potensi jamur pelapuk kayu isolat lokal makassar …
TRANSCRIPT

i
POTENSI JAMUR PELAPUK KAYU ISOLAT LOKAL MAKASSAR
DALAM MENDEKOMPOSISI KOMPONEN LIGNOSELULOSA
JERAMI PADI Oryza sativa L.
ERVIANI LESTARI
H411 09 271
JURUSAN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
UNIVERSITAS HASANUDDIN
MAKASSAR
2013

ii
POTENSI JAMUR PELAPUK KAYU ISOLAT LOKAL MAKASSAR
DALAM MENDEKOMPOSISI KOMPONEN LIGNOSELULOSA
JERAMI PADI Oryza sativa L.
Oleh :
ERVIANI LESTARI
H411 09 271
Skripsi ini dibuat untuk Melengkapi Tugas Akhir dan memenuhi Syarat untuk
Memperoleh Gelar Sarjana Sains Pada
Jurusan Biologi
JURUSAN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
UNIVERSITAS HASANUDDIN
MAKASSAR
2013

iii
LEMBAR PENGESAHAN
POTENSI JAMUR PELAPUK KAYU ISOLAT LOKAL MAKASSAR
DALAM MENDEKOMPOSISI KOMPONEN LIGNOSELULOSA
JERAMI PADI Oryza sativa L.
Disetujui Oleh :
Pembimbing Utama Pembimbing Pertama
Dr. Nur Haedar, S.Si, M.Si Prof. Dr. Ir. Tutik Kuswinanti, M.Sc NIP. 196801291997022001 NIP. 196503161989032002

iv
KATA PENGANTAR
Syukur dan pujian kehadirat Tuhan Yesus Kristus atas kebaikan, kekuatan
dan limpahan berkat serta kasih karuniaNya yang tak berkesudahan sehingga
skripsi dengan judul “Potensi Jamur Pelapuk Isolat Lokal Makassar Dalam
Mendekomposisi Komponen Lignoselulosa Jerami Padi Oryza sativa L” dapat
diselesaikan dengan baik. Skripsi ini disusun sebagai salah satu syarat untuk
memperoleh gelar sarjana sains (S.Si) pada Jurusan Biologi, Fakultas Matematika
dan Ilmu Pengetahuan Alam Universitas Hasanuddin.
Banyak kendala dan tantangan yang penulis hadapi selama proses
penyelesaian skripsi ini. Namun, berkat ketabahan, kesabaran, dan dukungan dari
berbagai pihak, penulis dapat menyelesaikan skripsi ini. Oleh karena itu, dengan
kerendahan hati dan rasa bangga, penulis mengucapkan terima kasih yang tak
terhingga kepada :
1. Ibu Dr. Nur Haedar, M.Si selaku pembimbing utama dan Ibu Prof. Dr. Ir.
Tutik Kuswinanti, M.Sc selaku pembimbing pertama yang telah dengan sabar
meluangkan waktu memberikan bimbingan, arahan dan petunjuk sehingga
skripsi ini dapat terselesaikan dengan baik.
2. Ibu Prof. Dr. Hj. Dirayah R. Husain, DEA, Bapak Drs. Ambeng, M.Si, Ibu
Dr. Magdalena Litaay, M.Sc, Ibu Dr. Rosana Agus, M.Si, dan Ibu Dr.
Juhriah, M.Si selaku tim penguji skripsi yang telah memberikan masukan dan
saran demi penyempurnaan penulisan skripsi ini.

v
3. Bapak Drs. Ambeng, M.Si selaku Penasehat Akademik atas motivasi,
bimbingan dan arahan kepada penulis selama masa perkuliahan.
4. Bapak Prof. Dr. H. Abd. Wahid Wahab. M.Sc Selaku Dekan Fakultas
Matematika dan Ilmu Pengetahuan Alam (FMIPA) Universitas Hasanuddin,
beserta seluruh staf atas bantuannya selama penulis menempuh pendidikan di
fakultas ini.
5. Bapak Dr. Eddy Soekandarsih, M.Sc selaku Ketua Jurusan Biologi Fakultas
Matematika dan Ilmu Pengetahuan Alam (FMIPA) Universitas Hasanuddin
dan para dosen yang telah memberikan ilmu pengetahuan yang sangat
berharga selama penulis menmpuh pendidikan serta pegawai yang telah
membantu dalam pengurusan administrasi penyusunan skripsi ini
6. Om Marten dan Tante Satriana sekeluarga, semua keluarga yang ada di
Toraja dan Manggarai terima kasih banyak buat dukungan dan bantuannya.
7. K’ Anto dan K’ Ahmad sebagai laboran PKP yang telah membantu selama
penelitian berlangsung.
8. Partner penelitianku Welsiliana, Nur Afni dan Yunianti Timang, terima kasih
sudah membuatku untuk lebih banyak bersabar. Saya bersyukur untuk semua
hal baik yang menyenangkan ataupun yang tidak selama penelitian dan
semuanya itu telah dilewati bersama.
9. Teman-teman seperjuangan MIPA 2009, khususnya teman-teman Bi09enesis,
yang senantiasa memberi semangat dan dukungan dalam penulisan skripsi ini.
10. Adikku Enrico dan Sary, terima kasih untuk doa, bantuan dan pengertiannya.

vi
11. Sahabat-sahabatku terkasih Roswita, Irene, Jeane, dan Ribka. Terima kasih
atas dukungannya.
12. Semua pihak yang tidak sempat disebutkan satu persatu.
Teristimewa skripsi ini saya persembahkan kepada kedua orang tua saya,
Ayahanda Petrus L (alm) dan Ibunda Selfia S. Payung Langi, serta adik-adikku
tersayang.
Penulis menyadari bahwa skripsi ini masih jauh dari kesempurnaan. Oleh
karena itu, penulis sangat mengharapkan kritik dan saran yang membangun dari
pembaca demi kesempurnaan skripsi ini.
Akhir kata, semoga skripsi ini bermanfaat bagi kita semua dan semoga
Tuhan Yang Maha Esa senantiasa menyertai kita dalam langkah hidup kita. Amin.
Makassar, Mei 2013
Penulis

vii
ABSTRAK
Jerami padi merupakan limbah pertanian yang keberadaanya sangat melimpah
pada saat panen, akan tetapi penggunaanya masih sangat terbatas dan proses
dekomposisi secara alamiah di alam berlangsung lama diakibatkan oleh
kandungan lignoselulosa terutama kandungan lignin. Proses dekomposisi dapat
dipercepat dan salah satunya adalah dengan menggunakan jamur pelapuk. Tujuan
dari penelitian ini adalah untuk mengetahui isolat jamur pelapuk yang
pertumbuhannya paling baik pada media jerami padi serta mengetahui
kemampuannya dalam mendekomposisi komponen lignin, hemiselulosa dan
selulosa jerami padi Oryza sativa L. Dalam penelitian ini digunakan 7 isolat
jamur pelapuk yaitu KSH, KSB, JM, MKS, B, C dan E. Ke tujuh isolat tersebut
hasil eksplorasi pada kayu lapuk di sekitar Makassar dan ditumbuhkan pada
baglog jerami padi selama 30 hari. Untuk mengetahui kemampuannya dalam
mendekomposisi komponen lignoselulosa jerami padi digunakan analisis Van
Soest (1976). Hasil penelitian menunjukkan bahwa isolat yang memiliki
kecepatan pertumbuhan terbaik adalah isolat KSH dan JM. Isolat yang paling
banyak mendekomposisi komponen lignin yaitu isolat JM sebesar 17, 18 %,
komponen hemiselulosa paling banyak didekomposisi oleh isolat KSH sebesar 61,
46 % dan isolat yang paling banyak dalam mendekomposisi komponen selulosa
adalah isolat E sebesar 41, 33 %.
Kata kunci : Jerami padi, dekomposisi, lignoselulosa, jamur pelapuk

viii
ABSTRACT
Rice straw is agricultural waste found in abundance during harvest season, but its use is still very limited. Its decomposition process also needs long time due to lignocellulose content, especially the lignin content. The decomposition process can be accelerated by, for instance, using rotting fungi. This study aims to find out : (1) rotting fungi isolates with the best growth on the media of rice straw ; and (2) to what extent the isolates are able to decompose lignin, hemicelluloses and straw cellulose of Oryza sativa L. The research used 7 isolates of rotting fungi including KSH, KSB, JM, MKS, B, C, and E. These isolates were obtained after exploring rotten woods arround Makassar. They were planted on baglogs of rice plant straw for 30 days. To find out whether the isolates were able to decompose lignocellulose of rice straw, Van Soest (1976) analysis was used. The result reveal that KSH and JM isolates have the best growth speed. JM isolate decomposes lignin component in the largest amount (17, 18%); KSH isolate decomposes hemicellulose component in the largest amount (61, 46%); and E isolate decomposes component in the largest amount (41, 33%). Keywords: Rice straw, decomposition, lignocellulose, rotting fungi

ix
DAFTAR ISI
HALAMAN SAMPUL ................................................................................ i
HALAMAN JUDUL ................................................................................... ii
HALAMAN PENGESAHAN ..................................................................... iii
KATA PENGANTAR ................................................................................. iv
ABSTRAK ................................................................................................... vii
ABSTRACT ................................................................................................ viii
DAFTAR ISI ............................................................................................... ix
DAFTAR TABEL ....................................................................................... xi
DAFTAR GAMBAR ................................................................................... xii
DAFTAR LAMPIRAN ............................................................................... xiii
BAB I PENDAHULUAN .......................................................................... 1
I.1 Latar Belakang ....................................................................... 1
I.2 Tujuan Penelitian ................................................................... 3
I.3 Manfaat Penelitian ................................................................. 4
I.4 Waktu dan Tempat Penelitian ................................................. 4
BAB II TINJAUAN PUSTAKA ................................................................ 5
II.1 Jerami Padi ............................................................................ 5
II.2 Komponen Penyusun Jerami Padi .......................................... 7
a. Lignin ............................................................................. 9
b. Hemiselulosa ................................................................... 10
c. Selulosa ........................................................................... 11
II.3 Dekomposisi Jerami Padi ....................................................... 12
II.4 Mikroorganisme Pendegradasi Lignin .................................... 12
a. Jamur Pelapuk Kayu ....................................................... 13
b. Jamur Pelapuk Putih (white rot fungi) ............................ 13
c. Jamur Pelapuk Cokelat (Brown rot fungi) ........................ 15
d. Soft rot Fungi .................................................................. 16
II.5 Degradasi Lignin, Hemiselulosa dan Selulosa ........................ 16

x
a. Degradasi Lignin ............................................................. 16
b. Degradasi Hemiselulosa ................................................. 17
c. Degradasi Selulosa .......................................................... 17
II.6. Analisis Kandungan Serat Kasar ............................................ 18
BAB III METODOLOGI PENELITIAN ................................................. 22
III.1 Alat ................................................................................... 22
III.2 Bahan ................................................................................ 22
III.3 Prosedur Kerja ................................................................... 22
III.3.1 Sterilisasi Alat ............................................................... 22
III.3.2 Pembuatan Medium ...................................................... 23
III.3.2.1 Medium Potato Dextrosa Agar (PDA) ........................... 23
III.3.2.2 Pembuatan Substrat Bahan Organik Sebagai Media
Tumbuh Isolat Jamur (baglog) ...................................... 24
III.3.3 Peremajaan Isolat .......................................................... 24
III.3.4 Seleksi Jamur Lignolitik ................................................ 25
III.3.4.1 Inokulasi Isolat Jamur Pada Substrat Bahan Organik .... 25
III.3.4.2 Analisa Lignin, Hemiselulosa dan Selulosa ................... 25
III.3.5 Analisis Data................................................................. 28
BAB IV HASIL DAN PEMBAHASAN .................................................... 29
IV.1 Pertumbuhan Isolat Jamur Pada Baglog Jerami Padi ............ 29
IV.2 Analisis Kandungan Lignin, Hemiselulosa dan Selulosa ...... 33
BAB V KESIMPULAN DAN SARAN ...................................................... 40
V.1 Kesimpulan .......................................................................... 40
V.2 Saran ................................................................................... 40
DAFTAR PUSTAKA .................................................................................. xiv
LAMPIRAN ................................................................................................ xix

