implikasi eksistensi chromolaena odorata (l.) … fasilitas yang tersedia di laboratorium. kepada...
TRANSCRIPT

IMPLIKASI EKSISTENSI Chromolaena odorata (L.) King & Robinson (ASTERACEAE) DAN AGENS HAYATINYA
Cecidochares connexa Macquart (DIPTERA: TEPHRITIDAE) TERHADAP STRUKTUR KOMUNITAS SERANGGA
DAN TUMBUHAN LOKAL
ALAL HUDA JAYA. S
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2006

SURAT PERNYATAAN
Dengan ini saya menyatakan bahwa tesis yang berjudul ”Implikasi Eksistensi Chromolaena odorata (L.) King & Robinson (Asteraceae) dan Agens Hayatinya Cecidochares connexa Macquart (Diptera: Tephritidae) terhadap Struktur Komunitas Serangga dan Tumbuhan lokal” adalah karya saya sendiri dan belum diajukan dalam bentuk apapun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini
Bogor, Desember 2006
Alal Huda Jaya. S NRP. A451030041

ABSTRAK
ALAL HUDA JAYA. S. Implikasi Eksistensi Chromolaena odorata (L.) King & Robinson (Asteraceae) dan Agens Hayatinya Cecidochares connexa Macquart (Diptera: Tephritidae) terhadap Struktur Komunitas Serangga dan Tumbuhan Lokal. Dibimbing oleh DAMAYANTI BUCHORI dan SOEKISMAN TJITROSEMITO.
Penelitian ini bertujuan untuk mempelajari distribusi dan kelimpahan populasi C. odorata dan agens hayatinya C. connexa, serta mempelajari struktur komunitas serangga dan tumbuhan lokal di daerah yang telah diinvasi oleh C. odorata. Kegiatan yang dilakukan adalah survei lokasi penelitian, analisis vegetasi, dan koleksi serangga. Penelitian ini berlokasi di Jawa Barat yang meliputi daerah Parung Panjang, Setu, Darmaga, Gunung Bunder, Gunung Salak, Gunung Halimun, Pakuwon, Ciawi, Cisarua, dan Cianjur. Lokasi penelitian tersebut ditentukan berdasarkan metode purposive sampling. Analisis vegetasi dilakukan dengan metode transek yang dikombinasikan dengan metode kuadrat. Koleksi serangga dilakukan di dalam petak contoh dengan memasang perangkap pitfall trap dan yellow pan trap, sedangkan perangkap malaise trap diletakkan secara diagonal diantara jalur transek.
Hasil penelitian memperlihatkan bahwa spesies tumbuhan eksotik invasif C. odorata terdapat disemua lokasi penelitian dan dapat tumbuh pada berbagai tipe habitat baik habitat perkebunan, hutan buatan, ladang dataran rendah, maupun ladang dataran tinggi dengan kelimpahan yang berbeda. Kelimpahan populasinya cenderung menurun seiring dengan bertambahnya ketinggian suatu lokasi. Keberadaan C. odorata diikuti pula dengan keberadaan musuh alaminya yaitu lalat puru C. connexa pada semua lokasi pengamatan, kecuali di Gunung Halimun dan Cianjur. Saat ini lalat puru C. connexa telah mapan dan menyebar secara alami. Adanya barier seperti pegunungan merupakan faktor pembatas terhadap distribusi lalat puru C. connexa.
Komunitas tumbuhan yang diperoleh pada habitat C. odorata di Parung Panjang, Setu, Darmaga, dan Gunung Bunder secara keseluruhan terdiri dari 131.132 individu tumbuhan yang termasuk ke dalam 21 famili dan 44 spesies. Sedangkan jumlah total komunitas serangga yang diperoleh pada habitat C. odorata di Parung Panjang, Setu, Darmaga, dan Gunung Bunder terdiri dari 24.213 individu serangga yang termasuk ke dalam 14 ordo, 132 famili, dan 568 spesies. Ordo Hymenoptera, Diptera, dan Hemiptera merupakan tiga ordo terbesar yang ditemukan dengan kelimpahan individu (species abundance) dan kekayaan spesies (species richness) paling tinggi.
Implikasi keberadaan C. odorata telah menyebabkan terjadinya pengambilalihan atau pergantian skala ruang oleh C. odorata terhadap spesies tumbuhan lokal serta penurunan keanekaragaman spesies tumbuhan yang berada disekitarnya. Introduksi tumbuhan eksotik invasif C. odorata dan agens hayatinya C. connexa telah menyebabkan terjadinya asosiasi dengan serangga-serangga lokal.

