potensi trichoderma spp. sebagai agens hayati dalam ... · yang menyerang tanaman sengon benyamin...
TRANSCRIPT

1
POTENSI Trichoderma spp. SEBAGAI AGENS HAYATI DALAM
PENGENDALIAN Ganoderma sp. YANG MENYERANG
TANAMAN SENGON
BENYAMIN DENDANG
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2013

2
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis Potensi Trichoderma spp. Sebagai
Agens Hayati dalam Pengendalian Ganoderma sp. yang Menyerang Tanaman Sengon
adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan
dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal
atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah
disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut
Pertanian Bogor.
Bogor, Juli 2013
Benyamin Dendang
NIM 451090111

3
SUMMARY
BENYAMIN DENDANG. The potency of Trichoderma spp. as biological agent in
controling Ganoderma sp. atacking sengon plant. Under direction of ACHMAD, ELIS
NINA HERLIYANA, AND DARMONO TANIWIRYONO.
Sengon (Falcataria mollucana) is one of the forestry plant that widely
developed by the community of Indonesia farmers especially in Java Island.
Nowadays, sengon is widely planted with a monoculture system, therefore
emerging many serious problems in silvi-culture. One of the serious problems
was attacked by the pathogenic fungi, Ganoderma sp., which mainly caused root
rot diseases. This research was aimed to investigate the ability of Trichoderma
spp. in controlling the in vitro growth of Ganoderma sp. and to evaluate the
capability of T. harzianum in inhibiting the growth of Ganoderma sp. on sengon
timber substrate at greenhouse. This research was carried out in three
experimental stages as follow: stage 1: study the growth activity of Ganoderma
sp.; stage 2: study the inhibition activity of Trichoderma spp. towards the growth
of Ganoderma sp. and stage 3: control the growth of Ganoderma sp. on Sengon
timber substrate at greenhouse.
Our results showed that the growth rate of Ganoderma sp. was expert on
PDA and MEA medium each 12 days and 14 days incubation, respectively.
Afterwards, the petridish was full of the Ganoderma sp. mycelium after 14 days
incubation. In this study, the average value of the growth of Ganoderma sp. on
PDA medium (7.09 mm day-1
) was significantly differs to the growth of
Ganoderma sp. on MEA medium (5.41 mm day-1
). Besides of that, the fastest
growth of Trichoderma spp. was shown by T. pseudokoningii such as 38.14 mm
day-1
and significantly differs to T. viride, 27.71 mm day-1
and T. harzianum,
26.31 mm day-1
. Based on antagonistic test, T. harzianum exhibited the best
inhibition to Ganoderma sp. on PDA medium up to 74.19% and significantly
differ to T. pseudokoningii 59.37% and T. viride 41.36%. Similarly on MEA
medium treatment T. harzianum exhibited the best inhibition to Ganoderma sp. up
to 73.00% and significantly differ to T. pseudokoningii 60.22% and T. viride
51.58%.
The inhibition growth of Ganoderma sp. on sengon timber substrate at
greenhouse showed that after 1 week, there was a decreasing growth of
Ganoderma sp. mycelium when the sengon timber was submerged (Xt1) and
obtained the highest value of inhibition up to 1.12 cm on sterile soil treatment + T.
harzianum, whilst on sterile soil treatment without T. harzianum, there was an
increasing growth of Ganoderma sp. mycelium up to 0.67 cm. Likewise, after 2
weeks of sengon timber submersion (Xt2) on sterile soil treatment + T. harzianum
there was a decreasing growth of Ganoderma’s mycelium up to 2.41 cm, whereas
on sterile soil treatment without T. harzianum, there was an increasing growth of
Ganoderma sp. mycelium up to 1.22 cm.
Keywords: biological agent, Ganoderma sp., sengon, Trichoderma spp.

4
RINGKASAN
BENYAMIN DENDANG. Potensi Trichoderma spp. Sebagai Agens Hayati dalam
Pengendalian Ganoderma sp. yang Menyerang Tanaman Sengon. Dibimbing oleh
ACHMAD, ELIS NINA HERLIYANA, DAN DARMONO TANIWIRYONO.
Sengon (Falcataria mollucana) merupakan tanaman hutan rakyat yang banyak
dikembangkan oleh petani hutan rakyat Indonesia, terutama di Pulau Jawa. Dewasa ini
penanaman sengon lebih banyak ditanam secara monokultur sehingga menimbulkan
berbagai permasalahan yang serius. Salah satu masalah yang muncul adalah serangan
penyakit Ganoderma sp. yaitu cendawan patogen yang dapat menyebabkan penyakit
busuk akar. Tujuan penelitian ini yaitu untuk mempelajari kemampuan Trichoderma spp.
dalam menghambat pertumbuhan Ganoderma sp. secara in vitro dan mengevaluasi
kemampuan Trichoderma spp. untuk mengendalikan Ganoderma sp. pada substrat balok
kayu sengon di rumah kaca. Penelitian ini dilakukan dengan tiga tahapan percobaan.
Penelitian I, studi pertumbuhan Ganoderma sp. dan Trichoderma spp. secara in vitro.
Penelitian II, uji penghambatan Trichoderma spp. terhadap pertumbuhan Ganoderma sp.
Penelitian III, pengendalian Ganoderma sp. pada balok kayu sengon di Rumah Kaca.
Hasil penelitian studi pertumbuhan diameter koloni Ganoderma sp. menunjukkan bahwa
laju pertumbuhan koloni Ganoderma sp. tercepat pada media PDA selama 12 hari dan
pada media MEA selama 14 hari cawan Petri terisi penuh dengan koloni. Rata-rata
pertumbuhan koloni isolat Ganoderma sp. sebesar 7.09 mm hari-1
pada media PDA nyata
lebih cepat dibanding pertumbuhan koloni isolat Ganoderma sp. pada media MEA
sebesar 5.41 mm hari-1
. Hasil penelitian studi pertumbuhan diameter koloni Trichoderma
spp. menunjukkan bahwa laju pertumbuhan diameter koloni Trichoderma spp. secara
berturut-turut tertinggi pada T. pseudokoningii sebesar 38.14 mm hari-1
nyata lebih cepat
dibanding T. viride sebesar 27.71 mm hari-1
dan T. harzianum 26.31 mm hari-1
. Uji
penghambatan Trichoderma spp. terhadap pertumbuhan Ganoderma sp. pada media PDA
diperoleh rata-rata persentase penghambatan terbaik pada T. harzianum sebesar 74.19%
nyata lebih cepat dibanding T. pseudokoningii sebesar 59,37% dan T. viride sebesar
41.36%. Demikian halnya pada media MEA diperoleh rata-rata persentase penghambatan
terbaik pada perlakuan T. harzianum (73.00%), berbeda nyata dengan T. pseudokoningii
(60.22%), dan T. viride (51.58%).
Hasil penelitian pengendalian Ganoderma sp. pada balok kayu sengon di rumah kaca
diperoleh penurunan pertumbuhan panjang koloni Ganoderma sp. satu minggu setelah
balok kayu dibenam (Xt1) diperoleh nilai penurunan panjang koloni tertinggi pada
perlakuan tanah steril + T. harzianum (1.12 cm), sedang pada perlakuan tanah steril tanpa
T. harzianum terjadi pertambahan panjang koloni sebesar 0.67 cm. Demikian halnya pada
dua minggu setelah balok kayu di benam (Xt2) pada perlakuan tanah steril + T. harzianum
terjadi penurunan panjang koloni tertinggi sebesar 2.41 cm, sedang pada perlakuan tanah
steril tanpa T. harzianum terjadi pertambahan panjang koloni Ganoderma sp. sebesar 1.22
cm.
Kata kunci : agensia hayati, Ganoderma sp., sengon, Trichoderma spp.

5
© Hak Cipta Milik IPB, Tahun 2013
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau
menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian,
penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu
masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis ini dalam
bentuk apa pun tanpa izin IPB
POTENSI Trichoderma spp. SEBAGAI AGENS HAYATI DALAM PENGENDALIAN
Ganoderma sp. YANG MENYERANG
TANAMAN SENGON

6
Tesis
sebagai salah satu syarat untuk memperoleh gelar
Magister Sains
pada
Program Studi Silvikultur Tropika
BENYAMIN DENDANG
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2013
POTENSI Trichoderma spp. SEBAGAI AGENS HAYATI DALAM PENGENDALIAN Ganoderma sp. YANG MENYERANG
TANAMAN SENGON

7
Penguji Luar Komisi pada Ujian Tesis: Dr Ir Bonny Purnama Wahyu Soekarno, MS

-
Judul Tesis : Potensi Trichoderma spp Sebagai Agens Hayati dalam Pengendalian Ganoderma sp. yang Menyerang Tanaman Sengon
Nama : Benyamin Dendang NIM : E451090111
Disetujui oleh
Dr Ir Achmad, MS Ketua
Dr Ir Elis Nina Herliyana, MSi Dr It Dannono Taniwiryono, MSc Anggota Anggota
Diketahui oleh
Ketua Program Studi Silvikultur Tropika
,
Dr Ir Basuki Wasis, MS
Tanggal Ujian : 28 Juni 2013 Tanggal Lulus: L 9 JUL LU 13
'"

8
Judul Tesis : Potensi Trichoderma spp. Sebagai Agens Hayati dalam Pengendalian
Ganoderma sp. yang Menyerang Tanaman Sengon
Nama : Benyamin Dendang
NIM : E451090111
Disetujui oleh
Komisi Pembimbing
Dr Ir Achmad, MS
Ketua
Dr Ir Elis Nina Herliyana, MSi
Anggota
Dr Ir Darmono Taniwiryono, MSc
Anggota
Diketahui oleh
.
Ketua Program Studi
Silvikultur Tropika
Dr Ir Basuki Wasis, MS
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
Tanggal Ujian : 28 Juni 2013 Tanggal Lulus :

9
PRAKATA
Puji dan syukur penulis panjatkan kepada Tuhan Yang Maha Kuasa atas
segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang
dipilih dalam penelitian yang dilaksanakan sejak bulan Januari 2011 sampai bulan
Juli 2012 ini dengan judul Potensi Trichoderma spp. Sebagai Agens Hayati Dalam
Pengendalian Ganoderma sp. yang Menyerang Tanaman Sengon.
Terima kasih penulis ucapkan kepada Bapak Dr Ir Achmad, MS, Ibu Dr Ir
Elis Nina Herliyana, MSi, dan Bapak Dr Ir Darmono Taniwiryono, MSc selaku
pembimbing. Di samping itu, penghargaan penulis sampaikan kepada Pusat Diklat
Kehutanan Kementrian Kehutanan yang telah memberikan waktu dan dana untuk
melaksanakan studi. terima kasih juga disampaikan kepada ayah, ibu, serta
seluruh keluarga, atas segala doa dan kasih sayangnya. Semoga karya ilmiah ini bermanfaat.
Bogor, Juli 2013
Benyamin Dendang

10
DAFTAR ISI
DAFTAR GAMBAR x
DAFTAR LAMPIRAN xi
PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
Manfaat Penelitian 2
TINJAUAN PUSTAKA 2 Sengon (F. moluccana) 2
Ganoderma sp. 4
Potensi Mikroorganisme Saprofitik Sebagai Agens Pengendali Hayati 5
Trichoderma spp. 5
METODE PENELITIAN 6
Tempat dan Waktu 6
Bahan dan Alat 6
Prosedur Penelitian 7
Percobaan I Pertumbuhan Diameter Koloni Ganoderma sp. 7
Pertumbuhan Diameter Koloni Trichoderma spp. 7
Percobaan II Uji penghambatan Trichoderma spp. 8
Percobaan III Pengendalian Ganoderma sp. pada balok kayu sengon di
Rumah Kaca 10
HASIL DAN PEMBAHASAN 12
Hasil 12
Pertumbuhan Diameter Koloni Ganoderma sp. 12
Pertumbuhan Diameter Koloni Trichoderma spp. 13
Penghambatan Trichoderma spp. terhadap Pertumbuhan Ganoderma sp. 15 Pengendalian Ganoderma sp. pada Balok Kayu Sengon di Rumah Kaca 17
Pembahasan 19
SIMPULAN DAN SARAN 23
Simpulan 23
Saran 23
DAFTAR PUSTAKA 24

11
DAFTAR GAMBAR
1 Metode biakan ganda Ganoderma sp. dan Trichoderma spp. pada cawan
konfrontasi berdiameter 9 cm 9
2 Pengukuran panjang koloni Ganoderma sp. pada substrat balok kayu
sengon 11
3 Pertumbuhan diameter koloni Ganoderma sp. pada media PDA dan
MEA 12
4 Rata-rata pertumbuhan diameter koloni Ganoderma sp. pada media PDA
dan MEA 12
5 Pertumbuhan diameter koloni Trichoderma spp. pada media PDA dan
MEA 14
6 Rata-rata pertumbuhan diameter koloni Trichoderma spp. pada media
PDA dan MEA 15
7 Penghambatan pertumbuhan diameter koloni Ganoderma sp. dengan
Trichoderma spp. pada media PDA dan MEA 16
8 Persentase penghambatan Trichoderma spp. terhadap pertumbuhan
diameter koloni Ganoderma sp. pada media PDA dan MEA 17
9 Pertumbuhan koloni Ganoderma sp. pada substrat balok kayu sengon di
rumah kaca 18
10 Rata-rata pertumbuhan panjang koloni Ganoderma sp. pada substrat
balok kayu sengon di rumah kaca 18
DAFTAR LAMPIRAN
1 Sidik ragam laju pertumbuhan diameter koloni Ganoderma sp. 28
2 Sidik ragam pertumbuhan diameter koloni Trichoderma spp. 28
3 Sidik ragam penghambatan Trichoderma spp. terhadap pertumbuhan diameter
koloni Ganoderma sp. 28
4 Sidik ragam panjang koloni Ganoderma sp. pada substrat balok kayu (Xt1) 28
5 Sidik ragam panjang koloni Ganoderma sp. pada substrat balok kayu (Xt2) 28
6 Komponen penyusun media PDA dan MEA 29

