon (Paraserianthes falcataria) di
Jawa Timur, Indonesia. Jurnal Ilmu
Kehutanan 1(1): 31-39.
Widyastuti, S. M., Harjono, H., & Surya, Z. A.
(2013). Initial infection of Falcataria
moluccana leaves and Acacia mangium
phyllodes by Uromycladium tepperianum
fungi in a laboratory trial. Jurnal Manajemen
Hutan Tropika, 19(3), 187–193.
Zulkipli, S., Marsuni, Y., & Rosa, H. O. (2018).
Uji lapangan beberapa pestisida nabati untuk
menekan perkembangan penyakit antraknosa
pada tanaman cabai besar. Proteksi Tanaman
Tropika 1(2): 31-34.

TEKNIK PENYIMPANAN BENIH MERANTI BALAU (Shorea seminis (de Vriese) Sloot)
Muhammad Zanzibar, Naning Yuniarti, dan Ratna Uli Damayanti S
Kontribusi penulis: Muhammad Zanzibar sebagai kontributor utama
© 2019 BPTPTH All rights reserved. Open access under CC BY-NC-SA license.doi: //doi.org/10.20886/bptpth.2019.7.2.113-125 113
TEKNIK PENYIMPANAN BENIH MERANTI BALAU (Shorea seminis (de Vriese) Sloot)
(The Technique of Storage of Meranti Balau (Shorea seminis (de Vriese) Sloot))
*Muhammad Zanzibar, Naning Yuniarti, dan/and Ratna Uli Damayanti S
Balai Penelitian dan Pengembangan Teknologi Perbenihan Tanaman Hutan
Jl. Pakuan Ciheuleut PO Box 105; Telp. (0251) 8327768, Bogor, Jawa Barat, Indonesia
e-mail: [email protected]
Naskah masuk: 25 Januari 2019; Naskah direvisi:30 Juli 2019; Naskah diterima: 29 November 2019
ABSTRACT
The problem faced by meranti balau (Shorea seminis (de Vriese) Sloot) seed is the recalcitrant seed
character so that the viability of the seeds rapidly decreases during the storage. The purpose of this research
was to determine the best storage technique for meranti balau seed. This research was used factorial
randomized complete design with three factors, namely the storage container factor (calico cloth and calico
cloth in a wooden box), the storage room factor (ambient and AC), and the storage period factor (0 day, 3
days, 6 days, 9 days, 12 days, 15 days and 18 days). Each treatment consisted of 25 seeds, repeated 4 (four)
times. The observed responses in this research were moisture content, germination percentage, and
germination value. The results showed that the container, the storage room, and period of storage influenced
the seed moisture content, germination percentage and germination value. The best storage technique is to
use a container of calico cloth in a wooden box inserted into the ambient room condition. The interaction of
the container of calico cloth in a wooden box with the ambient room can reduce the rate of meranti balau
seed deterioration during storage.
Keywords: Shorea seminis (de Vriese) Sloot), storage, seed viability
ABSTRAK
Permasalahan yang dihadapi dari benih meranti balau (Shorea seminis (de Vriese) Sloot) adalah benihnya
berwatak rekalsitran sehingga viabilitasnya cepat menurun selama penyimpanan. Tujuan penelitian ini
adalah untuk mengetahui teknik penyimpanan terbaik benih meranti balau. Penelitian menggunakan
rancangan acak lengkap (RAL) pola faktorial dengan tiga faktor, yaitu faktor wadah simpan (kain belacu dan
kain belacu dalam kotak kayu), faktor ruang simpan (kamar dan AC), dan faktor periode simpan (0 hari, 3
hari, 6 hari, 9 hari, 12 hari, 15 hari dan 18 hari). Ulangan sebanyak 4 (empat) kali, @ 25 butir benih.
Parameter yang diamati adalah kadar air, daya berkecambah, dan nilai perkecambahan. Hasil penelitian
menunjukkan bahwa wadah, ruang, dan periode simpan berpengaruh terhadap nilai kadar air, daya
berkecambah dan nilai perkecambahan. Teknik penyimpanan terbaik untuk benih meranti balau adalah
menggunakan wadah kain blacu dalam kotak kayu, dimasukkan ke dalam ruang suhu kamar. Interaksi
penggunaan wadah kain belacu dalam kotak kayu dengan ruang suhu kamar dapat menekan laju penurunan
viabilitas benih meranti balau selama penyimpanan. Kata kunci :Shorea seminis (de Vriese) Sloot), penyimpanan, viabilitas benih
I. PENDAHULUAN
Meranti balau (Shorea seminis (de Vriese)
Sloot) termasuk ke dalam famili
Dipterocarpaceae. Jenis ini sebaran alaminya
terdapat di Kalimantan, Sarawak, Sabah,
Brunei, dan Filipina. Tumbuh berkelompok
sepanjang daerah aliran sungai (Maharani,
Handayani, & Hardjana, 2013). Meranti balau
merupakan salah satu jenis meranti penghasil
tengkawang (Saridan, Fernandes, & Noor,
2013) Tengkawang merupakan marga dari
meranti (Shorea) yang bijinya dapat dipakai
sebagai sumber penghasil minyak nabati.
Kayunya mempunyai nilai ekonomi tinggi dan
sangat baik untuk mebel, panel, lantai, langit-
langit dan juga untuk kayu lapis (Puspitasari,

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019: 113-125
p-ISSN : 2354-8568
e-ISSN : 2527-6565
114
2011). Menurut Muslich dan Sumarni (2018)
kayu meranti balau mempunyai berat jenis 0,9
g.cm-3, kelas ketahanan IV dan kelas awet I-II.
Dilihat dari potensi yang dimiliki, baik
dari kegunaan kayunya, manfaat bijinya
maupun dari nilai ekonominya, maka meranti
balau sangat baik untuk dikembangkan.
Keberhasilan program penanaman dipengaruhi
oleh banyak faktor. Salah satu di antaranya
adalah pengadaan benih berkualitas yang
didukung penguasaan teknologi penanganan
benih secara tepat. Teknik penanganan benih
adalah semua tahap kegiatan penanganan
mulai dari benih dipanen sampai dengan
penyimpanan benih (Suita, 2013; Yuniarti,
Syamsuwida, & Aminah, 2013). Untuk
berhasilnya usaha penyimpanan benih, maka
watak dari benih harus diketahui. Berdasarkan
watak/karakteristiknya, benih dikelompokkan
menjadi 3, yaitu rekalsitran, ortodok dan
intermediate. Dilihat dari permasalahan yang
serius mengenai penyimpanannya, maka benih
rekalsitran yang paling bermasalah, karena
benih rekalsitran akan mengalami kemunduran
benih yang cepat selama penyimpanan
(Aminah, 2011). Salah satu famili yang
memiliki sifat rekalsitran adalah famili
Dipterocarpaceae (Schmidt, 2000), contohnya
benih meranti balau. Jadi permasalahan yang
dihadapi dari benih meranti balau adalah
benihnya berwatak rekalsitran sehingga
viabilitas benihnya cepat menurun selama
proses penyimpanan. Untuk menjamin
ketersediaan benih bermutu sampai waktu
penanaman, maka diperlukan teknik
penyimpanan secara tepat. Tujuan
penyimpanan benih adalah diperolehnya
ketersediaan benih yang berdaya hidup tinggi
dalam jangka waktu tertentu hingga saatnya
diperlukan untuk penanaman.
Penurunan mutu benih dapat diperlambat
melalui metode penyimpanan yang tepat
(Suita, 2013). Dalam penentuan metode
penyimpanan yang menjadi pertimbangan
utama adalah daya simpan benih (Yuniarti et
al., 2013). Teknik penyimpanan yang kurang
tepat akan mempercepat penurunan viabilitas
benih (Zanzibar & Widodo, 2011).
Pengemasan benih rekalsitran untuk
penyimpanan adalah menggunakan wadah
yang bersifat tidak kedap terhadap uap air dan
gas tetapi cukup dapat mempertahankan
kelembapan, misalnya kantong katun, karung
goni, kantong kertas, kantong plastik
berlubang, kotak kardus dan kotak kayu.
Bahan pencampur selama penyimpanan
tersebut dapat menggunakan serbuk gergaji
lembap, serbuk sabut kelapa lembap, batu
perlite dan bahan lainnya. Selain itu, benih
rekalsitran memerlukan penyimpanan yang
cukup lembap dan sejuk, dikombinasikan
dengan aerasi yang baik (pertukaran udara)
serta diupayakan tidak terjadi pemanasan yang
berlebihan akibat kelembapan benih dan

TEKNIK PENYIMPANAN BENIH MERANTI BALAU (Shorea seminis (de Vriese) Sloot)
Muhammad Zanzibar, Naning Yuniarti, dan Ratna Uli Damayanti S
115
respirasi. Ruang simpan yang dapat digunakan
adalah ruang simpan suhu kamar (suhu
27ºC―30ºC dan kelembapan nisbi udara 70
persen―80 persen) dan kering sejuk/ AC
(suhu 18ºC―20ºC, kelembaban nisbi 50
persen―60 persen) (Schmidt, 2000).
Tujuan penelitian ini adalah untuk
mengetahui teknik penyimpanan yang terbaik
untuk benih meranti balau (Shorea seminis (de
vriese) Sloot). Diharapkan dengan teknik
penggunaan kombinasi wadah dan ruang
simpan yang tepat dapat mempertahankan
viabilitas benih dengan waktu/periode simpan
yang lebih panjang.
II. BAHAN DAN METODE
A. Bahan dan Alat
Pengunduhan buah dilakukan di KHDTK
Haurbentes-Jasinga, Kabupaten Bogor, Jawa
Barat pada bulan Maret 2012. Pengujian benih
dan perkecambahan dilakukan di laboratorium
dan rumah kaca Fakultas Kehutanan IPB di
Darmaga. Pelaksanaan penelitian dilakukan
selama 3 (tiga) bulan, yaitu bulan Maret―Mei
2012.
Bahan dan alat yang digunakan dalam
penelitian ini adalah benih meranti balau,
media tanah dan pasir, bak kecambah, kotak
kayu, karung goni, kain belacu, oven, cawan
petri, timbangan analitik, desikator, cawan
porselen, ruang simpan suhu kamar, dan ruang
simpan suhu AC.
B. Prosedur Penelitian
1. Pengunduhan dan Ekstraksi
Buah diperoleh dengan cara memanjat
pohon induk dan dikumpulkan dari lantai
hutan. Sebelum pengumpulan buah, terpal
dibentangkan pada lantai hutan dan buah
dikumpulkan pada wadah karung goni. Buah
yang digunakan dalam penelitian ini adalah
buah yang sudah masak fisiologis yang
dicirikan dengan sayap berwarna cokelat.
Ekstraksi benih dilakukan dengan cara
memotong sayap hingga pangkal.
2. Perlakuan Benih
Benih dikemas dimasukkan ke dalam kain
belacu dan kain belacu dalam kotak kayu.
Kotak kayu dibuat dari kayu sengon,
berukuran 30 cm (panjang) x 20 cm (lebar) x
20 cm (tinggi), antar penyekat pada ke empat
sisinya dibuat renggang sebesar 2 cm sehingga
memungkinkan terjadinya pertukaran udara
dari dalam/keluar wadah kayu, atau ke dalam
benih. Wadah berisi benih kemudian
ditempatkan ke dalam ruang simpan suhu
kamar (suhu = 25ºC ―27ºC, kelembapan = 65
persen ―70 persen ), dan ruang ber-AC (suhu
= 18ºC ―22ºC, kelembapan = 70 persen ― 80
persen). Lama penyimpanan terdiri dari 0, 3, 6,
9, 12, 15 dan 18 hari. Metode pengujian kadar
air benih menggunakan suhu 105ºC, selama 18
jam (ISTA, 2010). Penaburan benih pada bak
kecambah dilakukan di rumah kaca dengan
menggunakan campuran media pasir dan tanah
yang telah disterilisasi (1 : 1, v/v).
Rancangan percobaan yang digunakan
dalam penelitian ini adalah rancangan acak