ABSTRACT
ALAL HUDA JAYA. S. Implication of existence both of Chromolaena odorata (L.) King & Robinson (Asteraceae) and its biological control agent - Cecidochares connexa Macquart (Diptera: Tephritidae) to local plant and insect community structure. Supervisors: DAMAYANTI BUCHORI and SOEKISMAN TJITROSEMITO.
The aims of this research were (1) to study density and population distribution of C. odorata and its biological control agent - C. connexa, and (2) to study local plant and insect community structure in invaded area of C. odorata. Several activities have been conducted i.e. survey of research sites, vegetation analysis, and insect sampling. The study sites were selected base on purposive sampling method. All area located in Bogor district (Parung Panjang, Setu, Darmaga, Gunung Bunder, Ciawi, and Cisarua), Sukabumi district (Gunung Salak, Gunung Halimun, and Pakuwon), and Cianjur district. Analysis of vegetation were conducted using combination of transect and quadrate method. Insect were sampled by setting up several traps inside the plots. Pitfall trap and yellow pan trap were set up inside the plot, whereas malaise trap were placed between two transect lines.
The result found that invasive exotic plant species (C. odorata) spread on whole study sites and well-growth on different habitat types (plantations, production forest, low land, and high land). Their population densities tend to decrease with increasing altitude. The occurrences of C. odorata co-exist with its biological control, the gall flies C. connexa. Almost all study sites (except Gunung Halimun and Cianjur) found these flies. Recently, C. connexa have established and wide spread, although the present of barrier (like mountain) is restriction factor for distribution of flies in West Java.
Over 131.132 individual of plants consist of 21 families and 44 species were identified co-exist with C. odorata in Bogor area (Parung Panjang, Setu, Darmaga, and Gunung Bunder). In the same area, over 24.213 individual of insect consist of 14 orders, 132 families, and 568 species were found in C. odorata habitat. Three dominance insect orders i.e. Hymenoptera, Diptera, and Hemiptera were collected with high species richness and abundance.
Implication of existence of C. odorata caused replacement local plant species and homogenization habitat on spatial scale. Introduction of C. odorata and its biological control agent caused and consequently associate with local insects.

© Hak cipta milik Institut Pertanian Bogor, tahun 2006 Hak cipta dilindungi
Dilarang mengutip dan memperbanyak tanpa izin tertulis dari Institut Pertanian Bogor, sebagian atau seluruhnya dalam bentuk apa pun,
baik cetak, fotokopi, mikrofilm, dan sebagainya

IMPLIKASI EKSISTENSI Chromolaena odorata (L.) King & Robinson (ASTERACEAE) DAN AGENS HAYATINYA
Cecidochares connexa Macquart (DIPTERA: TEPHRITIDAE) TERHADAP STRUKTUR KOMUNITAS SERANGGA
DAN TUMBUHAN LOKAL
ALAL HUDA JAYA. S
Tesis sebagai salah satu syarat untuk memperoleh gelar
Magister Sains pada Program Studi Entomologi/Fitopatologi
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2006

Judul Tesis : Implikasi Eksistensi Chromolaena odorata (L.) King &
Robinson (Asteraceae) dan Agens Hayatinya Cecidochares connexa Macquart (Diptera: Tephritidae) terhadap Struktur Komunitas Serangga dan Tumbuhan Lokal
Nama : Alal Huda Jaya. S NRP : A451030041 Program Studi : Entomologi/Fitopatologi
Disetujui
Komisi Pembimbing
Dr. Ir. Damayanti Buchori, M.Sc. Ketua
Dr. Soekisman Tjitrosemito, M.Sc. Anggota
Diketahui
Ketua Program Studi Entomologi/Fitopatologi
Dr. Ir. Sri Hendrastuti Hidayat, M.Sc.
Dekan Sekolah Pascasarjana
Prof. Dr. Ir. Khairil Anwar Notodiputro, MS
Tanggal Ujian: 28 September 2006 Tanggal Lulus: 18 Desember 2006

