kandidat vaksin iradiasi streptococcus agalactiae untuk ... · subclinical mastitis (scm) is a...
TRANSCRIPT

KANDIDAT VAKSIN IRADIASI Streptococcus agalactiae UNTUK
PENCEGAHAN MASTITIS SUBKLINIS PADA RUMINANSIA
BOKY JEANNE TUASIKAL
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
2012


PERNYATAAN MENGENAI DISERTASI DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi yang berjudul: “Kandidat Vaksin Iradiasi
Streptococcus agalactiae Untuk Pencegahan Mastitis Subklinis pada Ruminansia” adalah karya
saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada
perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang
diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan
dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Agustus 2012
Boky Jeanne Tuasikal
NRP B361070021


ABSTRACT
BOKY JEANNE TUASIKAL. Streptococcus agalactiae Irradiated Vaccine Candidate for
Subclinical Mastitis Prevention in Ruminants. Under supervision of Fachriyan H. Pasaribu, Sri
Estuningsih, I Wayan T. Wibawan.
Subclinical mastitis (SCM) is a disease that often infects dairy cow, which then decline
milk production. Streptococcus agalactiae is well known as a dominant causative agent of SCM.
An experiment to obtain gamma irradiated candidates vaccine of S. agalactiae for the prevention
of subclinical mastitis in ruminants have been conducted. S. agalactiae bacteria selected for the
irradiated vaccine material was isolated from dairy cows of subclinically mastitis from dairy
farm in Bogor and Garut area. The S. agalactiae used in this experiment was group B (GBS)
type, which is uncapsulated and has haemaglutynine (Hn+) property. Based on dose irradiation
orientation, 17 Gy dose was found to weaken the GBS bacteria to have 50% lethal dose (LD50),
which therefore, utilized to manufacture live vaccine. Irradiated GBS surface protein was
characterized by SDS-PAGE and resulted to find more than 75 kDa of the protein’s molecular
weight, where minimun of antigenically substance is 10 kDa. Infecting of the irradiated GBS
using mice resulted in lack pathogenic properties of the bacteria, which is histophatology drawn
that the mammary gland alveolly of mice grouped vaccinated with irradiated GBS and
vaccinated then challenge, were remain producing milk, and significantly not different as
compare to control animal. GBS vaccine irradiation did not cause inflammation of the mammary
glands of mice tested, evidences showed that number of udder interstitial inflammatory cells
around alveolly and gland epithelial cells in vaccinated mice groups were not significantly
different as compare to the control group. Most interstitial inflammatory cells in challenged
groups are found in mice that were challenged with virulent GBS and significantly different to be
found more as compare to vaccine and control groups. Enzyme Linked Immunosorbent Assay
(ELISA) to Ig-G serum showed levels of higher immune response in mice vaccinated group
compare to control group. The overall experiment confirmed that attenuated GBS using gamma
ray irradiation capable to induce immune response. This evidences were confirmed by an
observation result in the application of GBS irradiated vaccine at Etawah dairy-goat, where
immune responses are induced in those dairy-goats, and also were showed by the evidences
found of higher concentration of Ig-G serum by ELISA in the vaccinated group than the control
group. Milk production of vaccinated goat had no effect by vaccination. In summarize,
irradiated SGB vaccine does not affect milk yield.
Keywords: Subclinical mastitis, Group-B S. agalactiae, irradiated vaccine.


RINGKASAN
BOKY JEANNE TUASIKAL. Kandidat Vaksin Iradiasi Streptococcus agalactiae untuk
Pencegahan Mastitis Subklinis pada Ruminansia. Dibawah bimbingan Fachriyan H. Pasaribu, Sri
Estuningsih, I Wayan T. Wibawan.
Mastitis subklinis (MSK) adalah penyakit yang sering menyerang sapi perah sehingga
menyebabkan turunnya produksi susu. Streptococcus agalactiae dikenal sebagai bakteri dominan
yang umum menyebabkan MSK. Untuk mengantisipasi kasus ini, telah dilakukan suatu
penelitian untuk memperoleh kandidat vaksin iradiasi gamma S. agalactiae untuk pencegahan
MSK pada ruminansia. Sebanyak 65 ekor sapi perah yang terdiri dari 19 ekor berasal dari Kunak
Kabupaten Bogor, serta 25 ekor dari Cisurupan dan 21 ekor dari Bayongbong Kabupaten Garut,
telah diuji untuk MSK menggunakan reagen California Mastitis Test (CMT). Total sampel susu
yang diambil dari tiap puting sapi adalah sebanyak 240 sampel yang berasal dari Kunak (66
sampel), Cisurupan (97 sampel) dan Bayongbong (77 sampel). Hasil uji CMT menunjukkan
bahwa proporsi kejadian MSK berturut-turut dari daerah terinfeksi tertinggi ke daerah terendah
yaitu Kunak (100%), Cisurupan (94,84%) dan Bayongbong (93,50%). Dari sampel susu kasus
MSK tersebut diperoleh 57 genus Streptococcus sp. dari hasil uji Gram positif dan katalase
negatif dengan persentase kehadiran dalam sampel dari Kunak (40,90%), Cisurupan (15,22%)
dan Bayongbong (22,22%). Selanjutnya 34 isolat S. agalactiae diperoleh dengan uji keberadaan
faktor Christie, Atkins, Munch-Petersen (CAMP) pada media agar darah. Hasil pengelompokkan
dengan Streptococcal grouping kit diperoleh 14 isolat S. agalactiae group-B (SGB). Ekspresi
fenotip protein permukaan bakteri tanpa kapsul diidentifikasi dengan penanaman SGB pada
media soft agar (SA), sedangkan sifat adhesi SGB diuji dengan reaksi hemaglutinasi. Bakteri
terpilih untuk bahan vaksin iradiasi ini diperoleh dari kasus mastitis subklinis di peternakan
daerah Kunak Bogor, yaitu satu isolat S. agalactiae group-B (SGB) yang tidak berkapsul dan
memiliki sifat Hemaglutinin (Hn+). Berdasarkan orientasi dosis iradiasi gamma dengan sumber
60Co diketahui dosis sebesar 17 Gy untuk melemahkan bakteri SGB terpilih sampai taraf Lethal
dose 50% (LD50) untuk pembuatan vaksin hidup. Dosis iradiasi SGB tersebut diperoleh dari
kurva Pengaruh Iradiasi terhadap % Viabilitas SGB Hn+ dengan persamaan Y=95,414e
-0,0371X ;
R2=0,9979 (Y=%Viabilitas dan X= dosis iradiasi). Karakterisasi Protein permukaan SGB iradiasi
ini dengan metode SDS-PAGE mempunyai berat molekul lebih dari 75 KDa yang berarti

bersifat antigenik karena BM lebih dari 10 Kda (syarat BM minimal suatu protein bersifat
antigenik). Uji patogenitas dan imunitas pada hewan percobaan mencit betina jenis Balb-C,
berumur 8 minggu dengan bobot badan kira-kira 50 gram. Sebanyak 12 ekor mencit dibagi
kedalam 4 kelompok perlakuan, menjadi 3 ekor untuk tiap kelompok yaitu kelompok vaksin (V)
diberi SGB Hn+
iradiasi, kelompok vaksin dan tantang (VT) diberi vaksin iradiasi lalu ditantang
dengan SGB Hn+; kelompok tantang (T) diinfeksi dengan SGB Hn
+, dan kelompok kontrol (K)
tanpa vaksin dan tanpa tantang. Hasil perhitungan alveol produktif kelenjar mamae mencit
adalah 70,9±13,2; 77,1±13,7; 72,5±17,8; 60,4±14,0, masing-masing untuk kelompok hewan coba
K, V, VT, dan T. Hasil tersebut menunjukkan bahwa SGB iradiasi tidak patogen, terbukti dari
gambaran histopatologi (HP) bahwa sel-sel pada alveol kelenjar mamae dari kelompok yang
diberi vaksin SGB iradiasi (V) dan kelompok vaksin kemudian ditantang dengan SGB tanpa
iradiasi (VT), masih memproduksi susu dan jumlah alveol kelenjar tidak berbeda nyata
dibanding dengan kelompok kontrol (K) atau mencit normal; jumlah alveol kelenjar mamae yang
paling sedikit berproduksi ada pada kelompok mencit yang ditantang dengan SGB ganas tanpa
divaksin (T). Hasil perhitungan sel epitel terdeskuamasi dari kelenjar mamae mencit K=8,8±7,2;
V=8,1±0,7; VT=5,9±2,3; T=9,7±3,1 ; sedangkan perhitungan sel radang interstisial disekitar
kelenjar mamae mencit K=23,0±1,1; V=11,8±6,4; VT=14,5±10,7; T=41±5,9. Hasil pemeriksaan
kedua parameter tersebut menunjukkan bahwa vaksin SGB iradiasi juga tidak menimbulkan
peradangan pada kelenjar mamae mencit yang diuji, dibuktikan dari jumlah sel radang interstisial
disekitar alveol mamae dan sel epitel kelenjar yang terdeskuamasi pada kelompok mencit yang
divaksin (V) tidak berbeda nyata bahkan lebih sedikit dibanding dengan kelompok kontrol (K);
sel radang interstisial terbanyak pada kelompok mencit yang ditantang dengan SGB ganas (T),
bahkan berbeda nyata (p<0,05) dibanding kelompok vaksin (V) dan kontrol (K). Hasil
pemeriksaan Ig-G serum mencit dengan Enzyme Linked Immunosorbent Assay (ELISA)
menunjukkan tingkat respon imun yang lebih tinggi pada kelompok mencit yang divaksin
dibandingkan kelompok kontrol, hal tersebut membuktikan bahwa SGB yang telah dilemahkan
dengan iradiasi sinar gamma masih mampu menimbulkan respon imun. Aplikasi vaksin SGB
iradiasi pada kambing perah peranakan Etawah-pun dapat menimbulkan respon imun yang baik,
terbukti dari hasil pemeriksaan serum dengan ELISA pada kelompok kambing yang divaksin
mempunyai konsentrasi Ig-G lebih tinggi dibanding kelompok kontrol. Produksi susu kambing

perah kelompok vaksin tidak mengalami penurunan sehingga dapat disimpulkan pula bahwa
pemberian vaksin iradiasi SGB tidak mengganggu produksi susu.
Kata-kata kunci: Mastitis subklinis, S. agalactiae group-B, vaksin iradiasi.


© Hak Cipta milik IPB, tahun 2012
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan
atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan,
penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritikan, atau
tinjauan suatu masalah.
Pengutipan tidak merugikan kepentingan yang wajar IPB.
Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis
dalam bentuk apapun tanpa seizin IPB.


KANDIDAT VAKSIN IRADIASI Streptococcus agalactiae UNTUK
PENCEGAHAN MASTITIS SUBKLINIS PADA RUMINANSIA
BOKY JEANNE TUASIKAL
B361070021
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar
Doktor pada Mayor Ilmu Biomedis Hewan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
2012

Penguji Luar Komisi Pembimbing
Ujian Tertutup:
1. Drh. Kamaluddin Zarkasie, Ph.D.
2. Dr. drh. Agustin Indrawati M.Biomed
Ujian Terbuka:
1. Drh. Enuh Rahardjo Djusa, Ph.D.
2. Dr. drh. Koekoeh Santoso

Judul disertasi : Kandidat Vaksin Iradiasi Streptococcus agalactiae untuk
Pencegahan Mastitis Subklinis pada Ruminansia
Nama : Boky Jeanne Tuasikal
NRP : B361070021
Program Studi : Mayor Ilmu Biomedis Hewan
Disetujui
Komisi Pembimbing
Prof. Dr. drh. Fachriyan Hasmi Pasaribu
Ketua
Dr. drh. Sri Estuningsih, M.Si., APVet Prof. Dr. drh. I Wayan T. Wibawan, MS
Anggota Anggota
Diketahui
Ketua Program Studi Dekan Sekolah Pascasarjana IPB
Mayor Ilmu Biomedis Hewan
drh. Agus Setiyono, MS. PhD.APVet Dr. Ir. Dahrul Syah, M.Sc.Agr.
Tanggal Ujian : 27 Juli 2012 Lulus Tanggal : 09 Agustus 2012


KATA PENGANTAR
Penulis memanjatkan puji dan syukur ke Hadirat Allah SWT Yang Maha Kuasa, Maha
Pengasih dan Penyayang, atas berkat rahmat dan karunia-Nya sehingga penulis dapat
menyelesaikan pendidikan pascasarjana program doktor serta melaksanakan tugas penelitian dan
penulisan disertasi ini.
Pada kesempatan yang membahagiakan ini, penulis menyampaikan penghargaan yang
sangat dalam dan hormat, serta rasa terima kasih yang tulus kepada yang terhormat Bapak Prof.
Dr. drh. Fachriyan H. Pasaribu, selaku ketua Komisi Pembimbing yang telah memberikan
bimbingan dan pengarahan sejak penulis mempersiapkan judul disertasi, membuat proposal,
melaksanakan penelitian, dan membuat laporan berupa disertasi ini. Penghargaan yang dalam,
rasa hormat yang tinggi disertai rasa terima kasih yang tulus ditujukan pula kepada Ibu Dr. drh.
Sri Estuningsih, M.Si.APVet. dan Bapak Prof. I Wayan T. Wibawan, MS selaku anggota komisi
pembimbing atas segala bimbingan, arahan, nasihat, dan dorongan semangat yang telah
diberikan dengan penuh tanggung jawab selama penulis belajar, mempersiapkan, dan melakukan
penelitian di Institut Pertanian Bogor hingga penulisan disertasi ini selesai.
Penulis menyampaikan terima kasih dan penghargaan kepada Ketua Mayor Ilmu
Biomedis Hewan, Bapak drh. Agus Setiyono, MS. PhD. APVet. dan Ketua Departemen Klinik,
Reproduksi dan Patologi, Fakultas Kedokteran Hewan IPB, Bapak Prof. drh. Bambang Pontjo
Priosoeryanto, MS, PhD. yang telah berkenan memberikan ijin kepada penulis untuk mengikuti
program doktor pada Mayor Ilmu Biomedis Hewan di Departemen Klinik, Reproduksi dan
Patologi, Fakultas Kedokteran Hewan IPB.
Kepada yang terhormat Bapak Rektor Institut Pertanian Bogor beserta jajarannya,
Direktur Program Pascasarjana beserta staf, penulis menyampaikan rasa hormat dan terima kasih
serta penghargaan atas ijin dan kesempatan yang diberikan untuk mengikuti pendidikan program
doktor di IPB sejak tahun 2007.
Rasa terima kasih penulis sampaikan kepada Kepala Pusat Aplikasi Teknologi Isotop dan
Radiasi, Bapak Dr. Hendig Winarno dan Kepala Bidang Pertanian, Bapak Dr. Sobrizal atas ijin
yang diberikan, serta Prof. Ris. Dr. Ir. Soeranto Human, APU sebagai Pembimbing Karyasiswa.
Tak lupa terima kasih saya kepada para staf peneliti dan teknisi di Kelompok Kesehatan dan
Reproduksi Ternak, Badan Tenaga Nuklir Nasional atas pengertian dan dorongan semangat

maupun bantuan secara moril dan materil yang telah diberikan kepada penulis. Demikian pula
rasa terima kasih penulis sampaikan kepada yang terhormat Dekan Fakultas Kedokteran Hewan
IPB, Bapak drh. Srihadi Agungpriyono, PhD, PAVet., Penanggung jawab Laboratorium Terpadu
Fakultas Kedokteran Hewan IPB, Penanggung jawab Laboratorium Hewan Percobaan Bagian
Patologi Fakultas Kedokteran Hewan IPB, dan Penanggungjawab Laboratorium Mikrobiologi
Medis Fakultas Kedokteran Hewan IPB atas ijin dan kesempatan yang diberikan untuk
menggunakan fasilitas laboratorium selama penulis melakukan penelitian. Terima kasih penulis
sampaikan kepada semua staf pengajar, para teknisi laboratorium di lingkungan Institut Pertanian
Bogor yang telah membantu terselenggaranya penelitian ini.
Penulis sampaikan pula rasa terima kasih dan penghargaan kepada Deputi Bidang
Dinamika Masyarakat selaku penanggungjawab Program Pascasarjana Kementrian Negara Riset
dan Teknologi (KMNRT) yang telah memberikan beasiswa kepada karya siswa untuk
melanjutkan Program Doktor di Pascasarjana IPB.
Kepada ayahanda (alm) dan ibunda serta kakak dan adik-adik tercinta, penulis
menyampaikan rasa terima kasih yang sedalam-dalamnya dan penghargaan yang setinggi-
tingginya atas doa restu, bimbingan, didikan, serta dorongan semangat sejak lahir hingga penulis
mencapai jenjang pendidikan ini.
Terima kasih yang tulus dan penghargaan setinggi-tingginya penulis sampaikan kepada
suami dan anak-anak tercinta yang dengan setia, penuh kasih, sabar, dan penuh pengertian
mendoakan, memberikan dorongan semangat, berkorban, serta mendampingi penulis sehingga
penulis dapat menghargai waktu dan kesempatan yang diberikan dalam menyelesaikan penelitian
dan disertasi ini.
Penulis menyadari, bahwa karya ilmiah ini belum sempurna, oleh sebab itu dengan
rendah hati, penulis mengharapkan kepada seluruh pembaca untuk memberikan saran-saran yang
bermanfaat demi kesempurnaan karya tulis ini sehingga dapat bermanfaat bagi semua pihak yang
membutuhkannya.
Bogor, Agustus 2012
Penulis

RIWAYAT HIDUP
Penulis dilahirkan pada tanggal 13 Agustus 1963 di Jakarta, sebagai anak kedua dari
enam bersaudara dari ayah H. Umar Tuasikal (alm) dan ibu Hj. Syarifah Hadiyati. Penulis
menikah dengan drh. Zainul Zakir pada tahun 1989 dan dikaruniai seorang putra, Muhammad
Hafizh Dewantara, S.Ked. dan seorang putri, Hanifah Septianti.
Setelah lulus SMA Negeri 2 Bandung pada tahun 1982, penulis melanjutkan pendidikan
di Tingkat Persiapan Bersama Institut Pertanian Bogor. Pada tahun 1983, penulis melanjutkan
pendidikan di Fakultas Kedokteran Hewan Institut Pertanian Bogor dan lulus sebagai Sarjana
Kedokteran Hewan pada tahun 1986. Kemudian penulis mengikuti Program Pendidikan Profesi
Dokter Hewan dan lulus sebagai Dokter Hewan pada tahun 1987. Penulis melanjutkan
pendidikan Program Magister pada bidang studi Sains Veteriner Program Pascasarjana IPB dan
lulus pada tahun 2005.
Sejak tahun 1989 hingga sekarang, penulis adalah staf peneliti di Kelompok Kesehatan
dan Reproduksi Ternak, Bidang Pertanian, Pusat Aplikasi Teknologi Isotop dan Radiasi, Badan
Tenaga Nuklir Nasional. Pada tahun 2007, penulis melanjutkan pendidikan program Doktor pada
bidang Ilmu Biomedis Hewan di Sekolah Pascasarjana IPB. Selama studi S3 penulis mendapat
bantuan beasiswa dari Program Pascasarjana Kementrian Negara Riset dan Teknologi
(KMNRT).


DAFTAR ISI
Halaman
DAFTAR TABEL ………………………………………………… xvi
DAFTAR GAMBAR ……………………………………………… xvii
DAFTAR LAMPIRAN …………………………………………… xviii
PENDAHULUAN ………………………………………………… 1
1. Latar Belakang ………………………………………………………….. 1
2. Perumusan Masalah ……………………………………………………... 4
3. Tujuan Penelitian ………………………………………………………... 4
4. Hipotesis ………………………………………………………………… 5
5. Manfaat Penelitian ………………………………………………………. 5
6. Kebaruan ………………………………………………………………... 6
7. Ruang Lingkup Penelitian ………………………………………………. 6
TINJAUAN PUSTAKA ………………………………………… 7
1. Mastitis ………………………………………………………………….. 7
2. Patogenesa Mastitis ……………………………………………………... 8
3. Mastitis Subklinis ……………………………………………………….. 9
4. Streptococcus agalactiae ………………………………………………... 11
5. Sistem Imun ……………………………………………………………... 14
6. Radiasi …………………………………………………………………... 15
7. Vaksin Iradiasi Sinar Gamma …………………………………………... 15
BAHAN DAN METODE ………………………………………… 19
1. Tempat dan Waktu Penelitian …………………………………………... 19
2. Sampel Susu …………………………………………………………….. 19
3. Pemeriksaan Mastitis Subklinis Secara Tidak Langsung ……………….. 19
4. Pemeriksaan Mastitis Subklinis Secara Langsung ……………………… 20
5. Identifikasi dan Isolasi S. agalactiae…………………………………….. 20
5.1. Pemeriksaan Morfologi Koloni Bakteri ……………………………. 21
5.2. Pemeriksaan Mikroskopi …………………………………………… 21

5.3. Uji Katalase ………………………………………………………… 22
5.4. Uji Christie, Atkins and Munch-Petersen (CAMP) ..………………. 22
5.5. Uji Serogrup S. agalactiae …………………………………………. 23
5.6. Ekspresi Fenotipe SGB in vitro …………………………………….. 23
5.7. Uji Hemaglutinasi ………………………………………………….. 24
6. Orientasi Dosis Iradiasi Sinar Gamma SGB untuk LD50 ……………….. 24
7. Karakterisasi Antigenik Permukaan SGB dengan SDS PAGE …………. 25
8. Uji Patogenitas dan Imunitas dengan Hewan Model Mencit …………… 26
9. Aplikasi Kandidat Vaksin Iradiasi Terpilih pada Kambing Perah ……… 27
10. Rancangan Penelitian …………………………………………………… 27
HASIL DAN PEMBAHASAN …………………………………… 29
1. Kejadian Mastitis Subklinis di Daerah Pengambilan Sampel Susu …….. 29
2. Isolat SGB Terpilih sebagai Bahan Vaksin Iradiasi Mastitis Subklinis…. 33
3. Orientasi Dosis Iradiasi SGB untuk LD50 ................................................. 43
4. Hasil Uji Patogenitas dan Imunitas pada Hewan Model Mencit ……….. 45
5. Hasil Uji Respon Imun dan Produksi Susu pada Kambing Perah……….. 51
SIMPULAN DAN SARAN ……………………………………… 55
1. Simpulan ………………………………………………………………… 55
2. Saran …………………………………………………………………….. 55
DAFTAR PUSTAKA …………………………………………… 57
LAMPIRAN ……………………………………………………… 65

xvi
DAFTAR TABEL
Halaman
1. Kejadian MSK pada Wilayah Cisurupan, Bayongbong dan KUNAK…… 30
2. Jumlah Mikroba dalam Sampel Susu dari Uji Total Plate Count (TPC)…. 32
3. Presentasi Kehadiran Streptococcus sp. pada Susu dengan Uji Katalase… 36
4. Jumlah Isolat S. agalactiae dengan uji CAMP …………………………... 36
5. Jumlah Isolat S. agalactiae dari Uji Grouping Kit ………………………. 38
6. Pola Pertumbuhan S. agalactiae pada Media SA, SSA dan THB ……….. 39
7. Pengamatan persen viabilitas SGB akibat iradiasi sinar gamma pada
berbagai dosis untuk LD50 ………………………………………………..
43
8. Hasil perhitungan kelenjar mamae mencit pada empat kelompok
perlakuan ………………………………………………………………….
46


xvii
DAFTAR GAMBAR
Halaman
1. Phenomena “Gunung es” pada mastitis subklinis ………………………… 10
2. Hasil CMT memperlihatkan kekentalan yang terbentuk akibat
pencampuran susu dengan reagen uji ……………………………………...
30
3. Tampilan koloni Streptococcus sp. pada media BAP …………………...... 34
4. Pola hemolisis Streptococcus sp. pada media BAP …………………......... 34
5. Tampilan mikroskopik Streptococcus sp. Gram positif …………………... 35
6. Hasil uji CAMP positif dan negatif ……………………………………...... 37
7. Bentuk koloni S. agalactiae pada media SA dan SSA ………………......... 40
8. Perhitungan Dosis Iradiasi gamma untuk melemahkan SGB
sampai taraf LD50 ………………………………………………………….
43
9. Visualisasi pita protein SGB iradiasi 0 Gy dan 17 Gy
pada BM 75 – 225 kDa…………………………………………………….
44
10. Gambaran Histopatologi Kelenjar Mamae Mencit Kelompok Kontrol…… 47
11. Gambaran Histopatologi Kelenjar Mamae Mencit Kelompok Vaksin……. 47
12. Gambaran Histopatologi Kelenjar Mamae Mencit Kelompok
Vaksin+Tantang …………………………………………………………..
48
13. Gambaran Histopatologi Kelenjar Mamae Mencit Kelompok Tantang…... 48
14. Titer Ig-G kelompok mencit yang divaksin SGB iradiasi
dibanding kelompok mencit kontrol ………………………………………
50
15. Titer Ig-G kelompok vaksin SGB iradiasi kemudian ditantang
dibandingkan dengan kelompok kontrol …………………………………..
50
16. Konsentrasi Ig-G serum kambing yang telah divaksin
dengan SGB iradiasi dibandingkan dengan kontrol ……………………….
51
17. Produksi susu kambing pada kelompok vaksin iradiasi SGB post-partus… 52


xviii
DAFTAR LAMPIRAN
Halaman
1. Identifikasi S. agalactiae Grup-B ………………………………………… 67
2. Karakterisasi SGB tanpa kapsul, keberadaan Antigen Protein dan
Hemaglutinin positif (Hn+) …………………………………………...........
68
3. Pengamatan Pertumbuhan SGB Hn+ ………………………………............ 69
4. Prosedur Orientasi Dosis Iradiasi untuk Penentuan LD50 …………………. 70
5. Iradiator Gamma Chamber dengan Sumber Isotop 60
Co …………………... 71
6. Jadwal Perlakuan dan Parameter Penelitian pada Hewan Model Mencit...... 72
7. Pembuatan Preparat Histopatologi Kelenjar Mamae Mencit ………............ 73
8. Ekstraksi Antigen SGB untuk coating Ag pada ELISA …………………… 75
9. ELISA untuk Titer Antibodi Ig-G …………………………………………. 76


1
PENDAHULUAN
1. Latar Belakang
Usaha peternakan mempunyai potensi untuk dapat berkembang pesat di
Indonesia, mengingat cukupnya ketersediaan pakan dan keragaman jenis ternak yang
ada. Salah satu upaya pemerintah untuk meningkatkan konsumsi protein hewani bagi
penduduk Indonesia adalah dengan mengembangkan peternakan sapi perah, yang
telah dimulai sejak zaman Hindia Belanda hingga saat ini (Subandriyo dan Adiarto
2009). Hal ini terlihat dengan meningkatnya populasi sapi perah dari tahun ke tahun,
yaitu dari 369.008 ekor pada tahun 2006 menjadi 457.577 ekor pada tahun 2008, dan
terus meningkat menjadi 597.129 ekor pada tahun 2011 (Dirjen Peternakan dan
Kesehatan Hewan 2011). Kesadaran masyarakat yang meningkat tentang nilai gizi
serta kebutuhan konsumsi akan protein hewani, khususnya susu juga turut
mendukung berkembangnya usaha peternakan rakyat. Susu merupakan sekresi yang
dihasilkan oleh kelenjar mamae mamalia termasuk manusia, dan merupakan bahan
makanan sempurna serta mempunyai nilai gizi yang tinggi. Bayi membutuhkan susu
sebagai satu-satunya bahan makanan dalam beberapa bulan di awal kehidupannya.
Hingga saat ini susu sapi masih dianggap sebagai sumber makanan utama yang dapat
menggantikan air susu ibu (ASI). Susu sapi merupakan bahan makanan istimewa bagi
manusia karena kelezatan dan komposisinya yang ideal yaitu mengandung banyak zat
yang dibutuhkan oleh tubuh seperti lemak, protein terutama casein, laktosa, vitamin,
mineral seperti kalsium, dan lain-lain (De Laval 2008, Mirdhayati dkk. 2008).
Konsumsi susu penduduk Indonesia semakin meningkat dari tahun ke tahun, yaitu
6,50 kg/ kapita/ tahun pada tahun 2004 menjadi 16,42 kg/ kapita/ tahun pada tahun
2010. Meningkatnya konsumsi susu tentu saja diikuti oleh peningkatan permintaan,
tetapi pemenuhan permintaan susu dari produksi dalam negeri hanya dapat memasok
23% saja, dan selebihnya berasal dari substitusi impor (Dirjen Peternakan dan
Kesehatan Hewan 2011). Hal tersebut menunjukkan bahwa produksi susu di
Indonesia masih kurang.

