diagnosis penyebab mati meranggas pala di aceh...
TRANSCRIPT

LAPORAN AKHIR TAHUN
PENELITIAN DISERTASI DOKTOR
DIAGNOSIS PENYEBAB MATI MERANGGAS PALA
DI ACEH SELATAN
Tahun ke 1 dari rencana 1 tahun
Ir. Susanna, M.Si. 0030116801
Dibiayai oleh:
Direktorat Riset dan Pengabdian Masyarakat
Direktorat Jenderal Penguatan Riset dan Pengembangan
Kementerian Riset, Teknologi, dan Pendidikan Tinggi
Sesuai dengan Kontrak Penelitian
Nomor: 105/SP2H/LT/DPRM/IV/2017 tanggal 3 April 2017
UNIVERSITAS SYIAH KUALA
OKTOBER 2017


ii
RINGKASAN
Mati meranggas tanaman pala di Aceh Selatan telah memusnahkan ribuan hektar
perkebunan rakyat selama 15 tahun terakhir. Penyakit tersebut menyerang dan
menimbulkan kematian baik pada tanaman tua, muda, maupun pembibitan. Sampai saat
ini penyebab penyakit tersebut masih diperdebatkan dan kajian ataupun publikasi tentang
penyebabnya belum ada. Pengelolaan penyakit yang baik harus dibekali dengan
pengetahuan tentang patogen penyebab, begitu juga pada kasus mati meranggas pada
tanaman pala yang terdapat di Aceh Selatan. Penelitian bertujuan untuk membuktikan
penyebab penyakit mati meranggas tanaman pala di Aceh Selatan. Identifikasi patogen
dilakukan berdasarkan gejala tanaman terserang di lapangan dan yang muncul dari hasil
uji Postulat Koch berikut hasil pengamatan makro dan mikroskopik isolat yang berasal
dari berbagai bagian tanaman pala yang terserang serta analisa tanah. Selain itu juga
dilakukan analisa molekuler biakan murni patogen. Bagian-bagian tanaman terserang
berikut tanah diambil dari beberapa lokasi penanaman pala di Aceh Selatan. Postulat
Koch dilakukan dengan menggunakan miselia biakan murni patogen. Penularan pada uji
postulat Koch dilakukan dengan cara menempelkan miselia pada bibit pala yang berumur
satu tahun. Hasil penelitian menunjukkan bahwa terdapat empat genus cendawan patogen
(Botryodiplodia theobromae, Fusarium solani, Graphium euwallaceae., and Rigidoporus
microporus) dari enam cendawan yang berasosiasi dengan penyakit mati meranggas
sebagai penyebab penyakit. Namun, B. theobromae merupakan cendawan dominan yang
berasosiasi dengan penyakit mati meranggas pada tanaman pala di Aceh Selatan. Luaran
penelitian yang menjadi target adalah draf disertasi yang disetujui dan publikasi ilmiah
dalam jurnal internasional bereputasi tentang patogen penyebab penyakit mati meranggas
pada tanaman pala di Aceh Selatan.

iii
PRAKATA
Syukur Alhamdulillah penulis panjatkan ke Hadirat Allah Subhanahuwata’ala,
karena dengan rahmatNya penulis dapat menyelesaikan Penelitian Disertasi Doktor dan
penulisannya yang berjudul “Diagnosis Penyebab Mati Meranggas Pala di Aceh Selatan”.
Penelitian ini merupakan salah satu bagian dari disertasi penulis selama mengikuti
program doktoral (S3) di Institut Pertanian Bogor.
Selama melakukan penelitian banyak halangan dan rintangan yang penulis hadapi,
namun dengan kesabaran, kesungguhan dan keiklasan, akhirnya semua rintangan tersebut
dapat teratasi. Pada kesempatan ini penulis ingin mengucapkan terima kasih kepada
Rektor Universitas Syiah Kuala dan Dekan Fakultas Pertanian Universitas Syiah Kuala
yang telah member izin penulis untuk mengikuti program doktor (S3), Lembaga
Penelitian dan Pengabdian Pada Masyarakat Universitas Syiah Kuala yang telah memberi
kesempatan penulis melakukan Penelitian Disertasi Doktor pada Tahun 2017, Direktorat
Riset dan Pengabdian Masyarakat, Direktorat Jenderal Penguatan Riset dan
Pengembangan, Kementerian Riset, Teknologi, dan Pendidikan Tinggi yang telah
membiayai penelitian disertasi doktor ini.
Tak lupa pula penulis mengucapkan terima kasih yang tak terhingga kepada Staf
Pengajar pada Prodi Proteksi Tanaman Fakultas Pertanian Unsyiah beserta tenaga
administrasi yang telah memberikan dukungan kepada penulis, Staf Pengajar Podi
Fitopatologi Departemen Proteksi Tanaman Institute Pertanian Bogor, Staf/Karyawan
Laboratorium Perlindungan Hutan, Pusat Penelitian dan Pengembangan Hutan Bogor
yang telah memberikan fasilitas laboratorium, dan teman-teman seperjuangan sebagai
anggota Laboratorium Mikologi Tumbuhan Departemen Proteksi Tanaman, Institut
Pertanian Bogor yang telah membantu dan mendukung penulis selama penelitian sampai
penulisan laporan.
Akhir kata semoga hasil penelitian yang tertuang dalam laporan Penelitian
Disertasi Doktor ini dapat bermanfaat bagi pencinta ilmu pengetahuan terutama bagi
pemerintah dan masyarakat Aceh selatan yang sedang mengalami permasalahan dalam
bidang penyakit tanaman pala.
Banda Aceh, Okrober 2017
Penulis

iv
DAFTAR ISI
RINGKASAN ii
PRAKATA iii
DAFTAR ISI iv
DAFTAR TABEL v
DAFTAR GAMBAR v
DAFTAR LAMPIRAN v
BAB 1 PENDAHULUAN 1
BAB 2 TINJAUAN PUSTAKA 4
2.1 Diagnosis Penyakit 4
2.2 Teknik DNA untuk Identifikasi Cendawan 4
2.3 Penyebab Penyakit 6
BAB 3 TUJUAN DAN MANFAAT PENELITIAN 10
3.1 Tujuan 10
3.2 Manfaat Penelitian 10
BAB 4 METODE PENELITIAN 11
4.1 Tempat dan waktu 11
4.2 Bahan dan Metode 12
BAB 5 HASIL DAN PEMBAHASAN 15
5.1 Uji Postulat Koch 15
5.2 Identifikasi Morfologi dan Molekuler Patogen 17
BAB 6 KESIMPULAN DAN SARAN 19
6.1 Kesimpulan 21
6.2 Saran 21
DAFTAR PUSTAKA 19
LAMPIRAN 23

v
DAFTAR TABEL
Tabel 4.1 Sasaran, luaran, dan indikator capaian
kegiatan penelitian selama 2 tahun 11
Tabel 5.1 Asosiasi antar tanda dan gejala penyakit
pada pohon pala di Aceh Selatan 14 15
Tabel 5.2 Gejala yang ditimbulkan cendawan hasil isolasi 17
Tabel 5.3 Identifikasi cendawan patogen berdasarkan ITS
yang dibandingkan dengan data GenBank NCBI 20
DAFTAR GAMBAR
Gambar 5.1 Hasil inokulasi buatan pada bibit tanaman pala; inokulasi
Botryodiplodia sp.(A); inokulasi Graphium sp. (B);
inokulasi Fusarium solani (C); inokulasi Rigidoporus sp. (D);
Tanpa patogen (K). 16
Gambar 5.2 Morfologi koloni pada medium PDA dan struktur mikroskopis
cendawan patogen asal pala. 18
Gambar 5.3 Visualisasi elektroforesis hasil PCR 4 isolat cendawan dengan
menggunakan primer ITS1 dan ITS4. M= Marker;
1. Botryodiplodia sp.; 2. Graphium sp.; 3. F. solani;
dan 4. Rigidoporus sp. 19
DAFTAR LAMPIRAN
Lampiran 1 Instrumen Penelitian 25
Lampiran 2 Personalia Tenaga Peneliti 31
Lampiran 3 Publikasi Ilmiah Internasional 36
Lampiran 4 Artkel Jurnal Internasional 38

1
BAB 1. PENDAHULUAN
Pala merupakan salah satu komoditas perkebunan yang memiliki peran penting
dalam perekonomian nasional, yang memberikan kontribusi besar dalam memenuhi
kebutuhan minyak atsiri dalam negeri maupun dunia yaitu minyak atsiri pala. Sebagai
negara pemasok pala terbesar di dunia, Indonesia berkontribusi sebesar 70% -75%,
sedangkan sisanya 20% berasal dari Grenada dan 5% dari Sri Lanka, Trinidad, dan
Tobago. Sebagai pasar dunia bagi komoditas pala adalah Amerika Serikat, Inggris, dan
Jerman (Hadad dan Firman 2003). Oleh karena itu, upaya pengembangan dan
peningkatan produktivitas pala terus dilakukan. Namun dalam pembudidayaan tanaman
tersebut tidak terlepas dari gangguan hama dan penyakit, sehingga tanaman pala yang
dikembangkan oleh rakyat dapat diserang oleh berbagai patogen. Kasus ini terjadi pada
tanaman pala di Kabupaten Aceh Selatan.
Kabupaten Aceh Selatan merupakan salah satu sentra produksi pala di Indonesia.
Produksi pala dari wilayah ini berkontribusi besar dalam memenuhi kebutuhan pala
domestik maupun luar negeri. Luas area tanaman pala di Kabupaten Aceh Selatan pada
tahun 1994 mencapai 11 245 ha dengan rata-rata produktivitas buah pala utuh sebesar 8.2
ton/ha. Pada tahun 2003 luas arealnya menurun menjadi 9 843 ha, rata-rata total
produktivitasnya mengalami penurunan menjadi 1.1 ton/ha. Pada tahun 2014 nilai rata-
rata produktivitas pala Aceh semakin menurun menjadi 0.7 ton/ha (Dishutbun Kabupaten
Aceh Selatan, 2014). Umumnya perkebunan pala di Aceh Selatan adalah perkebunan
rakyat yang pengelolaan kebunnya kurang baik. Upaya peningkatan produksi pala
menciptakan pola tanam pala secara monokultur dengan pola dan jarak tanam (3 – 5 m)
yang tidak teratur. Kondisi tersebut menimbulkan ekosistem yang tidak seimbang,
sehingga ketersediaan makanan bagi hama dan penyakit cukup banyak.
Ketidakseimbangan ekosistem tersebut menyebabkan sering terjadi ledakan (outbreak)
hama dan penyakit. Banyak faktor yang menyebabkan menurunnya produktivitas
tanaman pala, antara lain: tehnik budidaya yang minimal, tanaman telah tua, juga
serangan hama dan penyakit yang akhir-akhir ini semakin meningkat dengan lingkungan
yang mendukung untuk infeksi menyebabkan penyakit terus berkembang. Sampai saat
ini beberapa cara pengendalian telah dilakukan, namun belum memperlihatkan hasil yang
signifikan.

2
Menurut laporan dinas kehutanan dan perkebunan Aceh Selatan, penyakit yang
lima belas tahun terakhir ini menyerang ribuan hektar perkebunan pala adalah mati
meranggas. Penyakit tersebut menyerang semua perkebunan pala yang ada di Aceh
Selatan sejak tahun 1998. Survey awal penulis menemukan dua macam gejala serangan,
yang pertama dimulai dengan layunya daun pada sebagian ranting, mengeriting, berwarna
kuning kemudian berubah menjadi coklat dan mengering yang sering diistilahkan mati
ranting (dieback). Kemudian berlanjut ke ranting-ranting dari satu cabang ke cabang
lainnya, mati secara bertahap, namun daun-daun tersebut masih tetap menggantung
selama beberapa minggu yang akhirnya gugur dan meranggas. Bila kulit kayu dikelupas
terlihat miselia cendawan, saat kambium disayat terlihat warna coklat kehitaman
(nekrotik). Selain itu ternyata pada tanaman tersebut juga terserang penggerek batang.
Pada akar, bila dibongkar terlihat hifa cendawan berwarna putih. Gejala yang kedua daun-
daun menguning berlanjut dari satu cabang ke cabang lain dan layu seluruhnya kemudian
daun gugur dan tanaman mati meranggas. Ketika pangkal batang dipotong melintang
terdapat lingkaran kambium berwarna coklat kehitaman. Akibat serangan penyakit ini
dapat menurunkan hasil sampai 70% (Harni et al., 2011). Penyakit tersebut menyerang
dan menimbulkan kematian baik pada tanaman di pembibitan, tanaman muda yang belum
dan telah menghasilkan, serta tanaman tua. Sampai tahun 2016 penyebab kematian
tanaman pala tersebut masih belum pasti dan diperdebatkan. Dikalangan petani penyakit
ini sudah sampai pada taraf meresahkan karena belum dapat dikendalikan dan kasus
serangannya cenderung bertambah luas. Pengetahuan mengenai patogen penyebab sangat
penting, terutama untuk penyakit-penyakit baru. Penyusunan strategi pengendalian yang
berkelanjutan biasanya harus didasarkan pada sifat patogen bersangkutan.
Laporan mengenai penyebab mati meranggas pada tanaman pala belum pernah
dipublikasikan, namun dari gejala yang terlihat dilapangan mirip dengan penyakit Elm
Belanda (Ducth Elm Disease) yang disebabkan oleh Ophiostoma novo-ulmi (Agrios,
2005), kanker batang cengkeh di Sulawesi yang disebabkan oleh Ceratocystis sp. (Van
Wyk et al., 2004), serangan Ceratocystis pirilliformis pada kayu putih di Australia
(Barnes et al., 2003), kanker batang yang disebabkan oleh Ceratosystis pada perkebunan
kopi, coklat, jeruk (Van Wyk et al., 2009), juga kayu putih di Venezuella (Van Wyk et
al., 2009). Serangan penyakit pada tanaman pala pernah terjadi di India yang dilaporkan
oleh Rahman et al. (1981) bahwa tanaman tersebut terserang penyakit akar disebabkan
oleh Cylindrocladium camelliae. Layu yang disebabkan oleh Ceratocystis merupakan

3
penyakit penting pada tanaman coklat di Amerika tengah dan Amerika Selatan
(Engelbrecht et al., 2007), pada Eucalyptus grandis di Afrika Selatan (Roux et al., 2004),
mangga di Oman dan Pakistan (Van Wyk, 2007; Fateh et al., 2006), juga spesies tanaman
hutan kayu putih dan akasia di Australia (Raux dan Wingfield 2009). Mamle dan Jolanda
(2015) menemukan Fusarium solani yang menyebabkan layu pada tanaman kapok randu
(Ceiba petandra) dan Botryosphaera sp. yang menyebabkan kanker batang pada kayu
putih di Ghana. Selanjutnya gejala serangan yang mirip dengan penyakit tersebut
diperlihatkan oleh tanaman yang terserang oleh patogen akar yang pernah dilaporkan oleh
Kaewchai et al. (2009), yang melakukan penelitian pada pertanaman karet di Thailand
Selatan dan di Sri Lanka (Madusha et al. 2013) yang disebabkan oleh R. microsporus,
juga pada tanaman akasia di Semenanjung Malaysia (Farid et al. 2007).
Berdasarkan uraian diatas ingin dilakukan serangkaian penelitian mengenai
penyebab mati meranggas pada tanaman pala di Kabupaten Aceh Selatan, identifikasi
patogen baik secara konvensional (postulat Koch) yang sering bergantung pada
identifikasi gejala penyakit, isolasi dan kultur dari organisme, juga deteksi dan
identifikasi cepat dengan teknik DNA secara molekuler. Hasil ini akan menjadi informasi
untuk mendapatkan teknik pengendalian yang tepat dalam mengendalikan penyakit yang
saat ini menjadi masalah besar pada tanaman pala di Aceh Selatan.