PRAKATA
Syukur Alhamdulillah penulis panjatkan kehadirat Allah SWT karena berkat rahmat dan karunia-Nya penelitian dan penulisan tesis ini berhasil diselesaikan. Tema yang dipilih dalam penelitian ini adalah spesies eksotik invasif, dengan judul Implikasi Eksistensi Chromolaena odorata (L.) King & Robinson (Asteraceae) dan Agens Hayatinya Cecidochares connexa Macquart (Diptera: Tephritidae) terhadap Struktur Komunitas Serangga dan Tumbuhan Lokal.
Penghargaan dan ucapan terima kasih penulis sampaikan kepada Dr. Ir. Damayanti Buchori, M.Sc. dan Dr. Soekisman Tjitrosemito, M.Sc. selaku komisi pembimbing, atas bimbingan, saran, dan motivasi yang telah diberikan selama persiapan penelitian sampai penulisan tesis. Kepada Akhmad Rizali, SP, M.Si terima kasih yang sebesar-besarnya atas bimbingan teknis ilmiahnya. Ucapan terima kasih juga disampaikan kepada Dirjen Pendidikan Tinggi Departemen Nasional RI melalui Hibah Tim Pascasarjana atas bantuan dana yang diberikan sehingga penelitian ini dapat terlaksana, kepada kepala Laboratorium Bioekologi Parasitoid dan Predator, Departemen Proteksi Tanaman, Fakultas Pertanian IPB yang telah memberikan izin untuk menggunakan fasilitas yang tersedia di laboratorium. Kepada kedua orang tua tercinta Ayahanda Banding Ratu (alm) dan Ibu Sundarti, semoga Allah SWT memberikan balasan amal baik mereka dengan pahala yang tak terhingga. Kepada kakakku Eteh Marina dan adik-adikku tercinta, Syukrida, Desrina, dan seluruh saudara serta Keluarga Besar di Lampung, terima kasih atas perhatian dan dukungannya selama ini. Terima kasih kepada semua pihak dan rekan-rekan sekalian, anggota tim Hibah Pascasarjana, anggota Laboratorium Bioekologi Parasitoid dan Predator, Departemen Proteksi Tanaman, Fakultas Pertanian IPB, serta seseorang yang akan mendampingiku kelak dikemudian hari, semoga Allah SWT membalasnya.
Akhir kata tak ada gading yang tak retak, semoga tulisan ini bermanfaat, Amin!!!.
Bogor, Desember 2006
Alal Huda Jaya. S

RIWAYAT HIDUP
Penulis dilahirkan di Menggala pada tanggal 7 Juni 1979 sebagai anak kedua dari empat bersaudara, dari Ayah Banding Ratu (alm) dan Ibu Sundarti.
Tahun 1997 penulis lulus dari Madrasah Aliyah Negeri (MAN) I Bandar Lampung dan pada tahun yang sama penulis diterima di Jurusan Proteksi Tanaman Program Studi Ilmu Hama dan Penyakit Tumbuhan, Fakultas Pertanian, Universitas Lampung melalui jalur Ujian Masuk Perguruan Tinggi Negeri (UMPTN), dan memperoleh gelar Sarjana Pertanian pada tahun 2002. Kesempatan untuk melanjutkan ke Program Magister Sains Entomologi /Fitopatologi Sekolah Pascasarjana Institut Pertanian Bogor diperoleh penulis pada tahun 2003.

DAFTAR ISI
Halaman
DAFTAR TABEL .......................................................................................... xi
DAFTAR GAMBAR ..................................................................................... xii
DAFTAR LAMPIRAN ................................................................................ xiii
PENDAHULUAN
Latar Belakang .................................................................................. 1
Tujuan Penelitian ................................................................................ 3
Manfaat Penelitian .............................................................................. 3
DISTRIBUSI, KELIMPAHAN POPULASI Chromolaena odorata (L.) King & Robinson (ASTERACEAE) DAN AGENS HAYATINYA Cecidochares connexa Macquart (DIPTERA: TEPHRITIDAE) DI BOGOR, JAWA BARAT
Pendahuluan ........................................................................................ 4
Bahan dan Metode .............................................................................. 6
Lokasi dan Waktu Penelitian ...................................................... 7
Metode Pengukuran Kelimpahan Populasi ....................... 7
Metode Survei Keberadaan Lalat Puru C. connexa .......... 8
Pengambilan Contoh Tumbuhan ................................................ 8
Estimasi Populasi C. odorata dan Lalat Puru C. connexa ......... 10
Analisis Data .............................................................................. 10
Hasil dan Pembahasan ........................................................................ 11
Distribusi dan Kelimpahan Populasi C. odorata ........................ 11
Distribusi dan Kelimpahan Populasi Lalat Puru C. connexa ..... 14
Kekayaan Spesies Tumbuhan di Habitat C. odorata ................. 18
Implikasi Keberadaan Tumbuhan Eksotik Invasif C. odorata terhadap Keberadaan Tumbuhan Lokal ..................................... 21
Kesimpulan ......................................................................................... 24

STRUKTUR KOMUNITAS SERANGGA PADA HABITAT TUMBUHAN EKSOTIK INVASIF: STUDI KASUS PADA HABITAT Chromolaena odorata (L.) KING & ROBINSON (ASTERACEAE) DI BOGOR, JAWA BARAT
Pendahuluan ........................................................................................ 26
Bahan dan Metode .............................................................................. 27
Lokasi dan Waktu Penelitian ...................................................... 27
Pengambilan Contoh Serangga .................................................. 27
Analisis Data .............................................................................. 30
Hasil dan Pembahasan ........................................................................ 30
Kekayaan Spesies Serangga di Habitat C. odorata .................... 30
Kekayaan Spesies Hymenoptera dan Diptera di Habitat C. odorata ................................................................................... 34
Hubungan Keanekaragaman Serangga dengan Kondisi Habitat ......................................................................................... 37
Asosiasi Serangga Lokal dengan Tumbuhan Eksotik Invasif C. odorata dan Agens Hayatinya C. connexa ............................ 41
Kesimpulan ................................................................................. 43
PEMBAHASAN UMUM ............................................................................. 44
KESIMPULAN DAN SARAN
Kesimpulan ......................................................................................... 47
Saran ................................................................................................... 48
DAFTAR PUSTAKA ................................................................................... 49
LAMPIRAN .................................................................................................. 52

