desain primer spesifik untuk deteksi dini penyakit
TRANSCRIPT

DESAIN PRIMER SPESIFIK UNTUK DETEKSI DINIPENYAKIT VIBRIOSIS PADA UDANG PENAEID
Ince Ayu Khairana Kadriah*), Endang Susianingsih*), Sukenda**),Munti Yuhana**), dan Enang Harris**)
*) Balai Penelitian dan Pengembangan Budidaya Air PayauJl. Makmur Dg. Sitakka No. 129, Maros 90512, Sulawesi Selatan
E-mail: [email protected]
**) Departemen Budidaya Perairan, Fakultas Perikanan dan Ilmu Kelautan,Institut Pertanian Bogor
Jl. Rasamala, Kampus IPB Darmaga, Bogor 16680
(Naskah diterima: 13 Juli 2012; Disetujui publikasi: 28 Februari 2013)
ABSTRAK
Serangan Vibriosis, yang disebabkan oleh Vibrio harveyi berpendar pada budidayaudang telah menyebabkan penurunan yang signifikan dalam produksi, baik padapembenihan maupun di tambak pembesaran. Pengembangan metode deteksi cepatberbasis PCR (Polymerase Chain Reaction) sangat penting untuk mencegah penularanvibriosis. Tujuan dari penelitian ini adalah untuk mengembangkan metode cepat deteksivibriosis pada udang penaeid dengan menggunakan penanda molekuler yang spesifik.PCR berbasis deteksi gen spesifik dilakukan menggunakan primer spesifik toxR,haemolysin (vvh), dan gyrB. Dari 35 isolat, 22 isolat yang terdeteksi memiliki genspesifik toxR, haemolysin (vvh) dan gen gyrB dan 9 isolat terdeteksi memiliki dua gentertentu. Penanda molekuler spesifik telah dirancang menggunakan data urutan genpenyandi protein haemolysin dan gyrase. Desain pasangan primer yang didasarkanpada program perangkat lunak dari Primer3 dan secara manual menggunakan programperangkat lunak Bioedit. Tiga pasangan primer untuk gen haemolysin dan dua primergyrase telah diperoleh dan dipilih sebagai primer.
KATA KUNCI: vibriosis, udang penaeid, PCR, deteksi cepat, penandamolekuler spesifik
ABSTRACT: Specific primer design for rapid detection of Vibriosis onpenaeid shrimp. By: Ince Ayu Khairana Kadriah, EndangSusianingsih, Sukenda, Munti Yuhana, and Enang Harris
Vibriosis occurrences, due to luminous Vibrio harveyi, on shrimp culture maycaused substantial declines in production, either in hatcheries or in grow out ponds.Development of Polymerase Chain Reaction (PCR)-based rapid detection methods isvery crucial in preventing vibriosis outbreaks. The aims of this study was to developrapid methods of detection of the vibriosis in penaeid shrimp by using specificmolecular markers. PCR-based detection of specific genes were performed employingspecific primers of toxR, haemolysin (vvh) and gyrB. Out of the 35 isolates, 22 isolateswere detected to have toxR, haemolysin (vvh) and gyrB specific genes and 9 isolateshave two out of these specific genes. Specific markers have been designed usingsequence data of the genes encoding the haemolysin protein and gyrase. Primerpairs design were based on the software program of Primer3 and manually aligned
Desain primer spesifik untuk deteksi dini penyakit ..... (Ince Ayu Khairana Kadriah)
131

by using Bioedit software program. Three of primer pairs for haemolysin gene andtwo of gyrase primers were obtained and selected as primer.
KEYWORDS: vibriosis, penaeid shrimp, PCR, rapid detection, specificmolecular marker
PENDAHULUAN
Kementerian Kelautan dan Perikanan masihmenempatkan udang sebagai komoditasunggulan perikanan budidaya selama 2010-2014. Selama periode 2010-2014 produksiudang diharapkan meningkat 74,75% ataudari 400 ribu ton menjadi 699 ribu ton yangterdiri atas udang vaname dan udang windu.Industri budidaya udang windu secara inten-sif dan transportasi udang windu ke seluruhdunia melalui perdagangan diketahui ber-hubungan erat dengan meningkatnya kejadianinfeksi penyakit yang menyerang udangwindu selama dua dekade ini (Saulnier et al.,2000). Bakteri vibrio berpendar adalah salahsatu penyebab penyakit yang cukup banyakmenyerang hewan budidaya seperti udang(Baticados et al., 1990; Karunasagar et al.,1994; Moriarty, 1998; Zhang & Austin, 2000),beberapa spesies ikan dan kekerangan (Aus-tin & Zhang, 2006) bahkan juga karang (Ben-Haim et al., 2003) di seluruh dunia.