xi
DAFTAR TABEL
Tabel 1 Perbedaan pertumbuhan miselia dan warna miselia dari ketujuh isolat setelah 3 hari sampai 30 hari inkubasi .................................. 31
Tabel 2 Persentase kadar NDF dan ADF pada jerami padi kontrol dan jerami padi setelah diinokulasi isolat jamur pelapuk ...................... 33
Tabel 3 Persentase kadar dan penurunan Lignin, Hemiselulosa, dan Selulosa setelah diinokulasi dengan 7 isolat jamur pelapuk dan pada kontrol jerami padi setelah 30 hari inkubasi. ......................... 34

xii
DAFTAR GAMBAR
Gambar 1 Jerami Padi ............................................................................... 5 Gambar 2 Konfigurasi Dinding Sel Tanaman ............................................ 8 Gambar 3 Lignoselulosa terdiri dari tiga komponen yaitu : selulosa,
hemiselulosa dan lignin yang memberikan struktur yang kuat pada dinding sel tanaman .......................................................... 19
Gambar 4 Pertumbuhan Isolat KSH (A), Isolat KSB (B), Isolat JM (C),
Isolat MKS (D), Isolat B (E) , Isolat C (F), Isolat E (G) dan (H) Kontrol pada baglog jerami padi 3 hari setelah inokulasi .... 29
Gambar 5 Pertumbuhan isolat KSH (A), Isolat KSB (B), Isolat JM (C),
Isolat MKS (D), Isolat B (E), Isolat C (F), Isolat E (F) dan
Kontrol (H) baglog jerami padi 30 hari setelah inokulasi .......... 30
Gambar 6 Grafik Persentase Penurunan Kandungan Lignin, Hemiselulosa dan Selulosa Pada Jerami Padi .................................................. 35

xiii
DAFTAR LAMPIRAN
Lampiran 1 Gambar koloni 7 isolat jamur pelapuk pada media PDA, 7 hari setelah inokulasi ............................................................. 46
Lampiran 2 Gambar Pengamatan ke-2 sampai ke-9 ................................... 47
Lampiran 3 Diagram alir analisis serat dengan metode Van Soest ............. 51

xiv
BAB I
PENDAHULUAN
I.1 Latar Belakang
Indonesia merupakan salah satu negara tropis terbesar di dunia. Keadaan
alam seperti ini menghasilkan iklim yang sangat mendukung bagi kelangsungan
hidup berbagai jenis hewan dan tumbuhan. Kondisi tersebut telah menjadikan
Indonesia sebagai negara agraris dan maritim yang subur. Indonesia sebagai
negara agraris yang memiliki potensi yang sangat besar dibidang pertanian.
Kebutuhan pangan dalam negeri sebagian besar dipenuhi sektor pertanian.
Sebagian masyarakat Indonesia masih menjadikan pertanian sebagai komunitas
utama sebagai usaha dan profesi. Produktivitas pertanian tanaman pangan di
Indonesia memiliki jumlah yang sangat besar setiap tahunnya. Meskipun
demikian, dalam setiap panen raya pertanian tanaman pangan di Indonesia selalu
membawa hasil samping atau limbah pertanian yang cukup besar pula hingga
mencapai jutaan ton. Limbah pertanian ini terdiri atas daun jagung, batang jagung,
daun kedelai, jerami padi, dan lain sebagainya (Jayanti dan Budiarti, 2010).
Jerami padi merupakan salah satu limbah pertanian yang sangat melimpah
pada saat panen dan belum sepenuhnya dimanfaatkan oleh masyarakat. Jerami
padi yaitu bagian dari batang padi tanpa akar yang tertinggal setelah diambil
butir buahnya (Komar, 1984). Menurut Hidanah (2007), pemanfaatan jerami padi
sebagai pakan ternak berkisar antara 31-39%, untuk industri 7 -16% dan sisanya
1

xv
36-62% dibiarkan sebagai limbah yang biasanya ditumpuk dan mengering lalu
kemudian di bakar.
Jerami padi seperti limbah pertanian lain pada umumnya, telah mengalami
lignifikasi lanjut yang menyebabkan terjadinya ikatan kompleks antara lignin,
selulosa dan hemiselulosa yang merupakan komponen utama dari dinding sel
tanaman (Eun et al, 2006). Berat kering dari jerami padi terdiri dari 26 %
hemiselulosa, 33 % selulosa dan 7 % lignin (Komar, 1984). Selulosa dan
hemiselulosa sebenarnya masih bisa dimanfaatkan oleh ternak, namun terselubung
oleh dinding keras yakni lignin sehingga pemanfaatan jerami padi sebagai pakan
ternak kurang optimal (Yunilas, 2009).
Jerami padi apabila dibiarkan begitu saja menumpuk akan mengganggu
manusia dan juga dapat menjadi inang bagi penyakit bagi manusia dan juga
tanaman itu sendiri, hal ini disebabkan karena proses dekomposisi limbah jerami
padi secara alamiah berlangsung sangat lama. Yang sering dilakukan petani untuk
menghilangkan tumpukan jerami padi adalah membiarkan jerami padi mengering
lalu kemudian di bakar. Akibat dari proses pembakaran jerami dapat mengganggu
aktivitas masyarakat karena asap tebal dan juga mencemari lingkungan karena
menghasilkan gas CO2 ke udara (Subiyatno, 2010).
Salah satu cara untuk mempercepat proses dekomposisi jerami padi yaitu
dengan memanfaatkan jasa mikroorganisme lignoselulolitik. Proses dekomposisi
jerami padi menggunakan mikroorganisme sangat menguntungkan, selain terjadi
konservasi hara juga mengurangi pencemaran lingkungan serta memberi nilai
tambah bagi petani. Kompos yang dikembalikan ke tanah akan melestarikan
2

xvi
kesuburan baik fisik, kimia, dan biologi tanah dengan demikian dapat mendukung
keberlanjutan produksi tanaman (Ekawati, 2003).
Komponen lignoselulosa dalam jerami padi dapat didegradasi oleh
beberapa jenis jamur. Banyak jenis jamur yang sudah diketahui mampu
mendegradasi komponen lignoselulosa dan umumnya merupakan jamur kelompok
Basidiomycetes yang paling efektif dalam perlakuan biologis pada bahan
berlignoselulosa (Sun dan Cheng, 2002; Zhang et al, 2007) .
Di alam terdapat tiga kelompok jamur yang dapat menguraikan komponen
kayu (lignoselulosa) yaitu jamur pelapuk coklat (brown rot), jamur pelapuk putih
(white rot) dan jamur pelapuk lunak (soft rot). Pengelompokan jamur pelapuk ini
didasarkan pada hasil proses pelapukan. Jamur pelapuk coklat menghasilkan sisa
hasil pelapukan berwarna coklat sedangkan jamur pelapuk putih menghasilkan
sisa hasil pelapukan yang berwarna putih (Fengel dan Wengener, 1995). Akan
tetapi banyak dari jamur ini selain mendegradasi lignin juga mendegradasi
selulosa dan hemiselulosa (Blanchete 1995). Jamur pelapuk putih banyak
dilaporkan memiliki kemampuan mendegradasi lignin yang tinggi dengan sedikit
kehilangan selulosa (Fengel dan Wengener, 1995).
Berdasarkan uraian di atas, maka dilakukanlah penelitian mengenai “Potensi
Jamur Pelapuk Kayu Isolat Lokal Makassar Dalam Mendekomposisi Komponen
Lignoselulosa Jerami Padi Oryza sativa L”
3

xvii
I.2 Tujuan Penelitian
Penelitian ini bertujuan untuk mengetahui isolat jamur pelapuk yang
paling cepat pertumbuhannya pada media jerami padi dan untuk mengetahui
kemampuannya dalam mendekomposisi komponen lignin, hemiselulosa dan
selulosa jerami padi Oryza sativa L.
I.3 Manfaat
Penelitian ini diharapkan dapat memberikan informasi awal tentang
kemampuan isolat jamur pelapuk lokal Makassar dalam mendekomposisi
komponen lignoselulosa (lignin, hemiselulosa dan selulosa) jerami padi Oryza
sativa L sehingga nantinya juga dapat digunakan dalam mendekomposisi
komponen lignoselulosa limbah pertanian lainnya .
I.4 Waktu dan Tempat
Penelitian ini dilaksanakan dari akhir bulan November sampai awal
Januari 2013, bertempat di Laboratorium Bioteknologi Pusat Kegiatan Penelitian
(PKP) dan Laboratorium Kimia dan Makanan, Fakultas Peternakan, Universitas
Hasanuddin, Makassar.
4

xviii
BAB II
TINJAUAN PUSTAKA
II.1 Jerami Padi
Padi Oryza sativa L merupakan salah satu tanaman pangan yang banyak
diusahakan oleh petani di Indonesia. Limbah panen dan olahan padi biasanya
berupa bekatul, sekam, jerami, dan merang. Jerami adalah sisa-sisa hijauan dari
tumbuhan sebangsa padi dan leguminosa setelah biji dan butir-butirnya dipetik
guna kepentingan manusia, sedangkan yang dimaksud dengan jerami padi
menurut Komar (1984) adalah bagian batang tumbuh yang telah dipanen bulir-
bulir buah bersama atau tidak dengan tangkainya dikurangi dengan akar dan
bagian batang yang tertinggal. Secara umum bagian utama jerami padi terdiri atas
helaian daun, pelepah daun, dan tangkai (Lubis,1992).
Gambar 1. Tumpukkan jerami padi (Sumber: http//epetani.deptan.go.id/)
5

xix
Sekitar 2,5 milyar residu pertanian diproduksi setiap tahun di seluruh
dunia (Grigoriou, 2000). Produksi beras Indonesia tahun 2009 sebesar 64,33 juta
ton (BPS, 2009). Perkiraan rasio beras-ke-jerami 1.0 (Yao et al., 2008), maka
produksi jeraminya sekitar 64 juta ton/tahun.
Di sejumlah besar daerah di Indonesia, jerami masih dianggap sebagai
sampah dan pada akhirnya hanya akan dibakar begitu saja tanpa ada pemanfaatan
lebih lanjut, padahal indonesia sebagai negara agraris merupakan penghasil
jerami yang sangat besar. Pemanfaatan jerami padi sebagai pakan ternak
berkisar antara 31-39%, untuk industri 7 -16% dan sisanya 36-62% dibiarkan
sebagai limbah yang biasanya ditumpuk dan mengering lalu kemudian di bakar
(Hidanah, 2007).
Berdasarkan kandungannya maka jerami padi termasuk dalam golongan
bahan yang kaya akan serat kasar, yaitu umumnya mencapai lebih dari 86% berat
kering (Lubis,1992). Akibat kandungan serat kasar yang tinggi kualitas nutrisi
menjadi rendah, hal tersebut merupakan faktor pembatas jerami padi
dimanfaatkan sebagai pakan ternak ruminansia, sehingga sehingga diberikan
perlakuan untuk menghilangkan atau memutuskan ikatan yang terjadi diantara
komponen serat. Jaringan-jaringan pada pakan yang berasal dari limbah telah
mengalami proses lignifikasi (pengerasan) sehingga terbentuk lignoselulosa dan
lingnohemiselulosa. Lignin dan selulosa sering membentuk senyawa
lignosellulosa dalam dinding sel tanaman dan merupakan suatu ikatan yang kuat
(Sutardi, 1980).
6

xx
Jerami padi merupakan limbah tanaman yang tua sehingga telah
mengalami lignifikasi bertaraf lajut yang menyebabkan terjadi ikatan yang erat
dan kompleks antara lignin, selulosa dan hemiselulosa. Kompleksitas kimia dan
struktural bahan ini akan mempersulit mikroorganisme dalam mendekomposisi
limbah jerami tersebut (Ekawati, 2003).
II.2 Komponen Penyusun Jerami Padi
Jerami padi mengandung 21% isi sel dan 79% dinding sel berdasarkan
berat kering. Dari 79% berat kering ini terdiri dari 26% hemiselulose, 33%
selulose, dan 7% lignin. kandungan dinding sel terutama lignin bertambah dengan
menigkatnya umur tanaman ( Komar, 1984 ).
Lignoselulosa merupakan komponen utama penyusun dinding sel pada
tanaman. Lignoselulosa atau biomassa kayu tersusun dari polimer karbohidrat
(selulosa dan hemiselulosa), lignin dan bagian yang lebih sedikit (ekstraktif, asam,
garam dan mineral) Semua komponen lignoselulosa terdapat pada dinding sel
tanaman. Susunan dinding sel tanaman terdiri dari lamela tengah (M), dinding
primer (P) serta dinding sekunder (S) yang terbentuk selama pertumbuhan dan
pendewasaan sel yang terdiri dari lamela transisi (S1), dinding sekunder utama
(S2) dan dinding sekunder bagian dalam (S3). Dinding primer mempunyai
ketebalam 0.1-0.2 μm dan mengandung jaringan mikrofibril selulosa yang
mengelilingi dinding sekunder yang relatif lebih tebal (Chahal dan Chahal 1998).
7