DAFTAR TABEL
Halaman
1 Deskripsi lokasi yang dipilih berdasarkan metode pengukuran populasi C. odorata dan lalat puru C. connexa ....................................................... 7
2 Deskripsi lokasi yang dipilih berdasarkan metode survei keberadaan lalat
puru C. connexa ........................................................................................ 8 3 Jumlah Famili (F), Spesies (S), Individu (N), Indeks Shannon (H’) dan
kemerataan (E) tumbuhan pada tiap habitat C. odorata ........................... 20 4 Indeks kemiripan Sorensen (Cs) seluruh spesies tumbuhan antar lokasi
penelitian pada habitat C. odorata ............................................................ 20 5 Indeks Nilai Penting (INP) spesies tumbuhan tegak pada empat habitat
C. odorata ................................................................................................. 22 6 Indeks Nilai Penting (INP) spesies tumbuhan merambat pada empat
habitat C. odorata ...................................................................................... 23 7 Deskripsi lokasi yang dipilih untuk penelitian .......................................... 28 8 Jumlah ordo (O), famili (F), spesies (S), Individu (N), Indeks Shannon
(H’) dan kemerataan (E) serangga pada tiap habitat C. odorata ............... 31 9 Jumlah famili, spesies (S), dan individu (N) ordo Hymenoptera pada tiap
habitat C. odorata ...................................................................................... 34 10 Jumlah famili, spesies (S), dan individu (N) ordo Diptera pada tiap
habitat C. odorata ..................................................................................... 36 11 Indeks kemiripan Sorensen (Cs) seluruh spesies serangga antar lokasi
penelitian pada habitat C. odorata ............................................................ 38

DAFTAR GAMBAR
Halaman
1 Metode analisis vegetasi ............................................................................ 9 2 Kelimpahan populasi C. odorata pada empat lokasi penelitian selama tiga
kali pengambilan sampel ........................................................................... 13 3 Distribusi lalat puru C. connexa dari titik pelepasan Parung Panjang-
Jasinga, Bogor, dan Parung Kuda-Sukabumi, Jawa Barat ........................ 14 4 Kelimpahan populasi lalat puru C. connexa pada empat lokasi penelitian
selama tiga kali pengambilan sampel ........................................................ 17 5 Kurva akumulasi spesies tumbuhan pada empat habitat C. odorata ........ 19 6 Dendogram pengelompokan spesies tumbuhan pada empat habitat
C. odorata ................................................................................................. 21 7 Perangkap serangga; (a) yellow pan trap (b) pitfall trap (c) malaise
trap ............................................................................................................ 29 8 Kurva akumulasi spesies serangga pada empat habitat C. odorata .......... 31 9 Kekayaan spesies (A) dan kelimpahan individu (B) serangga yang
diperoleh pada empat habitat C. odorata .................................................. 32 10 Kekayaan spesies (A) dan kelimpahan individu (B) serangga tanpa
Formicidae yang diperoleh pada empat habitat C. odorata ...................... 33 11 Dendogram pengelompokan seluruh spesies serangga pada empat
habitat C. odorata ..................................................................................... 39 12 Dendogram pengelompokan serangga predator dan parasitoid pada
empat habitat C. odorata ........................................................................... 40 13 Dendogram pengelompokan serangga herbivor pada empat habitat
C. odorata ................................................................................................. 40 14 Asosiasi serangga pada tumbuhan eksotik invasif C. odorata .................. 41 15 Serangga predator yang berasosiasi dengan lalat puru C. connexa .......... 42

DAFTAR LAMPIRAN
Halaman
1 Hasil analisis vegetasi kelompok spesies tumbuhan tegak pada habitat C. odorata di Parung Panjang ................................................................... 53
2 Hasil analisis vegetasi kelompok spesies tumbuhan tegak pada habitat
C. odorata di Setu ...................................................................................... 53 3 Hasil analisis vegetasi kelompok spesies tumbuhan tegak pada habitat
C. odorata di Darmaga .............................................................................. 54 4 Hasil analisis vegetasi kelompok spesies tumbuhan tegak pada habitat
C. odorata di Gunung Bunder ................................................................... 54 5 Hasil analisis vegetasi kelompok spesies tumbuhan merambat pada
habitat C. odorata di Parung Panjang ....................................................... 55 6 Hasil analisis vegetasi kelompok spesies tumbuhan merambat pada
habitat C. odorata di Setu ......................................................................... 55 7 Hasil analisis vegetasi kelompok spesies tumbuhan merambat pada
habitat C. odorata di Darmaga .................................................................. 55 8 Hasil analisis vegetasi kelompok spesies tumbuhan merambat pada
habitat C. odorata di Gunung Bunder ....................................................... 56 9 Jumlah spesies (S), Individu (N) dan peranan serangga yang diperoleh
pada empat habitat C. odorata .................................................................. 57