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019: 113-125
p-ISSN : 2354-8568
e-ISSN : 2527-6565
116
lengkap (RAL) pola faktorial dengan tiga
faktor. Faktor pertama adalah wadah simpan
(A), terdiri dari 2 perlakuan, yaitu: kain belacu
(A1) dan kain belacu dalam kotak kayu (A2).
Faktor kedua adalah ruang simpan (B), yang
terdiri dari 2 perlakuan, yaitu: ruang simpan
suhu kamar (B1) dan ruang simpan suhu AC
(B2). Sedangkan faktor ketiga adalah periode
simpan (C), terdiri dari 6 perlakuan, yaitu : 0
hari (C0), 3 hari (C1), 6 hari (C2), 9 hari, 12
hari (C3), 15 hari (C4) dan 18 hari (C5).
Ulangan sebanyak 4 (empat) kali, masing-
masing ulangan terdiri dari 25 butir benih.
Untuk perlakuan kontrol (0 hari), benih
langsung diukur kadar airnya dan benih
langsung dikecambahkan. Parameter yang
diamati adalah kadar air, daya berkecambah,
dan nilai perkecambahan. Rumus dari masing-
masing parameter adalah sebagai berikut :
a. Kadar air benih
Kadar air benih adalah banyaknya
kandungan air dalam benih yang diukur
berdasarkan hilangnya kandungan air tersebut
dan dinyatakan dalam persen (%) terhadap
berat asal contoh (ISTA, 2010).
(%) 100%b c
KA a Xb
−= − ..............................(1)
Keterangan:
KA = kadar air
a = berat wadah + tutup
b = berat wadah + tutup + berat contoh
awal
c = berat wadah + tutup + berat contoh
setelah pengeringan
b. Daya berkecambah
Daya berkecambah adalah kemampuan benih
untuk tumbuh dan berkembang menjadi
kecambah normal (ISTA, 2010). Kriteria
kecambah normal adalah telah tumbuh/muncul
sepasang daun.
(%) 100%KN
DB XJB
= .....................................(2)
Keterangan:
DB = daya berkecambah
KN = jumlah kecambah normal
JB = jumlah benih yang ditabur
c. Nilai perkecambahan
Nilai perkecambahan merupakan jumlah
kecambah yang diperkirakan akan diperoleh
dari penaburan benih di persemaian
(Djavanshir & Pourbeik, 1976).
( 10)KBH
NK PK XF
=∑ ……….................….(3)
Keterangan:
NK = Nilai Perkecambahan
= Kecepatan berkecambah harian
F = Frekuensi (jumlah kecepatan
berkecambah harian yang
dihitung selama pengujian)
PK = Proses berkecambah pada saat
pengujian
10 = Konstanta
C. Analisis Data
Data-data hasil penelitian (kadar air,
daya berkecambah, nilai perkecambahan)
dianalisis secara statistik dengan menggunakan
rancangan acak lengkap pola faktorial untuk
mendapatkan analisis ragam (ANOVA).
Analisis menggunakan software program
Statistical Analysis System (SAS) versi 9.1.3
TEKNIK PENYIMPANAN BENIH MERANTI BALAU (Shorea seminis (de Vriese) Sloot)
Muhammad Zanzibar, Naning Yuniarti, dan Ratna Uli Damayanti S
117
(SAS Institute, 2004). Apabila berpengaruh
nyata maka untuk mengetahui perbedaan
perlakuan dan interaksi lebih lanjut dilakukan
uji Duncan pada selang kepercayaan 95
persen.
III. HASIL DAN PEMBAHASAN
A. Hasil
1. Kadar air
Analisis ragam menunjukkan bahwa
perlakuan faktor utama periode simpan, ruang
simpan dan waktu simpan berpengaruh nyata
terhadap kadar air benih meranti balau.
Interaksi periode dan ruang, periode dan
wadah, ruang dan wadah serta interaksi
periode, ruang dan wadah juga memberikan
pengaruh yang nyata terhadap kadar air benih
meranti balau (Tabel 1). Untuk mengetahui
lebih lanjut perlakuan yang menimbulkan
perbedaan yang nyata, dilakukan uji beda
Duncan (Tabel 2).
Tabel (Table)1. Analisis ragam pengaruh wadah, ruang dan periode simpan terhadap nilai kadar air
benih meranti balau (Analysis of variance for the effect of the container, storage
room, and storage period on the moisture content of meranti balau seeds)
Sumber keragaman
(Source of variation)
Derajat
bebas
(Degree of
freedom)
Jumlah
kuadrat
(Sum of
square)
Kuadrat
tengah
(Mean of
square)
F hitung
(F-calculation)
F tabel (F-table)
5%
Wadah (container)/A
Ruang (room)/B
Interaksi (interaction)/AB
Periode (period)/C
Interaksi (interaction)/AC)
Interaksi(interaction)/BC
Interaksi(interaction)/ABC
Sisa (residual)
1
1
1
6
6
6
6
56
2399,80
1779,65
397,46
7214,42
834,02
675,12
256,32
10,77
1202,40
1779,65
397,46
1202,40
139,00
112,52
42,72
0,19
6251,63*
9252,88*
2066,50*
6251,63*
722,72*
585,03*
222,11*
2,27
4,01
4,01
2,27
2,27
2,27
2,27
Total (total) 83 13567.56
Keterangan (Remarks): * = berpengaruh nyata pada tingkat kepercayaan 95% (Significant at 95 percent confidence
level)
Tabel (Table) 2. Pengaruh wadah, ruang dan periode simpan terhadap kadar air benih meranti balau
(The effect of the container, the storage room, and the storage period on the
moisture content of meranti balau seed)
Periode simpan
(Period of storage)
Wadah (Container)
Kain belacu (Calico cloth)
Kain belacu dalam kotak kayu
(The calico cloth in a wooden box)
Ruang simpan (Room of storage) Ruang simpan (Room of storage)
AC (AC) Kamar (Ambient) AC (AC) Kamar (Ambient)
0 hari (day) 55,20 a 55,20 a 55,20 a 55,20 a
3 hari (days) 46,69 f 49,47 d 50,89 c 52,88 b
6 hari (days) 31,68 m 45,47 gh 48,66 e 52,62 b
9 hari (days) 27,58 n 44,17 i 46,02 fg 52,42 b
12 hari (days) 19,14 p 40,05 k 44,17 i 45,04 h
15 hari (days) 14,17 q 39,60 k 32,58 l 42,16 j
18 hari (days) 8,76 r 24,17 o 30,98 m 42,17 j
Keterangan (Remarks) : Angka-angka yang diikuti oleh huruf yang sama menunjukkan tidak adanya
perbedaan nyata pada tingkat kepercayaan 95% (Values followed by the same
letter are not significantly different at 95% confidence level)
Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019: 113-125
p-ISSN : 2354-8568
e-ISSN : 2527-6565
118
2. Daya berkecambah
Hasil analisis ragam menunjukkan bahwa
perlakuan faktor tunggal wadah, ruang dan
periode simpan, interaksi wadah dan ruang
simpan, interaksi ruang dan periode simpan,
serta interaksi wadah dan periode simpan,
berpengaruh nyata terhadap nilai daya
berkecambah benih meranti balau. Sedangkan
interaksi wadah, ruang dan periode simpan
tidak berpengaruh nyata (Tabel 3). Untuk
mengetahui lebih lanjut perlakuan yang
menimbulkan perbedaan yang nyata,
dilakukan uji beda Duncan, yang disajikan
pada Tabel 4, Tabel 5, dan Tabel 6.
Tabel (Table) 3. Analisis ragam pengaruh wadah, ruang dan periode simpan terhadap daya
berkecambah benih meranti balau (Analysis of variance for the effect of the
container, storage room, and storage period on the germination percentage of
meranti balau seed)
Sumber keragaman
(Source of variation)
Derajat
bebas
(Degree of
freedom)
Jumlah
kuadrat
(Sum of
square)
Kuadrat
tengah
(Mean of
square)
F hitung (F-
calculation)
F tabel
(F-table)
5%
Wadah (container)/A
Ruang (room)/ B
Interaksi A dan B (interaction A and B)
Periode (period)/C
Interaksi A dan C (interaction A and C)
Interaksi B dan C (interaction B and C)
Interaksi A, B dan C (interaction A, B and C)
Sisa (residual)
1
1
1
6
6
6
6
56
11386,1
2368,05
1491,86
52361,81
4189,62
1085,62
660,4
4076.67
8726,97
2368,05
1491,86
8726,97
698,27
180,94
110,08
72.80
119,88*
32,53*
20,49*
119,88*
9,59*
2,49*
1,51tn
2,27
4,01
4,01
2,27
2,27
2,27
2,27
Total (total) 83 77620.81 Keterangan (Remarks): * = berpengaruh nyata pada tingkat kepercayaan 95% (Significant at 95% confidence level), tn
= tidak berpengaruh nyata pada tingkat kepercayaan 95% (Not significant at 95% confidence
level)
Tabel (Table) 4. Pengaruh interaksi ruang simpan dan periode simpan terhadap daya berkecambah
benih meranti balau (The effect of storage room interaction and storage period on
the germination percentage of meranti balau seed) (Uji Duncan)
Periode simpan
(Period of storage)
Ruang simpan (Room of storage)
AC (AC) Kamar (Ambient)
0 hari (day) 96,00a 96,00a
3 hari (days) 83,33b 86,00ab
6 hari (days) 72,00c 80,00bc
9 hari (days) 46,00e 57,33d
12 hari (days) 34,67fg 56,00d
15 hari (days) 24,33gh 42,67ef
18 hari (days) 18,67h 31,33g Keterangan (Remarks) : Angka-angka yang diikuti oleh huruf yang sama menunjukkan tidak
adanya perbedaan nyata pada tingkat kepercayaan 95% (Values
followed by the same letter are not significantly different at 95%
confidence level)
TEKNIK PENYIMPANAN BENIH MERANTI BALAU (Shorea seminis (de Vriese) Sloot)
Muhammad Zanzibar, Naning Yuniarti, dan Ratna Uli Damayanti S
119
Tabel (Table) 5. Pengaruh interaksi wadah dan periode simpan terhadap daya berkecambah benih
meranti balau (The effect of container interaction and storage period on the
germination percentage of meranti balau seed) (Uji Duncan) Periode simpan
(Period of storage)
Wadah (Container)
Kain belacu dalam kotak kayu (The calico
cloth is in a wooden box)
Kain belacu (Calico cloth)
0 hari (day) 96,00a 96,00a
3 hari (days) 87,33ab 82,00b
6 hari (days) 86,00ab 66,00c
9 hari (days) 68,67c 34,67ef
12 hari (days) 64,67c 26,00fg
15 hari (days) 49,67d 17,33gh
18 hari (days) 41,33de 8,67h
Keterangan (Remarks) : Angka-angka yang diikuti oleh huruf yang sama menunjukkan tidak adanya
perbedaan nyata pada tingkat kepercayaan 95 persen (Values followed by the same
letter are not significantly different at 95 percent confidence level)
Tabel (Table) 6. Pengaruh interaksi wadah dan ruang simpan terhadap daya berkecambah benih
meranti balau (The effect of the interaction of the container and the storage room
on the germination percentage of meranti balau seeds) (Uji Duncan) Ruang simpan
(Storage room)
Wadah (Container)
Kain belacu dalam kotak kayu (The
calico cloth is in a wooden box)
Kain belacu (Calico cloth)
AC (AC) 69,43a 37,71c
Kamar (ambient) 71,62a 56,76b
Keterangan (Remarks) : Angka-angka yang diikuti oleh huruf yang sama menunjukkan tidak adanya
perbedaan nyata pada tingkat kepercayaan 95 persen (Values followed by the
same letter are not significantly different at 95 percent confidence level) 3. Nilai perkecambahan
Hasil analisis ragam menunjukkan bahwa
perlakuan faktor periode simpan, ruang simpan
dan waktu simpan berpengaruh nyata terhadap
nilai perkecambahan benih. Interaksi periode
dan wadah, ruang dan wadah serta periode,
ruang dan wadah, juga memberikan pengaruh
yang nyata terhadap nilai perkecambahan
benih, sementara interaksi periode dan ruang
tidak berpengaruh nyata terhadap nilai
perkecambahan benih (Tabel 7). Untuk
mengetahui lebih lanjut perlakuan yang
menimbulkan perbedaan yang nyata,
dilakukan uji beda Duncan (Tabel 8).
Tabel (Table) 7. Analisis ragam pengaruh wadah, ruang dan periode simpan terhadap nilai
perkecambahan benih meranti balau (Analysis of variance for the effect of the
container, storage room, and the storage period on the germination value of meranti
balau seed)
Sumber keragaman
(Source of variation)
Derajat bebas (Degree of
freedom)
Jumlah kuadrat
(Sum of square)
Kuadrat tengah
(Mean of square)
F hitung
(F-calculation)
F tabel
(table) 5%
Wadah (container)/A
Ruang (room)/ B
Interaksi (interaction)/ AB
Periode (period)/C)
Interaksi A dan C (interaction A and C)
Interaksi B dan C (interaction B and C)
Interaksi A, B dan C (interaction A, B, and C)
1
1
1
6
6
1
6
1390,72
253,31
158,24
15514,29
520,67
68,89
334,31
1390,72
253,31
158,24
2585,72
86,78
11,48
55,72
77,07*
14,04*
8,77*
143,3*
4,81*
0,64tn
3,09*
2,27
4,01
4,01
2,27
2,27
2,27
2,27
Sisa (residual) 56 1010,47 18,04
Total (total) 83 19250,90
Keterangan (Remarks): * = berpengaruh nyata pada tingkat kepercayaan 95 persen (Significant at 95persen convident
level) tn = tidak berpengaruh nyata pada tingkat kepercayaan 95 persen (Not significant at 95
percent confidence level)

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019: 113-125
p-ISSN : 2354-8568
e-ISSN : 2527-6565
120
Tabel (Table) 8. Pengaruh wadah, ruang dan periode simpan terhadap nilai perkecambahan benih
meranti balau (The effect of the container, storage room, and storage period on the
germination value of meranti balau seed) (Uji Duncan)
Periode simpan
(Period of storage)
Wadah (Container)
Kain belacu
(Calico cloth)
Kain belacu dalam kotak kayu
(The calico cloth is in a wooden box)
Ruang simpan (Storage room) Ruang simpan (Room of storage)
AC (AC) Kamar (Ambient) AC (AC) Kamar (Ambient)
0 hari (day) 44,09 a 44,09 a 44,09 a 44,09 a
3 hari (days) 20,12 cd 30,11 b 29,85 b 29,43 b
6 hari (days) 10,12 fgh 23,99 bc 38,13 a 29,19 b
9 hari (days) 2,06 ij 8,38 ghi 14,23 defg 18,64 cde
12 hari (days) 0,24 j 7,21 ghij 12,86 defgh 16,71 cdef
15 har (days) 0,00 j 5,12 hij 8,85 ghi 11,18 efgh
18 hari (days) 0,00 j 1,26 ij 4,79 hij 8,67 ghi
Keterangan (Remarks) : Angka-angka yang diikuti oleh huruf yang sama menunjukkan tidak adanya perbedaan
nyata pada tingkat kepercayaan 95% (Values followed by the same letter are not
significantly different at 95% confidence level).
B. Pembahasan
1. Kadar air
Benih meranti balau mempunyai nilai
rata-rata kadar air awal benih sebesar 55,20
persen. Benih rekalsitran merupakan benih
dengan kadar air tinggi dan peka terhadap
pengeringan, dan tidak dapat disimpan dalam
waktu lama. Dalam proses penyimpanan
benih, kadar air benih merupakan faktor kritis
yang berpengaruh terhadap proses penurunan
mutu benih (Raka, Astiningsih, Nyana, &
Siadi, 2012).
Perlakuan interaksi antara penggunaan
kain belacu dalam kotak kayu yang disimpan
pada ruang kamar, hingga periode
penyimpanan 9 hari, kadar air benih masih
tetap tinggi (> 52 persen) dan kehilangan
kadar air lebih kurang 3 persen dibanding
kadar air awal (55,20 persen) Setelah benih
disimpan (hingga 18 hari) laju penurunan
kadar benih terendah diperoleh pada interaksi
penggunaan wadah kain belacu dalam kotak
kayu dan benih disimpan pada ruang suhu
kamar (suhu = 25ºC―27ºC, kelembapan= 90
persen), sedangkan laju penurunan kadar air
tertinggi diperoleh pada interaksi antara
penggunaan kain belacu yang disimpan pada
ruang suhu AC (suhu = 18ºC―22ºC,
kelembapan = 70 persen ― 80 persen).
Benih yang berwatak rekalsitran seperti
benih meranti balau, penurunan kadar air yang
drastis dan kondisi penyimpanan dengan suhu
rendah akan menyebabkan kerusakan pada sel
sehingga hilangnya viabilitas benih.
Penurunan kadar air selama penyimpanan
merupakan faktor kritis yang mempengaruhi
viabilitas benih (Sukesh & Chandrashekar,
2013). Wadah simpan yang tepat untuk
penyimpanan benih meranti balau yang dapat
menekan laju penurunan kadar air adalah
wadah kain belacu dalam kotak kayu.
Sedangkan ruang simpannya yang cocok yaitu