12
PENDAHULUAN
Latar Belakang
Sengon (Falcataria mollucana) merupakan salah satu jenis tanaman hutan
rakyat yang paling banyak dikembangkan oleh masyarakat dalam sistem
agroforestry di Indonesia. Hal ini disebabkan oleh karena pertumbuhannya yang
cepat, nilai ekonominya tinggi dan pemasaran yang relatif mudah. Secara umum,
di Jawa terdapat hutan rakyat seluas hampir 400 000 hektar dan mampu memasok
895 000 m3
kayu pertahunnya. Jumlah tersebut terdapat 10% serapan kayu
berbagai industri di pulau jawa. Pasokan kayu sengon pada hutan rakyat di pulau
Jawa sebesar 2.29% ha-1
tahun-1
(Mile 2003). Sengon menghasilkan kayu yang
berwarna terang, umumnya sebagai bahan baku veneer dan juga dipakai sebagai
bahan untuk kayu pertukangan.
Dengan semakin maraknya penanaman sengon yang ditanam dengan pola
monokultur, maka menimbulkan berbagai permasalahan yang serius. Salah satu
masalah yang muncul adalah adanya serangan penyakit Ganoderma sp. Serangan
Ganoderma sp. di lapangan sulit dideteksi karena gejala yang ditimbulkan mirip
dengan gejala serangan penyakit akar lainnya termasuk gejala kekeringan.
Meskipun tanaman sudah menunjukkan gejala sakit, namun tubuh buah
Ganoderma sp. kadang-kadang belum terbentuk. Di lain pihak, pada tanaman
yang tampak sehat dapat ditemukan tubuh buah Ganoderma sp. pada pangkal
batang. Kerusakan hutan tanaman industri A. mangium di Sumatera dan
Kalimantan yang diakibatkan oleh Ganoderma sp. mencapai 3-28% (Irianto et al.
2006).
Ganoderma sp. telah dilaporkan menyerang tanaman tanaman sengon dan
pohon jenis penaung pada tanaman kopi dan kakao di Jawa Barat dan Jawa Timur
(Herliyana et al. 2012). Munculnya serangan penyakit yang disebabkan cendawan
Ganoderma sp. meresahkan semua pengelolah hutan, terutama untuk hutan rakyat
karena sulit untuk dikendalikan. Ketika gejala dan tanda serangannya sudah parah,
maka tanaman tersebut sudah tidak mungkin diselamatkan lagi. Serangan
Ganoderma sp. dapat terjadi apabila ada interaksi antara akar dengan Ganoderma
sp.. Interaksi tersebut mengakibatkan tunggul hasil tebangan dapat menjadi
sumber penyakit. Tunggul yang terserang Ganoderma sp. tidak mampu untuk
bertunas. Segala upaya pengendalian harus ditujukan terutama untuk melindungi
tanaman yang sehat pada semua tingkat umur dan melindungi tunggul tanaman
pasca penebangan pohon dari penularan serangan Ganoderma sp.. Dalam upaya
pengendalian penyakit pada tanaman sengon para pengelola hutan termasuk petani
hutan rakyat lebih memilih menggunakan fungisida sintetik dengan alasan praktis,
mudah diperoleh dan hasilnya lebih cepat terlihat. Oleh karena dampak
penggunaan fungisida sintetik yang merusak lingkungan maka perlu dilakukan
upaya pengendalian yang ramah lingkungan. Pengendalian yang ramah
lingkungan dapat dilakukan dengan menggunakan agens hayati yaitu dengan
memanfaatkan cendawan yang bersifat antagonistik terhadap cendawan patogen.
Pengendalian Ganoderma sp. pada sengon pada saat ini masih ke arah
efektivitas dan ekonomis, belum ke arah kelestarian terhadap lingkungan.
Perlindungan tanaman sebaiknya berwawasan terhadap keanekaragaman hayati
dan lingkungan hidup. Konsep tersebut sesuai dengan UU No. 12 Tahun 1992,

13
pasal 22 ayat 1 menyebutkan bahwa setiap orang dan/atau badan hukum dilarang
menggunakan sarana dan atau cara yang dapat mengganggu kesehatan dan/atau
mengancam keselamatan manusia, menimbulkan gangguan dan kerusakan sumber
daya alam dan atau lingkungan hidup. Konsep perlindungan hutan juga dijabarkan
dalam PP No. 6 Tahun 1995 yang menyatakan agar setiap orang tidak
sembarangan menggunakan cara dan sarana melakukan perlindungan tanaman.
Ayat ini memprioritaskan cara-cara perlindungan tanaman non-kimiawi agar tidak
mengganggu kesehatan, merusak sumber daya alam dan membunuh agens hayati.
Pengendalian Ganoderma sp. yang mempertimbangkan kelestarian
lingkungan membutuhkan agens hayati yang berpotensi sebagai pengendali. Salah
satu agens hayati adalah Trichoderma spp., yang merupakan salah satu dari agens
hayati dalam mencari bahan pengganti pestisida kimia untuk mengendalikan
penyakit patogen tular tanah termasuk Ganoderma sp.. Trichoderma dilaporkan
mempunyai sifat antagonistik terhadap patogen tular tanah (Widyastuti. 1998;
Widyastuti et al. 1999).
Tujuan Penelitian
Tujuan dari penelitian ini yaitu :
1. Mempelajari kemampuan Trichoderma spp. dalam menghambat pertumbuhan
Ganoderma sp. secara in vitro.
2. Mengevaluasi kemampuan Trichoderma spp. untuk mengendalikan
Ganoderma sp. pada balok kayu sengon di rumah kaca.
Manfaat Penelitian
Hasil penelitian ini diharapkan dapat memberikan manfaat sebagai
informasi mengenai potensi Trichoderma spp. sebagai agensia hayati terhadap
pengendalian Ganoderma sp. pada kayu sengon sehingga dapat mengurangi
kerugian ekonomis yang diakibatkannya.
TINJAUAN PUSTAKA
Sengon (F. mollucana)
Sengon dalam bahasa latin disebut F. mollucana termasuk famili
Mimosaceae, keluarga peta-petaian. Di Indonesia, sengon memiliki beberapa
nama daerah seperti jeunjing (Sunda), kalbi, sengon landi, sengon laut, atau
sengon sabrang (Jawa). Maluku : seja (Ambon), sikat (Banda), tawa (Ternate),
dan gosui (Tidore). Sengon dapat mencapai tinggi sekitar 30-45 meter dengan
diameter batang sekitar 7080 cm. Bentuk batang bulat dan tidak berbanir. Kulit
luarnya berwarna putih atau kelabu, tidak beralur dan tidak mengelupas. Berat
jenis kayu rata-rata 0.33 dan termasuk kelas awet IV-V. Kayu sengon digunakan
untuk tiang bangunan rumah, papan peti kemas, peti kas, perabotan rumah tangga,
pagar, kotak korek api, veneer, kertas dan lain-lainnya (Martawijaya et al. 1989).

14
Tajuk sengon berbentuk menyerupai payung, rimbun, dan daun yang tidak
terlalu lebat. Daun sengon tersusun majemuk menyirip ganda dengan anak daun
yang kecil-kecil dan mudah rontok. Warna daun sengon hijau pupus, berfungsi
untuk menyerap energi dari cahaya dan sekaligus sebagai penyerap karbon
dioksida dari udara bebas. Sengon memiliki akar tunggang yang cukup kuat
menembus ke dalam tanah, akar rambutnya tidak terlalu besar, tidak rimbun dan
tidak menonjol ke permukaan tanah. Bintil akar berfungsi untuk menyimpan zat
nitrogen, oleh karena itu tanah di sekitar tanaman sengon menjadi subur. Bunga
sengon tersusun dalam malai berukuran panjang 12 mm, berwarna putih
kekuningan dan sedikit berbulu, berbentuk seperti saluran atau lonceng. Bunganya
biseksual, terdiri dari bunga jantan dan bunga betina. Buah sengon berbentuk
polong, pipih, tipis, tidak bersekat-sekat dan berukuran panjang 10-13 dan lebar 2
cm. Setiap polong buah berisi 15-20 biji. Biji sengon berbentuk pipih, lonjong,
tidak bersayap, berukuran panjang 6 mm, berwarna hijau ketika masih muda dan
berubah menjadi kuning sampai coklat kehitaman jika sudah tua, agak keras dan
berlilin (Soerianegara dan Lemmens 1993).
Sengon dapat tumbuh optimal pada ketinggian antara 0-800 m dpl.
Walapun demikian, sengon masih dapat tumbuh sampai ketinggian 1500 m di atas
permukaan laut. Sengon termasuk jenis tanaman tropis, sehingga memerlukan
suhu sekitar 18-27 °C untuk pertumbuhannya. Curah hujan mempunyai beberapa
fungsi untuk tanaman, diantaranya sebagai pelarut zat nutrisi, pembentuk gula dan
pati, sarana transportasi hara dalam tanaman, pertumbuhan sel dan pembentukan
enzim, serta menjaga stabilitas suhu. Sengon membutuhkan batas curah hujan
minimum yang sesuai, yaitu 15 hari hujan dalam 4 bulan terkering dan memiliki
curah hujan tahunan yang berkisar antara 2000-4000 mm dengan kelembaban
sekitar 50-75%. Di Jawa, sengon dilaporkan dapat tumbuh di berbagai jenis tanah
kecuali tanah grumusol (Charomaini dan Suhaendi 1997). Pada tanah latosol,
andosol,aluvial dan podzolik merah kuning, sengon tumbuh sangat cepat. Pada
tanah marginal, pupuk mungkin diperlukan pada awal petumbuhan, selanjutnya
pertumbuhan sengon akan lebih cepat karena kemampuan untuk mengikat
nitrogen meningkat. Sengon termasuk jenis pionir yang dapat tumbuh di hutan
primer, hutan hujan dataran rendah sekunder dan hutan pegunungan, padang
rumput dan di sepanjang pinggir jalan dekat laut. Di habitat alaminya di Papua,
sengon berasosiasi dengan Agathis labillardieri, Celtis spp., Diospyros spp.,
Pterocarpus indicus, Terminalia spp., dan Toona sureni (Soerianegara dan
Lemmens 1993).
Peningkatan produktivitas persatuan luas (peningkatan produksi secara
vertikal), merupakan faktor penting yang berdampak nyata terhadap peningkatan
pendapatan petani. Awang et al. (2002) mengemukakan bahwa, kajian tentang
sosial ekonomi hutan rakyat sudah banyak dilakukan oleh para ilmuwan dan
peneliti namun upaya yang berkaitan dengan peningkatan produktivitas belum
banyak tersedia. Hal ini disebabkan oleh upaya peningkatan produktivitas per
satuan luas belum dikembangkan melalui uji coba pengembangan teknologi tepat
guna di lapangan. Mile (2003) mengemukakan bahwa tanaman sengon yang
dikembangkan di hutan rakyat saat ini banyak yang tidak optimal
pertumbuhannya, sekalipun berada pada kondisi yang sesuai baik dari segi
kesesuaian tanah maupun dari segi kesesuaian agroklimat. Hal ini disebabkan oleh
beberapa faktor yaitu semakin menurunnya kesuburan tanah, munculnya berbagai