2
Kendala yang sering menghambat keberhasilan suatu usaha peternakan adalah
serangan penyakit. Salah satu penyakit penting yang dapat menurunkan produksi susu
pada ternak sapi perah yaitu Mastitis yang dikenal juga sebagai penyakit radang
kelenjar air susu (mamae). Penurunan produksi susu merupakan masalah bagi
peternak sapi perah Indonesia yang umumnya hanya memelihara 3-4 ekor sapi tiap
peternak. Sebagian besar peneliti setuju bahwa kerugian ekonomi akibat mastitis pada
usaha sapi perah minimal sebanyak 70% yang diakibatkan oleh penurunan produksi
susu dan dibuangnya susu yang berasal dari sapi mastitis (Bogni et al. 2011).
Penyakit mastitis secara garis besar dibagi menjadi 2 tipe, pertama yaitu mastitis
klinis yang menunjukkan gejala kesakitan pada mamae ternak maupun kerusakan
pada susu, dan kedua adalah mastitis subklinis yang tidak menampakkan gejala
peradangan pada mamae kecuali bila dilakukan pemeriksaan pada air susu dengan uji
khusus contohnya California Mastitis Test (CMT) (Dirkeswan. 2001). Sebagaimana
negara-negara lain, di Indonesia-pun kasus yang lebih banyak terjadi yaitu mastitis
subklinis (Supar 1997, Subronto 2003, Hashemi et al. 2011). Kasus mastitis terutama
mastitis subklinis di Indonesia sampai akhir tahun 2006, tercatat sekitar 75% - 83%
(Sudarwanto dkk. 2006).
Mastitis dapat disebabkan oleh beberapa faktor, yakni cara pemerahan yang
salah, sanitasi yang buruk, kandang yang kurang bersih dan lantai kandang yang tidak
memenuhi persyaratan untuk sapi perah. Penyebaran penyakit ini dapat melalui
pemerahan yang tidak mengindahkan kebersihan, alat pemerahan, kain pembersih
puting dan pencemaran dari lingkungan. Kuman penyebab mastitis dari kandang yang
kotor akan masuk ke dalam mamae melalui lubang dan kanal puting saat pemerahan
atau ketika sapi duduk di lantai kandang. Kuman yang potensial menyebabkan
mastitis adalah Streptococcus agalactiae, Streptococcus digalactiae, Steptococcus
uberis dan Staphylococcus aures (Dirkeswan. 2001). Streptococcus agalactiae
adalah salah satu agen utama penyebab mastitis subklinis pada sapi perah. Hasil studi
Estuningsih dkk. (2002) menyebutkan bahwa 83 isolat bakteri yang diisolasi dari 3
area peternakan sapi perah di pulau Jawa, seluruhnya teridentifikasi sebagai
Streptococcus agalactiae. S. agalactiae memiliki nama lain yaitu Streptococcus

3
Grup- B (SGB). S. agalactiae lebih banyak dikenal di dalam dunia kedokteran hewan
sedangkan SGB populer di kedokteran manusia. Diketahui pula bahwa S. agalactiae
grup B (SGB) merupakan bakteri utama penyebab mastitis subklinis pada sapi perah
dan merupakan parasit obligat pada mamae (Wahyuni dkk. 2006). S. agalactiae juga
dilaporkan dapat menginfeksi anjing, babi, kelinci, kuda, dan merpati. Bakteri ini
menyebabkan abortus dan servitis pada kuda dan babi, bahkan dapat menyerang ikan
(Pasaribu et al. 1993, Evans et al. 2006).
Berbagai macam antibiotik untuk mengatasi permasalahan mastitis telah
banyak digunakan seperti penisilin, streptomisin, ampisilin, kloksasilin, dan lain
sebagainya. Pengobatan mastitis dengan menggunakan antibiotik terutama ditujukan
untuk membunuh bakteri penyebab mastitis, namun dengan banyaknya macam obat
antibiotik yang dipergunakan dan cara pemberian dosis yang tidak terkontrol, maka
dikhawatirkan menimbulkan permasalahan baru berupa resistensi kuman penyebab
mastitis (Wu et al. 2007) dan terdapat residu obat pada susu yang dihasilkan. Hasil
penelitian Sudarwanto dkk. (1992) menunjukkan 32,52% susu pasteurisasi dan
31,10% susu segar di wilayah Jakarta, Bogor dan Bandung mengandung residu
antibiotik dalam jumlah yang cukup tinggi. Keberadaan residu antibiotik pada susu
yang dikonsumsi dapat menyebabkan terjadinya reaksi alergis dan gangguan terhadap
mikroflora saluran pencernaan manusia yang mengkonsumsi susu tersebut. Susu yang
akan diekspor-pun diharuskan bebas dari residu antibiotik oleh negara pengimpor.
Penggunaan dan pemilihan antibiotik yang tidak tepat, ditinjau dari aspek indikasi,
dosis, waktu dimulainya dan lamanya terapi, serta penggunaan kombinasi antibiotik
dapat mempercepat terjadinya resistensi bakteri. Mastitis subklinis yang disebabkan
oleh bakteri Gram posisif semakin sulit ditangani dengan antibiotik karena bakteri ini
sudah banyak yang resisten terhadap berbagai jenis antibiotik (Wibawan 1998,
Maricato et al. 2005).
Peternak sudah mengetahui cara pencegahan penyakit mastitis, yaitu sistem
pemeliharaan dan manajemen pemerahan yang baik, termasuk melakukan teat
dipping pada sapi-sapi setelah pemerahan. Namun demikian kasus mastitis subklinis
masih saja banyak terjadi di Indonesia. Keberadaan masalah resistensi bakteri

4
terhadap antibiotik dan residunya pada susu, serta masih banyaknya kejadian mastitis
subklinis, maka menjadikan perlu untuk mencari alternatif lain dalam pencegahan
penyakit ini, salah satunya dengan pembuatan vaksin. Pemanfaatan iradiasi dalam
pembuatan vaksin dimaksudkan untuk melemahkan patogenitas S. agalactiae sebagai
bakteri dominan penyebab mastitis, namun bakteri ini diharapkan masih dapat
menimbulkan respon tanggap kebal sebagai bahan vaksin iradiasi. Sumber-sumber
sinar gamma yang digunakan untuk iradiasi kandidat vaksin dihasilkan dari
radioisotop 60
Co. Penggunaan sinar gamma memiliki keunggulan dibandingkan
dengan teknik konvensional seperti pemanasan atau penggunaan zat kimia. Sinar
gamma yang menembus tidak merusak dinding sel bakteri tempat protein antigenik
sehingga mampu mempertahankan imunogenitasnya (Kochman 2006). Sel yang
terpapar radiasi gamma akan kehilangan kemampuan replikasinya di dalam tubuh
inang, namun tidak menghentikan aktivitas metaboliknya. Sel tetap mempunyai
metabolik aktif, sehingga mampu menghasilkan senyawa imunogenik untuk
menstimulasi respon imun protektif pada inang (Sanakkayala 2005).
2. Perumusan Masalah
Berbagai aspek mengenai mastitis secara umum telah banyak diteliti tetapi
belum ada cara pencegahan mastitis yang efektif diaplikasikan pada ternak sapi
perah, sehingga kasus mastitis subklinis yang merugikan masih tetap terjadi di
Indonesia. Penelitian ini mencoba turut berkontribusi dalam penyelesaian masalah
tersebut dengan pembuatan bahan vaksin iradiasi dari bakteri terpilih S. agalactiae
untuk pencegahan mastitis subklinis pada ternak ruminansia.
3. Tujuan Penelitian
Penelitian ini bertujuan untuk mendapatkan kandidat vaksin guna pencegahan
mastitis subklinis pada ruminansia dengan memanfaatkan teknik radiasi terhadap
bakteri dominan penyebab mastitis subklinis S. agalactiae.
Tujuan ini dicapai dengan cara:

5
1. Mengisolasi S. agalactiae sebagai bakteri terpilih dengan sifat-sifat atau
karakter yang cocok untuk bahan vaksin iradiasi mastitis subklinis.
2. Orientasi dosis iradiasi sinar gamma untuk melemahkan S. agalactiae Group-
B sampai taraf lethal dose 50% (LD50).
3. Menguji pengaruh iradiasi SGB dengan mempelajari patogenitasnya pada
mencit.
4. Menguji imunitas hewan coba mencit dan kambing perah setelah divaksin
dengan SGB iradiasi.
4. Hipotesis Penelitian
Hipotesis yang diajukan dalam penelitian ini adalah sebagai berikut:
1. Diperoleh isolat lokal S. agalactiae dari kasus mastitis subklinis sebagai
bakteri terpilih untuk bahan vaksin iradiasi MSK.
2. Iradiasi sinar gamma dapat melemahkan bakteri S. agalactiae Group-B
sampai taraf LD50.
3. Kandidat vaksin iradiasi SGB tidak bersifat patogen pada hewan coba mencit
namun menimbulkan kekebalan.
4. SGB iradiasi mampu menginduksi timbulnya respon imun pada hewan coba
mencit dan kambing perah.
5. Manfaat Penelitian
Manfaat yang diharapkan adalah sebagai berikut:
1. Penelitian ini dapat memperoleh kandidat vaksin SGB terpilih dengan
karakter yang cocok sebagai bahan vaksin mastitis subklinis.
2. Teknik nuklir iradiasi sinar gamma dapat bermanfaat untuk melemahkan dan
menghilangkan patogenitas SGB tetapi masih mampu menimbulkan respon
imun pada hewan percobaan yang digunakan.
3. Hasil akhir penelitian ini diharapkan dapat bermanfaat untuk pencegahan
penyakit mastitis subklinis pada ternak perah, sehingga dapat meningkatkan

6
produksi susu sebagai sumber protein hewani dalam mendukung ketahanan
pangan di Indonesia.
6. Kebaruan
Kebaruan dari penelitian dalam disertasi ini adalah aplikasi teknik nuklir
untuk vaksin iradiasi S. agalactiae dalam pencegahan mastitis subklinis.
Pemanfaatan iradiasi sinar gamma dimaksudkan untuk mengatenuasi bakteri S.
agalactiae Group-B sebagai bakteri dominan penyebab mastitis subklinis, namun
kandidat vaksin ini diharapkan masih dapat menimbulkan respon imun pada
ternak perah.
7. Ruang Lingkup Penelitian
Ruang lingkup penelitian terdiri dari empat tahap. Pertama dilakukan isolasi
dan identifikasi isolat S. agalactiae yang termasuk ke dalam group-B (SGB) yang
tidak berkapsul dan memiliki sifat Hemaglutinin (Hn+). Bakteri terpilih diperoleh
dari sampel susu sapi perah yang mengalami mastitis subklinis dengan uji tapis
CMT. Tahap kedua yaitu atenuasi SGB terpilih dengan teknik iradiasi sinar
gamma dari sumber isotop Cobalt-60 (60
Co). Orientasi dosis iradiasi dilakukan
untuk melemahkan bakteri SGB sampai taraf Lethal dose 50% (LD50) dalam
pembuatan vaksin hidup. Karakterisasi Protein permukaan SGB iradiasi ini
menggunakan metode SDS-PAGE untuk mengetahui berat molekul (BM) yang
masih bersifat antigenik pada kandidat vaksin iradiasi SGB. Tahap ketiga adalah
uji patogenitas vaksin iradiasi SGB dan respon imun pada hewan coba mencit.
Tahap keempat merupakan aplikasi kandidat vaksin iradiasi SGB pada kambing
perah untuk uji respon imun dan pengaruhnya terhadap produksi susu.

7
TINJAUAN PUSTAKA
1.Mastitis
Mastitis merupakan peradangan di dalam parenkim kelenjar mamae, yang
dapat menurunkan produksi dan kualitas susu. Penyakit ini tersebar luas pada
peternakan perah di Indonesia maupun di berbagai belahan dunia, sehingga mastitis
menjadi penyakit utama pada sapi perah dan sangat merugikan (Supar dan Ariyanti
2008, Rinaldi et al. 2010).
Mastitis dapat disebabkan oleh berbagai hal, mulai dari trauma mekanis pada
kelenjar mamae atau puting susu, akibat mikroorganisme patogen, hingga kekurangan
unsur nutrisi pada pakan ternak. Sebagian besar kasus mastitis disebabkan oleh
mikroorganisme patogen, diantaranya Streptococcus agalactiae, S. disgalactiae, S.
uberis, S. zooepidemicus, Staphylococcus aureus, Escherichia coli, Enterobacter
aerogenes, Pseudomonas aeruginosa (Dirkeswan 2001).
Berdasarkan perubahan patologi anatomi kelenjar mamae, mastitis dapat
dibedakan menjadi dua tipe, yaitu mastitis klinis (MK) dan mastitis subklinis (MSK)
(Hashemi et al. 20011). Perubahan patologi mastitis klinis dapat jelas digambarkan
dari perubahan patologi anatomis kelenjar mamae dan air susu. Kelenjar mamae
membengkak edematus berisi cairan eksudat, disertai tanda-tanda peradangan lainnya
(kemerahan, terasa panas, kesakitan dan penurunan fungsi). Perubahan penting yang
terjadi di dalam susu mencakup perubahan warna, konsistensi dan penurunan jumlah
produksi susu.
Mastitis subklinis tidak menampakkan perubahan patologi anatomis, sehingga
perlu pemeriksaan khusus. Diagnosa mastitis subklinis hingga sekarang dilakukan
berdasarkan pada pemeriksaan sampel susu serta perubahan komposisi susu untuk
mengenali tanda peradangannya (Malinowski et al. 2006, Mirdhayati dkk. 2008).

8
2. Patogenesa Mastitis
Mastitis merupakan peradangan internal mamae yang sebagian besar
disebabkan oleh mikroorganisme (80% disebabkan oleh bakteri) terutama pada kasus
mastitis subklinis (Lukman et al. 2009). Menurut Jayarao (2006) ada 3 faktor utama
yang berperan dalam terjadinya mastitis yaitu kondisi hewan/ternak, kondisi
lingkungan dan agen penyebab.
Duval (1997) menjelaskan bahwa proses infeksi pada mastitis terjadi melalui
beberapa tahap, yaitu adanya kontak dengan mikroorganisme dimana sejumlah
mikroorganisme mengalami multiplikasi di sekitar lubang puting (sphincter),
kemudian dilanjutkan dengan masuknya mikroorganisme akibat lubang puting yang
terbuka ataupun karena adanya luka. Tahap berikutnya, terjadi respon imun pada
induk semang. Respon pertahanan pertama ditandai dengan berkumpulnya leukosit-
leukosit untuk mengeliminasi mikroorganisme yang telah menempel pada sel-sel
mamae. Apabila respon ini gagal, maka mikroorganisme akan mengalami
multiplikasi dan sapi dapat memperlihatkan respon yang lain, misalnya demam. Hal
ini kembali pada daya tahan tubuh ternak. Ternak yang dipengaruhi kondisi
lingkungan yang buruk dan berkembangnya mikroorganisme akan memudahkan
terjadinya mastitis.
Penularan mastitis dari mamae yang terinfeksi ke mamae yang sehat dapat
terjadi melalui peralatan pemerahan yang kotor, higiene personal yang buruk dari
pemerah dan urutan pemerahan yang salah (Jayarao, 2006).
Sori et al (2005) menyatakan bahwa saat periode kering kandang merupakan
masa awal kuman penyebab mastitis menginfeksi, karena pada saat itu terjadi
hambatan aksi fagositosis dari neutrofil pada mamae. Proses mastitis hampir selalu
dimulai dengan masuknya mikroorganisme ke dalam kelenjar melalui lubang puting
(sphincter puting). Sphincter puting berfungsi untuk menahan infeksi kuman. Pada
dasarnya, kelenjar mamae atau mammae sudah dilengkapi perangkat pertahanan,
sehingga air susu tetap steril. Namun tingkat pertahanan kelenjar mammae mencapai
titik terendah saat sesudah pemerahan, karena sphincter masih terbuka beberapa
saat, sedangkan sel darah putih, antibodi serta enzim juga habis karena ikut terperah.

9
Disamping faktor-faktor mikroorganisme yang terdiri dari berbagai jenis,
jumlah dan virulensinya, faktor ternak dan lingkungannya juga menentukan mudah
tidaknya terjadi radang mamae pada sapi perah. Faktor predisposisi radang mamae
dilihat dari segi ternak, meliputi : bentuk mamae, misalnya mamae yang sangat
menggantung, atau mamae dengan lubang puting terlalu lebar (Subronto 2003).
Demikian pula bentuk puting, ada dan tidaknya lesi pada puting mempengaruhi
kejadian mastitis. Hasil penelitian Sori et al. (2005) menunjukkan bahwa prevalensi
mastitis pada puting pendulous mencapai 77,78%, sedangkan pada puting non
pendulous mencapai 50%. Puting yang mengalami lesi memungkinkan prevalensi
mastitis sebesar 84%, sedangkan pada puting normal sebesar 47,74%.
Letak kuartir juga mempengaruhi kejadian mastitis. Kuartir kiri belakang dan
kanan depan lebih sering mengalami mastitis daripada kedua puting lainnya. Pada
kiri belakang, mastitis mencapai 34,3%, sedangkan kanan depan mencapai 30,06%
(Sori et al. 2005).
Faktor umur dan tingkat produksi susu sapi juga mempengaruhi kejadian
mastitis. Semakin tua umur sapi dan semakin tinggi produksi susu, maka semakin
mengendur pula spinchter putingnya. Puting dengan spincter yang kendur
memungkinkan sapi mudah terinfekesi oleh mikroorganisme, karena fungsi spinchter
adalah menahan infeksi mikroorganisme. Semakin tinggi produksi susu seekor sapi
betina, maka semakin lama waktu yang diperlukan spinchter untuk menutup
sempurna (Subronto 2003).
Faktor lingkungan dan pengelolaan peternakan yang banyak mempengaruhi
terjadinya radang mamae meliputi : pakan, perkandangan, banyaknya sapi dalam satu
kandang, ventilasi, sanitasi kandang dan cara pemerahan susu. Pada ventilasi yang
buruk, mastitis mencapai 87,5%, sedangkan ventilasi yang baik mencapai 49,39%
(Sori et al. 2005).
3. Mastitis Subklinis
Definisi mastitis subklinis menurut International Dairy Federation (IDF)
adalah mastitis yang ditandai dengan peningkatan jumlah sel somatik (> 400.000/ml

10
susu) dan ditemukan bakteri patogen pada susu yang berasal dari kwartir dalam masa
laktasi normal. Kejadian mastitis subklinis diperkirakan 20-50 kali kejadian mastitis
klinis. Hal ini sering digambarkan sebagai Fenomena Gunung Es atau Iceberg
Phenomena (Lukman et al. 2009).
Gambar 1. Phenomena “Gunung es” pada mastitis subklinis (Hurley and Morrin 2000).
Kejadian mastitis subklinis pada sapi perah di Indonesia sangat tinggi (97-98%) dan
menimbulkan banyak kerugian (Sudarwanto, 1999). Pada kejadian MSK tersebut
S.agalactiae merupakan agen penyebab yang sering ditemukan (Pasaribu et al. 1994). Hal
yang sama diiungkapkan Supar (1997) yang telah melakukan penelitian intensif pada
beberapa tempat peternakan sapi perah di Jawa Barat, Jawa Tengah, Jawa Timur dan Daerah
Istimewa Yogyakarta. Diketahui prevalensi mastitis subklinis berkisar antara 37% sampai
67%, sedangkan mastitis klinis lebih rendah yaitu 5% sampai 30%. Survei yang dilakukan
oleh Supar dan Ariyanti (2008) pada mastitis subklinis juga didominasi oleh bakteri S.
agalactiae yaitu sebanyak 60,6%. Diketahui pula bahwa S. agalactiae Group B merupakan
agen kontagius patogen yang menyebabkan mastitis subklinis pada sapi perah secara umum
di dunia (Bogni et al. 2011).

11
4. Streptococcus agalactiae
S. agalactiae merupakan jenis bakteri yang termasuk ke dalam famili
Streptococcaceae dari ordo Lactobacillales. Bakteri ini berbentuk bulat (coccus) dan
tersusun seperti rantai yang panjangnya bervariasi, diameter selnya sekitar 0,6-1,2
µm. Sifatnya Gram positif yang terlihat berwarna keunguan pada pewarnaan Gram,
non motil, tidak membentuk spora, dan dapat membentuk kapsul. Sifat lainnya
adalah katalase negatif, anaerob fakultatif, oksidase negatif, dapat memfermentasikan
beberapa jenis karbohidrat namun tidak menghasilkan gas (Carter 1984, Quinn et al.
2006).
Tahun 1896 Lehman dan Naumann mengklasifikasikan bakteri ini
berdasarkan taksonomi sebagai berikut (Bruckner and Colonna 1997):
Kingdom : Bacteria
Filum : Firmicutes
Kelas : Bacilli
Ordo : Lactobacillales
Famili : Streptococcaceae
Genus : Streptococcus
Spesies : S. agalactiae
Biakan S. agalactiae yang ditumbuhkan pada media agar darah terlihat
membentuk koloni-koloni halus, basah, konveks, terang tembus, dan menghemolisis
sel darah merah. Hemolisis yang ditimbulkan berupa β-hemolisis, dan beberapa tidak
menghemolisis (Daignault 2003). Strain yang menghemolisis darah hanya
mengasilkan zona hemolitik tidak lebih dari 1 mm pada agar darah, sedangkan yang
tidak menghemolisis dapat menghasilkan pigmen bewarna kehijauan pada media agar
darah.
Secara umum Streptococcus sp memiliki antigen terstruktur pada
permukaannya seperti kapsul, dinding sel, dan antigen protein. Keberadaan antigen
polisakarida pada dinding sel dijadikan sebagai dasar pengklasifikasian genus
Streptococcus sp. kedalam grup-grup. Penentuan grup-grup tersebut berdasarkan

12
serologis dari antigen polisakarida yang ditemukan oleh Lancefield pada 1933.
Terdapat sekitar 20 grup Streptococcus sp sampai saat ini yang dinamai sesuai abjad
mulai dari A-V; S. agalactiae dikelompokan kedalam grup B oleh Lancefield.
Pengelompokan ini didasari oleh substansi C polisakarida spesifik yang terdapat
pada diding sel (Cowan 2003). Secara serologis Grup ini dikelompokan lagi kedalam
serotipe-serotipe. Serotipe tersebut berdasarkan antigen polisakarida dan antigen
protein yang dihasilkan. Beberapa serotipe tersebut terdiri dari 9 serotipe yang
berbeda antara lain 1a, 1b, II, III, IV, V, VI, VII, VIII dari antigen polisakarida dan
c, R dan X berdasarkan antigen proteinnya (Henrichsen et al. 1984). Wibawan dan
Lammler (1991) menyatakan isolat S. agalactiae dapat memiliki serotipe dengan
antigen poliskarida dalam bentuk kombinasi atau hanya berdiri sendiri tanpa antigen
protein, misalnya Ia/c, II/X. Ada juga isolat yang belum bisa diklasifikasikan kedalam
serotipe yang ada disebut sebagai nontypeable (NT). Antigen protein X dominan
ditemukan pada S. agalactiae yang diisolasi dari sapi sedangkan antigen protein R
lebih banyak ditemukan pada S. agalactiae yang berasal dari manusia. Kedua antigen
ini erat hubungannya dengan patogenitas bakteri terhadap inangnya (Wibawan 1990).
Wahyuni et al. (2006) menyimpulkan dari beberapa tempat di pulau jawa S.
agalactiae yang berasal dari mamae yang menderita masititis subklinis ditemukan
sebanyak 52% didominasi oleh S. agalactiae serotipe NT dan protein antigen yang
muncul paling banyak adalah antigen protein X.
Keberhasilan bakteri melakukan infeksi terhadap sel inangnya ditentukan oleh
adanya faktor virulensi. Bakteri akan berpenetrasi dengan menggunakan faktor
virulrensi untuk mempengaruhi targetnya. Faktor virulensi yang penting bagi bakteri
antara lain adalah kapsul. Kapsul bakteri tersusun oleh asam hyaluronat seperti yang
ditemukan pada struktur dasar jaringan ikat pada mamalia. Kapsul polisakarida
sebagai antigen permukaan dan asam hyluronat sebagai produk ektraseluler akan
meningkatan kemampuan virulensi bakteri (Hayati dan Karmil 2009). Terhambatnya
kerja komplemen oleh kapsul S. agalactiae akan menghambat aktivitas fagositosis
oleh makrofag sehingga tidak dapat menfagosit bakteri. Peranan kapsul juga sebagai
antifagositik dan antigen yang tidak imunogenik.