TEKNIK PENYIMPANAN BENIH MERANTI BALAU (Shorea seminis (de Vriese) Sloot)
Muhammad Zanzibar, Naning Yuniarti, dan Ratna Uli Damayanti S
121
ruang suhu kamar. Kondisi suhu dan
kelembapan wadah kain belacu dalam kotak
kayu berbeda dengan wadah kain blacu. Pada
wadah kain belacu dalam kotak kayu
mempunyai suhu 30,4ºC dan kelembapan 63
persen. Sedangkan suhu pada wadah kain
belacu 30,7ºC dan kelembapan 61 persen. Jadi
wadah kain belacu dalam kotak kayu lebih
cocok untuk menyimpan benih balau, karena
mempunyai suhu lebih rendah dan kelembapan
yang lebih tinggi dibandingkan dengan wadah
kain belacu.
Penurunan kadar air selama proses
penyimpanan menyebabkan gangguan
metabolisme, terjadinya kemunduran benih
dan akhirnya benih mengalami kematian
(Halimursyadah, 2012). Penurunan kadar air
merupakan faktor penyebab terjadinya
kemunduran benih rekalsitran (Tresniawati,
Murniati, & Widajati, 2014). Selama periode
simpan, viabilitas benih dipengaruhi oleh
beberapa faktor, salah satunya adalah kadar
air, karena kadar air merupakan faktor yang
paling penting dalam kemunduran benih.
Sutopo (2010), dan Mahjabin dan Abidi
(2015) menjelaskan bahwa selama
penyimpanan, kadar air benih bertanggung
jawab terhadap kerusakan benih. Menurut
Nurhasybi dan Suita (2012), kadar air benih
dapat dipertahankan tetap tinggi dengan
menggunakan wadah yang sarang seperti kain
belacu yang tidak kedap, agar pertukaran
udara tetap terjadi dengan bebas sehingga
terhindar dari terjadinya panas yang
berlebihan. Lodong et al. (2015) menyatakan
bahwa usaha untuk mempertahankan viabilitas
benih rekalsitran agar tetap optimal, yaitu
dengan menyimpan benih pada wadah tidak
kedap dan media simpan yang berkelembapan
tinggi.
2. Daya berkecambah
Rata-rata daya berkecambah awal benih
meranti balau sebesar 89 persen. Ruang kamar
lebih dapat menekan laju penurunan daya
berkecambah pada semua wadah. Wadah
terbaik adalah kain belacu dalam kotak kayu.
Hingga hari ke-3 penyimpanan, ke dua wadah
tersebut relatif sama pengaruhnya, namun
setelahnya (hingga hari ke -18) wadah kain
belacu dalam kotak kayu selalu lebih baik
pengaruhnya dibanding hanya menggunakan
kain belacu. Laju penurunan daya
berkecambah menggunakan kain belacu dalam
kotak kayu dapat dipertahankan tetap tinggi
hingga hari ke 6, dan sampai akhir
penyimpanan hari ke 18 daya berkecambah
masih dapat dipertahankan hampir separuhnya
(41,33 persen).
Semakin lama disimpan, viabilitas benih
meranti balau semakin menurun. Solikin
(2016) juga menyebutkan bahwa lama
penyimpanan berpengaruh nyata terhadap
persentase perkecambahan. Siahaan (2017)
melaporkan bahwa mutu benih akan
mengalami penurunan secara linear dengan
waktu, sehingga tempat penyimpanan yang
tepat untuk menjaga mutu benih penting
diketahui. Menurut Sulaiman, Harun, dan

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019: 113-125
p-ISSN : 2354-8568
e-ISSN : 2527-6565
122
Kurniawan (2010) menurunnya daya
berkecambah dan semakin lamanya waktu
penyimpanan disebabkan oleh cadangan
makanan dalam benih yang semakin menurun
termasuk kadar air sebagai bahan dari proses
metabolisme.
3. Nilai perkecambahan
Nilai perkecambahan menunjukkan
jumlah benih berkecambah dalam persen hari
sampai akhir pengujian yang merupakan
pencerminan daya tumbuh benih (Payung et
al., 2012). Benih meranti balau mempunyai
rata-rata nilai perkecambahan awal sebesar
44,09±0,60. Nilai perkecambahan tertinggi
setelah 18 hari penyimpanan yaitu 8,67±3,08
diperoleh pada perlakuan benih yang disimpan
di ruang suhu kamar dengan menggunakan
wadah kain belacu dalam kotak kayu (Tabel
8). Sedangkan nilai perkecambahan yang
terendah (0) dihasilkan dari perlakuan benih
yang disimpan di ruang AC dengan
menggunakan wadah kain belacu.
Nilai perkecambahan yang tinggi
menunjukkan vigor benih yang baik. Nilai
perkecambahan mengindikasikan
kesempurnaan dari viabilitas benih. Nilai
puncak perkecambahan menunjukkan energi
(daya) kecambah maksimum yang dicapai
benih pada waktu tertentu. Pada prinsipnya
energi perkecambahan dari suatu lot benih
mengikuti pola kurva normal, pada fase awal
akan meningkat secara signifikan sampai
mencapai titik maksimal kemudian menurun
kembali. Nilai puncak perkecambahan tersebut
menunjukkan vigor dari benih. Benih yang
memiliki vigor yang bagus ditandai dengan
nilai puncak perkecambahan yang tinggi yang
dicapai pada waktu yang relatif cepat
(Hidayat, 2007).
Wadah kain belacu dalam kotak kayu,
baik pada ruang AC dan ruang kamar relatif
lebih dapat mempertahankan vigor benih
hingga akhir periode pengamatan dan masih
tetap tinggi bila dibandingkan dengan interaksi
lainnya. Setelah penyimpanan hari ke 6, nilai
perkecambahan benih pada semua interaksi
perlakuan menurun dan kehilangan nilai
perkecambahan benih, yang paling drastis
menurun yaitu pada perlakuan interaksi
penggunaan wadah kain belacu yang disimpan
pada ruang AC.
Wadah kain belacu dalam kotak kayu
merupakan wadah tidak kedap yang
merupakan wadah yang tepat untuk
menyimpan benih rekalsitran seperti pada
benih meranti balau. Wadah kain belacu
dalam kotak kayu lebih cocok untuk
menyimpan benih balau, karena mempunyai
suhu lebih rendah (30,4ºC) dan kelembapan
yang lebih tinggi (61 persen) dibandingkan
dengan wadah kain belacu (suhu 30,7ºC dan
kelembapan 78 persen). Hal ini disebabkan
karena wadah yang tidak kedap mampu
menjaga kelembaban yang baik pada

TEKNIK PENYIMPANAN BENIH MERANTI BALAU (Shorea seminis (de Vriese) Sloot)
Muhammad Zanzibar, Naning Yuniarti, dan Ratna Uli Damayanti S
123
penyimpanan, kebutuhan benih terhadap air
tetap terpenuhi sampai akhir periode
penyimpanan sehingga menghasilkan daya
berkecambah benih lebih cepat untuk tumbuh
dan mampu menghadapi kondisi lapangan
yang sub optimum serta menghasilkan vigor
benih semakin tinggi (Lodong, Tambing, &
Adrianto, 2015).
Pada umumnya semakin lama benih
disimpan maka viabilitasnya dan vigor akan
semakin menurun. Benih akan mengalami
kemunduran benih dengan bertambahnya
waktu penyimpanan. Kemunduran viabilitas
benih adalah sesuatu proses yang tidak dapat
dicegah (Widajati et al., 2012). Selama
penyimpanan, benih akan mengalami penuaan
dan kemunduran. Kemunduran benih dapat
ditinjau dari aspek fisiologi yang diindikasikan
dengan penurunan daya berkecambah dan
vigor (Rohandi & Widyani, 2016). Demikian
halnya dengan benih meranti balau.
Kemunduran benihnya juga dicirikan dengan
adanya penurunan daya berkecambah dan
vigor benih.
Interaksi faktor perlakuan yang meliputi
wadah, ruang, dan periode simpan
berpengaruh nyata terhadap nilai
perkecambahan. Wadah yang terbaik untuk
benih meranti balau adalah wadah kain belacu
dalam kotak kayu dan ruang simpannya yang
cocok adalah AC. Dengan perlakuan ini dapat
mempertahankan viabilitas benih meranti
balau selama penyimpanan 18 hari. Jadi untuk
teknik penyimpanannya, benih meranti balau
memerlukan wadah yang tidak kedap dan suhu
ruang penyimpanannya berkisar 18 ºC ―22ºC
dengan kelembabannya berkisar 70
persen―80 persen.
IV. KESIMPULAN
Interaksi perlakuan wadah, ruang, dan
periode simpan berpengaruh nyata terhadap
nilai kadar air, dan nilai perkecambahan benih
meranti balau (Shorea seminis (de Vriese)
Sloot). Sedangkan interaksi perlakuan periode
dan ruang simpan, periode dan wadah simpan,
serta ruang dan wadah simpan berpengaruh
nyata terhadap daya berkecambah benih
meranti balau. Teknik penyimpanan yang
terbaik untuk benih meranti balau adalah
menggunakan wadah kain belacu dalam kotak
kayu yang disimpan dalam ruang suhu kamar.
Interaksi penggunaan wadah kain belacu
dalam kotak kayu dengan ruang suhu kamar
dapat menekan laju penurunan daya
berkecambah benih meranti balau selama
penyimpanan. Dengan perlakuan tersebut
benih dapat disimpan dengan baik selama 18
hari.
UCAPAN TERIMA KASIH
Penulis mengucapkan terima kasih kepada
Agung Supriyanto yang telah banyak
membantu dalam pengambilan data penelitian
di lapangan.
DAFTAR PUSTAKA
Aminah, A. (2011). Pengaruh penyimpanan
terhadap perubahan fisiologis, biokimia dan

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019: 113-125
p-ISSN : 2354-8568
e-ISSN : 2527-6565
124
kandungan minyak benih keranji (pongamia
pinnata Merr). In teknologi perbenihan untuk
meningkatkan produktivitas hutan rakyat di
propinsi jawa tengah. Semarang: Balai
Penelitian Teknologi Perbenihan tanaman
Hutan.
Djavanshir, K., & Pourbeik, H. (1976).
Germination value- a new formula. Silvae
Genetica, 25(2), 79–83.
Halimursyadah. (2012). Pengaruh kondisi simpan
terhadap viabilitas dan vigor benih Avicenia
marina (Forsk.) Vierh pada beberapa periode
simpan. Jurnal Agrotropika, 17(2), 43–51.
Hidayat, Y. (2007). Pengaruh waktu penyimpanan
buah terhadap viabilitas benih gmelia
(Gmelina arborea Roxb). Jurnal Wana Mukti,
5(1), 27–36.
ISTA. (2010). Rules Proposals for the
International Rules for Seed Testing 2014
Edition (2010th ed.). Canada: Canadian Food
Inspection Agency.
Lodong, O., Tambing, Y., & Adrianto. (2015).
Peranan kemasan dan media simpan terhadap
ketahanan viabilitas dan vigor benih namgka
(Artocarpus heterophyllus Lamk) kultivar
tulo-5 selama penyimpanan. Jurnal
Agrotekbis, 3(3), 303–315.
Maharani, R., Handayani, P., & Hardjana, A.
(2013). Panduan Identifikasi Jenis Pohon
Tengkawang. Balai Besar Penelitian
Dipterokarpa, Badan Litbang Kehutanan.
Samarinda, Indonesia: Departemen
Kehutanan, Bekerjasama Dengan ITTO
PROJECT PD 586/10 Rev.1 (F).
Mahjabin, S. B., & Abidi, A. B. (2015).
Physiological and biochemical changes during
seed deterioration: a review. International
Journal of Recent Scientific Research, 6(4),
3416–3422.
Muslich, M. & Sumarni, G. (2008). Standardisasi
Mutu Kayu Berdasarkan Ketahanannya
Terhadap Penggerek di Laut. Prosiding PPI
Standardisasi, 25 November 2008.
Nurhasybi, & Suita, E. (2012). Perkecambahan dan
vigor benih suren (Toona sureni (blume))
pada berbagai metode perkecambahan dan
pengeringan. In Teknologi perbenihan jenis-
jenis potensial untuk rehabilitasi lahan bekas
tambang di provinsi kepulauan Bangka
Belitung. Pangkal Pinang: Balai Penelitian
Teknologi Perbenihan tanaman Hutan.
Payung, D., E. Prihatiningtyas & S. H. Nisa.
(2012). Uji Daya Kecambah Benih Sengon
(Paraserianthes falcataria (L.) Nielsen) di
Green House. Jurnal Hutan Tropis, 13 (2) :
133-138
Puspitasari, D. (2011). Identifikasi Jenis
Shorea Menggunakan Jaringan Syaraf
Tiruan Propogasi Balik Berdasarkan
Karakteristik Morfologi Daun. Skripsi.
Departemen Ilmu Komputer Fakultas
Matematika Dan Ilmu Pengetahuan Alam.
IPB. Bogor.
Raka, I. G. N., Astiningsih, A. A. M., Nyana, I. D.
N., & Siadi, I. K. (2012). Pengaruh dry heat
treatment terhadap daya simpan benih cabai
rawit (Capsicum frutescens L.). J. Agric. Sci.
and Biotechnol, 1(1), 1–11.
Rohandi, A., & Widyani, N. (2016). Perubahan
fisiologis dan biokimia benih tengkawang
selama penyimpanan. Jurnal Penelitian
Ekosistem Dipterokarpa, 2(1), 9–20.
Saridan, A., Fernandes, A., & Noor, M. (2013).
Sebaran dan potensi pohon tengkawang di
Hutan Penelitian Labanan, Kalimantan Timur.
Jurnal Penelitian Dipterokarpa, 7(2), 101–
108. Retrieved from http://ejournal.forda-
mof.org/ejournal-
litbang/index.php/JPED/article/view/1360
SAS Institute Inc. (2004). SAS/INSIGHT 9.1
User’s Guide. Volume 1 and 2. Cary, NC:
SAS Institute Inc.
Schmidt, L. (2000). Pedoman Penanganan Benih
Tanaman Hutan Tropis dan Sub tropis.
(Direktorat Jendral Rehabilitasi Lahan dan
Perhutanan Sosial- Indosesia Forest Seed
Project, Ed.). Jakarta: PT. Gramedia
Indobnesia.
Siahaan, F. A. (2017). Pengaruh kondisi dan
periode simpan terhadap perkecambahan
benih kesambi (Schleichera oleosa (Lour.)
Merr). Jurnal Perbenihan Tanaman Hutan,
5(1), 1–11.
Solikin. (2016). Pengaruh lama penyimpanan
terhadap perkecambahan biji sambiloto
(Andrographis paniculata (Burm.F.) Wallich

TEKNIK PENYIMPANAN BENIH MERANTI BALAU (Shorea seminis (de Vriese) Sloot)
Muhammad Zanzibar, Naning Yuniarti, dan Ratna Uli Damayanti S
125
ex Nees). Berita Biologi, 15(2), 201–206.
Suita, E. (2013). Pengaruh Pengusangan Terhadap
Viabilitas Benih Weru (Acacia procera
Benth.). Jurnal Perbenihan Tanaman Hutan,
1(1), 37–42.
Sukesh & K.R. Chandrashekar. (2013). Effect of
Temperature on Viability and Biochemical
Changes During Storage of Recalcitarant
Seeds of Vatica chinensis L. International
Journal of Botany, 9 (3) : 73-79.
Sulaiman, F., Harun, M.., & Kurniawan, A. (2010).
Perkecambahan benih tanaman karet (Hevea
brasiliensis Muell. Arg) yang disimpan pada
suhu dan periode yang berbeda. In Prosiding
Seminar Nasional (pp. 1593–1603).
Palembang.
Sutopo, L. (2010). Teknologi benih. Jakarta: PT.
Raja Grafindo Persada.
Tresniawati, C., Murniati, E., & Widajati, E.
(2014). perubahan fisik, fisiologi dan
biokimia selama pemasakan benih dan studi
rekalsitransi benih kemiri sunan. Jurnal
Agronomi Indonesia, 42(1), 74–79.
Widajati, E., Murniaty, E., Palupi, E.., Kartika, T.,
Suhartanto, M., & Qadir, A. (2012). Dasar
Ilmu dan Teknologi Benih. Bogor Indonesia:
IPB Press.
Yuniarti, N., Syamsuwida, D., & Aminah, A.
(2013). Dampak perubahan fisiologi dan
biokimia benih eboni (Diospyros celebica
Bakh.) selama penyimpanan. Jurnal
Penelitian Hutan Tanaman, 10(2), 65–71.
Zannzibar, M., & Widodo, W. (2011). Metoda
pengeringan dan penyimpanan benih mahoni
(Swietenia macrophylla King). In Teknologi
Perbenihan untuk meningkatkan produktivitas
hutan rakyat di Propinsi Jawa Tengah.
Semarang: Balai Penelitian Teknologi
Perbenihan tanaman Hutan.

