jurnal dr. yanuarman 1
TRANSCRIPT

Autophagi terkait Protein, LC3 dan Beclin - 1 , pada Plasenta dari Kehamilan yang dipersulit
oleh Preeklamsia
Tujuan dari penelitian ini adalah untuk mengetahui ekspresi autophagy yang terkait protein ( LC3 dan
Beclin - 1 ) pada plasenta manusia dan perubahan yang terjadi di plasenta dari kehamilan yang dipersulit oleh preeklamsia ( PE ) dan untuk mengeksplorasi mekanisme regulasi protein ini dalam sel JEG – 3 dalam menanggapi hipoksia atau perlakuan sitokin .Kehadiran dari autophagosomes dikonfirmasi dengan mikroskop elektron dan ekspresi LC3 dan Beclin - 1 oleh metode immunoimaging dalam trofoblas plasenta manusia. Dibandingkan dengan plasenta dari kehamilan normal , ekspresi LC3 - II
protein , tapi bukan Beclin - 1 , meningkat pada plasenta dari Preeklampsia parah . Dalam JEG - 3 sel , hipoksia
( O2 < 1 % ) memicu sedikit peningkatan namun tidak signifikan dalam ekspresi LC3 - II dengan penurunan ekspresi - Beclin 1 secara signifikan. Sementara itu, perlakuan TNF - α memicu peningkatan ekspresi LC3 - II yang signifikan tanpa perubahan signifikan dalam ekspresi Beclin - 1. Data kami menunjukkan bahwa yang meningkatkan aktivitas autophagic pada plasenta mungkin terlibat dalam patofisiologi PE.
Preeklamsia (PE) merupakan salah satu komplikasi kehamilan yang paling sering dan bertanggung jawab atas kematian dan morbiditas perinatal . Meskipun mekanisme yang mendasari PE tidak dimengerti dengan jelas, plasentasi abnormal dan disfungsi plasenta yang terlibat dalam etiologi gangguan ini. Selain itu, banyak peneliti telah melaporkan peningkatan apoptosis dari trofoblas di plasenta dari pasien dengan PE1 menunjukkan bahwa regulasi yang berubah dari trofoblas apoptosis dapat menyebabkan patofisiologi dari PE. Namun, mekanisme selain apoptosis mungkin berhubungan dengan disfungsi plasenta yang terkait dengan PE. Selain itu, ada bukti substansial untuk mendukung fakta bahwa plasenta dari wanita dengan PE ditandai oleh respon inflamasi yang berlebihan dan stres oksidatif dibandingkan dengan plasenta dari kehamilan normal. Autophagy juga disebut sebagai program kematian sel tipe II, merupakan sistem degradasi massal intraseluler yang bertanggung jawab atas degradasi lisosomal protein dan konstituen subselular lainnya. Seringkali diaktifkan sebagai respons terhadap kekurangan gizi dan stres intraseluler.
Autophagy adalah proses pelestarian evolusi yang terjadi pada semua sel eukariotik, dari ragi untuk mamalia. Autophagy telah terlibat dalam berbagai proses fisiologis termasuk diferensiasi selular dan kematian sel. Selain itu, autophagy dan apoptosis sering terkoaktivasi dalam respon terhadap stres. Dalam dekade terakhir, kemajuan pesat dalam memahami mekanisme molekuler dari autophagy telah dibuat dengan menggunakan teknik genetika untuk studi ragi Saccharomyces cerevisiae. Baru-baru ini, beberapa homolog mamalia ragi autophagy (ATG) gen telah diidentifikasi. Di antaranya, LC3, homolog mamalia

ragi Apg 8/Aut7, adalah protein pertama kali diidentifikasi dari membran autophagosome. Tiga isoform dari LC3 mRNA yang saat ini dikenal, LC3A, LC3B, dan LC3C.
Setelah disintesis, LC3 selanjutnya diproses untuk LC3-I (bentuk sitosolik) dan dimodifikasi ke _ LC3-II aktif (membran bentuk terikat _) dengan modifikasi posttranslational.
LC3-II, setelah konjugasi dan lipidasi dengan phosphatidylethanolamine, terikat pada membran luar autophagosome. Oleh karena itu, LC3 merupakan penanda yang dapat diandalkan pembentukan autophagosome dalam sel mamalia, dan jumlah relatif LC3-II mencerminkan aktivitas autophagic. Homolog penting lainnya dari Apg6/Vps ragi 30 adalah Beclin-1, yang dikenal untuk memicu autophagy dalam menanggapi kekurangan nitrogen. Awalnya, Beclin-1 diidentifikasi sebagai Bcl-2 interacting gene yang diperlukan untuk kematian sel nonapoptotic.
Berbeda dengan LC3, penanda pembentukan autophagosome akhir, interaksi Beclin-1 berpartisipasi dalam tahap awal autophagy, mempromosikan nukleasi dari vesikel autophagic dan merekrut protein dari sitosol. Telah dilaporkan bahwa interaksi Beclin-1 monoallelically dihapus dalam banyak keganasan termasuk kanker payudara. Konsisten dengan keterlibatannya dalam jalur autophagic, bukti terbaru menunjukkan bahwa interaksi Beclin-1-Bcl-2 _ memainkan peran penting dalam cross-talk antara apoptosis dan autophagy dalam respon terhadap stres dalam sel mamalia.