13
Virulensi dari Streptococcus sp dipengaruhi oleh pemukaan sel, sekresi
protein yang dihasilkan dan strukturnya yang secara langsung atau tidak langsung
dapat menghambat fagositosis, terlibat dalam proses adhesi dan mengakibatkan
pelepasan sitokin proinflamasi (Timoney and Kumar 2010).
Streptococcus juga memiliki komponen nonstruktural seperti protein antigen.
Protein tersebut dihasilkan selama masa perkembangbiakan dan membantu selama
proses infeksi. Protein ini berupa enzim ekstraseluler hasil metabolisme seperti
hemolisin, streptokinase, nuklease, protease, dan hyaluronidase. Hemaglutinin
sebagai adhesin pada permukaan bakteri berperan mempengaruhi tingkat virulensi
(Wahyuni dkk. 2005).
Adhesi adalah kemampuan menempelnya bakteri pada permukaan mukosa
yang bersifat irreversibel dan stabil. Kemampuan bakteri untuk menempel (adhesi)
pada sel inang diperantarai oleh komponen adesin bakteri yang membantu perlekatan
bakteri pada reseptor spesifik dari sel inang. Sifat hidrofobitas permukaan bakteri
berperan dalam mekanisme perlekatan dengan sel inang. Sifat ini dipengaruhi oleh
jumlah protein permukaan. Semakin hidrofobik permukaan sel maka akan semakin
tinggi kemampuan adhesi pada sel inang (Lämmler et al. 1998).
Derajat hidrofobitas permukaan S. agalactiae dapat ditentukan secara
langsung dengan melihat pertumbuhan koloni pada media padat, cair dan agar lunak
(soft agar). Koloni yang tumbuh dengan permukaan yang kasar pada media agar dan
kompak pada agar lunak merupakan bakteri yang bersifat hidrofob, sedangkan bakteri
yang tumbuh difus pada media agar lunak dan permukaan mukoid pada media padat
menunjukan sifat yang hidrofil (Wibawan dan Lämmler 1992).
Hasil kajian Wibawan et al. (1993) menyebutkan bahwa S. agalactiae yang
memilki hemaglutinin mempunyai kamampuan adhesi yang lebih besar daripada yang
tidak mempunyai hemaglutinin pada sel epitel mamae. Kemampuan menempel
bakteri tampaknya lebih penting dari pada kemampuan invasi bakteri ke dalam
jaringan dalam mekanisme infeksi, sehingga tidak dijumpai perubahan yang berarti
pada jaringan mamae (Wibawan et al. 1998). Keberadaan protein hemaglutinin
dengan kemampuan adhesi sel bakteri MSK pada sel epitel mamae mencit memiliki

14
keterkaitan, oleh karena itu ada peluang untuk pembuatan suatu produk pencegahan
mastitis yang cukup besar (Dian dkk. 2010).
5.Sistem Imun
Manusia dan hewan multiseluler memiliki suatu sistem imun yang melindungi
dirinya terhadap unsur-unsur patogen yang masuk ke dalam tubuh. Respon imun
tubuh terhadap unsur-unsur patogen sangat bergantung pada kemampuan sistem imun
untuk mengenal molekul-molekul asing atau antigen yang terdapat pada permukaan
unsur patogen, dan kemampuan dalam melakukan reaksi yang tepat untuk
menyingkirkan antigen. Kemampuan ini dimiliki oleh komponen-komponen sistem
imun yang terdapat dalam jaringan limforetikuler yang letaknya tersebar di seluruh
tubuh, misalnya di dalam sumsum tulang, kelenjar limfe, thymus, sistem saluran
nafas, saluran cerna, peredaran darah dan organ-organ lain. Bila antigen masuk ke
dalam tubuh, maka dapat terjadi dua macam reaksi imun yang berlainan, yaitu imun
humoral dan imun seluler (Baratawijaya dan Rengganis 2009). Dalam reaksi imun
humoral terjadi sintesa dan masuknya antibodi berupa Imunoglobulin (Ig) ke dalam
aliran darah dan cairan tubuh lainnya (antibodi humoral). Antibodi ini akan mengikat
dan menetralisir antigen, misalnya toksin kuman atau dapat membungkus kuman
untuk persiapan fagositosis. Bila kita menyuntikkan antigen misalnya vaksin ke tubuh
hewan percobaan, maka setelah beberapa hari sampai seminggu dapat ditemukan
antibodi di dalam darah. Sel utama dalam hal ini adalah Sel-B yang diproduksi oleh
sumsum tulang. Reaksi imun seluler ditengahi oleh sel-sel limfosit dan tidak
tergantung pada antibodi. Seri reaksi yang terlibat dalam jenis imunitas ini dikaitkan
dengan reaksi-reaksi sel sasaran efektor yang terlibat, terutama yang berkaitan dengan
penolakan tumor. Pada masing-masing keadaan ini, antigen berada intraseluler atau
karena tempatnya tidak dapat dicapai, sehingga reaksi antigen-antibodi tampak secara
relatif tidak efisien. Sel utama yang berperan dalam hal ini adalah Sel-T yang
dihasilkan oleh Thymus.
Pada masa-masa akhir kebuntingan hewan, antibodi dari darah dapat dikeluarkan
dan masuk ke dalam kelenjar mamae berupa kolostrum susu. Imunoglobulin yang

15
paling banyak dalam kolostrum dari semua hewan piara adalah Ig G, yang meliputi
65-95% dari semua imunoglobulin yang ada. Sementara laktasi berkembang,
kolostrum berubah menjadi susu, perbedaan terjadi diantara jenis hewan. Pada
primata dan manusia, Ig A merupakan bagian terbanyak baik dalam kolostrum
maupun susu. Pada ruminansia, Ig G1 merupakan imunoglobulin terbanyak baik
dalam kolostrum maupun dalam susu (Tizard 1988).
6. Radiasi
Radiasi merupakan energi yang dipancarkan dalam bentuk partikel atau
gelombang. Radiasi diklasifikasikan sebagai radiasi non pengion (energi rendah) atau
pengion (energi tinggi). Jenis radiasi non pengion seperti sinar ultraviolet, cahaya
tampak, radiasi inframerah, radiasi frekuensi radio dan gelombang mikro. Radiasi
pengion dari matahari (sinar kosmik), bahan radioaktif, dan mesin X-ray. Empat jenis
utama dari radiasi pengion (1) alpha partikel. bermuatan positif terdiri dari dua
neutron dan dua proton, relatif berat dan bergerak lambat dari emisi radioaktif
lainnya, (2) partikel beta merupakan partikel bermuatan negatif terdiri dari elektron
dan lebih ringan serta cepat dibandingkan partikel alpha, (3) sinar gamma adalah
radiasi gelombang elektromagnetik pendek yang dipancarkan oleh peluruhan
radioaktif yang dipancarkan oleh atom dengan kecepatan tinggi dan mempunyai
penetrasi yang tinggi pula, dan (4) sinar X mempunyai energi lebih rendah dan
kemampuan penetrasinya kurang daripada sinar gamma. Sinar X yang dipancarkan
berasal dari proses luar inti, sementara sinar gamma berasal dari inti (Soeminto
1989).
7. Vaksin Iradiasi Sinar Gamma
Sinar gamma berupa radiasi elektromagnetik panjang gelombang pendek
yang dipancarkan oleh isotop radioaktif sebagai inti tidak stabil dan meluruh untuk
mencapai bentuk stabil. DNA merupakan target selular yang mengatur hilangnya
viabilitas setelah terpapar sinar gamma. Suatu materi hidup seperti sel, bila terkena
sinar gamma akan mengalami kerusakan secara langsung atau tidak langsung. Efek

16
langsung adalah terjadinya pemutusan ikatan senyawa-senyawa penyusun sel. Efek
tidak langsung terjadi karena materi sel terbanyak adalah air yang apabila terkena
sinar gamma akan mengalami hidrolisis dan menghasilkan radikal bebas. Radikal
bebaslah yang akan menyebabkan kerusakan materi sel. Target utama bagian sel
adalah DNA yang merupakan sumber informasi genetik sel. Perubahan genetik sel
akan berakibat pada terganggunya kinerja atau kematian sel (Alatas 2007).
Pemanfaatan sinar gamma umumnya digunakan untuk sterilisasi alat-alat
medis dari cemaran bakteri (Trampuz et al. 2006), pengawetan makanan dan
pengolahan jaringan allografts dan komponen darah, dan menghindarkan kebutuhan
suhu tinggi yang dapat merusak suatu produk. Selain itu sinar gamma juga digunakan
untuk membuat vaksin yang lebih efektif daripada pemanasan atau inaktivasi
kimiawi.
Penelitian penggunaan vaksin yang dilemahkan dengan memberikan paparan
radiasi pengion terhadap suspensi mikroorganisme dilakukan pada metaserkaria
iradiasi yang mampu mengurangi jumlah Fasciola hepatica pada anak sapi,
penggunaan radiasi sinar X pada larva stadium kedua yang menetas dari telur cacing
paru Dictyocaulus viviparus penyebab pneumoni verminosa (Tizard 1988).
Penelitian vaksin iradiasi gamma untuk Venezuelan equine encephalitis
(VEE) telah menginduksi dengan tinggi serum netralisasi dan antibodi hemaglutinin-
inhibisi pada marmut dan kelinci. Percobaan vaksin iradiasi ini pada kultur jaringan
menyebabkan sel-sel dapat bertahan hidup sampai taraf Lethal dose-50 (LD50)
terhadap virus VEE tantangan (Reitman et al. 1970).
Vaksin lain dengan teknik iradiasi yaitu Vaksin bakteri Listeria
monocytogenes iradiasi yang dapat meningkatkan respon imun dibandingkan teknik
konvensional. Bakteri diiradiasi mendorong respon pelindung dari sel sistem
kekebalan sel T (Kochman 2006). Sinar gamma juga digunakan untuk melemahkan
Plasmodium (WHO 2001).
Tanggap kebal sapi terhadap Fasciolosis akibat inokulasi metaserkaria
Fasciola gigantica iradiasi sinar gamma dosis 45 Gy telah diteliti oleh Arifin (2006).
Hasil analisis ELISA dari penelitian tersebut menunjukkan titer Ig-G lebih tinggi

17
pada sapi yang diinokulasi metaserkaria F. gigantica iradiasi dibandingkan dengan
sapi kontrol. Sinar gamma yang dimanfaatan untuk inaktivasi yaitu radiasi
elektromagnetik panjang gelombang pendek yang memiliki kemampuan penetrasi
tinggi dan memiliki karakteristik tanpa memberikan radiaoaktivitas pada materi yang
terpapar (Hall & Giaccia 2006).
Prospek vaksin iradiasi gamma untuk influenza sangat baik karena vaksin
inaktif ini dapat diproduksi dengan aman. Inaktifasi vaksin dengan dosis 10 kGy
mempunyai potensi respon imun perlindungan silang dalam melawan heterotipe virus
tantangan. Iradiasi gamma ini juga mempunyai kemampuan tinggi dalam inaktifasi
virus-virus dengan keuntungan yaitu perubahan yang sangat minim pada protein
molekuler dan struktur virus. Dengan adanya perlindungan silang, maka produksi
vaksin bisa lebih murah. (Alsharifi dan Mullbacher 2009).
Efek metode inaktifasi dengan sinar gamma pada virus influenza-A yang
dapat menginduksi perlindungan silang pada beberapa subtipe virus dan persiapan
vaksin komersial ini juga dilakukan oleh Furuya et al. (2010).
Fenomena perlindungan silang terhadap beberapa subtipe virus influenza
akibat vaksin iradiasi gamma sebagaimana tersebut di atas, kemungkinan dapat
terjadi pula pada vaksin iradiasi SGB karena sama-sama memiliki beberapa subtipe.

18

19
BAHAN DAN METODE
1. Tempat dan Waktu Penelitian
Kegiatan dilaksanakan di Laboratorium Mikrobiologi Medik, Fasilitas Kandang
Bagian Hewan Coba untuk hewan model mencit dan Laboratorium Patologi, Fakultas
Kedokteran Hewan - IPB. Perlakuan iradiasi bahan vaksin SGB dilakukan dalam
iradiator Gamma chamber dengan sumber isotop 60
Co, di Balai Iradiasi, Pusat
Aplikasi Teknologi Isotop dan Radiasi (PATIR), Badan Tenaga Nuklir Nasional
(BATAN), Jakarta. Aplikasi kandidat vaksin iradiasi SGB dilakukan pada peternakan
kmamae perah Bangun Dioro Farm, desa Palasari, Kabupaten Bogor.
Penelitian dilakukan sejak Januari 2010 sampai Februari 2012.
2. Sampel Susu
Sampel susu diperoleh dari kasus mastitis subklinis sapi perah di peternakan
rakyat yaitu Kawasan Usaha Ternak (KUNAK) Cibungbulang Kabupaten Bogor dan
daerah Cisurupan serta Bayongbong Kabupaten Garut. Penapisan awal menggunakan
California Mastitis Test (CMT) pada 65 ekor sapi perah dengan total sampel susu
yang diambil dari tiap puting sapi adalah sebanyak 240 sampel, yang berasal dari
Kunak (66 sampel), Cisurupan (97 sampel) dan Bayongbong (77 sampel).
3. Pemeriksaan Mastitis Subklinis Secara Tak Langsung.
Pengujian sampel susu di lapang dilakukan dengan menggunakan uji
California Mastitis Test (CMT). Setiap sampel susu yang diambil dari setiap kuartir
melalui pemerahan langsung ditempatkan pada paddle sesuai urutan kuartir. Uji CMT
dimulai setelah susu dalam paddle ditambahkan dengan pereaksi CMT dengan
perbandingan yang sama, kemudian dihomogenkan campuran susu dan reagen
dengan cara menggoyang paddle secara searah selama 20 – 30 detik. Hasil campuran
segera diamati kekentalannya maksimal dalam 30 detik (reaksi cenderung mengarah
ke arah reaksi positif jika terlalu lama), kemudian diamati dengan bantuan tusuk gigi
yaitu dengan menarik hasil campuran keatas. Ukuran kekentalan diukur dengan

20
positif I (tebentuknya lendir tipis), II (terbentuk lendir lebih kental), dan III (lendir
sangat kental seperti massa gelatin). Sampel dengan kekentalan positif I, II dan III
dipakai sebagai sampel untuk kandidat yang dikultur pada agar darah (Ruegg 2005).
4. Pemeriksaaan Mastitis Subklinis Secara Langsung
Uji dilakukan dengan metode Total Plate Count (TPC). Pemeriksaan sampel
susu terhadap jumlah total bakteri yang dapat ditemukan dalam media Plate Count
Agar (PCA). Sebanyak 1 ml sampel susu dimasukkan ke dalam pengencer NaCl
fisiologis 9 ml, kemudian dihomogenkan selama 1 menit, campuran ini merupakan
pengenceran ke-1 (10-1
). Sebanyak 1 ml pengenceran 10-1
dipindahkan ke dalam 9 ml
pengencer NaCl fisiologis pada tabung yang berbeda, dan campuran ini merupakan
pengenceran ke-2 (10-2
). Langkah ini terus dilakukan hingga pengenceran ke-6 (10-6
).
Pada penelitian ini digunakan metode hitungan cawan dengan cara tuang
(pour plate method). Sebanyak 1 ml dari masing-masing pengenceran tersebut di atas
dipupuk di dalam cawan petri steril yang telah diberi label sesuai dengan urutan
pengenceran, kemudian ditambah dengan media PCA yang telah dicairkan sebanyak
18 ml (suhu 450C). Campuran dihomogenkan dengan membuat gerakan angka 8 pada
cawan petri di tempat yang datar lalu didiamkan hingga memadat dan diinkubasi
didalam inkubator pada suhu 350C selama 24-48 jam dengan posisi terbalik.
Pencatatan terhadap pertumbuhan koloni dilakukan setelah 48 jam pada setiap cawan
yang mengandung 25-250 koloni. Langkah ini dilanjutkan dengan penghitungan
angka lempeng total dalam cawan tersebut dengan mengalikan jumlah rata-rata
koloni pada cawan sesuai dengan faktor pengenceran yang digunakan (Lay 1994).
5. Identifikasi dan Isolasi S. agalactiae
Isolasi S. agalactiae berasal dari air susu yang menunjukkan reaksi CMT
positif. Identifikasi terhadap keberadaan S. agalactiae berdasarkan morfologi koloni,
morfologi sel bakteri dengan pemeriksaan mikroskopis dan pewarnaan Gram, uji

21
katalase dan keberadaan faktor Christie, Atkins dan Muence Petersen (CAMP) pada
media agar darah (Quinn et al. 2006).
5.1. Pemeriksaan Morfologi Koloni Bakteri
Uji dilakukan pada media agar darah (Blood agar plate/ BAP. Kultur bakteri pada
media BAP dilakukan secara aseptik untuk mencegah terjadinya
pencemaran/kontaminasi pada media yang digunakan. Langkah pertama adalah
memberikan label pada cawan-cawan petri yang telah berisi media Blood Agar Plate
(BAP). Langkah berikutnya sampel susu kuartir diambil sebanyak 2 mata öse dan
digoreskan pada agar darah. Penanaman bakteri secara aseptis dilakukan dalam
kabinet laminair flow dekat nyala api bunsen. Agar darah yang telah ditanami sampel
susu kuartir, lalu diinkubasi dalam inkubator selama 24 jam pada suhu 37˚C.
Pengamatan dilakukan terhadap pertumbuhan koloni bakteri dan wilayah jernih di
sekitar koloni (hemolisis). Bakteri Streptococcus memiliki bentuk koloni-koloni bulat
halus, basah, cembung, terang tembus, dan menghemolisis sel darah merah.
Hemolisis yang terjadi di sekeliling koloni bakteri dapat dibedakan menjadi tiga jenis,
yaitu hemolisis alpha (α) berupa hemolisis sebagian yang ditunjukkan dengan zona
kehijauan disekitar koloni; hemolisis beta (β) berupa hemolisis komplit yang
ditunjukkan dengan zona bening di sekeliling koloni, dan hemolisis gamma (γ) yang
tidak menunjukkan perubahan warna di seputar koloni bakteri (Quinn et al. 2006).
5.2. Pemeriksaan Mikroskopi
Untuk menunjang pemeriksaan mikroskopi maka dilakukan pewarnaan Gram.
Langkah awal dari pewarnaan Gram adalah mempersiapkan preparat yang akan
diwarnai. Larutan NaCl fisiologis secukupnya diteteskan diatas gelas objek.
Kemudian koloni bakteri sebanyak 1 Öse diambil dari media Blood Agar Plate (BAP)
dan dicampurkan bersama NaCl fisiologis diatas gelas objek tadi. Pencampuran ini
dilakukan dengan cara menggerakkan Öse memutar dari arah dalam dan makin lama

22
makin keluar. Selanjutnya campuran yang terdapat diatas objek gelas tadi difiksasi
diatas bunsen yang menyala. Kemudian preparat ini diteteskan dengan zat warna
kristal violet diatas gelas objek hingga preparat tertutup, langkah ini dilakukan selama
1 menit. Setelah itu ditambahkan lugol diatas preparat tadi selama 1 menit. Preparat
kemudian dicuci dengan aquadest/air kran diatas bak pewarnaan. Pencucian
dilanjutkan dengan aceton alkohol selama 15 detik. Preparat kemudian dibilas sekali
lagi dengan aquadest/air kran hingga bersih. Pewarnaan kemudian dilanjutkan dengan
meneteskan safranin selama 15 detik pada preparat tersebut dan kemudian dibilas lagi
dengan aquadest/air kran hingga bersih. Preparat yang telah dibilas kemudian
dikeringkan, lalu ditetesi dengan minyak emersi dan siap untuk diamati dibawah
mikroskop dengan perbesaran 10x100 (Carter 1984). Koloni Streptococccus sp. akan
terlihat berbentuk bulat (coccus), bergerombol membentuk rantai, berwarna ungu.
5.3 Uji Katalase
Uji ini dilakukan sebagai acuan untuk identifikasi bakteri yang
berkarakteristik dapat memproduksi enzim katalase. Langkah yang pertama yaitu
menyiapkan gelas objek atau cawan petri yang sebelumnya telah dibersihkan dengan
alkohol 70% dan dikeringkan diatas api Bunsen. Pada gelas objek ini kemudian
diteteskan reagen H2O2 3% sebanyak 1 tetes. Langkah berikutnya yaitu mengambil
koloni bakteri yang telah tumbuh dari Blood Agar Plate (BAP) sebanyak 1 Öse dan
kemudian dicampurkan pada reagen H2O2 tadi. Reaksi memberikan hasil positif
apabila terbentuk gelembung-gelembung udara, sebaliknya reaksi memberikan hasil
negatif apabila tidak terbentuk gelembung. Bakteri Streptococcus sp. akan
memberikan hasil berupa katalase negatif (Lay 1994).
5.4. Uji Christie, Atkins, and Munch-Petersen (CAMP)
Uji CAMP dilakukan pada media agar darah. Koloni bakteri yang akan diuji
adalah koloni bakteri yang telah dikultur ulang selama satu malam. Koloni S. aureus
diambil dengan öse, digoreskan vertikal ditengah-tengah media agar darah sehingga
membagi agar darah menjadi dua bagian sama besar. Koloni Streptococcus diambil

23
dengan ose kemudian digoreskan horizontal di sebelah kanan dan kiri goresan
Staphylococcus aureus membentuk seperti garis tegak lurus. Jarak antara kedua
goresan kira-kira 0.5 cm dari goresan Staphylococcus aureus. Setelah selesai, agar
darah diinkubasi selama 18-24 jam pada suhu 37 ο
C. Uji positif akan menunjukan
zona bening seperti anak panah (arrow head) diantara kedua goresan bakteri tersebut
(Allen-Mierl et al. 2006).
5.5. Uji Serogrup S. agalactiae
S. agalctiae dari mastitis subklinis umumnya termasuk Grup-B. Identifikasi
dilakukan dengan menggunakan Streptococcal Grouping kit (Oxoid®, England). Tiap
isolat bakteri CAMP positif yang telah diremajakan pada media agar darah diambil 3-
5 koloni menggunakan öse secara aseptis, lalu dimasukan ke dalam tabung-tabung
kecil ujung lancip bertutup ukuran 1 ml yang telah berisi ekstrak enzim streptococcal
masing-masing sebanyak 200 µl. Suspensi tersebut dipanaskan di dalam penangas air
pada suhu 37 οC selama 10 menit. Setiap lima menit tabung-tabung tersebut dikocok.
Langkah berikutnya suspensi diambil dengan pipet Pasteur lalu diteteskan sebanyak
dua tetes di atas kertas latex. Tambahkan latex reagen group B pada suspensi dan
keduanya dicampurkan, dibiarkan selama 10-15 detik sambil diamati reaksi
aglutinasi yang terjadi.
5.6. Ekspresi Fenotipe SGB in vitro
Pengujian ekspresi fenotip dilakukan dengan menanam SGB pada media Soft
Agar (SA) dan media cair Todd Hewit Broth (THB) untuk diamati pola
pertumbuhannya. Isolat SGB yang telah diremajakan pada media agar darah ditanam
secara aseptis sebanyak satu öse kedalam tabung reaksi yang berisi 5 ml media brain
heart infusion (BHI) broth. Suspensi diinkubasi selama 18-24 jam pada suhu 37 οC,
kemudian biakan segar diambil dengan öse dan diencerkan dalam 10 ml larutan NaCl
0.14 M. Langkah berikutnya, batang öse ujung lurus dimasukan kira-kira nya ke