PENDAHULUAN
Latar Belakang
Banyak biota yang secara langsung maupun tidak langsung terbawa oleh
manusia ke bagian dunia yang lain. Hal ini dimungkinkan dengan dimulainya era
eksplorasi yang dapat menghilangkan penghalang biogeografi yang sebelumnya
mengisolasi biota benua selama jutaan tahun (Mooney & Cleland 2001). Setelah
keluar dari habitat alaminya, biota asing tersebut segera menjadi penginvasi
agresif yang lebih kompetitif daripada biota lokal dan mengakibatkan pengaruh
merusak dalam ekosistem serta menimbulkan kerugian yang nyata bagi produksi
pertanian (Schoonhoven et al. 1996). Sifat invasif biota eksotik pada habitat
baru disebabkan karena tidak ada musuh alami (natural enemy) yang mampu
mengendalikan seperti di habitat aslinya.
Spesies eksotik invasif pada habitat baru dapat menyebabkan terjadinya
homogenisasi biotik yaitu pengantian spesies lokal oleh spesies pendatang (Olden
et al. 2004). Sebagai contoh, Bromus tectorum merupakan tanaman yang
diintroduksi ke Amerika Utara sekitar tahun 1890. Dalam kurun waktu 20 tahun
B. tectorum telah mapan di daerah pegunungan bagian barat Amerika Utara. Pada
tahun 1930 B. tectorum telah berhasil menginvasi dan mendominasi lebih dari
200.000 km2 habitat pegunungan di bagian barat Amerika Utara karena
peningkatan kemampuan migrasi dan populasinya (Mooney & Cleland 2001).
Contoh lain adalah Eichhornia crassipes atau eceng gondok merupakan tumbuhan
eksotik invasif pada ekosistem perairan di Indonesia dan beberapa negara di Asia
Tenggara (Kasno et al. 2001). Dampak keberadaan gulma ini sangat nyata akibat
adanya akumulasi biomassa, penutupan permukaan danau dan sungai secara cepat,
serta dapat menyebabkan terjadinya pendangkalan danau dan sungai.
Keberadaan spesies eksotik invasif cenderung merugikan karena merupakan
ancaman yang serius terhadap ekosistem dan keanekaragaman hayati (Wittenberg
& Cock 2003). Spesies eksotik invasif mampu beradaptasi dan mendominasi
suatu habitat baru yang awalnya di dominasi oleh spesies lokal sehingga bisa
memicu terjadinya homogenisasi biotik dan perubahan struktur komunitas pada
habitat baru tersebut. Selain itu, kemampuan kompetisi spesies eksotik invasif

2
mampu mengalahkan spesies lokal sehingga dapat menyebabkan spesies lokal
kalah bersaing dan akhirnya bisa memicu terjadinya kepunahan (Olden et al.
2004; Untung 2005).
Chromolaena odorata (L.) King & Robinson (Asteraceae) atau kirinyuh
merupakan spesies tumbuhan eksotik invasif dan merupakan gulma penting pada
habitat perkebunan di Indonesia (Anonim 1991). Tumbuhan ini berasal dari
Amerika yang diintroduksi ke kebun raya di Dakka (India), Jawa, dan Peradeniya
(Srilanka) pada abad ke-19 (Binggeli 1997). Di Indonesia tumbuhan ini pertama
kali dilaporkan pada tahun 1934 di Lubuk Pakam, Sumatera Utara (Tjitrosemito
1999). Saat ini persebaran C. odorata sudah sangat luas, meliputi Sumatera, Jawa,
Kalimantan, Nusa Tenggara Timur, dan Irian Jaya (Sipayung & Chenon 1995).
Tjitrosemito (1998) melaporkan bahwa persebaran tumbuhan ini sangat cepat
karena kemampun dan efisiensinya, bahkan di laboratorium pertumbuhan C.
odorata sangat cepat pada 8 minggu pertama.
Pengendalian C. odorata dengan memanfaatkan agens hayatinya telah
dilakukan di Indonesia. Upaya ini diawali dengan mengintroduksi serangga
herbivor Pareuchaetes pseudoinsulata Rego Barros (Lepidoptera: Arctiidae) dari
Guam, Amerika Serikat dan pertama kali di lepaskan pada tahun 1992. Namun
P. pseudoinsulata hanya berhasil menekan populasi tumbuhan tersebut di
Sumatera Utara, sedangkan di Jawa dilaporkan tidak berhasil. Selanjutnya pada
tahun 1993 kembali diintroduksi lalat puru Cecidochares connexa Macquart
(Diptera: Tephritidae) dari Columbia dan mulai dilepas pada tahun 1995 setelah
memperoleh izin pelepasan yang dikeluarkan oleh Menteri Pertanian
(Tjitrosemito 1998; Chenon et al, 2002). Sampai saat ini lalat puru C. connexa
keberadaannya telah mapan dan menyebar secara alami khususnya di Jawa Barat.
Sayangnya sampai saat ini, kajian terhadap lalat puru C. connexa sebagai
agens hayati C. odorata masih terbatas pada keberhasilan pelaksanaan introduksi,
perbanyakan massal, dan pelepasannya di lapangan tanpa diikuti kajian secara
komprehensif. Belum pernah dilakukan penelitian untuk memonitor distribusi,
kelimpahan populasi, dan bagaimana dampak keberadaan spesies eksotik invasif
C. odorata dan agens hayati C. connexa terhadap struktur komunitas serangga dan