Pengembangan metode deteksi cepat,tepat, akurat dan murah sangat bermanfaatkarena dapat digunakan dalam upaya pen-cegahan penyakit vibriosis di lapangan baikdi panti benih maupun pada pembesaranudang di tambak. Upaya pencegahan ini harusdilakukan sebelum koloni bakteri mencapaiquorum. Penelitian yang dilakukan olehDefoirdt (2007) menyimpulkan bahwa ke-mampuan bakteri vibrio untuk melakukanquorum sensing sangat dipengaruhi olehpopulasi bakteri tersebut di alam. Upaya untukdeteksi cepat secara molekular salah satu-nya dengan mengisolasi gen spesifik yangdimiliki oleh bakteri vibrio berpendar dandigunakan sebagai penanda molekular dalamdiagnosis cepat untuk penyakit vibriosis ber-pendar (kunang-kunang) pada budidaya udang.
Gen haemolysin diketahui merupakan genspesifik yang dimiliki bakteri Vibrio berpendar.Gen haemolysin adalah gen yang bertang-gung jawab pada penghancuran membran seldarah atau proses hemolisis. Gen yang meng-kode haemolysin ini dilaporkan ditemukanpada beberapa spesies bakteri di antaranya
adalah V. harveyi (Nishibuchi et al., 1990;Nishibuchi & Kaper, 1995; Zhang et al., 2001)dan V. parahaemolyticus (Bej et al., 1999).
Selain gen haemolysin gen toxR yangsecara spesifik mengkode protein trans-membran juga memegang peranan pentingpada regulasi gen toksin ctx dan beberapagen-gen toksin lainnya. Gen toxR akan meng-aktifkan gen-gen lainnya untuk menghasilkantoksin (Pang et al., 2006). Gen gyrB diketahuiberperan untuk mengkode protein subunit Bdari DNA gyrase (topoisomerase type II). DNAgyrase mengatur superkoiling pita ganda DNA.Gen gyrB sangat diperlukan untuk replikasiDNA di mana gen ini berperan dalam pem-bentukan protein yang mengkode enzimgyrase.
Pendekatan yang memanfaatkan kemajuanbioinformatika dan teknik PCR dapat diguna-kan untuk mengembangkan penanda spesifiktersebut. Metode PCR digunakan untuk meng-amplifikasi sekuen spesifik dari rantai DNA.Primer rantai pendek oligonukleotida yangdidesain akan berkomplemen dengan masing-masing ujung dari daerah target pada rantaiDNA dan kemudian memperpanjangnya padasisi yang berlawanan dari DNA template.Karena fungsi primer sebagai inisiator seka-ligus pembatas daerah yang akan diamplifi-kasi, maka idealnya primer memiliki urutanbasa nukleotida yang tepat berpasangandengan urutan basa DNA target yang akandiamplifikasi, dan tidak menempel di bagianlainnya. Desain primer yang bagus merupakanhal esensial bagi keberhasilan reaksi PCR.
Untuk mendesain suatu primer memer-lukan data-data sekuen gen yang menyandi-kan protein sejenis dengan gen yang akandiamplifikasi melalui PCR. Data-data sekuengen dapat diperoleh pada basis data gen darilembaga-lembaga penyedia informasi gen danprotein atau hasil sekuen gen tertentu yangbersifat spesifik yang telah dihasilkan. Prosesamplifikasi dari gen-gen yang mengkode sifat-sifat tertentu utamanya sifat patogen dapatmenjadi suatu pendekatan baru dalam metodedeteksi cepat penyakit (Cunningham, 2002).
132
J. Ris. Akuakultur Vol. 8 No. 1 Tahun 2013: 131-143

BAHAN DAN METODE
Penelitian dilakukan di Laboratorium Ke-sehatan Ikan dan Lingkungan Balai Penelitiandan Pengembangan Perikanan Budidaya AirPayau, Maros. Analisis sekuensing dilaksana-kan di Laboratorium Bioteknologi UNIKAAtmajaya, Jakarta dan Laboratorium 1st BaseSingapura. Tahapan Penelitian terdiri atas:
Isolasi Bakteri Vibrio Patogen dariPanti Benih dan Tambak Udang Windu
Sampel bakteri dan udang windu diko-leksi dari tambak percobaan Balai Penelitiandan Pengembangan Budidaya Air Payau,Maros di Maranak dan Takalar serta tambak-tambak udang windu di Barru, Pangkep,Pinrang, Banyuwangi, dan Bali. Bakteri diiso-lasi dari air tambak, sedimen tambak, danudang sakit. Sampel air diambil dengan meng-gunakan botol steril kemudian dibawa kelaboratorium. Sedangkan sampel sedimendiambil menggunakan sudip steril dan di-bawa ke laboratorium menggunakan botolsteril. Isolasi bakteri dilakukan dengan caramengambil 1 mL air sampel dan 1 g sedimentambak kemudian diencerkan secara ber-tingkat dalam larutan fisiologis (Benson, 1985)dan dikultur pada media TCBSA (ThiosulfateCitrate Bile Sucrose Agar). Bakteri juga diisolasidari hepatopankreas udang windu.