xxi
Gambar 2. Konfigurasi Dinding Sel Tanaman (Perez et al, 2002).
Selulosa pada setiap lapisan dinding sekunder terbentuk sebagai lembaran
tipis yang tersusun oleh rantai panjang residu β-D-glukopiranosa yang berikatan
melalui ikatan β-1,4 glukosida yang disebut serat dasar (elementary fiber).
Sejumlah serat dasar jika terjalin secara lateral akan membentuk mikrofibril.
Mikrofibril mempunyai struktur dan orientasi yang berbeda pada setiap lapisan
dinding sel. Lapisan dinding sekunder terluar (S1) mempunyai struktur serat
menyilang, lapisan S2 mempunyai mikrofibril yang paralel terhadap poros lumen
dan lapisan S3 mempunyai mikrofibril yang berbentuk heliks. Mikrofibril
dikelilingi oleh hemiselulosa dan lignin. Bagian antara dua dinding sel disebut
lamela tengan (M) dan diisi dengan hemiselulosa dan lignin. Hemiselulosa
dihubungkan oleh ikatan kovalen dengan lignin. Selulosa secara alami terproteksi
dari degradasi dengan adanya hemiselulosa dan lignin (Perez et al. 2002).
8

xxii
a. Lignin
Lignin adalah senyawa kompleks yang membentuk ikatan ether dengan
selulosa dan hemiselulosa, protein dan komponen lainnya dalam jaringan tanaman
membentuk bagian struktural dan sel tumbuhan (Young, 1986). Lebih dari 30
persen tanaman tersusun atas lignin yang memberikan bentuk yang kokoh dan
memberikan proteksi terhadap serangga dan patogen (Orth et al. 1993).
Lignin merupakan polimer dengan struktur aromatik yang terbentuk melalui
unit-unit penilpropan yaitu: alkohol kumaril, alkohol koniferil, dan alkohol
sinapil (Sjorberg 2003). Senyawa ini juga merupakan senyawa yang tidak
mudah larut dalam air (Srebotnik et al. 1998). Senyawa tersebut berhubungan
secara bersama oleh beberapa jenis ikatan yang berbeda (Perez et al. 2002).
Lignin sulit didegradasi karena strukturnya yang kompleks dan heterogen.
Fungsi lignin disamping memberikan bentuk yang kokoh terhadap tanaman, lignin
juga member kekakuan pada jaringan pengangkut tumbuhan serta membentuk
ikatan yang kuat dengan polisakarida (selulosa dan hemiselulosa) dan melindungi
polisakarida dari degradasi mikroba sehingga lignin bersifat rekalsitran karena
tahan terhadap degradasi atau tidak terdegradasi dengan cepat di lingkungan
(Hammel, 1997).
Lignin terutama terkonsentrasi pada lamela tengah dan lapisan S2 dinding
sel yang terbentuk selama proses lignifikasi jaringan tanaman (Chahal dan Chahal
1998; Steffen 2003). Lignin tidak hanya mengeraskan mikrofibril selulosa, juga
berikatan secara fisik dan kimia dengan hemiselulosa.
9

xxiii
Pembentukan lignin terjadi secara intensif setelah proses penebalan dinding
sel terhenti. Pembentukan dimulai dari dinding primer dan dilanjutkan ke dinding
sekunder. Faktor lignin dalam membatasi permeabilitas dinding sel tanaman dapat
dibedakan menjadi efek kimia dan efek fisik. Efek kimia, yaitu hubungan lignin-
karbohidrat dan asetilisasi hemiselulosa. Lignin secara fisik membungkus
mikrofibril dalam suatu matriks hidrofobik dan terikat secara kovalen dengan
hemiselulosa. Hubungan antara lignin karbohidrat tersebut berperan dalam
mencegah hidrolisis polimer selulosa (Chahal dan Chahal 1998).
b. Hemiselulosa
Hemiselulosa merupakan kelompok polisakarida heterogen dengan berat
molekul rendah. Hemiselulosa rantainya pendek dibandingkan selulosa dan
merupakan polimer campuran dari berbagai senyawa gula, seperti xilosa,
arabinosa, dan galaktosa. Jumlah hemiselulosa biasanya antara 15 dan 30 persen
dari berat kering bahan lignoselulosa (Taherzadeh 1999).
Dari penelitian yang dilakukan Morrison (1986) mendapatkan bahwa
hemiselulosa lebih erat terikat dengan lignin dibandingkan dengan selulosa,
sehingga selulosa lebih mudah dicerna dibandingkan dengan hemiselulosa.
Hemiselulosa mengikat lembaran serat selulosa membentuk mikrofibril yang
meningkatkan stabilitas dinding sel. Hemiselulosa juga berikatan silang dengan
lignin membentuk jaringan kompleks dan memberikan struktur yang kuat.
Hemiselulosa relatif lebih mudah dihidrolisis dengan asam menjadi monomer
yang mengandung glukosa, mannosa, galaktosa, xilosa dan arabinosa.
10

xxiv
c. Selulosa
Selulosa merupakan suatu polisakarida yang mempunyai formula umum
seperti pati (C6H10O5)n. Sebagain besar selulosa terdapat pada dinding sel dan
bagian-bagian berkayu dari tumbuhan-tumbuhan dan merupakan komponen utama
penyusun dinding sel tanaman. Kandungan selulosa pada dinding sel tanaman
tingkat tinggi sekitar 35-50% dari berat kering tanaman (Lynd et al. 2002).
Selulosa merupakan polimer glukosa dengan ikatan β-1,4 glukosida dalam
rantai lurus. Bangun dasar selulosa berupa suatu selobiosa yaitu dimer dari
glukosa. Rantai panjang selulosa terhubung secara bersama melalui ikatan
hidrogen dan gaya van der Waals (Perez et al. 2002).
Selulosa mengandung sekitar 50-90% bagian berkristal dan sisanya bagian
amorf. Ikatan β-1,4 glukosida pada serat selulosa dapat dipecah menjadi
monomer glukosa dengan cara hidrolisis asam atau enzimatis. Kesempurnaan
pemecahan selulosa pada saluran pencernaan ternak tergantung pada ketersediaan
enzim pemecah selulosa yaitu selulase. Saluran pencernaan manusia dan ternak
non ruminansia tidak mempunyai enzim yang mampu memecah ikatan β-1,4
glukosida sehingga tidak dapat memanfaatkan selulosa. Ternak ruminansia
dengan bantuan enzim yang dihasilkan mikroba rumen dapat memanfaatkan
selulosa dengan terlebih dahulu merombaknya menjadi monomer kemudian
digunakan sebagai sumber energi. Pencernaan selulosa dalam sel merupakan
proses yang kompleks yang meliputi penempelan sel mikroba pada selulosa,
hidrolisis selulosa dan fermentasi yang menghasilkan asam lemak terbang (Aziz
et al. 2002).
11

xxv
II.3 Dekomposisi Jerami Padi
Dekomposisi merupakan merupakan suatu proses penguraian
mikrobiologis alami dari bahan buangan organik. Metode ini mempunyai prinsip
dasar menurunkan atau mendegradasi bahan-bahan organik secara terkontrol
menjadi bahan-bahan anorganik dengan menggunakan aktifitas mikroorganisme.
Bahan – bahan organik seperti selulosa, hemiselulosa dan lignin didegradasi oleh
mikroorganisme baik berupa bakteri, jamur atau yang lainnya (Murbandono,
2006).
Lignin merupakan polimer alami dan tergolong ke dalam senyawa
rekalsitran karena tahan terhadap degradasi, atau tidak terdegradasi dengan cepat
di lingkungan, sehingga diperlukan mikroorganisme yang mampu mendegradasi
lignin secara efektif menjadi air (H2O) dan karbondioksida (CO2) (Nasrul dan
Maimun, 2009).
II.4 Mikroorganisme Pendegradasi Lignin
Degradasi lignin membutuhkan enzim ekstraseluler yang tidak spesifik
karena lignin mempunyai struktur yang kompleks dengan berat molekul yang
tinggi. Lignin biasanya terakumulasi selama proses degradasi lignoselulosa.
Lignin selain dapat didegradasi oleh sekelompok mikroorganisme, dalam kondisi
lingkungan tertentu dapat juga didegradasi oleh faktor abiotik seperti dengan
senyawa alkali atau radiasi ultraviolet (Vahatalo et al 1999), namun hanya kapang
pelapuk putih yang banyak dilaporkan mampu mendegradasi lignin secara efektif
(Blanchette, 1995). Degradasi lignin oleh bakteri seperti Streptomyces sp.
(Crawford et al 1983) dan Actinomycetes (Kirk dan Farrell, 1987) terjadi seperti
12

xxvi
oksidasi yang dilakukan oleh kapang pelapuk putih, namun bakteri hanya mampu
mendegradasi sebagian kecil molekul lignin. Spesies kapang yang mampu
mendegradasi lignin dapat dikelompokkan menjadi White rot fungi, Brown rot
fungi dan Soft rot fungi.
a. Jamur Pelapuk Kayu
Kapang/jamur pelapuk kayu yang digolongkan dalam kelas Basidiomycetes
merupakan mikroorganisme yang mampu mendegradasi lignin. Berkenaan
dengan proses perombakan kayu oleh jamur, Brauns (1952) mengemukakan
bahwa ada dua jenis jamur yang berperan aktif, yaitu jamur pelapuk coklat (brown
rot fungi) dan jamur pelapuk putih putih (white rot fungi).
Menurut (Fengel and Wegener 1995) kapang pelapuk kayu dibedakan atas
tiga berdasarkan mekanisme degradasinya yaitu kapang pelapuk putih (white rot
fungi), kapang pelapuk coklat (brown rot fungi), dan kapang pelapuk lunak (soft
rot fungi) yang masing-masing memiliki metabolisme degradatif yang berbeda.
Kapang pelapuk putih menyerang lignin maupun polisakarida. Kayu yang
terdegradasi menjadi putih dan lunak. Berbeda dengan kapang pelapuk putih,
kapang pelapuk coklat mendegradasi polisakarida kayu dan mendegradasi sedikit
lignin sehingga kayu menjadi coklat dan rapuh. Sedangkan kapang pelapuk lunak
lebih menyukai selulosa dan hemilselulosa sebagai substratnya.
b. Jamur Pelapuk Putih (white rot fungi)
Jamur pelapuk putih (white rot fungi) adalah kelompok jamur
Bassidiomycetes dan Ascomycetes yang mampu menguraikan lignin, selulosa dan
hemiselulosa. Substrat bagi pertumbuhan jamur pelapuk putih ini adalah selulosa
13

xxvii
dan hemiselulosa dan degradasi lignin terjadi pada akhir pertumbuhan primer
melalui metabolisme sekunder dalam kondisi defisiensi nutrien seperti nitrogen,
karbon atau sulfur (Hatakka 2001).
Kapang ini ada yang mampu mendegradasi lignin secara selektif dan ada
pula yang non selektif. Kapang pelapuk putih selektif (contoh: Ceriporiopsis
subvermispora, Dichomitus squalens, Phanerochaete chrysosporium, Phlebia
radiata), lignin dan hemiselulosa didegradasi lebih banyak dibanding selulosa,
sedangkan kapang non selektif (contoh: Trametes versicolor and Fomes
fomentarius), mendegradasi semua komponen lignoselulosa dalam jumlah yang
sama (Blanchette 1995; Hatakka 2001).
Jamur white rot selain menguraikan lignin diduga juga dapat menguraikan
senyawa polutan lain (Hammel, 1997). Jamur ini menghasilkan enzim
ekstraseluler sehingga tahan terhadap bahan beracun atau bahan kimia
mutagenik. Beberapa jamur white rot telah digunakan dalam penguraian
lignin, misalnya Bjerkandera adusta mampu mendegradasi lignin 40% dan
pengurangan warna lignin sekitar 70% pada inkubasi selama 40 jam. Jamur
pelapuk putih (white rot) menguraikan lignin melalui proses oksidasi
menggunakan enzim phenol oksidase menjadi senyawa yang lebih sederhana
sehingga dapat diserap oleh mikroorganisme (Sanchez, 2009).
Kapang pelapuk putih menggunakan selulosa sebagai sumber karbon.
Kapang mendegradasi lignin secara keseluruhan menjadi karbon dioksida
untuk masuk ke polisakarida kayu yang dilindungi oleh lignin-karbohidrat
kompleks (Wilson dan Walter 2002).
14