Stres sitokin yang berlebihan serta hipoksia biasanya dikaitkan memiliki beberapa peran dalam patofisiologi PE17 dan kondisi ini juga diketahui berhubungan dengan penyimpangan di lingkungan rahim dan kinetika sel trofoblas. Kami menghipotesiskan bahwa lingkungan mikro hipoksia dan peningkatan stres sitokin dalam plasenta pasien dengan PE dapat menyebabkan aktivitas autophagic dalam plasenta yang memberikan kontribusi untuk patofisiologi PE.
Kami meneliti ekspresi autophagi terkait protein (LC3 dan Beclin1) pada plasenta manusia dan perubahan mereka dalam plasenta dari kehamilan dengan komplikasi PE parah. Untuk menjelajahi mekanisme regulasi autophagi terkait protein ini dalam trofoblas, kita lebih lanjut menganalisis perubahan protein ini dalam menanggapi hipoksia atau perlakuan sitokin menggunakan kultur JEG3 sel.
BAHAN DAN METODE
Studi Kependudukan dan Tissue Collection The Institutional Review Board untuk Penelitian Klinis di Samsung Medical Center menyetujui penelitian ini dan informed consent diperoleh dari masing-masing peserta. Plasenta dikumpulkan setelah melahirkan sesar, tanpa pekerja, dari wanita dengan kehamilan jangka normal (n = 8) serta dari wanita dengan PE berat (n = 11). Diagnosis PE berat didasarkan pada data berikut.
Tekanan darah 160/110 mm Hg atau lebih tinggi 2 kali setidaknya 6 jam terpisah, proteinuria (≥ 4 g dalam spesimen urin 24 jam atau ≥ 2 + pada 2 dipsticks random), disertai dengan adanya sakit kepala, visual yang terganggu, epigastrik, nyeri kuadran kanan atas , oliguria (<500 mL dalam 24 jam), edema paru, gangguan fungsi hati, trombositopenia, atau IUGR .
Setelah terkumpulnya sampel dari pertengahan bagian tengah antara lempeng basal dan membran chorionic seperti yang dijelaskan oleh Wyatt et al, jaringan plasenta yang patah dibekukan dalam nitrogen cair dan disimpan pada -80 º C atau ditanam dalam parafin untuk Immunostaining. Untuk menyelidiki

perubahan dalam protein-autophagy terkait selama memajukan usia kehamilan, plasenta dari aborsi atau kelahiran prematur (n = 16) juga diperoleh. Setiap kasus dengan kehamilan ganda, diabetes gestasional, hipertensi, dan korioamnionitis histologis dikeluarkan dari populasi penelitian.
Electron Microscopy
Jaringan plasenta difiksasi selama 12 jam dalam 2,5% glutaraldehid, dicuci dalam 0,1 M bufer fosfat (pH 7,4) dua kali selama 15 menit, dan difiksasi lagi dalam air 1% osmium tetroksida selama 1 jam. Setelah fiksasi, mereka didehidrasi dalam etanol, ditanamkan pada ok812, dan diinkubasi dalam oven semalam pada 37 º C. Jaringan kemudian dipotong (50-60 nm) dan diwarnai dengan uranil asetat selama 30 menit dan sitrat lead selama 10 menit. Pemeriksaan mikroskopik elektron dilakukan dengan menggunakan Hitachi 7100 transmisi elektron mikroskop (Jepang).
Reverse Transcriptase Polymerase Chain Reaction (RT-PCR) dan Real-Time PCR Kuantitatif
Total RNA diekstraksi dari jaringan plasenta menggunakan Trizol (Invitrogen Life Technologies, Carlsbad, CA) sesuai dengan instruksi pabrik. Satu mikrogram RNA total digunakan untuk transkripsi balik dengan hexamers acak menggunakan
Superscript III RT (Invitrogen). Kondisi berikut ini digunakan untuk PCR: denaturasi awal pada 94 º C selama 5 menit, diikuti oleh 30 sampai 35 siklus denaturasi pada
94 º C selama 30 detik, annealing pada 55 º C selama 30 detik, dan ekstensi pada 68 º C selama 10 menit dengan menggunakan polimerase Taq DNA Accuprime (Invitrogen).