15
serangan hama dan penyakit dan kurangnya monitoring yang dilakukan oleh para
petani hutan rakyat.
Ganoderma sp.
Ganoderma sp. termasuk dalam kelas Basidiomycetes yang dapat
menyebabkan busuk akar pada berbagai jenis tanaman keras, melalui
kemampuannya dalam menghancurkan lignin, menghancurkan selulosa dan
berkaitan dengan pengaruhnya terhadap polisakarida (Hepting 1971; Blanchette
1984; Adaskaveg dan Ogawa 1990). Ganoderma sp. pertama kali dilaporkan oleh
Karsten (1881) dengan G. lucidum sebagai satu-satunya jenis.
Ganoderma sp. digolongkan ke dalam Ganodermataceae,
Aphylophorales, Basidiomycetes, dan Basidiomycotina. Ganoderma sp.
mempunyai tubuh buah yang berpori pada bagian bawahnya dengan bentuk,
ukuran, dan warna yang beragam. Ganoderma sp. dijumpai secara luas menyerang
tanaman inang berdaun lebar (Phillips dan Burdekin 1989), namun gejala
terjadinya busuk akar yang disebabkan oleh Ganoderma sp. telah ditemukan pada
tanaman akasia di Australia bagian utara, di Peninsular Malaysia dan Sumatera
Utara (Lee 1996). Penyakit busuk akar yang disebabkan oleh Ganoderma sp. telah
tercatat sebagai penyakit yang paling berbahaya menyerang A. mangium di
Bengal Barat, India (Sharma dan Florensce 1996). Di Indonesia, Ganoderma sp.
merupakan penyakit utama pada berbagai tanaman kehutanan termasuk sengon
(Basset dan Peters 2003; Salomon et al. 1993). Ganoderma sp. juga menyerang
tanaman kelapa sawit dengan tingkat kematian 50% (Turner 1981).
Gejala yang timbul akibat penyakit Ganoderma sp. adalah daun
menguning, kering dan akhirnya rontok (Semangun 2000). Sedangkan pada akar
terlihat adanya selaput miselium berwarna merah bata. Miselium yang baru
tumbuh umumnya berwarna putih krem dan warna merah yang khas apabila
miselium menjadi tua. Pada tingkatan serangan lebih lanjut, Ganoderma sp.
membentuk badan buah (basidiokarp) pada pangkal batang (Rahayu 1994).
Infeksi Ganoderma sp. diakibatkan oleh adanya kontak antara akar yang
sehat dengan akar yang sakit. Meskipun cendawan membentuk rizomorf, tetapi
rizomorfnya tidak dapat berjalan bebas dalam tanah terlepas dari alas
makanannya. Spora tidak dapat menginfeksi tanaman yang sehat, tetapi spora
dapat menginfeksi tunggul-tunggul yang segar dari tanaman yang rentan dan
dapat menjadi sumber infeksi baru. Sengon terinfeksi dari tunggul dan sisa-sisa
akar pohon atau pohon-pohon yang tua. Ganoderma sp. tumbuh dengan baik pada
medium yang mempunyai pH 6.0-7.0. dapat bertahan lebih lama pada akar
tanaman dan pada tonggak-tonggak kayu di dalam tanah dibanding dengan
cendawan patogen lainnya (Semangun 2000). Perkembangan Ganoderma sp.
berlangsung dalam waktu yang cukup lama dan biasanya lebih dari delapan tahun
(Allen 1994).

16
Potensi Mikroorganisme Saprofitik Sebagai Agensia Pengendali Hayati
Ganoderma sp.
Pengendalian hayati merupakan penurunan jumlah inokulum atau aktivitas
menghasilkan penyakit suatu patogen yang dilaksanakan dengan atau melalui satu
atau lebih mikroorganisme selain manusia (Cook dan Baker 1983). Pengendalian
hayati dapat dilaksanakan melalui praktek budidaya dan pemuliaan tanaman untuk
meningkatkan resistensi terhadap patogen atau kesesuaian tanaman inang untuk
aktivitas antagonis; melalui introduksi massal antagonis, ras nonpatogenik,
organisme atau agens berguna lainnya (Lewis dan Papavizas 1991).
Aktivitas penyakit termasuk didalamnya adalah pertumbuhan, virulensi,
dan agresifitas. Faktor lain dari patogen adalah termasuk 1) individu atau populasi
avirulen atau hipovirulen dari spesies patogen itu sendiri, 2) manipulasi genetik
tanaman inang, kultur teknis, atau dengan menggunakan mikroorganisme untuk
meningkatkan ketahanan tanaman inang terhadap patogen, dan 3) pemanfaatan
antagonis patogen yang diartikan sebagai mikroorganisme yang menginterfensi
pertahanan atau aktivitas produksi penyakit dari patogen. Pengendalian hayati
dapat berupa kultur teknis (pengelolaan habitat) sehingga membuat lingkungan
mendukung untuk pertumbuhan antagonis, penggunaan tanaman inang yang
resisten, atau keduanya, persilangan tanaman untuk meningkatkan ketahanan
terhadap patogen atau keadaan tanaman inang yang mendukung (disukai) untuk
aktivitas antagonis, introduksi antagonis, strain non-patogenik, dan agen atau
organisme lain yang mempunyai manfaat yang sama.
Salah satu contoh pengendalian hayati adalah dengan memanfaatkan
Trichoderma spp. sebagai organisme yang mempunyai kemampuan antagonistik
dalam mengendalikan penyakit tanaman. Trichoderma spp. merupakan cendawan
yang sangat umum dijumpai dalam tanah dan merupakan cendawan yang bersifat
antagonistik terhadap cendawan.
Trichoderma spp.
Trichoderma spp. merupakan cendawan inperfekti (tidak sempurna) dari
Subdivisio Deuteromycotina, Kelas Hyphomycetes, Ordo Moniliaceae.
Konodiofor tegak, bercabang banyak, agak berbentuk kerucut, dapat membentuk
klamidiospora, pada umumnya koloni dalam biakan tumbuh dengan cepat,
berwarna putih sampai hijau (Cook dan Baker 1989). Bentuk sempurna dari
cendawan ini secara umum dikenal sebagai Hipocreales atau kadang-kadang
Eurotiales, Clacipitales dan Spheriales. Morfologi beberapa spesies Trichoderma
menurut Cook dan Baker (1989) sebagai berikut: 1) Trichoderma viride,
konidiofor berakhir pada fialid, fialospora mempunyai dinding yang kasar,
berwarna hijau, berukuran antara 2.8-5.0 X 2.8-4.5 mm, dan koloni cepat tumbuh.
2) Trichoderma harzianum, konidiofor berakhir pada fialid, fialospora halus,
berwarna hijau, berukuran antara 2.4-3.2 X 2.2-2.8 mm, dan koloni cepat tumbuh.
3) Trichoderma pseudokoningii; konidiofor berakhir pada fialid, fialospora halus,
berwarna hijau, eliptik-silindris, berukuran 3-4.8 X 1.9-2.8 mm, dan koloni cepat
tumbuh.
Trichoderma spp. merupakan salah satu cendawan tanah yang dominan
yang bersifat saprotrof sehingga secara ekologis mampu berkompetisi dengan
cendawan yang lain dan mampu mengkolonisasi berbagai substrat yang ada di

17
hutan, sehingga Trichoderma spp. dapat dikembangkan sebagai agensia
pengendali hayati cendawan patogen tular tanah (Reese dan Mendels 1959; Hadar
et al. 1979; Elad et al. 1983). Trichoderma spp. menghasilkan 3 tipe propagul
yang dapat digunakan sebagai bahan formula, yaitu : hifa, klamidiospora dan
konidia (Papavizas 1985). Trichoderma spp. mempunyai daya antagonis yang
tinggi dan dapat menghasilkan racun, sehingga dapat menghambat dan mematikan
cendawan lain (Webster dan Dennis 1971).
Widyastuti et al. (2001) menyatakan bahwa isolat Trichoderma spp. telah
menghambat secara penuh miselia cendawan patogen skala in vitro. Daya hambat
Trichoderma spp. tersebut sebesar 91.13-93.49% dengan menggunakan
konsentrasi 103 dan 10
5 .
Mekanisme Trichoderma spp. sebagai agens hayati berlangsung setelah
konidianya tumbuh dan berkembang di sekitar perakaran tanaman yang berfungsi
sebagai mikoparasitik dan akan menekan populasi cendawan patogen yang ada
pada akar tanaman. Pengendalian penyakit yang disebabkan oleh cendawan
patogen dengan menggunakan cendawan Trichoderma spp. selain dapat menekan
pertumbuhan penyakit akar putih, juga diduga dapat mempengaruhi keragaman
serta kepadatan populasi cendawan tanah. Mekanisme pengendalian Trichoderma
spp. dengan cara membelit atau tumbuh disepanjang hifa inang dan membentuk
sruktur semacam kait yang membentuk penetrasi ke dalam dinding sel inang (Chet
et al. 2004).
METODE PENELITIAN
Tempat dan Waktu
Penelitian ini dilaksanakan pada bulan Januari 2011 sampai dengan bulan
Juli 2012 di Laboratorium Patologi Hutan dan di Rumah Kaca Ekologi,
Departemen Silvikultur Fakultas Kehutanan IPB.
Bahan dan Alat
Bahan yang digunakan adalah : Biakan Trichoderma harzianum yang
diperoleh dari Laboratorium Bioteknologi Perkebunan Bogor, Trichoderma viride
yang diperoleh dari Laboratorium Mikologi UGM, Trichoderma pseudokoningii
yang diperoleh dari Seameo Biotrop, tubuh buah Ganoderma sp. diperoleh dari
tunggul sengon di Desa Baregbeg Kabupaten Ciamis, aquades, media Potato
Dextrose Agar (PDA), media Malt Extact Agar (MEA), tanah steril, tanah tidak
steril, alkohol, spirtus, dan balok kayu sengon. Alat yang digunakan dalam
penelitian ini adalah : otoklaf, Laminar Air Flow, oven, inkubator, cawan Petri,
bunsen, sundip, kapas, timbangan analitik, mikroskop, polibag, kamera digital,
mistar, dan alat tulis.

18
Prosedur Penelitian
Penelitian dilakukan dengan 3 percobaan. Percobaan 1: Pertumbuhan
koloni Ganoderma sp., dan pertumbuhan koloni Trichoderma spp. Percobaan 2:
Uji penghambatan Trichoderma spp. terhadap pertumbuhan Ganoderma sp., dan
Percobaan 3: Pengendalian Ganoderma sp. pada balok kayu sengon di Rumah
Kaca.
Percobaan 1
Pertumbuhan Diameter Koloni Ganoderma sp.
Ganoderma sp. diisolasi dari tubuh buah yang diambil dari tunggul
sengon. Tubuh buah dipotong secara melintang menjadi beberapa bagian,
kemudian mengambil daging buah (konteks) untuk ditanam pada media PDA di
dalam cawan Petri yang berdiameter 9 mm (Steyaert 1967). Setiap cawan Petri
ditanam tiga potongan konteks. Setelah isolat tumbuh, maka dilanjutkan dengan
pemurnian, yaitu mengambil bagian dari isolat yang tidak terkontaminasi oleh
cendawan atau mikroorganisme lain, kemudian diisolasi kembali pada cawan
Petri. Selanjutnya biakan hasil pemurnian diisolasi pada dua macam media yaitu
media PDA dan MEA dengan menggunakan kok bor 6 mm dan diletakkan pada
bagian tengah cawan Petri.
Pengamatan dan Analisis Data
Pengambilan data dilakukan dengan mengukur pertumbuhan diameter
koloni isolat Ganoderma sp. sampai cawan Petri terisi penuh dan dilakukan setiap
hari. Penelitian ini dilakukan dengan menggunakan Rancangan Acak Lengkap
(RAL) dimana faktor perlakuannya adalah dua macam media yaitu media PDA
dan MEA dengan tiga kali ulangan. Model linier dari pertumbuhan diameter
koloni isolat Ganoderma sp. yaitu:
Yijk = μ + δi + εij
Keterangan :
Yij : Diameter koloni isolat Ganoderma sp. pada media ke-i, ulangan ke-j
μ : Nilai tengah pengamatan
δi : Pengaruh media ke-i
εij : Pengaruh galat percobaan perlakuan ke-i pada ulangan ke-j
Data kuantitatif dari diameter pertumbuhan koloni dianalisis dengan
menggunakan SAS 9.00. Apabila terdapat data yang signifikan akan dilanjutkan
dengan uji lanjut Duncan pada taraf kepercayaan 95%.
Pertumbuhan Diameter Koloni Trichoderma spp.
Percobaan ini dilakukan untuk mengetahui pertumbuhan diameter koloni
dari tiga jenis Trichoderma (T. harzianum, T. viride, dan T. pseudokoningii) pada
media PDA dan MEA. Pengujian masing-masing dilakukan pada ketiga isolat

19
Trichoderma yaitu dengan meletakkan potongan koloni isolat yang diambil dari
pada tengah cawan Petri berdiameter 9 cm yang berisi media PDA dan MEA
dengan menggunakan kok bor 6 mm Biakan tersebut diinkubasi pada suhu
ruangan sampai cawan Petri terisi penuh.
Pengamatan dan Analisis Data
Peubah yang diamati pada percobaan ini adalah mengukur pertumbuhan
diameter koloni masing-masing jenis Trichoderma sampai cawan Petri terisi
penuh dan pengukuran dilakukan setiap hari. Penelitian ini disusun dengan
menggunakan Rancangan Acak Lengkap (RAL) Faktorial dimana faktor
perlakuannya adalah T. harzianum, T. viride, T. pseudokoningii dan dua macam
media yaitu media PDA dan MEA. masing-masing perlakuan diulang sebanyak
tiga kali.
Model linier dari pertumbuhan koloni Trichoderma spp. adalah sebagai berikut:
Yijk = μ + αi + βj + (αβ)ij + εijk
Keterangan :
Yijk : Diameter koloni isolat jenis Trichoderma ke-i, media ke-j dan ulangan
ke-k
μ : Nilai tengah pengamatan
αi : Pengaruh jenis Trichoderma ke-i
βj : Pengaruh jenis media ke-j
(αβ)ij : Pengaruh interaksi jenis Trichoderma ke-i, media ke-j
εijk : Pengaruh galat pada jenis Trichoderma ke-i, media ke-j, dan ulangan ke-
k
Data kuantitatif dari diameter pertumbuhan koloni dianalisis dengan
menggunakan SAS 9.00. Apabila terdapat data yang signifikan akan dilanjutkan
dengan uji lanjut Duncan pada taraf kepercayaan 95%.
Percobaan 2
Uji Penghambatan Trichoderma spp. terhadap Pertumbuhan Ganoderma sp.
Penelitian ini dilakukan dengan biakan ganda (Coșkuntuna dan Özer
2007). Uji penghambatan dilakuan pada ketiga jenis Trichoderma dengan
menggunakan media PDA dan MEA. Petri (9 cm) diinokulasikan dengan biakan
Ganoderma sp. diameter 6 mm usia 10 hari. Setelah biakan Ganoderma sp.
berumur 5 hari, kemudian masing-masing biakan Trichoderma dengan ukuran
yang sama di kulturkan dari arah yang berlawanan (Gambar 1). Petri disimpan
dalam inkubator 23 0c. Kontrol yang digunakan adalah isolat tanpa Trichoderma
dengan ulangan sebanyak 3 kali. Perlakuan yang memberikan penghambatan
yang lebih bagus akan digunakan untuk penelitian selanjutnya di rumah kaca.