xxviii
Pada proses degradasi lignin, kapang kapang pelapuk putih memproduksi
enzim oksidatif ekstraselular yang unik. Sistem enzim hasil sekresi
mikroorganisme inilah yang berfungsi sebagai agen biodegradasi yang mampu
memecah bahan berlignoselulosa menjadi molekul-molekul yang lebih sederhana.
Enzim ini juga sangat baik mendegradasi senyawa pestisida dan limbah beracun
(Srebotnik et at. 1998). Hal tersebut dikarenakan untuk mendepolimerisasi
dan memineralisasi lignin, kapang pelapuk putih memiliki system oksidatif non
spesifik meliputi beberapa ekstraselular oksidoreduktase, metabolit dengan bobot
molekul rendah, dan kerja oksigen yang sangat efektif (Saparrat et al 2002).
Selain itu kapang pelapuk putih memiliki kemampuan mendepolimerisasi lignin
dan memetabolisme lignin menjadi CO2 dan H20 (Kaal et al. 1995).
c. Jamur Pelapuk Cokelat (Brown rot fungi)
Brown-rot fungi terutama termasuk dalam kelas Basidiomycetes. Kapang
ini mendegradasi selulosa dan hemiselulosa sangat efeisien dan mendegradasi
sedikit lignin sehingga kayu menjadi coklat dengan mekanisme yang berbeda dari
organisme lain yang melibatkan reaksi non enzimatik dan tanpa enzim
eksoglukonase (Blanchette 1995).
Keberadaan lignin memacu degradasi selulosa oleh brown-rot fungi
meskipun lignin didegradasi dalam tingkat yang lebih kecil terutama pada lamela
tengah dinding sel yang kaya lignin (Blanchette 1995; Hatakka 2001). Kapang
Polyporus ostreiformis mampu menghasilkan enzim MnP and LiP, tetapi
kemampuannya dalam degradasi lignin lebih rendah dibanding P. chrysosporium
(Dey et al. 1994).
15

xxix
d. Soft rot Fungi
Soft-rot fungi terutama hanya terdapat pada daerah dengan lingkungan
yang ekstrim bagi kapang pelapuk dari kelas basiodiomycetes seperti lingkungan
yang terlalu basah atau terlalu kering (Blanchette 1995). Kapang ini juga
mempunyai tingkat toleransi yang lebih baik terhadap temperatur, pH dan
keterbatasan oksigen dibanding kapang pelapuk lain (Blanchette 1995, Daniel
dan Nilsson 1998).
Soft-rot fungi dapat berkembang pada tanah, kompos, kayu, hay, jerami
dan daerah perairan (Tuomela 2002). Penambahan nitrogen dalam substrat
mampu meningkatkan laju perombakan lignin, berlawanan dengan sifat kapang
pelapuk putih dan coklat (Daniel dan Nilsson 1998).
II.5 Degradasi Lignin, Hemiselulosa , dan Selulosa
a. Degradasi Lignin
Degradasi lignin tidak seperti selulosa dan hemiselulosa. Lignin pinsipnya
tidak berikatan linear tetapi merupakan senyawa kompleks. Polimer heterogen,
dengan senyawa aromatik non-stereoregular yang disusun oleh unit
fenilpropanoid (Boyle et al. 1992).
Ligninolitik berhubungan dengan produksi enzim ekstraseluler pendegradasi
lignin yang dihasilkan oleh jamur pelapuk putih. Dua enzim yang berperan
dalam proses tersebut adalah fenol oksidase (lakase) dan peroksidase (lignin
peroksidase (LiP) dan manganese peroksidase (MnP)) (Kirk and Farrell. 1987).
LiP merupakan katalis utama dalam proses ligninolisis oleh kapang karena
mampu memecah unit non fenolik yang menyusun sekitar 90 persen struktur
16

xxx
lignin (Srebotnik et al. 1994). MnP mengoksidasi Mn2+ menjadi Mn3+ yang
berperan sebagai dalam pemutusan unit fenolik lignin. LiP mengkatalis oksidasi
senyawa aromatik non fenolik. Jamur pelapuk putih adalah satu-satunya
organisme yang dikenal mampu mendegradasi lignin secara sempurna menjadi
menjadi produk yang larut dalam air dan CO2 (Boyle et al. 1992).
b. Degradasi Hemiselulosa
Degradasi hemiselulosa menjadi monomer gula dan asam asetat dengan
bantuan enzim hemiselulase. Hemiselulase seperti kebanyakan enzim lainnya
yang dapat menghidrolisis dinding sel tanaman merupakan protein multi-domain.
Xilan merupakan karbohidrat utama penyusun hemiselulosa dan Xylanase
merupakan hemiselulase utama yang menghidrolisis ikatan β-1,4 rantai xilan
menjadi oligosakarida (Perez et al. 2002).
c. Degradasi Selulosa
Degradasi selulosa oleh fungi merupakan hasil kerja sekelompok enzim
selulolitik yang bekerja secara sinergis (Howard et al. 2003). Sistem enzim
selulolitik terdiri dari tiga kelompok utama yaitu (Perez et al. 2002; Howard et al.
2003 dalam Suparjo, 2008):
Endoglucanases atau 1,4-β-D-glucan-4-glucanohydrolases yang berfungsi
menghidrolisis secara acak bagian amorf selulosa serat menghasilkan
oligosakarida dengan panjang yang berbeda dan terbentuknya unjung rantai
baru
Exoglucanases, yang meliputi 1,4-β-D-glucan glucanohydrolases atau
cellodextrinase dan 1,4-β-D-glucan cellobiohydrolases atau cellobio-
17

xxxi
hydrolases yang bekerja terhadap ujung pereduksi dan non pereduksi rantai
polisakarida selulosa dan membebaskan glukosa yang dilakukan oleh enzim
glucanohydrolases atau selobiosa yang dilakukan oleh enzim
cellobiohydrolases sebagai produk utama
β-glucosidases atau β-glucoside glucohydrolases, enzim ini tergantung pada
sumber karbon yang tersedia.
Hasil kerja sinergis endoglucanases dan exoglucanases menghasilkan
molekul selobiosa. Hidrolisis selulosa secara efektif memerlukan enzim (β-
glucosidases yang memecah selobiosa menjadi 2 molekul glukosa.
II.6. Analisis Kandungan Serat Kasar
Dalam pangan/pakan yang berasal dari tanaman terdapat serat makanan
yaitu bahan yang tahan terhadap penguraian oleh enzim dalam saluran pencernaan
dan karenanya tidak diabsorpsi. Serat makanan ini terdiri dari selulosa dan
senyawa lainnya dari polisakarida atau yang berkaitan dengan polisakarida seperti
lignin dan hemiselulosa (Gaman dan Sherrington, 1992).
Bagian terbesar komponen serat kasar yaitu polisakarida yang terdapat pada
dinding sel tanaman. Degradasi polisakarida bervariasi tergantung pada jaringan
tanaman, jenis tanaman dan umur tanaman (Hatfield, 1989). Chesson (1988)
melaporkan bahwa penyusun utama dinding sel tanaman lebih mungkin dapat
dicerna daripada bagian yang kedua yang lebih tebal dari dinding sel.
Karakteristik serat terutama struktur fisika dan kimia, mempunyai peranan
penting dalam mempengaruhi kecepatan dan tingkat degradasi serat kasar
tersebut. Adanya ikatan ester dan ikatan kovalen antara lignin, polisakarida dari
18
xxxii
protein serat kasar, secara alamiah membentuk ikatan intrinsik pada sebagian
besar struktur serat kasar, dan merupakan pembatas utama dalam degradasi, baik
degradasi selulosa maupun hemiselulosa (Chesson, 1988; Hatfield, 1989; dan
Jung,1989).
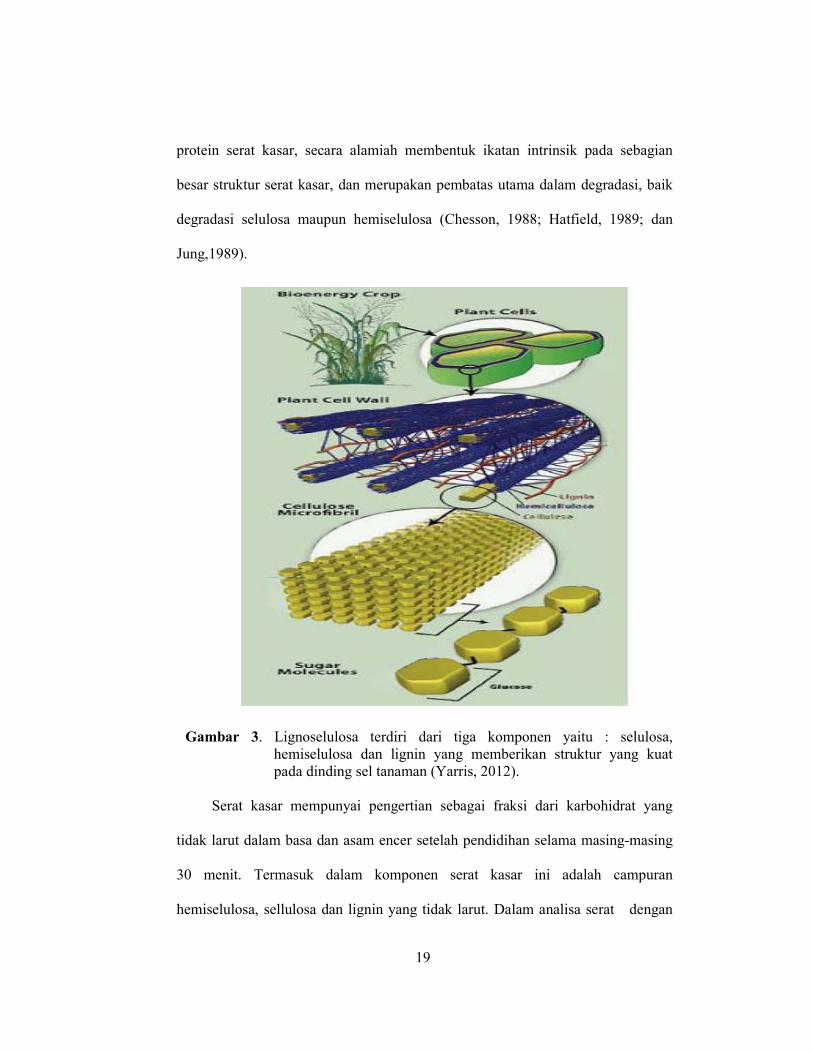
Gambar 3. Lignoselulosa terdiri dari tiga komponen yaitu : selulosa, hemiselulosa dan lignin yang memberikan struktur yang kuat pada dinding sel tanaman (Yarris, 2012).
Serat kasar mempunyai pengertian sebagai fraksi dari karbohidrat yang
tidak larut dalam basa dan asam encer setelah pendidihan selama masing-masing
30 menit. Termasuk dalam komponen serat kasar ini adalah campuran
hemiselulosa, sellulosa dan lignin yang tidak larut. Dalam analisa serat dengan
19