Deteksi Gen Spesifik pada BakteriVibrio Berpendar
Pada penelitian ini digunakan primerspesifik untuk mendeteksi gen-gen spesifiktoxR gene, haemolysin (vvh) gene dan GyrBgene dengan metode PCR (Tabel 1). Primer-primer ini digunakan untuk mendeteksi gen-gen spesifik pada isolat bakteri yang diisolasidari tambak dan panti benih di berbagai daerahdi Sulawesi Selatan dan Jawa.
Isolasi Genom Bakteri Kandidat
Metode yang digunakan dalam isolasigenom bakteri adalah metode phenol-chloro-form yang dikembangkan oleh Parenrengi(2000). Bakteri dikultur di dalam nutrient brothselama empat jam kemudian dipanen dengancara sentrifugasi. Sebanyak 1 mL biakan bakteridipindahkan ke dalam tabung eppendorf steril1,5 mL dan disentrifugasi (6.000 rpm; 10 menit).Proses sentrifugasi diulang sebanyak dua kalidan kemudian dilakukan pencucian denganlarutan fisiologis juga dengan sentrifugasi.
Pelet bakteri yang dihasilkan kurang lebih50 mg kemudian dicampur dengan 500 μLlysis buffer, 20 μL proteinase-K (stok 20 mg/mL), dan 40 μL sodium dodecyl sulfate (SDS)10%. Setelah itu, lisat bakteri diinkubasi dalamwaterbath selama 1-3 jam pada suhu 55oC.Penambahan 12,5 μL RNAse dilakukan sebe-lum lisat disimpan pada suhu ruang selama 15-30 menit. Selanjutnya ditambahkan Phenol:Chloroform:Isoamyl alcohol (PCIA 25:24:1)sebanyak 500 μL. Tabung eppendorf di-homogenkan dengan menggunakan minimixer secara perlahan sampai homogen dandisimpan pada suhu ruang selama 10 menit.Selanjutnya disentrifugasi pada kecepatan13.000 rpm selama delapan menit. Lapisanpaling atas diambil dan dipindahkan ke tabungeppendorf baru dan dilakukan penambahanPCIA seperti sebelumnya.
Setelah lapisan paling atas dipindahkan ketabung eppendorf baru, kemudian dilakukanpenambahan satu bagian larutan Chloroform:Isoamyl alcohol (CIA 24: 1) dan disentrifugasiselama empat menit dengan kecepatan 13.000rpm. Lapisan paling atas dipindahkan ke tabungeppendorf baru. Kemudian ditambahkan duabagian ethanol absolut dingin dan dicampurperlahan sampai homogen selanjutnya di-lakukan sentrifugasi dengan kecepatan 6.000rpm selama 30 menit. Cairan dibuang kemudianpelet DNA dicuci dengan 1 mL ethanol 70%kemudian disentrifugasi dengan kecepatan6.000 rpm selama 15 menit. Pellet DNA di-keringkan selama satu malam dan setelahkering ditambahkan 100 μL Buffer Tris-Etilendiamin Tetra Acetic Acid (EDTA) (TE) danselanjutnya disimpan pada suhu -20oC sampaidigunakan (Parenrengi, 2000).
Proses PCR
Deteksi gen spesifik dilakukan denganmelakukan amplifikasi DNA menggunakanteknik Polymerase Chain Reaction (PCR).Proses PCR dilakukan menggunakan Kit PCRReady To Go (RTG) dengan primer spesifikkomersil yang sudah ada dan dilakukanoptimasi pada beberapa tingkatan suhu an-nealing.
Proses ampifikasi DNA untuk primer toxRadalah sebagai berikut: larutan master mixdibuat dengan mencampur 20 μL aquadestmilliQ ke dalam 1 tube RTG (GE Healthcare UKLimited Little Chalfont Buckinghamshire, UK),kemudian ditambahkan masing-masing 1 μLPrimer toxRR1 dan toxRF1 (Pang et al., 2006).
Desain primer spesifik untuk deteksi dini penyakit ..... (Ince Ayu Khairana Kadriah)
133

Tab
el 1
.Pr
imer
spes
ifik
yan
g d
igunak
an u
ntu
k m
endet
eksi
keb
erad
aan g
en s
pes
ifik
Tab
le 1
.Sp
ecif
ic p
rim
ers
wer
e use
d t
o det
ect
the
pre
sence
of
spec
ific
gen
es
Sum
ber
(So
urc
e):
Can
o-G
om
ez e
t al. (
20
09
)
Ge
nPr
od
uk
gen
Na
ma
pri
mer
Se
ku
en
pri
me
r (5
' - 3
')Pa
nja
ng
Refe
ren
siG
enes
Gen
es p
rod
uct
Pri
mer
na
me
Pri
mer
sec
uen
ce (
5' - 3')
Len
gth
(b
p)
Ref
eren
ce
Su
b u
nit
B d
ari D
NA
gy
rase
A2
TC
TA
AC
TAT
CC
AC
CG
CG
G
Sub u
nit
B f
rom
gyra
se D
NA
B3
AG
CAA
TG
CC
ATC
TT
CA
CG
TT
C
tox
RF1
GA
AG
CA
GC
AC
TC
AC
CG
AT
tox
RR
1G
GT
GAA
GA
CT
CAT
CA
GC
A
VH
F1A
TC
AT
GA
AT
AA
AA
CT
ATT
AC
GT
TA
CT
VH
R1
GA
AA
GG
AT
GG
TT
TG
AC
AA
T
Vh
hem
oR
GC
TTG
AT
AAC
AC
TT
TG
CG
GT
308
Co
neje
ro &
He
dre
yd
a (2
004
)
gyrB
363
Th
aith
on
gn
um
et
al.