24
dalam suspensi biakan segar SGB yang telah diencerkan, lalu ditanam kedalam 10 ml
media soft agar (SA). Setiap suspensi tersebut ditanam dalam dua media SA. Salah
satu dari media SA ditambahkan serum kelinci sebanyak 200 µl (serum soft
agar/SSA) selanjutnya diinkubasi selama 18-24 jam pada suhu 37οC. Pengujian pola
tumbuh pada media cair dilakukan dengan menanam 1 öse biakan segar SGB ke
dalam 10 ml media cair THB, kemudian diinkubasi pada suhu 37oC selama 18-24 jam
dan diamati pola pertumbuhannya (Wibawan and Lämmler, 1990).
5.7. Uji Hemaglutinasi
Isolat S. agalactiae Group B (SGB) selanjutnya diuji hemaglutinasi
menggunakan 1% eritrosit sapi perah dalam NaCl fisiologis. Pengujian hemaglutinasi
dilakukan dengan mencampur secara homogen 50 µl suspensi bakteri 108
sel/ml
dengan 50 µl eritrosit sapi perah ke dalam mikroplate. Reaksi hemaglutinasi dapat
dibaca jika kontrol eritrosit mengendap. Reaksi dikatakan positif apabila terjadi
aglutinasi di dasar tabung dan negatif apabila terjadi endapan seperti pada kontrol
eritrosit (Wahyuni dkk. 2005).
6. Orientasi Dosis Iradiasi Sinar Gamma SGB untuk LD50
Isolat SGB terpilih (yang mempunyai hemaglutinin positif/ Hn+) ditumbuhkan
dalam media BHI broth dan diinkubasi 37oC pada inkubator. Jika telah mencapai fase
log dengan waktu pembelahan sel tertinggi pada kurva pertumbuhan perlakuan, lalu
bakteri dicuci memakai aquades steril 3 kali dengan sentrifus 10.000 rpm. Setelah
pencucian, bakteri diencerkan menjadi 108
cfu/ml, lalu diiradiasi dengan dosis iradiasi
bertingkat dengan laju dosis 112,504 krad/ jam, di dalam iradiator Gamma chamber
di PATIR, BATAN. Selanjutnya bakteri ditanam pada media BHI agar plat dan
diinkubasi di dalam inkubator 37oC selama 1 malam, lalu keesokan harinya koloni
SGB yang tumbuh dihitung untuk menentukan nilai LD50 bakteri SGB iradiasi yang

25
masih bertahan hidup sebanyak 50% sebagaimana penelitian terdahulu (Tuasikal dkk.
2003).
7. Karakterisasi Protein Antigenik Permukaan SGB dengan SDS PAGE
Karakterisasi protein permukaan SGB iradiasi dilakukan dengan dua tahap,
yaitu persiapan protein antigenik dan teknik SDS-PAGE (Sodium Dodecyl Sulphate
Polyacrylamid Gel Electrophoresis) untuk mengukur berat molekulnya (Walker JM
2002).
Protein antigenik diperoleh dari isolat SGB yang sudah ditanam dalam BHI
Broth dan diberi perlakuan iradiasi dengan dosis 0 dan 17 Gy, lalu disentrifuge
dengan kecepatan 10.000 rpm selama 10 menit. Pelet sel bakteri diambil kemudian
dicuci dengan NaCl fisiologis, selanjutnya diencerkan dengan NaCl fisiologis sampai
volume 1,5 ml. Suspensi sel dimasukkan ke dalam tabung sonikator untuk memecah
dinding sel dan melepas protein. Sonikasi dilakukan selama tiga menit. Sel hasil
sonikasi dipindahkan ke dalam tabung mini steril, kemudian disentrifuge pada 10.000
rpm selama 3 menit. Supernatan yang diperoleh dipisahkan dan disimpan pada suhu
0-4 oC sampai akan digunakan dalam pengukuran konsentrasi protein menggunakan
metode Bradford.
Penentuan berat molekul (BM) protein dianalisis dengan metode SDS PAGE.
Pembuatan agar akrilamid dilakukan dengan bantuan dua lempeng kaca berukuran 13
x 15,5 cm (Pharmacia - Biothec® yang telah dibersihkan dengan alkohol 70%. Kedua
lempeng kaca di himpit dan dijepit, serta diberi celah pada kedua sisi tepi bagian
dalam. Di bagian atas lempeng kaca disisipkan sisir pembuat jalur yang diisi gel
pemisah (12% poliakrilamid) sampai 1 cm di bawah ujung sisir dengan bantuan
mikropipet, lalu dibiarkan sekitar 60 menit, kemudian diisi gel pengumpul (4%
poliakrilamid) hingga mencapai permukaan lempeng kaca. Gel yang sudah dicetak
kemudian dibuka sisirnya, selanjutnya dipasang pada tangki elektroforesis, dan
ditambahkan buffer elektroforesis (Tris-glysin) sampai gel terendam. Sumuran
dibersihkan sebelum digunakan dengan menyemprotkan bufer elektroforesis ke

26
dalamnya sehingga gel-gel yang tersisa di dalam sumuran dapat keluar. Setiap
sumuran diisi dengan 10 µl campuran sampel protein dan sampel buffer yang sudah
dipersiapkan sebelumnya. Selain sampel digunakan juga marker protein sebagai
penanda. Marker protein yang digunakan mempunyai berat molekul 25 sampai 225
kDa. Visualisasi berat molekul protein antigen dilakukan dengan perangkat
elektroforesisi yang dihubungkan ke arus listrik pada tegangan 100 volt dengan arus
50 mA selama kurang lebih 3 jam sampai sampel buffer terlihat pada bagian bawah
gel (kurang lebih 1 cm di atas batas bawah gel). Elektroforesis dilakukan pada
kondisi suhu 4oC. Setelah elektroforesis berakhir, gel diangkat dari lempeng kaca dan
direndam di dalam pewarnaan Commasie Brilliant Blue selama 30 menit pada suhu
ruang sambil diagitasi perlahan. Pewarna yang tidak terikat pada protein dihilangkan
dengan merendam gel pada larutan pemucat metanol dan asam asetat sehingga gel
berwarna bening atau pita-pita protein telah terlihat jelas. Mobilitas relatif protein
dihitung dengan membandingkan jarak migrasi protein dari garis awal gel pemisah
dengan jarak migrasi pewarna, atau dibandingkan terhadap pewarna marker.
8. Uji Patogenitas dan Imunitas dengan Hewan Model Mencit
Hewan model yang digunakan adalah mencit betina jenis Balb-C, berumur 8
minggu dengan bobot badan kira-kira 50 gram. Sebanyak 12 ekor mencit dibagi
dalam 4 kelompok perlakuan, yaitu kelompok vaksin (V) diberi SGB Hn+
iradiasi,
kelompok vaksin dan tantang (VT) diberi vaksin SGB Hn+ iradiasi lalu ditantang
dengan SGB Hn+
tanpa iradiasi; kelompok tantang (T) diinfeksi dengan SGB Hn+
tanpa iradiasi, dan kelompok kontrol (K) tanpa vaksin dan tanpa tantang. Bakteri
yang diberikan mempunyai kepadatan 108 cfu / ml. Vaksin SGB Hn+ iradiasi
diinjeksikan dengan route intraperitoneal dengan dosis 0,3 – 0,4 cc/ ekor. Vaksin
booster diberikan setelah hari ke 7 dan ke 14, dan tantangan pada hari ke 21 (post
partus). Infeksi tantang SGB Hn+ tanpa iradiasi diberikan melalui parenteral (tetes)
pada 5 pasang putting sebanyak 50 µl/ mencit. Suspensi bakteri tantangan diteteskan
di atas orificium externa puting susu mencit secara bertahap (satu tetes sebanyak 5

27
µl), kemudian ditunggu hingga terabsorbsi. Nekropsi dilakukan sehari setelah
tantangan. Penilaian respon terhadap vaksin yang diberikan, dilakukan dengan
pemeriksaan Histopatologi jaringan kelenjar mamae yang difiksasi formalin dengan
pewarnaan hematoksilin dan eosin (HE) (Kiernan, 1990). Titer antibodi Ig-G
diperiksa dengan teknik Enzyme Linked Immunosorbent Assay secara tak langsung
(indirect ELISA) dari sampel serum darah yang dikoleksi tiap minggu (Crowther
2010).
9. Aplikasi Kandidat Vaksin Iradiasi Terpilih pada Kambing Perah
Hewan percobaan yang digunakan sebanyak 6 ekor kambing perah peranakan
etawah yang dibagi menjadi 2 kelompok perlakuan, yaitu 4 ekor untuk kelompok
vaksin (V) yang diberi SGB Hn+ iradiasi secara subkutan dengan dosis 108
cfu/ml
sebanyak 2 ml/ kambing, dan 2 ekor untuk kelompok kontrol (K) yaitu hewan normal
yang tidak diberi vaksin SGB. Vaksin diberikan pada masa kering kandang dengan
pemberian booster 3 kali prepartus. Pemeriksaan konsentrasi Ig-G dilakukan dengan
teknik ELISA dari serum sampel yang dikoleksi tiap minggu (Crowther 2010).
Produksi susu postpartus dicatat setiap hari.
10. Rancangan Penelitian
Informasi tentang efektifitas bahan vaksin ini akan dilakukan dengan cara
membandingkan kondisi hewan percobaan antara yang hanya divaksin (V) yang
diberi vaksin dengan tantangan (VT), yang diberi tantangan tanpa vaksin (T); dan
hewan kontrol tanpa vaksin maupun tantangan (K). Signifikansi dari efektifitas bahan
vaksin akan ditelusuri dengan membandingkan kelompok perlakuan tersebut
menggunakan rancangan acak lengkap (RAL). Gambar histopatologi diambil dengan
kamera digital Electronic Eyepiece MD-130® menggunakan mikroskop cahaya.
Gambar diolah menggunakan software NIH Image-J®. Data pengamatan dianalisis

28
pada analisis ragam (ANOVA), post hoc Duncan test menggunakan software SPSS®
versi 16 untuk mengetahui perbedaan setiap perlakuan pada p<0,05.

29
HASIL DAN PEMBAHASAN
1.Kejadian Mastitis Subklinis di Daerah Pengambilan Sampel Susu
Sampel susu yang digunakan berasal dari 3 wilayah pada 2 daerah yang berbeda
yaitu daerah Bogor dengan wilayah Kawasan Usaha Peternakan (Kunak)
Cibungbulang dan Garut dengan wilayah Kecamatn Cisurupan dan Kecamatan
Bayongbong. Sampel susu dari hewan penderita mastitis subklinis dapat diketahui
dengan CMT (California Mastitis Test). Dengan uji ini didapatkan hasil dengan
mengamati parameter derajat konsistensi kekentalan dari campuran susu dan reagen
uji yang membentuk materi gelatinous kental. Parameter ini diukur dengan melihat
kekentalan yang terjadi untuk positif I akan terbentuk lendir tipis, positif II akan
terbentuk lendir yang lebih kental, dan positif III lendir yang terbentuk sangat kental
seperti massa gelatin. CMT bekerja berdasarkan 3 prinsip yaitu jumlah leukosit akan
meningkat drastis saat jaringan mamae mengalami luka atau infeksi, leukosit
terutama polymorphonuclear leukosit (PMNs) mempunyai inti sel yang besar (DNA)
bila dibandingkan dengan sel lain atau bakteri yang terdapat didalam susu, selain itu
penyusun utama dinding sel leukosit adalah lipid (lemak). Semakin tebal gel yang
terbentuk di dalam piring uji CMT, maka semakin banyak juga sel darah putih yang
terdapat di dalam sampel susu. Peningkatan penebalan dari gel ini menandakan
terdapat peningkatan dalam hal infeksi. Reagen dari CMT merupakan suatu detergen
dengan penambahan indikator pH yang menyebabkan terbentuknya warna ungu pada
pencampuran dengan sampel susu. Saat sampel susu dan reagen MT dicampur
dengan jumlah yang setara, reagen CMT akan melarutkan dinding sel terluar dan
dinding sel nuklear dari leukosit yang penyusun utamanya adalah lemak, dimana
detergen akan melarutkan lemak. Hal ini mengakibatkan DNA lepas dari inti yang
mengarah kepada pembentukan gel oleh sesama DNA ntuk membentuk suatu massa
seperti serabut. Seiring dengan penambahan jumlah leukosit pada setiap quarter akan
berbanding lurus dengan peningkatan jumlah pembentukan gel (Ruegg 2005). Hasil
dari uji ini pada ketiga wilayah dapat dilihat pada gambar 2.
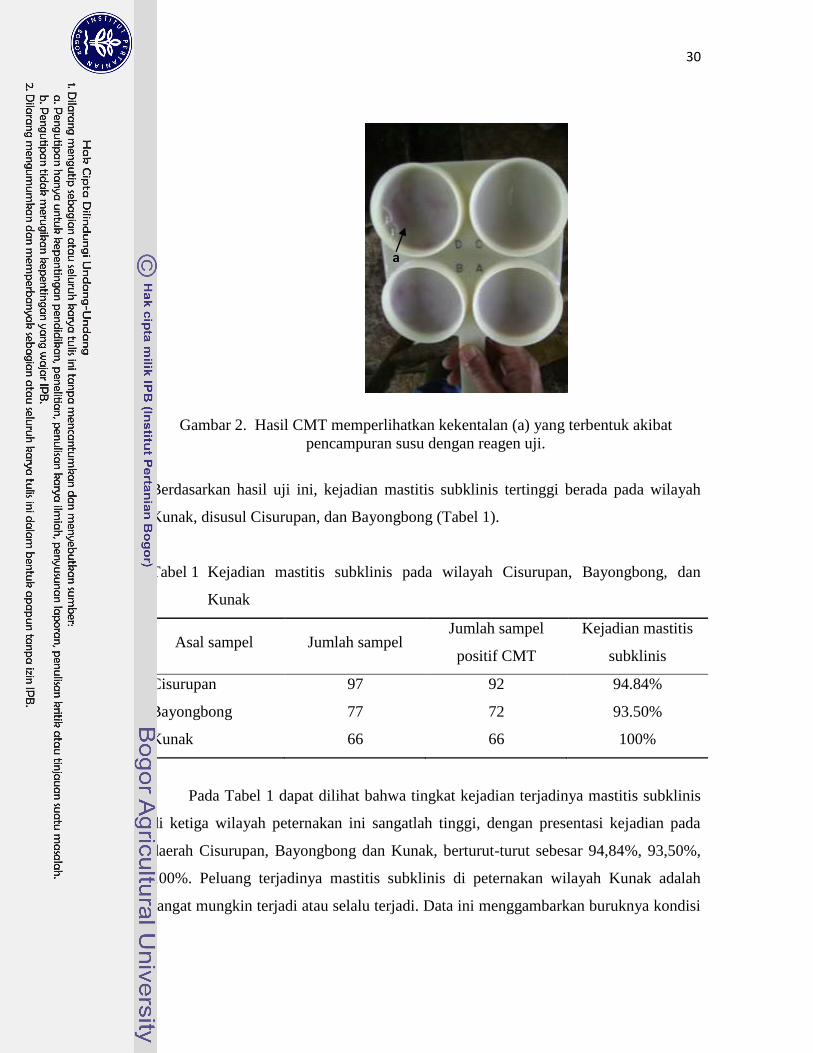
30
Gambar 2. Hasil CMT memperlihatkan kekentalan (a) yang terbentuk akibat
pencampuran susu dengan reagen uji.
Berdasarkan hasil uji ini, kejadian mastitis subklinis tertinggi berada pada wilayah
Kunak, disusul Cisurupan, dan Bayongbong (Tabel 1).
Tabel 1 Kejadian mastitis subklinis pada wilayah Cisurupan, Bayongbong, dan
Kunak
Asal sampel Jumlah sampel Jumlah sampel
positif CMT
Kejadian mastitis
subklinis
Cisurupan 97 92 94.84%
Bayongbong 77 72 93.50%
Kunak 66 66 100%
Pada Tabel 1 dapat dilihat bahwa tingkat kejadian terjadinya mastitis subklinis
di ketiga wilayah peternakan ini sangatlah tinggi, dengan presentasi kejadian pada
daerah Cisurupan, Bayongbong dan Kunak, berturut-turut sebesar 94,84%, 93,50%,
100%. Peluang terjadinya mastitis subklinis di peternakan wilayah Kunak adalah
sangat mungkin terjadi atau selalu terjadi. Data ini menggambarkan buruknya kondisi
a

31
lingkungan, rendahnya tingkat sanitasi perkandangan, dan cara pemerahan
mencerminkan higienitas personal yang buruk. Faktor manusia menjadi masalah
utama sehingga hal-hal buruk tersebut dapat terjadi.
Tingginya presentasi kejadian mastitis subklinis pada peternakan sapi perah di
ketiga daerah ini dapat dikaitkan dengan kondisi lingkungan dan pengelolaan
peternakan diantaranya yaitu sanitasi kandang dan cara pemerahan susu (Sori et al.
2005). Sanitasi kandang yang buruk, peralatan yang tidak bersih dan ditambah lagi
dengan cara pemerahan susu yang juga tidak bersih kemungkinan besar merupakan
faktor utama penyebab terjadinya mastitis subklinis pada ketiga peternakan sapi perah
tersebut.
Hasil pemeriksaan CMT positif yang lebih dari 50% pada penelitian ini
menunjukkan masih tingginya kejadian mastitis subklinis. Penelitian intensif Balai
Penelitian Veteriner di Jawa Barat, Jawa Tengah, Jawa Timur dan Daerah Istimewa
Yogyakarta prevalensi mastitis subklinis berkisar antara 37 sampai 67%, sedangkan
mastitis klinis antara 5 sampai 30% (Supar 1997). Kasus mastitis terutama mastitis
subklinis di Indonesia sampai akhir tahun 2006, tercatat sekitar 75% - 83%
(Sudarwanto dkk. 2006). Sebagaimana Indonesia, di negara-negara lainpun kasus
mastitis subklinis selalu lebih tinggi dibandingkan mastitis klinis. Kejadian mastitis
subklinis di Polandia sebesar 25,35% sedangkan mastitis klinis hanya 0,41%
(Hameed et al. 2007). Prevalensi mastitis klinis dan subklinis di Iran berturut-turut
2,2 dan 42,5% (Hashemi et al. 2011). Mastitis subklinik adalah mastitis yang tidak
menampakkan perubahan fisik pada mamae dan susu yang dihasilkan, tetapi dapat
menurunkan produksi susu, ditemukannya mikroorganisme patogen dan terjadi
perubahan komposisi susu (Petrovski and Emanuel, 2006). Hal-hal tersebut di atas
dapat menyebabkan kerugian ekonomis yang cukup besar dalam usaha peternakan
sapi perah. Kerugian ekonomis yang diakibatkan oleh mastitis antara lain: Penurunan
produksi susu perkwartir per hari antara 9% – 45,5% (Sudarwanto 1999); Penurunan
kualitas susu yang mengakibatkan penolakan susu mencapai 30% - 40% (Sudarwanto
dan Sudarnika 2008) dan penurunan kualitas hasil olahan susu (Hamman 2005);

32
Peningkatan biaya perawatan dan pengobatan serta pengafkiran ternak lebih awal
(Seegers et al. 2003, Shim et al. 2004).
Selain kondisi lingkungan dan pengelolaan peternakan sebagai faktor utama
penyebab mastitis subklinis di ketiga peternakan, faktor penunjang dari sisi ternak
juga berperan dalam terjadinya radang mamae ini. Kondisi kesehatan ternak yang
kurang baik, luka/lecet pada puting susu yang diakibatkan pemerahan kasar, lantai
kasar, lap yang kotor dan sikat yang keras, bentuk mamae yang sangat menggantung,
serta mamae dengan lubang puting yang terlalu lebar merupakan faktor predisposisi
terjadinya mastitis subklinis dari segi ternak. Disamping itu terdapat pula faktor
penyebab mastitis subklinis dari segi mikroorganisme penyebab, diantaranya adalah
jenis dan jumlah mikroorganisme patogen. Faktor-faktor penyebab tersebut dapat
berpengaruh terhadap kerusakan struktur dan gangguan fungsi kelenjar mamae pada
ruminansia yang terserang mastitis, sehingga berujung pada penurunan produksi susu
pada induk pasca melahirkan (Akers and Nickerson, 2011).
Kondisi lingkungan (kandang) tempat sampel susu diperoleh maupun sanitasi
saat pemerahan dapat diukur dengan menggunakan beberapa parameter diantaranya
adalah jumlah mikroba. Rata-rata jumlah mikroba dalam sampel susu yang diperoleh
dari daerah Cisurupan, Bayongbong dan Kunak disajikan dalam Tabel 2.
Tabel 2 Jumlah mikroba dalam sampel susu dari uji Total Plate Count (TPC)
Asal sampel Rata-rata jumlah mikroba (cfu/ml)
Cisurupan 1.302.893
Bayongbong 1.300.987
Kunak 1.764.613
Data dari hasil tabel diatas menunjukkan bahwa rata-rata jumlah mikroba pada
setiap perwakilan sampel di ketiga wilayah sangat tinggi karena berada diatas nilai
kisaran normal total cemaran mikroba pada susu segar yaitu 106 cfu/ml sesuai dengan
SNI 01-3141-1998. Hal ini mendukung data dari hasil CMT dimana prevalensi

33
mastitis subklinis pada peternakan sapi perah di ketiga wilayah sangat tinggi, yang
dibuktikan dengan tingginya rata-rata jumlah mikroba pada sampel susu yang
dikumpulkan secara aseptis langsung ditampung dalam tabung steril. Menurut
Lukman dkk (2009) pencemaran dapat berasal dari mamae atau masuk melalui puting
susu. Jumlah mikroba bertambah dengan adanya cemaran dari tangan dan baju
pemerah, alat perah dan lingkungan kandang. Jumlah mikroba dalam susu akibat
kontaminasi melalui udara sekitar 100-1.500 koloni/ml susu, melalui kontaminasi
mamae dan sekitarnya ditemukan 300-4.000 koloni/ml. melalui sanitasi yang buruk
pertambahan mikroba mencapai 500-15.000 koloni/ml. kontaminasi dari mamae yang
sakit mencapai 25.000 koloni/ml. jumlah mikroba dalam susu akan meningkat
melalui kontaminasi dari peralatan susu sampai dengan lebih dari 1.000.000
koloni/ml (Lukman dkk. 2009).
2. Isolat S. agactiae Group B Terpilih sebagai Bahan Vaksin Iradiasi Mastitis
Subklinis
Isolasi dan identifikasi bakteri Streptococcus sp pada sampel susu yang
dinyatakan positif mastitis subklinis berdasarkan hasil CMT, dilakukan dengan
mengkultur pada media Blood Agar Plate (BAP). Media BAP ini merupakan media
diferensial yang dapat membedakan bakteri normal dan bakteri pathogen berdasarkan
kemampuannya melisiskan sel darah merah. Selain itu pada media ini dapat pula
diamati bentuk koloni dari bakteri yang tumbuh. Koloni Streptococcus sp pada
permukaan BAP akan tampak sebagai koloni yang halus, kecil, cembung, transparan
sampai putih (Gambar 3). Koloni ini akan membentuk pola hemolisis α, hemolisis β,
atau γ (tanpa hemolisis) seperti yang ditunjukkan pada Gambar 4 (Cowan 2003).
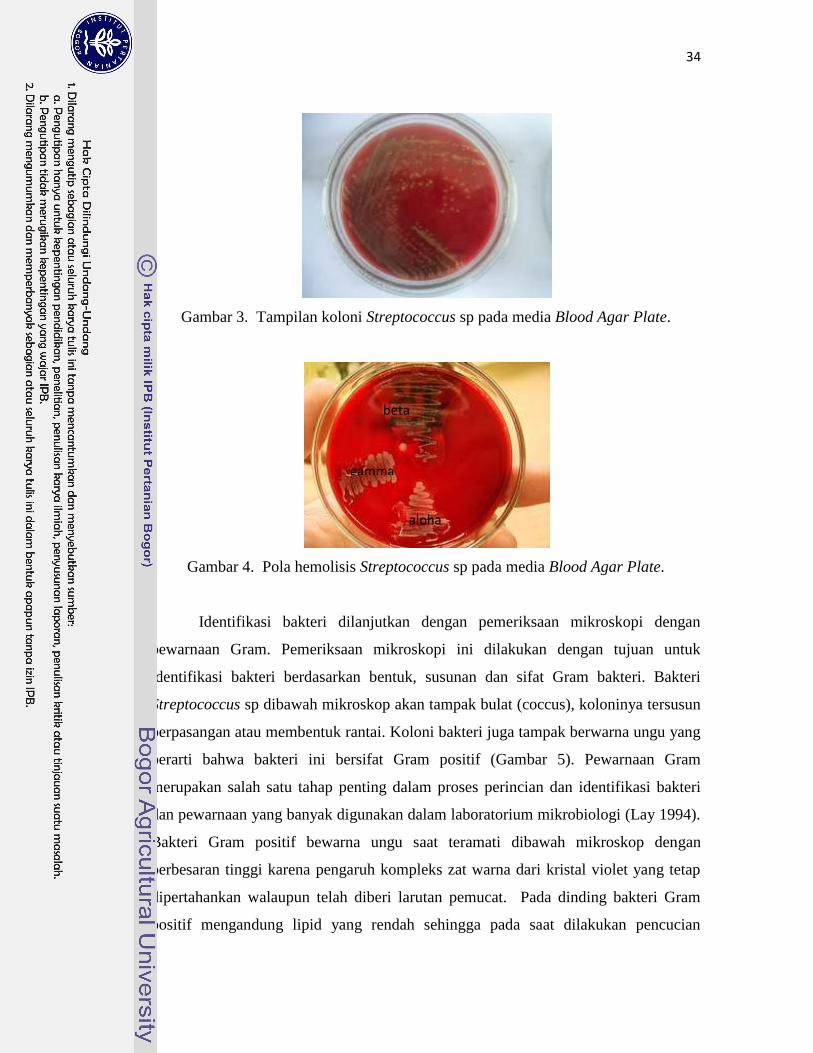
34
Gambar 3. Tampilan koloni Streptococcus sp pada media Blood Agar Plate.
Gambar 4. Pola hemolisis Streptococcus sp pada media Blood Agar Plate.
Identifikasi bakteri dilanjutkan dengan pemeriksaan mikroskopi dengan
pewarnaan Gram. Pemeriksaan mikroskopi ini dilakukan dengan tujuan untuk
identifikasi bakteri berdasarkan bentuk, susunan dan sifat Gram bakteri. Bakteri
Streptococcus sp dibawah mikroskop akan tampak bulat (coccus), koloninya tersusun
berpasangan atau membentuk rantai. Koloni bakteri juga tampak berwarna ungu yang
berarti bahwa bakteri ini bersifat Gram positif (Gambar 5). Pewarnaan Gram
merupakan salah satu tahap penting dalam proses perincian dan identifikasi bakteri
dan pewarnaan yang banyak digunakan dalam laboratorium mikrobiologi (Lay 1994).
Bakteri Gram positif bewarna ungu saat teramati dibawah mikroskop dengan
perbesaran tinggi karena pengaruh kompleks zat warna dari kristal violet yang tetap
dipertahankan walaupun telah diberi larutan pemucat. Pada dinding bakteri Gram
positif mengandung lipid yang rendah sehingga pada saat dilakukan pencucian
beta
a
gamma
alpha