xxxiii
metode proksimat tidak dapat secara terpisah fraksi lignin, sellulosa dan
hemiselulosa yang justru perlu diketahui komposisinya untuk hijauan pakan
atau umumnya pakan berserat. Untuk memperoleh data yang lebih akurat tentang
fraksi lignin dan sellulosa dapat dilakukan analisa yang lebih spesifik dengan
metode Van Soest (Sutardi, 1980).
Pada metode Van Soest membagi pakan menjadi beberapa fraksi
berdasarkan kelarutannya dalam detergen. Mula-mula jerami dimasak dalam
larutan detergen netral (NDS) pada pH 6,9-7,0. Tiap liter larutan ini mengandung
30 grNa-Lauryl Sulfate, 15,61 gr Disodium-Dihidrogen-ethylendiamin-tetraacetat
dihidrate, 4, 56 gr disodium-hidrogen phosphate, 6,81 gr Na-Borat Decahidrat,
dan 10 ml Ethilen-Glikol-monomethyl eter. Larutan detergen memisahkan
komponen lignoselulosa dengan bahan penyusun dinding sel lainnya protein,
karbohidrat dan mineral-mineral mudah larut serta lemak. Komponen lain ini
bersifat mudah larut dalam larutan detergen netral, sedangkan komponen
lignoselulosa penyusun dinding sel tidak. Setelah dikeringkan, bobot dinding sel
ditimbang. Selanjutnya jerami padi diuji kelarutannya dalam detergent asam
(ADS), tiap liter larutan ini mengandung 49,04 gr H2SO4 , dan 20 gr Cetyl-
trimetil-amonium-Bromida (CTAB dan CETAB). Pemasakan dalam larutan
detergent asam ini akan membagi dinding sel menjadi fraksi yang larut dan tak
larut. Fraksi yang larut sebagian besar terdiri atas hemiselulosa dan sedikit protein
dinding sel. Fraksi yang tidak larut adalah lignoselulosa yang lazim disebut “Acid
Detergent Fiber” (ADF). Kemudian setelah penyaringan dan pengeringan, ADF
20

xxxiv
bahan makanan ditimbang dan ADF yang diperoleh dari analisis ini digunakan
pada tahap berikutnya untuk penentuan kadar lignin (Sutardi, 1980).
Untuk memisahkan selulosa dari lignin, ADF ditambahi H2SO4 dingin,
sehingga selulosanya akan larut. Selanjutnya residu yang tertinggal dicuci dengan
air hangat (85-95oC) sampai bebas dari asam. Lalu dikeringkan, kemudian
ditimbang lagi dan selisih bobot antara ADF dengan residu tersebut adalah
selulosa. Setelah residu ditimbang, lalu dibakar pada tanur dengan suhu 500oC.
Abu sisanya setelah dingin ditimbang dan selisih antara residu dengan abu adalah
lignin (Sutardi, 1980).
21

xxxv
BAB III
METODE PENELITIAN
III.1 Alat
Alat yang digunakan adalah gunting, timbangan, autoklaf, ember,
pengaduk, pisau, ose lurus, cawan petri, batang pengaduk, Laminary Air Flow
(LAF), bunsen, enkas, penangas, hand sprayer, pisau scalpel, jarum inokulum,
kaca masir (sintered glass), pompa vakum, oven, desikator, gelas piala dan
kamera.
III.2 Bahan
Bahan yang digunakan adalah jerami padi, isolat jamur pelapuk, kapur,
dedak halus, air, alumanium foil, tisu, kapas, plastik polipropilena (PP), alkohol
70%, spiritus, karet gelang, pipa paralon dengan panjang 3 cm diameter 3 cm,
cloramfenikol, plastik parafilm, aquades steril, PDA (Potato Dekstrosa agar), air
panas, H2SO4 72%, larutan ADS dan NDS.
III.3 Prosedur Kerja
III.3.1 Sterilisasi Alat
Semua alat yang digunakan dalam penelitian ini harus dalam keadaan
steril dan bebas dari segala bentuk kehidupan terutama mikroorganisme. Untuk
alat yang terbuat dari bahan gelas dicuci menggunakan sabun dan dibilas dengan
air lalu dikering-anginkan, kemudian disterilkan dengan menggunakan oven pada
suhu 180°C selama 2 jam. Sedangkan alat-alat non gelas yang tidak tahan panas,
22

xxxvi
dicuci dan dikering-anginkan lalu disterilkan dengan menggunakan otoklaf pada
suhu 121°C dengan tekanan 2 atm selama 15 - 30 menit. Alat-alat yang terbuat
dari logam seperti ose bulat, ose lurus, jarum preparat dan pinset disterilkan
dengan cara dibilas dengan alkohol lalu dipanaskan di atas nyala api Bunsen
hingga pijar. Alat lain yaitu enkas disterilkan dengan menyemprotkan alkohol
70% pada seluruh bagian dalam enkas, lalu diberikan pemanasan dengan
menyalakan api Bunsen kemudian segera pintu enkas ditutup dan dibiarkan
selama 30 menit sebelum digunakan.
III.3.2 Pembuatan Medium
III.3.2.1 Medium Potato Dextrosa Agar (PDA)
Bahan yang digunakan adalah kentang 200 g, agar 20 g, dan dextrosa 15 g.
Bahan terlebih dahulu ditimbang sesuai dengan jumlah yang dibutuhkan. Kentang
direbus dalam 1 liter aquades hingga mendidih, kemudian mengukur volume
ekstrak kentang menggunakan gelas ukur lalu menambahkan aquades steril untuk
mencukupkan volume hingga 1 liter, untuk mengganti volume air yang hilang saat
pemanasan. Ekstrak kentang dimasukkan ke dalam labu Erlenmeyer, lalu
ditambahkan agar dan dextrosa kemudian dipanaskan di atas penangas hingga
semua bahan larut dan homogen. Setelah semua bahan larut dan homogen, labu
Erlenmeyer kemudian ditutup dengan kapas dan aluminium foil, selanjutnya
medium siap disterilkan di dalam otoklaf pada suhu 121°C dan tekanan 2 atm
selama 15 menit. Untuk mencegah pertumbuhan bakteri, ke dalam medium
ditambahkan antibiotik chloramphenicol 500 mg pada saat medium akan
digunakan (Dwyana dan Gobel, 2011).
23

xxxvii
III.3.2.2 Pembuatan Substrat Bahan Organik Sebagai Media Tumbuh Isolat Jamur (baglog) Jerami padi yang digunakan diperoleh dari Kabupaten Maros Provinsi
Sulawesi Selatan. Di laboratorium jerami padi di cacah menjadi bagian-bagian
yang lebih kecil. Jerami padi yang telah dipotong ditimbang sebanyak 3 kg,
selanjutnya dicampurkan dengan dedak sebanyak 600 gram dan kapur sebanyak
30 gram lalu dicampur rata dengan ditambahkan air sedikit demi sedikit sampai
jeraminya basah. Jerami yang sudah tercampur rata tersebut kemudian
dimasukkan ke dalam 15 kantong plastik tahan panas dan tiap kantongnya
sebanyak 150 gram kemudian ditutup menggunakan kapas steril lalu diautoklaf
selama 2 x 3 jam (Achmad dkk, 2011).
III.3.3 Peremajaan Isolat.
Pada penelitian ini digunakan isolat jamur pelapuk koleksi Laboratorium
Bioteknologi Pusat Kegiatan Penelitian (PKP), Universitas Hasanuddin,
Makassar. Isolat jamur ini diambil dari kayu lapuk di sekitar daerah Makassar.
Media PDA (Potatoes Dextrose Agar) steril dituang ke cawan petri yang
steril di Laminar Air Flow secara aseptik. Media dibiarkan dingin dan memadat.
Setelah media PDA padat, isolat jamur yang telah tersedia dipotong dadu dengan
ukuran 1x1 cm, kemudian dipindahkan satu potong ke dalam media PDA secara
aseptik. Setelah selesai, cawan petri disegel dengan parafilm dan diinkubasi
pada suhu kamar (28 0C) selama 3-5 hari hingga terbentuk miselia. Untuk melihat
pertumbuhan miselia pada media, cukup dilakukan pengamatan secara visual
24

xxxviii
karena penampakan miselia pada media sangat khas seperti serat-serat (Sigit,
2008).
III.3.4 Seleksi Jamur Lignolitik.
Tahapan-tahapan seleksi jamur lignolitik adalah sebagai berikut (Achmad
dkk, 2011) :
III.3.4.1 Inokulasi Isolat Jamur Pada Substrat Bahan Organik
Agar dan miselia yang berasal dari cawan petri dipotong kecil dengan
ukuran 1cm x1 cm, kemudian 5 potongan kecil dimasukkan ke dalam 14 kantong
plastik tahan panas yang berisi substrat bahan organik (baglog) yang telah
disterilkan, lalu dicampurkan dengan cara diaduk-aduk dengan substratnya.
Plastik kemudian ditutup kembali dengan sumbat kapas steril kemudian
diikat dengan karet lalu ditutup dengan menggunakan plastik parafilm. Pengerjaan
dilakukan secara aseptik di dalam Laminary Air Flow (LAF). Satu kantong plastik
sebagai kontrol, tidak diinokulasi dengan isolat jamur pelapuk.
Media cacahan jerami yang sudah diinokulasi dengan bibit jamur
kemudian diinkubasikan selama 30 hari. Pengamatan dilakukan setiap tiga hari
sekali dengan melihat pertumbuhan dari setiap isolat pada baglog dan untuk
melihat isolat yang paling cepat memenuhi baglog.
III.3. 4. 2 Analisis Lignin, Hemiselulase dan Selulase
Jerami padi yang telah difermentasi dikeluarkan dari plastik, hal yang
sama juga dilakukan pada kontrol, kamudian dilakukan pengamatan terhadap
tekstur produk fermentasi serta analisis kandungan serat kasar (CF). Untuk
25

xxxix
menentukan kadar lignin, hemiselulosa dan selulosa terlebih dahulu ditentukan
kadar ADF dan NDF menggunakan metode Van Soest (1976).
a. Penentuan Kadar Acid Detergent Fiber (ADF)
Sampel sebanyak 0,5 gram (a gram) dimasukkan ke dalam gelas piala
kemudian ditambahkan 50 ml larutan ADS. Dipanaskan selama 1 jam diatas
penangas air. Kemudian dilakukan penyaringan dengan menggunakan penyaring
kaca masir yang sudah di timbang sebagai b gram sambil diisap pompa vakum.
Pencucian di lakukan dengan menggunakan air panas dan alkohol. Selanjutnya
dilakukan pengeringan dengan memasukkan hasil penyaringan tersebut dalam
oven pada suhu 105 oC, setelah itu dimasukkan lagi di dalam desikator untuk
melakukan pendinginan selama lebih kurang ½ jam kemudian ditimbang sebagai c
gram.
c - b %ADF = --------- x 100% a Ket : a = berat sampel b = berat kaca masir
c = berat kaca masir + berat sampel setelah ditambah larutan ADS, dioven dan didinginkan dalam desikator
b. Penentuan Neutral Detergent Fiber (NDF)
Sampel sebanyak 0,5 gram (a gram) di masukkan ke dalam gelas piala
berukuran 500 ml, kemudian di tambahkan dengan 50 ml larutan NDS lalu
dipanaskan selama 1 jam. Kemudian dilakukan penyaringan dengan menggunakan
penyaring kaca masir yang sudah di timbang sebagai b gram sambil diisap pompa
vakum. Pencucian di lakukan dengan menggunakan air panas. Selanjutnya
dilakukan pengeringan dengan memasukkan hasil penyaringan tersebut dalam
26

xl
oven pada suhu 105 oC, setelah itu dimasukkan lagi di dalam desikator untuk
melakukan pendinginan selama lebih kurang ½ jam kemudian ditimbang sebagai c
gram.
c - b %NDF = ---------- x 100% a
Ket : a = berat sampel b = berat kaca masir
c = berat kaca masir + berat sampel setelah ditambah larutan NDS, dioven dan didinginkan dalam desikator
c. Penentuan Persentase Lignin, Selulosa dan Hemiselulosa
% Hemisellulosa = %NDF - % ADF
Persentase Lignin dan Selulosa
Untuk menghitung pesentase lignin dan selulosa dilakukan melalui cara
yaitu residu ADF (c gram) yang berada di dalam kaca masir diletakkan di atas
nampan yang berisi air setinggi kira-kira 1 cm lalu ditambahkan H2S04 72%
setinggi ¾ bagian gelas kaca masir dan dibiarkan selama 3 jam sambil
diaduk-aduk. Penyaringan dilakukan dengan kaca masir dan dengan bantuan
pompa vakum serta pencucian juga dilakukan seperti analisis sebelumnya.
Selanjutnya dilakukan pengeringan dengan menggunakan oven dengan suhu
105°C dan selanjutnya dilakukan pendinginan dengan desikator dan ditimbang
sebagai berat akhir, yaitu e gram.
c - e % Selulosa = -------------- x 100% a
Ket : a = berat sampel c = residu ADF
e = berat kaca masir + berat residu ADF setelah ditambahakan H2S04 72%, kemudian dioven dan didinginkan dalam desikator
27