KERAGAMAN PERTUMBUHAN BIBIT NYAMPLUNG (Calophyllum inophyllum) ASAL SUMENEP, MADURA
Aam Aminah, Danu dan Yulianti Bramasto
Kontribusi penulis: Aam Aminah, Danu dan Yulianti Bramasto sebagai kontributor utama
© 2019 BPTPTH All rights reserved. Open access under CC BY-NC-SA license.doi: //doi.org/10.20886/bptpth.2019.7.2.127-138 127
KERAGAMAN PERTUMBUHAN BIBIT NYAMPLUNG (Calophyllum inophyllum) ASAL
SUMENEP, MADURA
(Growth Diversity of Nyamplung (Calophyllum inophyllum) Seedling from Sumenep, Madura)
*Aam Aminah, *Danu, dan/and *Yulianti Bramasto
Balai Penelitian dan Pengembangan Teknologi Perbenihan Tanaman Hutan
Jl. Pakuan Ciheuleut PO Box 105; Telp. (0251) 8327768, Bogor, Jawa Barat, Indonesia
e-mail: [email protected]
Naskah masuk: 14 Agustus 2018; Naskah direvisi: 26 November 2018; Naskah diterima: 12 Desember 2019
ABSTRACT
One source of potential biofuel feedstock in Indonesia is nyamplung (Calophyllum inophyllum). Superiority
of nyamplung is having a high oil content of 40 percent ―73percent. The purpose of this study was to
identify growth variation and heritability value for the height and diameter characters of nyamplung
seedlings at six months old from 26 parent trees from Sumenep, Madura. The research used Randomized
Complete Block Design with 26 families from Sumenep Madura as treatment, each treatment consisted of 3
replications,each replication consisted of 8 seedlings, so the number of seedlings used was 624 seedlings.
The results showed that there were diversity in growth (height and diameter) at the level of nyamplung
seedling from Sumenep, Madura. The height and diameter of nyamplung seedlings of 26 families tested
varied between 12.52 cm ― 21.27 cm and 3.04 mm ―4.54 mm. Individual heritability values for seedling
height and diameter are 0.27 and 0.16, while family heritability values are 0.65 for height and 0.52 for
diameter
Keyword: Calophyllum inophyllum , family, genetic, heritability
ABSTRAK
Salah satu sumber bahan baku biofuel yang berpotensi di Indonesia adalah nyamplung (Calophyllum
inophylum). Kelebihan nyamplung adalah memiliki rendemen minyak yang tinggi sebesar 40 persen―73
persen. Tujuan dari penelitian ini adalah untuk mengidentifikasi keragaman pertumbuhan dan nilai
heritabilitas untuk karakter tinggi dan diameter bibit nyamplung umur enam bulan dari 26 pohon induk asal
Sumenep, Madura. Penelitian menggunakan Rancangan Lengkap Berblok dengan 26 famili asal Sumenep
Madura sebagai perlakuan, setiap perlakuan terdiri dari 3 ulangan, setiap ulangan terdiri dari 8 bibit,
sehingga jumlah bibit yang digunakan sebanyak 624 bibit. Hasil penelitian menunjukkan bahwa terdapat
keragaman pertumbuhan (tinggi dan diameter) pada tingkat bibit tanaman nyamplung asal Sumenep,
Madura. Pertumbuhan tinggi dan diameter bibit nyamplung dari 26 famili yang diuji bervariasi antara 12,52
cm ―21,27 cm dan 3,04 mm ― 4,54 mm. Nilai heritabilitas individu pada tingkat bibit untuk karakter tinggi
dan diameter adalah 0,27 dan 0,16, sedangkan nilai heritabilitas famili adalah yaitu 0,65 untuk karakter
tinggi dan 0,52 untuk diameter. Kata kunci : Calophyllum inophyllum, famili, genetik, heritabilitas
I. PENDAHULUAN
Kebutuhan manusia akan energi saat ini
semakin meningkat sejalan dengan
pertumbuhan penduduk yang semakin pesat.
Sumber energi yang saat ini digunakan adalah
sumberdaya alam yang keberadaannya
semakin berkurang di alam (minyak bumi, gas
bumi, batu bara dan lain-lain), sedangkan
sumber daya alam terbarukan masih belum
optimal dalam pengelolaan dan
penggunaannya. Pemerintah merencanakan
penggunaan biodiesel, sebagai energi baru
terbarukan untuk mengurangi ketergantungan
terhadap bahan bakar fosil. Menurut
Tampubolon (2008) penggunaan energi
terbarukan dalam konteks diversifikasi energi
sangat strategis karena sejalan dengan
pembangunan berkelanjutan dan ramah

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019: 127-138
p-ISSN : 2354-8568
e-ISSN : 2527-6565
128
lingkungan (emisi gas rumah kaca relatif
rendah). Pemanfaatan sumber daya alam
terbarukan ini memungkinkan untuk dikelola
dalam jangka waktu yang relatif panjang
dibandingkan dengan penggunaan minyak
bumi dan bahan tambang sejenisnya
(Bustomi, Rostiwati, Sudrajat, Kosasih &
Effendi, 2009).
Nyamplung sebagai bahan baku biofuel
memiliki kelebihan di antaranya memiliki
rendemen minyak yang tinggi. Bustomi et al.
(2009) melaporkan bahwa nyamplung
memiliki rendemen minyak 40 persen―73
persen. Leksono Windyarini & Hasnah (2014)
melaporkan kandungan minyak buah
nyamplung dari Sumenep 53 persen dan
Gunung Kidul (DIY) 50 persen. Kandungan
minyak nyamplung lebih besar bila
dibandingkan dengan Tanaman lain, yaitu
malapari yang mengandung 23 persen ―26
persen minyak (Aminah, 2017). Selain itu
dalam pemanfaatannya, buah nyamplung tidak
berkompetisi dengan kebutuhan pangan.
Sebagai energi terbarukan, ketersediaan bahan
baku baik dari buah, maupun biomas atau
limbah kayu memiliki jaminan kelestarian.
Pengembangan energi alternatif terbarukan
akan mampu menciptakan lapangan kerja bagi
masyarakat di sekitar hutan. Dari berbagai
tahap kegiatan dapat menyerap tenaga kerja
cukup tinggi: pembangunan hutan tanaman,
pemeliharaan tahunan, pengolahan
biodiesel/biokerosin, pembangunan pabrik,
instalasi mesin, pasokan bahan baku,
tumbuhnya industri pendukung (kompos,
briket, gliserol dsb).
Mengingat potensi yang dimiliki jenis
tersebut maka pengembangan jenis nyamplung
sebagai jenis alternatif penghasil energi
merupakan salah satu solusi untuk mengatasi
ketidakseimbangan antara kebutuhan bioenergi
dengan produktivitasnya. Bustomi et al.
(2009) mengemukakan apabila seluruh
kebutuhan biodesel dipenuhi dari nyamplung,
akan dibutuhkan biodesel 720.000 kilo liter
atau setara dengan 5,1 juta ton biji nyamplung
dengan asumsi 2,5 kg biji nyamplung
menghasilkan 1 liter minyak nyamplung.
Bustomi et al. (2009) melaporkan bahwa
nyamplung memiliki produktivitas buah yang
tinggi yaitu 20 ton/ha. Untuk memenuhi
kebutuhan 5,1 jt ton biji nyamplung, maka
diperlukan tegakan nyamplung seluas 255.000
ha.
Untuk menyediakan bahan baku biodiesel
berbasis buah nyamplung, maka diperlukan
kebun benih nyamplung yang memiliki
produktivitas tinggi. Nyamplung yang berasal
dari Sumenep merupakan salah satu provenan
nyamplung yang memiliki kandungan minyak
yang tinggi (Leksono et al., 2014).
Pembangunan hutan tanaman nyamplung yang
mempunyai produktivitas tinggi memerlukan
bibit unggul, dalam hal ini adalah

KERAGAMAN PERTUMBUHAN BIBIT NYAMPLUNG (Calophyllum inophyllum) ASAL SUMENEP, MADURA
Aam Aminah, Danu dan Yulianti Bramasto
129
menghasilkan produksi buah yang tinggi
dengan kandungan minyak yang tinggi pula.
Untuk mengetahui variasi genetik dapat
dilakukan studi marka genetik maupun sifat-
sifat kuantitatif. Tujuan penelitian ini untuk
mengidentifikasi keragaman pertumbuhan
tinggi dan diameter serta menduga nilai
heritabilitas bibit nyamplung umur 6 bulan di
persemaian yang berasal dari 26 pohon induk
asal Sumenep, Madura.
II. BAHAN DAN METODE
A. Bahan dan Alat
Bahan yang digunakan adalah buah
nyamplung yang berasal dari 26 pohon induk
yang tumbuh tersebar dibeberapa Kecamatan
di wilayah Kabupaten Sumenep, Madura.
Letak geografis Kabuapaten Sumenep adalah
pada 113º 32’ ― 116º 16’ Bujur Timur dan 4º
55’ ― 7º 24’ Lintang Selatan. Pohon induk
yang diunduh buahnya terletak di Kecamatan
Batuputih, Kecamatan Batang, Kecamatan
Gapura, Kecamatan Saronggi, Kecamatan
Manding, Kecamatan Dasuk, dan Kecamatan
Ambunten, Kabupeten Sumenep, Madura.
Tegakan nyamplung yang diunduh buahnya
sebanyak 40 pohon induk yang memiliki tinggi
pohon antara 9 m ―15 m dan diameter batang
antara 21,02 cm ― 63,69 cm, namun yang
dianalisis dalam penelitian adalah bibit
nyamplung yang berasal dari 26 pohon induk.
Adapun bahan lainnya yang digunakan adalah
media perkecambahan dan semai, yaitu pasir,
tanah, pupuk kandang, arang sekam padi dan
polibag ukuran 12 cm x 15 cm.
Peralatan yang digunakan adalah bak
kecambah, penggaris, kaliper, kamera dan alat-
alat tulis. Penelitian dilakukan di persemaian
Stasiun Penelitian Nagrak, Balai Penelitian
dan Pengembangan Teknologi Perbenihan
Tanaman Hutan Bogor pada ketinggian tempat
200 m dpl. Pembuatan persemaian dimulai
pada bulan Maret 2017 dan pengamatan
dilakukan saat tanaman berumur 6 bulan.
B. Prosedur Penelitian
1. Rancangan penelitian
Penelitian ini menggunakan Rancangan
Acak Lengkap Blok (Randomized Complete
Blok Design) dengan 26 famili asal Sumenep,
Madura dengan ulangan sebanyak 3 blok.
Setiap ulangan terdiri atas 8 bibit, sehingga
jumlah bibit yang digunakan sebanyak 624
bibit.
Family
Family
Family
1 2 3 . . . . 26 1 2 3 . . . . 26 1 2 3 . . . . 26
Tree plot
1
2
3
.
.
.
8
Blok 1
Blok 2
Blok 3
Gambar (Figure) 1. Desain plot semai nyamplung di persemaian (Plot design of
nyamplung seedling in nursery)