4
BAB 2 TINJAUAN PUSTAKA
2.1 Diagnosis Penyakit
Diagnosis merupakan suatu proses untuk mengidentifikasi suatu penyakit tanaman
melalui gejala dan tanda penyakit yang khas, termasuk faktor-faktor lain yang
berhubungan dengan proses pembentukan penyakit tersebut. Gejala merupakan
karakteristik fisik dari penyakit yang diekspresikan oleh tanaman. Gejala dapat termasuk
layu, puru, kanker, busuk, nekrosis, klorosis, dan decline. Tanda adalah struktur nyata
dari patogen yang menyebabkan penyakit. Tanda dapat termasuk tubuh buah cendawan
(seperti mushroom atau piknidia), miselia, lendir bakteri, keberadaan nematode atau
serangga, atau adanya lubang-lubang serangga yang tertimbun oleh serbuk kayu.
Diagnosis penyakit yang benar diperlukan untuk merekomendasikan cara pengendalian
yang tepat dan harus dilakukan dalam suatu survey penyakit tanaman (Sinaga, 2003).
Agar hasil diagnosis akurat, diperlukan pembuktian dengan menggunakan postulat
Koch, sebagai berikut:
1. Patogen yang diduga harus selalu berasosiasi pada tanaman yang sakit.
2. Patogen tersebut harus dapat diisolasi dan ditumbuhkan sebagai biakan murni.
3. Biakan murni tersebut jika diisolasikan ke tanaman sehat, harus menghasilkan
gejala dan tanda penyakit yang sama.
4. Bila penyebab penyakit di reisolasi dari tanaman yang diinokulasi tersebut, akan
dihasilkan biakan murni yang sama dengan penyebab yang diisolasi dari tanaman
sakit yang di diagnosis.
2.2 Teknik DNA untuk Identifikasi Cendawan
Teknik DNA merupakan alat yang berharga dalam mengidentifikasi cendawan,
yang sangat berguna untuk sarana penentuan identifikasi cendawan yang tidak
memproduksi tubuh buah, karakter taksonomi morfologi yang membentuk dasar deskripsi
spesies. Bila beberapa cendawan dapat tumbuh pada biakan dengan mudah dan
menghasilkan karakteristik fitur mikroskopik yang membantu dalam identifikasi
taksonomi, yang susah tumbuh dalam biakan atau ketika tumbuh pada biakan gagal
memproduksi fitur sebagai kunci perbedaan. Kebutuhan untuk identifikasi cepat, sebagai
aplikasi karantina dan biosekuriti dapat juga menggunakan teknik DNA.

5
Ada beberapa teknik yang dapat digunakan untuk identifikasi cendawan.
Pemilihan metode yang paling tepat tergantung pada aplikasi dan jumlah sampel. Namun,
dasar untuk semua metode adalah menggunakan penanda DNA untuk menghubungkan
cendawan yang belum diketahui untuk deskripsi secara penuh. Identifikasi berbagai
cendawan tergantung pada seperangkat specimen acuan yang telah diperiksa dan di
identifikasi dengan taksonomi mikologi. Metode untuk identifikasi DNA cendawan terdiri
dari: Southern blotting; PCR (polymerase chain reaction); PCR-RFLP (PCR restriction
fragment length polymorphism); RAPD (random amplified polymorphic DNA); T-RFLP
(terminal restriction fragment length polymorphism); Sekuensing DNA; dan microarrays.
Langkah pertama untuk metode ini adalah ekstraksi DNA dan purifikasi (Glen, 2006).
PCR dikembangkan pada tahun 1980-an dan memanfaatkan enzim DNA
polymerase termostabil dari bakteri tahan panas sehingga enzim dapat bertahan pada
siklus suhu yang diperlukan untuk PCR. PCR digunakan untuk mengkopi bagian kecil,
biasanya sampai 1 kbp dari genom DNA. Dua primer, sekitar 20 bp oligonukleotida,
dengan urutan basa komplementer terhadap fragmen kecil dari DNA target. Pemisahan
untaian DNA template diperlukan sebelum primer tersebut mengikat template, dan ini
dicapai dengan pemanasan campuran reaksi, kemudian didinginkan untuk mempromot
penempelan primer dan dipanaskan lagi untuk mencapai suhu optimum untuk DNA
polymerase terhadap perpanjangan primer, membuat komplemen reverse mengkopi
untaian DNA template (Yuwono, 2006).
Saat ini, metode yang paling cocok untuk mendeteksi/mengidentifikasi cendawan
adalah dengan menggunakan PCR dan sekuensing DNA, sebagai primer spesifik spesies
telah dikembangkan hanya untuk beberapa dari banyak spesies busuk akar atau busuk
pangkal. Taksonomi spesies polypore berasosiasi dengan busuk hati, busuk pangkal dan
busuk akar, dalam banyak kasus membutuhkan klarifikasi dan sekuensing DNA untuk
penyelesaian penentuan beberapa spesies. Menurut Toju et al., 2012, primer ITS (Internal
Transcibed Spacer) cocok digunakan dalam mengidentifikasi sampel lapangan untuk
cendawan dari kelompok Ascomycetes dan Basidiomycetes berbasiskan pada untaian
DNA dengan primer ITS 4 dan ITS 5. Wilayah ITS adalah studi yang berguna untuk
karakterisasi dalam cendawan karena empat alasan yaitu: 1) wilayah ITS yang relative
singkat (500 – 800 bp) dan dapat dengan mudah di amplifikasi dengan PCR
menggunakan primer tunggal universal; 2) multicopy dari rDNA membuat wilayah ITS
mudah di amplifikasi dari sampel DNA yang kecil atau encer; 3) wilayah ITS mungkin

6
sangat bervariasi antar spesies berbeda secara morfologi; dan 4) PCR menghasilkan
probe-probe spesies spesifik ITS dapat diproduksi secara cepat. Wilayah ITS ini telah
digunakan untuk karakterisasi, identifikasi, dan deteksi cendawan seperti R. microporus
yang diisolasi dari pertanaman karet di Thailand (Kaewchai et al. 2010), Ceratocystis
fimbriata pada Eucalyptus di Afrika Tengah (Raux et al. 2000), C. fimbriata pada family
Araceae di Hawaii (Thorpe et al. 2005), C. fimbriata yang menyerang Tectona grandis di
Brazil (Firmino et al. 2012), penyebab busuk buah dari Passiflora edulis di Brazil
(Firmino et al. 2013), C. fimbriata pada tanaman mango di Oman (Van Wyk et al. 2005),
C. larium pada Styrax benzoin di Indonesia (Van Wyk et al. 2009).
2.3 Penyebab Penyakit
Botryodiplodia theobromae
Cendawan ini termasuk filum Ascomycota (telemorf Botryosphaeria rhodinae).
Koloni tumbuh menyebar denga cepat, berwarna gelap kehitaman, konidia hialin pada
umur muda dan berubah coklat gelap pada umur tua bersekat satu yang sering disebut
binukleat, berbentuk elips, dan diproduksi pada piknidia askostroma. Piknidia
mengandung sel konidiogen yang berkembang secara parafisis. Konidia memiliki
dinding yang tebal dan berukuran 22,5 x 12,5 µm. Kondisi tumbuh yang optimum untuk
pertumbuhan B. theobromae pada media adalah pada suhu 280C (Ismail et al. 2012).
Cendawan ini memiliki kisaran inang lebih dari 280 spesies tanaman. Beberapa contoh
tanaman yang menjadi inang dari B. theobromae adalah jeruk, eukaliptus, mangga, kakao,
pinus, mimba, dan kacang mete.
Penetrasi B. theobromae ke dalam jaringan tanaman menggunakan enzim dan
toksin yang dihasilkan patogen. Enzim utama yang dihasilkan oleh cendawan ini adalah
β-glukosidase dan selulase. Enzim dimanfaatkan olah B. theobromae untuk melunakkan
jaringan batang tanaman inang sedangkan toksin dimanfaatkan untuk mematikan sel
tanaman. Toksin memengaruhi permeabilitas membrane sel dengan cara menonaktifkan
dan menghambat enzim tanaman atau menghentikan aktivitas enzim tanaman yang
terkait. Enzim dan toksin yang diproduksi oleh B. theobromae digunakan untuk
menembus titik infeksi yang berada pada epidermis dan sel parenkim pada korteks (Al-
Saadoon et al.2012).
Gejala yang timbul akibat infeksi B. theobromae bergantung pada jenis tanaman
dan kondisi iklim. Beberapa gejala yang dapat dtimbulkan adalah nekrotik pada batang

7
dan cabang, hawar, mati pucuk, pembusukan pada batang, umbi, dan akar (Shahbaz et al.
2009; Adandonon 2014). Menurut Latha et al. (2009), tanaman yang terinfeksi B.
theobromae mengalami penguningan dan kelayuan paa daun terutama daun-daun bagian
bawah. Daun-daun tersebut menghitam dan membusuk, diikuti pembusukan akar dan
kematian tanaman. Pembelahan akar dan batang yang terinfeksi menunjukkan perunahan
warna menjadi coklat kehitaman pada jaringan vaskuler. Gejala pada batang akan
muncul sekitar satu bulan setelah patogen melakukan inokulasi. Kondisi fisik yang cocok
untuk penyebaran dan perkembangan B. theobromae dari sumber inokulumnya adalah
kondisi kelembaban yang tinggi (RH di atas 80%, titik kritis 40%) terutama pada keadaan
curah hujan yang tinggi dan suhu yang hangat (25 - 30 oC, titik kritis 20
oC).
Fusarium Solani
F. solani memiliki koloni berwarna putih seperti kapas, namun dapat juga
berkembang menjadi warna pink atau violet seperti kebanyakan genus Fusarium. Koloni
floccose, menyebar, berlendir dan relatif. F. solani memiliki hifa udara yang
menghasilkan konidiofor lateral, dan cabang konidiofor yang terbentuk akan membentuk
monofialid tipis dan memanjang yang menhasilkan konidia. Konidia dari F. solani terdiri
dari dua jenis yaitu makrokonidia dan mikrokonidia. Makrokonidia berbentuk sedikit
melengkung dan hialin yang terdiri dari 3 - 5 septa. Mikrokonidia memiliki sel lebih
tebal, berbentuk oval atau silindris, hialin dan smooth serta tiak bersepta. Namun
mikrokonidia juga ada yang melengkung dan memiliki septa sampai 2. F. solani
membentuk klamidospora yang umumnya terjadi pada kondisi sub optimal.
Graphium sp.
Graphium sp. merupakan genus cendawan dari family Microascaceae. Cendawan
ini termasuk cendawan patogen tumbuhan. Graphium sp. termasuk dalam grup
Hypomycetes dan memiliki sekitar 20 spesies. Cendawan ini terdapat di dalam tanah,
sisa-sisa tanaman, substrat berkayu, pupuk kandang, dan air tercemar. Struktur sporulasi
dari Graphium sp. membentuk sinema, yang terdiri dari ikatan konidifor seperti buket
bunga, memiliki konidia hialin yang dihasilkan dari annelida-annelida. Cendawan ini
menimbulkan warna kebiruan pada kayu-kayuan . fase sempurna dari Graphium sp. yang
telah ditemukan adalah Ceratocystis sp.

8
Rigidoporus microporus (sinonim: R. lignosus, Fomes lignosus)
Tubuh buah besar (lebar sampai 20 cm), relatif tipis, annual jarang perennial,
kasar, bergerombol, sering imbricate, permukaan atas konsentris berkerut, awalnya
orange berubah merah kemudian coklat, beludru samar-samar, halus, dan memudar,
permukaan bawah terang, orange berubah coklat, akhirnya memucat, pori-pori halus (6 -
9 per mm). Minomitik, hifa generative tipis atai berdinding tebal, dengan dinding silang
(septa), tanpa clam, hialin. Hymenium dengan sistidiol, basidiospora sub-globuse,
berdinding tipis, berwarna, halus, 3,5 - 4,5 x 3,5 - 4 µm. Tanda penyakit akar pohon yang
ditanam, putih, miselium bercabang dan busuk putih. Koloni cendawan di media PDA
(Potato Dextrose Agar) berwarna putih an tipis dengan hifa hialin, bersepta, tanpa clam
conection (Kaewchai et al. 2010). Menurut Liyanage (1997), R. microporus adalah
cendawan yang bersifat parasit fakultatif, artinya dapat hidup sebagai saprofit yang
kemudia menjadi parasit. R. microporus tidak dapat bertahan hidup apabila tidak ada
sumber makanan. Bila belum ada inang, cendawan ini bertahan di sisa-sisa tunggul.
Menurut Jayasuriya dan Thennakoon (2007), R. microporus (Sw.) diketahui sebagai
penyebab penyakit yang merugikan di perkebunan karet pada kebanyakan negara seperti
India, Indonesia, Malaysia, Srilanka, Thailand, Afrika Tengah, dan Afrika Barat.
Gejala serangan R. Microporus pada tanaman karet ditandai dengan adanya
perubahan warna daun. Daun berwarna hijau kusam, permukaan daun lebih tebal dari
yang normal. Setelah itu daun-daun menguning dan rontok. Pada pohon dewasa
gugurnya daun yang disertai dengan matinya ranting menyebabkan pohon mempunyai
mahkota yang jarang. Ada kalanya tanaman membentuk bunga/buah lebih awal. Pada
tanaman muda gejalanya mirip dengan tanaman yang mengalami kekeringan. Daun-daun
berwarna hijau kusam dan lebih tebal dari yang normal. Daun tersebut akhirnya menjadi
cokelat dan mengering. Pohon akhirnya tumbang dengan daun yang masih menggantung.
Ada kalanya pohon tiba-tiba tumbang tanpa menimbulkan gejala kematian tajuk, karena
akar tanaman telah busuk dan mati. Apabila leher akar tanaman yang terserang dibuka,
akan tampak rizomorf jamur berwarna putih, baik diakar tunggang ataupun di akar lateral.
Akar-akar tersebut akan busuk dan tanaman akan mati (Semangun, 2000).
Pada permukaan akar yang sakit terdapat benang-benang miselium cendawan
(Rizomorf) berwarna putih menjalar di sepanjang akar. Di sini benang-benang meluas
atau bercabang seperti jala. Pada ujungnya benang meluas seperti bulu, benang-benang
melekat erat pada permukaan akar. Kadang-kadang berwarna kekuningan, dalam tanah

9
merah tanahnya dapat kemerahan atau kecokelatan, kulit yang sakit akan busuk dan
warnanya cokelat. Kayu dari akar yang baru saja mati tetap keras, berwarna cokelat,
kadang-kadang agak keabu-abuan. Pada pembusukan yang lebih jauh, kayu berwarna
putih atau krem, tetapi padat dan kering, meskipun di tanah basah kayu yang terserang
dapat busuk dan hancur (Semangun, 2000). Serangan lebih lanjut penyakit akar putih
akan membentuk badan buah, berbentuk setengah lingkaran yang tumbuh pada pangkal
batang. Badan buah berwarna pink dengan tepi kuning mudah atau keputihan. Badan
buah berisi spora-spora cendawan yang akan berkembang dan keluar dari tubuh buah.
Spora tersebut akan berpencar dan menyerang tanaman karet yang masih sehat.