3
tumbuhan lokal. Tanpa adanya usaha monitoring, dikhawatirkan terjadi penurunan
tingkat keanekaragaman hayati pada ekosistem tersebut tidak dapat dipantau,
terutama tempat atau habitat yang jauh dari titik pelepasan (Simberloff 1996).
Bertitik tolak dari uraian di atas, kajian terhadap spesies eksotik invasif C.
odorata dan agens hayatinya C. connexa dianggap penting sehingga nantinya
dapat digunakan untuk menganalisis dampak yang terjadi akibat introduksi
spesies eksotik invasif tersebut dan agens hayatinya.
Tujuan Penelitian
Penelitian ini bertujuan untuk (1) mempelajari distribusi dan kelimpahan
populasi C. odorata dan agens hayatinya lalat puru C. connexa, (2) mempelajari
struktur komunitas serangga dan tumbuhan di daerah yang telah diinvasi oleh C.
odorata dengan mengambil studi kasus di daerah Bogor, Jawa Barat.
Manfaat Penelitian
Hasil yang diperoleh melalui penelitian ini diharapkan dapat memberikan
informasi mengenai (1) distribusi dan kelimpahan populasi spesies tumbuhan
eksotik invasif C. odorata dan agens hayatinya lalat puru C. connexa, (2) struktur
komunitas serangga dan tumbuhan di daerah yang telah diinvasi oleh C. odorata,
(3) fenomena ekologi introduksi spesies eksotik dapat menyebabkan terjadinya
asosiasi antara spesies eksotik tersebut dengan serangga lokal dalam struktur
komunitas baru, sehingga menjadi rekomendasi bagi pemerintah bahwa perlunya
upaya perhatian dan penyaringan yang ketat terhadap spesies eksotik yang
didatangkan dari luar.

DISTRIBUSI, KELIMPAHAN POPULASI Chromolaena odorata (L.) King & Robinson (ASTERACEAE) DAN AGENS
HAYATINYA Cecidochares connexa Macquart (DIPTERA: TEPHRITIDAE) DI BOGOR, JAWA BARAT
PENDAHULUAN
Invasi spesies eksotik merupakan ancaman yang serius terhadap suatu
ekosistem alami dan keanekaragaman hayati. Penghalang alami seperti samudera,
pegunungan, dan padang pasir telah terlampaui sekitar abad ke dua puluh,
sehingga memberikan peluang suatu spesies untuk mengalami koevolusi dan
berkembang dalam suatu ekosistem baru. Keadaan ini disebabkan karena adanya
aktivitas manusia seperti perdagangan dan perjalanan, sehingga dapat
menyebabkan perpindahan suatu spesies baik disengaja maupun tidak disengaja
ke berbagai bagian dunia lainnya (Wittenberg & Cock 2003). Setelah keluar dari
habitat alaminya, spesies eksotik tersebut berkembang dan beradaptasi dengan
lingkungan sekitar sehingga menjadi penginvasi yang agresif. Selain itu, spesies
eksotik tersebut mampu berkompetisi dengan spesies lokal, menggeser
keberadaannya, menyebabkan kerusakan ekosistem alami, serta menimbulkan
kehilangan yang nyata bagi produksi pertanian (Schoonhoven et al. 1996).
C. odorata merupakan tumbuhan eksotik invasif yang berasal dari Amerika
Selatan dan Amerika Tengah yang telah menyebar ke Asia, Asia Tenggara, dan
beberapa daerah di Afrika (Anonim 1991). Tumbuhan ini sengaja diintroduksi ke
Kalkuta (India) sebagai tanaman hias pada tahun 1840-an (Ramachandra 1920
dalam Tjitrosemito 1997). Persebaran tumbuhan tersebut saat ini telah mencapai
Asia Tenggara termasuk Indonesia dan Australia yang telah berubah status
menjadi gulma yang tumbuh secara mapan dan sangat merugikan (Tjitrosemito
1997). Di indonesia persebaran C. odorata sudah sangat luas, meliputi Sumatera,
Jawa, Kalimantan, Nusa Tenggara Timur, dan Irian Jaya (Sipayung & Chenon
1995). Spesies ini dapat menyebar secara cepat karena kemampuan dan efisiensi
persebarannya (Binggeli 1997).