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019: 127-138
p-ISSN : 2354-8568
e-ISSN : 2527-6565
130
2. Pembuatan bibit
Buah nyamplung setiap famili diekstraksi
dengan cara dipukul untuk mengeluarkan biji
dari cangkang buahnya. Benih dikecambahkan
dalam bak kecambah berisi campuran media
pasir dan tanah (1:1v/v), kemudian diletakkan
di rumah kaca dan diberi label nomor pohon
induk atau famili. Pemeliharaan semai dalam
bentuk penyiraman dan penyiangan gulma
dilakukan setiap hari hingga benih
berkecambah.
Semai yang telah berumur 5 minggu ―6
minggu disapih ke dalam polibag ukuran 12
cm x 15 cm yang berisi media campuran tanah
+ pupuk kandang + arang sekam padi (3:1:1,
v/v/v) dan diberi label nomor famili. Bibit
disimpan di bedeng semai selama 6 bulan dan
disiram satu kali setiap hari.
3. Karakter yang diamati
Karakter yang diamati adalah tinggi dan
diameter bibit, tinggi bibit diukur dari pangkal
batang sampai dengan pucuk dan diameter
diukur pada pangkal batang di permukaan
tanah di polibag.
C. Analisis Data
Data hasil pengukuran dianalisis
menggunakan analisis varian untuk
memperoleh data pengaruh famili (Fj) dan
blok terhadap pertumbuhan tinggi dan
diameter semai (µ). Model analisis varian yang
digunakan sebagai berikut (O’Neill, 2010):
ijk i j ijkY µ B F E= + + + ..................................1)
dimana:
Yijk : pengamatan pada individu ke-k dari
famili ke-j, dalam blok ke-i;
µ : rerata umum hasil pengukuran;
Bi : pengaruh blok ke-i
Fj : pengaruh famili ke-j;
Eijk : galat
Apabila terdapat pengaruh famili terhadap
karakter yang diamati yang nyata, maka
dilanjutkan dengan uji jarak berganda Duncan
untuk mengetahui perbedaan antar famili yang
diuji. Untuk mengetahui pengaruh faktor
genetik terhadap fenotipe dilakukan dengan
menghitung heritabilitas. Pendugaan nilai
heritabilitas famili dan individu dihitung
berdasarkan komponen ragam pada setiap
parameter yang diukur yaitu tinggi dan
diameter bibit nyamplung. Komponen ragam
famili (σ2f), komponen ragam plot (σ2p) dan
komponen ragam galat (σ2e) merupakan
turunan dari nilai kuadrat tengah harapan hasil
analisis ragam. Komponen ragam famili
diasumsikan sebesar 1/4 ragam genetik aditif
(σ2A), karena benih dikumpulkan dari pohon
induk dengan penyerbukan terbuka secara
alami di hutan rakyat. Nilai heritabilitas
individu (h2i) dan heritabilitas famili (h2f)
dihitung menggunakan rumus heritabilitas
yang telah disusun oleh Zobel & Talbert,
(1984):
2 2
2 2 2
4
fh i
f f p e
σ
σ σ σ=
+ +
dan2
2
2 2 2
fh f
f f pe
n jn
σ
σ σσ
=
+ +
...2)
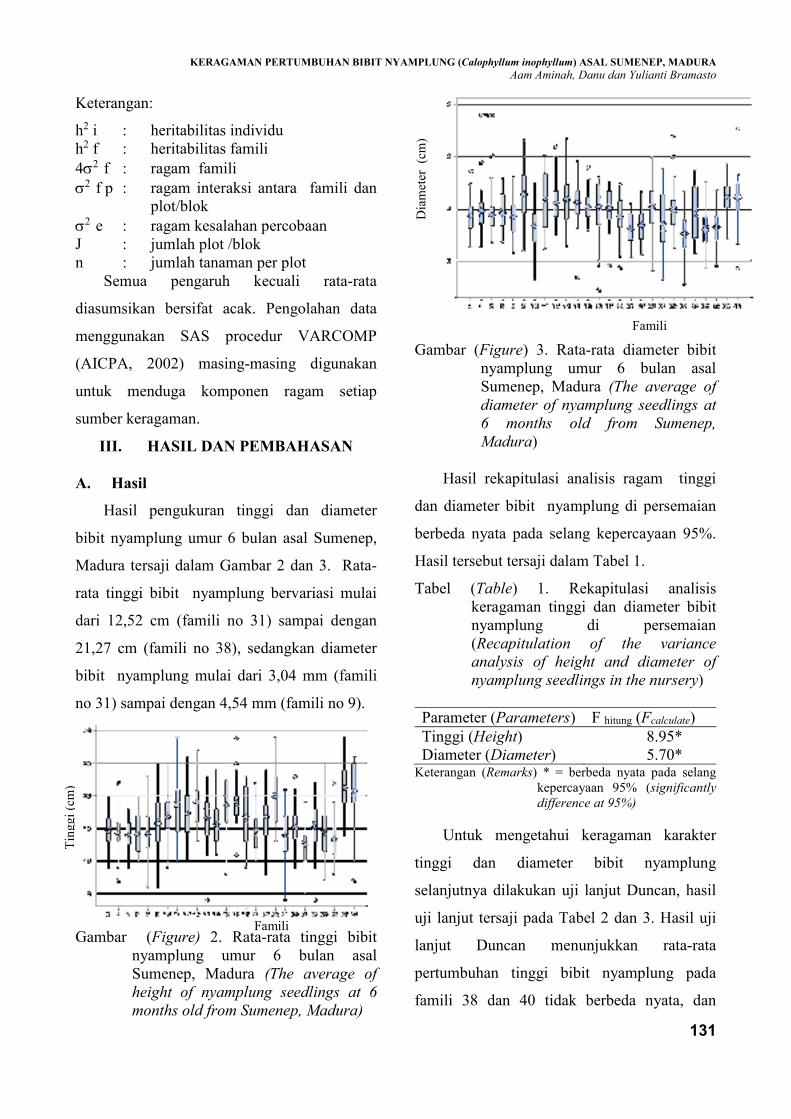
KERAGAMAN PERTUMBUHAN BIBIT NYAMPLUNG (Calophyllum inophyllum) ASAL SUMENEP, MADURA
Aam Aminah, Danu dan Yulianti Bramasto
131
Tin
gg
i (c
m)
Famili
Keterangan:
h2 i : heritabilitas individu
h2 f : heritabilitas famili
4σ2 f : ragam famili
σ2 f p : ragam interaksi antara famili dan
plot/blok
σ2 e : ragam kesalahan percobaan
J : jumlah plot /blok
n : jumlah tanaman per plot
Semua pengaruh kecuali rata-rata
diasumsikan bersifat acak. Pengolahan data
menggunakan SAS procedur VARCOMP
(AICPA, 2002) masing-masing digunakan
untuk menduga komponen ragam setiap
sumber keragaman.
III. HASIL DAN PEMBAHASAN
A. Hasil
Hasil pengukuran tinggi dan diameter
bibit nyamplung umur 6 bulan asal Sumenep,
Madura tersaji dalam Gambar 2 dan 3. Rata-
rata tinggi bibit nyamplung bervariasi mulai
dari 12,52 cm (famili no 31) sampai dengan
21,27 cm (famili no 38), sedangkan diameter
bibit nyamplung mulai dari 3,04 mm (famili
no 31) sampai dengan 4,54 mm (famili no 9).
Gambar (Figure) 2. Rata-rata tinggi bibit
nyamplung umur 6 bulan asal
Sumenep, Madura (The average of
height of nyamplung seedlings at 6
months old from Sumenep, Madura)
Gambar (Figure) 3. Rata-rata diameter bibit
nyamplung umur 6 bulan asal
Sumenep, Madura (The average of
diameter of nyamplung seedlings at
6 months old from Sumenep,
Madura)
Hasil rekapitulasi analisis ragam tinggi
dan diameter bibit nyamplung di persemaian
berbeda nyata pada selang kepercayaan 95%.
Hasil tersebut tersaji dalam Tabel 1.
Tabel (Table) 1. Rekapitulasi analisis
keragaman tinggi dan diameter bibit
nyamplung di persemaian
(Recapitulation of the variance
analysis of height and diameter of
nyamplung seedlings in the nursery)
Parameter (Parameters) F hitung (Fcalculate)
Tinggi (Height) 8.95*
Diameter (Diameter) 5.70* Keterangan (Remarks) * = berbeda nyata pada selang
kepercayaan 95% (significantly
difference at 95%)
Untuk mengetahui keragaman karakter
tinggi dan diameter bibit nyamplung
selanjutnya dilakukan uji lanjut Duncan, hasil
uji lanjut tersaji pada Tabel 2 dan 3. Hasil uji
lanjut Duncan menunjukkan rata-rata
pertumbuhan tinggi bibit nyamplung pada
famili 38 dan 40 tidak berbeda nyata, dan
Dia
met
er
(cm
)
Famili
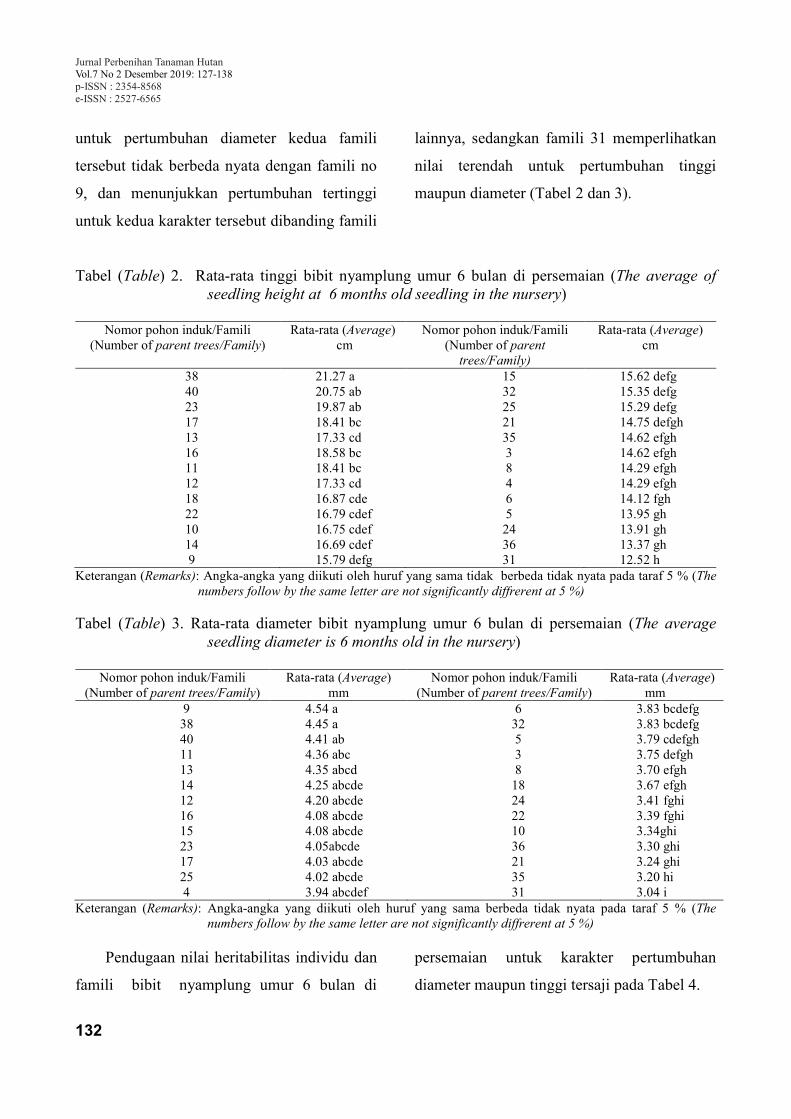
Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019: 127-138
p-ISSN : 2354-8568
e-ISSN : 2527-6565
132
untuk pertumbuhan diameter kedua famili
tersebut tidak berbeda nyata dengan famili no
9, dan menunjukkan pertumbuhan tertinggi
untuk kedua karakter tersebut dibanding famili
lainnya, sedangkan famili 31 memperlihatkan
nilai terendah untuk pertumbuhan tinggi
maupun diameter (Tabel 2 dan 3).
Tabel (Table) 2. Rata-rata tinggi bibit nyamplung umur 6 bulan di persemaian (The average of
seedling height at 6 months old seedling in the nursery)
Nomor pohon induk/Famili
(Number of parent trees/Family)
Rata-rata (Average)
cm
Nomor pohon induk/Famili
(Number of parent
trees/Family)
Rata-rata (Average)
cm
38 21.27 a 15 15.62 defg
40 20.75 ab 32 15.35 defg
23 19.87 ab 25 15.29 defg
17 18.41 bc 21 14.75 defgh
13 17.33 cd 35 14.62 efgh
16 18.58 bc 3 14.62 efgh
11 18.41 bc 8 14.29 efgh
12 17.33 cd 4 14.29 efgh
18 16.87 cde 6 14.12 fgh
22 16.79 cdef 5 13.95 gh
10 16.75 cdef 24 13.91 gh
14 16.69 cdef 36 13.37 gh
9 15.79 defg 31 12.52 h
Keterangan (Remarks): Angka-angka yang diikuti oleh huruf yang sama tidak berbeda tidak nyata pada taraf 5 % (The
numbers follow by the same letter are not significantly diffrerent at 5 %)
Tabel (Table) 3. Rata-rata diameter bibit nyamplung umur 6 bulan di persemaian (The average
seedling diameter is 6 months old in the nursery)
Nomor pohon induk/Famili
(Number of parent trees/Family)
Rata-rata (Average)
mm
Nomor pohon induk/Famili
(Number of parent trees/Family)
Rata-rata (Average)
mm
9 4.54 a 6 3.83 bcdefg
38 4.45 a 32 3.83 bcdefg
40 4.41 ab 5 3.79 cdefgh
11 4.36 abc 3 3.75 defgh
13 4.35 abcd 8 3.70 efgh
14 4.25 abcde 18 3.67 efgh
12 4.20 abcde 24 3.41 fghi
16 4.08 abcde 22 3.39 fghi
15 4.08 abcde 10 3.34ghi
23 4.05abcde 36 3.30 ghi
17 4.03 abcde 21 3.24 ghi
25 4.02 abcde 35 3.20 hi
4 3.94 abcdef 31 3.04 i
Keterangan (Remarks): Angka-angka yang diikuti oleh huruf yang sama berbeda tidak nyata pada taraf 5 % (The
numbers follow by the same letter are not significantly diffrerent at 5 %)
Pendugaan nilai heritabilitas individu dan
famili bibit nyamplung umur 6 bulan di
persemaian untuk karakter pertumbuhan
diameter maupun tinggi tersaji pada Tabel 4.

KERAGAMAN PERTUMBUHAN BIBIT NYAMPLUNG (Calophyllum inophyllum) ASAL SUMENEP, MADURA
Aam Aminah, Danu dan Yulianti Bramasto
133
Tabel (Table) 4. Nilai Heritabilitas Tinggi dan diameter bibit nyamplung umur 6 bulan di
persemaian (Heritabity value of height and diameter of 6 months old nyamplung
seedling in nursery)
Karakter
(Characters)
Komponen Varian
(Variance Component)
Nilai heritabilitas (Heritability values)
Individu (h2 i) Famili (h2 f)
Diameter
(diameter)
Var (Rep) -0.002996 0.159513 0.519012
Var (Family) 0.034073
Var (Rep *Fam) 0.04919
Var (Error) 0.77115
Tinggi
(height)
Var (Rep) -0.05319 0.274094 0.654305
Var (Family) 1.169645
Var (Rep *Fam) 0.97991
Var (Error) 14.91968
Nilai heritabilitas individu untuk diameter
adalah 0,16 sedangkan untuk tinggi adalah
0,27 dan nilai heritabilitas famili untuk
diameter adalah 0,52 sedangkan untuk tinggi
sebesar 0,65.
B. Pembahasan
Berdasarkan hasil analisis ragam (Tabel
1), terdapat adanya keragaman pertumbuhan
tinggi dan diameter pada bibit nyamplung
umur 6 bulan yang benihnya berasal dari 26
pohon induk di Kabupaten Sumenep Madura.
Untuk selanjutnya dalam pembahasan ini,
istilah pohon induk disesuaikan menjadi
famili. Keragaman pertumbuhan antar famili
ditunjukkan pada Gambar 2 dan Gambar 3,
terlihat bahwa bibit yang berasal dari famili no
31 mempunyai pertumbuhan tinggi dan
diameter rata-rata terendah bila dibandingkan
dengan famili-famili yang lain. Sedangkan
pertumbuhan bibit yang benihnya berasal dari
famili no 38 dan 40 mempunyai pertumbuhan
tinggi dan diameter tertinggi. Tinggi bibit
nyamplung umur 6 bulan di persemaian
berkisar antara 12,52 cm hingga 21,27 cm dan
diameter berkisar antara 3,04 mm hingga 4,54
mm. Keragaman pertumbuhan pada tingkat
bibit merupakan hasil interaksi antara
pengaruh lingkungan (suhu, kelembaban,
media bibit) dan genetik (asal benih atau
pohon induk). Adanya keragaman
pertumbuhan pada bibit yang benihnya berasal
dari 26 famili atau pohon induk nyamplung,
diduga karena adanya pengaruh lingkungan
dan pengaruh genetik yang diturunkan dari
induknya. Pohon induk yang digunakan dalam
penelitian ini tumbuh tersebar di 7 Kecamatan,
di Kabupaten Sumenep Madura. Menurut
pendapat Zobel & Talbert (1984) dikatakan
bahwa adanya perbedaan kondisi tempat
tumbuh setiap pohon induk dalam provenan
akan mempengaruhi sifat genetiknya.,
terutama sifat tinggi dan diameter. Untuk
menduga seberapa besar faktor lingkungan dan
genetik mempengaruhi karakter pertumbuhan
tinggi dan diameter bibit nyamplung umur 6
bulan di persemaian, dapat diduga dengan
menghitung nilai heritabitabilitas
Nilai heritabilitas adalah parameter yang
menunjukkan kuat atau lemahnya suatu
karakter dipengaruhi oleh faktor lingkungan