10
BAB 3 TUJUAN DAN MANFAAT PENELITIAN
3.1 Tujuan
Penelitian ini bertujuan untuk membuktikan penyebab penyakit mati meranggas
tanaman pala di Aceh Selatan.
3.2 Manfaat Penelitian
Mati meranggas merupakan penyakit yang telah menyerang ribuan hektar
tanaman pala di Kabupaten Aceh Selatan selama lima belas tahun terakhir. Penurunan
hasil produksi pala dapat mencapai 70% akibat serangan penyakit ini. Penyakit tersebut
menyerang dan menimbulkan kematian baik pada tanaman tua, muda, bahkan telah
dijumpai di pembibitan. Sampai saat ini penyebab kematian tanaman pala tersebut masih
belum pasti dan diperdebatkan. Dikalangan petani penyakit ini sudah sampai pada taraf
meresahkan karena belum dapat dikendalikan dan kasus serangannya cenderung
bertambah luas. Pengetahuan mengenai patogen penyebab sakit penting, terutama untuk
penyakit-penyakit baru. Penyusunan strategi pengendalian yang berkelanjutan biasanya
harus didasarkan pada sifat patogen bersangkutan. Pada kenyataannya sampai saat ini
memang belum ada metoda pengendalian yang dapat dilakukan untuk menghambat
perkembangan dan penyebaran patogen penyakit tersebut. Oleh karena itu perlu
dilakukan serangkaian penelitian untuk membuktikan penyebab penyakit mati meranggas
yg menyerang tanaman pala di Aceh Selatan, sehingga menjadi langkah awal dalam
menentukan tehnik pengendalian yang tepat dan bijaksana.
11
BAB 4 METODE PENELITIAN
4.1 Tempat dan Waktu
Kegiatan survey kejadian penyakit dan pengambilan sampel bagian-bagian
tanaman pala yang terserang beserta tanahnya akan dilakukan di 6 kecamatan yang berada
di kabupaten Aceh Selatan. Isolasi dan peremajaan isolate akan dilakukan di laboratorium
Mikologi Tumbuhan, Institut Pertanian Bogor. Pengujian dengan teknik Postulat Koch
akan dilakukan di Rumah Kaca Cikabayan, Bogor. Identifikasi secara molekuler akan
dilakukan di laboratorium Virologi Tumbuhan. Institut Pertanian Bogor. Kegiatan
penelitian ini akan dimulai dari Januari – Desember 2017.
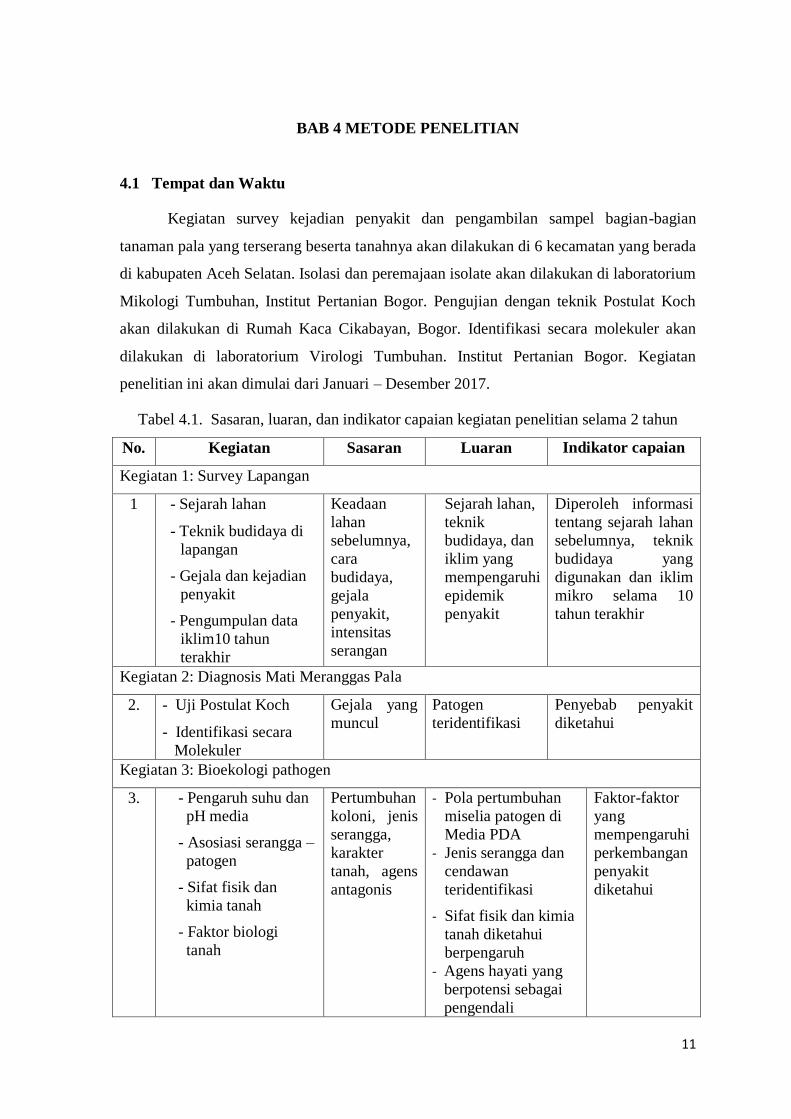
Tabel 4.1. Sasaran, luaran, dan indikator capaian kegiatan penelitian selama 2 tahun
No. Kegiatan Sasaran Luaran Indikator capaian
Kegiatan 1: Survey Lapangan
1 - Sejarah lahan
- Teknik budidaya di
lapangan
- Gejala dan kejadian
penyakit
- Pengumpulan data
iklim10 tahun
terakhir
Keadaan
lahan
sebelumnya,
cara
budidaya,
gejala
penyakit,
intensitas
serangan
Sejarah lahan,
teknik
budidaya, dan
iklim yang
mempengaruhi
epidemik
penyakit
Diperoleh informasi
tentang sejarah lahan
sebelumnya, teknik
budidaya yang
digunakan dan iklim
mikro selama 10
tahun terakhir
Kegiatan 2: Diagnosis Mati Meranggas Pala
2. - Uji Postulat Koch
- Identifikasi secara
Molekuler
Gejala yang
muncul
Patogen
teridentifikasi
Penyebab penyakit
diketahui
Kegiatan 3: Bioekologi pathogen
3. - Pengaruh suhu dan
pH media
- Asosiasi serangga –
patogen
- Sifat fisik dan
kimia tanah
- Faktor biologi
tanah
Pertumbuhan
koloni, jenis
serangga,
karakter
tanah, agens
antagonis
- Pola pertumbuhan
miselia patogen di
Media PDA
- Jenis serangga dan
cendawan
teridentifikasi
- Sifat fisik dan kimia
tanah diketahui
berpengaruh
- Agens hayati yang
berpotensi sebagai
pengendali
Faktor-faktor
yang
mempengaruhi
perkembangan
penyakit
diketahui

12
Untuk mencapai tujuan penelitian Hibah Disertasi Doktor yang diusulkan,
pelaksanaan riset akan dibagi menjadi tiga tahapan kegiatan penelitian yang satu sama
lain saling terkait sebagai satu kesatuan yang utuh. Kegiatan yang diusulkan dalam Hibah
Disertasi Doktor adalah kegiatan 2.
4.2 Bahan dan Metode
Identifikasi dengan Teknik Postulat Koch
Identifikasi terhadap penyebab penyakit tanaman pala dilakukan berdasarkan
karakter morfologi. Pengamatan dilakukan terhadap spesimen gejala penyakit yang
diambil dari lapangan dan hasil isolasi pada media PDA. Pengamatan makromorfologi
dan mikroskopik dari tubuh buah menggunakan mikroskop stereo dan kompon.
Pengamatan mikroskopik juga dilakukan terhadap cendawan hasil isolasi dari media
biakan. Sampel tubuh buah patogen diambil dari bagian pangkal batang. Sebagian dari
lapisan himenium dibuka untuk melihat bentuk pilar, subikulum serta rizomorf. Untuk
bagian himenium dan pilar dari tubuh buah diambil menggunakan pinset dan diletakkan
pada gelas objek untuk digunakan dalam pengamatan secara mikroskopik. Pengamatan
mikroskopik ini menggunakan bantuan mikroskop, dimaksudkan untuk mengamati spora,
hifa pathogen, dan tubuh buah (askokarb), serta jalinan miselia pada pilar. Sampel tubuh
buah yang dijadikan objek pengamatan diambil dari beberapa tempat kebun pala di
Kabupaten Aceh Selatan seperti kecamatan Labuhan Haji Timur, Meukek, Sawang,
Samadua, Tapak Tuan, dan Pasie Raja.
Pengamatan mikroskopik juga dilakukan terhadap miselia hasil isolasi patogen
pada media PDA (Potato Dextrose Agar). Isolasi patogen dilakukan dari jaringan ranting,
batang, akar yang bergejala, bagian tubuh buah dan tanah sekitar akar. Sebelumnya
bagian-bagian tanaman pala tersebut dibersihkan dan dipotong-potong sepanjang 3 cm,
kemudian dilakukan sterilisasi permukaan dengan merendamnya dalam klorok 1%,
dilanjutkan dengan larutan alkohol 70% selama 3 menit. Setelah itu potongan bagian-
bagian tanaman pala tersebut dibilas menggunakan aquades steril dan dikeringkan
menggunakan kertas isap. Bagian-bagian tersebut kemudian diletakkan pada media PDA
dalam cawan petri. Inkubasi dilakukan dalam inkubator pada suhu 28 oC selama 3 hari.
Setiap jenis cendawan yang tumbuh di permukaan atau bagian tepi potongan sampel

13
bagian-bagian tanaman pala tersebut di reisolasi. Hasil reisolasi ini akan digunakan dalam
identifikasi. Identifikasi cendawan menggunakan Watanabe (2002), Streets (1980), dan
Leslie and Summerel (2006).
Selanjutnya isolasi juga dilakukan dari bagian tubuh buah (himenium dan pilar).
Potongan tubuh buah tersebut sebelum diisolasi terlebih dahulu direndam dalam larutan
klorok 1%, selanjutnya dalam alkohol 70% selama 2 menit dan dibilas dengan
merendamnya dalam aquades steril sambil diguncang. Untuk menumbuhkan patogen
digunakan media PDA. Inkubasi dilakukan dalam inkubator pada suhu 28 oC selama 3
hari. Isolasi juga dilakukan dari tanah sekitar perakaran dengan teknik pengenceran
berseri dan ditumbuhkan pada media PDA, kemudian diinkubasi dalam inkubator pada
suhu 28 oC selama 3 hari.
Jenis cendawan yang tumbuh dari hasil isolasi, baik dari jaringan ranting, batang,
akar, ataupun tubuh buah digunakan untuk melakukan postulat Koch mengikuti
Brathwaite (1981), dengan langkah-langkah sebagai berikut: 1. mengidentifikasi patogen
yang berasosiasi dengan gejala dari beberapa sampel, 2. melakukan isolasi terhadap
patogen, baik dari tubuh buah ataupun dari jaringan tanaman. Isolasi menggunakan media
PDA, 3. penularan pada tanaman sehat menggunakan patogen hasil isolasi. Gejala yang
terbentuk hasilnya sama dengan gejala di lapangan, dan 4. dilakukan reisolasi dari gejala
yang terbentuk, dan hasilnya sama seperti hasil isolasi pertama.
Cendawan hasil isolasi dari jaringan kayu yang memperlihatkan kesamaan dengan
hasil isolasi dari tubuh buah digunakan sebagai sumber inokulum dalam postulat Koch.
Sumber inokulum yang digunakan pada postulat Koch berupa potongan miselia biakan
murni ukuran 1 cm2
yang diinokulasikan pada bagian akar bibit pala atau penempelan
miselia ke batang yang dilukai. Untuk pengujian menggunakan bibit pala yang berumur 1
tahun, masing-masing menggunakan 10 bibit untuk pengulangan. Pengamatan dilakukan
setiap hari sampai menunjukkan gejala penyakit.
Identifikasi Molekuler dengan ITS (Internal Transcribe Spacer)
Isolat cendawan yang didapat digunakan untuk identifikasi molekuler. Ekstraksi DNA
cendawan sebelumnya dibiakan pada media PDB (Potato Dextrose Broth) menggunakan
reagen nucleon PHYTOpure, dilanjutkan dengan amplifikasi PCR menggunakan primer ITS
1 dan ITS 4 (Harrington et al. 2000; Thorpe et al. 2005; Chungu et al. 2010). Untuk lebih
jelas dapat dilihat sebagai berikut:

14
a. Isolasi dan Amplifikasi DNA Cendawan
Ekstraksi DNA dilakukan berdasarkan pada metode Castillo et al. (1994) yang telah
dimodifikasi, yaitu dengan penambahan polyvinylpirolydone (PVP) pada bufer
ekstraksi CTAB dan natrium asetat pada tahap pengendapan DNA. Pelet yang di-
peroleh dicuci dengan 500 μL etanol 70%, dikeringkan, kemudian diresuspensi
dengan 100 μL bufer TE. Suspensi DNA disimpan pada –20 °C atau dapat langsung
digunakan untuk proses selanjutnya. Amplifikasi DNA cendawan menggunakan
pasangan primer ITS1 (5’ TCCGTAGG TGAACCTGCGG 3’) dan ITS4 (5’
TCCTCC GCTTATTGATATGC 3’). Reaksi PCR (volume 25 μL) terdiri atas 1 μL
DNA sampel dengan konsentrasi 25-50 ng μL-1; 18.8 μL air bebas nuklease; 2.5 μL
10x bufer PCR (10 mM KCl, 20 mM Tris HCl pH 8.8, 10 mM (NH4)2SO4, 2 mM
MgCl2, dan 0.1% Triton X-100) (Fermentas, USA); 0.5 μL dNTP 10 mM (Fermentas,
USA); masing-masing 1 μL primer ITS1 dan ITS4 dengan konsentrasi 10 μM; 0.2 μL
enzim Taq DNA polimerase rekombinan 5U μL-1 (Dream Taq DNA Polymerase
Fermentas, USA). Amplifikasi didahului dengan denaturasi awal selama 5 menit pada
94 °C, dilanjutkan sebanyak 35 siklus melalui tiga tahapan meliputi denaturasi selama
1 menit pada 94 °C, penempelan primer (annealing) selama 1 menit pada 55 °C,
sintesis selama 2 menit pada 72 °C, pada tahap akhir ditambah 10 menit pada 72 °C.
Analisis DNA hasil amplifikasi dilakukan dengan elektroforesis menggunakan gel
agarosa 1% dalam bufer Tris Boric EDTA (TBE 0.5x) dan visualisasi menggunakan
sinar UV (Sambrook dan Russel 2001; Barnes et al. 2003; Van Wyk et al. 2007; Van
Wyk et al. 2012).
b. Analisis Perunutan DNA
DNA hasil amplifikasi digunakan untuk tahapan perunutan DNA berdasarkan pada
metode dideoxy nucleotide chain termination (Macrogen Inc., Korea Selatan). Hasil
pe-runutan DNA selanjutnya disusun dengan program Bioedit dan dianalisis
menggunakan program BLASTN dengan memanfaatkan informasi dari Genbank
(http://www.ncbi.nlm.nih.gov).