35
dengan larutan pemucat terjadi dehidrasi dan lubang pori-porinya mengecil, itu
sebabnya zat warna tetap dipertahankan. Ditemukan juga pada dindingnya senyawa
Mg-ribonukleat yang mengikat kristal violet sehingga tidak mudah dilarutkan (Lay
1994).
Gambar 5. Tampilan mikroskopik Streptococcus sp. Gram positif menggunakan
mikroskop cahaya (perbesaran 10x100).
Untuk membedakan lebih lanjut bakteri bulat Gram positif maka dilakukan uji
katalase. Uji ini dilakukan sebagai acuan untuk identifikasi bakteri yang
berkarakteristik dapat memproduksi enzim katalase. Katalase yang diproduksi oleh
bakteri ini dapat mengubah hidrogen peroksida menjadi gas hidrogen dan air. Hasil
positif mengindikasikan adanya cytochrome oxidase (Quinn et al. 2006). Bakteri
Streptococcus sp akan memberikan hasil katalase negatif yang ditandai dengan tidak
terbentuknya gas hidrogen dan air. Hal ini menunjukkan salah satu karakteristik
bakteri tersebut yaitu tidak mempunyai enzim cytochrome oxidase. Uji katalase
positif pada Staphylococcus sp, dengan reaksi yang terjadi sebagai berikut:
H2O2 enzim katalase H2O(l) + O2(g)
Berdasarkan pada prosedur isolasi dan identifikasi bakteri yang telah
dilaksanakan maka diperoleh sejumlah sampel dari ketiga daerah yang memenuhi
kriteria sebagai Streptococcus sp penyebab mastitis subklinis. Jumlah sampel yang
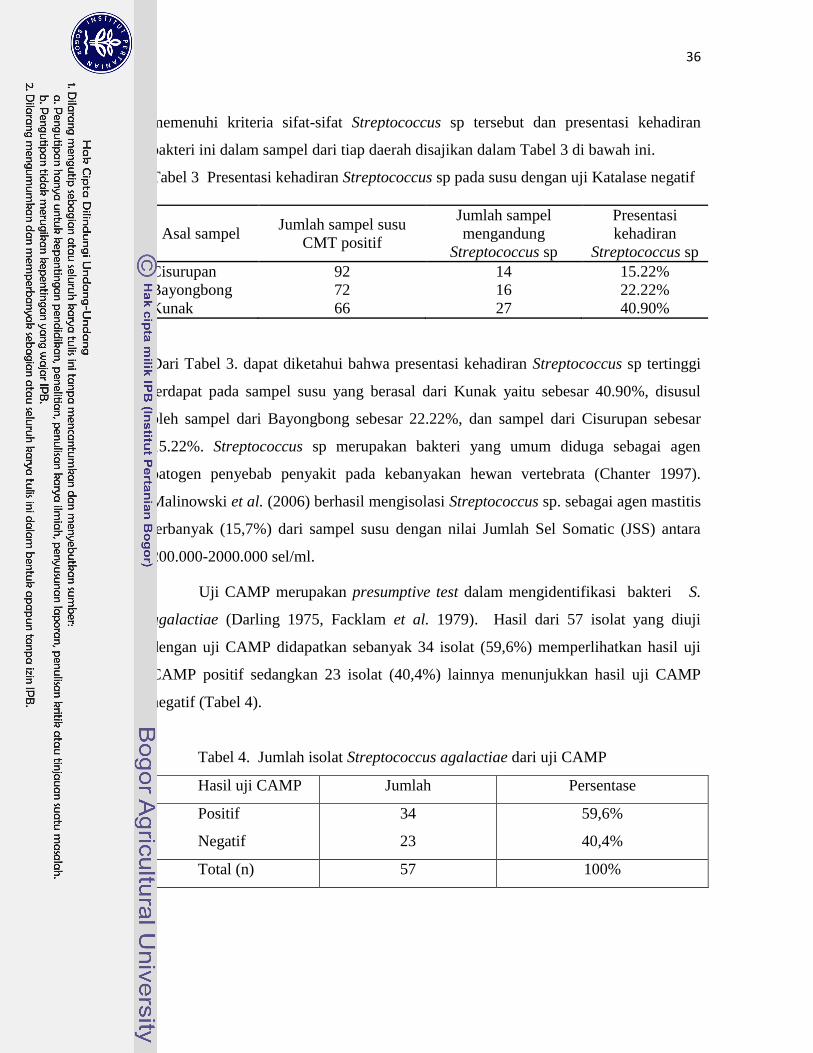
36
memenuhi kriteria sifat-sifat Streptococcus sp tersebut dan presentasi kehadiran
bakteri ini dalam sampel dari tiap daerah disajikan dalam Tabel 3 di bawah ini.
Tabel 3 Presentasi kehadiran Streptococcus sp pada susu dengan uji Katalase negatif
Asal sampel Jumlah sampel susu
CMT positif
Jumlah sampel
mengandung
Streptococcus sp
Presentasi
kehadiran
Streptococcus sp
Cisurupan 92 14 15.22%
Bayongbong 72 16 22.22%
Kunak 66 27 40.90%
Dari Tabel 3. dapat diketahui bahwa presentasi kehadiran Streptococcus sp tertinggi
terdapat pada sampel susu yang berasal dari Kunak yaitu sebesar 40.90%, disusul
oleh sampel dari Bayongbong sebesar 22.22%, dan sampel dari Cisurupan sebesar
15.22%. Streptococcus sp merupakan bakteri yang umum diduga sebagai agen
patogen penyebab penyakit pada kebanyakan hewan vertebrata (Chanter 1997).
Malinowski et al. (2006) berhasil mengisolasi Streptococcus sp. sebagai agen mastitis
terbanyak (15,7%) dari sampel susu dengan nilai Jumlah Sel Somatic (JSS) antara
200.000-2000.000 sel/ml.
Uji CAMP merupakan presumptive test dalam mengidentifikasi bakteri S.
agalactiae (Darling 1975, Facklam et al. 1979). Hasil dari 57 isolat yang diuji
dengan uji CAMP didapatkan sebanyak 34 isolat (59,6%) memperlihatkan hasil uji
CAMP positif sedangkan 23 isolat (40,4%) lainnya menunjukkan hasil uji CAMP
negatif (Tabel 4).
Tabel 4. Jumlah isolat Streptococcus agalactiae dari uji CAMP
Hasil uji CAMP Jumlah Persentase
Positif
Negatif
34
23
59,6%
40,4%
Total (n) 57 100%

37
Isolat yang menunjukkan hasil CAMP positif membentuk zona hemolisis
seperti anak panah (arrowhead) pada media BAP diantara biakan Staphylococcus
aureus dan isolat yang diuji, sedangkan hasil uji CAMP negatif tidak menunjukkan
perubahan apapun pada media BAP seperti yang terlihat pada Gambar 6.
Gambar 6. Hasil uji CAMP positif (zona bening seperti anak panah dibagian tengah
BAP), dan CAMP negatif (tidak ada hemolisis dibagian paling atas dan
paling bawah).
Fenomena yang terjadi pada uji CAMP karena adanya interaksi antara faktor
CAMP yang dihasilkan isolat yang diuji (dalam hal ini Streptococcus agalactiae)
dengan produk ekstraseluler berupa β-lysin stpahylococcal yang dihasilkan oleh
Staphylococcus aureus β-hemolisis sehingga hemolisis pada media BAP semakin
jelas karena daya hemolisis yang ditimbulkan meningkat (Hansen and Sørensen
2003) .
Bahan vaksin yang dipilih adalah Streptococcus agalactiae karena merupakan
salah satu agen utama penyebab mastitis subklinis pada sapi perah dan dapat
menyebabkan kerugian ekonomi untuk industri susu (Keefe et al. 1997, Keefe 1997).
Hasil studi Estuningsih dkk (2002) menyebutkan bahwa 83 isolat bakteri yang
diisolasi dari 3 area di pulau Jawa, seluruhnya teridentifikasi sebagai Streptococcus
agalactiae. Sesuai dengan hasil isolasi bakteri yang dilakukan Supar dan Ariyanti
(2008) dari sampel susu kuartir di daerah Bandung, Bogor dan Sukabumi diperoleh
60,6% Streptococcus agalactiae sebagai bakteri dominan penyebab mastitis
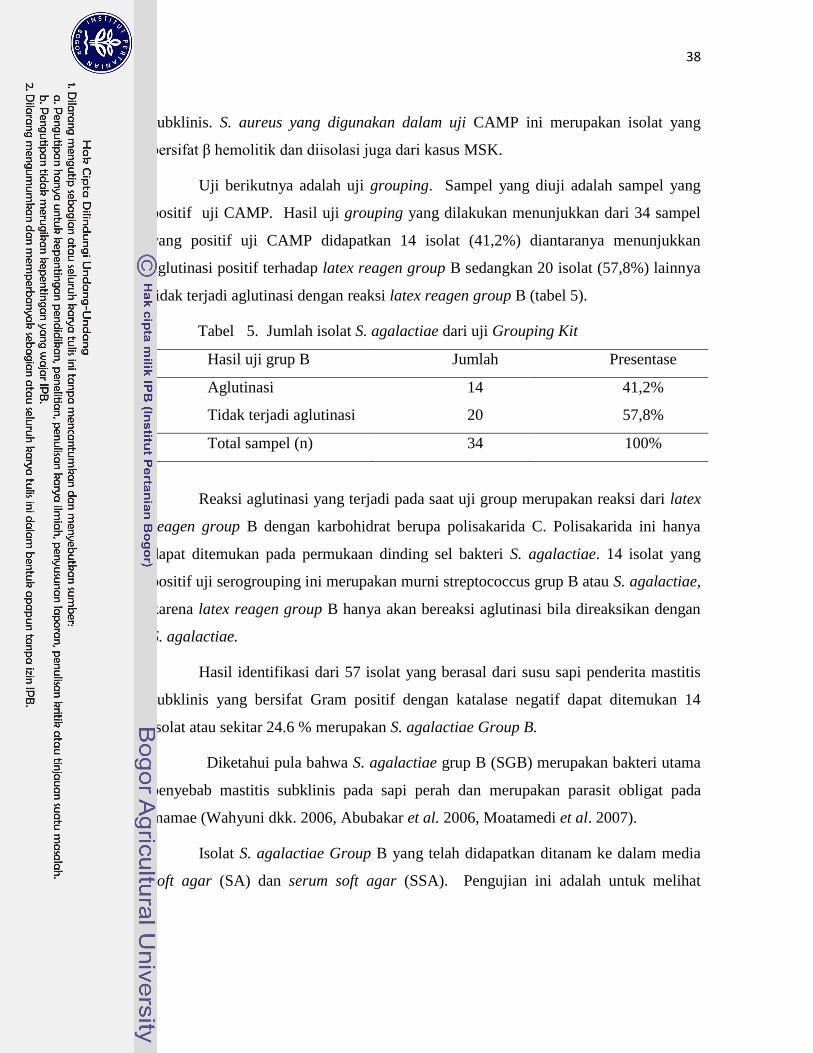
38
subklinis. S. aureus yang digunakan dalam uji CAMP ini merupakan isolat yang
bersifat β hemolitik dan diisolasi juga dari kasus MSK.
Uji berikutnya adalah uji grouping. Sampel yang diuji adalah sampel yang
positif uji CAMP. Hasil uji grouping yang dilakukan menunjukkan dari 34 sampel
yang positif uji CAMP didapatkan 14 isolat (41,2%) diantaranya menunjukkan
aglutinasi positif terhadap latex reagen group B sedangkan 20 isolat (57,8%) lainnya
tidak terjadi aglutinasi dengan reaksi latex reagen group B (tabel 5).
Tabel 5. Jumlah isolat S. agalactiae dari uji Grouping Kit
Hasil uji grup B Jumlah Presentase
Aglutinasi
Tidak terjadi aglutinasi
14
20
41,2%
57,8%
Total sampel (n) 34 100%
Reaksi aglutinasi yang terjadi pada saat uji group merupakan reaksi dari latex
reagen group B dengan karbohidrat berupa polisakarida C. Polisakarida ini hanya
dapat ditemukan pada permukaan dinding sel bakteri S. agalactiae. 14 isolat yang
positif uji serogrouping ini merupakan murni streptococcus grup B atau S. agalactiae,
karena latex reagen group B hanya akan bereaksi aglutinasi bila direaksikan dengan
S. agalactiae.
Hasil identifikasi dari 57 isolat yang berasal dari susu sapi penderita mastitis
subklinis yang bersifat Gram positif dengan katalase negatif dapat ditemukan 14
isolat atau sekitar 24.6 % merupakan S. agalactiae Group B.
Diketahui pula bahwa S. agalactiae grup B (SGB) merupakan bakteri utama
penyebab mastitis subklinis pada sapi perah dan merupakan parasit obligat pada
mamae (Wahyuni dkk. 2006, Abubakar et al. 2006, Moatamedi et al. 2007).
Isolat S. agalactiae Group B yang telah didapatkan ditanam ke dalam media
soft agar (SA) dan serum soft agar (SSA). Pengujian ini adalah untuk melihat
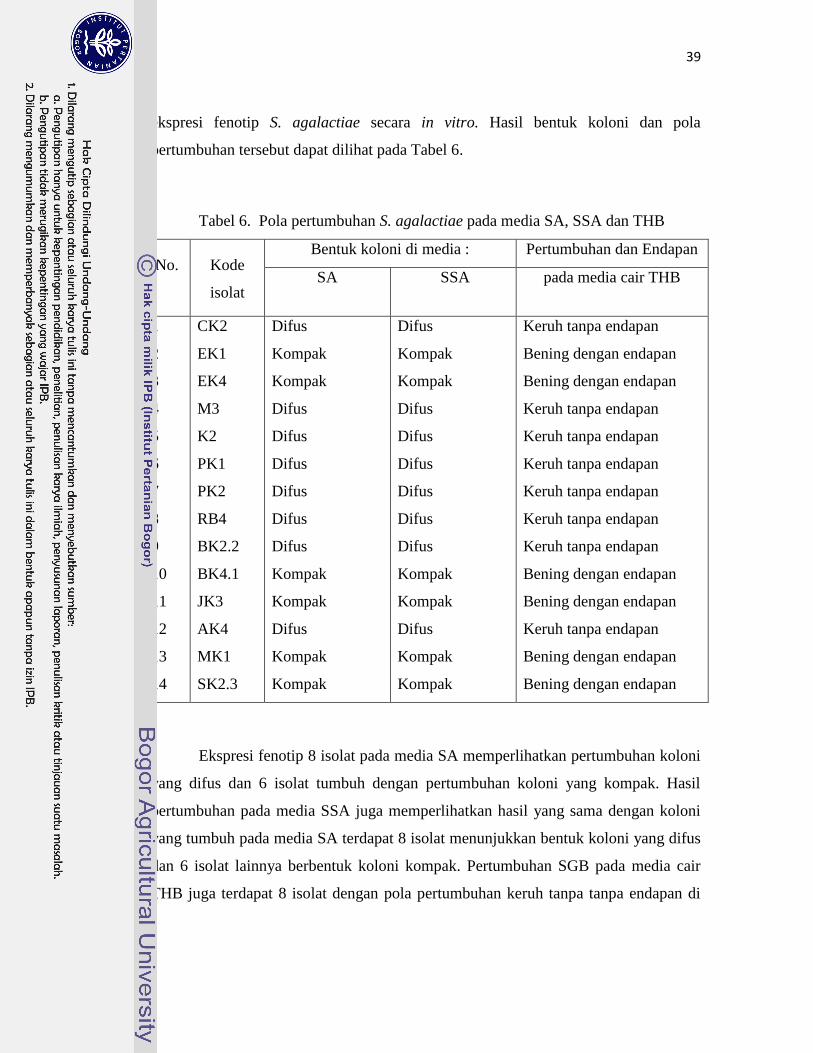
39
ekspresi fenotip S. agalactiae secara in vitro. Hasil bentuk koloni dan pola
pertumbuhan tersebut dapat dilihat pada Tabel 6.
Tabel 6. Pola pertumbuhan S. agalactiae pada media SA, SSA dan THB
No. Kode
isolat
Bentuk koloni di media : Pertumbuhan dan Endapan
SA SSA pada media cair THB
1
2
3
4
5
6
7
8
9
10
11
12
13
14
CK2
EK1
EK4
M3
K2
PK1
PK2
RB4
BK2.2
BK4.1
JK3
AK4
MK1
SK2.3
Difus
Kompak
Kompak
Difus
Difus
Difus
Difus
Difus
Difus
Kompak
Kompak
Difus
Kompak
Kompak
Difus
Kompak
Kompak
Difus
Difus
Difus
Difus
Difus
Difus
Kompak
Kompak
Difus
Kompak
Kompak
Keruh tanpa endapan
Bening dengan endapan
Bening dengan endapan
Keruh tanpa endapan
Keruh tanpa endapan
Keruh tanpa endapan
Keruh tanpa endapan
Keruh tanpa endapan
Keruh tanpa endapan
Bening dengan endapan
Bening dengan endapan
Keruh tanpa endapan
Bening dengan endapan
Bening dengan endapan
Ekspresi fenotip 8 isolat pada media SA memperlihatkan pertumbuhan koloni
yang difus dan 6 isolat tumbuh dengan pertumbuhan koloni yang kompak. Hasil
pertumbuhan pada media SSA juga memperlihatkan hasil yang sama dengan koloni
yang tumbuh pada media SA terdapat 8 isolat menunjukkan bentuk koloni yang difus
dan 6 isolat lainnya berbentuk koloni kompak. Pertumbuhan SGB pada media cair
THB juga terdapat 8 isolat dengan pola pertumbuhan keruh tanpa tanpa endapan di

40
dasar tabung, dan 6 isolat dengan pola pertumbuhan bening dengan endapan di dasar
tabung.
Bentuk koloni pada media SA dan SSA dapat dilihat pada isolat PK1 dan
MK1 dalam Gambar 7. Isolat PK1 pada media SA menunjukkan bentuk koloni difus
halus, sedangkan pada media SSA terlihat bentuk koloni difus yang lebih besar.
Koloni yang tumbuh kompak dapat terlihat pada isolat MK1. Koloni MK1 pada
media SA terlihat kompak dengan koloni yang kecil, sedangkan pada media SSA
terlihat kompak dengan koloni lebih besar. Penambahan serum ke dalam media SA
dapat memperkaya media tersebut akan unsur hara yang dibutuhkan untuk
pertumbuhan bakteri. Koloni dari isolat yang sama di media SA yang semula kecil
menjadi koloni yang tampak lebih jelas dan besar pada media SSA.
Gambar 7. Bentuk koloni S. agalactiae pada media SA dan SSA menunjukkan
pertumbuhan bentuk koloni difus (isolat PK1 sebelah kiri) dan
pertumbuhan bentuk koloni kompak (isolat MK1 sebelah kanan).
Media SA digunakan untuk melihat ekspresi fenotip dari komponen yang
terdapat pada permukaan bakteri, diantaranya untuk membedakan bakteri yang
membentuk kapsul dengan yang tidak membentuk kapsul pada permukaannya.
Kondisi ini akan ditunjukkan oleh pola pertumbuhan koloni yang tumbuh pada media

41
SA. S. agalactiae yang memiliki kapsul pada permukaannya akan diekspresikan
dengan pola pertumbuhan koloni yang difus di media SA sedangkan bentuk koloni
yang lebih kompak menandakan tidak adanya kapsul pada permukaan dinding sel
(Wibawan & Lämmler 1991). Kapsul sebagian besar penyusunnya merupakan
karbohidrat dalam bentuk polisakarida yang menutupi protein permukaan sel bakteri,
sehingga tidak terekspresikan pada SA yang bahan penyusunnya juga karbohidrat
(Harlina. 1999). Oleh karena itu, dalam media SA bakteri berkapsul akan
terekspresikan dengan bentuk pertumbuhan yang difus sedangkan bakteri tidak
berkapsul terekspresikan dalam bentuk kompak.
Bentuk pertumbuhan koloni pada media SA juga dapat menentukan
keberadaan antigen protein R dan X. Permukaan bakteri diselimuti oleh antigen
protein R atau X akan terekspresikan dengan tumbuh menjadi koloni yang kompak
pada media SA sedangkan yang tidak memiliki antigen protein akan tumbuh menjadi
difus pada media SA (Wibawan & Lämmler 1990).
Selain mengekspresikan bentuk pertumbuhan koloni pada media SA dapat
juga terlihat ekspresi pertumbuhan koloni yang ditanam pada media cair Todd-Hewitt
broth (THB). Penanaman pada media cair untuk menentukan ukuran panjang rantai
yang tersusun membntuk koloni. Pertumbuhan koloni yang keruh pada media cair
menggambarkan koloni dengan rantai pendek (<20 bakteri/rantai), sebaliknya koloni
yang tumbuh membentuk sedimen didasar tabung dan supernatan yang jernih
merupakan koloni dengan rantai yang lebih panjang (>40 bakteri/rantai). Sedimen
terbentuk karena koloni degan rantai panjang cenderung mengendap di dasar tabung
(Wibawan and Lämmler 1990).
Adanya antigen protein R dan X ini juga memberikan pengaruh terhadap
pertumbuhan pada media cair THB. Bakteri dengan antigen protein akan tumbuh
dengan supernatan yang jernih dan membentuk sedimen didasar tabung sedangkan
bakteri tanpa antigen protein tumbuh keruh di media cair (Wibawan and Lämmler
1990).