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019: 127-138
p-ISSN : 2354-8568
e-ISSN : 2527-6565
134
atau faktor genetik. Pendugaan nilai
heritabilitas individu dan famili untuk
diameter bibit nyamplung masing-masing
sebesar 0,16 dan 0,52, sedangkan untuk tinggi
sebesar 0,27 dan 0,65 (Tabel 4). Merujuk
pendapat (Satriawan, Sugiharto & Ashari,
2017), maka nilai heritabilitas individu untuk
tinggi bibit nyamplung termasuk kategori
rendah (0,16), dan untuk diameter bibit
termasuk kategori sedang (0,27), sedangkan
nilai heritabilitas famili termasuk kategori
tinggi (0,52 dan 0,65). Pendugaan nilai
heritabilitas merupakan bagian dalam
pemuliaan, karena informasi ini dibutuhkan
dalam seleksi tanaman, sehingga akan
diperoleh hasil yang sesuai dengan yang
diinginkan (Zobel & Talbert, 1984). Tinggi
rendahnya heritabilitas ini berhubungan
dengan strategi seleksi, dan kemajuan genetik
yang diperoleh dari masing-masing strategi
yang nanti digunakan. Menurut Akhtar, Oki ,
Adachi & Khan (2007) dikatakan bahwa nilai
heritabilitas yang rendah berpengaruh
terhadap respon seleksi, rendahnya nilai
heritabilitas maka rendah pula responnya
terhadap seleksi.
Nilai heritabilitas merupakan suatu
petunjuk seberapa besar suatu karakter atau
sifat dipengaruhi oleh faktor genetik atau
lingkungan (Herawati, Purwoko & Dewi,
2009). Nilai heritabilitas dapat digunakan
untuk mengetahui tingkat keterwarisan sifat
tetua kepada keturunannya dan untuk
mengetahui proporsi genetik terhadap fenotip.
Menurut Putri, Aswidinnoor & Asmono
(2009) nilai duga heritabilitas yang tinggi
untuk suatu karakter menggambarkan karakter
tersebut penampilannya lebih ditentukan oleh
faktor genetik dibandingkan dengan faktor
lingkungan.
Penelitian tentang keragaman
pertumbuhan pada tingkat bibit telah
dilakukan pada beberapa jenis tanaman hutan,
diantaranya untuk jenis gempol (Putri, Yulianti
& Danu, 2016), meranti (Sulistyawati, P.,
Widyatmoko, AYPB & Nurtjahjaningsih,
2014), araucaria (Setiadi, 2018) dan jabon
putih (Yudohartono & Herdiyanti, 2013)
Penelitian tersebut sebagai awal untuk
mengetahui ada tidaknya pengaruh pohon
induk terhadap pertumbuhan tingkat bibit,
yang selanjutnya dapat menjadi bahan
pertimbangan dalam kegiatan seleksi. Seleksi
merupakan salah satu proses dalam program
pemuliaan tanaman untuk perbaikan karakter.
Kegiatan seleksi sangat ditentukan oleh
tersedianya keragaman genetik yang luas
dengan heritabilitas tinggi. Dalam suatu
populasi tanaman, hanya nilai fenotipik yang
dapat diamati dan diukur secara langsung.
Ragam fenotip dari suatu populasi
menunjukkan tingkat perbedaan fenotipik
antar kelompok individu yang timbul akibat
adanya ragam genetik dan atau lingkungan

KERAGAMAN PERTUMBUHAN BIBIT NYAMPLUNG (Calophyllum inophyllum) ASAL SUMENEP, MADURA
Aam Aminah, Danu dan Yulianti Bramasto
135
(Falconer & Mackay, 1996). Jika ragam
genetik yang membentuk suatu karakter
diketahui, maka heritabilitas dari karakter
tersebut dapat diduga. Heritabilitas merupakan
penduga yang penting dari derajat respons
suatu populasi terhadap seleksi alami maupun
seleksi buatan. Pendugaan heritabilitas sangat
berguna untuk melihat nilai relatif dari seleksi
yang dilakukan berdasarkan ekspresi fenotipik
dari karakter-karakter yang berbeda (Razzaq
Hammad, Tahir & Sadaqat, 2014). Falconer &
Mackay (1996) menyatakan bahwa suatu
karakter yang mempunyai nilai duga
heritabilitas tinggi menandakan bahwa
penampilan karakter tersebut kurang
dipengaruhi oleh lingkungan. Seleksi dapat
berlangsung lebih efektif pada karakter yang
memiliki nilai duga heritabilitas tinggi karena
pengaruh lingkungan sangat kecil. Namun
demikian nilai heritabilitas dapat berubah
sesuai dengan kondisi pertumbuhan dan
perlakuan (Sumardi, 2016). Dalam penelitian
ini karena yang dianalisis adalah pada tingkat
bibit, maka nilai heritabilitas yang telah
diperoleh belum dapat digunakan sebagai
dasar dalam kegiatan seleksi di lapangan.
Selain itu karakter atau sifat yang akan
diseleksi pada nyamplung adalah produktivitas
buah serta kandungan minyaknya, dan karakter
ini baru bisa terlihat setelah uji keturunan di
lapangan.
Berdasarkan hasil analisis ragam terdapat
pengaruh yang nyata pada karakter tinggi dan
diameter bibit nyamplung yang diduga
diakibatkan oleh perbedaan antara famili,
sehingga terdapat keragaman yang tinggi pada
karaketer tinggi dan diameter bibit nyamplung
di persemaian. Hal ini diduga adanya
keragaman antara 26 famili atau pohon induk
nyamplung yang digunakan dalam penelitian
ini, sehingga berpengaruh nyata terhadap
tinggi dan diameter bibit nyamplung. Pohon
induk nyamplung yang digunakan berasal dari
7 populasi yang berbeda, dalam hal ini adalah
7 kecamatan di Kabupaten Sumenep, Madura.
Tinggi dan diameter pohon induk yang
tersebar di 7 kecamatan tersebut berkisar
antara 9 m – 15 m dan 21,02 cm - 63,69 cm
sehingga diduga hal ini yang mempengaruhi
ekspresi pertumbuhan yang berbeda pada
tingkat bibit. Keragaman pada tingkat bibit
juga terlihat pada jenis gempol, meranti dan
araukaria yang disebabkan oleh perbedaan
pohon induk atau sumber benih (Putri, et al.,
2016; Sulistyawati, et al., 2014 dan Setiadi,
2018). Keragaman yang timbul akibat
perbedaan famili atau pohon induk, juga
terlihat dari nilai pendugaan heritabilitas famili
(h2f) yang cukup tinggi untuk karakter tinggi
dan diameter bibit nyamplung umur 6 bulan di
persemaian. Nilai h2f untuk tinggi dan
diameter adalah 0,65 dan 0,52, yang berarti
bahwa lebih dari 50 persen faktor famili atau
pohon induk, dalam hal ini adalah faktor
genetik mempengaruhi ekspresi sifat tinggi
dan diameter bibit nyamplung umur 6 bulan.
Hasil serupa juga diperlihatkan pada bibit
nyawai umur 8 bulan di persemaian (Haryjanto

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019: 127-138
p-ISSN : 2354-8568
e-ISSN : 2527-6565
136
& Prastyono, 2014), yaitu nilai heritabilitas
famili untuk karakter tinggi dan diameter bibit
masing-masing adalah 0.98 dan 0,91.
Demikian pula hasil perhitungan nilai
heritabilitas famili pada bibit mahoni umur 5
bulan di persemaian (Mashudi, Susanto &
Darwo, 2017), nilai heritabilitas famili untuk
karakter tinggi dan diameter adalah 0,74 dan
0,75. Pendugaan keragaman genetik telah
dilakukan pula pada tanaman eboni umur 12
bulan di persemaian (Kinho, Halawane,
Irawan & Kafiar, 2015), nilai heritabilitas
famili untuk tinggi dan diameter eboni umur
12 bulan adalah 0,97 dan 0,89. Tingginya
nilai heritabilitas famili pada bibit
dipersemaian, diduga karena faktor lingkungan
di persemaian relatif seragam, sehingga variasi
yang terjadi lebih dominan dipengaruhi oleh
pohon induk.
Informasi tentang keragaman dan nilai
heritabilitas untuk karakter tinggi dan diameter
di tingkat bibit pada nyamplung dapat
digunakan sebagai informasi awal untuk
mengetahui adanya keragaman serta
mengetahui berapa besar sifat-sifat induk
diturunkan kepada keturunanya (faktor
genetik). Informasi ini diperlukan untuk
membangun tanaman uji keturunan yang
nantinya dapat dikonversi menjadi sumber
benih nyamplung. Keragaman pertumbuhan
yang terekspresikan pada tingkat bibit
merupakan hasil interaksi antara pengaruh
lingkungan dan genetik, namun untuk
mengetahui pengaruh yang dominan, dapat
diduga dari nilai heritabilitas. Nilai
heritabilitas individu untuk sifat tinggi dan
diameter bibit nyamplung umur 6 bulan
tergolong rendah yaitu 0,27 dan 0,16, hal ini
mengindikasikan keragaman tersebut kuat
dipengaruhi oleh faktor lingkungan (kondisi
lingkungan persemaian) dan pengaruh faktor
genetiknya lemah. Rendahnya nilai
heritabilitas indvidu dapat disebabkan
beberapa faktor, antara lain sistem
penyerbukan atau sistem perkawinan pada
jenis nyamplung yang terbatas. Hasil
penelitian (Yudohartono, T.P & Fambayun,
2012) menyatakan bahwa dalam program
pemuliaan jenis binuang keragaman genetik
yang tinggi sangat penting karena optimalisasi
perolehan genetik akan dapat dicapai dengan
semakin besarnya peluang untuk seleksi
terhadap sifat-sifat yang diinginkan. Oleh
karena itu strategi seleksi yang harus
dilakukan tergantung sifat yang diinginkan.
Seleksi dapat dilakukan pada tanaman uji
keturunan, khusus pada nyamplung seleksi
dapat dilakukan pada saat pohon tersebut
sudah berbuah. Data yang diperoleh tersebut
dapat menunjukkan pohon induk mana yang
mempunyai produksi buah dan kandungan
minyak yang paling tinggi sehingga dapat
dikembangkan dan dipilih pohon-pohon induk
yang mempunyai potensi yang paling baik dari

KERAGAMAN PERTUMBUHAN BIBIT NYAMPLUNG (Calophyllum inophyllum) ASAL SUMENEP, MADURA
Aam Aminah, Danu dan Yulianti Bramasto
137
segi produktivitas buah dan kandungan
minyaknya.
IV. KESIMPULAN
Pertumbuhan tinggi dan diameter bibit
nyamplung umur 6 bulan di persemaian yang
berasal dari Sumenep, Madura, menunjukkan
keragaman yang cukup tinggi. Variasi
pertumbuhan tinggi diantara 26 famili yang
diuji adalah berkisar antara 12,52 cm – 21, 27
cm dan variasi pertumbuhan diameter adalah
3,04 mm hingga 4,54 mm. Terlihat ada
kecenderungan bahwa famili yang terendah
pertumbuhan tinggi, juga terendah dalam
pertumbuhan diameternya, demikian pula
terlihat konsistensi untuk famili dengan
pertumbuhan tinggi dan diameter tertingginya.
Keragaman tersebut sangat dipengaruhi oleh
perbedaan pohon induk atau famili, yang
ditunjukkan pula dengan tingginya nilai
heritabiltas famili dibandingkan heritabilitas
individu pada tingkat bibit, akan tetapi
kegiatan seleksi untuk pencarian famili terbaik
untuk nyamplung sebaiknya dilakukan pada
tingkat lapang dengan mempertimbangkan
produktivitas buah dan kandungan minyak
pada buah dari setiap famili karena hal ini
sesuai dengan tujuan dari penanaman
nyamplung, yaitu sebagai bahan baku biofuel.
UCAPAN TERIMA KASIH
Ucapan terima kasih disampaikan
kepada Ir. Rina Kurniaty, Prof. Riset. Dr. Dida
Syamsuwida, Ir. Naning Yuniarti, Ir.
Nurmawati Siregar, M.Si., Bapak Ateng
Rahmat Hidayat dan Bapak Sutrisno yang
telah bekerja sama dalam Tim Energi.
DAFTAR PUSTAKA
Akhtar, M.S., Oki Y., Adachi, T. & Khan, H.
(2007). analyses of the genetic parameters (
variability, heritability, genetic advance,
relationship of yield and yield contributing
characters) for some plant traits among
brassica cultivars under phosphorus starved
environmental cues. Journal of the Faculty of
Environmental Science and Technology, 12(l),
91–98.
American Institute of Certified Public Accountant
(AICPA). (2002). Statement on Auditing
Standards (SAS) No. 99: Consideration of
Fraud in a Financial Staetement Audit,
AICPA”. New York.
Aminah, A. (2017). Karakterisasi morfologi,
genetik, kandungan minyak dan evaluasi awal
pertumbuhan bibit pongamia (Pongamia
pinnata (L.) Pierre) di Pulau Jawa. Disertasi.
Sekolah Pascasarjana. Bogor: Institut
Pertanian Bogor,
Bustomi, S., Rostiwati, T., Sudrajat, R., Kosasih,
S., …. & Effendi, R. (2009). Nyamplung
(Calophyllum inophyllum L) Sumber Energi
Biofuel yang Potensial. Ed revisi. Bogor:
Badan Litbang Kehutanan.
Falconer, Douglas S. & Mackay, T. F. C. (1996).
Introduction to Quantitative Genetics (4th
Edition). Longman.
Haryjanto, L & Prastyono (2014) Pendugaan
parameter genetik semai Nyawai (Ficus
variegata blume) asal pulau Lombok. Jurnal
Penelitian Kehutanan Wallacea, 3(1), 37-45
Herawati, R., Purwoko, B. S., & Dewi, I. S.
(2009). Keragaman genetik dan karakter
agronomi galur haploid ganda padi gogo
dengan sifat-sifat tipe baru hasil kultur antera.
J. Agron. Indonesia, 37(2), 87–94.
Kinho, J., Halawane, J., Irawan, A., & Kafiar, Y.
(2015, Juli). Prosiding Seminar Nasional
Masyarakat Biodiversitas Indonesia.
Evaluation of plant growth on progeny test