(20
06)
Re
gu
lato
r u
ntu
k tr
ansk
rip
tor
tran
smem
bra
ne (R
egul
ato
r fo
r tr
an
scri
pto
r tr
an
smem
bra
ne
)
vvh
Pro
tein
haem
oly
sin
Ha
emol
ysi
n p
rote
in1,2
57
Zh
ang
et
al.
(20
01
)
tox R
382
Pan
g e
t a
l. (2
006
)
134
J. Ris. Akuakultur Vol. 8 No. 1 Tahun 2013: 131-143

Setelah larutan dihomogenkan selanjutnyadimasukkan template DNA sebanyak 3 μL. Kon-disi proses amplifikasi untuk primer spesifiktoxR diatur sebanyak 30 siklus pada suhudenaturasi 94oC selama satu menit, annealing57oC selama satu menit dan elongasi 72oCselama satu menit serta tahap ekstra elongasi72oC selama sepuluh menit pada mesin PCR(Applied Biosystems 2720 Thermal Cycler).Proses amplifikasi DNA untuk primer spesifikvvh gen (haemolysin) diatur sebanyak 30 sikluspada suhu denaturasi 94oC selama satu menit,annealing 53oC selama satu menit danelongasi 72oC selama satu menit (Conejero &Hedreyda, 2004). Sedangkan untuk amplifikasigen gyrB proses PCR juga diatur sebanyak 30siklus dengan suhu denaturasi 94oC selamasatu menit, annealing 60oC selama satu menitdan elongasi 72oC selama dua menit sertatahap ekstra elongasi 72oC selama tujuh menit(Thaithongnum et al., 2006).
Elektroforesis
Hasil PCR kemudian diaplikasikan padagel agarosa 2% untuk diobservasi dan di-dokumentasikan. Elektroforesis minigel hanyaoptimum untuk pemisahan DNA berukurankecil (ratusan bp hingga sekitar 10 kb). Proseselektroforesis menggunakan minigel yangdialiri listrik dengan voltase 150 V dan kuatampere 70 waktu running selama 30 menit.Larutan yang digunakan adalah buffer elek-troforesis yang diperlukan untuk menciptakankondisi stabil selama proses berlangsung. Padaumumnya buffer yang digunakan berupa TrisAceticacid EDTA (TAE) atau Tris Boric EDTA (TBE).Untuk pewarna gel digunakan red gel sebagaipengganti etidhium bromide yang sudah tidakdianjurkan lagi karena bersifat karsinogenik.
Desain Primer Spesifik
Gen-gen spesifik yang sudah berhasildiamplifikasi dengan primer terpublikasikemudian dianalisis urutan basa DNA-nyadengan metode sekuensing. Hasil sekuensingini menjadi rujukan untuk mendesain primerspesifik. Setelah hasil sekuensing sudah di-peroleh selanjutnya adalah mengumpulkansekuen gen target sebagai referensi. Sekuenreferensi dapat diperoleh dari databaseGenBank di situs NCBI (http://www.ncbi.nlm.nih.gov/). Sekuen referensi akan digunakandalam uji BLAST (http://blast.ncbi.nlm. nih.gov/Blast) untuk mengetahui kemiripansekuen gen yang dimiliki dengan sekuen dari
gen sejenis yang sudah terdeposit pada NCBI.Urutan basa dari gen target dalam formatnotepad dapat langsung diunduh setelahlaman NCBI/BLAST/blastn suite terbuka.Tahapan selanjutnya adalah menentukanperangkat lunak yang akan digunakan untukdesain primer. Pada dasarnya sembarangdaerah tertentu pada sekuen referensi dapatdiambil untuk dijadikan primer, tanpa perlubantuan perangkat lunak khusus. Namun padapenelitian ini digunakan bantuan perangkatlunak Primer3 Plus (http://frodo.wi.mit.edu/cgi-bin/primer3). Setelah software sudahberhasil diunduh, maka proses desain primersudah dapat dimulai.
HASIL DAN BAHASAN
Hasil Deteksi Gen-Gen Spesifik padaVibrio Berpendar
Dari 35 isolat yang diisolasi dari berbagaidaerah, 22 isolat terdeteksi memiliki genspesifik, di mana sembilan di antaranya terde-teksi memiliki dua gen spesifik (Tabel 2). Isolatbakteri yang memiliki gen spesifik ini kemudiandiuji virulensinya dengan uji patogenisitassecara invivo menggunakan hewan uji udangwindu. Isolat kode 1, 120, 170, dan 275 ter-bukti memiliki tingkat patogenisitas lebihtinggi dibandingkan isolat bakteri vibrioberpendar lainnya (Tabel 2). Isolat yangmemiliki tingkat patogenisitas tinggi kemudiandipilih untuk dikarakterisasi gen spesifikhaemolysin dan gyrase-nya dengan prosessekuensing. Untuk gen spesifik ToxR tidakdapat disekuensing disebabkan hasil PCRyang divisualisasikan pada gel elektroforesismenunjukkan adanya penempelan yang tidakspesifik dari primer (unspecific annealing)sehingga tidak terbentuk pita tunggal DNA.