5
Berbagai upaya pengendalian telah dilakukan untuk mengatasi pesatnya
perkembangan populasi C. odorata di Indonesia. Salah satu teknik yang
dikembangkan adalah pengendalian hayati secara klasik dengan mengintroduksi
lalat puru Cecidochares connexa Macquart (Diptera: Tephritidae) dari Columbia
pada tahun 1993 dan mulai dilepas pada tahun 1995 setelah memperoleh izin
pelepasan yang dikeluarkan oleh Menteri Pertanian. Pelepasan lalat puru C.
connexa telah dilakukan di Jawa Barat yang meliputi daerah Hutan Tanaman
Industri Parung Panjang-Jasinga, Bogor pada tahun 1995, Pakuwon-Parung Kuda,
Sukabumi tahun 1996, dan padang pengembalaan Cagar Alam Pangandaran,
Ciamis pada tahun 1999. Selain itu, pada tahun 1996 pelepasan lalat puru C.
connexa juga dilakukan di Saradan, Madiun, Jawa Timur (Tjitrosemito 1998;
Tjitrosemito 2000; Widayanti et al. 2001). Sampai saat ini, lalat puru C. connexa
telah menyebar secara alami khususnya di daerah Bogor, Jawa Barat .
Lalat puru C. connexa merupakan lalat berwarna hitam yang berukuran
panjang 6.9 mm dan lebar 2 mm dengan rentang sayap 11.2 mm untuk imago
betina, sedangkan imago jantan berukuran panjang 5.6 mm dan lebar 1.8 mm
dengan rentang sayap 10 mm (Widayanti et al. 1999). Lalat puru C. connexa
meletakkan telur pada pucuk terminal maupun lateral daun C. odorata yang belum
membuka. Telur menetas 4-7 hari setelah oviposisi dan larva yang baru menetas
masuk ke dalam jaringan batang. Menjelang instar akhir, larva akan membuat
saluran keluar yang disebut dengan jendela puru. Stadia pupa membutuhkan
waktu 15-25 hari. Lama hidup lalat puru C. connexa dewasa berkisar antara 5-11
hari (McFadyen et al. 2003), sedangkan siklus hidupnya antara 47-73 hari dengan
rata-rata 60 hari (Chenon et al. 2002)
Penggunaan lalat puru C. connexa sebagai agens hayati untuk
mengendalikan C. odorata pada beberapa daerah khususnya di Bogor, Jawa Barat
kurang memberikan hasil yang memuaskan. Hasil survei di lapangan
memperlihatkan bahwa kelimpahan populasi C. odorata tetap tinggi meskipun
telah terinvestasi oleh lalat puru C. connexa. Tjitrosemito (1999b) melaporkan
bahwa pada musim kemarau populasi C. odorata sebagai inang bagi lalat puru C.
connexa menjadi kering dan sangat rendah sehingga telur yang diletakkan oleh