Jurnal Perbenihan Tanaman Hutan Vol.7 No 2 Desember 2019: 127-138
p-ISSN : 2354-8568
e-ISSN : 2527-6565
138
ebony (Diospyros rumphii) age one year in
the nursery (Vol. 1, No. 4, pp. 800-804).
Leksono, B., Windyarini, E., & Hasnah, T. M.
(2014). Budidaya Nyamplung (Calophyllum
inophyllum L.) untuk Bioenergi dan Prospek
Pemanfaatan Lainnya. Bogor: IPB Press.
Mashudi, M., Susanto, M., & Darwo, D. (2017).
Keragaman dan estimasi parameter genetik
bibit mahoni daun lebar (Swietenia
macropylla King.) di Indonesia. Jurnal
Penelitian Hutan Tanaman, 14(2), 115-125.
O’Neill, M. (2010). Anova & reml A Guide to
linear mixed models in an experimental
design context. Training.
Putri, L.A.P., Aswidinnoor, H., & Asmono, D.
(2009). Keragaan genetik dan pendugaan
heritabilitas pada komponen hasil dan
kandungan β-karoten progeni kelapa sawit
genetics performance and heritability
estimations on yield component and β-
carotene content of oil palm progenies. J.
Agron. Indonesia, 37(2), 145–151.
Putri, KP., Yulianti & Danu. (2016). Keragaman
pertumbuhan bibit gempol (Nauclea orientalis
L.) dari beberapa pohon induk. Jurnal Hutan
Tropis,4(1),8-13.
DOI:http://dx.doi.org/10.20527/jht.v4i1.2876
Razzaq, H., Hammad, M., Tahir, N., & Sadaqat, H.
A. (2014). Genetic variability in sunflower
(Helianthus annuus L .) for achene yield and
morphological characters. International
Journal of Science and Nature, 5(4), 669–676.
Satriawan I.B, Sugiharto, A. N. & Ashari S.
(2017). Heritabilitas dan kemajuan genetik
harapan populasi F 2 pada tanaman cabai
besar (Capsicum annuum L.). Jurnal Produksi
Tanaman, 5(2), 343–348.
Setiadi, D. (2018, Oktober). In Proceeding Biology
Education Conference: Biology, Science,
Enviromental, and Learning. Keragaman
genetik araucaria cunninghamii sumber asal
benih kepulauan Papua pada pertumbuhan di
tingkat semai. (Vol. 15, No. 1, pp. 785-790).
Sumardi, S. (2016). Variasi genetik pada
pertumbuhan tanaman konservasi sumberdaya
genetik cendana (Santalum album Linn.)
populasi pulau Timor bagian timur. Jurnal
Ilmu Lingkungan, 14(1), 27.
https://doi.org/10.14710/jil.14.1.27-31
Sulistyawati, P., Widyatmoko, AYPB. &
Nurtjahjaningsih. (2014). Keragaman genetik
anakan Shorea leprosula berdasarkan penanda
mikrosatelit. Jurnal Pemuliaan Tanaman
Hutan, 8(3), 171-183.
Tampubolon, A. P. (2008). Kajian kebijakan energi
biomassa kayu bakar. Analisis Kebijakan
Kehutanan, 5(1), 29–37.
Yudohartono, T.P & Fambayun, R. A. (2012).
Karakteristik pertumbuhan semai binuang asal
provenan Pasaman Sumatera Barat. Jurnal
Pemuliaan Tanaman Hutan, 6(3), 143–156.
Yudohartono, T. P., & Herdiyanti, P. R. (2013).
Variasi karakteristik pertumbuhan bibit jabon
dari dua provenanberbeda. Jurnal Penelitian
Hutan Tanaman, 10(1), 7-16.
Zobel, B.J. & Talbert, J. (1984). Applied Forest
Tree Improvement. New York: Jonhn Wiley.

JURNAL PERBENIHAN TANAMAN HUTAN
ISI VOLUME 7
Nomor 1
Ganis Citra Purmadewi, Arum Sekar Wulandari dan/and Ratna Uli Damayanti
PENGARUH METODE PENGAKARAN DAN MEDIA AKLIMATISASI TERHADAP
KEBERHASILAN AKLIMATISASI TEMBESU (Fagraea fragrans (Roxb.) Miq.)................ 1-12
Rina Laksmi Hendrati and/ dan Siti Husna Nurrohmah
BREEDING FOR BETTER GROWTH OF Calliandra calothyrsus Meissn., A FAST
GROWING LEGUME FOR WOOD-ENERGY ......................................................................... 13-20
Nurmawati Siregar dan/and Aam Aminah
PENGARUH WAKTU PENGUNDUHAN DAN WARNA KULIT BUAH TERHADAP
DAYA BERKECAMBAH DAN PERTUMBUHAN BIBIT MINDI (Melia azadarech Linn ... 21-30
Dewi Ayu Lestari
TEHNIK PENYIMPANAN BENIH REKALSITRAN Mesua ferrea L. DAN Swinglea
glutinosa (Blanco) Merr............................................................................................................... 31-44
Y.M.M. Anita Nugraheni, Ratna Uli Damayanti Sianturi dan/and Yulianti Bramasto
PENGARUH PRA-STERILISASI TERHADAP KEBERHASILAN INISIASI EKSPLAN
YANG BERASAL DARI PUCUK SENGON ............................................................................ 45-54
Arif Irawan dan/and Hanif Nurul Hidayah
PENGARUH KONDISI DAN PERIODE SIMPAN TERHADAP PERKECAMBAHAN
BENIH CEMPAKA WASIAN (Magnolia tsiampaca (Miq.) Dandy) ........................................ 55-65
Nomor 2
Dwi Kartikaningtyas, Teguh Setyaji, dan/and Surip
EFEKTIVITAS PENGGUNAAN MEDIA TANAM DAN PUPUK TERHADAP
KEMAMPUAN BERTUNAS TANAMAN PANGKAS KESS (Lophostemon suaveolens
(Sol.ex Gaertn.) Peter G.Wilson & J.T. Waterh) ......................................................................... 67-76
Nurmawati Siregar dan/and Kurniawati Purwaka Putri
PENGARUH UMUR TUNAS PANGKASAN TERHADAP PERTUMBUHAN SETEK
Hopea odorata Roxb. .................................................................................................................. 77-85
Asep Rohandi dan/and Dede J. Sudrajat
KARAKTERISASI PERTUMBUHAN AWAL BEBERAPA FAMILI MANGLID PADA UJI
KETURUNAN DI SUKAMANTRI, KABUPATEN CIAMIS ................................................... 87-100
Tati Suharti, Kurniawati Purwaka Putri, dan/and Yulianti Bramasto
KETAHANAN BIBIT SENGON (Falcataria moluccana (Miq) Barneby & J.W. Grimes)
TERHADAP CENDAWAN Uromycladium falcatarium BERDASARKAN ASAL BENIH
DAN JENIS PENGENDALI .................................................................................................. 101-111
Muhammad Zanzibar, Naning Yuniarti, dan/and Ratna Uli Damayanti S
TEKNIK PENYIMPANAN BENIH MERANTI BALAU (Shorea seminis (de Vriese) Sloot) 113-125
Aam Aminah, Danu dan/and Yulianti Bramasto
KERAGAMAN PERTUMBUHAN BIBIT NYAMPLUNG (Calophyllum inophyllum) ASAL
SUMENEP, MADURA .............................................................................................................. 127-138

INDEX KATA KUNCI VOLUME 7
Anthocepalus macrophyllus
arabic gum
asal benih
benih
bibit
biocontrol
budidaya
busuk pangkal batang
Calophyllum inophyllum
CMC
daun
daya berkecambah
daya simpan
dormansi benih
dosis
epidermis
Eucalyptus pellita
family
Falcataria moluccana
fotosintesis
gaharu
genetik
gypsum
heritabilitas
Hibiscus macrophyllus
Hopea
hormon
hutan penelitian Parungpanjang
in vitro
indeks stomata
juvenilitas
karakteristik pertumbuhan
karat puru
kayu energi
keragaman
kerapatan stomata
kemampuan bertunas
Kilemo
klon unggul
41
131
101
173
101, 173
131
145
131
127
131
159
49
49, 173
49
145
101
1
127
101
159
119
127
131
127
49
77
145
61
1
159
77
87
101
15
87
159
67
103
1
kompos
kultur mata tunas
Leucaena leucochepala
Litsea cubeba
Lophostemon suaveolens
Mancozeb
manglid
materi genetik
media tanam
modal sosial
naungan
penyimpanan
pemberdayaan masyarakat
pemuliaan
perbanyakan vegetatif
perkecambahan
pertumbuhan
petani penggarap
pohon induk
produktivitas
pupuk
rotary shaker
sekam
setek
setek pucuk
Shorea seminis (de Vriese) Sloot
sumber benih bersertifikat
Syzygium cumini
Trema orientalis L.
Trichoderma sp.
tunas pangkasan
variasi
viabilitas benih
zat pengatur tumbuh
41
1
15
103
67
41
87
15
67, 103, 119
61
103
113
61
15
119
173
103, 173
61
77
87
67
1
41
77
31
113
159
145
31
41
77
15
113
31

INDEX PENULIS VOLUME 7
Aam Aminah
Agus Astho Pramono
Aris Sudomo
Asep Rohandi
Budi Leksono
Danu
Dede J. Sudrajat
Desmiwati
Dida Syamsuwida
Dwi Kartikaningtyas
Endah Retno Palupi
Eni Widajati
Evayusvita Rustam
Ganik Jawak
Kun Estri Maharani
Kurniawati Purwaka Putri
Maman Turjaman
Mirna Aulia Pribadi
Muhammad Zanzibar
127
159
145
87
1
31, 127
31, 87, 173
61
159
67
131
131
173
131
61
77, 101, 119, 159
145
61
113
Naning Yuniarti
Nur Hidayati
Nurmawati Siregar
Nutrita Toruan Mathius
Rahmad Suhartanto
Ratna Uli Damayanti S
Retno Agustarini
Rina Laksmi Hendrati
Surip
Tati Suharti
Tatiek K. Suharsi
Teguh Setyaji
Toni Herawan
Y.M.M. Anita Nugraheni
Yetti Heryati
Yulianti Bramasto
49, 113
15
31, 77
131
173
113
103
15
67
49, 101
173
67
1
119
103
49, 101, 127

JUDUL
Penulis
© 2017 BPTPTH All rights reserved. Open access under CC BY-NC-SA license.doi: //doi.org/10.20886/bptpth.2017.5.2.95-102 95
Times New Roman 9,
italic, huruf kecil
Times New Roman 8,
Bold, huruf kapital
JUDUL
( Title)
Penulis Pertama1, Penulis Kedua2, dan/and Penulis Ketiga3
1)Institusi asal penulis 2)Institusi asal penulis 3)Institusi asal penulis
Alamat; Telp/Fax, Kota, Negara e-mail: salah satu penulis sebagai koresponden
Naskah masuk: ....; Naskah direvisi: ...........; Naskah diterima: ..........(diisi oleh sekretariat redaksi)
ABSTRACT
Abstract should be written in Indonesia and English using Time New Roman font, size 11 pt, italic, single space. Abstract is not a merger of several paragraphs, but it is a full and complete summary that describe
content of the paper it should contain background, objective, paragraph and should be no more than 200 words
in English.
Keyword: 3-5 keywords
ABSTRAK
Abstrak ditulis dalam bahasa Indonesia dan bahasa Inggris dengan jenis huruf Times New Roman, ukuran
11 pt, spasi tunggal. Abstrak bukanlah penggabungan beberapa paragraf, tetapi merupakan ringkasan yang
utuh dan lengkap yang menggambarkan isi tulisan. Sebaiknya abstrak mencakup latar belakang, tujuan,
metode, hasil, serta kesimpulan dari penelitian. Abstrak tidak berisi acuan atau tidak menampilkan
persamaan matematika dan singkatan yang tidak umum. Abstrak terdiri dari satu paragraf dengan jumlah kata paling banyak 250 kata dalam bahasa Indonesia dan bahasa Inggris.
Kata kunci : 3-5 kata kunci
I. PENDAHULUAN
Pendahuluan mencakup hal-hal berikut ini: Latar Belakang, berisi uraian permasalahan dan
alasan pentingnya masalah tersebut diteliti. Permasalahan dirumuskan secara jelas, penjelasan
ditekankan pada rencana pemecahan masalah dan keterkaitan dengan pencapaian luaran yang
telah ditetepkan. Tujuan, berisi pernyataan secara jelas dan singkat tentang hasil yang ingin
dicapai dari serangkaian kegiatan penelitian yang akan dilakukan. Sasaran atau luaran
menjelaskan secara spesifik yang merupakan hasil antara dalam rangka mencapai tujuan penelitian.
Hasil yang dicapai, dijelaskan kaitannya dengan kegiatan yang dilaksanakan (khusus untuk
kegiatan penelitian lanjutan).
Kosong satu spasi tunggal
Kosong 2 (dua) spasi tunggal
Kosong satu spasi tunggal
Kosong 1 (satu )spasi tunggal
Kosong 1 (satu) spasi tunggal
Kosong 1 (satu) spasi tunggal
Kosong 2 (dua) spasi tunggal
Kosong 1 (satu) spasi tunggal
Kosong 1 (satu) spasi tunggal
Kosong 2 (dua) spasi tunggal
Commented [U1]: Times New Roman 12, bold, centered, huruf
kapital, spasi tunggal, maksimum dua baris, ≤ 15kata
Commented [U2]: Times New Roman 12, italic, centered, huruf
kecil diawali huruf kapital tiap kata, spasi tunggal,tanda buka dan
tutup kurung
Commented [U3]: Times New Roman 11, tegak, centered, huruf
kecil,spasi tunggal, tanda 1) 2) dst digunakan hanya jika penulis satu
dengan yang lainnya berbeda asal instasi, jika masih satu instansi
tidak perlu menggunakan tanda 1) 2) dst
Commented [U4]: Times New Roman11, huruf kapital,
italic,bold
Commented [U5]: Times New Roman 11, spasi tunggal, italic,
apabila ada nama ilmiah menjadi tegak & underline
Commented [U6]: Times New roman 11, bold, italic, urutkan
sesuai abjad
Commented [U7]: Times New Roman 11,huruf kapital,tegak,
bold
Commented [U8]: Times New Roman 11, bold, tegak, urutkan
sesuai abjad
Commented [U9]: Times New Roman 12, tegak, bold, centered
Commented [U10]: Times New Roman 12, spasi 1,5 fist line 0,75
cm

Jurnal Perbenihan Tanaman Hutan Vol.5 No 2 Desember 2017: 95-102
p-ISSN : 2354-8568
e-ISSN : 2527-6565
96
II. BAHAN DAN METODE
Metode Penelitian yang digunakan harus ditulis sesuai dengan cara ilmiah, yaitu rasional,
empiris dan sistematis. Tanaman dan binatang ditulis lengkap dengan nama ilmiah.
Menggunakan tolak ukur internasional, system matrix dan standar nomenklatur. Metode
penelitian dijelaskan sesuai dengan penelitian yang dilaksanakan. Jika metode merupakan kutipan
harus dicantumkan dalam referansi. Jika dilakukan perubahan terhadap metode kutipan atau
standar harus disebutkan perubahannya. Bila diperlukan dengan disajikan dalam tabel..
Untuk Bab dan Sub Bab secara konsisten ditulis rata di batas kiri tulisan, sebagaimana berikut:
A. Bahan dan Alat
Mengemukakan semua bahan yang digunakan seperti tumbuhan kayu, bahan kimia, alat dan
lokasi penelitian, waktu penelitian.
B. Prosedur Penelitian
Mengemukakan tahapan kerja dan beberapa pengujian yang dilakukan. Pelaksanaan penelitian
disusun berurutan menurut waktu, ukuran dan kepentingan. Untuk Sub Sub Bab secara konsisten
ditulis rata di batas kiri tulisan, sebagaimana berikut:
1. Penyiapan contoh kerja
2. Pengujian perkecambahan benih standar di laboratorium
Untuk Sub Sub Sub Bab secara konsisten ditulis, sebagaimana berikut:
a. Indeks perkecambahan (Gi)
b. Uji tetrazolium
C. Analisis Data
Metode statistik (bila ada) harus disebutkan dengan singkat.
III. HASIL DAN PEMBAHASAN
A. Hasil
Hasil disajikan dalam bentuk uraian umum. Disusun secara berurutan sesuai dengan tujuan
penelitian. Jika tujuan penelitian tidak tercapai perlu dikemukakan alasan dan penyebabnya.
Tabulasi, grafik, analisis statistik dilengkapi dengan tafsiran yang benar. Judul, keterangan tabel
dan gambar dilengkapi dengan terjemahan bahasa Inggris dengan huruf miring atau sebaliknya.
Angka yang tercantum dalam tabel tidak perlu diuraikan lagi, tetapi cukup dikemukakan makna
atau tafsiran masalah yang diteliti; dalam bagian ini juga dapat disajikan ilustrasi dalam bentuk
grafik bagan, pictogram dan sebagainya. Dapat mengemukakan perbandingan hasil yang
Kosong 2 (dua) spasi tunggal
Commented [U11]: Times New Roman12, tegak bold, centered
Commented [U12]: Times New Roman 12, spasi 1,5 fist line 0,75
cm
Commented [U13]: Times New Roman 12, bold, huruf kecil,
awal huruf besar kecuali kata hubung
Commented [U14]: Times New Roman 12, huruf kecil semua,
hanya awal kalimat huruf besar, bold
Commented [U15]: Times New Roman 12, huruf kecil semua,
hanya awal kalimat huruf besar