xli
Jika dibakar dalam tanur 500°C kemudian didinginkan dalam desikator
serta disimpan kembali sebagai berat akhir, yaitu f gram.
e - f % Lignin = ---------------- x 100% a
Ket : a = berat sampel e = berat kaca masir + berat residu ADF setelah ditambahakan H2S04 72% f = berat residu ADF setelah + H2S04 72% lalu dibakar dalam tanur 500 °C
III.3.5 Analisis Data
Kemampuan isolat jamur pelapuk kayu dalam mendekomposisi jerami
padi dapat diketahui dari pertumbuhan koloni jamur. Isolat jamur yang paling
cepat tumbuh dan memenuhi baglog yaitu isolat yang mempunyai kemampuan
dalam memanfaatkan jerami padi sebagai sumber makanannya. Untuk mengetahui
pengurangan kadar lignin, selulosa dan hemiselulosa pada jerami padi maka
setelah diinkubasi selama 30 hari, jerami padi dikeluarkan dari plastik dan
dianalisis kadar lignin, selulosa dan hemiselulosa di Laboratorium Kimia dan
Makanan, Fakultas Peternakan, Universitas Hasanuddin, Makassar.
28

xlii
BAB IV
HASIL DAN PEMBAHASAN
IV.1 Pertumbuhan Isolat Jamur Pada Baglog Jerami Padi
Jamur pelapuk kayu isolat KSH, KSB, MKS, JM, B, C dan E yang
digunakan pada penelitian merupakan isolat terpilih hasil skrining yang
menunjukan potensi lignoselulolitik (Lampiran 1). Isolat ini diperoleh dari kayu
lapuk di sekitar daerah Makassar. Masing-masing isolat diinokulasi dalam setiap
baglog jerami padi dan kemudian diinkubasi pada suhu ruangan di dalam
inkubator. Pertumbuhan jamur diamati secara visual setiap 3 hari sekali selama 30
hari tanpa membuka baglog. Berikut ini merupakan gambar pengamatan pertama
setelah inkubasi selama 3 hari.
A B C D
E F G H
Gambar 4. Pertumbuhan Isolat KSH (A), Isolat KSB (B), Isolat JM (C), Isolat
MKS (D), Isolat B (E) , Isolat C (F), Isolat E (G) dan (H) Kontrol pada baglog jerami padi 3 hari setelah inokulasi.
29

xliii
Pertumbuhan ketujuh isolat jamur pada baglog jerami padi selama 3 hari
setelah proses inokulasi menunjukkan kecepatan pertumbuhan yag berbeda-beda.
Isolat KSH merupakan isolat yang paling cepat pertumbuhannya, namun miselia
yang tumbuh belum memenuhi seluruh baglog. Untuk 6 isolat lainnya yaitu KSB,
JM, MKS, B, C, dan E miselianya sudah tumbuh tetapi belum terlalu banyak
(Gambar 4 dan Tabel 1).
Hasil pengamatan ke-2 sampai pengamatan ke-9 dapat dilihat pada
Lampiran 2. Pada pengamatan terakhir yaitu pengamatan ke-10 (30 hari inkubasi),
ketujuh isolat jamur memperlihatkan kemampuan bertumbuh yang baik pada
media organik jerami padi. Isolat yang paling bagus pertumbuhannya adalah isolat
KSH, JM, MKS dan isolat E (Gambar 5 dan Tabel 1).
A B C D
E F G H
Gambar 5. Pertumbuhan isolat KSH (A), Isolat KSB (B), Isolat JM (C), Isolat
MKS (D), Isolat B (E), Isolat C (F), Isolat E (G) dan Kontrol (H)
baglog jerami padi 30 hari setelah inokulasi.
30

xliv
Hasil pengamatan pertumbuhan miselia yaitu setiap 3 hari sekali sampai
pada pengamatan terakhir (30 hari inkubasi), dari ketujuh isolat yang digunakan
pada penelitian ini, isolat KSH mempunyai kecepatan pertumbuhan yang cepat,
diikuti oleh isolat JM, lalu isolat E, kemudian isolat MKS, isolat KSB, isolat C
dan yang terakhir adalah isolat B. Warna miselia dari ke tujuh isolat juga berbeda-
beda. Isolat KSH, isolat C, dan isolat B dan memiliki warna miselia kehijauan,
isolat KSB, JM dan MKS berwarna putih sedangkan isolat E warna miselianya
hitam (Gambar 5 dan Tabel 1).
Tabel 1. Perbedaan pertumbuhan dan warna miselia dari ketujuh isolat setelah 3 hari sampai 30 hari inkubasi
Keterangan : + = Miselia sudah tumbuh tetapi belum terlalu banyak ++ = Miselia yang tumbuh sudah banyak tetapi belum memenuhi baglog +++ = Miselia yang tumbuh sangat banyak dan memenuhi baglog.
Ketujuh isolat dapat tumbuh pada substrat jerami padi karena mampu
menggunakan komponen lignoselulosa yaitu lignin, selulosa dan hemiselulosa
jerami padi sebagai nutrisi untuk pertumbuhannya. Hal ini dapat diketahui dengan
Isolat
3 hari 30 hari
Pertumbuhan Warna Pertumbuhan Warna
KSH ++ Kehijauan +++ Kehijauan
KSB + Putih ++ Putih
JM + Putih +++ Putih
MKS + Putih +++ Putih
B + Kehijauan ++ Kehijauan
C + Kehijauan ++ Kehijauan
E + Hitam +++ Hitam
31

xlv
tumbuh dan berkembangnya miselium jamur pada substrat jerami padi. Menurut
Hendritomo (2002), senyawa karbon yang dapat digunakan oleh jamur
diantaranya monosakarida, oligosakarida, asam organik, alkohol, selulosa dan
lignin. Sumber karbon dibutuhkan untuk keperluan energi dan struktural jamur
(Chang dan Miles, 1989).
Pada Gambar 5 dan Tabel 1, isolat KSB, B dan C, menunjukan
pertumbuhan yang agak lambat bila dibandingkan dengan 4 isolat lainnya (KSH,
JM, MKS dan E), tidak memenuhi baglog sampai pada pengamatan terakhir
selama inkubasi. Pertumbuhan jamur dapat dipengaruhi oleh beberapa faktor yaitu
faktor media tumbuh dan faktor lingkungan. Faktor media tumbuh salah satunya
adalah nutrisi yang merupakan faktor yang sangat penting dalam pertumbuhan
jamur. Media tumbuh harus memiliki unsur C, N, dan S. Menurut Stamets dan
Chilton (1983) kandungan nitrogen pada substrat mempengaruhi pertumbuhan
miselium. Miselium jamur tidak dapat tumbuh pada media yang kekurangan unsur
nitrogen, tetapi kelebihan nitrogen pada substrat dapat menyebabkan
terakumulasinya amonia yang dapat meningkatkan pH sehingga menghambat
pertumbuhan miselium dan pembentukan tubuh buah.
Faktor lingkungan yang mempengaruhi pertumbuhan jamur yaitu yaitu
faktor diantaranya suhu, kelembaban ruangan, cahaya , dan sirkulasi udara. Suhu
inkubasi jamur berkisar antara 22-28oC dengan kelembaban 60-80%. Selain suhu
dan kelembaban, faktor cahaya dan sirkulasi udara juga sangat menentukan..
Sirkulasi udara harus cukup, tidak terlalu besar tetapi tidak pula terlalu kecil.
32

xlvi
Intensitas cahaya yang diperlukan pada saat pertumbuhan sekitar 10%.
(Yuniasmara et al. , 2004).
IV.2 Analisis Kandungan Lignin, Hemiselulosa dan Selulosa
Komponen lignin, hemiselulosa dan selulosa pada dinding sel tanaman
selalu berikatan membentuk suatu ikatan kompleks. Untuk menentukan kadar
lignin, selulosa dan hemiselulosa maka terlebih dahulu ditentukan kadar NDF dan
ADF.
Tabel 2. Persentase kadar NDF dan ADF pada jerami padi kontrol dan jerami padi setelah diinokulasi isolat jamur pelapuk
Sampel
Komposisi (%)
NDF ADF
Kontrol 81,05 52,40
KSB 73, 26 52, 30
KSH 58, 92 47, 88
JM 66, 57 52, 18
MKS 77, 37 55, 10
B 75, 72 53, 62
C 61, 45 50, 60
E 59, 26 45, 95
NDF : Neutral Detergent insoluble Fiber ADF : Acid Detergent insoluble Fiber
Jerami padi Oryza sativa L yang di gunakan pada penelitian ini memiliki
kandungan hemiselulosa 28, 65 %, selulosa 40, 89 % dan lignin 9, 89 % yaitu
merupakan kandungan kontrol tanpa pemberian isolat jamur pelapuk. Setelah
jerami padi dinokulasi 7 isolat jamur pelapuk yang berbeda selama 30 hari maka
terjadi penurunan komponen lignin, hemiselulosa dan selulosa, dapat dilihat pada
Tabel 3.
33

xlvii
Tabel 3. Persentase kadar dan penurunan Lignin, Hemiselulosa, dan Selulosa setelah diinokulasi dengan 7 isolat jamur pelapuk dan pada kontrol jerami padi setelah 30 hari inkubasi
Perlakuan
Lignin (%) Hemiselulosa (%) Selulosa (%)
Kadar Penurunan Kadar Penurunan Kadar Penurunan
Kontrol 9, 89 - 28, 65 - 40, 89 -
Isolat KSH 9, 63 2.62 11, 04 61.46 36, 68 10.29
Isolat KSB 9, 47 4.24 20, 96 26.84 38, 55 5.72
Isolat JM 8, 19 17.18 14, 39 49.77 34, 38 15.92
Isolat MKS 9, 19 7.07 22, 27 22.26 31, 42 23.15
Isolat B 8, 99 9.1 22, 10 22.86 37, 93 7.23
Isolat C 8, 70 12.03 14, 85 48,16 34, 12 16.55
Isolat E 8, 26 16.48 13, 31 53.54 23, 99 41.33
Lignin secara umum merupakan komponen yang belum mengalami
penurunan yang berarti pada semua perlakuan isolat jamur. Penurunan komponen
lignin tertinggi yaitu jerami padi yang diinokulasi isolat JM dari 9, 89%
kandungan lignin pada jerami padi kontrol tanpa pemberian isolat jamur dan
tersisa sekitar 8, 19 % (Tabel 2) atau terjadi penurunan sebesar 17,18%, dan yang
terendah yaitu pada jerami padi yang diinokulasi isolat KSH yaitu sebesar 2, 62%,
kemudian berturut-turut adalah isolat E, C, B MKS, dan KSB masing-masing
sebesar 16, 48 %, 12, 03 %, 9, 10 %, 7, 07 %, 4, 24 % ( Tabel 3 dan Gambar 6).
34