20
Gambar 1 Metode biakan ganda Ganoderma sp. dan Trichoderma spp. pada
cawan konfrontasi berdiameter 9 cm.
Untuk mengetahui daya hambat Trichoderma spp. digunakan rumus Fokkema
(1973) sebagai berikut:
I = a - b
x 100% a
Keterangan :
I = Persentase penghambatan
a = Jari-jari koloni patogen (Ganoderma sp.) menjauhi antagonis
(Trichoderma spp.)
b = Jari-jari koloni patogen ke arah antagonis
Pengamatan dan Analisis Data
Pengamatan dilakukan dengan mengukur pertumbuhan koloni ganoderma
sp. secara radial dan mengukur diameter perkembangan koloni tersebut pada
perlakuan kontrol (tanpa Trichoderma spp.). Data tersebut akan digunakan untuk
menghitung perkembangan miselium dan persen penghambat jenis Trichoderma
terhadap pertumbuhan diameter koloni Ganoderma sp.. Uji penghambatan
dilakukan dengan Rancangan Acak Lengkap Faktorial dengan dua faktor yaitu
faktor jenis cendawan (T. harzianum, T. viride, T. pseudokoningii) dan jenis
media (PDA dan MEA). Model linier dari uji penghambatan Trichoderma spp.
adalah sebagai berikut:
Yijk = µ + άi +βj + (άβ)ij + εijk
i, = 1,2,3,4 j = 1,2
Keterangan :
Yijk : Persentase penghambatan jenis Trichoderma untuk jenis ke-i, media ke-j,
dan ulangan ke-k
µ : Nilai tengah pengamatan
άi : Pengaruh jenis Trichoderma ke-i
βj : Pengaruh medi ke-j
(άβ)ij : Pengaruh interaksi jenis Trichoderma ke-i pada media ke- j
εijk : Pengaruh galat pada jenis Trichoderma ke-i, media ke-j, dan ulangan ke-
k
Trichoderma spp. Ganoderma sp.
b a

21
Data kuantitatif dari diameter pertumbuhan koloni dianalisis dengan
menggunakan SAS 9.00. Apabila terdapat data yang signifikan akan dilanjutkan
dengan uji lanjut Duncan pada taraf kepercayaan 95%.
Percobaan 3
Pengendalian Ganoderma sp. pada Balok Kayu Sengon di Rumah Kaca
Percobaan ini mengacu kepada metode yang dikembangkan oleh
Hadiwiyono (1996) yang terdiri dari tiga tahapan, yaitu :
a. Pembuatan Formula Trichoderma sp.
Formula Trichoderma sp. dilakukan dengan menggunakan media jagung +
dedak (1:1). Media jagung direbus terlebih dahulu selama 30 menit, kemudian
diaduk dengan dedak dan ditambahkan air secukupnya sampai media agak lengket
kemudian dimasukkan ke dalam botol. Selanjutnya media disterilisasi dengan
menggunakan otoklaf pada suhu 121 0C selama 30 menit. Media yang sudah steril
diinokulasikan dengan Trichoderma sp. yang memberikan hasil penghambatan
terbaik pada cawan Petri. Formula tersebut di inkubasi selama 12 hari
(Hadiwiyono 1996).
b. Persiapan Patogen Uji
Substrat yang digunakan adalah balok kayu sengon berukuran 3 x 2 x 10
cm. Balok kayu sengon di bungkus dengan plastik kemudian disterilisasi dengan
otoklaf pada suhu 121 0C selama 30 menit. Setelah disterilisasi, balok kayu
didinginkan dan segera diinokulasi dengan biakan Ganoderma sp. yang di
kulturkan di dalam toples. Balok kayu diletakkan secara tegak lurus pada
permukaan koloni Ganoderma sp. yang berumur 10 hari di dalam toples. Setiap
toples diberi 4 balok kayu, kemudian diinkubasi pada suhu ruangan selama 7 hari.
c. Pembenaman Balok Kayu pada Polibag di Rumah Kaca
Balok kayu yang telah diinokulasi dengan Ganoderma sp. dibenam pada
polibag yang telah diisi dengan media tanah ditambah formula Trichoderma sp.
dan media tanah tanpa formula Trichoderma sp. Tanah yang digunakan terdiri
dari dua macam, yaitu tanah steril dan tanah tidak steril. Tanah disterilisasi
dengan menggunakan otoklaf pada suhu 121 0C selama 30 menit. Perlakuan
formula Trichoderma sp. pada polibag dilakukan bersamaan dengan pembenaman
balok kayu dengan dosis 10 gram per polibag. Balok kayu dibenam secara tegak
lurus sedalam 3 cm dan masing-masing polibag dibenam 3 balok kayu. Setelah
balok kayu dibenam pada polibag, disungkup dengan plastik bening dan di simpan
di rumah kaca.
Pengamatan dan Analisi Data
Pengamatan dilakukan dengan mengukur panjang koloni Ganoderma sp.
pada permukaan balok kayu yaitu saat balok kayu dibenam (Xt0), 6 hari setelah
balok kayu di benam (Xt1), dan 12 hari setelah balok kayu di benam (Xt2).
Pengamatan laju pertumbuhan koloni Ganoderma sp. pada balok kayu
menggunakan rumus Van Der Plank (1963) sebagai berikut:

22
r = 2.3
log Xt
t X0
Keterangan :
r = Laju pertumbuhan patogen (unit-1
hari-1
)
Xt = Panjang koloni patogen pada balok kayu setelah waktu t
X0 = Panjang koloni patogen pada balok kayu saat t = 0
t = Waktu selang pengukuran Xt dan X0 (hari)
Gambar 2 Pengukuran panjang koloni Ganoderma sp. pada substrat balok kayu
sengon.
Penelitian pada balok kayu di rumah kaca disusun berdasarkan Rancangan
Acak Lengkap (RAL) 2 faktor dengan 3 ulangan. Faktor pertama (A) adalah tanah
dengan dua taraf yaitu A0 (tanpa sterilisasi tanah) dan A1 (sterilisasi tanah).
Faktor kedua (B) adalah aplikasi Trichoderma sp. dengan dua taraf yaitu B0
(tanpa Trichoderma) dan B1 (dengan Trichoderma sp.). Model statistik pada
penelitian ini adalah Model Faktorial RAL, sebagai berikut :
Yijk = μ + άi +βj + (άβ)ij + εijk
Keterangan :
Yijk : Nilai pengamatan peubah yang dipengaruhi oleh sterilisasi tanah ke-i,
Trichoderma sp. ke-j, dan ulangan ke-k
μ : Rataan umum
άi : Pengaruh sterilisasi tanah ke-i
βj : Pengaruh Trichoderma sp. ke-j
(άβ)ij : Pengaruh interaksi sterilisasi tanah ke-i dan jenis Trichoderma sp. ke-j
εijk : Pengaruh galat sterilisasi tanah ke-i, Trichoderma sp. ke-j dan ulangan
ke-k
Data kuantitatif dari diameter pertumbuhan koloni dianalisis dengan
menggunakan SAS 9.00. Apabila terdapat data yang signifikan akan dilanjutkan
dengan uji lanjut Duncan pada taraf kepercayaan 95%.
Xo Xt
23
HASIL DAN PEMBAHASAN
HASIL
Pertumbuhan Diameter Koloni Ganoderma sp.
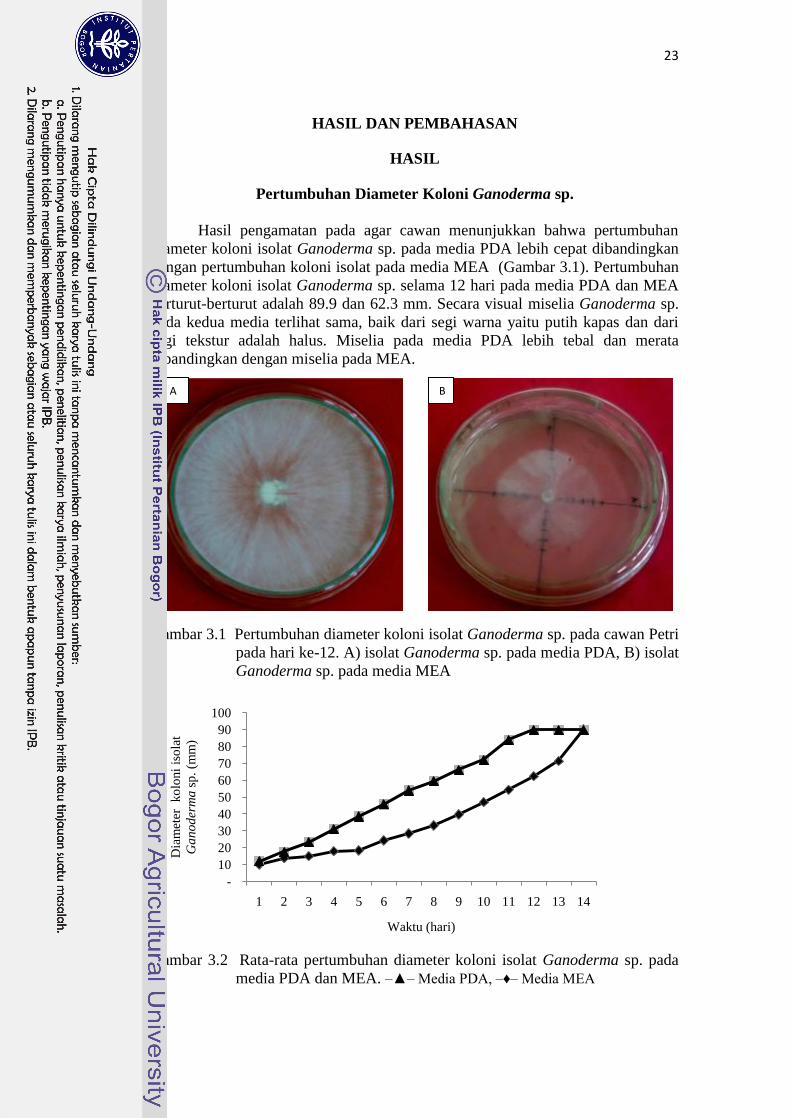
Hasil pengamatan pada agar cawan menunjukkan bahwa pertumbuhan
diameter koloni isolat Ganoderma sp. pada media PDA lebih cepat dibandingkan
dengan pertumbuhan koloni isolat pada media MEA (Gambar 3.1). Pertumbuhan
diameter koloni isolat Ganoderma sp. selama 12 hari pada media PDA dan MEA
berturut-berturut adalah 89.9 dan 62.3 mm. Secara visual miselia Ganoderma sp.
pada kedua media terlihat sama, baik dari segi warna yaitu putih kapas dan dari
segi tekstur adalah halus. Miselia pada media PDA lebih tebal dan merata
dibandingkan dengan miselia pada MEA.
Gambar 3.1 Pertumbuhan diameter koloni isolat Ganoderma sp. pada cawan Petri
pada hari ke-12. A) isolat Ganoderma sp. pada media PDA, B) isolat
Ganoderma sp. pada media MEA
Gambar 3.2 Rata-rata pertumbuhan diameter koloni isolat Ganoderma sp. pada
media PDA dan MEA. ‒▲‒ Media PDA, ‒♦‒ Media MEA
-
10
20
30
40
50
60
70
80
90
100
1 2 3 4 5 6 7 8 9 10 11 12 13 14
Dia
met
er
ko
loni
iso
lat
Ga
no
der
ma
sp
. (m
m)
Waktu (hari)
A B