Primer ( National Center for Biotechnology Information nomor aksesi dan ukuran produk ) yang digunakan untuk gen LC3 manusia adalah 5' - CAGCATGGTGAGTGTGTCCA - 3 ' (forward ) dan 5' - GCAGCTCAGTTCAGGAACCA - 3 ' (reverse ) untuk LC3A ( NM_181509 , 195 bp ) , 5' - GAGAGCAGCATCCAACCAAA - 3 ' (forward ) dan 5' - TCCGTTCACCAACAGGAAGA - 3 ' (reverse ) untuk LC3B ( NM_022818 , 185 bp ) , 5' - GCAATCAGACAAGAGGAAGT - 3 ' (forward ) dan 5' - TAGAGAGGATTGCAGGGTC - 3 ' ( terbalik ) untuk LC3C ( NM_001004343 , 386 bp ) , dan 5' - GGCGGACTATGACTTAGTTG -
3 ' (forward ) dan 5' - AAACAACAATGTGCAATCAA - 3 ' (reverse ) untuk β - aktin ( 238 bp ) . Semua hasil dinormalisasikan untuk β-aktin untuk mengkonfirmasi jumlah keseragaman RNA Template. Untuk real-time PCR kuantitatif, ABI Prism 7000 Sequence Detection System (PE Terapan Biosystems, Foster City, CA) digunakan dengan SYBR sistem deteksi hijau seperti yang dijelaskan sebelumnya. PCR dilakukan dalam rangkap tiga dengan volume reaksi 25 uL dari 5 uL cDNA, 12,5 uL SYBR Hijau PCR Master Mix, dan 0,3 mmol / L masing-masing primer. PCR dilakukan dengan menggunakan parameter siklus berikut: 2 menit pada 50 º C, 10 menit pada 95 º C, 40 siklus 15 detik pada 95 º C, dan 1 menit pada 60 º C. Primer (dan ukuran produk) yang digunakan untuk real-time PCR kuantitatif adalah 5'-ATCATCGAGCGCTACAAGGGTG-3 'maju) dan 5'-TCATGTTGACATGGTCCGGG-3' (reverse) untuk LC3A (85 bp), 5'-GCAAAACGCATTTGCCATCAC-3 '(depan) dan 5'-CAGTTTACAGTCAGGGCCGTTTTC-3 '(reverse) untuk LC3B (80 bp), dan 5'-CGCCGCTAGAGGTGAAATTC-3' (forward) dan 5'-CGAACCTCCGACTTTCGTTCT-

3 '(reverse) untuk 18S rRNA (100 basis poin). Semua hasil dinormalisasikan untuk 18S untuk memastikan jumlah keseragaman RNA Template.
Western Blotting Analisis, Imunohistokimia, dan Pewarnaan Protein Immunofluorescence diekstraksi menggunakan bufer RIPA (50 mmol / L Tris-Cl, 150 mmol / L NaCl, 1% NP-40, 0,5% natrium deoksikolat, dan 0,1% sodium dodesil sulfat [SDS] , pH 7,5) yang mengandung 1 mmol / L phenylmethylsulfonyl fluoride dan dihitung dengan Protein Bio-Rad Assay (Bio-Rad Laboratories, Hercules, CA). Sampel protein (30 mg) dipisahkan dengan elektroforesis gel SDSpolyacrylamide (12% untuk LC3 dan 8% untuk Beclin-1) dan kemudian dielectrotransfer ke membran nitroselulosa. Setelah memblokir dengan 5% susu kering tanpa lemak di TBS-T (1 TBS dengan 0,1% Tween 20), membran diinkubasi dengan antibodi primer (1:1000 untuk masing-masing) pada 4 º C dalam semalam. Untuk menilai LC3 ekspresi isoform dalam plasenta, kami juga menggunakan antibodi spesifik untuk LC3A dan LC3B di 1:500 dilusi untuk masing-masing (Novus Biologicals, Littleton, CO).
Setelah pencucian, blot diinkubasi dengan anti rabbit horseradish peroxidase antibodi sekunder terkonjugasi pada pengenceran 1:5000 selama 1 jam, dan dikembangkan dengan ECL (Amersham Biosciences Little Chalfont, UK). Immunostaining dari LC3 dilakukan pada parafin bagian tertanam menggunakan antibodi poliklonal kelinci terhadap LC3 (MBL, Nagoya, Jepang). Setelah deparaffination dan rehidrasi, bagian diinkubasi dalam 3% H2O2 dalam metanol selama 30 menit, dan kemudian dimicrowave. Setelah memblokir dengan 10% serum nonimmune, bagian diinkubasi dengan antibodi untuk LC3 pada pengenceran 1:1000 pada 4 º C semalaman dan diikuti dengan inkubasi dengan antibodi sekunder biotinylated (Santa Cruz Biotechnology, Santa Cruz, CA) selama 1 jam pada pengenceran 1:2000. Setelah inkubasi dalam peroksidase konjugasi streptavidin, kompleks antibodi divisualisasikan dengan diaminobenzidin tetrahydrochloride chromogen selama 10 detik.
Bagian itu di counterstained dengan hematoxylin, didehidrasi dan dipasang. Mikroskop immunofluorescence of-Beclin 1 telah dilakukan pada parafin bagian tertanam menggunakan antibodi poliklonal kelinci untuk Beclin-1 (Cell Signaling, Beverley, MA). Bagian yang diblokir diinkubasi dengan antibodi untuk Beclin-1 pada pengenceran 1:1000 pada 4 º C semalam dan diikuti dengan inkubasi dengan Alexa 594-kelinci IgG antibodi sekunder (Santa Cruz Biotechnology) selama 1 jam pada pengenceran 1:400 pada ruangan bagian suhu ruang itu di counterstained dengan DAPI, didehidrasi, dan dipasang.