Gambar 6. Grafik Persentase Penurunan Kandungan Lignin, Hemiselulosa dan Selulosa Pada Jerami Padi.
Komponen hemiselulosa jerami padi merupakan komponen yang paling
banyak terdegradasi. Komponen hemiselulosa jerami padi pada kontrol sebesar
28, 65 %, kemudian tersisa sekitar
terjadi penurunan sekitar 61, 4
paling tinggi (Gambar 6). Hampir semua isolat memperlihatkan kemampuan yang
baik dalam mendegradasi hemiselulosa dan terendah pada perlakuan dengan
menggunakan isolat MKS yaitu sebesar
dan B masing- masing sebesar 53, 54 %, 49, 77 %, 48, 16
% (Tabel 3 dan Gambar 6).
Selulosa merupakan komponen yang paling banyak terdapa
padi. Pada Tabel 3 dan Gambar 6 dapat dilihat bahwa dengan pemberian
jamur pelapuk maka terjadi penurunan kandungan selulosa, dan yang paling tinggi
0
10
20
30
40
50
60
70
80
90
100
KSH KSB
%
xlviii
Persentase Penurunan Kandungan Lignin, Hemiselulosa dan Selulosa Pada Jerami Padi.
Komponen hemiselulosa jerami padi merupakan komponen yang paling
banyak terdegradasi. Komponen hemiselulosa jerami padi pada kontrol sebesar
%, kemudian tersisa sekitar 11, 04% setelah diinokulasi isolat KSH
terjadi penurunan sekitar 61, 46 %, ini merupakan persentase penurunan yang
paling tinggi (Gambar 6). Hampir semua isolat memperlihatkan kemampuan yang
baik dalam mendegradasi hemiselulosa dan terendah pada perlakuan dengan
menggunakan isolat MKS yaitu sebesar 22. 26 %. Untuk isolat E, JM , C
masing sebesar 53, 54 %, 49, 77 %, 48, 16 %, 26, 84%
dan Gambar 6).
Selulosa merupakan komponen yang paling banyak terdapat pada jerami
dan Gambar 6 dapat dilihat bahwa dengan pemberian
jamur pelapuk maka terjadi penurunan kandungan selulosa, dan yang paling tinggi
JM MKS B C EIsolat
Lignin
Hemiselulosa
Selulosa
35
Persentase Penurunan Kandungan Lignin, Hemiselulosa dan
Komponen hemiselulosa jerami padi merupakan komponen yang paling
banyak terdegradasi. Komponen hemiselulosa jerami padi pada kontrol sebesar
11, 04% setelah diinokulasi isolat KSH atau
6 %, ini merupakan persentase penurunan yang
paling tinggi (Gambar 6). Hampir semua isolat memperlihatkan kemampuan yang
baik dalam mendegradasi hemiselulosa dan terendah pada perlakuan dengan
JM , C, KSB
dan 22, 86
t pada jerami
dan Gambar 6 dapat dilihat bahwa dengan pemberian inokulan
jamur pelapuk maka terjadi penurunan kandungan selulosa, dan yang paling tinggi
Lignin
Hemiselulosa
Selulosa

xlix
adalah jerami padi yang dinokulasi dengan isolat E sebesar 41, 33 % dan terendah
adalah isolat KSB sebesar 5, 72 % , kemudian 23, 15 %, 16, 55 %, 15, 92 %, 10,
29 % dan 7, 23 %, masing-masing untuk isolat MKS, C, JM, KSH, dan B.
Kemampuan isolat jamur pelapuk dalam mendegradasi komponen
lignoselulosa dapat diketahui dari nilai presentase tingkat penurunan kandungan
lignin, hemiselulosa dan selulosa. Semakin tinggi nilai presentase tingkat
penurunan kandungan lignin, hemiselulosa dan selulosa suatu bahan maka
kemampuan jamur dalam menguraikan bahan tersebut semakin baik. Pada
Gambar 6 memperlihatkan kemampuan dari masing-masing isolat jamur dalam
mendekomposisi komponen lignin, hemiselulosa dan selulosa jerami padi.
Komponen hemiselulosa merupakan komponen yang tinggi didegradasi oleh
hampir semua isolat jamur kecuali isolat MKS (komponen yang paling banyak
didegradasi adalah selulosa), kemudian diikuti oleh komponen selulosa,
sedangkan lignin belum memperlihatkan penurunan yang nyata hingga akhir
pengamatan.
Hasil yang diperoleh sejalan dengan penelitian yang dilakukan oleh
Ermawar, dkk., (2006) yang melaporkan bahwa semakin lama waktu inkubasi
jamur maka semakin banyak komponen lignin yang terdegradasi dan pada saat
yang bersamaan komponen holoselulosa (alfaselulosa dan hemiselulosa) pada
jerami padi juga ikut terdegradasi. Selain itu Anita dkk., (2009) mengungkapkan
bahwa pada perlakuan menggunakan jamur Trametes versicolor komponen
holoselulosa yang hilang pada umumnya adalah hemiselulosa sedangkan pada
36

l
perlakuan menggunakan jamur Pleurotus ostreatus, komponen terbesar
holoselulosa yang hilang adalah alfa selulosa.
Pada penelitian ini komponen hemiselulosa jerami padi merupakan
komponen yang lebih banyak didegradasi oleh isolat jamur pelapuk. Menurut
Taherzadeh (1999) hal ini disebabkan karena hemiselulosa yang memiliki yang
memiliki rantai bercabang, merupakan kelompok polisakarida yang terdiri dari
berbagai senyawa gula, seperti xilosa, arabinosa, dan galaktosa yang berantai
pendek serta memiliki berat molekul dan derajat polimerisasi yang rendah. Selain
itu menurut Perez et al (2002) komponen hemiselulosa dapat didegradasi karena
isolat jamur menghasilkan enzim hemiselulase. Hemiselulase seperti kebanyakan
enzim lainnya yang dapat menghidrolisis dinding sel tanaman merupakan protein
multi-domain. Xilan merupakan karbohidrat utama penyusun hemiselulosa dan
Xylanase merupakan hemiselulase utama yang menghidrolisis ikatan β-1,4 rantai
xilan menjadi oligosakarida.
Selulosa merupakan komponen kedua yang banyak didegradasi oleh isolat
jamur pelapuk pada penelitian ini. Selulosa merupakan polimer glukosa dengan
ikatan β-1,4 glukosida dalam rantai lurus. Bangun dasar selulosa berupa suatu
selobiosa yaitu dimer dari glukosa. Rantai panjang selulosa terhubung secara
bersama melalui ikatan hidrogen dan gaya van der Waals (Perez et al. 2002).
Isolat jamur pelapuk dapat mendegradasi selulosa karena mampu
mengasilkan sekelompok enzim selulolitik yang bekerja secara sinergis yaitu
enzim 1,4-β-D-glucan- 4- glucanohydrolases (endoglucanases), enzim 1,4-β-D-
glucan glucanohydrolases atau cellodextrinase dan 1,4-β-D-glucan
37

li
cellobiohydrolases atau cellobiohydrolases (exoglucanases). Hasil kerja sinergis
endoglucanases dan exoglucanases menghasilkan molekul selobiosa. Hidrolisis
selulosa secara efektif memerlukan enzim (β-glucosidases yang memecah
selobiosa menjadi 2 molekul glukosa (Perez et al. 2002; Howard et al. 2003).
Hasil penelitian ini juga menunjukkan bahwa komponen lignin merupakan
komponen yang paling lambat didegradasi. Hal ini disebabkan karena lignin
merupakan komponen yang memiliki struktur yang sangat kompleks dan
heterogen serta bersifat rekalsitran karena tahan terhadap degradasi atau tidak
terdegradasi dengan cepat , selain itu diperkirakan karena isolat jamur pelapuk
lebih banyak menggunakan komponen hemiselulosa yang lebih mudah
didegradasi, sehingga isolat jamur dapat tumbuh dengan baik tanpa harus
memecah lignin sebagai sumber energinya. Menurut Kirk and Farrell (1987)
terjadinya proses degradasi lignin harus diawali dengan berkurangnya kandungan
N dan C pada substrat sehingga aktivasi enzim meningkat. Adapun enzim yang
berperan dalam proses degradasi lignin tersebut adalah fenol oksidase (lakase)
dan peroksidase (lignin peroksidase (LiP) dan manganese peroksidase (MnP).
MnP mengoksidasi Mn2+ menjadi Mn3+ yang berperan sebagai dalam
pemutusan unit fenolik lignin. LiP mengkatalis oksidasi senyawa aromatik non
fenolik dan jamur pelapuk putih merupakan organisme yang mampu
mendegradasi lignin secara sempurna menjadi menjadi produk yang larut dalam
air dan CO2. Pada proses inkubasi selama 30 hari tidak menunjukkan adanya kerja
enzim pendegradasi lignin yang dihasilkan oleh isolat jamur pelapuk sehingga
komponen lignin tidak banyak mengalami degradasi. Hal ini juga didukung oleh
38

lii
Jung et al (1992) yang menyatakan bahwa perubahan komponen kimia berupa
lignin sangatlah bergantung pada jenis jamur yang digunakan dan juga jenis
substratnya.
39

liii
BAB V
KESIMPULAN DAN SARAN
V.1 Kesimpulan
1. Isolat KSH, KSB, JM, MKS, B, C, E dapat tumbuh pada media jerami padi
tetapi kecepatan pertumbuhan setiap isolat berbeda-beda, namun isolat yang
paling cepat pertumbuhannya adalah isolat KSH dan JM.
2. Isolat yang paling banyak mendekomposisi komponen lignin adalah isolat
JM (17, 18 %) dan komponen hemiselulosa adalah isolat KSH (61, 46 %),
sedangkan selulosa adalah isolat E (41, 33 %).
V.2 Saran
Perlu adanya penelitian lebih lanjut mengenai proses dekomposisi
menggunakan campuran isolat jamur pelapuk dengan bakteri selulolisis sehingga
dapat mempercepat proses dekomposisi. Untuk memperoleh hasil yang baik
dalam mendegradasi lignin sebaiknya sebelum di inokulasi pada media organik
isolat jamur harus terlebih dahulu dibuat dalam bentuk spawn sehingga
beradaptasi dengan media berlignin, dan juga penambahan waktu inkubasi.
40

liv
DAFTAR PUSTAKA
Achmad, M., T. Arlianti dan C. Azmi. 2011. Panduan Lengkap Jamur. Penebar Swadaya. Jakarta.
Anita, S. H., T.Fajriutami, Fitria, R. A. Ermawar, D. H. Y. Yanto, E.Hermiati. 2009. Perlakuan awal bagasse menggunakan kultur tunggal dan kultur campuran jamur pelapuk putih Trametes versicolor dan Pleurotus ostreatus. Dipresentasikan di Seminar Nasional “Biomass Utilization for Alternative Energy and Chemicals”. Universitas Parahyangan. Bandung.
Anonim. 2012. Jerami Padi. https:// epetani.deptan.go.id/. Diakses pada tanggal 24 September 2012.
Aziz A.A., M. Husin and A. Mokhtar. 2002. Preparation of cellulose from oil palm empty fruit bunches via ethanol digestion: effect of acid and alkali catalysts. Journal of Oil Palm Research. 14 (1):9-14.
Blanchette R.A. 1995. Degradation of lignocellulose complex in wood. Can. J. Bot. 73 (Suppl. 1): S999-S1010.
Boyle C.D., B.R. Kropp and I.D. Reid. 1992. Solubilization and mineralization of lignin by white rot fungi. Appl. Environ. Microbiol. 58:3217-3224.
BPS. 2009. http://www.bps.go.id/tnmn_pgn.php. Diakses pada tanggal 24 September 2012.
Brauns, F.E. 1952. The Chemistry of Lignin. Academic Press. New York.
Chahal P.S. and D.S. Chahal. 1998. Lignocellulosic Waste: Biological Conversion. In: Martin, A.M. [eds]. Bioconversion of Waste Materials to Industrial Products. Ed ke-2. London: Blackie Academic & Professional. pp. 376-422.
Chang S.T. and P.G. Miles. 1989. Edible Mushrooms and Their Cultivation. CRC Press Inc. Florida.
Chesson, A and C.W. Forssberg. 1988. Polysaccharide Degradation by Rumen Mikroflora. In P>N. Hobson Ed. The Rumen Microbial Ecosystem. Elsevier Applied Science. London.
Crawford D.L., A.L. Pometto III and R.L. Crawford. 1983. Lignin degradation by Streptomyces viridosporus: Isolation and characterization of new polymeric lignin degra-dation intermediate. Appl. Environ. Microbiol. 45:898-904.
41