24
Pertumbuhan diameter koloni isolat Ganoderma sp. pada media PDA lebih
cepat dibanding dengan pertumbuhan diameter koloni isolat pada media MEA
(Gambar 1.2). Pada media PDA diameter pertumbuhan koloni Ganoderma sp.
mampu memenuhi cawan Petri pada hari ke-12, sedang pada media MEA
pertumbuhan diameter koloni Ganoderma sp. memenuhi cawan Petri pada hari
ke-14.
Hasil analisis ragam menunjukkan adanya perbedaan rata-rata
pertumbuhan diameter koloni Ganoderma sp. isolat pada kedua media. Rata-rata
pertumbuhan diameter koloni isolat Ganoderma sp. sebesar 7.09 mm h-1
pada
media PDA berbeda nyata dengan pertumbuhan diameter koloni isolat pada media
MEA dengan nilai 5.41 mm h-1
.
Pertumbuhan Diameter Koloni Trichoderma spp.
Hasil pengamatan pada agar cawan menunjukkan bahwa pertumbuhan
ketiga koloni isolat Trichoderma spp. pada masing-masing media memiliki
perbedaan secara umum baik dari segi pertumbuhan diameter koloni miselia,
warna, tekstur dan ketebalan miselia. Secara umum pertumbuhan diameter koloni
yang tercepat pada T. viride adalah di hari kedua pada media PDA, sedangkan
pada T. pseudokoningii adalah pada media MEA (Gambar 4.2). Pengamatan
visual pertumbuhan T. harzianum pada media PDA terlihat sirkuler, bagian
tengah berwarna putih kehijauan dan bagian luar berwarna hijau tua (Gambar
4.1A). Pengamatan visual koloni isolat T. harzianum pada media MEA terlihat
sirkuler, dan terdiri dari lapisan warna yang berbeda yaitu bagian dalam berwarna
hijau tua, lapisan ke-2 agak transparan, lapisan ke-3 berwarna hijau tua, lapisan
ke-4 berwarna putih tipis dan lapisan paling luar berwarna hijau tua (Gambar
4.1B). Koloni isolat T. pseudokoningii pada media PDA tumbuh menyebar,
berwarna hijau muda dan berukuran tipis (Gambar 4.1C). Koloni isolat T.
pseudokoningii pada media MEA terlihat menyebar, berwarna hijau muda, dan
agak tebal (Gambar 4.1D). Koloni isolat T. viride pada media PDA berbentuk
kapas tebal, bagian tengah berukuran tipis, dan berwarna putih (Gambar 4.1E),
sedangkan koloninya pada media MEA membentuk titik-titik berwarna hijau
muda, dan tebal (Gambar 4.1F).
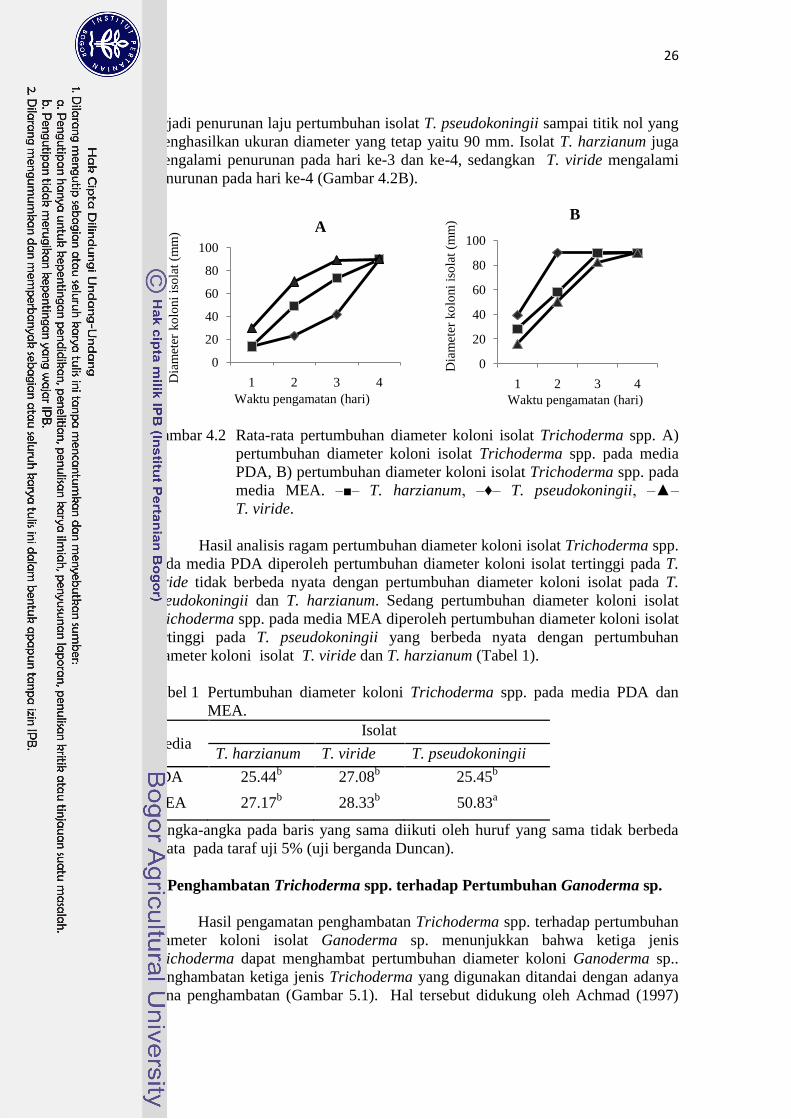
Isolat T. pseudokoningii dan T. harzianum pada media PDA di hari
pertama memiliki pertumbuhan diameter yang sama yaitu 13.67 mm, namun
untuk isolat T. viride pertumbuhan diameter koloninya lebih besar dari kedua
isolat, yaitu 29.50 mm. Pada hari ke-2 terjadi penambahan pertumbuhan diameter
koloni pada ketiga isolat yaitu T. pseudokoningii sebesar 9.30 mm, T. harzianum
sebesar 35.50 mm dan T. viride sebesar 40.70 mm. Hasil tersebut menunjukkan
bahwa pertumbuhan isolat T. viride pada hari ke-2 lebih cepat dibandingkan
kedua isolat lainnya. Memasuki hari ke-3 terjadi penurunan laju pertumbuhan
isolat T. harzianum menjadi 23.90 mm dan T. viride menjadi 18.80 mm,
sedangkan T. pseudokoningii meningkat dengan nilai 18.50 mm. Ketiga jenis
isolat memenuhi cawan Petri pada hari ke-4 (Gambar 4.2A).
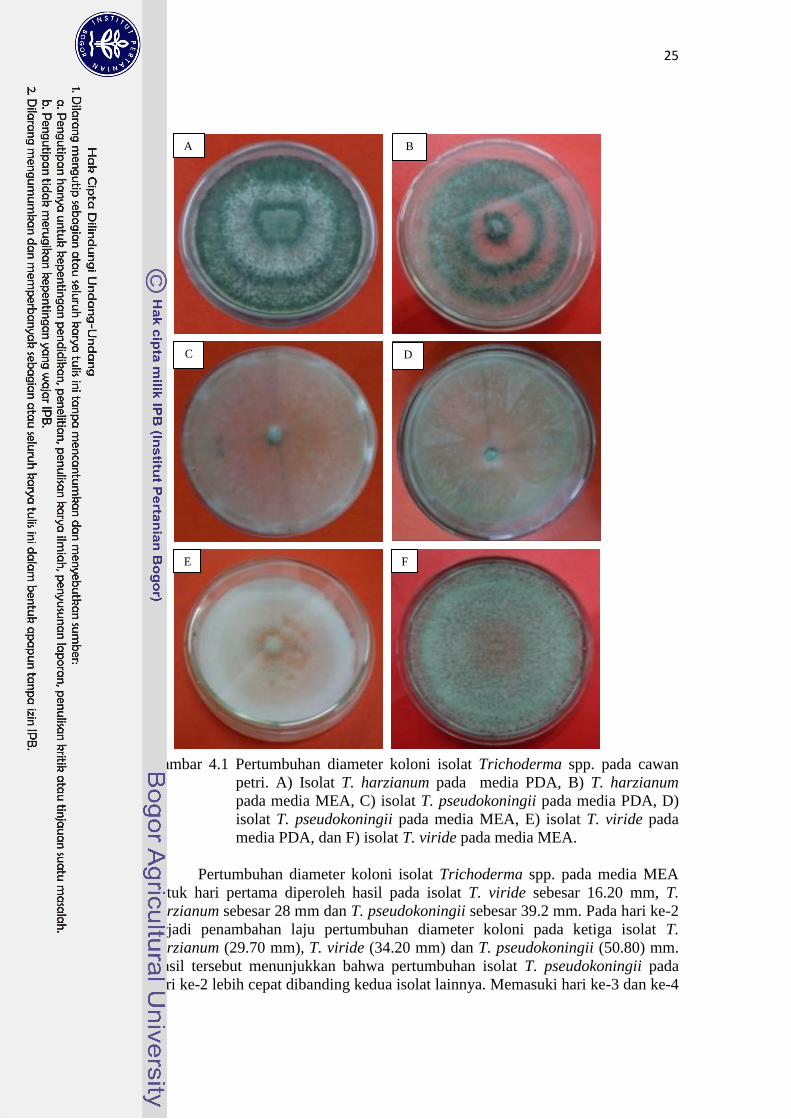
25
Gambar 4.1 Pertumbuhan diameter koloni isolat Trichoderma spp. pada cawan
petri. A) Isolat T. harzianum pada media PDA, B) T. harzianum
pada media MEA, C) isolat T. pseudokoningii pada media PDA, D)
isolat T. pseudokoningii pada media MEA, E) isolat T. viride pada
media PDA, dan F) isolat T. viride pada media MEA.
Pertumbuhan diameter koloni isolat Trichoderma spp. pada media MEA
untuk hari pertama diperoleh hasil pada isolat T. viride sebesar 16.20 mm, T.
harzianum sebesar 28 mm dan T. pseudokoningii sebesar 39.2 mm. Pada hari ke-2
terjadi penambahan laju pertumbuhan diameter koloni pada ketiga isolat T.
harzianum (29.70 mm), T. viride (34.20 mm) dan T. pseudokoningii (50.80) mm.
Hasil tersebut menunjukkan bahwa pertumbuhan isolat T. pseudokoningii pada
hari ke-2 lebih cepat dibanding kedua isolat lainnya. Memasuki hari ke-3 dan ke-4
A
C
B
E
D
F
26
0
20
40
60
80
100
1 2 3 4Dia
met
er k
olo
ni
iso
lat
(mm
)
Waktu pengamatan (hari)
A
0
20
40
60
80
100
1 2 3 4
Dia
met
er k
olo
ni
iso
lat
(mm
)
Waktu pengamatan (hari)
B
terjadi penurunan laju pertumbuhan isolat T. pseudokoningii sampai titik nol yang
menghasilkan ukuran diameter yang tetap yaitu 90 mm. Isolat T. harzianum juga
mengalami penurunan pada hari ke-3 dan ke-4, sedangkan T. viride mengalami
penurunan pada hari ke-4 (Gambar 4.2B).
Gambar 4.2 Rata-rata pertumbuhan diameter koloni isolat Trichoderma spp. A)
pertumbuhan diameter koloni isolat Trichoderma spp. pada media
PDA, B) pertumbuhan diameter koloni isolat Trichoderma spp. pada
media MEA. ‒■‒ T. harzianum, ‒♦‒ T. pseudokoningii, ‒▲‒
T. viride.
Hasil analisis ragam pertumbuhan diameter koloni isolat Trichoderma spp.
pada media PDA diperoleh pertumbuhan diameter koloni isolat tertinggi pada T.
viride tidak berbeda nyata dengan pertumbuhan diameter koloni isolat pada T.
pseudokoningii dan T. harzianum. Sedang pertumbuhan diameter koloni isolat
Trichoderma spp. pada media MEA diperoleh pertumbuhan diameter koloni isolat
tertinggi pada T. pseudokoningii yang berbeda nyata dengan pertumbuhan
diameter koloni isolat T. viride dan T. harzianum (Tabel 1).
Tabel 1 Pertumbuhan diameter koloni Trichoderma spp. pada media PDA dan
MEA.
Media Isolat
T. harzianum T. viride T. pseudokoningii
PDA 25.44b 27.08
b 25.45
b
MEA 27.17b 28.33
b 50.83
a
aAngka-angka pada baris yang sama diikuti oleh huruf yang sama tidak berbeda
nyata pada taraf uji 5% (uji berganda Duncan).
Penghambatan Trichoderma spp. terhadap Pertumbuhan Ganoderma sp.
Hasil pengamatan penghambatan Trichoderma spp. terhadap pertumbuhan
diameter koloni isolat Ganoderma sp. menunjukkan bahwa ketiga jenis
Trichoderma dapat menghambat pertumbuhan diameter koloni Ganoderma sp..
Penghambatan ketiga jenis Trichoderma yang digunakan ditandai dengan adanya
zona penghambatan (Gambar 5.1). Hal tersebut didukung oleh Achmad (1997)

27
yang menyatakan bahwa Trichoderma sp. bersifat sebagai antagonis secara in
vitro dengan terbentuknya zona penghambatan yang merupakan indikasi awal
terlibatnya antibiotik dan antagonisme baik pada media PDA maupun MEA.
Gambar 5.1 Penghambatan pertumbuhan diameter koloni Ganoderma sp. oleh
Trichoderma spp. pada media PDA dan MEA. G = Ganoderma sp.
Th = T. harzianum, Tv = T. viride, Tp = T. pseudokoningii
Persentase penghambatan Trichoderma spp. terhadap pertumbuhan
diameter koloni Ganoderma sp. pada media PDA diperoleh rata-rata
penghambatan tertinggi pada perlakuan T. harzianum (74.19%), T pseudokoningii
(59.37%), dan T. viride (41.36%) (Gambar 5.2). Persentase penghambatan pada
media MEA secara berturut-turut diperoleh rata-rata penghambatan tertinggi
yaitu T. harzianum (73.00%), T. pseudokoningii (60.22%), dan T. viride
(51.58%).
MEA
PDA MEA
PDA MEA
G
G
G
G
Th Th
Tv
Tp Tp
G
PDA
G Tv