15
BAB 5 HASIL DAN PEMBAHASAN
Pada tanaman pala terinfeksi di lapangan memerlihatkan gejala mati ranting, yaitu
layu pada sebagian ranting, mengering, kemudian menjalar dari satu cabang ke cabang
lainnya sampai keseluruhan bagian tanaman, selanjutnya daun menjadi kecoklatan,
mengeriting, dan menggantung yang akhirnya gugur sampai mati meranggas. Disisi lain
terdapat tanaman layu secara keseluruhan, menguning, mengering, gugur daun, dan mati.
Tabel 5.1 Asosiasi antar tanda dan gejala penyakit pada pohon pala di Aceh Selatan
Gejala
penyakit L MR DM D K PB R MM
L 1.000
MR -0.173**
1.000
DM -0.222**
0.717**
1.000
D 0.309**
0.394**
0.550**
1.000
K -0.320**
0.350** 0.581
** 0.719
** 1.000
PB -0.183**
0.401**
0.608**
0.797**
0.893**
1.000
R -0.094 -0.089 -0.142**
0.214**
0.247**
0.242**
1.000
MM -0.149**
-0.199**
-0.289**
0.376**
0.491**
0.469**
0.549**
1.000
** = berasosiasi sangat nyata, Layu (L), Mati ranting (MR), Daun menggantung (DM), Diskolorasi (D), Kanker (K),
Penggerek batang (PB), Rizomorf (R), Mati meranggas (M); n = 360 pohon
Berdasarkan pengamatan lapangan pada tanaman pala, diketahui bahwa setiap
gejala yang terjadi pada tanaman terinfeksi berasosiasi sangat nyata dengan mati
meranggas, baik secara positif maupun negatif. Gejala mati meranggas berasosiasi sangat
nyata dengan diskolorasi, kanker, penggerek, dan rizomorf. Sedangkan gejala-gejala yang
berasosiasi sangat nyata negatif berarti bila satu gejala muncul maka gejala lainnya tidak
ada, sebagai contoh apabila satu tanaman tersebut memerlihatkan gejala layu maka tidak
memerlihatkan gejala mati ranting, daun menggantung, kanker, penggerek, maupun mati
meranggas (Tabel 5.1).
5.1 Uji Postulat Koch
Berdasarkan uji Postulat Koch, diketahui ada empat jenis cendawan yang
memerlihatkan gejala sama, mulai dari terbentuknya kanker pada bagian bibit tanaman
yang diinokulasi, mati pucuk/ranting, daun menggantung sampai akhirnya mati
meranggas, dan saat batang dipotong secara melintang maupun disayat terlihat nekrosis

16
pada bagian pembuluh batang (Gambar 5.1). Hasil uji membuktikan bahwa cendawan A,
B, C, dan D merupakan cendawan penyebab penyakit (Tabel 5.1).
Gambar 5.1 Hasil inokulasi buatan pada bibit tanaman pala; inokulasi Botryodiplodia sp.(A); inokulasi
Graphium sp. (B); inokulasi Fusarium solani (C); inokulasi Rigidoporus sp. (D); Tanpa
patogen (K).
Gejala awal hasil inokulasi buatan adalah kanker dengan rata-rata masa inkubasi
tiga puluh dua hari. Gejala yang terbentuk ini awalnya tidak berkembang namun setelah
bibit mendapat perlakuan kekeringan, sebulan kemudian mulai muncul gejala mati pucuk
maupun cabang lalu menjalar ke seluruh bagian tanaman, berlanjut dengan daun
mengering, dan menggantung selama empat sampai delapan minggu, kemudian gugur,

17
dan tanaman mati meranggas. Sementara bibit tanaman pala yang diinokulasi dengan
isolat cendawan D tidak memerlihatkan gejala kanker tetapi gejala busuk akar karena
cendawan tersebut merupakan cendawan yang diisolasi dari bagian akar tanaman pala
yang memerlihatkan hifa (rizomorf) yang melilit bagian akar tanaman, namun gejala yang
terbentuk pada bagian atas tanaman memerlihatkan gejala yang sama dengan yang
disebabkan oleh cendawan uji lainnya.
Frekuensi cendawan saat diisolasi dari jaringan tanaman terserang asal tiga puluh
enam kebun sampel ternyata sangat bervariasi. Cendawan A yaitu Botryodiplodia sp.
adalah jenis cendawan yang paling dominan ditemukan , bahkan dapat ditemukan secara
murni dari hasil isolasi dengan teknik trapping (perangkap) spora langsung menggunakan
media MA dan PDA di sekitar tanaman terserang. Empat cendawan (A, B, C, dan D)
yang berasosiasi dengan tanaman mati meranggas positif pada semua tahapan postulat
Koch. Hal ini membuktikan keempat cendawan tersebut bisa menyebabkan gejala mati
meranggas. Namun cendawan A merupakan cendawan yang memiliki frekuensi isolasi
yang tertinggi yaitu 36/36 (100%). Sedangkan cendawan B, C, dan D frekuensinya lebih
rendah yaitu 4/36 (11%), 5/36 (13,9%), dan 2/36 (5%). Oleh karena itu dapat
disimpulkan bahwa penyebab mati meranggas tanaman pala adalah cendawan A
(Botryodiplodia sp.). Setelah reisolasi dari tanaman uji, empat cendawan menunjukkan
hasil yang sama dengan cendawan patogen hasil isolasi awal (Tabel 5.2).
Tabel 5.2 Gejala yang ditimbulkan cendawan hasil isolasi
No. Cendawan Hasil postulat Koch Hasil identifikasi
Frekuensi isolasi
Batang
(36)*
Ranting
(36)*
Akar
(36)*
1 A + Botryodilodia sp. 36 36 36
2 B + Graphium sp. 4 4 -
3 C + Fusarium solani 2 2 5
4 D + Rigidoporus sp. - - 2
5 E - Endomelanconiopsis sp. 1 - -
6 F - Clonotachys sp. 2 - - Keterangan : + = ada (positif pada semua tahapan postulat Koch) ; - = tidak ada ; * = angka dalam kurung
menunjukkan jumlah total sampel
5.2 Identifikasi Morfologi dan Molekuler Patogen
Cendawan yang berhasil diisolasi ada enam jenis, namun hanya empat isolat yang
patogenik. Keempat isolat tersebut memiliki karakteristik yang sesuai dengan genus pada
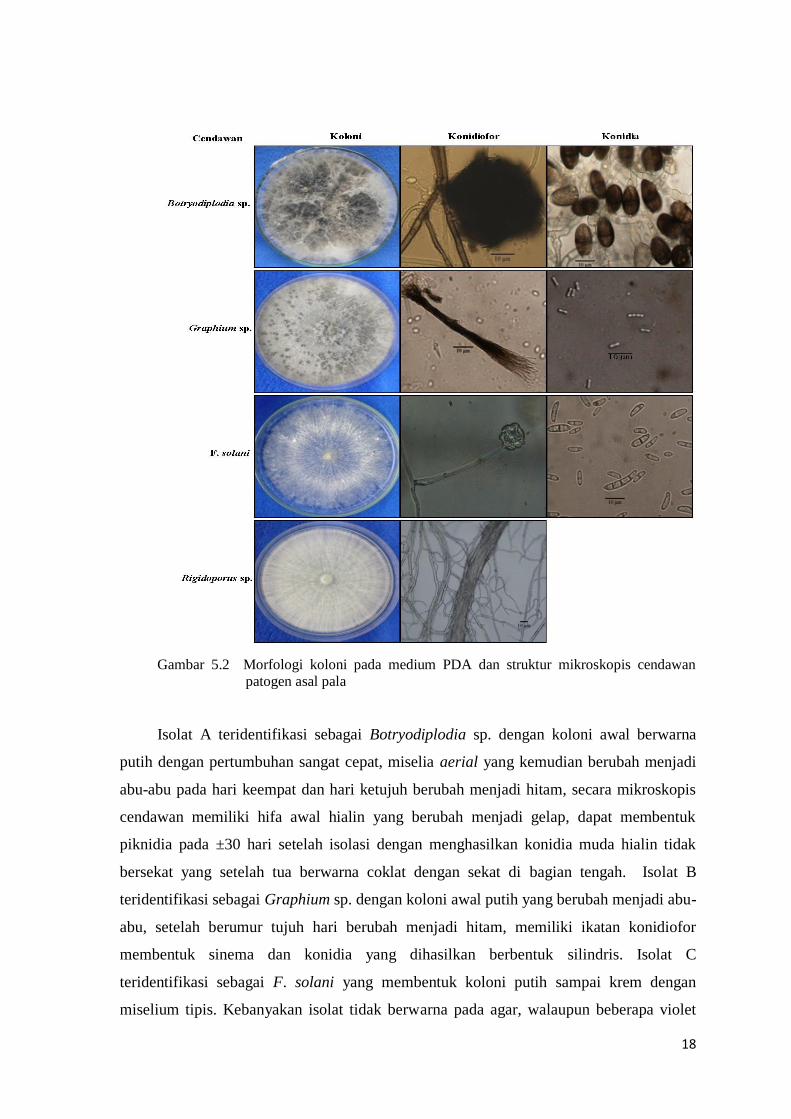
kunci identifikasi yang digunakan (Gambar 5.2).
18
Gambar 5.2 Morfologi koloni pada medium PDA dan struktur mikroskopis cendawan
patogen asal pala
Isolat A teridentifikasi sebagai Botryodiplodia sp. dengan koloni awal berwarna
putih dengan pertumbuhan sangat cepat, miselia aerial yang kemudian berubah menjadi
abu-abu pada hari keempat dan hari ketujuh berubah menjadi hitam, secara mikroskopis
cendawan memiliki hifa awal hialin yang berubah menjadi gelap, dapat membentuk
piknidia pada ±30 hari setelah isolasi dengan menghasilkan konidia muda hialin tidak
bersekat yang setelah tua berwarna coklat dengan sekat di bagian tengah. Isolat B
teridentifikasi sebagai Graphium sp. dengan koloni awal putih yang berubah menjadi abu-
abu, setelah berumur tujuh hari berubah menjadi hitam, memiliki ikatan konidiofor
membentuk sinema dan konidia yang dihasilkan berbentuk silindris. Isolat C
teridentifikasi sebagai F. solani yang membentuk koloni putih sampai krem dengan
miselium tipis. Kebanyakan isolat tidak berwarna pada agar, walaupun beberapa violet

19
atau coklat muda. Cendawan ini memiliki konidiofor panjang yang menghasilkan dua tipe
konidia yaitu makrokonidia yang relatif lebar (luas), lurus, ramping, melenkung, yang
terdiri dari 3 sampai 6 septa dengan ujung yang membulat, sedangkan mikrokonidia
berbentuk oval yang terdiri dari 0 sampai 1 septa yang diproduksi dari monofialid yang
panjang. Klamidospora diproduksi pada hifa, hialin, berbentuk bundar, oval dengan
dinding yang halus dan kasar. Pertumbuhan klamidospora cepat dan banyak (berlimpah)
yang berada pada bagian interkalar hifa atau ujung cabang lateral yang pendek, yang
terdiri dari satu atau berpasangan. Isolat D teridentifikasi sebagai Rigidoporus sp. karena
membentuk koloni putih tipis, dengan hifa hialin dan bercabang-cabang namun tidak
memiliki clam conection.
Selanjutnya proses identifikasi secara molekuler dilakukan untuk mengonfirmasi
spesies cendawan yang teridentifikasi secara morfologi. Visualisasi hasil PCR
menunjukkan amplifikasi pita yang dilakukan menggunakan primer ITS1 dan ITS4
tampak sesuai harapan. Amplifikasi pita DNA yang terbentuk pada agarose mempunyai
ukuran sekitar ~ 500 - 650 bp (Gambar 5.3). Ukuran tersebut merupakan kelompok
cendawan.
Gambar 5.3 Visualisasi elektroforesis hasil PCR 4 isolat cendawan dengan menggunakan primer
ITS1 dan ITS4. M= Marker; 1. Botryodiplodia sp.; 2. Graphium sp.; 3. F. solani;
dan 4. Rigidoporus sp.
Menurut Toju et al. (2012), primer ITS cocok digunakan dalam mengidentifikasi
sampel lapangan dari cendawan kelompok Ascomycetes dan Basidiomycetes berbasiskan
pada untaian DNA. Namun, cendawan dari kelompok Deuteromycetes juga dapat
terdeteksi dengan primer ITS.
M 1 2 3 4
500 bp 650 bp
20
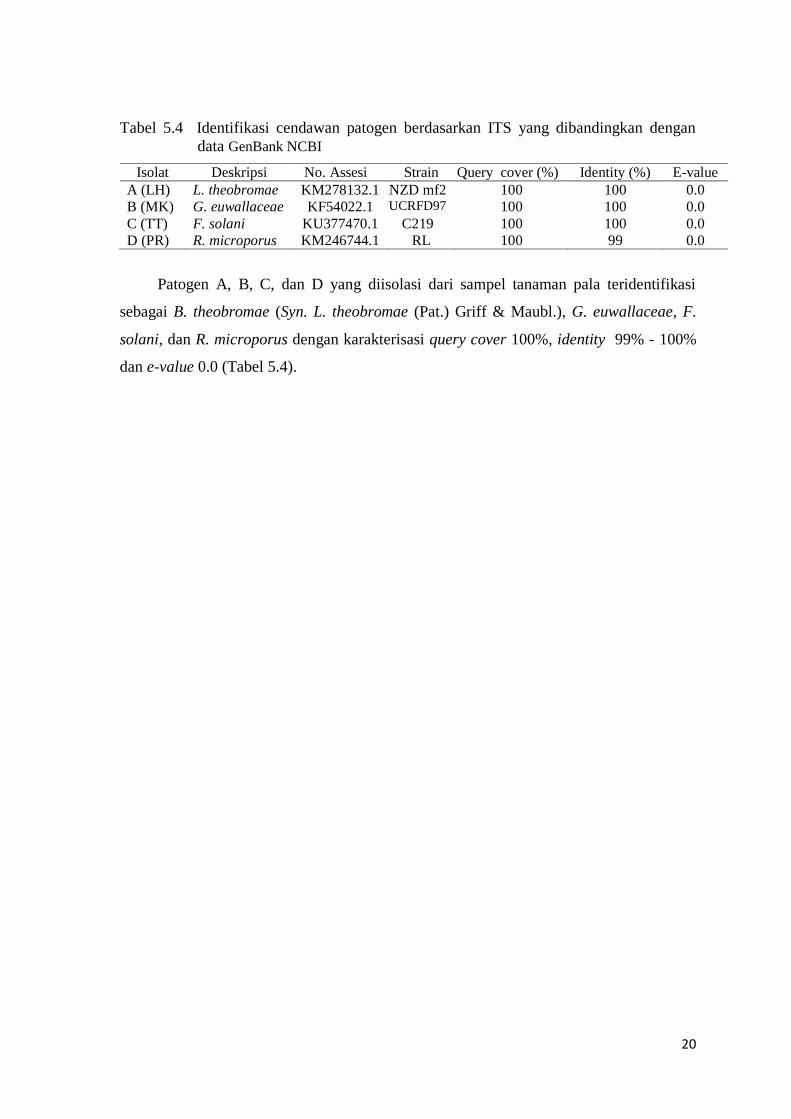
Tabel 5.4 Identifikasi cendawan patogen berdasarkan ITS yang dibandingkan dengan
data GenBank NCBI
Isolat Deskripsi No. Assesi Strain Query cover (%) Identity (%) E-value
A (LH) L. theobromae KM278132.1 NZD mf2 100 100 0.0
B (MK) G. euwallaceae KF54022.1 UCRFD97 100 100 0.0
C (TT) F. solani KU377470.1 C219 100 100 0.0
D (PR) R. microporus KM246744.1 RL 100 99 0.0
Patogen A, B, C, dan D yang diisolasi dari sampel tanaman pala teridentifikasi
sebagai B. theobromae (Syn. L. theobromae (Pat.) Griff & Maubl.), G. euwallaceae, F.
solani, dan R. microporus dengan karakterisasi query cover 100%, identity 99% - 100%
dan e-value 0.0 (Tabel 5.4).
M 1 2 3 4