42
Umumnya komponen yang terdapat pada permukaan sel adalah faktor virulen
yang membantu bakteri dalam proses infeksi. Kapsul menentukan tingkat virulensi
dari S. agalactiae. Kehilangan kapsul asam sialat akan menghilangkan daya virulensi
(Wibawan & Lämmler 1991). Kapsul pada bakteri akan membantu menurunkan
penyingkiran bakteri terhadap fagositosis sel-sel polimorfonuklear (PMN) (Wibawan
& Lämmler 1991). Bahan pembangun kapsul merupakan polisakarida yang
merupakan antigen yang tidak imunogenik terhadap inang menyebabkan sel-sel PMN
tidak mengenalinya sebagai benda asing bagi tubuh. Namun, bakteri yang tidak
memliki kapsul akan memiliki daya adhesi yang lebih tinggi untuk melekat pada sel
inang dibanding yang memiliki kapsul (Utama et al. 1997). Adanya antigen protein R
atau X menimbulkan permukaan sel memiliki daya hidrofobitas yang tinggi dan
mempunyai peran penting terhadap daya adesi dari S. agalactiae untuk melekat pada
sel epitel inangnya (Wibawan and Lämmler 1991).
Hemaglutinin mempunyai peran dalam proses adhesi. Hasil kajian Wibawan
(1998) menyebutkan bahwa S. agalactiae yang memilki hemaglutinin mempunyai
kamampuan adhesi yang lebih besar daripada yang tidak mempunyai hemaglutinin
pada sel epitel mamae. Pada mastitis subklinis kemampuan adhesi mempunyai peran
lebih penting dari pada kemampuan invasi (Wahyuni, 2005). Kebanyakan strain
bakteri yang bereaksi hemaglutinin mempunyai tipe protein antigen-X pada S.
agalactiae yang diisolasi dari kasus mastitis pada sapi perah (Rainard dkk. 1991).
Lewat kemampuan adhesi ini S. agalactiae terbebas dari pengaruh pembasuhan
organ-organ sekresi, sehingga terhindar dari efek basuh aliran susu. Kondisi ini
menjawab alasan bakteri ini menyebabkan masititis subklinis.
Isolat bakteri terpilih untuk kandidat vaksin iradiasi pada penelitian ini adalah
Streptococcus agalactiae yang diperoleh dari susu asal mastitis subklinis dan
termasuk ke dalam kelompok Group-B (SGB). Sebagai bahan vaksin iradiasi telah
dipilih SGB dengan karakter mempunyai antigen protein tanpa kapsul yang tumbuh
dengan bentuk koloni kompak dalam media soft agar dan mempunyai Hemaglutinin
(Hn+).
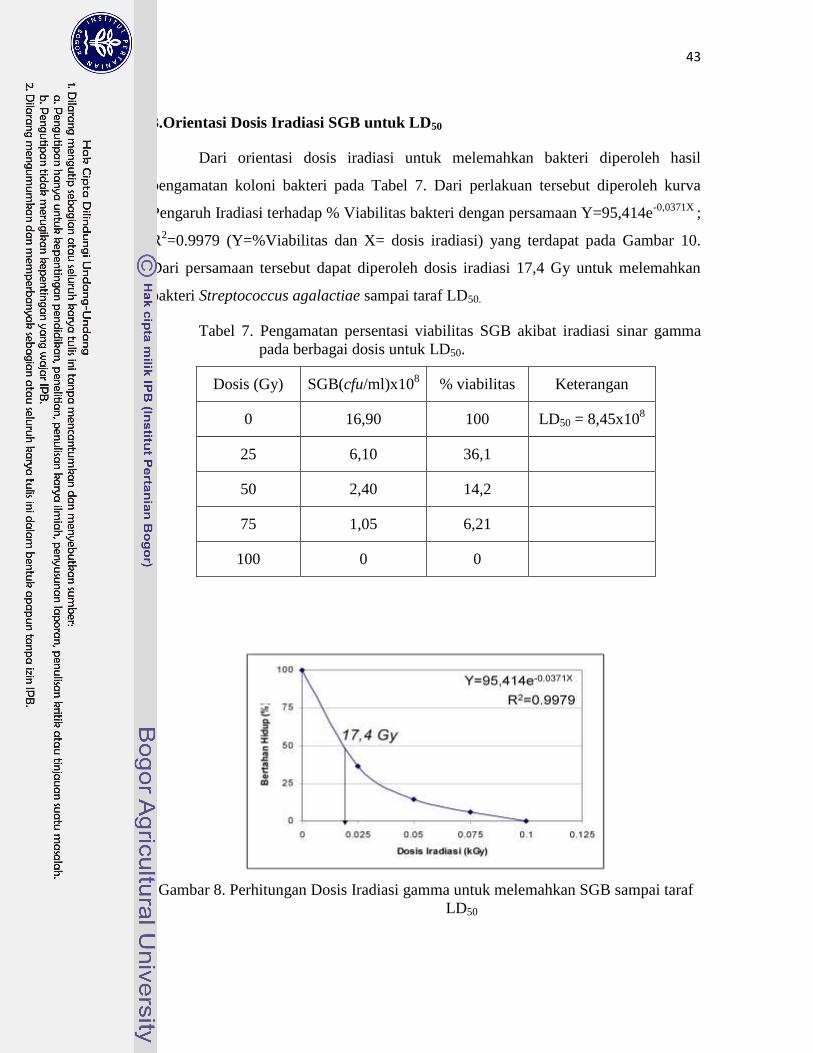
43
3.Orientasi Dosis Iradiasi SGB untuk LD50
Dari orientasi dosis iradiasi untuk melemahkan bakteri diperoleh hasil
pengamatan koloni bakteri pada Tabel 7. Dari perlakuan tersebut diperoleh kurva
Pengaruh Iradiasi terhadap % Viabilitas bakteri dengan persamaan Y=95,414e-0,0371X
;
R2=0.9979 (Y=%Viabilitas dan X= dosis iradiasi) yang terdapat pada Gambar 10.
Dari persamaan tersebut dapat diperoleh dosis iradiasi 17,4 Gy untuk melemahkan
bakteri Streptococcus agalactiae sampai taraf LD50.
Tabel 7. Pengamatan persentasi viabilitas SGB akibat iradiasi sinar gamma
pada berbagai dosis untuk LD50.
Dosis (Gy) SGB(cfu/ml)x108 % viabilitas Keterangan
0 16,90 100 LD50 = 8,45x108
25 6,10 36,1
50 2,40 14,2
75 1,05
6,21
100 0 0
Gambar 8. Perhitungan Dosis Iradiasi gamma untuk melemahkan SGB sampai taraf
LD50
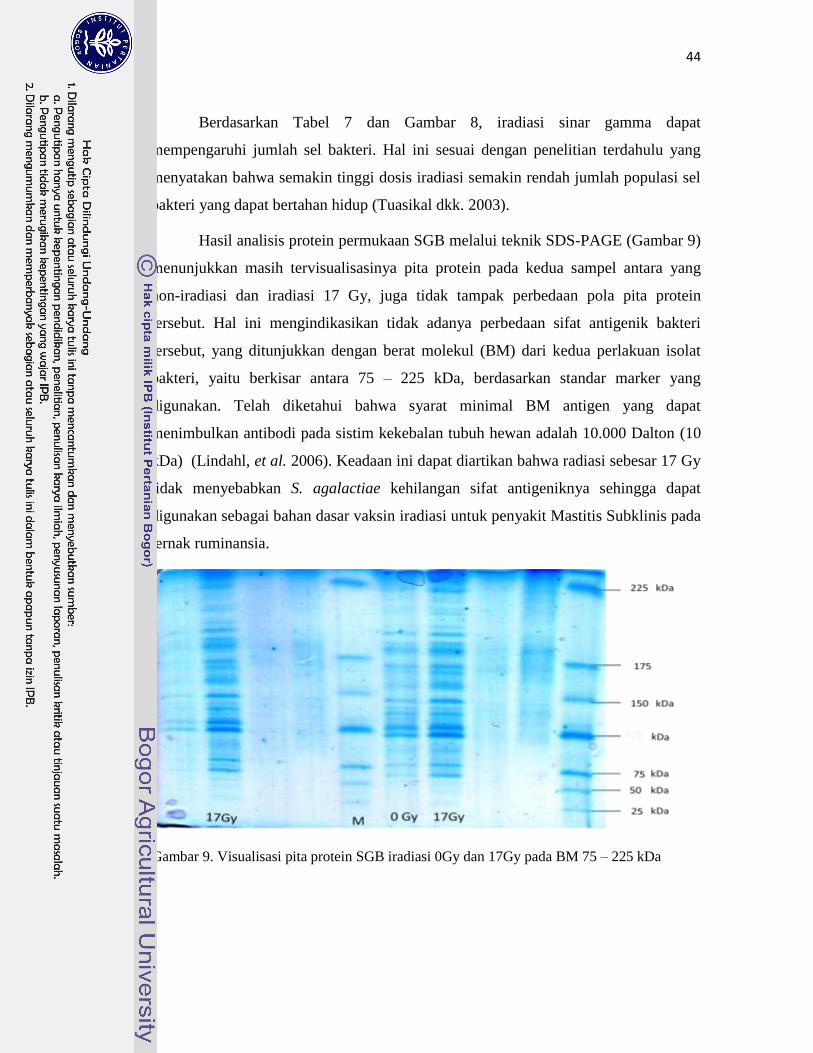
44
Berdasarkan Tabel 7 dan Gambar 8, iradiasi sinar gamma dapat
mempengaruhi jumlah sel bakteri. Hal ini sesuai dengan penelitian terdahulu yang
menyatakan bahwa semakin tinggi dosis iradiasi semakin rendah jumlah populasi sel
bakteri yang dapat bertahan hidup (Tuasikal dkk. 2003).
Hasil analisis protein permukaan SGB melalui teknik SDS-PAGE (Gambar 9)
menunjukkan masih tervisualisasinya pita protein pada kedua sampel antara yang
non-iradiasi dan iradiasi 17 Gy, juga tidak tampak perbedaan pola pita protein
tersebut. Hal ini mengindikasikan tidak adanya perbedaan sifat antigenik bakteri
tersebut, yang ditunjukkan dengan berat molekul (BM) dari kedua perlakuan isolat
bakteri, yaitu berkisar antara 75 – 225 kDa, berdasarkan standar marker yang
digunakan. Telah diketahui bahwa syarat minimal BM antigen yang dapat
menimbulkan antibodi pada sistim kekebalan tubuh hewan adalah 10.000 Dalton (10
kDa) (Lindahl, et al. 2006). Keadaan ini dapat diartikan bahwa radiasi sebesar 17 Gy
tidak menyebabkan S. agalactiae kehilangan sifat antigeniknya sehingga dapat
digunakan sebagai bahan dasar vaksin iradiasi untuk penyakit Mastitis Subklinis pada
ternak ruminansia.
Gambar 9. Visualisasi pita protein SGB iradiasi 0Gy dan 17Gy pada BM 75 – 225 kDa

45
Penelitian Hughes et al. (2002) berhasil mengidentifikasi 2 macam
antigen permukaan SGB dari kasus meningitis bayi yang baru lahir. Protein
permukaan SGB tersebut adalah ornithine carbamoyltransferase dan
phosphoglycerate kinase dengan visualisasi protein masing-masing 38 kDa dan
42 kDa. Antisera terhadap kedua protein ini pada hewan model mencit dapat
melindungi hewan baru lahir terhadap infeksi S. agalactiae.
Diketahui pula bahwa protein permukaan SGB bersifat imunoreaktif
yang dapat menimbulkan respon imun berupa Ig G1 pada sapi. Diduga protein
extraseluler ini mempunyai peranang penting dalam patogenesis S. agalactiae
dalam menginduksi mastitis (Trigo et al. 2008).
Park et al. (2002) mengidentifikasi Lactoferrin-Binding Protein (LBP)
Streptococcus spp. yang diisolasi dari sapi perah mastitis. LBP ini memegang
peranan dalam patogenesis mastitis sapi. Seluruh strain S. agalactiae yang
diteliti menunjukkan LBP dengan pita-pita protein dari 70 kDa sampai 112 kDa.
LBP dengan BM yang bersifat antigenik ini juga ideal untuk menjadi kandidat
vaksin di masa depan.
4.Hasil Uji Patogenitas dan Imunitas pada Hewan Model Mencit
Hasil pemeriksaan gambaran histopatologi pada kelenjar mamae mencit yang
diuji ditunjukkan pada Tabel 8. Uji patogenitas pada hewan percobaan mencit
menunjukkan bahwa SGB iradiasi tidak patogen, terbukti dari gambaran histopatologi
(HP) menunjukkan bahwa sel-sel pada alveol kelenjar mamae dari kelompok yang
diberi vaksin SGB iradiasi dan kelompok vaksin kemudian ditantang dengan SGB
tanpa iradiasi, masih memproduksi susu dan jumlah alveol kelenjar tidak berbeda
nyata dibanding dengan kelompok kontrol (mencit normal); jumlah alveol kelenjar
mamae yang paling sedikit berproduksi ada pada kelompok mencit yang ditantang
dengan SGB ganas tanpa divaksin. Vaksin SGB iradiasi juga tidak menimbulkan
peradangan pada kelenjar mamae mencit yang diuji, dibuktikan dari jumlah sel
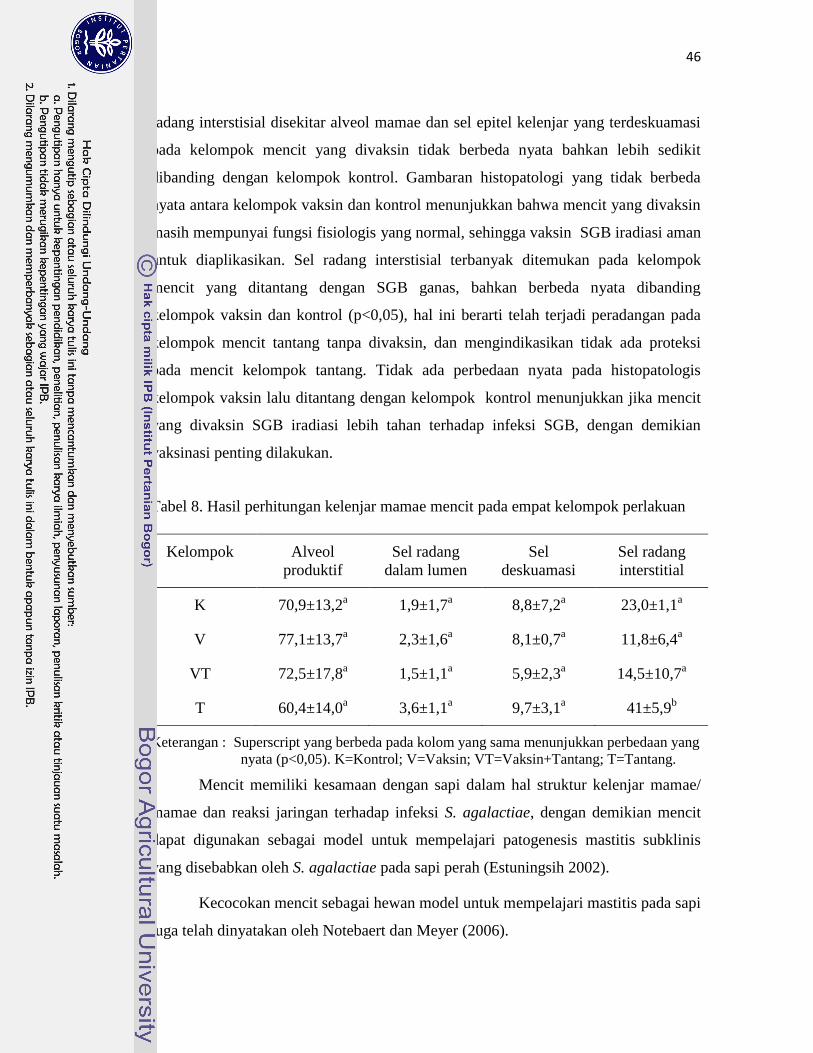
46
radang interstisial disekitar alveol mamae dan sel epitel kelenjar yang terdeskuamasi
pada kelompok mencit yang divaksin tidak berbeda nyata bahkan lebih sedikit
dibanding dengan kelompok kontrol. Gambaran histopatologi yang tidak berbeda
nyata antara kelompok vaksin dan kontrol menunjukkan bahwa mencit yang divaksin
masih mempunyai fungsi fisiologis yang normal, sehingga vaksin SGB iradiasi aman
untuk diaplikasikan. Sel radang interstisial terbanyak ditemukan pada kelompok
mencit yang ditantang dengan SGB ganas, bahkan berbeda nyata dibanding
kelompok vaksin dan kontrol (p<0,05), hal ini berarti telah terjadi peradangan pada
kelompok mencit tantang tanpa divaksin, dan mengindikasikan tidak ada proteksi
pada mencit kelompok tantang. Tidak ada perbedaan nyata pada histopatologis
kelompok vaksin lalu ditantang dengan kelompok kontrol menunjukkan jika mencit
yang divaksin SGB iradiasi lebih tahan terhadap infeksi SGB, dengan demikian
vaksinasi penting dilakukan.
Tabel 8. Hasil perhitungan kelenjar mamae mencit pada empat kelompok perlakuan
Kelompok Alveol
produktif
Sel radang
dalam lumen
Sel
deskuamasi
Sel radang
interstitial
K 70,9±13,2a 1,9±1,7
a 8,8±7,2
a 23,0±1,1
a
V 77,1±13,7a 2,3±1,6
a 8,1±0,7
a 11,8±6,4
a
VT 72,5±17,8a 1,5±1,1
a 5,9±2,3
a 14,5±10,7
a
T 60,4±14,0a 3,6±1,1
a 9,7±3,1
a 41±5,9
b
Keterangan : Superscript yang berbeda pada kolom yang sama menunjukkan perbedaan yang
nyata (p<0,05). K=Kontrol; V=Vaksin; VT=Vaksin+Tantang; T=Tantang.
Mencit memiliki kesamaan dengan sapi dalam hal struktur kelenjar mamae/
mamae dan reaksi jaringan terhadap infeksi S. agalactiae, dengan demikian mencit
dapat digunakan sebagai model untuk mempelajari patogenesis mastitis subklinis
yang disebabkan oleh S. agalactiae pada sapi perah (Estuningsih 2002).
Kecocokan mencit sebagai hewan model untuk mempelajari mastitis pada sapi
juga telah dinyatakan oleh Notebaert dan Meyer (2006).
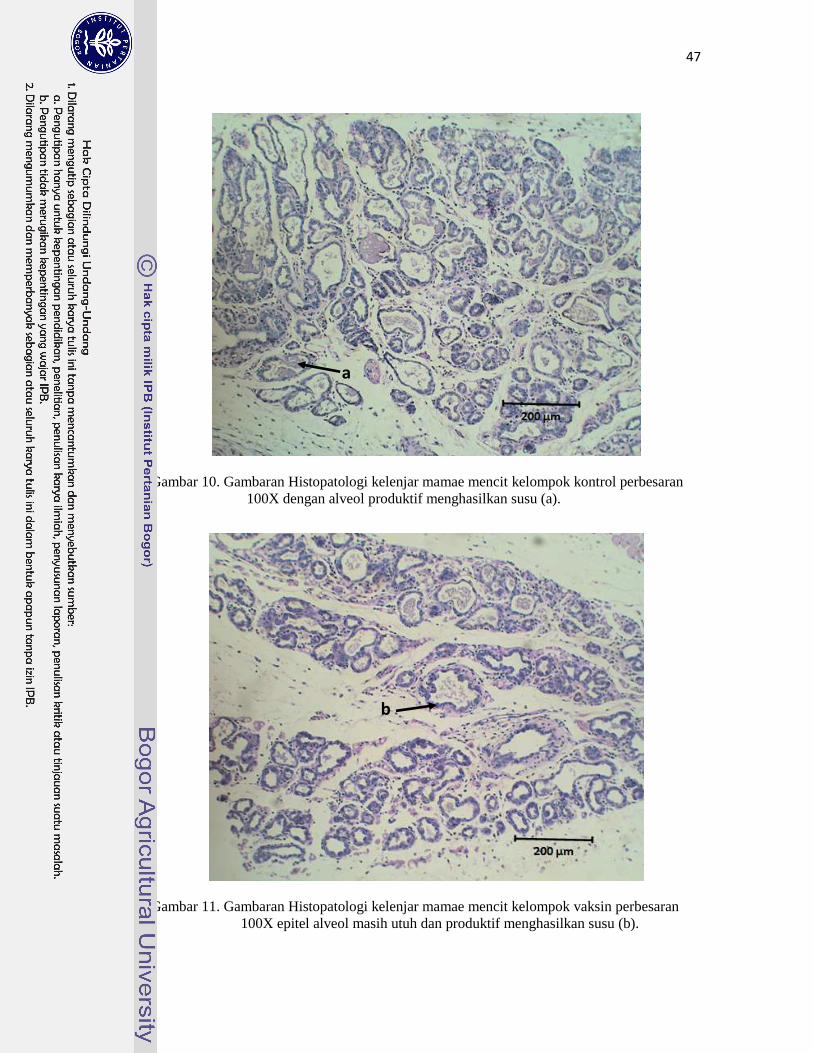
47
Gambar 10. Gambaran Histopatologi kelenjar mamae mencit kelompok kontrol perbesaran
100X dengan alveol produktif menghasilkan susu (a).
Gambar 11. Gambaran Histopatologi kelenjar mamae mencit kelompok vaksin perbesaran
100X epitel alveol masih utuh dan produktif menghasilkan susu (b).
a
b
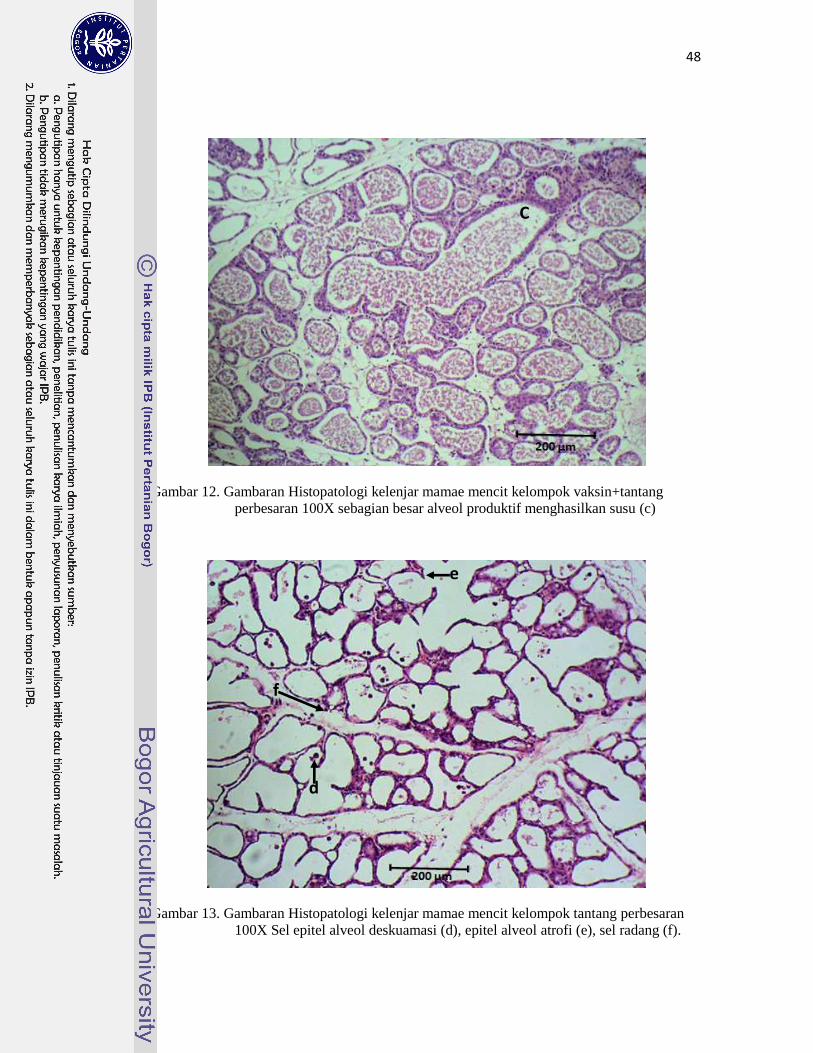
48
Gambar 12. Gambaran Histopatologi kelenjar mamae mencit kelompok vaksin+tantang
perbesaran 100X sebagian besar alveol produktif menghasilkan susu (c)
Gambar 13. Gambaran Histopatologi kelenjar mamae mencit kelompok tantang perbesaran
100X Sel epitel alveol deskuamasi (d), epitel alveol atrofi (e), sel radang (f).
C
d
e
f