28
0
10
20
30
40
50
60
70
80
Th Tp Tv
Per
sen
tase
pen
gh
am
ba
tan
(%)
Jenis Trichoderma
Gambar 5.2 Persentase penghambatan Trichoderma spp. terhadap pertumbuhan
diameter koloni Ganoderma sp. pada media PDA dan MEA. Th = T.
harzianum, Tp = T. pseudokoningii, Tv = T. viride. (■) Media PDA,
( ) Media MEA.
Pengendalian Ganoderma sp. pada Balok Kayu Sengon di Rumah Kaca
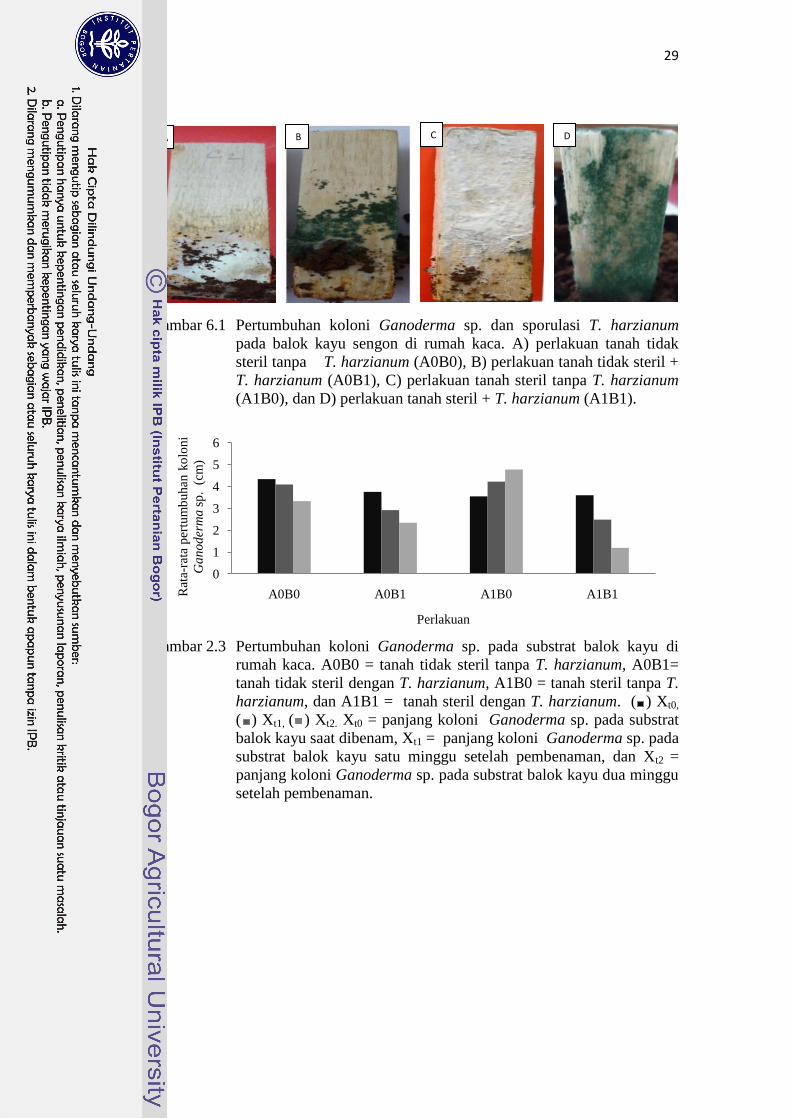
Hasil penelitian menunjukkan bahwa pertumbuhan koloni Ganoderma sp.
dan T. harzianum pada substrat balok kayu secara visual terlihat jelas (Gambar
6.1). Pada perlakuan tanah tidak steril tanpa T. harzianum terlihat koloni
Ganoderma sp. berwarna putih terang (Gambar 6.1A). Pada perlakuan tanah tidak
steril + T. harzianum telihat sporulasi T. harzianum pada permukaan balok kayu
sengon yang berwarna hijau, bila diperhatikan terlihat banyak konidia yang
berwarna hijau, dan terlihat koloni Ganoderma sp. berwarna putih dengan luasan
yang lebih kecil dibanding dengan perlakuan yang lain (Gambar 6.1B). Perlakuan
tanah steril tanpa T. harzianum terlihat koloni Ganoderma sp. yang hampir
memenuhi permukaan balok kayu sengon dengan warna putih kapas (Gambar
6.1C). Pada perlakuan tanah steril + T. harzianum terlihat sporulasi T. harzianum
yang berwarna hijau dan koloni Ganoderma sp. pada permukaan balok kayu
sengon dengan warna putih (Gambar 6.1D).
Rata-rata nilai pertumbuhan panjang koloni Ganoderma sp. pada balok
kayu dapat dilihat pada Gambar 6.2. Penurunan pertumbuhan panjang koloni
Ganoderma sp. saat satu minggu setelah balok kayu dibenam (Xt1) diperoleh nilai
penurunan panjang koloni tertinggi sebesar 1.12 cm pada perlakuan tanah steril +
T. harzianum, sedang pada perlakuan tanah steril tanpa T. harzianum terjadi
pertambahan panjang koloni sebesar 0.67 cm. Demikian halnya pada saat dua
minggu setelah balok kayu di benam (Xt2) pada perlakuan tanah steril + T.
harzianum terjadi penurunan panjang koloni sebesar 2.41 cm, sedang pada
perlakuan tanah steril tanpa T. harzianum justru terjadi pertambahan panjang
koloni Ganoderma sp. sebesar 1.22 cm. Weller (1988) menyatakan bahwa
pengendalian hayati sering dipengaruhi oleh faktor biotik maupun abiotik. Hal
tersebut diduga diakibatkan oleh kemampuan pengaruh sterilisasi tanah yang
mengakibatkan T. harzianum tidak berkompetisi dengan mikroorganisme tanah
lain sehingga mampu menghambat pertumbuhan koloni Ganoderma sp. pada
substrat balok kayu.
29
0
1
2
3
4
5
6
A0B0 A0B1 A1B0 A1B1Rat
a-ra
ta p
ertu
mb
uhan
ko
loni
Ga
no
der
ma
sp.
(cm
)
Perlakuan
Gambar 6.1 Pertumbuhan koloni Ganoderma sp. dan sporulasi T. harzianum
pada balok kayu sengon di rumah kaca. A) perlakuan tanah tidak
steril tanpa T. harzianum (A0B0), B) perlakuan tanah tidak steril +
T. harzianum (A0B1), C) perlakuan tanah steril tanpa T. harzianum
(A1B0), dan D) perlakuan tanah steril + T. harzianum (A1B1).
Gambar 2.3 Pertumbuhan koloni Ganoderma sp. pada substrat balok kayu di
rumah kaca. A0B0 = tanah tidak steril tanpa T. harzianum, A0B1=
tanah tidak steril dengan T. harzianum, A1B0 = tanah steril tanpa T.
harzianum, dan A1B1 = tanah steril dengan T. harzianum. ( ) Xt0,
( ) Xt1, ( ) Xt2. Xt0 = panjang koloni Ganoderma sp. pada substrat
balok kayu saat dibenam, Xt1 = panjang koloni Ganoderma sp. pada
substrat balok kayu satu minggu setelah pembenaman, dan Xt2 =
panjang koloni Ganoderma sp. pada substrat balok kayu dua minggu
setelah pembenaman.
B
V A C D

30
PEMBAHASAN
Secara visual miselia Ganoderma sp. pada media PDA dan MEA terlihat
sama, baik dari segi warna yaitu putih kapas dan dari segi tekstur adalah halus.
Miselia Ganoderma sp. pada media PDA lebih tebal dan merata dibandingkan
dengan miselia pada media MEA. Pertumbuhan diameter koloni isolat
Ganoderma sp. pada media PDA lebih cepat dibanding dengan pertumbuhan
diameter koloni isolat pada media MEA. Perbedaan tersebut diduga terjadi karena
kedua jenis media memiliki kandungan nutrisi yang berbeda. Chang dan Miles
(1989), menyatakan bahwa cendawan untuk dapat tumbuh membutuhkan
beberapa elemen nutrisi dalam jumlah yang spesifik dalam media sesuai dengan
spesies dari cendawan tersebut. Media yang banyak digunakan di laboratorium
terbuat dari ekstrak bahan alami yang mengandung karbohidrat dan hara lain.
Bahan alami yang banyak digunakan untuk menumbuhkan cendawan secara in-
vitro adalah ekstrak kentang, tepung jagung, dan malt ekstrak atau kecambah
gandum (Agrios 1997).
Media PDA memiliki kandungan nutrisi karbohidrat, air, dan protein yang
berasal dari ekstrak kentang, glukosa dan agar. Menurut Achmad (1997) bahwa
PDA merupakan media kaya dengan gula sederhana sebagai sumber karbon. Dari
setiap 100 g dalam bentuk utuh mengandung protein 1.6 g, serat 0.6 g, fosfor 40
mg, vitamin C 17 mg, vitamin B2 0.01 mg, magnesium 30 mg, kalium 2.47 mg,
karbohidrat 22.6 g, lemak 0.1 g, kalsium 10 mg, zat besi 0.1 mg, vitamin B1 0.1
mg, niacin 1.2 mg, dan sodium 11 mg (Al-Weshahy dan Rao 2012).
Bagaimanapun juga kandungan beberapa unsur tersebut tersedia di dalam ekstrak
kentang sudah barang tentu lebih rendah. Media MEA mengandung nitrogen,
karbohidrat, dan sodium klorida. Disamping malt ekstrak juga vitamin yang
berbeda jumlahnya dengan ekstrak kentang, baik dalam jenis maupun jumlahnya.
Sumber karbon pada PDA berasal dari gula sederhana. Karbohidrat diperlukan
untuk pertumbuhan koloni cendawan, pembentukan struktur dan keperluan energi
bagi sel cendawan. Dalam ekstrak kentang terkandung senyawa-senyawa asam
amino, asam organik, enzim, fenol, solanin dan unsur (Smith 1968).
Nitrogen digunakan oleh cendawan dalam sistesis protein, purine,
pirimidin dan komponen kitin pada dinding sel cendawan (Chang dan Miles
1989). Nitrogen dibutuhkan oleh semua organisme untuk mensintesa asam amino
dan membentuk protein yang dibutuhkan untuk membentuk protoplasma. Tanpa
protein, pertumbuhan tidak dapat terjadi. Cendawan dapat menggunakan nitrogen
anorganik untuk pembentukan nitrat, nitrit, ammonia atau nitrogen organik untuk
pembentukan asam amino. Tidak semua cendawan menggunakan sumber nitrogen
dengan jenis yang sama dan setiap cendawan membutuhkan nitrogen dalam
bentuk yang berbeda-beda (Moore 1982).
Seperti telah disebutkan di atas bahwa ekstrak malt dan ekstrak kentang
juga mengandung mineral dan vitamin. Mineral berfungsi sebagai aktivator enzim
dan vitamin berfungsi sebagai katalisator di dalam sel yaitu sebagai koenzim atau
merupakan bagian yang menyusun koenzim (Chang dan Miles 1989; Hadi 1999).
Moore (1972) menjelaskan akan pentingnya unsur karbon bagi cendawan karena
cendawan membutuhkan unsur karbon dalam jumlah yang besar daripada unsur-
unsur esensial yang lain dan karbon merupakan nutrisi yang pokok dan terpenting
pada cendawan.