21
BAB 6 KESIMPULAN DAN SARAN
6.1 Kesimpulan
Empat Penyebab penyakit mati meranggas pohon pala di Aceh Selatan yang
teridentifikasi, yaitu Lasiodiplodia theobromae, Graphium euwallacea, Fusarium solani,
dan Rigidoporus microporus. Penyebab penyakit yang dominan adalah Botryodiplodia
theobromae Pat. (sinonim Lasiodiplodia theobromae (Patouillard) Griffon & Maubland).
6.2 Saran
Perlu penelitian lanjutan untuk melihat kisaran inang dari B. theobromae, dan
interaksi antara patogen dengan serangga hama

22
DAFTAR PUSTAKA
Adandonon A, Datinon B, Baimey J, Toffa J. 2014. First report of Lasiodiplodia
theobromae (Pat.) Griffon & Maubl. Causing root rot and collar rot disease of
Jatropha curcas L. in Benin. J Appl Biosci. 79(1):6873-6877.
Agrios GN. 2005. Plant Pathology. 5th
. San Diego (US). New York. Academic Pr.
Al-Saadoon AH, Amen MKM, Al-Rubaie EMA. Histopathology of grapevine inoculated
with Lasiodiplodia theobromae. Basrah J of Agric Sci. 25(1):1-12.
Barnes I, Roux J, Wingfield BD, Dudzinski MJ, Old KM, Wingfield MJ. 2003.
Ceratocystis pirilliformis, a new species from Eucalyptus nitens in Australia.
Mycologia 95(5) : 865 – 871.
Brathwaite CWD. 1981. An Introduction to the Diagnosis of Plant Disease. Inter-Am Inst
Corp Agric San Jose (CR). Edit IICA
Castillo CO, Chalmers KJ, Waugh R, Powell W. 1994. Detection of genetic diversity and
selective gene in coffea using RAPD markers. Theor Appl Genet. 87(8):934-940.
doi: 10.1007/BF00225787.
Chungu D, Muimba-Kankolongo A, Wingfield MJ, Roux J. 2010. Identification of
fungal pathogens occurring in eucalypt and pine plantations in Zambia by
comparing DNA sequences. J. Forestry 83 (5) : 507 – 515.
Dishutbun Aceh Selatan. 2014. Dinas Perkebunan dan Kehutanan; Pedoman
Pengamatan dan Pengendalian Hama pada Tanaman Pala. Tapaktuan, Aceh
Selatan.
Engelbrecht CJ, Harrington TC, Alfenas A. 2007. Ceratocystis Wilt of Cacao – A
Disease of Increasing Importance. Phytopathology 97 : 1648 – 1649.
Faridh AM, Lee SS, Maziah Z, Rosli H, Norwati M. 2007. Root rot in tree spesies other
than Acacia. Di dalam: Potter K, Rimbawanto A, Beadle C, editor. Heart rot and
root rot in tropical Acacia plantations. Proceeding of a workshop help in
yokyakarta. 2007 February 7-9; Camberra, ACIAR. Proceeding 124 : 60 – 66.
Fateh FS, Kazmi MR, Ahmad I, Ashraf M. 2006. Ceratocystis fimbriata isolated from
vascular bundles of declining mango trees in Sindh, Pakistan. Pak. J. Bot. 38 (4) :
1257 – 1259.
Firmino AC, Novaes QS, Tozze HJ, Sobrinho GGR, Santos A, Bezerra JL, Furtado. 2013. First report of Ceratocystis fimbriata causing fruit-rot of Passiflora edulis in
Brazil. New Disease Reports 27 : 4.
Firmino AC, Tozze HJ, Furtado EL. 2012. First report of Ceratocystis fimbriata causing
wilt in Tectona grandis in Brazil. New Disease Reports 25 : 24.

23
Glen M. 2006. The use of DNA techniques to indentify fungi. Di dalam: Potter K,
Rimbawanto A, Beadle C, editor. Heart rot and root rot in tropical Acacia
plantations. Proceeding of a workshop help in yokyakarta. 2007 February 7-9;
Camberra, ACIAR. Proceeding 124 : 46 - 54.
Hadad EA, Firman C. 2003. Budidaya Pala. Balittro. Bogor (ID).
Harni R, Trisawa IM. 2011. Observasi dan identifikasi penyebab matinya pala di daerah
Aceh Selatan. Laporan Kerjasama Dinas Kehutanan dan Perkebunan dengan Balai
Penelitian Tanaman Rempah dan Aneka Tanaman Industri. Sukabumi. 36 p
Harrington TC, Steimel J, Workneh F, Yang XB. 2000. Molecular identification of fungi
associated with vascular discoloration of soybean in the north central United
States. Plant Dis. 84 : 83 – 89.
Ismail AM. Cirvilleri G, Polizzi G, Crous PW, Groenewald JZ, Lombard L. 2012.
Lasiodiplodia species associated with dieback disease of mango (Mangifera
indica) in Egypt. Austral P Pathol. 41(2012):649-660. doi: 10.1007/s13313-012-
0163-1.
Kaewchai S, Kai Wang H, Lin FC, Hyde KD, Soytong K. 2009. Genetic Variation
among Isolates of Rigidoporus microporus causing White Root Disease of Rubber
Trees in Southern Thailand Revealed by ISSR Markers and Pathogenicity. African
Journal of Microbiology Research 3 (10) : 641 – 648.
Kaewchai S, Lin FC, Wang HK, Soytong K. 2010. Characterization of Rigidoporus
microporus isolated from rubber trees based ob morphology and ITS sequencing.
Journal of Agricultural Technology 6 (2) : 289 – 298.
Latha P, Prakasam V, Kamalakannan A, Gopalakrishnan C, Raguchander T, Paramathma
M, Samiyappan R. 2009. First report of Lasiodiplodia thebromae (Pat.) Griffon &
Maubl causing root rot and colar rot disease of physic nut (Jatropha curcas L.) in
India. Austral Plant Dis Notes. 2009(4):19-20.
Leslie JS, Summerel BA. 2006. The Fusarium Laboratory Mannual. 1st ed. Blackwell
Publishing. Ltd. (US): Iowa.
Liyanage A De S. 1997. Rubber. In Hillocks RJ, Walter JM, editor. Soil borne diseases
of tropical crops. Wallingford, UK, CAB International, 331–347.
Mamble AM, Jolanda R. 2015. Diseases of plantation forestry trees in Southern Ghana.
Int. J. Phytopathol. 04 (01) : 05-13
Rahman MU, Sankaran KV, Leelavathy KM, Zachariah S. 1981. Cylindrocladium Root
Rot of Nutmeg in South India. Plant Disease 65 : 514 – 515.
Roux J, Van Wyk M, Hatting H, Wingfield MJ. 2004. Ceratocystis species infecting stem
wounds on Eucalyptus grandis in South Africa. Plant Pathology 53 : 414 – 421.

24
Roux J, Wingfield MJ, Bouillet JP, Wingfield BD, Alfenas AC. 2000. A serious new
wilt disease of Eucalyptus caused by Ceratocystis fimbriata in Central Africa.
Path. 30: 175 – 184.
Roux J, Wingfield MJ. 2009. Ceratocystis species: emerging pathogens of non-native
plantation Eucalyptus and Acacia species. Southern Forests 71 (2) : 115 – 120.
Sambrook J, Russel DW. 2001. Molecular Cloning a Laboratory Manual. Ed ke-3. New
York (US): Cold Spring Harbor Laboratory Pr.
Sinaga MS. 2003. Dasar-Dasar Ilmu Penyakit Tumbuhan. Jakarta. Penebar Swadaya.
Shahbaz M, Iqbal Z, Saleem A, Anjum MA. 2009. Association of Lasiodiplodia
theobromae with different decline disorders in mango (Mangifera indica L.). Pak
J Bot. 41(1):259-368.
Streets RB. 1972. The Diagnosis of Plant Diseases: A Field and Laboratory Manual
Emphasizing The Most Practical Methode for Rapid Identification. The University
of Arizona Press. (US): Tuscon – Arizona.
Thorpe DJ, Harrington TC, Uchida JY. 2005. Pathogenicity, internal transcribed spacer-
rDNA variation, and human dispersal of Ceratocystis fimbriata on the family
Araceae. Phytopathology 95 : 316 – 323.
Toju H, Tanabe AS, Yamamoto S, Sato H. 2012. High-covarage ITS primers for the
DNA-based identification of Ascomycetes and Basidiomycetes in environmental
samples. Plos ONE 7(7) : 1-11.
Van Wyk M, Al Adawi AO, Khan IA, Deadman ML, AlJahwari AA, Wingfield BD,
Ploetz R, Wingfield MJ. 2007. Ceratocystis manginecans sp. nov., causal agent
of a destructive mango wilt disease in Oman and Pakistan. Fungal Diversity 27 :
213 – 230.
Van Wyk M, Al Adawi AO, Wingfield BD, Al-Subhi AM, Deadman ML, Wingfield MJ.
2005. DNA based characterization of Ceratocystis fimbriata isolates associated
with mango decline in Oman. Australastan Plant Pathology 34 : 587 – 590.
Van Wyk M, Roux J, Barnes I, Wingfield BD, Liew ECY, Assa B, Summerell BA,
Wingfield MJ. 2004. Ceratocystis polychrome sp. nov., a new species from
Syzygium aromaticum in Sulawesi. Studies in Mycology 50 : 273 – 282.
Van Wyk M, Wingfield BD, Mohali S, Wingfield MJ. 2009. Ceratocystis
fimbriatomima, a New Species in The C. fimbriata sensu lato Complex Isolated
from Eucalyptus Trees in Venezuela. Fungal Diversity 34 : 175 – 185.
Van Wyk M, Roux J, Nkuekam GK, Wingfield BD, Wingfield MJ. 2012. Ceratocystis
eucalypticola sp. nov. from Eucalyptus in South Africa and comparison to global
isolate from this tree. IMA fungus 3 (1) : 45 – 58.

25
Van Wyk M, Wingfield BD, Clegg PA, Wingfield MJ. 2009. Ceratocystis larium sp.
nov., a new species from Styrax benzoin wounds associated with incense
harvesting in Indonesia. Persoonia 22 : 75 – 82.
Van Wyk M, Wingfild BD, Marin M, Wingfild MJ. 2010. New Ceratocystis species
infecting coffee, cocoa, citrus, and native trees in Colombia. Fungal Diversity 40:
103-117.
Watanabe T. 1994. Pictorial Atlas of Soil and Seed Fungi: Morphologies of Cultured
Fungi and Key to Species. Lewis Publishers. (JP): Tokyo
Yuwono T. 2006. Teori dan Aplikasi Polymerase Chain Reaction. Yogyakarta: Penerbit
Andi.

26
LAMPIRAN
Lampiran 1. Instrumen Penelitian
Form Questioner
Kecamatan : Tanggal wawancara :
Desa : Tempat wawancara :
Pewawancara : Waktu wawancara :
Karakteristik petani
1. Nama :
2. Umur :
3. Pendidikan :
Tidak sekolah
SD
SMP
SMU
Perguruan Tinggi 4. Pekerjaan utama :
5. Pekerjaan sampingan :
6. Pengalaman usaha pala
< 5 tahun
> 5 – 10 tahun
> 10 – 15 tahun
> 15 – 20 tahun
> 20 tahun
Keadaan lahan
7. Topografi lahan
Datar
Berlereng
Berbukit
Lainnya …………………
8. Sejarah lahan
Tanaman yang ditanam sebelum pala adalah ………………….
Lama penanaman tanaman tersebut …………………..
Budidaya Pala
9. Asal bibit
bibit sendiri
Beli dari perusahaan pembibitan
Instansi pemerintah
Beli dari petani lain
Lainnya …………………… 10. Bila melakukan pembibitan sendiri, cara penyiapan bibit:

27
Secara generative dengan biji
Lainnya …………………… 11. Umur bibit yang ditanam :
12. Umur tanaman saat ini :
< 2 tahun
2 – 5 tahun
5 – 10 tahun
> 10 tahun 13. Luas lahan :
14. Jumlah pohon pala yang ada :
15. Pola tanam :
monokultur
Tumpang sari
Lainnya ………………………..
16. Pemeliharaan:
a. Pemupukan
Jenis pupuk Frekwensi/tahun Waktu pemupukan Dosis
Urea
TSP
KCL
NPK
Ca
b. Jenis gulma yang tumbuh :
c. Pengendalian gulma
Cara pengendalian Frekwensi/tahun Waktu Jenis alat/kimia
Mekanik
Herbisida
17. Gulma yang telah dibersihkan diletakkan di :
sekitar pohon
dibakar
dibuat pupuk kompos
Lainnya …………………………. 18. Apakan tanaman pala bapak/ibu terserang penyakit mati meranggas?
Ya Tidak
19. Sejak kapan tanaman bapak/ibu terserang penyakit tersebut ……………….
20. Berapa banyak pohon yang terserang ………………
21. Menurut bapak/ibu berapa persen kerusakan akibat penyakit tersebut?
< 10% > 25 – 50%
10 – 25% > 50%
22. Apakah serangan penyakit mati meranggas tersebut mengganggu pertumbuhan
23. Apakah serangan penyakit mati meranggas pada tanaman pala bapak/ibu telah
menyebabkan kematian ?

28
24. Berapa persen kematian tanaman pala bapak/ibu sampai saat ini?
< 10% > 25 – 50%
10 – 25% > 50%
25. Berapa persen kerugian ekonomi akibat serangan penyakit tersebut?
< 10% > 25 – 50%
10 – 25% > 50%
26. Apakah bapak/ibu melakukan pengendalian terhadap penyakit tersebut?
Ya Tidak
27. Pengendalian yang telah dilakukan
a. …………………
b. …………………………
c. ………………………….
d. …………………………
e. ………………………….
28. Jenis pestisida yang digunakan
Jenis/nama pestisida Frekwensi/tahun Waktu Dosis/takaran
29. Bagaimana hasilnya setelah pengendalian dilakukan?
Tidak ada pengaruh
Dapat mengurangi serangan penyakit
Lainnya …………………
30. Apa harapan bapak/ibu ke depan terkait dengan masalah yang dihadapi saat ini?
a. ……………………………
b. ……………………………
c. ……………………………
d. …………………………...