Kultur dari JEG-3 Cell Line
Koriokarsinoma manusia JEG-3 garis sel diperoleh dari Culture Collection American Type (Rockville, MD). Garis sel dipertahankan dalam medium RPMI 1640 (Gibco BRL, Burlington, Ontario, Kanada) ditambah dengan 10% serum janin sapi yang mengandung campuran antibiotik pada 37 º C di bawah 5% CO2 atmosfer dilembabkan. Sekitar 1 105 sel per sumur, dari piring-6 juga tumbuh di RPMI 1640 medium, yang ditambah dengan 10% fetal bovine serum dan digunakan untuk transfeksi.
Medium kultur diganti dengan 10% serum janin sapi yang mengandung media MEM sekitar 4 jam sebelum transfeksi dan sel-sel ditempatkan dalam larutan 10% CO2 incubator.The terdiri dari setengah volume yang 2X BBS (N, N-bis [2 - hidroksietil asam-buffered]-2-aminoethanesulfonic garam, pH 6,95) (Sigma-Aldrich, St Louis, MO) dan setengah Volume 0.25 M CaCl2 (Sigma-Aldrich).

Larutan 2X BBS ditambahkan ke 5 ug GFP-LC3 penampungan DNA plasmid LC3B (microtubuleassociated protein 1 light chain 3 beta, Map1lc3b, baik yang disediakan oleh Dr Tamotsu Yoshimori, National Institute of Genetics di Mishima, Jepang) dan diinkubasi selama 30 menit. Campuran transfeksi kemudian ditambahkan ke sel-sel di bawah suasana CO2 5% pada 37 º C. Setelah 16 jam transfeksi, sel diterapi dengan 10 ng / mL tumor necrosis factor-α (TNF-α; Biosource International Inc, Camarillo, CA) atau kultur dalam ruang hipoksia (Thermo Electron, Marietta, OH) memberikan suasana hipoksia didefinisikan sebagai ~ 1% O2 (5% CO2, 10% H2, dan 85% N2) dengan pO2 <15 mm Hg pada 37 º C.
Media kultur sel berubah setiap 24 jam. Pada waktu tertentu, aktivitas autophagic dinilai dengan observasi punktuasi GFP-LC3 pewarnaan menggunakan mikroskop confocal laser scanning (CLSM, Radiance 2100 Rainbow, Bio-Rad, Hercules, CA).
Untransfected JEG-3 sel juga diobati dengan 10 ng / mL TNF-α atau kultur dalam ruang hipoksia untuk waktu tertentu dan perubahan LC3 dan Beclin-1 dinilai menyatakan analisis imunoblot.
Analisis Statistik
The Mann-Whitney U test dan uji Fisher digunakan untuk membandingkan karakteristik klinis dan perbedaan antara kelompok dengan mRNA dan analisis protein. Uji Spearman uji korelasi peringkat dilakukan untuk menilai perubahan LC3 dan Beclin-1 kadar protein selama memajukan kehamilan. Untuk data yang diperoleh dari budaya JEG-3 cell, kami menggunakan uji Kruskal-Wallis untuk membandingkan kadar protein-Beclin 1 dan LC3-II pada 12, 24, dan 48 jam dalam menanggapi hipoksia atau TNF-α dengan tingkat referensi pada waktu 0 poin. Hasilnya dianggap signifikan secara statistik ketika nilai P kurang dari 0,05.
HASIL
Karakteristik klinis dari peserta disajikan pada Tabel 1. Tidak ada perbedaan dalam usia ibu, indeks massa tubuh ibu atau neonatal gender. Wanita multipara lebih umum pada kelompok kontrol, mencerminkan fakta bahwa kebanyakan wanita hamil pada kelompok kontrol telah menjalani kelahiran sesar berulang. Seperti yang diharapkan, usia kehamilan saat melahirkan dan berat lahir neonatal secara signifikan lebih rendah pada kelompok PE parah dibandingkan dengan kelompok kehamilan normal.