31
Pertumbuhan serta perkembangan cendawan akan sangat dipengaruhi oleh
sejumlah faktor. Menurut Dhingra dan Sinclair (1985), faktor tersebut di
antaranya ialah suhu, cahaya, udara, pH serta nutrisi. Apabila kandungan nutrisi
kurang dan kondisi ruang tumbuh sempit maka pertumbuhan Trichoderma spp.
akan melambat dan tidak mampu untuk membentuk konodium. Danielson dan
Davey (1973), menyatakan bahwa konidia Trichoderma spp. memerlukan nutrisi
dari luar agar berkecambah.
Uji penghambatan Trichoderma spp. terhadap pertumbuhan koloni isolat
Ganoderma sp. menunjukkan bahwa ketiga jenis Trichoderma spp. yang di uji
dapat menghambat pertumbuhan diameter koloni Ganoderma sp.. Penghambatan
ketiga jenis Trichoderma yang digunakan terlihat dengan adanya zona
penghambatan (Gambar 5.1). Trichoderma sp. bersifat sebagai antagonis secara
in vitro dengan terbentuknya zona penghambatan yang merupakan indikasi awal
terlibatnya antibiotik dan antagonisme baik pada media PDA maupun MEA
(Achmad 1997). Hasil penelitian yang sama juga dilaporkan oleh Abadi (1987)
dan Dharmaputra (1989) yang menyatakan bahwa Trichoderma spp. dapat
menghambat pertumbuhan Ganoderma boninense.
Pertumbuhan koloni Trichoderma spp. pada media PDA dan MEA lebih
cepat dari pada pertumbuhan koloni Ganoderma sp. sehingga Trichoderma spp.
mempunyai kemampuan kompetisi lebih tinggi. Dennis dan Webster (1971)
menyatakan bahwa Trichoderma sp. mempunyai daya antagonis yang tinggi dan
dapat mengeluarkan racun (mikotoksin) yaitu senyawa yang dapat menghambat
bahkan dapat mematikan cendawan lain. Penghambatan pada perlakuan T.
harzianum lebih kuat dibanding dengan T. viride, dan T. pseudokoningii. Hal ini
didukung oleh Achmad et al. (2010) yang mengemukakan bahwa T.harzianum
lebih kuat menghambat pertumbuhan patogen lodoh secara in vitro pada patogen
lodoh Pinus merkusii, mengakibatkan terbentuknya zona hambatan yang lebih
besar dan menghasilkan kitinase yang lebih efektif mendegradasi kitin dibanding
T. pseudokoningii. Terbentuknya zona penghambatan antagonisme pada media
padat menunjukkan bahwa cendawan antagonis mendifusikan metabolit yang
dapat menghambat pertumbuhan cendawan patogen (Achmad 1997). Lebih lanjut
Baker dan Scher (1987) menyatakan bahwa pengujian antibiosis melalui
pembentukan zona hambatan pada media agar, dapat disebabkan oleh mikroba
yang menghasilkan antibiotik yang larut atau tidak larut.
Mekanisme dalam antagonisme antar jasad renik, yaitu antibiosis,
kompetisi, dan mikoparasitisme (Baker dan Cook 1974). Terbentuknya zona
penghambatan pada media padat merupakan indikasi bekerjanya mekanisme
antibiosis. Bekerjanya mekanisme antibiosis tersebut dikuatkan oleh tertekannya
pertumbuhan cendawan patogen pada media padat. Terbentuknya penghambatan
terhadap pertumbuhan diameter koloni Ganoderma sp. diduga karena adanya
enzim dan senyawa metabolit yang diproduksi oleh Trichoderma spp. yang
mungkin mampu merusak dinding sel Ganoderma sp.. Kerusakan pada dinding
sel mengakibatkan rusaknya susunan dan perubahan mekanisme permeabilitas
dari mikrosom, lisosom dan dinding sel. Kerusakan pada membran ini
memungkinkan ion anorganik yang penting, nukleotida, koenzim dan asam amino
berosmosis ke luar sel. Selain itu, kerusakan membran dapat mencegah masuknya
bahan-bahan penting ke dalam sel karena membran sitoplasma juga
mengendalikan pengangkutan aktif dalam sel (Volk dan Wheeler 1993).

32
Mekanisme antibiosis dapat melibatkan metabolit beracun (toksin) atau
enzim ekstraseluler yang dihasilkan oleh cendawan antagonis. Trichorderma sp.
menghasilkan toksin trikhor dermin yang merupakan suatu senyawa sesquiterpen,
dermadin yaitu asam berbasa tunggal yang aktif terhadap cendawan dengan
kisaran yang luas dan meliputi bakteri gram positif dan gram negatif, serta dua
senyawa peptida yang bersifat antifungal sekaligus anti bakterial. Tertekannya
pertumbuhan cendawan patogen menunjukkan mekanisme kompetisi dalam
antagonisme, dalam hal ini cendawan antagonis lebih kompetitif dalam
memanfaatkan ruang tumbuh dan nutrisi. Selama Trichoderma spp. tumbuh aktif
menghasilkan sejumlah besar enzim ekstra selular ß (1.3) glukonase, dan kitinase,
yang dapat melarutkan dinding sel patogen (Lewis dan Papavizas 1984).
Hasil pengamatan pengendalian Ganoderma sp. pada balok kayu sengon
di rumah kaca menunjukkan adanya penekanan pertumbuhan koloni Ganoderma
sp. sebagai akibat perlakuan T. harzianum. Kemampuan T. harzianum dalam
menekan pertumbuhan Ganoderma sp. diduga merupakan bentuk dari
kemampuannya sebagai kompetitor yang unggul dalam mendapatkan ruang dan
nutrisi atau merupakan bentuk dari kemampuannya sebagai antagonis. Hal ini
sesuai dengan pendapat Wells (1988) yang menyatakan bahwa T. harzianum
dapat tumbuh dengan cepat pada berbagai substrat dan memiliki kemampuan
kompetisi yang baik terhadap makanan dan ruang. Pada permukaan balok kayu
sengon dengan perlakuan Ganoderma sp. terjadi perubahan warna dari putih
menjadi putih kecoklatan. Ini terjadi karena adanya kemampuan Ganoderma sp.
menghasilkan enzim hidrolitik terutama selulase sebagai pelapuk (Domsch et al.
1980).
Kemampuan T. harzianum yang secara konsisten melakukan kompetisi
tidak terlepas dari kemampuannya dalam merubah strategi berdasarkan kondisi
lingkungannya. Dalam kondisi lingkungan yang sesuai T. harzianum mampu
menghasilkan konidia yang melimpah dan menyebar dengan cepat sehingga
memiliki daya kompetitif yang tinggi. Karena itu T. harzianum diklasifikasikan
sebagai ruderal dan oportunis (Williams et al 2003).
Mekanisme interaksi antara T. harzianum dan Ganoderma sp. pada
substrat balok kayu terjadi melalui kompetisi terhadap sumber makanan.
Kompetisi biasanya terjadi terhadap nutrisi dan ruang atau faktor-faktor
pertumbuhan penting lainnya (Achmad. 1997). Selain mekanisme kompetisi,
antagonisme yang terjadi pada substrat balok kayu, kemungkinan terjadi melalui
mikoparasitisme sekrotrofik (Papavizas 1985). Hasil pengamatan pada substrat
balok kayu yang diperlakukan dengan T. harzianum menunjukkan terjadinya
sporulasi yang menutupi semua permukaan substrat. Untuk menghancurkan
inokulum, maka antagonis yang efektif memarasit patogen, dengan kemampuan
saprofitiknya kuat dan beradaptasi dengan baik pada lingkungan fisik tanah
tempat antagonis tersebut diinfestasikan (Achmad 1997).
Pertumbuhan Ganoderma sp. pada balok kayu sengon pada perlakuan
tanah tidak steril + T. harzianum lebih rendah dibandingkan dengan perlakuan
tanah tidak steril tanpa T. harzianum. Pada balok kayu nampak adanya perubahan
warna dari putih menjadi coklat. Hal tersebut diduga adanya pengaruh perlakuan
T. hazianum dan mikroorganisme tanah pada tanah tidak steril yang bekerja
sebagai agens antagonis yang mampu menghambat pertumbuhan koloni
Ganoderma sp. Pertumbuhan Ganoderma sp. pada balok kayu sengon dengan

33
perlakuan tanah steril tanpa T. harzianum yang ditandai dengan perubahan warna
dari putih menjadi coklat kehitaman. Hal ini diduga pada tanah steril tanpa T.
harzianum tidak terjadi kompetisi terhadap nutrisi yang dibutuhkan Ganoderma
sp. dalam pertumbuhannya sehingga Ganoderma sp. mampu menghasilkan enzim
hidrolitik terutama selulase sebagai pelapuk (Domsch et al. 1980). Selanjutnya
Abadi (1987) menyatakan bahwa G. boninense dapat tumbuh pada tanah steril dan
tidak pada tanah tidak steril dalam cawan petri. Kerry dan Bourne (1996)
menegaskan bahwa tanah tidak steril berisi mikroba lain yang telah berada
sebelumnya dalam hal ini dapat berperan sebagai kompetitor. Pada tanah tidak
steril diperkirakan menghadapai kompetisi nutrisi atau mikrohabitat dengan
mikroba lain, mengalami predasi oleh protozoa (Hossain dan Alexander 1984)
atau lisis karena bakteriofage (Keel et al. 2002; Janowitz 2004) dapat terjadi
sehingga mempengaruhi kemampuan antagonis dalam memberikan penekanan
terhadap patogen.
Trichoderma spp. adalah salah satu jenis cendawan yang digunakan
sebagai pengendali hayati karena terdapat di mana-mana, mudah diisolasi dan
dibiakkan, tumbuh dengan cepat pada beberapa macam substrat, mempengaruhi
patogen tanaman, jarang bersifat patogenik pada tanaman tingkat tinggi, bereaksi
sebagai mikoparasit, bersaing dengan baik dalam hal makanan, tempat dan
menghasilkan 24 antibiotik (Wells 1988). Trichoderma sp. dapat digunakan
sebagai agen biokontrol melawan beberapa cendawan petogenik tular tanah
(Anggraeni 2004). Mekanisme pengendalian T. harzianum bersifat khusus
terhadap sasaran, sehingga tidak menimbulkan musnahnya organisme yang bukan
sasaran. Darmono (1994) mengemukakan bahwa penggunaan cendawan antagonis
sebagai pengendali patogen merupakan salah satu alternatif yang dianggap aman
dan dapat memberikan hasil yang cukup memuaskan. Pengendalian hayati
terhadap patogen dengan menggunakan mikroorganisme antagonis dalam tanah
memiliki harapan yang baik untuk dikembangkan karena pengaruh negatif
terhadap lingkungan tidak ada. T. harzianum mempunyai kemampuan untuk
menghasilkan enzim hidrolitik β-1-3 glukanase, kitinase dan selulase. Enzim-
enzim inilah yang secara aktif merusak sel-sel cendawan lain yang sebagian besar
tersusun dari 1,3 glukan (linamirin) dan kitin sehingga dengan mudah T.
harzianum dapat melakukan penetrasi ke dalam hifa cendawan inangnya (Elad et
al. 1983). Degradasi kitin T. harzianum dilakukan secara bertahap, dan hal
tersebut menunjukkan dihasilkannya kitinase secara terus-menerus (Achmad
1997).

34
SIMPULAN DAN SARAN
Simpulan
Trichoderma spp. dapat menghambat pertumbuhan diameter koloni
Ganoderma sp. pada cawan Petri secara in vitro. Persentase daya hambat
Trichoderma spp. terhadap pertumbuhan koloni Ganoderma sp. pada media PDA
diperoleh persentase penghambatan tertinggi pada perlakuan T. harzianum
(74.19%), T. pseudokoningii (59.37%), dan T. viride (41.36%). Sedangkan
Persentase penghambatan terhadap pertumbuhan koloni Ganoderma sp. pada
media MEA secara berturut-turut diperoleh persentase penghambatan tertinggi
yaitu T. harzianum (73.00%), T. pseudokoningii (60.22%), dan T. viride
(51.58%). Penghambatan tersebut terjadi melalui mekanisme antagonisme dengan
terbentuknya zona penghambatan. Kemampuan T. harzianum pada tanah steril
mampu menurunkan pertumbuhan koloni Ganoderma sp. pada balok kayu sengon
di rumah kaca sebesar 1.12 cm pada satu minggu setelah balok kayu dibenam dan
2.41 cm pada dua minggu setelah balok kayu dibenam.
Saran
Trichoderma spp. dapat dipertimbangkan sebagai salah satu agens hayati
dalam pengendalian Ganoderma sp. yang menyerang tanaman sengon secara in
vitro dan skala rumah kaca.

35
DAFTAR PUSTAKA
Abadi AL. 1987. Biologi Ganoderma boninense Pat. pada kelapa sawit (Elaesis
guineensis Jacg.) dan pengaruh beberapa mikroba tanah antagonistik
terhadap pertumbuhannya. [disertasi]. Bogor: Institut Pertanian Bogor.
Achmad. 1997. Mekanisme serangan patogen dan ketahanan inang serta
Pengendalian Hayati Penyakit Lodoh pada Pinus merkusii. [disertasi].
Bogor: Institut Pertanian Bogor.
Achmad, Hadi S, Harran S, Gumbira SE, Satiawiharja B, Kosim. 2010. Aktivitas
antagonisme in vitro T. harzianum dan T. pseudokoningii terhadap patogen
lodoh Pinus merkusii. Jurnal Penelitian Hutan Tanaman. 7(5): 233-240.
Adaskaveg JE, Ogawa JM. 1990. Wood decay pathology of fruit and nut trees in
California. Plant Disease. 74: 341-352.
Agrios GN. 1997. Plant Pathology. New York (US): Academic Press.
Allen GU. 1994. Pedology. London: Trans By T.R.Patton.
Al-Weshahy A, Rao VA. 2012. Potato Peel as a Source of Important
Phytochemical Antioxidant Nutraceuticals and Their Role in Human
Health - A Review. Di dalam: Venketeshwer Rao, editor. Phytochemicals
as Nutraceuticals - Global Approaches to Their Role in Nutrition and
Health. Chapter 11. InTech Europe. Rijeka, Croatia: 207-224.
Anggraeni I. 2004. Identifikasi dan patogenitas penyakit akar pada Acacia
mangium Willd. Buletin Penelitian Hutan. 645: 61-73.
Awang, San Afri W, Andayani B, Himmah WT. Widayanti A, Afianto. 2002.
Hutan rakyat, sosial ekonomi, dan Pemasaran. Fakultas Ekonomi
Universitas Gaja Mada, Yogyakarta.
Bassett K, Peters RN. 2003. Ganoderma; a significant root pathogen).
Arborilogical Services Inc. Publication.
Baker KF, Cook RJ. 1974. Biological control of plant pathogens. Freeman WH
and Co. San Fransisco. 433 p.
Baker RY, Scher FM. 1987. Enhancing the activity of biological control agents.
New York. PP. 1-8
Blanchette RA. 1984. Screening wood decayed by white rot fungi for preferential
lignin degradation. Applied Environmental Microbiology. 48: 647-653.
Chang ST, Miles PG. 1989. Edible Mushrooms and Their Cultivation. Boca
Raton: CRC Press, Inc.
Charomaini M, Suhaendi H. 1997. Genetic variation of Paraserianthes falcataria
seed sources in Indonesia and its potential in tree breeding programs.
Dalam: Zabala, N. (ed.) Workshop international tentang spesies Albizia
dan Paraserianthes, 151–156. Prosiding workshop, 13–19 November 1994,
Bislig, Surigao del Sur, Filipina. Forest, Farm, and Community Tree
Research Reports (tema khusus). Winrock International, Morrilton,
Arkansas, AS.
Chet I. 1987. Trichoderma-application, mode of action, and potential as
biocontrol agent of soil-borne plant pathogenic fungi. Innovative
approaches to plant diseases control. John Wiley and Sons, A Wiley-
Interscience Publication, USA. pp. 11-210.