Haemolysin adalah eksotoksin yang ber-tanggung jawab dalam proses penyerapanmembran eritrosit atau proses hemolisis seldarah. Gen yang mengkode haemolysin inidilaporkan ditemukan pada beberapa spesiesbakteri yang termasuk genus vibrio (Conejero& Hedreyda, 2004). Bakteri vibrio patogenyang memiliki gen haemolysin diketahui dapatmenyebabkan terjadinya lysis pada sel darahinang.
Enzim gyrase terdistribusi hampir padasemua spesies dalam genus vibrio. Enzim iniyang mengurangi tekanan saat double-stranded DNA sedang tidak terikat oleh ikatanhelikase hal ini menyebabkan terjadinya
Desain primer spesifik untuk deteksi dini penyakit ..... (Ince Ayu Khairana Kadriah)
135

superkoiling DNA. Banyak antibiotik bekerjadengan menyerang gyrase. Gyrase DNAbakteri hadir di prokariota dan beberapaeukariota, tetapi enzim ini tidak sepenuhnyamirip dalam struktur atau urutan, dan memi-liki kedekatan yang berbeda untuk setiapmolekul yang berbeda. Enzim ini tidak dite-mukan pada manusia. Hal ini membuat gyrasesebagai target yang baik untuk antibiotik.Bakteri patogen memiliki struktur gen Gyr-Byang spesifik dibandingkan bakteri lainnya(Thaithongnum et al., 2006).
Hasil Sekuensing Gen-Gen SpesifikVibrio Berpendar
Hasil sekuensing gen haemolysin daribakteri hasil koleksi menunjukkan kemiripan94% dengan gen haemolysin bakteri V. harveyiyang terdeposit pada NCBI (Gambar 1).
Sedangkan hasil sekuensing gen gyrasememiliki kemiripan 99% dengan gen gyrasebakteri V. harveyi (Gambar 3). Kemiripan gentarget yang diisolasi dibandingkan denganbeberapa sekuen referensi menentukanberhasil tidaknya isolasi gen spesifik. Jumlahsekuen referensi yang diperlukan tergantungdari sampel target. Semakin banyak referensisekuen gen yang kita peroleh akan lebih baikagar kita dapat mendesain primer di daerahyang benar-benar sama (conserved region)setelah sekuen tersebut disejajarkan (align-ment). Pada dasarnya satu data sekuen sudahdapat dijadikan referensi dengan syarat bahwasampel target nantinya memiliki kesamaanspesies atau dengan kata lain secara genetiksangat mirip. Hasil sekuensing selanjutnyadigunakan untuk mendesain primer spesifikbakteri Vibrio patogen.