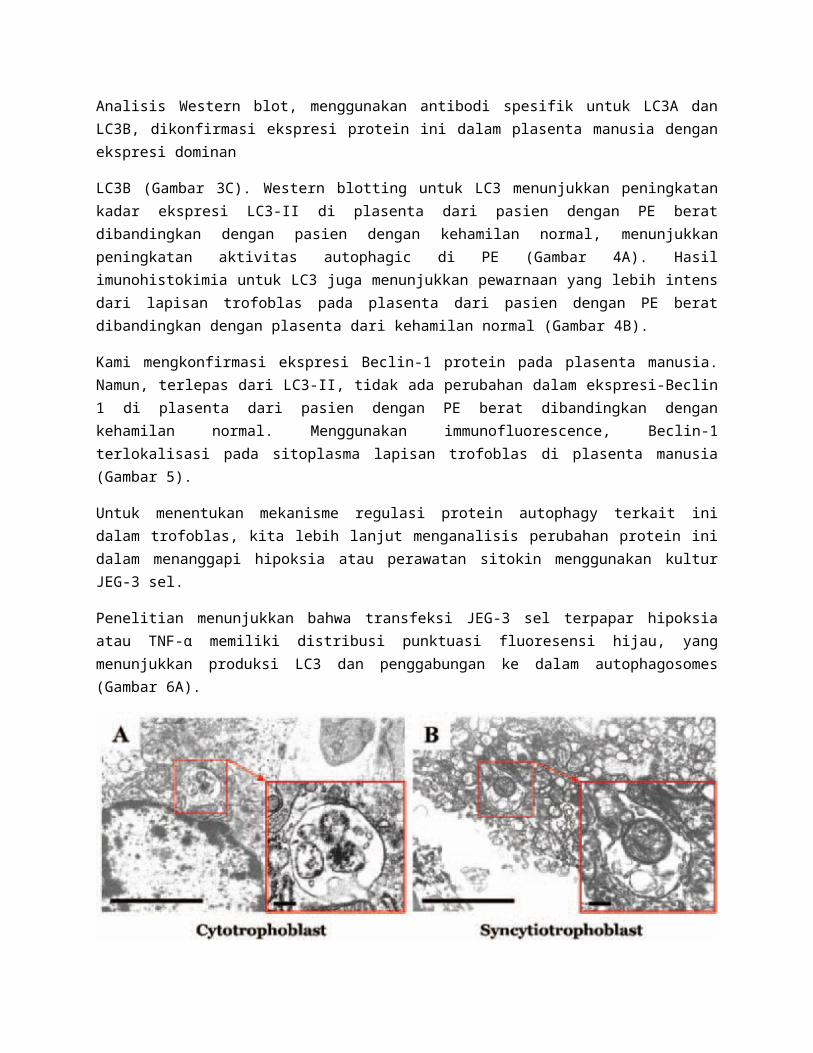
Dengan mikroskop elektron, vakuola autophagic diamati di kedua sitotrofoblas dan sinsitiotrofoblas (Gambar 1). Vakuloa Autophagic ini mengandung organel intracytoplasmic termasuk mitokondria. Morfologi vakuola autophagic agak bervariasi tergantung pada tahap pembentukan vesikel autophagic dan degradasi bahan intravacuolar.
Pertama, untuk menyelidiki apakah ada perubahan dalam tingkat LC3 dan ekspresi Beclin-1, pada plasenta manusia selama kehamilan, plasenta dari berbagai usia
kehamilan setelah terapeutik aborsi atau kelahiran prematur dinilai untuk protein ini. Plasenta manusia menyatakan baik LC3 dan Beclin-1 protein selama kehamilan dari trimester kedua . Tidak ada perubahan

yang signifikan dari tingkat LC3 dan Beclin-1 dalam plasenta yang diteliti sesuai dengan usia kehamilan (Gambar 2).
Kami mengevaluasi produksi isoform LC3 dalam plasenta manusia. Menggunakan RT-PCR, kami mengamati ekspresi mRNA dan LC3A LC3B mRNA (Gambar 3A) tapi tidak bisa mendeteksi LC3C mRNA dalam plasenta manusia (data tidak ditampilkan). Meskipun ekspresi mRNA LC3A di plasenta tidak berbeda antara 2 kelompok, ekspresi mRNA LC3B secara signifikan meningkat pada plasenta dari pasien dengan PE dibandingkan dengan kehamilan normal (Gambar 3B).
Analisis Western blot, menggunakan antibodi spesifik untuk LC3A dan LC3B, dikonfirmasi ekspresi protein ini dalam plasenta manusia dengan ekspresi dominan
LC3B (Gambar 3C). Western blotting untuk LC3 menunjukkan peningkatan kadar ekspresi LC3-II di plasenta dari pasien dengan PE berat dibandingkan dengan pasien dengan kehamilan normal, menunjukkan peningkatan aktivitas autophagic di PE (Gambar 4A). Hasil imunohistokimia untuk LC3 juga menunjukkan pewarnaan yang lebih intens dari lapisan trofoblas pada plasenta dari pasien dengan PE berat dibandingkan dengan plasenta dari kehamilan normal (Gambar 4B).
Kami mengkonfirmasi ekspresi Beclin-1 protein pada plasenta manusia. Namun, terlepas dari LC3-II, tidak ada perubahan dalam ekspresi-Beclin 1 di plasenta dari pasien dengan PE berat dibandingkan dengan kehamilan normal. Menggunakan immunofluorescence, Beclin-1 terlokalisasi pada sitoplasma lapisan trofoblas di plasenta manusia (Gambar 5).
Untuk menentukan mekanisme regulasi protein autophagy terkait ini dalam trofoblas, kita lebih lanjut menganalisis perubahan protein ini dalam menanggapi hipoksia atau perawatan sitokin menggunakan kultur JEG-3 sel.
Penelitian menunjukkan bahwa transfeksi JEG-3 sel terpapar hipoksia atau TNF-α memiliki distribusi punktuasi fluoresensi hijau, yang menunjukkan produksi LC3 dan penggabungan ke dalam autophagosomes (Gambar 6A).
Gambar 1. Gambar mikroskop elektron dari plasenta manusia.

Scanning kedua sitotrofoblas (A) dan sinsitiotrofoblas (B) dengan bidang daya rendah dan tinggi. Panah menunjukkan vakuola autophagic. Bentuk vakuola autophagic bervariasi tergantung pada tahap pembentukan autophagosome. Pada panel sebelah kanan, dalam vesikel membran ganda, organel sitoplasma termasuk mitokondria sedang dilisiskan. Pada panel sebelah kiri, autolysosome dibentuk dengan mitokondria rusak menyatakan tahap berikutnya adalah pembentukan autophagosome.