49
Gambaran Histopatologi kelenjar mamae mencit kelompok vaksin (Gambar
10) mirip dengan kelompok kontrol (Gambar 11) yaitu mencit normal. Mencit-mencit
dari kedua kelompok perlakuan ini sama-sama mempunyai epitel alveol yang masih
utuh dan produktif menghasilkan susu. Hal tersebut membuktikan jika pemberian
vaksin iradiasi SGB masih aman dilakukan karena kelompok mencit yang divaksin
menunjukkan fungsi fisiologis normal sebagaimana kelompok mencit kontrol.
Gambaran Histopatologi kelenjar mamae mencit kelompok vaksin kemudian
diberi tantangan dengan SGB ganas atau tidak diradiasi (Gambar 12) menunjukkan
bahwa sebagian besar alveol kelenjar masih produktif menghasilkan susu. Hal
tersebut menunjukkan bahwa mencit yang divaksin dengan SGB iradiasi dapat
menahan infeksi tantangan dan tetap memproduksi susu.
Gambaran Histopatologi kelenjar mamae mencit kelompok yang tidak
divaksin tetapi hanya diberikan uji tantang dengan SGB ganas tanpa iradiasi (Gambar
13) memperlihatkan kerusakan berupa atrofi epitel alveol dan berdeskuamasinya sel
epitel ke dalam lumen alveol. Hal tersebut terjadi akibat peradangan yang
ditunjukkan dengan sel-sel radang yang terlihat baik di dalam lumen alveol maupun
interstisial. Keadaan patologis pada kelompok tantang ini mengakibatkan alveol tidak
dapat berproduksi dan tampak lumen alveol banyak yang tidak berisi susu.
Hasil pemeriksaan Ig-G serum mencit dengan ELISA (Gambar 14.)
menunjukkan tingkat respon imun yang lebih tinggi pada kelompok mencit yang
divaksin dibandingkan kelompok kontrol, hal tersebut membuktikan bahwa SGB
yang telah dilemahkan dengan iradiasi sinar gamma masih mampu menimbulkan
respon imun. Gambar 15 juga menunjukkan bahwa titer Ig-G pada kelompok Vaksin
dan tantang masih lebih tinggi dari pada kelompok kontrol.
Hewan model mencit dapat menunjukkan respon imun bawaan maupun
dapatan terhadap Streptococcus agalactiae penyebab mastitis (Trigo et al., 2009).
Hewan model mencit juga digunakan untuk uji vaksin rekombinan S. agalactiae dan
S. aureus dari kasus mastitis sapi. Uji ELISA serum mencit menunjukkan antibodi
dominan berasal dari subtipe Ig G1 (Xu et al. 2011).
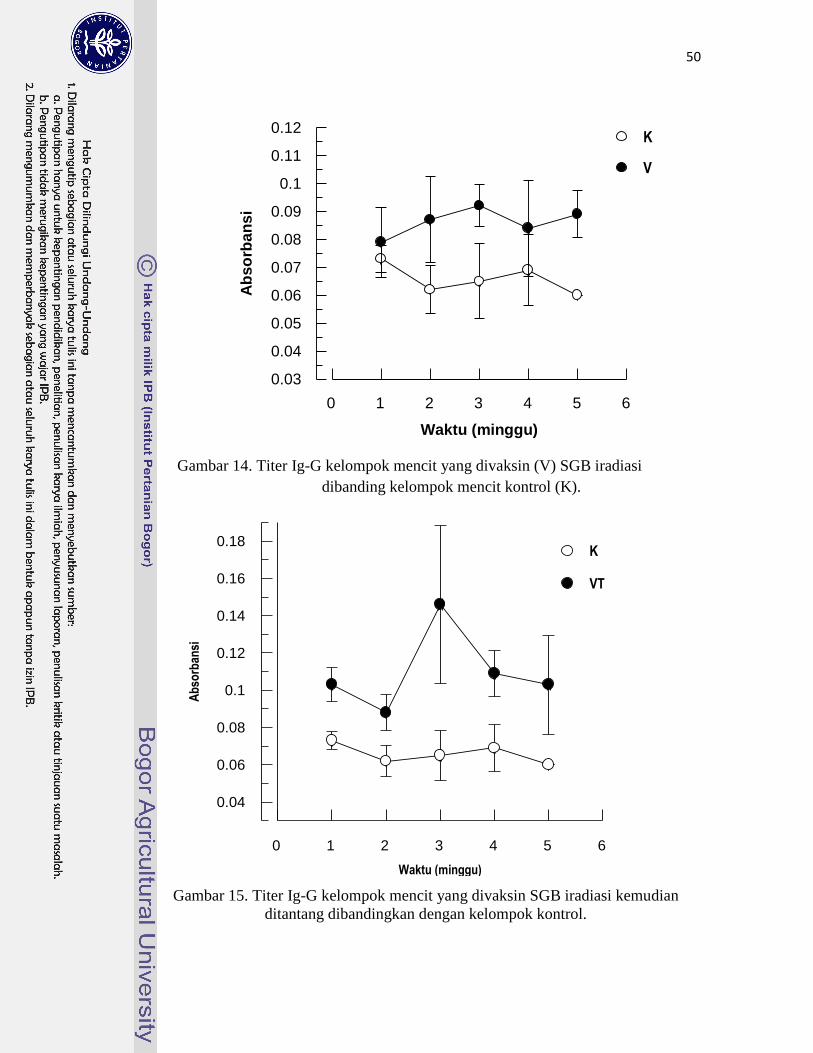
50
Gambar 14. Titer Ig-G kelompok mencit yang divaksin (V) SGB iradiasi
dibanding kelompok mencit kontrol (K).
Gambar 15. Titer Ig-G kelompok mencit yang divaksin SGB iradiasi kemudian
ditantang dibandingkan dengan kelompok kontrol.
6543210
0.12
0.11
0.1
0.09
0.08
0.07
0.06
0.05
0.04
0.03
Waktu (minggu)
Ab
so
rba
ns
i
K
V
6543210
0.18
0.16
0.14
0.12
0.1
0.08
0.06
0.04
Waktu (minggu)
Ab
sorb
ansi
K
VT
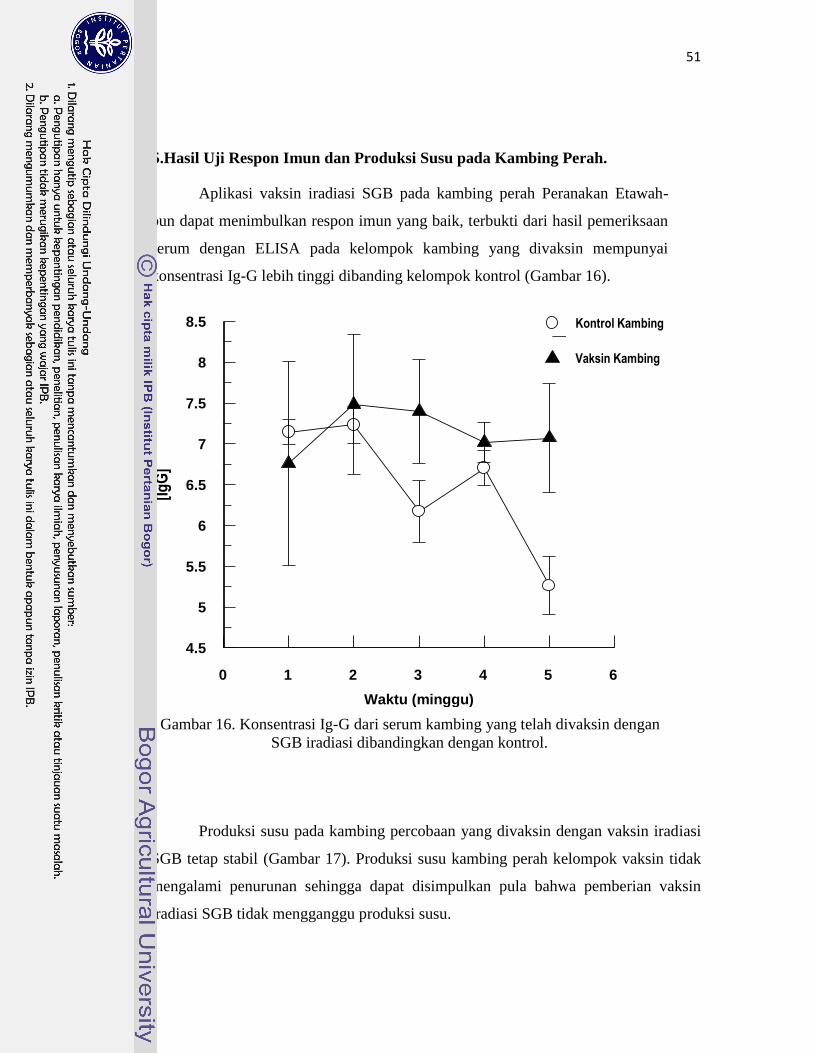
51
5.Hasil Uji Respon Imun dan Produksi Susu pada Kambing Perah.
Aplikasi vaksin iradiasi SGB pada kambing perah Peranakan Etawah-
pun dapat menimbulkan respon imun yang baik, terbukti dari hasil pemeriksaan
serum dengan ELISA pada kelompok kambing yang divaksin mempunyai
konsentrasi Ig-G lebih tinggi dibanding kelompok kontrol (Gambar 16).
Gambar 16. Konsentrasi Ig-G dari serum kambing yang telah divaksin dengan
SGB iradiasi dibandingkan dengan kontrol.
Produksi susu pada kambing percobaan yang divaksin dengan vaksin iradiasi
SGB tetap stabil (Gambar 17). Produksi susu kambing perah kelompok vaksin tidak
mengalami penurunan sehingga dapat disimpulkan pula bahwa pemberian vaksin
iradiasi SGB tidak mengganggu produksi susu.
6543210
8.5
8
7.5
7
6.5
6
5.5
5
4.5
Waktu (minggu)
[Ig
G]
Kontrol Kambing
Vaksin Kambing
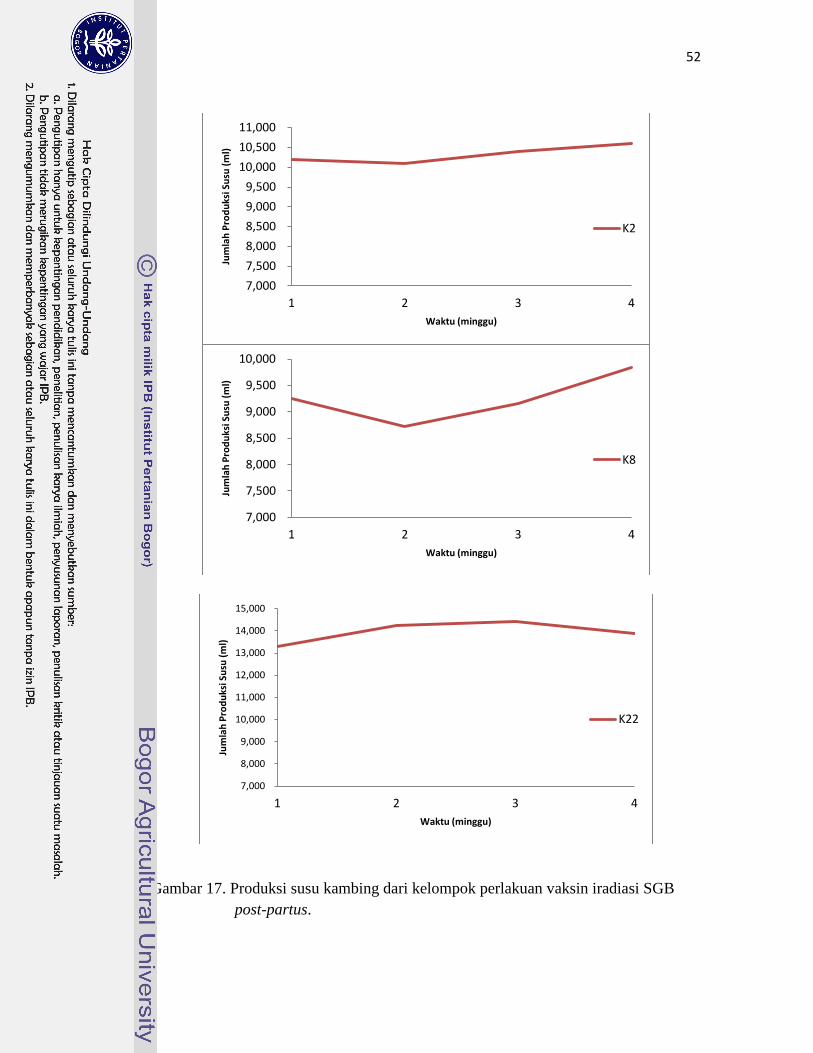
52
Gambar 17. Produksi susu kambing dari kelompok perlakuan vaksin iradiasi SGB
post-partus.
7,000
7,500
8,000
8,500
9,000
9,500
10,000
10,500
11,000
1 2 3 4
Jum
lah
Pro
du
ksi S
usu
(m
l)
Waktu (minggu)
K2
7,000
7,500
8,000
8,500
9,000
9,500
10,000
1 2 3 4
Jum
lah
Pro
du
ksi S
usu
(m
l)
Waktu (minggu)
K8
7,000
8,000
9,000
10,000
11,000
12,000
13,000
14,000
15,000
1 2 3 4
Jum
lah
Pro
du
ksi S
usu
(m
l)
Waktu (minggu)
K22

53
Penanganan kasus mastitis pada kambing perah pada intinya sama dengan
pengendalian mastitis pada sapi perah. Deteksi MSK juga dilakukan dengan cara tak
langsung seperti uji dengan CMT maupun penghitungan sel somatik (Shearer and
Haris 2003). Kambing perah dapat digunakan sebagai hewan model yang paling tepat
sebelum dilakukan uji lapang pada sapi perah.

54

55
SIMPULAN DAN SARAN
1. Simpulan
Berdasarkan hasil penelitian ini telah diperoleh kandidat vaksin iradiasi S.
agalactiae yang termasuk kedalam Group-B (SGB) tanpa kapsul dengan
hemaglutinin positif (Hn+) yang dapat digunakan sebagai bahan vaksin mastitis
subklinis. Dapat disimpulkan pula bahwa iradiasi sinar gamma dosis 17 Gy dapat
melemahkan Isolat SGB bahan vaksin hidup tersebut sehingga tidak bersifat patogen
dan masih mempunyai sifat antigenik dengan berat molekul lebih besar dari 75 kDa.
Vaksin iradiasi SGB tidak bersifat patogen pada mencit dan dapat menimbulkan
respon imun yang lebih baik dibanding kelompok mencit kontrol yang tidak divaksin.
Aplikasi vaksin iradiasi SGB pada kambing perah dapat menimbulkan respon imun,
sehingga besar harapan untuk dapat membuat vaksin iradiasi SGB dalam mencegah
mastitis subklinis pada ternak ruminansia. Produksi susu kambing perah kelompok
vaksin tidak mengalami penurunan sehingga dapat disimpulkan pula bahwa
pemberian vaksin iradiasi SGB tidak mengganggu produksi susu.
2. Saran
Saran dari penelitian ini adalah perlu dilakukan uji lapang aplikasi vaksin
SGB iradiasi pada hewan ruminansia khususnya sapi perah.

56

57
DAFTAR PUSTAKA
Alsharifi M and Mullbacher A. 2009. The γ-irradiated influenza vaccine and the
prospect of producing safe vaccines in general. Immunol Cell Biol: 1-2.
Abrar M. 2001. Isolasi, karakterisasi dan aktifitas biologi hemaglutinin
Staphylococcus aureus dalam proses adhesi pada permukaan sel epitel mamae
sapi perah. IPB. Bogor.
Abubakar M, Muhammad G, Ibrahim K. 2006. Primary and secondary response to
formalin inactivated Streptococcus agalactiae isolates in rabbits. Pakistan Vet J
26(3): 115-117.
Akers RM and Nickerson SC. 2011. Mastitis and its impact on structure and function
in the ruminant mammary gland. J Mammary Gland Biol. Neoplasia. Springer.
Publish online 04 October 2011.
Alatas Z. 2007. Efek kesehatan pajanan radiasi. Cermin Dunia Kedokteran No. 154.
Allen-Mierl D, Buxton R, Jones LK, Madl D, Mason A, Malendez J, Milligan P.
2006. http://www.microbelibrary.org/component/resource/laboratory-test/3086-
camp-test-protocols [27Juni2010]
Arifin M. 2006. Tanggap kebal sapi terhadap Fasciolosis akibat inokulasi
metaserkaria Fasciola gigantica iradiasi. J Apl Isotop dan Radiasi. 2(1): 18–29.
Baratawidjaya KG dan Rengganis I. 2009. Imunologi Dasar. Ed 8. Balai Penerbit.
Fakultas Kedokteran Universitas Indonesia. Jakarta.
Bogni, C, Ordieno L, Raspanti C, Giraudo J, Larriestra A, Reinoso E, Lasagno M,
Ferrari M, Ducrós E, Frigerio C, Bettera S, Pellegrino M, Frola I, Dieser S,
Vissio C. 2011. “War against mastitis: Current concepts on controlling bovine
mastitis pathogens.”, Science against pathogens: communicating current
research and technological advances., A. Mendez-Vilaz (Ed.) : 483 – 494,
Cordoba Argentina.
Bruckner DA and Colonna P. 1997. Nomenclatur for Aerobic and Facultative
Bacteria. Clin Infect Dis 25 : 1 – 10.
Carter GR. 1984. Diagnostic Procedures in Veterinary Bacteriology and Mycology.
Publisher Springfield. Illinois. 4th
Ed.
Chanter N. 1997. Streptococci and Enterococci as animal pathogens. J Applied
Microbiol. Symposium Supplement. 83 : 100S – 109S.
Cowan ST. 2003. Manual for The Identification of Medical Bacteria. Cambridge
University Press. Cambridge London New York Melbourne Sydney. pp: 52-66.
Crowther JR. 2010. The ELISA Guidebook, 2nd
ed., Humana Press, International
Atomic Czarnecki L. 2009. Nuclear physics. http://www.hpwt.de/Kern2e.htm.
[12 Juni 2011].

58
Daignault D. 2003. Serotype of Streptococcus agalactiae cultured from dairy milk
sample in Quebec. Can Vet J 44:217-220.
Darling CL. 1975. Standardization and evaluation of CAMP reaction for the prompt,
presumptive identification of Streptococcus agalactiae (Lancefield Group B) in
clinical material. J Clin Microbiol : 171 – 174.
De Laval, 2008, Milking Technology, www.delaval.com /.../ Milking _ Technology.
htm [27 Juni 2009].
Dian AS, Lusia W, Franky, Rosana SD, Riyan P. 2010. Karakterisasi kemampuan
adesi bakteri penyebab mastitis subklinis pada sapi perah serta peluang
pembuatan anti adesi untuk pencegahannya. J Saintifika Vol.II No.2 : 48 – 54.
[Dirjenak & Keswan]. 2011. Statistik Peternakan dan Kesehatan Hewan. Direktorat
Jenderal Peternakan dan Kesehatan Hewan. Kementerian Pertanian Republik
Indonesia.
[Dirkeswan] 2001. Manual Penyakit Hewan Mamalia: Mastitis. Dirjen Bina Produksi
Peternakan. Direktorat Kesehatan Hewan. Departemen Pertanian:175-177.
Duval J. 1997. Treating mastitis without antibiotics. Ecological Agriculture Projects.
http://www.eap.mcgill.ca/Publications/EAP69.htm. [15 Juli 2010].
Estuningsih S, Soedarmanto I, Fink K, Lämmler C, Wibawan IWT. 2002. Studies on
Streptococcus agalactiae isolated from bovine mastitis in Indonesia, J Vet Med
49 (4): 185-187.
Estuningsih S. 2002. Patogenesis mastitis subklinis pada sapi perah: Pendekatan
histopatologis mastitis subklinis akibat infeksi Streptococcus agalactiae
hemaglutinin positif pada mencit, Disertasi Doktor Pascasarjana, IPB.
Evans JJ, Pasnik DJ, Klesius PH, Al-Ablani S. 2006. First Report of Streptococcus
agalactiae and Lactococcus garvieae from a wild bottlenose dolphin (Tursiups
truncatus). J Wildlife Dis 42(3) : 561–569.
Facklam RR, Padula JF, Wortham EC, Cooksey RC, Rountree HA. 1979.
Presumptive identification of Group A, B, and D Streptococci on Agar Plate
Media. J Clin Microbiol: 665 – 672.
Furuya Y, Regner M, Lobigs M, Koskinen A, Mullbacher A, Alsharifi M. 2010.
Effect of inactivation method on the cross-protective immunity induced by
whole „killed‟ influenza A viruses and commercial vaccine preparations. J Gen
Virol (91): 1450-1460.
Hall EJ, Giaccia AJ. 2006. Radiobiology for the Radiologist. 6th
ed. Philadelphia, PA:
Lippincott Williams & Wilkins.
Hameed KGA, Sender G, Korwin-Kossakowska A. 2007. Public health hazard due to
mastitis in dairy cow. Animal Science Papers and Report vol.25, no.2: 73-85.

59
Hamman J. 2005. Diagnosis of mastitis and indicators of milk quality. Proceeding of
IDF Congress on Mastitis in Dairy Production: Current Knowledge and Future
Solutions. H. Hogeveen (Ed.). Wageningen Academic Publishers, The
Netherlands p. 89 – 90.
Hansen SM and Sørensen UBS. 2003. Method for quantitative detection and
presumptive identification of Group B Streptococci on primary platting. J Clin
Microbiol. Vol.41 No.4: 1399 – 1403.
Harlina E. 1999. Studi Patogenitas Fase Varian Streptococcus equi subsp.
Zooepidimicus. Penyebab Wabah Penyakit pada Babi di Indonesia Berdasarkan
Gambaran Patologi dan Virulensinya [Tesis]. Program Pascasarjana, Institut
Pertanian Bogor. Bogor.
Hashemi M, Kafi M, Safdarian M. 2011. The prevalence of clinical and subclinical
mastitis in dairy cows in the central region of fars province, South of Iran.
Iranian J Vet Res, Shiraz University 12 (3) Ser. No. 36: 236 – 241.
Hayati Z, Karmil TF. 2009. Patogenitas streptokakus grup B pada mencit neonatus.
J Vet 10(4):184-188.
Henrichen G, Ferrieri P, Jelinkova J, Kohler W, Maxted WR. 1984. Nomenclature of
antigens of group B streptococci. Int J Syst Bacteriol 34:500.
Hughes MJG, Moore JC, Lane JD, Wilson R, Pribul PK, Younes ZN, Dobson RJ,
Everest P, Reason AJ, Redfern JM, Greer FM, Paxton T, Panico M, Morris
HR, Feldman RG, Santangelo JD. 2002. Infect Immun pp: 1254-1259.
Hurley WL and Morrin DE. 2000. Mastitis Lesson A. Lactation Biology, ANSCI
308, http://classes aces.uiuc.edu/Ansci 308 [27 Juli 2009].
Keefe GP, Dohoo IR, Spangler E. 1997. Physiology and Management: Herd
prevalence and incidence of Streptococcus agalactiae in the dairy industry of
Prince Edward island. J Dairy Sci 80 : 464 – 470.
Keefe GP. 1997. Streptococcus agalactiae mastitis: A Review. Can Vet J (38): 429-
437.
Kiernan JA. 1990. Histological and Histochemical Method: Theory and Practice. 2nd
ed. UK. Pergenan Press.
Kochman M. 2006. Gamma radiation proves effective in vaccine development.
BioPharm Bulletin. http://biopharminternational.findpharma.com. [5 Februari
2011].
Lämmler C, Wibawan IWT, Pasaribu FH. 1998. Relation between encapsulation of
streptococci of serogical group B and adherence properties of the bacteria to
DEAE-sephacel. Media veteriner 5(4):1-5.
Lang S. and Palmer M. 2003. Characterization of Streptococcus agalactiae CAMP
Factor as a Pore-forming Toxin. J Biol Chem 278 (10): 38167 – 38173.

60
Lay BW. 1994. Analisis Mikroba di Laboratorium. Jakarta: Rajawali Press
Lindahl G, Carlemalm MS, Areschoug T. 2006. Surface proteins of Streptococcus
agalactiae and related proteins in Other bacterial pathogens. Clin Microbiol Rev:
102-127.
Lukman DW, Sudarwanto M, Sanjaya AW, Purnawarman T, Latief H, Soejoedono
RR. 2009. Higiene Pangan. Fakultas Kedokteran Hewan. Institut Pertanian
Bogor.
Malinowski E, Lassa H, Klosowska A, Markiewicz H, Kaczmarowski M, Smulski S.
2006. Relationship between mastitis agents and Somatic Cell Count in Foremilk
Samples. Bull Vet Inst Pulawy 50 : 349 – 352.
Maricato E, Lange CC, Brito MAVP, Brito JRF, Cerqueira MMOP. 2005.
Characterization and antibiotic susceptibility patterns of catalase-negative Gram-
positive Cocci isolated from bovine mastitis in Brazil. ISAH – Warsaw Poland
Vol 1: 288-291.
Mirdhayati I, Handoko J, Putra KU. 2008. Mutu susu segar di UPT Ruminansia Besar
Dinas Peternakan Kabupaten Kampar Provinsi Riau. J Peternakan 5 (1): 14 –
21.
Moatamedi H, Shapouri, Ghorbanpoor M, Jamshidin M, Gooraninejad S. 2007. A
Polymerase Chain Reaction based study on the subclinical mastitis caused by
Streptococcus agalactiae, S. dysgalactiae and S. uberis in Cattle in Ahvaz.,
Iranian J Vet Res University of Shiraz Vol.8, No.3, Ser. No. 20: 260-265.
Notebaert S and Meyer E. 2006. Mouse models to study the pathogenesis and control
of bovine mastitis. A review. Vet Quart 28: 12 -13.
Park HM, Almeida RA, Oliver SP. 2002. Identification of Lactoferin-Binding
Proteins in Streptococcus species Isolated from Cows with Mastitis. FEMS
Microbiol Letters vol 207 Issue 1 pp:87-90.
Pasaribu FH, Wibawan IWT, Warsa UC. 1993. Distribusi tipe antigen Streptococcus
agalactiae isolat sapi dan manusia di Jakarta, Bogor dan Sekitarnya.
Disampaikan dalam KONAS Permi ke-VI di Surabaya.
Petrovski KR and Emanuel S. 2006. Milk composition changes during mastitis.
Published in http://www.milkproduction.com/Library/Articles/Milk composition
changes during mastitis [3/17/2009].
Quin PJ, Markey BK, Carter MF, Donnelly WJC, Leonard FC. 2006. Veterinary
Microbiology and Microbial Disease. Section II Pathogenic Bacteria:
Streptococci. pp: 49-54. Blackwell Science Asia Pty.Ltd. Victoria. Australia.
Reitman M, Tribble HR Jr, Green L. 1970. Gamma-irradiated Venezuelan Equine
Encephalitis Vaccines. App Microbiol: 763-767.