36
Chet I, Viterbo A, Shoresh M. 2004. Enhancement of plant disease resistance by
the biocontrol agent T. asperellum. Departement of Biology. Chemistry.
www.weizmann.ac.il. 2013.
Cook RJ, Baker KF. 1983. The nature and practice of biological control of plant
pathogens. The American Phytopathological Society, St. Paul, Minnesota
539 p.
Coșkuntuna A, Özer N. 2007. Biological control of onion rot disease using
Trichoderma harzianum and induction of antifungal compounds in onion
set following seed treatmen. Departement of plant protection, Faculti of
Agriculture, Namik Kemal University, Tekirdağ 59030, turkey 27 : 330-
336.
Darmono T. 1994. Kemampuan beberapa isolat Trichoderma spp. dalam menekan
inokulum Phytophthora sp. di dalam jaringan buah kakao. Menara
Perkebunan. 62(2):25-29.
Dennis C, Webster J. 1971. Antagonistic properties of species groups of
Trichoderma. III. Hyphal interaction. Trans. Br. Mycol. Soc. 57 : 363-369.
Dharmaputra OS. 1989. Fungi antagonistik terhadap ganoderma boninense Pat.
penyebab penyakit busuk batang kelapa sawit. Sumatera Utara. Laporan
Tahunan kerjasama penelitian PP Marihat-Biotrop. P.28-45.
Dhingra OD, Sinclair JB. 1985. Basic plant pathology methods. Boca Raton,
Florida. CRC Press, Inc
Domsch KH, Anderson TH. 1980. Compendium of soil fungi. Vol. I. Academic
Press. London.
Elad Y, Chet I, Boyle P, Henis Y. 1983. Parasitism of Trichoderma spp. on
Rhizoctonia Solani and Sclerotium rolfsii. Scanning electron microscopy
and fluorescense microscopy. Phytopathology. 73:85-88.
Hadar Y, Henis Y, Chet I. 1979. Biological control of Rhizoctonia solani
Dumping-off with bran culture of Trichoderma harzianum.
Phytopathology. 69:64-68.
Hadiwiyono. 1996. Pengendalian hayati Ganoderma boninense Pat. (Penyebab
busuk pangkal batang kelapa sawit) Oleh Trichoderma, Gliocladium, dan
Pseudomonas Kelompok fluoresen. Tesis Program Pascasarjana Institut
Pertanian Bogor.
Hepting GH. 1971. Diseases of forest dan shade trees of the United states. US
Department of Agriculture, Agricultural Handbook 386; 1- 658.
Herliyana EN, Darmono T, Minarsih, Hayati. 2012. Root diseases Ganoderma sp.
on the Sengon in West and East Java. Journal of Tropical Forest
Management 18 (2):94-99. DOI:10.7226/jtfm.18.2.94.
Hossain AKM, Alexander M. 1984. Enhancing soybean rhizosphere colonization
by Rhizobium japonicum. Appl Environ Microbiol. 48: 468-472.
Janowitz J. 2004. Culture independence characterization of soil bacteriophage in
agroecosystems. CASNR. Honors research and creative achievement
project. University of Tennessee, Knoxville.
Karsten P. 1881. Numeratio boletinearum et Polyporearum fennicarum,
Systemate novo dispositarum. Rev. Mycol. 3: 16-19.

37
Keel C, Ucurum Z, Michaux P, Adrian M, Haas D. 2002. Deleteroius impact of a
virulent bacteriophage on survival and biocontrol activity of Pseudomonas
fluorescens strain CHA0 in natural soils. Mol Plant Microbe Interact.
15: 567-576.
Kerry BR, Bourne JM. 1996. The important of rhizosphere interaction in the
biological control of plant-parasitic nematodes-case study using
Verticillium chlamydosporium. Pestic Sci. 47: 69-75.
Irianto RSB, Barry K.N, Hidayati, Ito S, Fiani A, Rimbawanto A, Mohammed
C. 2006. Incidence and spatial analysis of root rot of Acacia mangium in
Indonesia. Journal of Tropical Forest Science. 18(3): 157-165.
Lee SS. 1996. Diseases of some tropical plantation Acacias in Peninsular
Malaysia, in K. M. Old, Lee S. S. and J. K. Sharma. Diseases of tropical
Acacias. Proc. Internat. Workchop Subanjeriji (South Sumatra). CIFOR,
Jakarta.
Lewis JR, Papavizas GC. 1991. Biocontrol of plant disease: The approach for
tomorrow. Crop Protect. 10:95-105.
Martawijaya A, Kartasujana I, Mandang YI, Prawira SA, Kadir K. 1989. Atlas
Kayu Indonesia Jilid II. Pusat Penelitian dan Pengembangan Hasil Hutan,
Bogor, Indonesia.
Mile Y. 2003. Penilaian tingkat produktivitas dan kelestarian hutan rakyat.
Prosiding seminar sehari prospek pengembangan hutan rakyat di era
otonomi daerah, Loka Litbang Hutan Monsoon. Badan Litbang
Departemen Kehutanan.
Papavizas GC. 1985. Trichoderma and Gliocladium: bioecology, ecology, and
potential for biocontrol. Ann. Rev. Phytopathol. 23:23-54.
Phillips DH, Burdekin DA. 1989. Diseases of forest and ornamental trees. The
Macmillan Press LTD. London. 435p.
Rahayu S. 1994. Pola Penyebaran Penyakit Akar pada Tegakan Sengon
(Paraserianthes falcataria) di Hutan Rakyat pada Berbagai Ketinggian
Tempat. Laporan Penelitian. DPP Fakultas Kehutanan Universitas Gadjah
Mada. Yogyakarta.
Reese ET, Mendels M. 1959. β_1.3 Glucanases in fungi. Can. J. Microbiology. 5:
173-185.
Salomon JD, Leiniger TD, Anderson RL, Thompson LC, McCracken FI. 1993.
Ash Pesta: A guide to major insects, diseases, air pollution injury and
chemically injury. Gen. Tech. Rep. SO-96. New Orleans, LA; U.S.
Department of Agriculture, Forest Service, Southern Forest Experiment
Station. 45 p.
Semangun H. 2000. Penyakit-penyakit tanaman perkebunan di Indonesia. Gadjah
Mada University Press.Yogyakarta. 808p.
Sharma JK, Florence EJ. 1996. Fungal pathogens as potential threat to tropical
Acacias-Case Study of India in K. M. Old, Lee s. s. and J. K. Sharma.
Diseases of Tropical Acacias. Proc. Internat. Workshop Subanjeriji South
Sumatra). Cifor. Jakarta.
Soerianegara I, Lemmens RH. 1993. Plant resources of South-East Asia 5(1):
Timber trees:major commercial timbers. Pudoc Scientific Publishers,
Wageningen, Belanda.

38
Steyaert RL. 1967. Les Ganoderma palmicoles.Bull.Jard. Bot. Nat. Belg. :J7: 465
- 492.
Turner PD. 1981. Oil palm diseases and disorder. Oxford University Press.
Kualalumpur. 280p.
Van der Plank JE. 1963. Plant disease. Epidemics and control. Academic Press.
New York. 344p.
Webster J, Dennis. 1971. Antagonistic properties of the different species group of
Trichoderma. II. Production of Volatile Antibiotics. Trans. Mycol.Soc. 57
(I): 41-48.
Weller DM. 1988. Biological control of soil borne plant pathogens in rhizosphere
with bacteria. Ann. Rev. Phytopathol. 26:379-407.
Wells HD. 1988. Trichoderma as a biocontrol agent. Pp. 71-79. in Murkeji KG
(EDS). Biocontrol of Plant Diseases. Vol. I. CRC Press, Inc. Boca Raton.
Florida.
Widyastuti SM. 1998. Trichoderma spp. As decomposing and biological control
agens isolated from Dipterocarp forest in Jambi. Prosiding internasional
seminar ecology aproach for productivity and sustainability of dipterocarp
Forest. Yogyakarta. Indonesia. 58-60.
Widyastuti SM, Sumardi, Harjono. 1999. Potensi antagonistik tiga Trichoderma
spp. terhadap delapan penyakit akar tanaman kehutanan. Buletin
Kehutanan. 36:24-38.
Widyastuti SM, Sumardi, Sumantoro P. 2001. Efektivitas Trichoderma spp.
sebgai pengendali hayati terhadap tiga patogen tular tanah pada beberapa
jenis tanaman kehutanan. Jurnal Perlindungan Hutan Tanaman.
Indonesia. 7(2):98-107.
Williams J, Clarkson JM, Mills PR, Cooper RM. 2003. Saprotrophic and
mycoparasityc components of aggressiveness of Trichoderma harzianum
groups towards the commersial mushroom Agaricus bisporus. Appl
Environ Microbiol. 96(7): 4192-4199.
Yates IE, Meredith F, Smart W, Bacon CW, Jaworski AJ. 1999. Trichoderma
viride suppresses fumonisin B1 production by Fusarium moniliforme, J.
Food Prot. 62(11): 1326 – 1332.

39
LAMPIRAN
Lampiran1 Sidik ragam diameter koloni Ganoderma sp.
Source DF Sum of
Squares
Mean
Square
F Value Pr > F
media 1 4.25041667 4.25041667 17.46 0.0139
Error 4 0.97386667 0.24346667
Corrected
Total
5 5.22428333
Lampiran 2 Sidik ragam diameter koloni Trichoderma spp.
Source DF Sum of
Squares
Mean Square F
Value
Pr > F
Media 1 402.1448000 402.1448000 23.22 0.0004
trichoderma 2 501.7124778 250.8562389 14.49 0.0006
media*trichoderma 2 571.3780333 285.6890167 16.50 0.0004
Error 12 207.818000 17.318167
Corrected Total 17 1683.053311
Lampiran 3 Sidik ragam penghambatan Trichoderma spp. terhadap
pertumbuhan diameter koloni Ganoderma sp.
Lampiran 4 Sidik ragam panjang koloni Ganoderma sp. pada substrat balok
kayu (Xt1)
SK db JK KT F -hit p-value
perlakuan 3 0.50340150 0.16780050 13.24 0.0018
Error 8 0.10141583 0.01267698
Total 11 0.60481733
Lampiran 5 Sidik ragam panjang koloni Ganoderma sp. pada substrat balok
kayu (Xt2)
SK db JK KT F -hit p-value
perlakuan 3 0.78091902 0.26030634 42.31 <.0001
Galat 8 0.04922138 0.00615267
Total 11 0.83014040
Source DF Sum of
Squares
Mean Square F
Value
Pr > F
Media 1 48.972006 48.972006 1.78 0.2072
trichoderma 2 2207.278978 1103.639489 40.06 <.0001
edia*trichoderma 2 111.086178 55.543089 2.02 0.1758
Error 12 330.563733 27.546978
Corrected Total 17 2697.900894

40
Lampiran 6 Komponen penyusun media PDA dan MEA
Media Komponen penyusun
PDA
Karbohirdat
Protein
Air
MEA
Nitrogen
Karbohirat
Sodium klorida

41
RIWAYAT HIDUP
Penulis dilahirkan di Tiropadang, Kabupaten Tana Toraja, Provinsi Sulawesi
Selatan pada tanggal 30 Desember 1976 dari ayah Thomas Dendang dan ibu Germana
Palanda. Pendidikan sarjana ditempuh di Program Studi Ilmu Hama dan Penyakit
Tumbuhan, Fakultas Pertanian UNHALU, lulus pada tahun 2003. Pada tahun 2009,
penulis diterima di Program Studi Silvikultur Tropika pada Program Pascasarjana IPB.
Beasiswa pendidikan pascasarjana diperoleh dari Kementrian Kehutanan Republik
Indonesia.
Penulis bekerja sebagai Calon Peneliti di Balai Penelitian Kehutanan sejak tahun
2005 dan di tempatkan Balai Teknologi Agroforestry Ciamis.