Perhatikan sinsitiotrofoblas yang berisi sitoplasma vacuolized dan banyak lagi autophagosomes dari sitotrofoblas. Bar skala, 1 um, skala bar insets, 100 nm.
Gambar 2. Ekspresi LC3 dan Beclin-1 pada plasenta manusia selama kehamilan.
Dengan blot Western, ditemukan bahwa plasenta manusia dinyatakan mengekspresikan LC3 dan Beclin-1 di semua usia kehamilan . Setelah analisis densitometri dan normalisasi untuk β-aktin, disimpulkan bahwa tidak ada perubahan signifikan dalam tingkat protein dari LC3 dan Beclin-1 sesuai dengan usia kehamilan dari trimester kedua kehamilan jangka panjang (P = tidak signifikan, oleh Spearman uji korelasi rank ). Blot representatif ditampilkan.

Gambar 3. Ekspresi LC3A dan LC3B dalam plasenta manusia dari kehamilan normal dan mereka dengan preeklamsia: reverse transcriptase polymerase chain reaction, RT-PCR (A), real-time qPCR (B), dan imunoblotting (C).
Ekspresi LC3B mRNA, tetapi bukan LC3A mRNA, secara signifikan meningkat pada plasenta dengan PE berat dibandingkan dengan kehamilan normal (data disajikan sebagai rata-rata ± standar error dari mean).
Analisis Western blot, menggunakan antibodi spesifik untuk LC3A dan LC3B, dikonfirmasi ekspresi protein ini dalam plasenta manusia dengan ekspresi dominan LC3B. Perhatikan bahwa bentuk belum diproses dari protein LC3A nyaris terdeteksi (C). Kasus representatif ditunjukkan untuk RT-PCR dan Western blot LC3A dan LC3B. * P <.05.
Selain itu, kami menemukan bahwa untransfected JEG-3 sel terkena hipoksia atau TNF-α menunjukkan perubahan yang berbeda dari LC3 dan Beclin-1 protein dalam program waktu tertentu. Dalam JEG-3 sel, hipoksia (O2 <1%) memicu peningkatan dalam ekspresi LC3-II (tanpa signifikansi statistik, P = .15) dengan penurunan yang signifikan dalam ekspresi-Beclin 1 (P <. 05). Sementara itu, perawatan TNF-α
menginduksi peningkatan yang signifikan dalam ekspresi LC3-II (P <.05) tanpa perubahan signifikan dalam ekspresi-Beclin 1 (Gambar 6B dan C).
Gambar 4. Ekspresi LC3 dalam plasenta manusia dari kehamilan normal dan preeklampsia.
A, analisis imunoblotting menunjukkan bahwa tingkat ekspresi LC3-II secara signifikan meningkat pada PE berat dibandingkan dengan kehamilan normal. Data disajikan sebagai rata-rata ± SEM. Blot representatif ditampilkan. * P <.05. B, Immunostaining untuk LC3 dalam vili plasenta. (a) Tidak ada kontrol antibodi primer, (b) kontrol negatif dengan antibodi primer preabsorbed dengan memblokir peptida, (c) plasenta jangka normal, dan (d) Plasenta dengan preeklamsia. LC3 Imunostaining dominan dalam sitoplasma trofoblas. Perhatikan perbedaan ditandai dalam ekspresi LC3 dalam plasenta dari PE dibandingkan dengan kehamilan normal (panel kiri, pembesaran asli 400;? Bar = 15 m;? Panel sebelah kanan, pembesaran asli 1000).

Gambar 5. Ekspresi-Beclin 1 pada plasenta manusia dari kehamilan normal dan preeklampsia.
A, analisis imunoblotting menunjukkan bahwa tingkat Beclin-1 ekspresi dalam plasenta preeklamsia mirip dengan kehamilan normal.
Data disajikan sebagai rata-rata ± SEM. Blot representatif ditampilkan. B, pewarnaan immunofluorescence menunjukkan Beclin-1 (merah) yang difus diterjemahkan ke sitoplasma lapisan sel trofoblas pada plasenta manusia (perbesaran asli? 400).

Gambar 6. Ekspresi LC3 dan Beclin-1 di JEG-3 sel setelah terapi dengan hipoksia dan TNF-α. A, GFP-LC3 fluoresensi di JEG-3 sel. JEG-3 sel ditransfeksi dengan GFP-LC3 dan kemudian dibiakkan dalam ruang hipoksia (O2 <1%) atau diolah dengan 10 ng / mL TNF-α. Sedangkan JEG-3 sel ditransfeksi dengan plasmid kosong tidak menunjukkan distribusi GFP-LC3, sel-sel transfected dengan GFP-LC3 plasmid menunjukkan punktuasi fluorescent hijau lebih banyak dan lebih kuat setelah 24 jam dalam menanggapi hipoksia atau TNF-α dibandingkan dengan nol, menunjukkan lebih banyak produksi LC3 dan penggabungan ke dalam autophagosomes. B, analisis Western blot LC3 di JEG-3 sel dalam menanggapi hipoksia atau TNF-α. Meskipun hipoksia menginduksi sedikit peningkatan dalam ekspresi LC3-II (P = .15, uji Kruskal-Wallis), TNF-α diinduksi peningkatan yang signifikan dalam ekspresi LC3-II dalam 48 jam terapi (* P <.05 , uji Kruskal-Wallis). C, analisis Western blot-Beclin 1 di JEG-3 sel dalam menanggapi hipoksia atau TNF-α.