JUDUL Penulis
97
berlainan dan beberapa perlakuan. Metode statistik yang digunakan dalam pengolahan data harus
dikemukakan, sehingga tingkat kebenaran harus dapat ditelusuri. Prinsip dasar metode
harus diterangkan dengan mengacu pada referensi atau keterangan lain mengenai masalah ini.
Penulis mengemukakan pendapatnya secara objektif dengan dilengkapi data kuantitatif.
TABEL: Diberi nomor, judul, dan keterangan yang diperlukan, ditulis dalam bahasa Indonesia
dan Inggris. Tabel ditulis dengan Times New Roman ukuran 12 pt dan berjarak satu spasi di
bawah judul tabel. Judul tabel ditulis dengan huruf berukuran 12 pt, rata kiri dan ditempatkan di
atas tabel. Penomoran tabel menggunakan angka (1, 2, ......). Apabila tabel memiliki lajur/kolom
cukup banyak, dapat digunakan format satu kolom atau satu halaman penuh. Apabila judul pada
lajur tabel terlalu panjang, maka lajur diberi nomor dan keterangannya di bawah tabel.
Keterangan (Remarks) dan sumber (Source) ditulis di kiri bawah tabel ditulis dengan Times New
Roman ukuran 10 pt dan berjarak satu spasi. Tabel diletakkan segera setelah disebutkan dalam
naskah.
GAMBAR: Gambar, grafik, dan ilustrasi lain yang berupa gambar harus berwarna kontras (hitam
putih atau arsir), masing-masing harus diberi nomor, judul dan keterangan yang jelas dalam
bahasa Indonesia dan Inggris. Gambar diletakkan pada posisi paling atas atau paling bawah dari
setiap halaman. Gambar diletakkan simetris dalam kolom. Apabila gambar cukup besar, bisa
digunakan format satu kolom. Penomoran gambar menggunakan huruf Times New Roman ukuran
12 pt dan berjarak satu spasi rata kiri dan ditempatkan di bagian bawah, seperti pada contoh di
bawah. Gambar diletakkan segera setelah disebutkan dalam naskah.
Tabel (Table) 1. Hasil uji beda Duncan pengaruh lama pengeringan terhadap kadar air polong,
kadar air benih dan daya berkecambah sengon laut (The results of Duncan test of
the effect drying treatment on the pod, seed moisture content and germination
percentage of sengon laut)
Perlakuan pengeringan(Drying treatment)
Kadar air
polong(Pod moisture content
%
Kadar air
benih(Seed moisture content)
%
Daya
berkecambah/%
(Germination
percentage) %
Kontrol (Control) (7,76 + 0,13 )a (7,93 + 0,36) a (77,00+4,36) Penjemuran 1 hari (Sun drying for 1 day) (7,78+0,65) a (7,95 + 0,29) a (78,66+ 4,16)
Penjemuran 2 hari (Sun drying for 2 days) (5,72+ 0,69) b (5,98 + 0,10) b (73,66 + 1,53)
Penjemuran 3 hari (Sun drying for 3 days) (5,52 + 0,47 ) b (5,77 + 0,09) b (76,00 + 1,00)
Penjemuran 4 hari (Sun drying for 4 days) (5,53+0,19) b (5,77 + 0,19) b (77,33+2,31) Penjemuran 5 hari (Sun drying for 5 days) (5,53 + 0,23) b (5,59 + 0,51) b (78,66+ 1,53)
Alat pengering 4 jam (Seed drier for 4 hours) (7,70+ 0,09) a (7,94 + 0,05) a (76,33+9,07)
Alat pengering 8 jam (Seed drier for 8 hours) (7,43 + 0,35) a (7,52 +0,03) a (81,66+3,21)

Jurnal Perbenihan Tanaman Hutan Vol.5 No 2 Desember 2017: 95-102
p-ISSN : 2354-8568
e-ISSN : 2527-6565
98
Alat pengering 12 jam (Seed drierfor 12 hours) (7,46+ 0,14) a (7,56 + 0,11) a (74,44+ 3,21)
Alat pengering 16 jam (Seed drier for 16 hours) (7,47 + 0,41) a (7,49+ 0,29) a (79,00+ 6,24)
Alat pengering 20 jam (Seed drier for 20 hours) (7,51+ 0,44) a (7,56 + 0,12) a (77,66 + 6,81)
Alat pengering 24 jam (Seed drier for 24 hours) (7,52 + 0,42) a (7,53 + 0,21) a (78,00+ 7,21) Alat pengering 28 jam (Seed drier for 28 hours) (7,44 + 0,11) a (7,59 + 0,08) a (77,33+ 4,04)
Alat pengering 32 jam (Seed drier for 32 hours) (5,73 + 0,38) b (6,04 +0,03) b (78,66+ 4,62)
Alat pengering 36 jam (Seed drier for 36 hours) (5,79+ 0,16) b (6,05+ 0,07) b (78,66+ 3,06)
Alat pengering 40 jam (Seed drier for 40 hours) (5,75+ 0,43) b (6,11+0,14) b (77,00+ 1,53)
Alat pengering 44 jam (Seed drier for 44 hours) (5,77+ 0,45) b (6,07 + 0,09) b (77,66+ 4,62) Alat pengering 48 jam (Seed drier for 48 hours) (5,74+ 0,69) b (6,09 +0,11) b (76,66+ 9,07)
Rata-rata (average) 6,70 6,79 77,50
SD 0,95 0,89 4,44
Nilai F hitung (F test) 25,48** 74,14** 0,39
Keterangan (Remarks): Nilai-nilai yang diikuti oleh huruf yang sama pada kolom yang sama tidak berbeda nyata pada
selang kepercayaan 99% (Values followed by the same letters on the same colm are not
significantly different : a > b > c < d, etc.P = 99%). ** berbeda sangat nyata pada selang
kepercayaan 99% (significant effect, P = 99%)
Gambar (Figure) 1. Pengaruh skarifikasi benih terhadap waktu berkecambah, kecepatan
berkecambah dan daya berkecambah pada benih kayu kuku (Effect of seed scarification to
germination time, germination speed and germination rate of P. mooniana seeds)
B. Pembahasan
Pembahasan dapat menjawab apa arti hasil yang dicapai dan apa implikasinya. Dapat
menafsirkan hasil dan menjabarkannya, sehingga dapat dimengerti pembaca. Mengemukakan
hubungan dengan hasil penelitian sebelumnya. Bila berbeda tunjukkan, bahas dan jelaskan
penyebab perbedaan tersebut. Hasil penelitian ditafsirkan dan dihubungkan dengan hipotesis dan
tujuan penelitian. Mengemukakan fakta yang ditemukan dan alasan mengapa hal tersebut terjadi.
Menjelaskan kemajuan penelitian dan kemungkinan pengembangan selanjutnya. Simbol/lambang
ditulis dengan jelas dan konsisten. Istilah asing ditulis dengan huruf italic. Singkatan harus
dituliskan secara lengkap pada saat disebutkan pertama kali, setelah itu dapat ditulis kata
Kosong 1 (satu) spasi tunggal

JUDUL Penulis
99
singkatannya.
Apabila terdapat persamaan reaksi atau matematis, diletakkan simestris pada kolom.
Nomor persamaan diletakkan diujung kanan dalam tanda kurung dan penomoran dilakukan secara
berurutan. Apabila terdapat rangkaian persamaan yang lebih dari satu baris, maka penulisan
nomor diletakkan pada baris terakhir. Penunjukan persamaan dalam naskah dalam bentuk
singkatan, seperti persamaan berikut.
).................................................(1)
Keterangan:
Gt = persen kecambah hari ke-n
Tt = hari uji perkecambahan
IV. KESIMPULAN
Kesimpulan memuat hasil yang telah dibahas. Hal yang perlu diperhatikan adalah segitiga
konsistensi (masalah-tujuan-kesimpulan harus konsisten). Saran dapat dikemukakan untuk
dipertimbangkan pembaca.
UCAPAN TERIMA KASIH
Merupakan bagian yang wajib ada dalam sistematika karya tulis ilmiah. Suatu penelitian
tidak akan berhasil tanpa melibatkan pihak- pihak yang telah membantu, baik berperan secara
finansial, teknis, maupun substantif. Ucapan terima kasih merupakan sebuah kewajiban, bukan
pilihan (opsional).
DAFTAR PUSTAKA
Daftar Pustaka merupakan referensi yang dirujuk dalam naskah. Format penulisan Daftar
Pustaka mengacu pada American Psychological Association (APA) style. Referensi terdiri dari
acuan primer dan/atau acuan skunder. Sumber acuan primer adalah sumber acuan yang langsung
merujuk pada bidang ilmiah tertentu, sesuai topik penelitian dan sudah teruji. Sumber acuan primer
dapat berupa: tulisan dalam makalah ilmiah dalam jurnal internasional maupun nasional
terakreditasi, hasil penelitian di dalam disertai, tesis, maupun skripsi. Buku (textbook), termasuk
dalam sumber acuan sekunder. Semua karya yang dikutip dalam penulisan karya tulis harus
dimuat dalam daftar pustaka (dan sebaliknya).
Pustaka minimal 15,80% dari pustaka merupakan acuan primer, dan 80% dari acuan primer
Commented [A16]: Times New Roman12, spasi 1

Jurnal Perbenihan Tanaman Hutan Vol.5 No 2 Desember 2017: 95-102
p-ISSN : 2354-8568
e-ISSN : 2527-6565
100
merupakan publikasi 10 tahun terakhir. Pengelolaan pustaka dalam Jurnal Perbenihan Tanaman
Hutan menggunakan aplikasi software Mendeley, untuk itu disarankan agar penulis menggunakan
software yang sama. Jarak antar pustaka (after spacing) adalah 6 pt. Inden (hanging) pada baris
kedua dengan jarak 0,75 cm. Daftar pustaka harus disusun berdasarkan alphabet nama
pengarang. Penulisan situasi dan daftar pustaka diharuskan menggunakan aplikasi referensi
seperti Mendeley. Contoh Penulisan Daftar Pustaka Berdasarkan APA style:
1. Paper dalam jurnal
a. Artikel dalam jurnal ilmiah dengan volume dan nomor (1 penulis)
Bonner, F. T. (1998). Testing tree seeds for vigor: A review. Seed Tehcnology, 20(1), 5–17.
b. Artikel dalam jurnal ilmiah dengan volume dan nomor (2-6 penulis)
Vieira, R. D., Paiva, A. J. A., & Perecin, D. (1999). Electrical conductivity and field performance of soybean seeds. Seed Technology, 21, 15–24.
2. Buku
Chakraverty, A., & Singh, R. (2001). Postharvest Technology Cereals, Pulses, Fruit, and Vegetables. New
Hampshire (US): Science Publishers, Inc.
3. Prosiding
Gill, N. S., & Delouche, J. (1973). Proceedings of the Association of Official Seed Analysts 63. In
Deterioration of seed corn during storage. (pp. 35–50).
4. Makalah Seminar dan Lokakarya
DBPTH. (2014). Lokakarya penyusunan Standar Mutu Benih dan Mutu Bibit Tanarnan Hutan. In Kebijakan
pengujian benih. Solo, 4-7 November 2014: Direktorat Bina Perbenihan Tanaman Hutan. Jakarta.
5. Skripsi, tesis dan disertasi
Sudrajat, D. J. (2014). Keragaman populasi, uji provenansi dan adaptasi jabon (Neolamarckia cadamba
(Roxb.) Bosser). Disertasi. Sekolah Pascasrjana. Bogor: Insitut Pertanian Bogor.
6. Laporan penelitian
Aminah, A., & Budiman, B. (2009). Teknik penanganan benih kranji (Pongamia pinnata) sebagai sumber
energi terbarukan. Laporan Penelitian Balai Penelitian Teknologi Perbenihan Tanaman Hutan. Bogor (ID): Kementerian Kehutanan.
7. Artikel dari internet
Graham, P., Reedman, L., Rodriguez, L., Raison, J., Braid, A., Haritos, V., Adams, P. (2011). Sustainable
aviation fuels road map: Data assumptions and modelling, (May), 1–104. Retrieved from
http://www.csiro.au/en/Outcomes/Energy/Powering-Transport/Sustainable-Aviation-Fuels.aspx#
CATATAN:
1. Petunjuk penulisan ini dibuat untuk keseragaman format penulisan dan kemudahan bagi penulis dapat diakses di http://ejournal.forda-mof.org/ejournal-litbang/index.php/BPTPTH.
2. Naskah ditulis dalam format kertas berukuran A4 (210 mmx 297 mm) dengan margin atas 2,5 cm,
margin bawah 2,5 cm, margin kiri dan kanan masing-masing 2 cm. Bentuk naskah berupa 2 kolom
dengan jarak antar kolom 1 cm. Panjang naskah hendaknya maksimal 12 halaman, termasuk
lampiran Times New Roman, font 12, kecuali Abstrak, kata kunci dan daftar Pustaka font 11.
Pengutipan pustaka di dalam naskah berdasarkan sistem penulisan referensi APA Style,
sebagai berikut :

JUDUL Penulis
101
· Karya dengan dua pengarang.
....seperti yang dilakukan oleh Gill dan Delouche (1973)..... atau (Gill & Delouche, 1973)
· Karya tiga sampai lima pengarang.
(Kernis, Cornel, Sun, Berry, & Harlow, 1993) atau Kernis, Cornel, Sun, Berry, & Harlow (1993) menjelaskan... Dalam kutipan berikutnya, (Kernis et al., 1993) atau Kernis et al. (1993) argued...
· Enam pengarang atau lebih.
Harris et al. (2001) mengasumsikan... atau (Harris et al., 2001)
3. Penggunaan titik dan koma dalam penulisan angka : Naskah (teks) bahasa Indonesia: titik (.) menunjukkan kelipatan ribuan dan koma (,) menunjukkan pecahan.
4. Dewan Redaksi berhak mengubah naskah tanpa mengurangi isi yang terkandung di dalamnya dan juga berhak menolak naskah yang dianggap tidak memenuhi ketentuan yang disyaratkan. Penulis wajib Authorship Ethical Statement dan Copyright Agreement Form.