61
Rainard P, Lautrou Y, Sarradin P, Poutrel B. 1991. Protein X of Streptococcus
agalactiae induces opsonic antibodies in cows. J Clin Microbiol 29(9): 1842-
1846.
Rinaldi M, Li RW, Capuco AV. 2010. Mastitis associated transcriptomic disruption
in cattle. Vet Immunol and Immunopathol 138 : 267 -279.
Ruegg, P. L. 2005. California Mastitis Test (CMT) Fact Sheet 2.
http://www.uwex.edu/milkuality/PDF/Vol 3 pdf/Pg 3 16-
18_CMT_factsheet_1.pdf [28 Juli 2010].
Sanakkayala N et al. 2005. Induction of Antigen-Specific Th1 Type Immune
Responses by Gamma-Irradiated Recombinant Brucella abortus RB51. Clin
Diagn Lab Immunol 12 (12) : 1429–1436.
Seegers H, Fourichon C, Beaudeu F. 2003. Production effect related to mastitis and
mastitis economics in dairy cattle herds. Vet Res 34: 475 – 491.
Shearer JK and Harris B Jr. 2003. Mastitis in dairy goats. University of Florida, Ifas
extention. http://edis.ifas.ufl.edu. [28 Juli 2010].
Shim EH, Shanks RD, Morin DE. 2004. Milk loss and treatment cost associated with
two treatment protocols for clinical mastitis in dairy cows. J Dairy Sci 87: 2702
– 2708.
Soeminto B. 1989. Manfaat Tenaga Atom untuk kesejahteraan manusia: Radiasi.
Karya Indah: 32-36.
Sori H, Zerihum A, Abdicho S. 2005. Dairy cattle mastitis in and around Sebeta
Ethiopia. Int J Appl Res Vet Med 3 : 332-338.
Subandriyo dan Adiarto. 2009. Sejarah perkembangan peternakan sapi perah. Profil
Usaha Peternakan Sapi Perah di Indonesia, Pusat Penelitian dan Pengembangan
Peternakan. Badan Penelitian dan Pengembangan Pertanian Bogor : 1 – 25.
Subronto. 2003. Ilmu Penyakit Ternak I. Edisi Kedua. Gadjah Mada University Press.
Yogyakarta : 309 – 351.
Sudarwanto M. 1999. Usaha peningkatan produksi susu melalui program
pengendalian mastitis subklinik. Orasi Ilmiah, 22 Mei 1999. FKH – IPB.
Sudarwanto M., Latif MH, Noordin M. 2006. The Relationship of the somatic cell
counting to subclinical mastitis and to improve milk quality., 1st International
AAVS Scientific Conference. Jakarta, July 12-13, 2006.
Sudarwanto M dan Sudarnika E. 2008. Hubungan antara pH susu dengan Jumlah Sel
Somatik sebagai parameter mastitis subklinis. Media Peternakan: 107-113
Supar. 1997. Mastitis subklinis pada sapi perah di Indonesia: masalah dan
pendekatannya, Wartazoa 6 (2) : 48 – 52.

62
Supar, Ariyanti T. 2008. Kajian pengendalian mastitis subklinis pada sapi perah.
Pros. Semiloka Nas. Prospek Industri Sapi Perah Menuju Perdagangan Bebas –
2020. PUSLITBANGNAK : 360 – 366.
Timoney JF, Kumar P. 2010. Streptococcus. Di dalam Carlton LG, JF Prescott, G
songer, Charles OT, editor. Pathogenesis Of Bacterial Infections In Animals
4th Edition. Danvers-Massachusetts USA: Blackwell Publishing. Hlm 51-74
Tizard I.1988. Pengantar Imunologi Veteriner. Edisi ke 2. Airlangga University
Press.
Trampuz A, Piper KE, Steckelberg JM, Patel R. 2006. Effect of gamma irradiation on
viability and DNA of Staphylococcus epidermidis and Escherichia coli. J
Med Microbiol 55: 1271-1275.
Trigo G, Ferreira P, Ribeiro N, Dinis M, Andrade EB, Cristino JM, Ramirez M,
Tavares D. 2008. Identifications of immunoreactive extracelluler proteins of
Streptococcus agalactiae in bovine mastitis. Canadian J Microbiol 54 (11):
899-905.
Trigo G, N, Dinis M, Franca A, Andrade EB, Rui M, da Costa G, Ferreira P, Tavares
D. 2009. Leukocyte populations and cytokine expression in the mammary gland
in a mouse model of Streptococcus agalactiae matitis. J Med Microbiol 58: 951
– 958.
Tuasikal BJ, Sugoro I, Tjiptosumirat T, Lina M. 2003. Pengaruh iradiasi sinar gamma
pada pertumbuhan Streptococcus agalactiae sebagai bahan vaksin penyakit
Mastitis pada sapi perah. J Sains Tek Nuklir Ind Vol. IV, Edisi Khusus (2): 137-
149.
Tubiana M, Dutreix J, Wambersie A. 1990. Introduction to Radiobiology. London :
Taylor & Francis.
Walker J. M. 2002. The Protein Protocols Handbook: Human Press. Second ed.
Wahyuni AETH, Wibawan IWT, Wibowo MH. 2005. Karakterisasi hemaglutinin
Streptococcus agalactiae dan Staphylococcus aureus penyebab mastitis
subklinis sapi perah. J Sain Vet 23 (2): 79-86.
Wahyuni AETH, Wibawan IWT, Pasaribu FH, Priosoeryanto BP. 2006. Distribusi
serotipe Streptococcus agalactiae penyebab mastitis subklinis pada sapi perah
di Jawa Timur, Jawa Tengah dan Jawa Barat. J Vet Vol 7 (1).
[WHO] World Health Organization. 2001. Malaria early warning systems – concepts,
indicators and partners. A framework for field research in Africa. Geneva : The
organization.
Wibawan IWT, Lämmler C. 1990. Properties of group B Streptococci with protein
surface antigens X and R. J Clin Microbiol 28(12):2834-2836.

63
Wibawan IWT, Lämmler C. 1991. Influence of capsuler neuramnic acid on
properties of Streptococci of serogical group B. J Gen Microbiol 137:2721-
2725.
Wibawan IWT, Lammler C, Pasaribu FH. 1992. Role of hydrophobic surface proteins
in mediating adherence of group B streptococci to epithelial cells. J Gen
Microbiol 138 (6): 1237-1242.
Wibawan IWT, Lammler C, Seleim R, Pasaribu FH. 1993. A haemaglutinin adhesin
of group B Streptococci isolated from cases of bovine mastitis mediates
adherence to Hela cells. J Gen Microbiol 139(9): 2173-2178.
Wibawan IWT. 1998. The possibility of using vaccine to control bovine subclinical
mastitis and human neonatal infection caused by group B Streptococci. Media
Veteriner 5: 1-6.
Wu J, Hu S, Cao L. 2007. Therapeutic effect of Nisin Z on subclinical mastitis in
lactating cows., Antimicrob Agents and Chemoth 51 (9) : 3131 – 3135.
Xu H, Hu C, Gong R, Chen Y, Ren N, Xiao G, Xie Q, Zhang M, Liu Q, Guo A, Chen
H. 2011. Clin Vacc Immunol 18(6): 893-900.

64

65
LAMPIRAN

66
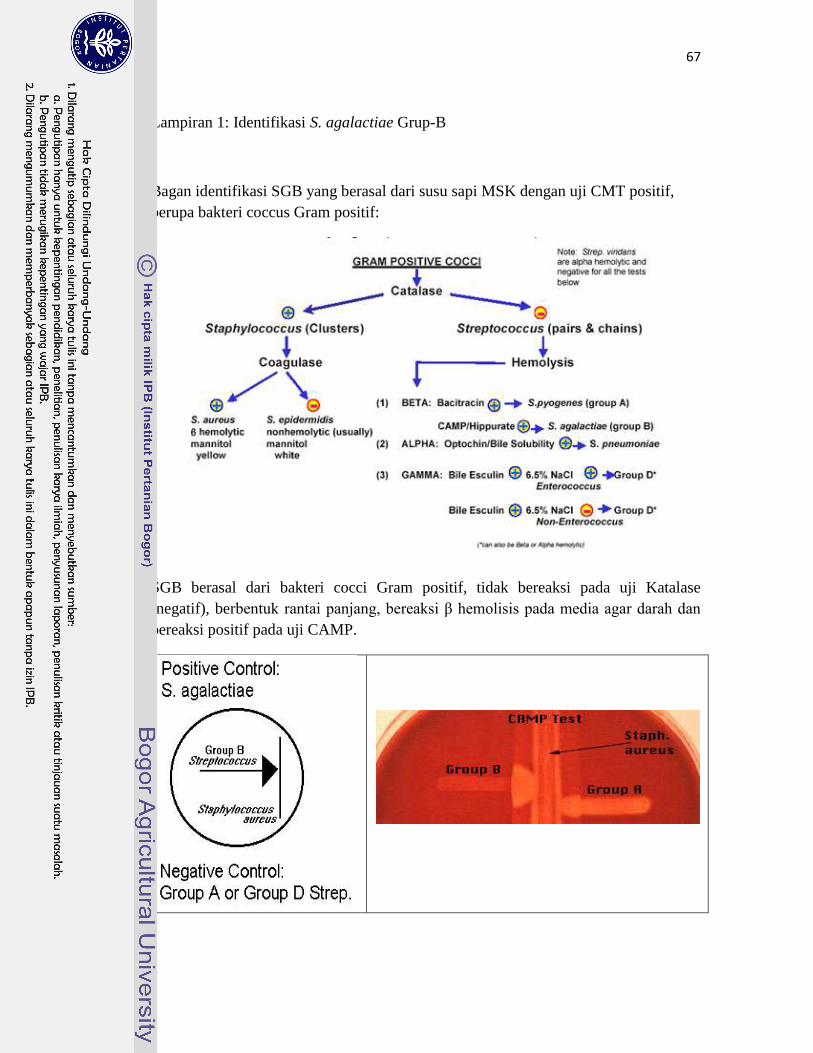
67
Lampiran 1: Identifikasi S. agalactiae Grup-B
Bagan identifikasi SGB yang berasal dari susu sapi MSK dengan uji CMT positif,
berupa bakteri coccus Gram positif:
SGB berasal dari bakteri cocci Gram positif, tidak bereaksi pada uji Katalase
(negatif), berbentuk rantai panjang, bereaksi β hemolisis pada media agar darah dan
bereaksi positif pada uji CAMP.

68
Lampiran 2. Karakterisasi SGB tanpa kapsul, keberadaan antigen protein dan
Hemaglutinin positif (Hn+).
Karakterisasi SGB isolat no.2 dengan kode isolat EK1:
1. Karakter tanpa kapsul ditunjukkan dengan pola pertumbuhan kompak pada
media soft agar (SA) di tabung kanan.
2. SGB dengan antigen protein tumbuh dengan supernatan jernih dan
membentuk sedimen di dasar tabung pada media cair Todd Hewitt Broth.
3. Hemaglutinin SGB menunjukkan jika bersifat adhesi pada kelenjar mamae
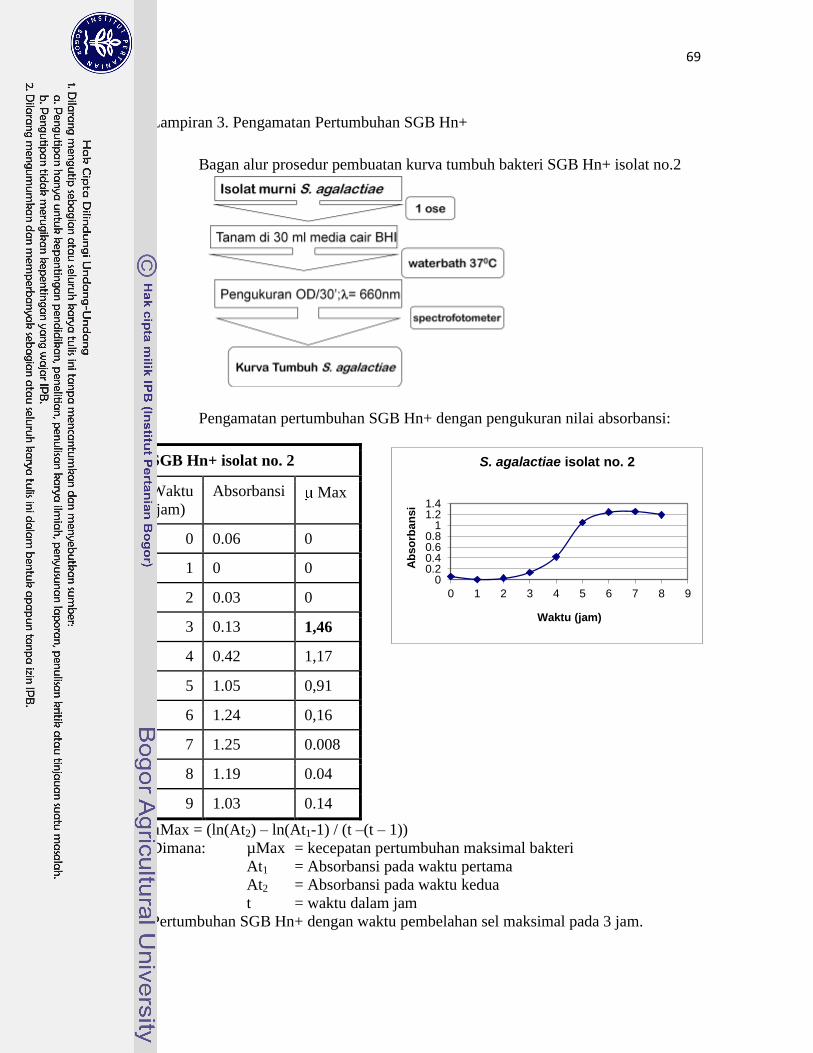
69
Lampiran 3. Pengamatan Pertumbuhan SGB Hn+
Bagan alur prosedur pembuatan kurva tumbuh bakteri SGB Hn+ isolat no.2
Pengamatan pertumbuhan SGB Hn+ dengan pengukuran nilai absorbansi:
SGB Hn+ isolat no. 2
Waktu
(jam)
Absorbansi Max
0 0.06 0
1 0 0
2 0.03 0
3 0.13 1,46
4 0.42 1,17
5 1.05 0,91
6 1.24 0,16
7 1.25 0.008
8 1.19 0.04
9 1.03 0.14
µMax = (ln(At2) – ln(At1-1) / (t –(t – 1))
Dimana: µMax = kecepatan pertumbuhan maksimal bakteri
At1 = Absorbansi pada waktu pertama
At2 = Absorbansi pada waktu kedua
t = waktu dalam jam
Pertumbuhan SGB Hn+ dengan waktu pembelahan sel maksimal pada 3 jam.
0 0.2 0.4 0.6 0.8
1 1.2 1.4
0 1 2 3 4 5 6 7 8 9
Ab
so
rba
nsi
Waktu (jam)
S. agalactiae isolat no. 2
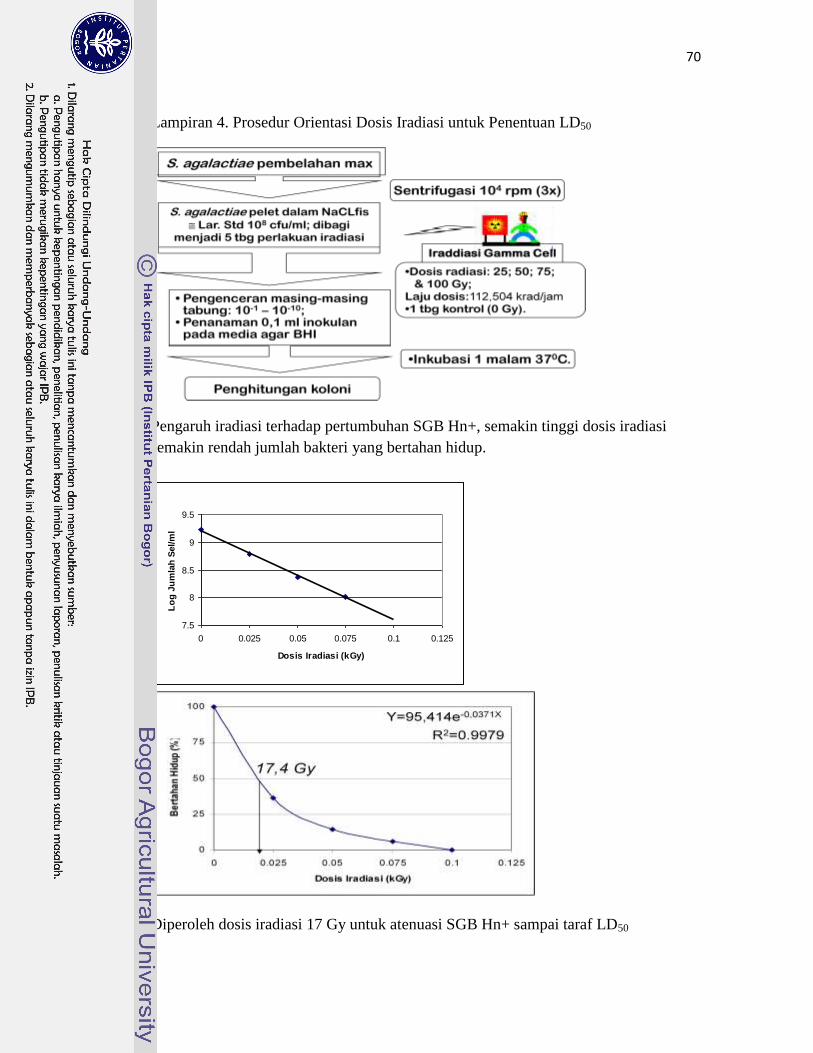
70
Lampiran 4. Prosedur Orientasi Dosis Iradiasi untuk Penentuan LD50
Pengaruh iradiasi terhadap pertumbuhan SGB Hn+, semakin tinggi dosis iradiasi
semakin rendah jumlah bakteri yang bertahan hidup.
Diperoleh dosis iradiasi 17 Gy untuk atenuasi SGB Hn+ sampai taraf LD50
7.5
8
8.5
9
9.5
0 0.025 0.05 0.075 0.1 0.125
Dosis Iradiasi (kGy)
Lo
g J
um
lah
Sel/m
l

71
Lampiran 5. Iradiator Gamma Chamber dengan Sumber Isotop Cobalt-60 ( 60
Co).
Iradiasi SGB Hn+ dilakukan dalam iradiator Gamma Chamber dengan sumber isotop 60
Co menggunakan fasilitas Balai Iradiasi Pusat Aplikasi Teknologi Isotop dan
Radiasi (PATIR), Badan Tenaga Nuklir Nasional (BATAN) Jakarta.
Perhitungan waktu paparan iradiasi untuk masing-masing dosis serap iradiasi :
Waktu = Dosis / Laju dosis
Dosis 50 Gy = 0,050 KGy = 5 Krad
Laju dosis = 112,504 Krad/jam
Waktu = 5 Krad = 0,044443 jam = 2 menit 40 detik
112,504 Krad/jam
untuk selanjutnya seperti tersaji dalam tabel berikut,
Dosis serap iradiasi Waktu paparan iradiasi
Gy menit detik
0 0 0
25 1 20
50 2 40
75 4 0
100 5 20

72
Lampiran 6. Jadwal Perlakuan dan Parameter Penelitian pada Hewan Model Mencit
Persiapan Hewan Percobaan Mencit:
Mencit yang digunakan dalam penelitian adalah mencit betina dalam keadaan
bunting. Sebelum dikawinkan, seluruh mencit diberi obat cacing (albendazole50%)
dengan dosis 10mg/kg BB dan diulang seminggu kemudian, lalu diberi antibiotik
(clavamox) dengan dosis 25mg/kg BB selama lima hari berturut-turut setelah
pemberian obat cacing yang pertama dengan jeda waktu 2 hari. Diberikan pula anti
jamur (flagyl) dengan dosis 30mg/kgBB setelah jeda dua hari dari pemberian obat
cacing yang kedua.
2 hari 5 hari 2 hari 2 hari 2 hari
albendazole clavamox albendazole flagyl kawin
Perlakuan Hewan Percobaan Mencit:
Pengamatan parameter:
1.Histopat kelenjar mamae dengan pewarnaan Haematoksilin Eosin (HE)
Alveol kelenjar produktif
Sel epitel kelenjar deskuamasi
Sel radang dalam lumen alveol kelenjar
Sel radang interstisial
2.Antibodi Ig-G dengan ELISA secara tidak lansung.

73
Lampiran 7. Pembuatan Preparat Histopatologi Kelenjar Mamae Mencit
1. Kelenjar mamae yang telah difiksasi dalam BNF 10% kurang lebih dua hari
kemudian dipotong tipis. Potongan kelenjar mamae dengan ketebalan kurang
lebih 3mm dimasukkan ke dalam tissue basket dan di dehidrasi dengan
merendam sediaan secara berturut-turut pada alkohol 70%, 80%, 90%,
alkohol absolut I, alkohol absolut II, lalu dijernihkan dalam xilol I, xilol II,
dan diinfiltrasi pada parafin parafin I, dan parafin II. Masing-masing proses
perendaman dilakukan selama 2 jam dalam mesin automatic tissue processor,
Sakura (Japan).
2. Jaringan dimasukkan ke dalam pencetak berisi parafin cair dengan bentuk
blok dan disusun di tengah parafin. Setelah mulai membeku, parafin ditambah
kembali hingga alat pencetak penuh dan dibiarkan hingga parafin mengeras.
Selanjutnya, parafin yang berisi jaringan, dipotong menggunakan mikrotom
dengan ketebalan 5 mikron. Hasil pemotongan yang berbentuk pita diletakkan
diatas permukaan air hangat bersuhu 450celcius dengan tujuan menghilangkan
lipatan-lipatan. Sediaan diangkat dari permukaan air dengan gelas obyek yang
telah diolesi larutan albumin yang berguna untuk merekatkan sediaan. Setelah
itu, preparat dikeringkan dalam inkubator bersuhu 600celcius semalam.
3. Sediaan yang telah kering dimasukkan ke dalam xilol dua kali selama 2 menit.
Kemudian sediaan direhidrasi yang dimulai dari alkohol absolut sampai
alkohol 80% dengan waktu masing-masing 2 menit. Selanjutnya sediaan
dicuci pada air mengalir dan dikeringkan.
4. Sediaan yang telah siap, diwarnai dengan pewarnaan Mayer‟s Hematoksilin
selama 8 menit, kemudian dibilas dengan air mengalir dan dicuci dengan
lithium karbonat selama 15-30 detik, dibilas dengan air lagi, dan akhirnya
diwarnai dengan pewarna eosin selama 2 menit. Untuk menghilangkan warna
eosin yang berlebihan, sediaan dicuci dengan air mengalir dan dikeringkan.
Kemudian sediaan dicelup ke dalam alkohol 90% sebanyak 10 kali celupan,

74
alkohol absolut I 10 kali celupan, alkohol absolut II selama 2 menit, xilol I
selama 1 menit, dan xylol II selama 2 menit.
5. Sediaan dikeringkan dahulu, lalu ditetesi dengan perekat permount dan
kemudian ditutup dengan cover glass dan dibiarkan selama beberapa menit
hingga melekat sempurna. Setelah itu, preparat siap untuk diamati dan diambil
gambar menggunakan camera mikroskop. Identifikasi dan pengambilan
gambar preparat dilakukan di Bagian Patologi, Departemen Klinik,
Reproduksi dan Patologi, Fakultas Kedokteran Hewan, Institut Pertanian
Bogor.

75
Lampiran 8. Ekstraksi Antigen SGB untuk coating Antigen pada ELISA
Metode ekstraksi untuk mempersiapkan antigen untuk coating Ag pada
ELISA mengikuti metode yang dikembangkan oleh Wibawan (1993). Ekstraksi Ag
dilakukan dengan menggunakan Asam Klorida (HCl).
1. Sebanyak 40 ml biakan SGB Hn+ yang telah ditumbuhkan dalam media BHI
broth selama 1 malam disentrifus (10000 rpm selama 10 menit, pada suhu
40C);
2. Sedimen bakteri yang terbentuk kemudian dicuci sebanyak 2 kali
menggunakan 10 ml 0,14 mol/l NaCl dengan sentrifugasi seperti metode
penyiapan Ag di atas.
3. Sedimen bakteri yang terbentuk diresuspensikan dalam 350 µl larutan 0,2
mol/l asam klorida (HCl) dan diinkubasikan dalam penangas air pada suhu
52oC, selama 2 jam.
4. Suspensi bakteri kemudian dinetralisasi oleh 1 mol/l NaOH dan diberi Phenol
Red sebagai indikator.
5. Antigen untuk coating plat ELISA dikoleksi dengan cara sentrifugasi suspensi
bakteri tersebut dengan kecepatan 13000 rpm, selama 5 menit pada suhu 4oC.
Supernatan yang terbentuk dipergunakan sebagai Antigen untuk melapisi
sumuran pada plat ELISA dengan metode tidak langsung (Indirect ELISA)..

76
Lampiran 9. ELISA untuk Titer Antibodi Ig-G
Prosedur ELISA (indirect) untuk pengukuran titer antibodi Ig-G sebagai berikut:
1. Coating plat ELISA dg Ag SGB dalam buffer carbonat bicarbonat pH 9,6
masing-masing 50 µl tiap sumur, bungkus alumunium foil, lalu simpan satu
malam dalam refreegerator 4oC
2. Cuci plat dengan PBS Tween20 (0,05%) 4 kali lalu bilas dengan aquabidest
3. Blocking dg Fetal Bovine Serum (FBS) 2% dalam PBS masing-masing 50 µl
tiap sumur , lalu inkubasi 1jam dlm inkubator 37oC
4. Cuci plat seperti langkah 2
5. Serum sampel yg sdh diinaktifasi pada penangas air 54oC selama 30 menit
dan diencerkan dgn PBS 1:30 , diisikan kedlm sumuran plat sesuai pola
nomor sampel, lalu diinkubasi 1 jam dlm inkubator 37oC
6. Cuci plat seperti langkah 2
7. Penambahan conjugate Peroxidase-Anti mouse Ig-G (1:10.000) masing-
masing 50 µl tiap sumur, lalu diinkubasi 1jam dlm inkubator 37oC
8. Cuci plat seperti langkah 2
9. Masukkan substrat TMB (dalam ruang gelap) masing-masing 50 µl tiap sumur
lalu diinkubasi dalam inkubator 37oC selama 30 menit
10. Pembacaan titer ELISA dalam microplate reader Benchmark Biorad©
dengan panjang gelombang (λ) = 655 nm.