Ekspresi-Beclin 1 dalam sel secara signifikan menurun selama 12 sampai 48 jam kultur dalam ruang hipoksia (* P <0,05, uji Kruskal-Wallis). TNF-α tidak menyebabkan perubahan signifikan dalam ekspresi-Beclin 1 pada waktu tertentu. Hasil yang sama diperoleh dalam 3 Data experiments.The independen disajikan sebagai rata-rata ± SEM.
Sementara itu, pengobatan TNF-α diinduksi peningkatan yang signifikan dalam ekspresi LC3-II (P <.05) tanpa perubahan signifikan dalam ekspresi-Beclin 1 (Gambar 6B dan C).
Komentar
Bagi pengetahuan kita, ini adalah studi pertama untuk mengidentifikasi autophagosomes dan protein-autophagy terkait (LC3 dan Beclin-1) pada plasenta manusia dan untuk menunjukkan ekspresi mereka berubah dalam hubungan dengan PE. Kami juga menunjukkan modifikasi protein ini dalam garis sel trofoblas dalam menanggapi hipoksia atau pengobatan sitokin.
Autophagy adalah proses dinamis yang memungkinkan sel untuk menghilangkan organel yang rusak dan mendaur ulang asam amnio. Metode standar untuk menilai autophagy adalah dengan visualisasi langsung dari autophagosomes menggunakan mikroskop elektron. Secara morfologi, kami mengamati bahwa konstituen sitoplasma trofoblas termasuk mitokondria yang diasingkan ke dalam membran ganda dari autophagosomes, yang kemudian menyatu dengan lisosom dan kemudian isinya terdegradasi. Namun, mikroskop elektron adalah prosedur memakan waktu dan tunduk pada interpretasi bias. Oleh karena itu, imunoblotting LC3-II lebih disukai karena metode yang sederhana dan kuantitatif untuk menentukan aktivitas khusus autophagic dalam sel mamalia.
Data kami menunjukkan bahwa tingkat LC3B mRNA dan ekspresi protein dari LC3-II meningkat pada plasenta dari pasien dengan PE berat dibandingkan dengan kehamilan normal, menunjukkan peningkatan aktivitas autophagic di PE.
Keterbatasan yang melekat dari penelitian ini dapat menjadi fakta bahwa usia kehamilan koleksi plasenta berbeda antara kelompok normal dan PE . Karena sebagian besar plasenta prematur diperoleh setelah persalinan prematur atau prematur ketuban pecah dini, di mana peradangan yang mendasari mungkin ada, sulit untuk mendapatkan plasenta prematur normal kontrol. Oleh karena itu, kami mengevaluasi ekspresi LC3 serta Beclin-1 pada plasenta dari pasien dengan PE berat pada trimester ketiga kehamilan dan digunakan plasenta _ sebagai kelompok kontrol. Keterbatasan kedua dari studi kami terletak bahwa jumlah relatif kecil dari plasenta untuk studi tentang kehamilan perubahan terkait usia.
Oleh karena itu, tampaknya lebih bijaksana untuk menyertakan lebih plasenta prematur yang normal di masa depan untuk memperkuat in vivo hasil ini dan untuk menjangkau berbagai kesimpulan.
LC3 awalnya copurified dengan mikrotubulus terkait 1A protein dan IB dari otak tikus. Ia memiliki asam amino 28% urutan homologi dengan ragi Apg 8/Aut7, yang penting untuk ragi autophagy. LC3 adalah protein pertama kali diidentifikasi pada membran autophagosome.

Tiga jenis urutan LC3 telah diidentifikasi sampai saat ini, LC3A, LC3B, dan LC3C. Berdasarkan hasil penelitian kami, ekspresi mRNA dari LC3 terbatas pada LC3A dan LC3B, tetapi tidak untuk LC3C. Setelah sintesis, LC3 mengalami modifikasi posttranslational ubiquitin-seperti sebelum pembentukan autophagosome. segera setelah
sintesis, daerah C-terminal dari LC3 dibelah untuk membentuk LC3-I, yang memiliki residu glisin pada akhir C-terminal. LC3-I terletak di sitosol, melainkan diproses lebih lanjut dan dimasukkan ke dalam membran autophagosome sebagai LC3-II.