29
1 Survey lapangan di salah satu kebun petani di Kabupaten Aceh selatan
2 Isolat-isolat murni hasil isolasi dari bagian-bagian tanaman pala terserang

30
3 Teknik isolasi cendawan dari bagian tanaman pala terserang penyakit
Postulat Koch: Inokulasi Cendawan
4 Perlakuan postulat Koch pada bibit tanaman pala 1 tahun

31
5 Hasil pengujian postulat Koch
Isolasi DNA : Column based methods (teknik molekuler)
hifa homogenisasi + lisis bufer Inkubasi 60oC Ambil supernatan
+ binding buffer binding DNA
+ washing buffer I
+ washing buffer II
pengeringan resuspensi DNA
- Tahap Pretreatment
- Tahap Isolasi
6 Teknik molekuler untuk identifikasi cendawan

32
Lampran 2. Personalia Tenaga Peneliti
I. IDENTITAS DIRI
1.1 Nama Lengkap (dengan gelar) Ir. Susanna, M.Si.
1.2 Jabatan Fungsional Lektor Kepala
1.3 NIP/NIDN 196811301994032001 / 0030116801
1.4 Tempat dan Tanggal Lahir Langsa, 30 November 1968
1.5 Alamat Rumah Jl. Mesjid Inayatullah no.5, Kp. Cimanggis RT
03/01, Mekarwangi, Tanah Sereal, Bogor
1.6 Nomor Telepon/Faks 02517539210
1.7 Nomor HP 085261251136
1.8 Alamat Kantor Fakultas Pertanian Unsyiah
1.9 Nomor Telepon/Faks 0651-51977 Ps. 4245
1.10 Alamat e-mail [email protected]
1.11 Lulusan yang Telah Dihasilkan S-1= 31 orang S-2= - orang
S-3= - orang
1.12 Mata Kuliah yang Diampu 1. Botani
2. Mikrobiologi Pertanian
3. Organisme Pengganggu Tanaman
4. Pengelolaan Organisme Penganggu
Tanaman
5. Pengendalian Hayati dan Pengelolaan
Habitat
II. RIWAYAT PENDIDIKAN
S-1 S-2 S-3
Nama PT Universitas Syiah Kuala IPB- Bogor
Bidang Ilmu Penyakit Tanaman Fitopatologi
Tahun Masuk-Lulus 1987 - 1992 1996 - 2000
Judul Tugas Akhir Pengujian Ekstrak Daun
Sirih terhadap Jamur
Colletotrichum capsici In
Vitro
Analisis Introduksi
Mikroorganisme
Antagonis untuk
Pengendalian Hayati
Penyakit layu
(Fusarium oxysporum
f.sp. cubense) pada
Pisang
Nama Pembimbing/
Promotor
1. Ir. Azwir Mirin
2. Ir. Helmi A.Gani
1. Dr. Ir. Meity
Sinaga, M.Sc.
2. Dr. Ir. Sientje
Mandang Sumarau,
M.Sc.
3. Prof. Dr. Ir. Ahmad
Surkati, M.Sc.
33
III. PENGALAMAN PENELITIAN
No. Tahun Judul Penelitian Pendanaan
Sumber Jumlah (Juta Rp)
1.
1992
Pengujian Ekstrak Daun Sirih
terhadap Jamur Colletotrichum
capsici In Vitro.
Mandiri
2.500.000
2.
2000
Analisis Introduksi
Mikroorganisme Antagonis untuk
Pengendalian Hayati Penyakit
layu (Fusarium oxysporum f.sp.
cubense) pada Pisang
Mandiri
7.500.000
3.
2006
Analisis Introduksi Trichoderma
harzianum dan Pupuk Bokhasi
untuk Pengendalian Layu
Sklerotium pada Tanaman
Kedelai
PDM
10.000.000,-
4. 2008 Aplikasi Urine Sapi sebagai
Biofungisida Pyricularia oryzae
pada Tanaman Padi
PDM
10.000.000,-
5.
2008
Pengendalian Biologi Layu
Sklerotium pada Tanaman
Kedelai dengan Pseudomonas
fluorescens dan Trichoderma
harzianum pada Waktu Berbeda
PDM
10.000.000,-
6.
2009
Aplikasi Pupuk Hayati dan Pupuk
Kandang untuk Pengendalian
Hama Lalat Bibit (Ophiomyia
phaseoli) pada Tanaman Kedelai.
PDM
15.000.000,-
7.
2009
Keragaman dan Kesesuaian Inang
Jamur Endofitik yang Berasosiasi
pada Area Daun Tanaman Kakao
di Aceh
Hibah
Kompetitif
150.000.000,-
8. 2011 Peran Cendawan Trichoderma
virens dalam Merangsang
Perkecambahan benih dan
Hubungannya dengan
Peningkatan Auksin
PDM
15.000.000,-
9. 2011 Keanekaragaman Fungi Tanah
Per-kebunan Kopi Arabika dan
Potensinya sebagai Pengendali
Hayati Penyakit Busuk Akar Kopi
di Kabupaten Bener Meriah
Hibah
Bersaing
44.000.000,-
IV. PENGALAMAN PENGABDIAN KEPADA MASYARAKAT
No. Tahun Judul Pengabdian Kepada
Masyarakat
Pendanaan
Sumber Jumlah (Rp)
1. 1996 Pembinaan sistem manajemen pada DPP 2.500.000,-
34
pesantren terpadu di Desa Ulee Lhee
2. 1996 Pembinaan petani dalam
pengembangan tanaman kacang tanah
(Arachis hypogea L.) pada lahan
sawah tadah hujan di Desa Tungkop
Kecamatan Darussalam Kabupaten
Aceh besar
DPP 2.500.000,-
3. 2005 Tehnik aplikasi insektisida Fipronil
pada padi sawah di Desa Lambitra
Kecamatan Darussalam
DPP 3.000.000,-
4. 2007 Pengenalan dan Pembuatan
Pestisida Organik untuk
Pengendalian Penyakit-Penyakit
pada tanaman Padi
IPTEKS 7.500.000,-
5. 2008 Pemanfaatan Pestisida Nabati untuk
Pengendalian Penyakit pada
Tanaman Sayuran
IPTEKS
7.500.000,-
6. 2009 Pemanfaatan Ekstrak Nimba
sebagai Pengendali Hama ulat
Grayak pada Tanaman Sawi
IPTEKS
7.500.000,-
7.
2010 Potensi Pemanfaatan Trichoderma
harzianum dan Kascing untuk
Pengendalian Penyakit Layu pada
Budidaya Tanaman Tomat di Desa
Tanjong Selamat Kecamatan
Darussalam Aceh Besar
IbM
48.000.000,-
8. 2010 Penerapan Metode Pengendalian
Hama Terpadu pada Budidaya
Tanaman Cabai di Desa Lambaro
Seubun Kecamatan Lhok Nga
Kabupaten Aceh Besar
IbM
50.000.000,-
V. PENGALAMAN PENULISAN ARTIKEL ILMIAH
No. Tahun Judul Artikel Ilmiah Volume/
Nomor
Nama Jurnal
1.
2005
Analisis inroduksi Cendawan Antagonis
untuk Pengendalian Hayati Penyakit layu
(Fusarium oxysporum f.sp. cubense) pada
Pisang
11/2
Agrista
2.
2006
Pemanfaatan Bakteri Antagonis sebagai
Agen Biokontrol Penyakit Layu (Fusarium
oxsysporum f.sp. cubense) pada Tanaman
Pisang.
2/2
Floratek
3.
2007
Analisis Introduksi Trichoderma harzianum
dan Pupuk Bokhasi untuk Pengendalian
Layu Skelrotium Pada Tanaman Kedelqai
Varietas Lokal.
11/2
Agrista
35
4. 2008 Aplikasi Urine Sapi sebagai Biofungisida Pyricularia oryzae pada Tanaman Padi.
1/1 Rona Lingkungan
Hidup
5.
2008
Kompatibilitas Spodoptera litura Nuclear
Polyhedrosis Virus (SlNPV) dengan Ekstrak
Gadung Racun (Dioscorea hispida Denst)
terhadap Mortalitas Larva Spodoptera litura
Fab.
12/2
Agrista
6.
2009
Pemanfaatan Kascing untuk Menghambat
Perkembangan Fusarium oxysporum pada
Tanaman Tomat
13/3
Agrista
7.
2010
Aplikasi Pupuk Hayati dan Pupuk Kandang
untuk Pengendalian Hama Lalat Bibit
(Ophiomyia phaseoli) pada Tanaman
Kedelai.
5/2
Floratek
8.
2010
Dosis dan Frekuensi Kascing untuk
Pengendalian Penyakit Layu Fusarium pada
Tanaman Tomat
5/2
Floratek
9.
2010
Kemampuan Fungi Mikoriza Arbuskula
(FMA) dalam Menekan Perkembangan
Colletotricum capsici Penyebab Antraknosa
pada Cabai Merah (Capsicum annum L.)
12/2
Penelitian
Universitas
Jambi
(Seri Sains)
10. 2010 Detected and Characterize the Endophytic
Fungal Associated on Leaf Area Cacao
(Theobroma cacao L) Tree in East Aceh
Proceeding
ISHSFS
11. 2011 Pengaruh Dosis Fungi Mikoriza Arbuscular
(FMA) dan Trichoderma virens untuk
Mengendalikan Sclerotium rolfsii pada
Tanaman kedelai
4/2
Rona
Lingkungan
Hidup
Semua data yang saya isi dan tercantum dalam biodata ini adalah benar dan dapat
dipertanggungjawabkan secara hukum. Apabila di kemudian hari ternyata dijumpai
ketidak-sesuaian dengan kenyataan, saya sanggup menerima risikonya. Demikian biodata
ini saya buat dengan sebenarnya untuk memenuhi salah satu persyaratan dalam pengajuan
Hibah Penelitian Disertasi Doktor Tahun Anggaran 2017.
Banda Aceh, Oktober 2017
Hormat saya,
Ir. Susanna, M.Si.
NIP. 196811301994032001
Susanna Hasan <[email protected]>
[WJST] Submission Acknowledgement1 pesan
Editor of Walailak J Sci & Tech <[email protected]> 7 September 2017 14.47Kepada: Susanna Hasan <[email protected]>
Dear Professor Susanna Hasan:
Thank you for submitting the manuscript, "DIAGNOSIS OF DIEBACK DISEASE OFTHE NUTMEG TREE IN ACEH SELATAN" to Walailak Journal of Science andTechnology (WJST). With the online journal management system that we areusing, you will be able to track its progress through the editorial processby logging in to the journal web site:
Manuscript URL: http://wjst.wu.ac.th/index.php/wjst/author/submission/4379Username: terataikami
If you have any questions, please contact me. Thank you for considering thisjournal as a venue for your work and hope you will enjoy publishing in thisjournal, and look forward to receiving articles from you in future.
PS. We encourage authors to submit the names of 5-7 referees suitable toreview the work to aid in the peer review process.
Editor of Walailak J Sci & TechWalailak Journal of Science and Technology (WJST)2014 SCImago Journal Rank (SJR): 0.207________________________________________________________________________Walailak Journal of Science and Technology (WJST)http://wjst.wu.ac.th2015 SCImago Journal Rank (SCOPUS): 0.199 (Q2)
Gmail - [WJST] Submission Acknowledgement https://mail.google.com/mail/u/0/?ui=2&ik=4951de0b96&jsver=9hi...
1 of 1 18/09/2017 20:57

Active Submissions http://wjst.wu.ac.th/index.php/wjst/author
1 of 2 18/09/2017 21:01

Active Submissions http://wjst.wu.ac.th/index.php/wjst/author
2 of 2 18/09/2017 21:01

http://wjst.wu.ac.th What are types of article? (MiniReview, Article)
Walailak J Sci & Tech 2016; 13(xx): xxx-xxx.
DIAGNOSIS OF DIEBACK DISEASE OF THE NUTMEG TREE IN
ACEH SELATAN
Susanna*, Meity S. Sinaga
**, Suryo Wiyono**, Hermanu Triwidodo
**
*Studi Program of Plant Protection, Faculty of Agriculture, Syiah Kuala University, Banda Aceh, Aceh
**Department of Plant Protection, Faculty of Agriculture, Bogor Agriculture University
(*Corresponding author’s e-mail: [email protected])
Received: xxx, Revised: xxx, Accepted: xxx (Times New Roman, 10 pt, Italic)
Running title
Diagnosis of Dieback Diesease of The Nutmeg Tree
Abstract
Effective control disease measures require a great deal of knowledge about the pathogen, which is
also applied to dieback disease of the nutmeg tree in Aceh Selatan District. The objectives of this study
were to identify the causal agent of dieback disease on the nutmeg plantation and to assess the effect of
drought stress on the development of dieback disease. Pathogen identification was carried out by
observing the symptoms of infected plants, morphological characters of the isolated pathogens, Koch’s
postulates, as well as molecular analysis and pure pathogen cultures. The samples were taken from an
infected part of the nutmeg plant (twigs, stems, and roots) in six subdistricts at the center of the nutmeg
plantation in Aceh Selatan. Koch's postulate used the mycelia of pathogens on one-year-old nutmeg
seedlings. Drought stress was simulated everyday and then once every two weeks, to be able to study the
effect of drought stress on dieback. The results showed that there were four genera of fungal pathogens
(Botryodiplodia theobromae, Fusarium solani, Graphium euwallaceae, and Rigidoporus microporus) out
of six fungi associated with dieback disease. However, B. Theobromae Pat. turned out to be the dominant
fungi associated with dieback on the nutmeg tree in Aceh Selatan. Drought stress was proven to be a
predisposing factor for explaining this problem epidemic in Aceh Selatan.

Diagnosis of Dieback Disease of The Nutmeg Tree Susanna, Meity S.Sinaga, Suryo Wiyono, and Hermanu Triwidodo
http://wjst.wu.ac.th
Walailak J Sci & Tech 2016; 13(xx)
2
Keywords: Drought stress, Koch’s postulates, pathogen, Botryodiplodia theobromae
Introduction
Nutmeg (Myristica fragrans) is an important plantation commodity for increasing Indonesia’s
foreign exchange and the livelihood of Indonesian society in general. As the world's largest nutmeg
supplier, Indonesia contributes 70% -75%, while the remaining 20% comes from Grenada and 5% from
Sri Lanka, Trinidad and Tobago. The most significant users of nutmeg products are the United States,
Britain, and Germany [4].
Aceh Selatan District is one of the centers of nutmeg production in Indonesia. The nutmeg
production of this region contributes greatly to the fulfillment of domestic and foreign nutmeg needs. The
area of nutmeg plantations in Aceh Selatan District in 1994 reached 11,245 ha with an average
productivity of whole nutmeg of 8.2 ton/ha. In 2003 the total area had decreased to 9,843 ha and the
average total productivity decreased to 1.1 tons/ha. In 2014, the average value of Aceh nutmeg
productivity had decreased to 0.7 tons/ha [3].
One of the factors that cause the declining productivity of nutmeg plants in Aceh Selatan is the so-
called ‘die-back’ disease. Cases of this disease continue to spread throughout the nutmeg production
centers in Aceh Selatan district. This has led to a production decline up to 70% [5]. Until 2016, the cause
of the disease was not known for certain. Symptoms of die-back disease appear as necrosis of leaves,
stems, branches, and twigs in the form of die-back that occur gradually and eventually result in complete
die-back. Pathogens can attack and cause death in Nutmeg plants from the nursery stage, young plants
that are not yet or already in production, or old ones [2, 6, 7].
The development of this disease may have been affected by prolonged drought impacts in
Indonesia's land areas and, at the same time, changes in the ratio between the length of the dry and rainy
seasons. This phenomenon has a negative impact on the agricultural sector, especially in terms of setting
the planting time and also causing the explosion of pest and plant diseases. The district of Aceh Selatan is
located in the West/South coast area, which is predicted to be affected by drought caused by the dipole