136
J. Ris. Akuakultur Vol. 8 No. 1 Tahun 2013: 131-143
Tabel 2. Hasil deteksi gen-gen spesifik bakteri vibrio berpendar
Table 2. The results of the detection of specific genes luminous Vibrio
tox-R haemo gyr-B
1 Banyuwangi
2 Bali
3 Bali
5582 Maros
5584 Maros
5585 Maros
109 Maros
120 Maros
128 Maros
133 Maros
137 Maros
139 Maros
154 Maros
159 Maros
671 Maros
672 Maros
673 Maros
676 Maros
168 Takalar
170 Takalar
275 Pinrang
276 Pinrang
Kode isolat uji Code of isolates
Asal Origin
Gen-gen spesifik (Specific genes )

Sekuen gen haemolysin
>1st_BASE_399953_1hem_VhF1
GTAGTGCTATACTACATTAATCTTGCTCCGCAGCCGACTCATCAGAGCCTTCTTACCTGCTAAATGCCTCAGAAGTGAGAAGCGCACAACAAAAGCAAACATACACCTACGTACGATGCTGGTATCGAACTAGTTATTCACATGATGACCCAGAAACCGACTGGGAGTGGGCAGAAAATCCAGATGGCAGTTATTTCACTATCGAAGGCTATTGGTGGAACGCACTCTCGTTTAAAAACATGTTCTATACCAATACACCGCAAAGTGTTATCAAGCAAAANCCCATTNAG
Gambar 1. Hasil sekuensing dan alignment gen haemolysin
Figure 1. Result of sequencing and alignment of haemolysin gene
Desain primer spesifik untuk deteksi dini penyakit ..... (Ince Ayu Khairana Kadriah)
137
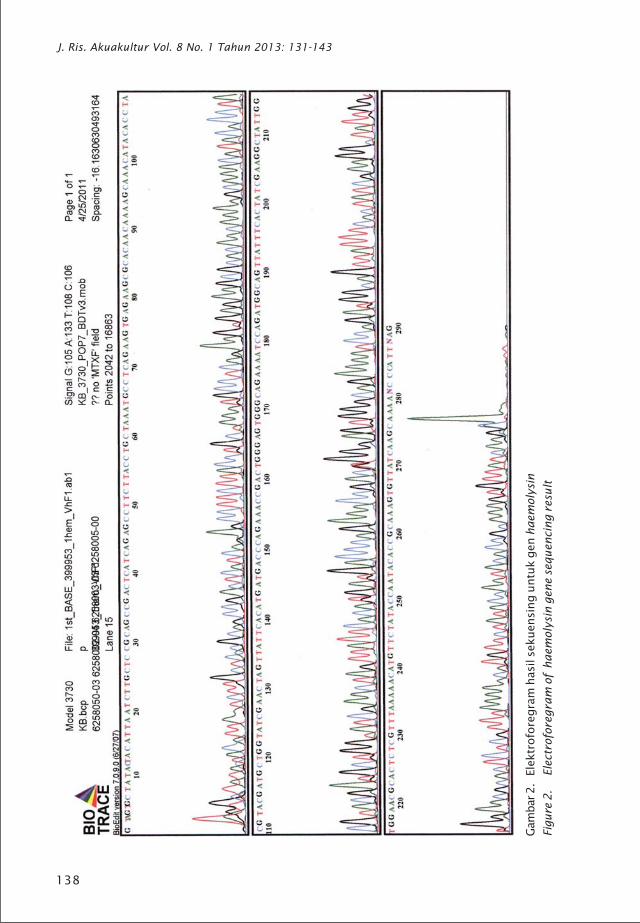
Gam
bar 2
.El
ektr
ofo
regra
m h
asil
seku
ensi
ng u
ntu
k gen
haem
olys
in
Figu
re 2
.El
ectr
ofor
egra
m o
f h
aem
olys
in g
ene
sequen
cing r
esult
138
J. Ris. Akuakultur Vol. 8 No. 1 Tahun 2013: 131-143
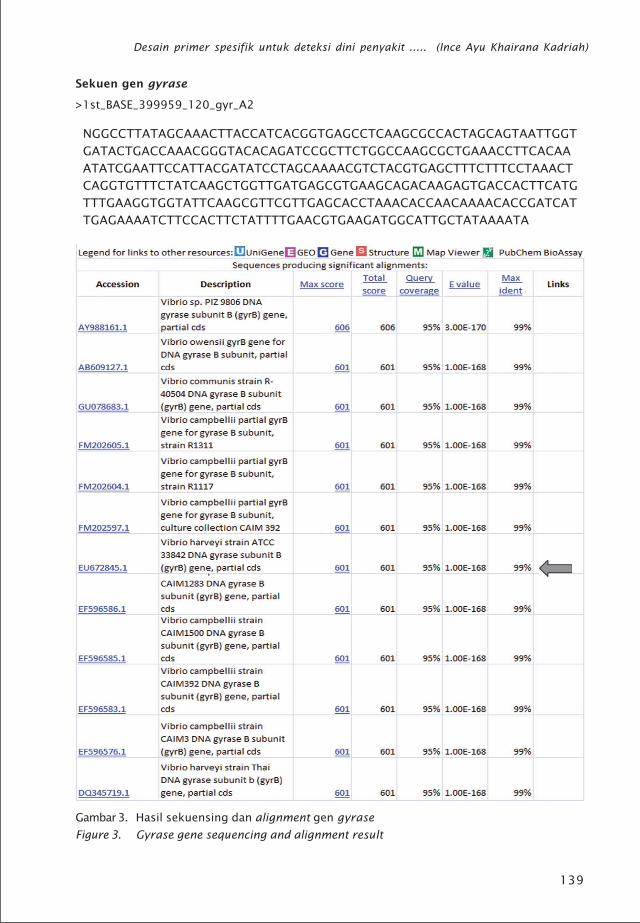
Gambar 3. Hasil sekuensing dan alignment gen gyrase
Figure 3. Gyrase gene sequencing and alignment result
Sekuen gen gyrase
>1st_BASE_399959_120_gyr_A2
NGGCCTTATAGCAAACTTACCATCACGGTGAGCCTCAAGCGCCACTAGCAGTAATTGGTGATACTGACCAAACGGGTACACAGATCCGCTTCTGGCCAAGCGCTGAAACCTTCACAAATATCGAATTCCATTACGATATCCTAGCAAAACGTCTACGTGAGCTTTCTTTCCTAAACTCAGGTGTTTCTATCAAGCTGGTTGATGAGCGTGAAGCAGACAAGAGTGACCACTTCATGTTTGAAGGTGGTATTCAAGCGTTCGTTGAGCACCTAAACACCAACAAAACACCGATCATTGAGAAAATCTTCCACTTCTATTTTGAACGTGAAGATGGCATTGCTATAAAATA
Desain primer spesifik untuk deteksi dini penyakit ..... (Ince Ayu Khairana Kadriah)
139

Gam
bar 4
.El
ektr
ofo
regra
m h
asil
seku
ensi
ng u
ntu
k gen
gyr
ase
Figu
re 4
.El
ectr
ofor
egra
m o
f gyr
ase
gen
e se
quen
cing r
esult
140