Oleh karena itu, LC3-II secara khusus terkait dengan pembentukan autophagosome. Data kami menunjukkan bahwa tingkat LC3 diproses dalam plasenta manusia relatif rendah (Gambar 3C) dibandingkan dengan orang-orang dari LC3-I dan LC3-II (Gambar 4A). Temuan ini menunjukkan proses yang cepat dari LC3 terutama di PE setelah sintesis awal (Gambar 3B). The inducer yang paling umum dari LC3-II adalah kekurangan gizi, tapi stres selular lainnya seperti hipoksia juga bisa memicu autophagy. Selain itu, diferensiasi sel normal dikaitkan dengan peningkatan LC3-II. Dari catatan adalah bahwa LC3-II meningkat selama diferensiasi podocyte murine cells.24 Hasil penelitian kami menunjukkan bahwa hipoksia atau sitokin pengobatan meningkatkan ekspresi LC3-II di garis sel trofoblas. Bahkan, hipoksia menghambat fosforilasi dari jalur mTOR (target mamalia dari rapamycin), yang merupakan jalur sinyal hulu penting yang negatif mengatur autophagy.
Beclin-1 merupakan regulator penting dari autophagy.
Ekspresi sel mamalia Beclin-1 di MCF7 meningkatkan respon autophagic mereka terhadap kekurangan gizi. Penekanan-Beclin 1 dalam sel mamalia mengganggu autophagy dan sensitisasi sel menjadi kelaparan-diinduksi autophagy. Dalam penelitian kami, kami awalnya berspekulasi bahwa ekspresi-Beclin 1 di plasenta dari wanita dengan PE akan meningkat bila dibandingkan dengan plasenta normal.
Namun, tidak seperti protein LC3-II, kami menemukan bahwa tidak ada perubahan dalam tingkat plasenta Beclin-1 ekspresi dalam plasenta PE berat dibandingkan dengan plasenta normal. Untuk mengeksplorasi mekanisme yang mendasari untuk menjelaskan kami ex vivo data, kami melakukan percobaan in vitro menggunakan garis sel trofoblas dan menunjukkan perubahan signifikan dalam ekspresi LC3-II dan Beclin-1 tergantung pada jenis stimulasi (hipoksia atau TNF-α) .
Data kami menunjukkan bahwa TNF-α daripada hipoksia memicu up-regulasi yang signifikan dari LC3-II sejalan sel trofoblas. Sebaliknya, hipoksia daripada TNF-α memicu downregulation signifikan-Beclin 1. Pengaruh hipoksia terhadap ekspresi LC3-II dan Beclin-1 adalah serupa setelah pengobatan JEG-3 sel dengan CoCl2 (data tidak ditampilkan).
Meskipun Beclin-1 diketahui menginduksi autophagy, itu diperdebatkan apakah ekspresi-Beclin 1 meningkatkan respon autophagic. Sebagai contoh, Yan et al23 menunjukkan
bahwa ekspresi Beclin-1, bersama dengan LC3-II, meningkat ketika miokardium iskemik kronis babi diinduksi autophagy, sedangkan Martinet et AL21 menemukan bahwa kelaparan-diinduksi autophagy tidak terkait dengan perubahan dalam ekspresi Beclin-1 di HepG2 sel.
Selain itu, Tsujimoto et al mengusulkan bahwa penarikan asam amino atau stimulasi limfokin mempertahankan tingkat rendah atg5/atg6 selama autophagy untuk bertahan hidup, ini konsisten dengan

kami in vitro data. Oleh karena itu, aktivitas autophagic meningkat di plasenta dari wanita dengan PE mungkin mencerminkan respon kelangsungan hidup trofoblas terhadap rangsangan inflamasi berlebihan dan stres oksidatif pada plasenta PE.
Namun, dalam data kami, tetap ada pertanyaan mengapa ekspresi Beclin-1 penurunan JEG-3 sel dalam menanggapi hipoksia meskipun ekspresi Beclin-1 pada plasenta
tidak berbeda antara normal dan PE kehamilan.
Kami tidak yakin persis mengapa perbedaan ini terjadi. Namun demikian, ini mungkin sebagian disebabkan karena perbedaan antara in vivo oksigen lingkungan dan in vitro kondisi eksperimental kami. Telah disarankan oleh peneliti lain yang mungkin ada variasi spasial dan temporal dari tekanan oksigen dalam ruang intervillous dalam PE plasenta terganggu.
Selain itu, Tsujimoto et al mengusulkan bahwa penarikan asam amino atau stimulasi limfokin mempertahankan tingkat rendah atg5/atg6 selama autophagy untuk bertahan hidup, ini konsisten dengan kami in vitro data. Oleh karena itu, aktivitas autophagic meningkat di plasenta dari wanita dengan PE mungkin mencerminkan respon kelangsungan hidup trofoblas terhadap rangsangan inflamasi berlebihan dan stres oksidatif pada plasenta PE.
Singkatnya, kami menunjukkan bahwa ekspresi LC3, tetapi tidak Beclin-1, meningkat pada plasenta dari kehamilan dengan komplikasi PE berat dibandingkan dengan
plasenta yang normal menunjukkan peningkatan aktivitas autophagic di plasenta dari pasien dengan PE.
Selain itu, temuan hasil in vitro menunjukkan bahwa respon autophagic ini dapat diatur oleh trofoblas mikro (yaitu, hipoksia atau sitokin). Secara keseluruhan, ekspresi diubah protein-autophagy terkait ini di PE dapat terlibat dalam patofisiologi gangguan ini dan ini menjamin agenda penelitian mendatang penyelidikan intensif.