Diagnosis of Dieback Disease of The Nutmeg Tree Susanna, Meity S.Sinaga, Suryo Wiyono, and Hermanu Triwidodo
http://wjst.wu.ac.th
Walailak J Sci & Tech 2016; 13(xx)
3
mode phenomenon [1]. In some cases, drought conditions will have an impact on the disease by making
the environment more or less favorable for infection, disease progression or disease spread. On the other
hand, drought exacerbates the damage caused by disease in drought-affected plants.
Knowledge of the cause of the disease and the trigger factor of its development is needed to develop
its control strategy. An understanding of the pathogens, as well as the factors driving the occurrence of
the epidemic, is not yet complete. Therefore, a research programme has been conducted to identify the
cause of the disease and to find out the effect of drought stress on die-back disease on the nutmeg tree in
Aceh Selatan.
Materials and methods
Research was conducted at Laboratorium Mikologi Tumbuhan Fakultas Pertanian IPB,
Laboratorium Perlindungan Hutan, PusLitbang Hutan Bogor, and in the experimental grounds at Kp.
Cimanggis, Bogor, from September 2015–September 2016.
Disease Diagnosis and Identification of the Disease Cause
Infected plant samples (twigs, stems and roots) were taken from thirty six nutmeg plantations in six
sub-districts (Labuhan Haji Timur, Meukek, Sawang, Samadua, Tapaktuan and Pasie Raja) in Aceh
Selatan. Samples were taken from plant tissues (stems, twigs, and roots) showing symptoms of die-back,
leaf yellowing and necrosis. The samples of plant parts were surface sterilized, isolated, and incubated on
PDA media for three days at room temperature. The pathogen that grows is isolated and purified on the
PDA, then used to perform Koch’s postulates. Koch’s postulates were also performed on year-old
Myristica fragrans nutmeg seedlings. Inoculation was achieved by attaching cultures to stems or roots
that have been injured and then wrapped with sterile cotton. Each treatment uses five nutmeg seedlings.
Observations were made daily on the development of symptoms in seedlings.
Identification of pathogenic morphology refers to Street [12]; Leslie and Summerrel [9]; and
Watanabe [15]. Molecular identification refers to Sambrook and Russell [11]. The amplification product

Diagnosis of Dieback Disease of The Nutmeg Tree Susanna, Meity S.Sinaga, Suryo Wiyono, and Hermanu Triwidodo
http://wjst.wu.ac.th
Walailak J Sci & Tech 2016; 13(xx)
4
produced was sent to Bioneer (Korea) to rank the nucleotides. The DNA sequence was identified using
BLAST and the data was compared with the NCBI GenBank data.
Drought Relation with Disease Occurrence
The association of drought with disease occurrence was analyzed based on rainfall data in Aceh
Selatan during the last fifteen years. Climate data was obtained from local BMKG and disease data for six
years was obtained from the Plantation General Directorate. Simulation of artificial drought stress was
achieved in the laboratory by watering once every two weeks, compared with daily watering. Artificial
inoculation uses pathogens detected in previous studies on one-year-old plant seedlings. Observations
were made daily; the observed variables were the latent period and the development of symptoms on
nutmeg seedlings. The water content of the soil was also measured during the treatment.
Results and discussion
Infected nutmeg plants in the field usually show initial symptoms of dieback, i.e. dried and withered
leaves on some twigs, which then spread from one branch to another until the whole part of the plant is
affected. The leaves become brownish, curling, and hanging and eventually fall. However there are plants
that wilt as a whole, showing yellowing, drying, falling leaves, and ultimately complete death.
Based on field observations on nutmeg plant, it is known that every symptom that occurs in infected
plants is clearly associated with die-back, either positively or negatively. Symptoms of die-back are
clearly associated with leaf discoloration, cancer, borers, and fungal rhizomorphs. The associated
symptoms are clearly exclusive, thus if one symptom occurs, the other symptoms are absent, for example
if one of these plants show wilting symptoms it does not show other symptoms of dieback, such as leaves
hanging, cancer, stem borers, or leaf loss (Table 1).
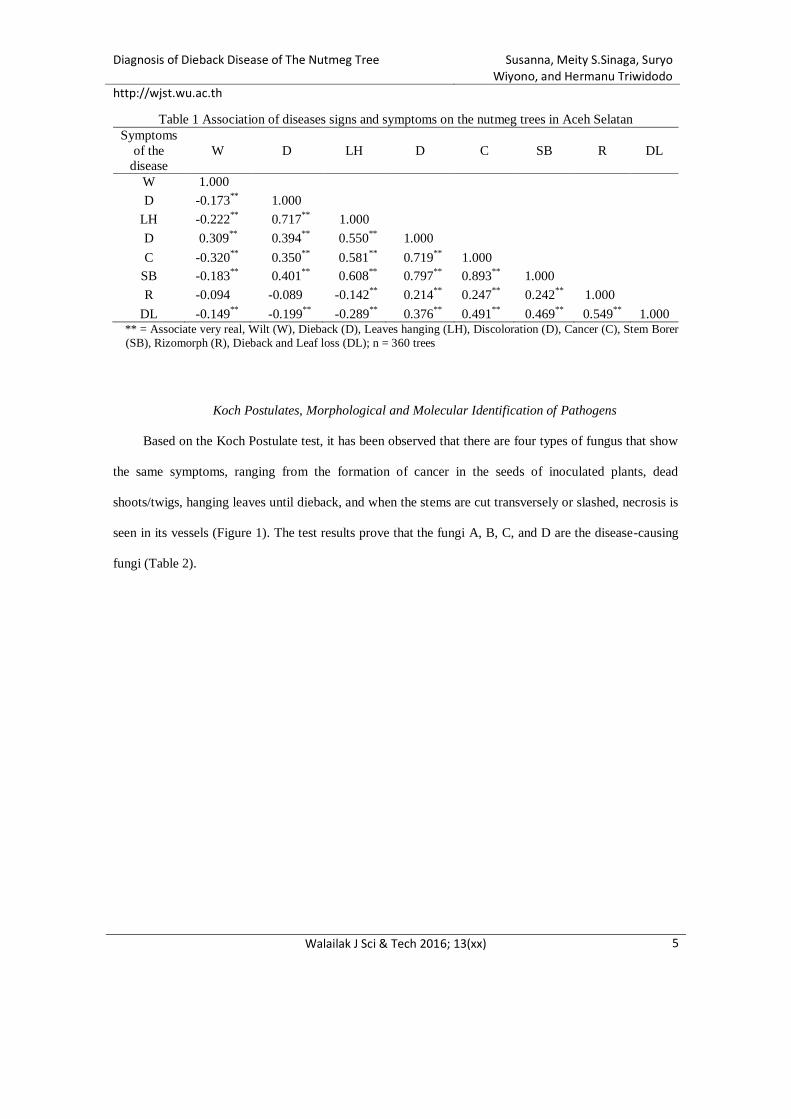
Diagnosis of Dieback Disease of The Nutmeg Tree Susanna, Meity S.Sinaga, Suryo Wiyono, and Hermanu Triwidodo
http://wjst.wu.ac.th
Walailak J Sci & Tech 2016; 13(xx)
5
Table 1 Association of diseases signs and symptoms on the nutmeg trees in Aceh Selatan
Symptoms
of the
disease
W D LH D C SB R DL
W 1.000
D -0.173**
1.000
LH -0.222**
0.717**
1.000
D 0.309**
0.394**
0.550**
1.000
C -0.320**
0.350** 0.581
** 0.719
** 1.000
SB -0.183**
0.401**
0.608**
0.797**
0.893**
1.000
R -0.094 -0.089 -0.142**
0.214**
0.247**
0.242**
1.000
DL -0.149**
-0.199**
-0.289**
0.376**
0.491**
0.469**
0.549**
1.000 ** = Associate very real, Wilt (W), Dieback (D), Leaves hanging (LH), Discoloration (D), Cancer (C), Stem Borer
(SB), Rizomorph (R), Dieback and Leaf loss (DL); n = 360 trees
Koch Postulates, Morphological and Molecular Identification of Pathogens
Based on the Koch Postulate test, it has been observed that there are four types of fungus that show
the same symptoms, ranging from the formation of cancer in the seeds of inoculated plants, dead
shoots/twigs, hanging leaves until dieback, and when the stems are cut transversely or slashed, necrosis is
seen in its vessels (Figure 1). The test results prove that the fungi A, B, C, and D are the disease-causing
fungi (Table 2).

Diagnosis of Dieback Disease of The Nutmeg Tree Susanna, Meity S.Sinaga, Suryo Wiyono, and Hermanu Triwidodo
http://wjst.wu.ac.th
Walailak J Sci & Tech 2016; 13(xx)
6
Infection of seedlings Cancer/root rot Discoloration/rot
Figure 1 Results of artificial inoculation of the nutmeg seedlings; Inoculation of Botryodiplodia
sp. (A); Inoculation of Graphium sp. (B); inoculation of Fusarium solani (C); Inoculation of Rigidoporus sp. (D); Without pathogens (K).
The initial symptom of artificial inoculation is cancer, with an average incubation period of thirty-
two days. The symptoms formed were initially not developed but after the seeds received drought
treatment, a month later symptoms of died shoots and twigs began to appear and then spread to all parts
of the plant, continued with the drying leaves, which were left hanging for four to eight weeks, then died,
and the plant suffered full dieback. The seeds of nutmeg plants inoculated with isolates of fungus D did
not show symptoms of cancer but displayed root rot symptoms because the pathogen was a fungus

Diagnosis of Dieback Disease of The Nutmeg Tree Susanna, Meity S.Sinaga, Suryo Wiyono, and Hermanu Triwidodo
http://wjst.wu.ac.th
Walailak J Sci & Tech 2016; 13(xx)
7
isolated from the root parts of the nutmeg plant, showing the fungal hyphae (rhizomorph) that wrapped
around the root parts of the plant, but the symptoms formed on the upper parts of the plant showing
similar symptoms to those caused by other fungus tests.
Table 2 Symptoms of an isolated fungus
No. Fungus Result of postulat
Koch Result of identification
Frequency of isolates
Stem
(36)*
Twig
(36)*
Root
(36)*
1 A + Botryodilodia sp. 36 36 36
2 B + Graphium sp. 4 4 -
3 C + Fusarium solani 2 2 5
4 D + Rigidoporus sp. - - 2
5 E - Endomelanconiopsis sp. 1 - -
6 F - Clonotachys sp. 2 - -
Description: + = exists (positive at all stages of Koch postulate); - = none; * = The numbers in brackets
indicate the total number of samples
The frequency of the fungus when isolated from stricken plant tissue from thirty six sample sites
was very variable. Botryodiplodia sp. is the most dominant type of fungus found; it can even be found by
the direct spore trapping technique using MA and PDA media around the stricken plants. Four fungi (A,
B, C, and D) found in association with dieback plants give positive results at all stages of Koch
postulates. This proves that all four fungi can cause die-back symptoms. After the re-isolation of the test
plant, the four fungi showed similar results to the initially isolated pathogenic fungi (Table 2).
The isolated fungi were from six genera but only four isolates were pathogenic. Isolate A was
identified as Botryodiplodia sp., with white early colonies and very rapid growth of aerial mycelia, which
then turn gray on the fourth day and turn black on the seventh day. Microscopically the fungus has a
hyaline as initial hyphae that turns dark, can form pycnidia within ± 30 days after isolation by producing a
not insulated young hyaline conidia that later show a color change to brown, with partition in the center.
Isolate B is identified as Graphium sp. with white early colonies turning gray, and after seven days
turning black. This fungus has a conidiophore bond forming synema and conidia produced in cylindrical
form. Isolate C was identified as F. solani that forms a white colony with thin mycelium, has a long
conidiophore which produces two types of conidia, i.e. microconidia with either a partition or not; and

Diagnosis of Dieback Disease of The Nutmeg Tree Susanna, Meity S.Sinaga, Suryo Wiyono, and Hermanu Triwidodo
http://wjst.wu.ac.th
Walailak J Sci & Tech 2016; 13(xx)
8
macroconidia with a partition between two and four, hyaline chlamydospores that are formed on the
intercalary or hyphae tip. Isolate D was identified as Rigidoporus sp. because it forms a thin white colony,
with hyaline hyphae and branches but does not have clam connection. The four isolates have
characteristics corresponding to the genus on the identification keys used (Figure 2).
Fungi Colony Conidiophore Conidia
Figure 2 Morphology of colonies on PDA medium and microscopic structure of pathogenic fungi
from nutmeg.
Furthermore, the molecular identification process was performed to confirm the morphologically
identified fungal species. Visualization of PCR results showed that band amplification performed using
primers ITS1 and ITS4 looked as expected. Amplification of DNA bands formed on agarose has a size of
about ~ 500-650 bp (Figure 3). The size corresponds to a group of fungi.