J. Ris. Akuakultur Vol. 8 No. 1 Tahun 2013: 131-143
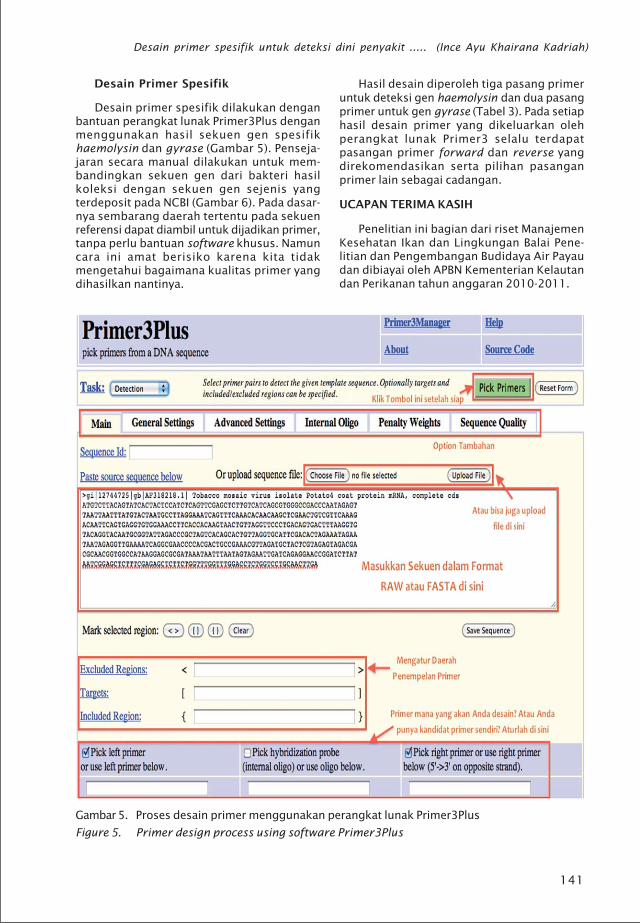
Desain Primer Spesifik
Desain primer spesifik dilakukan denganbantuan perangkat lunak Primer3Plus denganmenggunakan hasil sekuen gen spesifikhaemolysin dan gyrase (Gambar 5). Penseja-jaran secara manual dilakukan untuk mem-bandingkan sekuen gen dari bakteri hasilkoleksi dengan sekuen gen sejenis yangterdeposit pada NCBI (Gambar 6). Pada dasar-nya sembarang daerah tertentu pada sekuenreferensi dapat diambil untuk dijadikan primer,tanpa perlu bantuan software khusus. Namuncara ini amat berisiko karena kita tidakmengetahui bagaimana kualitas primer yangdihasilkan nantinya.
Hasil desain diperoleh tiga pasang primeruntuk deteksi gen haemolysin dan dua pasangprimer untuk gen gyrase (Tabel 3). Pada setiaphasil desain primer yang dikeluarkan olehperangkat lunak Primer3 selalu terdapatpasangan primer forward dan reverse yangdirekomendasikan serta pilihan pasanganprimer lain sebagai cadangan.
UCAPAN TERIMA KASIH
Penelitian ini bagian dari riset ManajemenKesehatan Ikan dan Lingkungan Balai Pene-litian dan Pengembangan Budidaya Air Payaudan dibiayai oleh APBN Kementerian Kelautandan Perikanan tahun anggaran 2010-2011.
Gambar 5. Proses desain primer menggunakan perangkat lunak Primer3Plus
Figure 5. Primer design process using software Primer3Plus
Desain primer spesifik untuk deteksi dini penyakit ..... (Ince Ayu Khairana Kadriah)
141

DAFTAR ACUAN
Austin, B. & Zhang, X.-H. 2006. Vibrio harveyi: asignificant pathogen of marine vertebratesand invertebrates. Lett. Appl. Microbiol., 43:119-124.
Baticados, M.C.L., Lavilla-Pitogo, C.R., Cruz-Lacierda, E.R., de la Pena, L.D., & Sunaz,N.A. 1990. Studies on the chemical con-trol of luminous bacteria Vibrio harveyi
and V. splendidus isolated from diseasedPenaeus monodon larvae and rearing wa-ter. Diseases of Aquatic Organism., 9: 133-139.
Bej, A.K., Patterson, D.P., Brasher, C.W., Vickery,M.C., Jones, D.D., & Kaysner, C.A. 1999.Detection of total and haemolysin-produc-ing Vibrio parahaemolyticus in shellfishusing multiplex PCR amplification of tl, tdhand trh. J. Microbiol. Methods, 36: 215-225.