lv
Daniel G. and T. Nilsson. 1998. Developments in the study of soft rot and bacterial decay. In: Bruce A.and J.W Palfreyman [eds]. Forest Products Biotechnology. Great Britain: Taylor & Francis pp. 37-62.
Dey S., T.K. Maiti and B.C. Bhattacharyya. 1994. Production of some extracellular enzymes by a lignin peroxidase-producing brown rot fungus, Polyporus ostreiformis, and its comparative abilities for lignin degradation and dye decolorization. Appl. Environ. Microbiol. 60:4216-4218.
Dwyana, Z., dan Gobel, R. B., 2011. Penuntun Praktikum Mikrobiologi Umum. Laboratorium Mikrobiologi, Jurusan Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam, Universitas Hasanuddin, Makassar.
Ekawati, I. 2003. Pengaruh Pemberian Inokulum Terhadap Kecepatan Pengomposan Jerami Padi. Tropika. 11(2):144.
Ermawar, R.A., D.H.Y.Yanto, Fitria, and E.Hermiati. 2006. Lignin degradation content in rice straw pre-treated by white-rot fungi. Jurnal Widya Riset. 9 (3): 197-202.
Eun J.S., K.A. Beauchemin, S.H Hong, and M.W. Bauer. 2006. Exogenous enzymes added to untreated or ammoniated rice straw : Effect on in vitro fermentation characteristic and degradability. J. Anim. Sci. and Tech. 131 : 86‐101.
Fengel, D and G. Wegener. 1995. Kayu : Kimia, Ultrastruktur, Reaksi-reaksi. (Terjemahan). Gadjah Mada Univ. Peress. Yogyakarta.
Gaman, P.M. dan K.B. Sherrington. 1992. Pengantar ilmmu Pangan, Nutrisi dan Mikrobiologi. Edisi 2. Gadjah Mada University Press. Yogyakarta.
Grigoriou A.H. 2000. Straw-wood composites bonded with various adhesive systems. Wood Science and Technology.34(4):355-365.
Hadinah, S. 2007. Isolasi Bakteri dan Jamur Selulolitik sebagai Inokulum Untuk Meningkatkan Jerami Padi dan Produktivitas Domba [Disertasi] Pascasarjana Universitas Airlangga, Surabaya.
Hammel K.E. 1997. Fungal Degradation of Lignin. Di Dalam: Cadisch G, Giller KE, Editor. Driven By Nature: Plantt Litter Quality And Decompostion. London: CAB International. pp. 33-45.
Hataka, A. 2001. Biodegradation of lignin. In: Steinbüchel A. [ed] Biopolimers. Vol 1: Lignin, Humic Substances and Coal Germany: Wiley VCH. pp. 129 – 180.
Hatfield, R.D. 1989. Structural Polysacharides in Forage and Their Degradability. Agron, J. 81.
42

lvi
Hendritomo, H.I. 2002. Biologi Jamur Pangan. Pusat Pengkajian dan Penerapan Teknologi Bio Industri. Jakarta.
Howard R.L., P. Masoko and E. Abotsi. 2003a. Enzymeactivity of Phanerochaete chrysosporium cellobiohydrolase (CBHI.1) expressed as a heterologous protein from Escherichia coli. African J. Biotechnol. Vol. 2.No. 9. pp 296-300.
Jayanti, C. M. T., dan Aprilia B. 2010, Pabrik Pulp dari Jerami Padi dengan Proses Biochemical Pulping [Tugas Akhir], http://digilib.its.ac.id/public/ITS-NonDegree-12799-pabrik-pulp-dari-jerami-padi-dengan-proses-biochemical-pulping.pdf, Diakses pada tanggal 24 September 2012.
Jung, H.G. 1989. Forage Lignins and Their Effects on Feed Digestibility. Agron. J. 81.
Kaal, E.E.J., J.A. Field and T.W. Joice. 1995. Increasing Ligninolitic Enzyme Activities in Several White Rot Basiddiomycetess by Nitrogen Sufficient Media. Biosource Technology 53.
Kirk T.K. and R.L. Farrell. 1987. Enzymatic “combustion”: the microbial degradation of lignin. Ann. Rev. Microbiol. 41:465-505.
Komar, A. 1984. Teknologi Pengolahan Jerami sebagai Makanan Ternak. Cetakan pertama. Yayasan Dian Grahita. Bandung.
Lubis, D. A. 1992. Ilmu Makanan Ternak. P.T. Pembangunan. Jakarta.
Lynd L.R., P.J. Weimer, W.H. van Zyl and I.S. Pretorius. 2002. Microbial Cellulose Utilization: Fundamentals and Biotechnology. Microbiol. Mol. Biol. Rev.66(3):506-577.
Morrison, F.B. 1986. Feed and Feeding. 21th Ed. The Iowa State University Press, Iowa.
Murbandono, L. 2006. Membuat Kompos. Penebar Swadaya. Jakarta.
Nasrul, dan T.Maimun. 2009. Pengaruh Penambahan Jamur Pelapuk Putih (White Rot Fungi) pada Proses Pengomposan Tandan Kosong Kelapa Sawit. Jurnal Rekayasa Kimia dan Lingkungan.7(2):194-199.
Orth A.B., D.J. Royse, M. Tien. 1993. Ubiquity of lignin-degrading peroxidases among various wood-degrading fungi. Appl Environ Microbiol 59:4017-4023.
43

lvii
Perez J., J. Munoz-Dorado, T. de la Rubia and J. Martinez. 2002. Biodegradation and biological treatments of cellulose, hemicellulose and lignin: an overview. Int. Microbiol. 5:53-63.
Sanchez, C. 2009. Lignocellulosic Residues : Biodegradation and Bioconversion by Fungi. Biotechnology Advances 27.
Saparrat, M.C.N., F. Guillen, A.M. Arambarri, A.T. Martinez and M.J Martinez. 2002. Induction, Isolation and characterization of Two Laccases from The White Rot Basidiomycete Coriolosis rigida. Applied and Environmental Microbiology 68.
Sigit, A. M. 2008. Pola Aktivitas Enzim Lignolitik Jamur Tiram Pleurotus ostreatus Pada Media Sludge Industri Kertas. [Skripsi]. Program Studi Biokimia, Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor. Bogor.
Sjöberg, G. 2003. Lignin degradation: Long-term effects of nitrogen addition on decomposition of forest soil organic matter. [disertasi]. Uppsala: Dep. Soil Sci. Swedish University of Agricultural Sciences.
Srebotnik E., K.A. Jensen and K.E. Hammel. 1998. Fungal degradation of recalcitrant nonphenolic lignin structure without lignin peroxidase. Proc Natl Acad Sci 91:12794-12797.
Staments P. and J.S. Chilton. 1983. The Mushroom Cultivator. Agaricon Press. Olympia, Washington.
Steffen, K.T. 2003. Degradation of recalcitrant biopolymers and polycyclic aromatic hydrocarbons by litter-decomposing basidiomycetous fungi. [disertasi]. Helsinki: Division of Microbiology Department of Applied Chemistry and Microbiology Viikki Biocenter, University of Helsinki.
Subiyatno. 2010. Aplikasi Informasi Paten Untuk Perintisan Industri Baru Berbasis Sumber Daya Alam, Studi Kasus Informasi Paten Di Bidang Pemanfaatan Jerami. Pusat Inovasi Lembaga Ilmu Pengetahuan Indonesia.
Suparjo. 2008. Degradasi Komponen Lignoselulosa oleh Kapang Pelapuk Putih. http//Jajo 66.wordpress.com/. Diakses pada tanggal 24 September 2012.
Sun, Y. and J. Cheng. 2002. Hydrolysis of lignocellulosic materials for ethanol production: a review. 2002. Bio resource Technology 83: 1 – 11.
Sutardi, T. 1980. Landasan Ilmu Nutrisi Departemen Ilmu Makanan Ternak, Fakultas Peternakan Institut Pertanian Bogor.
44

lviii
Taherzadeh M.J. 1999. Ethanol from Lignocellulose: Physiological Effects of Inhibitors and Fermentation Strategies. [thesis]. Göteborg: Department of Chemical Reaction Engineering, Chalmers University Of Technology.
Tuomela, M. 2002. Degradation of lignin and other 14C-labelled compounds in compost and soil with an emphasis on white-rot fungi. Helsinki: Dep. Appl. Chem. Microbiol. Division of Microbiology University of Helsinki.
Vähätalo A.V., K. Salonen, M. Salkinoja-Salonen and A. Hatakka. 1999. Photochemical mineralization of synthetic lignin in lake water indicates rapid turnover of aromatic organic matter under solar radiation. Biodegradation 10:415-420.
Van Soest P. J. 1976. New Chemical Methods for Analysis of Forages for The Purpose of Predicting Nutritive Value. Pref IX International Grassland Cong.
Wilson KB and M. Walter. 2002. Development of Biotechnology Tool Using New Zealand White Rot Fungi to Degrade Pentachorophenol. Hasil Presentasi pada Waste Management Institute New Zealand. http://www.hortresearch.co.nz/files/2002/biorem-wasteminz.pdf.
Yarris, L. (2012). The Evolutionary Road to Biofuels. Retrieved Feb 25, 2012, From Berkeley Lab: Lawrence Berkeley National Laboratory: http://www.Lbl.Gov/Publications/Yos/Assets/Img/Biofuels_Evolution.Jpg
Yao F, Q. Wu, Y. Lei,Y. Xu. 2008. Rice straw .. fiber-reinforced high-density polyethylene composite: Effect of fiber type and loading . Industrial Crops and Products.28:63-72.
Young, R. 1986. Cellulosa Strukture Modification and Hydrolysis. New York.
Yuniasmara C, Muchrodji, M.Bakrun. 2004. Jamur Tiram. Penebar Swadaya. Jakarta.
Yunilas, 2009. Bioteknologi Jerami Padi Melalui Fermentasi Sebagai Bahan Pakan Ternak Ruminansia [Karya Ilmiah]. Departemen Peternakan, Fakultas Pertanian, Universitas Sumatra Utara. Medan.
Zhang, X., H.Yu, H. Huang,Y. Liu. 2007. Evaluation of biological pretreatment with white rot fungi for the enzymatic hydrolysis of bamboo culms. International Biodeterioration & Biodegradation 60:159–164.
45

lix
LAMPIRAN
Lampiran 1. Penampakan koloni 7 isolat jamur pelapuk pada media PDA, 7 hari setelah inokulasi.
JM MKS E
Isolat JM Isolat MKS Isolat E
KSB KSH B
Isolat KSB Isolat KSH Isolat B
C
Isolat C
46

lx
Lampiran 2. Gambar Pengamatan ke-2 sampai ke-9 Gambar pengamatan ke-2 (6 hari) setelah inokulasi
Isolat KSH Isolat KSB Isolat JM Isolat MKS
Isolat B Isolat C Isolat E Kontrol
Gambar pengamatan ke-3 (9 hari) setelah inokulasi
Isolat KSH Isolat KSB Isolat JM Isolat MKS
Isolat B Isolat C Isolat E Kontrol
47

lxi
Gambar pengamatan ke-4 (12 hari) setelah inokulasi
Isolat KSH Isolat KSB Isolat JM Isolat MKS
Isolat B Isolat C Isolat E Kontrol
Gambar pengamatan ke-5 (15 hari) setelah inokulasi
Isolat KSH Isolat KSB Isolat JM Isolat MKS
Isolat B Isolat C Isolat E Kontrol
48

lxii
Gambar pengamatan ke-6 (18 hari) setelah inokulasi
Isolat KSH Isolat KSB Isolat JM Isolat MKS
Isolat B Isolat C Isolat E Kontrol
Gambar pengamatan ke-7 (21 hari) setelah inokulasi
Isolat KSH Isolat KSB Isolat JM Isolat MKS
Isolat B Isolat C Isolat E Kontrol
49

lxiii
Gambar pengamatan ke-8 (24 hari) setelah inokulasi
Isolat KSH Isolat KSB Isolat JM Isolat MKS
Isolat B Isolat C Isolat E Kontrol
Gambar pengamatan ke-9 (27 hari) setelah inokulasi
Isolat KSH Isolat KSB Isolat JM Isolat MKS
Isolat B Isolat C Isolat E Kontrol
50

lxiv
Lampiran 3. Diagram alir analisis serat dengan metode Van Soest
Di + larutan ADF Di + larutan ADF
Dibilas dengan air panas Dibilas air panas
Dioven 105 oC Dioven 105 oC Ditimbang (ADF)
Ditambah H2SO4 75 ml
Dicuci air panas
Dioven 105 oC Ditimbang (selulosa)
Filtrat Residu
Residu untuk ananlisis lignin
Dipanaskan dengan tanur 500 oC
Ditimbang (lignin)
Residu untuk ananlisis selulosa
Disaring
Ditimbang (NDF)
Disaring Disaring
Filtrat Residu Filtrat Residu
Sampel jerami padi Oryza sativa L. setelah diinokulasi 7 isolat jamur
Analisis ADF Analisis NDF
0,5 g sampel 0, 5 g sampel
Dipanaskan Dipanaskan
51