Diagnosis of Dieback Disease of The Nutmeg Tree Susanna, Meity S.Sinaga, Suryo Wiyono, and Hermanu Triwidodo
http://wjst.wu.ac.th
Walailak J Sci & Tech 2016; 13(xx)
9
Figure 3 Electrophoresis visualization of PCR results of 4 fungi isolates using primary ITS1 and ITS4.
M = Marker; 1. Botryodiplodia sp .; 2. Graphium sp .; 3. F. solani; and 4. Rigidoporus sp.
According to Toju et al. [14], primary ITS is suitable for identifying field samples from the
Ascomycetes and Basidiomycetes fungi based on DNA strands. However, the fungus of the
Deuteromycetes group can also be detected with the primary ITS.
Pathogens A, B, C, and D, which were isolated from nutmeg samples, were identified as
Botryodiplodia theobromae (syn. Lasiodiplodia theobromae), Graphium euwallaceae, Fusarium solani,
and Rigidoporus microporus with 100% query cover characterization, 99% -100% identity and e-value
0.0 (Table 3).
Table 3 Identification of pathogenic fungi based on ITS compared with NCBI GenBank data
Isolate Description No. Assesion Strain Query cover
(%)
Identity
(%) E-value
A (LH) L. theobromae KM278132.1 NZD mf2 100 100 0.0
B (MK) G. euwallaceae KF54022.1 UCRFD97 100 100 0.0
C (TT) F. solani KU377470.1 C219 100 100 0.0
D (PR) R. microporus KM246744.1 RL 100 99 0.0
Description: - = is being re-sequenced
The Effect of Drought Stress on Dieback and Leaf Loss
What is important in understanding the epidemic of this disease is what triggers the epidemic.
Climatic data over the past fifteen years indicates that rainfall in South Aceh district has declined by as
much as 50.5% (Figure 4).
M 1 2 3 4
500 bp 650 bp
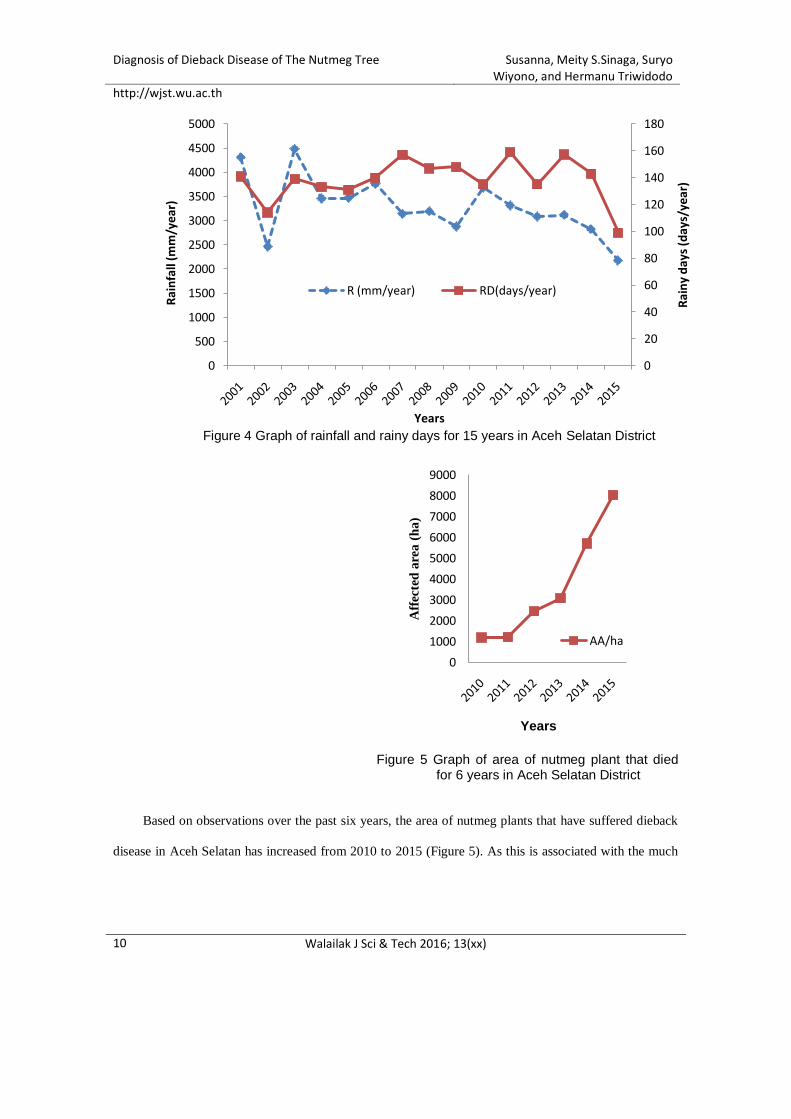
Diagnosis of Dieback Disease of The Nutmeg Tree Susanna, Meity S.Sinaga, Suryo Wiyono, and Hermanu Triwidodo
http://wjst.wu.ac.th
Walailak J Sci & Tech 2016; 13(xx)
10
Figure 4 Graph of rainfall and rainy days for 15 years in Aceh Selatan District
Figure 5 Graph of area of nutmeg plant that died
for 6 years in Aceh Selatan District
Based on observations over the past six years, the area of nutmeg plants that have suffered dieback
disease in Aceh Selatan has increased from 2010 to 2015 (Figure 5). As this is associated with the much
0
20
40
60
80
100
120
140
160
180
0
500
1000
1500
2000
2500
3000
3500
4000
4500
5000
Rai
ny
day
s (d
ays/
year
)
Rai
nfa
ll (m
m/y
ear)
Years
R (mm/year) RD(days/year)
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
Aff
ecte
d a
rea (
ha)
AA/ha
Years
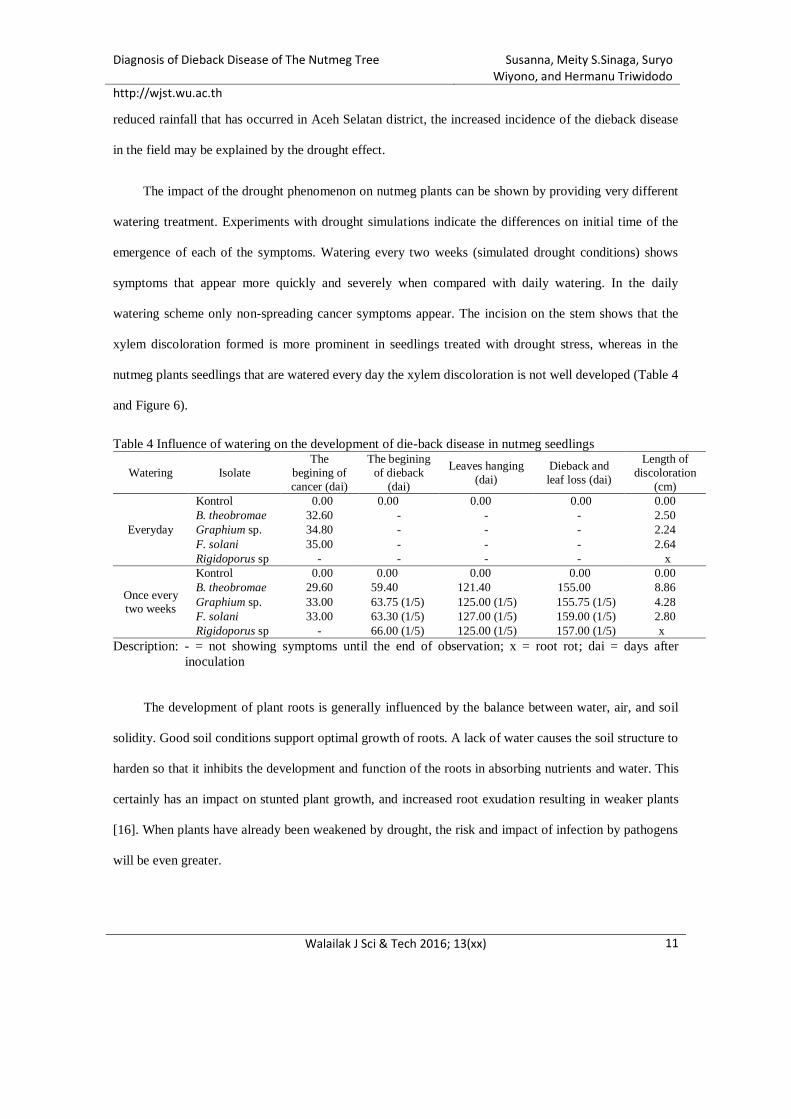
Diagnosis of Dieback Disease of The Nutmeg Tree Susanna, Meity S.Sinaga, Suryo Wiyono, and Hermanu Triwidodo
http://wjst.wu.ac.th
Walailak J Sci & Tech 2016; 13(xx)
11
reduced rainfall that has occurred in Aceh Selatan district, the increased incidence of the dieback disease
in the field may be explained by the drought effect.
The impact of the drought phenomenon on nutmeg plants can be shown by providing very different
watering treatment. Experiments with drought simulations indicate the differences on initial time of the
emergence of each of the symptoms. Watering every two weeks (simulated drought conditions) shows
symptoms that appear more quickly and severely when compared with daily watering. In the daily
watering scheme only non-spreading cancer symptoms appear. The incision on the stem shows that the
xylem discoloration formed is more prominent in seedlings treated with drought stress, whereas in the
nutmeg plants seedlings that are watered every day the xylem discoloration is not well developed (Table 4
and Figure 6).
Table 4 Influence of watering on the development of die-back disease in nutmeg seedlings
Watering Isolate
The
begining of
cancer (dai)
The begining
of dieback
(dai)
Leaves hanging
(dai)
Dieback and
leaf loss (dai)
Length of
discoloration
(cm)
Everyday
Kontrol 0.00 0.00 0.00 0.00 0.00
B. theobromae 32.60 - - - 2.50
Graphium sp. 34.80 - - - 2.24
F. solani 35.00 - - - 2.64
Rigidoporus sp - - - - x
Once every two weeks
Kontrol 0.00 0.00 0.00 0.00 0.00
B. theobromae 29.60 59.40 121.40 155.00 8.86
Graphium sp. 33.00 63.75 (1/5) 125.00 (1/5) 155.75 (1/5) 4.28
F. solani 33.00 63.30 (1/5) 127.00 (1/5) 159.00 (1/5) 2.80
Rigidoporus sp - 66.00 (1/5) 125.00 (1/5) 157.00 (1/5) x
Description: - = not showing symptoms until the end of observation; x = root rot; dai = days after
inoculation
The development of plant roots is generally influenced by the balance between water, air, and soil
solidity. Good soil conditions support optimal growth of roots. A lack of water causes the soil structure to
harden so that it inhibits the development and function of the roots in absorbing nutrients and water. This
certainly has an impact on stunted plant growth, and increased root exudation resulting in weaker plants
[16]. When plants have already been weakened by drought, the risk and impact of infection by pathogens
will be even greater.

Diagnosis of Dieback Disease of The Nutmeg Tree Susanna, Meity S.Sinaga, Suryo Wiyono, and Hermanu Triwidodo
http://wjst.wu.ac.th
Walailak J Sci & Tech 2016; 13(xx)
12
Symptoms of discoloration on the longitudinal incision of the stem Treatments
Figure 6 Symptoms of discoloration due to the effects of drought stress
Watering every two weeks, as a simulation of drought stress, induces very dry soil conditions at
about 25% of field capacity. This condition causes the plant to be severely stressed and weakened. The
district of Aceh Selatan has experienced a serious decline in rainfall over the last fifteen years. This is
likely to cause the plants to become stressed and thus more vulnerable to attack by pathogens. As a result
the occurrence of dieback disease has increased. For certain diseases such as stem and root rot, the
intensity of the attack is strongly influenced by the drought stress factor: the drier the condition is, the
higher disease’s intensity [7, 8, 13].
Drought affects the biochemical balance and concentration of many components in plant tissues,
including nitrogen compounds such as amino acids and nitrates; osmolites such as sugars and inorganic
ions; and chemical alleles, such as cyanogenic glycosides, terpenoids, and alkaloids. The increasing
concentration of these compounds may result in inhibition of plant growth, photosynthesis, and nutrient
uptake, so plants become more vulnerable to attack by pathogenic organisms. Additional important
factors for determining disease severity under dry conditions are pathogen status, plant part, and water
stress intensity [7].
Conclusions
The cause of die-back disease of nutmeg trees in South Aceh is Botryodiplodia theobromae Pat.
(syn. Lasiodiplodia theobromae (Patouillard) Griffon & Maubland). The epidemic of the die-back disease
Without pathogens on watering every two
weeks
Watering every two weeks
Watering everyday
Without pathogen on watering everyday

Diagnosis of Dieback Disease of The Nutmeg Tree Susanna, Meity S.Sinaga, Suryo Wiyono, and Hermanu Triwidodo
http://wjst.wu.ac.th
Walailak J Sci & Tech 2016; 13(xx)
13
began from 2003. The dramatic decline of rainfall over the last fifteen years is a factor in triggering the
dieback epidemic of nutmeg trees. The lower the rainfall is, the more likely the disease to happen.
Acknowledgements
The authors are grateful to the Ministry of Research, Technology and High Education, Indonesia for
their financial to conduct this research work.
References
[1] [BMKG] Badan Meteorologi, Klimatologi, dan Geofisika. 2017. Informasi Perubahan Iklim.
Stasiun Klimatologi Kelas IV Aceh Besar. Aceh (ID).
[2] Corcobado T, Cubera E, Juarez E, Moreno G, Solla A. 2014. Drought events determine
performance of Quercus ilex seedlings and increase their susceptibility to Phytophthora cinnamomi.
Agric. For. Meteor. 1 (8): 192 – 193.
[3] [Dishutbun] Dinas Kehutanan dan Perkebunan. 2014. Pedoman Pengamatan dan Pengendalian
Hama pada Tanaman Pala. Tapaktuan, Aceh Selatan.
[4] [Ditjenbun] Direktorat Jenderal Perkebunan. 2016. Statistik Perkebunan Indonesia 2016. Jakarta
(ID). Dirjenbun Indonesia.
[5] Hadad EA, Firman C. 2003. Budidaya Pala. Balittro. Bogor (ID).
[6] Harni R, dan Trisawa IM. 2011. Observasi dan identifikasi penyebab matinya pala di daerah Aceh
Selatan. Laporan Kerjasama Dinas Kehutanan dan Perkebunan dengan Balai Penelitian Tanaman
Rempah dan Aneka Tanaman Industri. Sukabumi.
[7] Jactel H, Petit J, Loustau ML, Delzon S, Piou D, Battisti A, and Koricheva J. 2012. Drought effects
on damage by forest insects and pathogens: a meta – analysis. Global Change Biol. 18: 267–276.
DOI: 10.1111/j.1365-2486.2011.02512.x

Diagnosis of Dieback Disease of The Nutmeg Tree Susanna, Meity S.Sinaga, Suryo Wiyono, and Hermanu Triwidodo
http://wjst.wu.ac.th
Walailak J Sci & Tech 2016; 13(xx)
14
[8] Kolb TE, Fettig CJ, Ayres MP, Bentz BJ, Hicke JA, Mathiasen R, Stewart JE, and Weed AS. 2016.
Observed and anticipated impacts of drought on forest insects and disease in the United States. For.
Ecol. and Management 380: 321 – 334.
[9] Leslie JS. and Summerel BA. 2006. The Fusarium Laboratory Mannual. 1st ed. Blackwell
Publishing. Ltd. (US): Iowa.
[10] Lipiec J, Doussan C, Nosalewicz A, and Kondracka K. 2013. Effect of drought and heat stress on
plant growth and yield: a review. Int. Agrophys. 27: 463 – 477. DOI: 10.2478/intag-2013-0017
[11] Sambrook J. and Russel DW. 2001. Molecular Cloning a Laboratory Manual. 3th ed. Cold Spring
Harbor Laboratory Pr. (US): New York.
[12] Streets RB. 1972. The Diagnosis of Plant Diseases: A Field and Laboratory Manual Emphasizing
The Most Practical Methode for Rapid Identification. The University of Arizona Press. (US):
Tuscon – Arizona
[13] Sturrock RN, Frankel SJ, Brown AV, Hennon PE, Kliejunas JT, Lewis KJ, Worrall JJ, and Woods
AJ. 2011. Climate change and forest diseases. Plant Pathol. 60:133 - 149. Doi: 10.1111/j.1365-
3059.2010.02406.x
[14] Toju H, Tanabe AS, Yamamoto S, and Sato H. 2012. High-covarage ITS primers for the DNA-
based identification of Ascomycetes and Basidiomycetes in environmental samples. Plos. ONE 7(7)
: 1-11.
[15] Watanabe T. 1994. Pictorial Atlas of Soil and Seed Fungi: Morphologies of Cultured Fungi and
Key to Species. Lewis Publishers. (JP): Tokyo
[16] Whitmore AP. and Whalley WR. 2009. Physical effects of soil drying on roots and crop growth. J.
Exp. Bot. 60 (10): 2845 – 2857.

Diagnosis of Dieback Disease of The Nutmeg Tree Susanna, Meity S.Sinaga, Suryo Wiyono, and Hermanu Triwidodo
http://wjst.wu.ac.th
Walailak J Sci & Tech 2016; 13(xx)
15
Names of 5-9 referees
1. Dr. Evi Toding Tondok, S.P., M.Sc.Agr., Departemen Proteksi Tanaman Fakultas Pertanian
Institut Pertanian Bogor, Jawa Barat, e-mail: [email protected]
2. Dr. Ir. Kikin Hamzah M., M.Sc., Departemen Proteksi Tanaman Fakultas Pertanian Institut
Pertanian Bogor, Jawa Barat , e-mail: [email protected]
3. Prof. Dr. Ir. Sri Hendrastuti Hidayat, M.Sc., Departemen Proteksi Tanaman Pertanian institute
Pertanian Bogor, Jawa Barat, e-mail: [email protected]
4. Dr. Ir. Bonny Poernomo Wahyu Soekarno, M.S., Departemen Proteksi Tanaman Institut
Pertanian Bogor, Jawa Barat, e-mail: [email protected]
5. Thomas E. Kolb., School of Forestry, Northern Arizona University, flagstaff, AZ, United States,
e-mail: [email protected]
6. Matthew P. Ayres, Department of Biological Sciences, Dartmounth College, Hanover, NH,
United States, e-mail: [email protected]

Diagnosis of Dieback Disease of The Nutmeg Tree Susanna, Meity S.Sinaga, Suryo Wiyono, and Hermanu Triwidodo
http://wjst.wu.ac.th
Walailak J Sci & Tech 2016; 13(xx)
16

Diagnosis of Dieback Disease of The Nutmeg Tree Susanna, Meity S.Sinaga, Suryo Wiyono, and Hermanu Triwidodo
http://wjst.wu.ac.th
Walailak J Sci & Tech 2016; 13(xx)
17