Tabel 3. Hasil desain primer spesifik
Table 3. The result of specific primer design
Nama oligo Sekuen (5’ - 3’ ) PanjangOligo name Secuence (5’ - 3’ ) Length (bp)
IAVhF1 CAAAAGTGAAAAGCGCACAA 20
IAVhF2 TGAAAAGCGCACAACAAAAG 20
IAVhF3 GAAGTGAGAAGCGCACAACA 20
IAVhR1 TTCCACCAATAGCCTTCGAT 20
IAVhR2 TTAAACGAGAGTGCGTTCCA 20
IAVhR3 AACACTTTGCGGTGTATTGG 20
IAGyrF1 TGACCAAACGGGTACACAGA 20
IAGyrF2 CCAAACGGGTACACAGATCC 20
IAGyrR1 CAACGAACGCTTGAATACCA 20
IAGyrR2 CGAACGCTTGAATACCACCT 20
Gambar 6. Proses pensejajaran hasil sekuen secara manual menggunakan perangkat lunak Bioedit
Figure 6. The process sequence alignment results manually using software Bioedit
142
J. Ris. Akuakultur Vol. 8 No. 1 Tahun 2013: 131-143

Ben_Haim, Y., Thompson, F. L., Thompson, C.C., Cnockaert, M.C., Hoste, B., Swings, J., &Rosenberg, E. 2003. Vibrio coralliilyticus sp.nov., a temperature-dependent pathogenof the coral Pocillopora damicornis. Int. J.Syst. Evol. Microbiol., 53: 309-315.
Benson, H.J. 1985. Microbiological Applica-tions: A. Laboratory Manual in GeneralMicrobiology. Fourth Edition. Wm. C. BrownPublishers. Dubuque, Iowa, 450 pp.
Cano-Gomez, A., Bourne, D.G., Hall, M.R., Owens,L. & Hoj, L. 2009. Molecular identification,typing and tracking of Vibrio harveyi inaquaculture systems: Current methods andfuture prospects. Aquaculture, 287: 1-10.
Conejero, M.J.U. & Hedreyda, C.T. 2004. PCRdetection of haemolysin (vhh) gene inVibrio harveyi. J. Gen. Appl. Microbiol., 50:137-142.
Cunningham, C.O. 2002. Molecular diagnosisof fish and shellfish diseases: present sta-tus and potential use in disease control.Aquaculture, 206: 19-55.
Defoirdt, T. 2007. Quorum sensing disruptionand the use of short-chain fatty acids andpolyhydroxyalkanoates to control lumines-cent Vibriosis. PhD thesis, Ghent Univer-sity, Belgium, 228 pp.
Karunasagar, I., Pai, R., Malathi, G.R., &Karunasagar, I. 1994. Mass mortality ofPenaeus monodon larvae due to antibiotic-resistant Vibrio harveyi infection. Aquacul-ture, 128: 203-209.
Moriarty, D.J.W. 1998. Control of luminousVibrio species in penaeid aquacultureponds. Aquaculture, 168: 351-358.
Nishibuchi, M., Khaeomanee-iam, V., Honda, T.,Kaper, J.B., & Miwatani, T. 1990. Compara-
tive analysis of the haemolysin genes ofVibrio cholerae non-O1, Vibrio mimicus, andVibrio hollisae that are similar to the tdhgene of Vibrio parahaemolyticus. FEMSMicrobiol. Lett., 67: 251-256.
Nishibuchi, M. & Kaper, J. 1995. Thermostabledirect haemolysin gene of Vibrio para-haemolyticus: A virulence gene acquiredby a marine bacterium. Infect. Immun., 63:2,093-2,099.
Pang, L., Zhang, X.H., Zhong, Y., Chen, J., Li, Y.,& Austin, B. 2006. Identification of Vibrioharveyi using PCR amplification of the toxRgene. Lett. Appl. Microbiol., 43: 249-255.
Parenrengi, A. 2000. Studies on genetic vari-ability of groupers (Genus: Epinephelus)from Indo-Malaysian waters using PCR-RAPD Analysis. Thesis master of Science,Kolej University Terengganu, UniversitiPutra Malaysia, 174 pp.
Saulnier, D., Haffner, P., Goarant, C., Levy, P., &Ansquer, D. 2000. Experimental infectionmodels for shrimp Vibriosis studies: a re-view. Aquaculture, 191: 133-144.
Thaithongnum, S., Ratanama, R., Weeradechapol,K., Sukhoom, A., & Vuddhakul, V. 2006. De-tection of Vibrio harveyi in shrimp post-lavae and hatchery tank water by the mostprobable number technique with PCR.Aquaculture, 261: 1-9.
Zhang, X.H. & Austin, B. 2000. Pathogenicity ofVibrio harveyi to salmonids. J. Fish Dis., 23:93-102.
Zhang, X.H., Meaden, P.G., & Austin, B. 2001.Duplication of haemolysin genes in a viru-lent isolate of Vibrio harveyi. Appl. Environ.Microbiol., 67: 3,161-3,167.
Desain primer spesifik untuk deteksi dini penyakit ..... (Ince Ayu Khairana Kadriah)
143