6
imago betina lalat puru C. connexa tidak dapat menyelesaikan siklus hidupnya.
Ketika musim hujan, C. odorata tumbuh kembali secara serentak yang berasal
dari populasi sebelumnya dimana pucuk C. odorata bertambah banyak dan
meningkat dengan pesat. Keadaan ini menyebabkan kelimpahan populasi lalat
puru C. connexa tidak mampu mengimbangi populasi inangnya yang tumbuh
dengan populasi yang sangat berlimpah sehingga mengakibatkan kurang
efektifnya lalat puru tersebut untuk menekan populasi C. odorata. Selain itu,
hasil pengamatan di lapangan menunjukkan bahwa batang C. odorata yang
terinvestasi oleh puru justru memiliki jumlah pucuk yang lebih banyak
dibandingkan yang tidak terinvestasi oleh puru.
Penelitian di Indonesia hanya terfokus pada keberhasilan pelaksanaan
introduksi lalat puru C. connexa dan pengembangan metode pembiakan massal
dalam upaya pemanfaatan serangga ini untuk mengendalikan C. odorata. Belum
pernah dilakukan evaluasi distribusi dan kelimpahan populasi lalat puru C.
connexa sebagai agens hayati untuk mengendalikan spesies eksotik invasif C.
odorata. Penelitian ini bertujuan untuk mengetahui distribusi dan kelimpahan
populasi C. odorata dan lalat puru C. connexa, serta mempelajari struktur
komunitas tumbuhan di daerah yang telah di invasi oleh C. odorata dengan
mengambil studi kasus di daerah Bogor, Jawa Barat.
BAHAN DAN METODE
Penelitian ini menggunakan dua metode yaitu (1) metode pengukuran
kelimpahan populasi C. odorata dan lalat puru C. connexa, (2) metode survei
keberadaan lalat puru C. connexa. Metode pertama difokuskan untuk mempelajari
secara spesifik kelimpahan populasi C. odorata dan lalat puru C. connexa
berdasarkan perbedaan habitat dan ketinggian tempat. Metode kedua hanya
difokuskan untuk melihat distribusi dan keberadaan lalat puru C. connexa pada
suatu lokasi berdasarkan gradien jarak dari titik pelepasan, ketinggian tempat,
serta adanya barier berupa pegunungan.
7
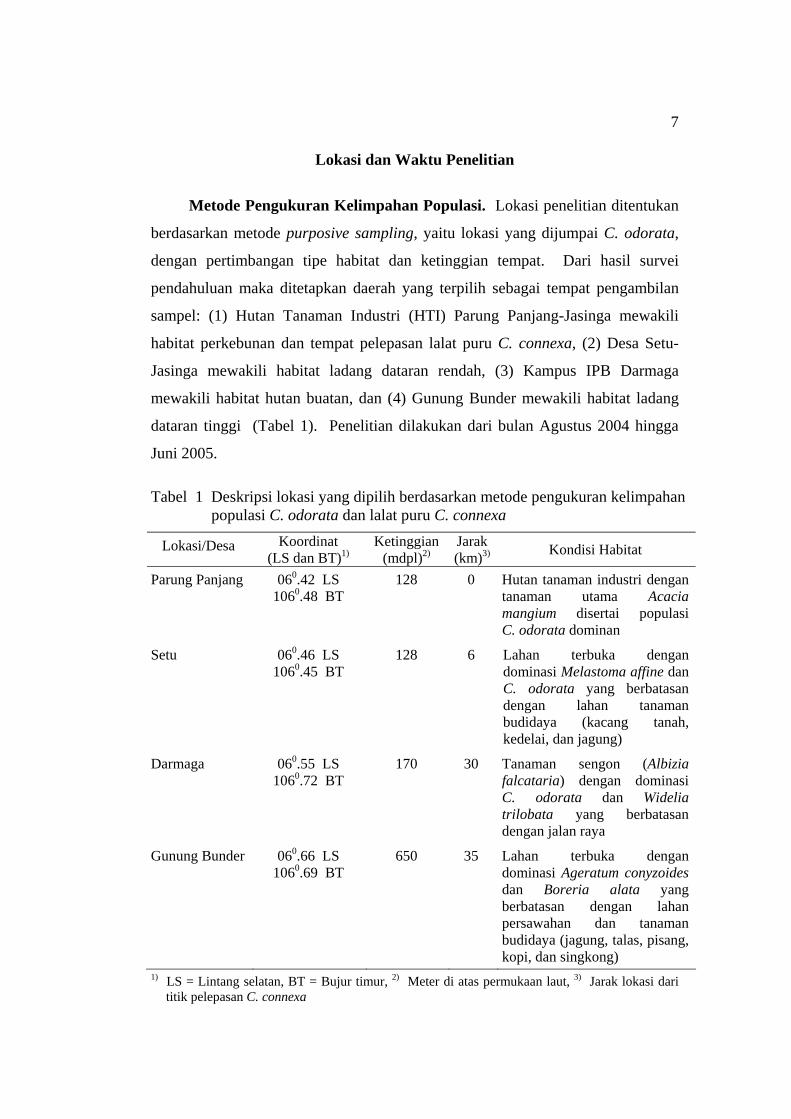
Lokasi dan Waktu Penelitian
Metode Pengukuran Kelimpahan Populasi. Lokasi penelitian ditentukan
berdasarkan metode purposive sampling, yaitu lokasi yang dijumpai C. odorata,
dengan pertimbangan tipe habitat dan ketinggian tempat. Dari hasil survei
pendahuluan maka ditetapkan daerah yang terpilih sebagai tempat pengambilan
sampel: (1) Hutan Tanaman Industri (HTI) Parung Panjang-Jasinga mewakili
habitat perkebunan dan tempat pelepasan lalat puru C. connexa, (2) Desa Setu-
Jasinga mewakili habitat ladang dataran rendah, (3) Kampus IPB Darmaga
mewakili habitat hutan buatan, dan (4) Gunung Bunder mewakili habitat ladang
dataran tinggi (Tabel 1). Penelitian dilakukan dari bulan Agustus 2004 hingga
Juni 2005.
Tabel 1 Deskripsi lokasi yang dipilih berdasarkan metode pengukuran kelimpahan populasi C. odorata dan lalat puru C. connexa
Lokasi/Desa Koordinat (LS dan BT)1)
Ketinggian (mdpl)2)
Jarak (km)3) Kondisi Habitat
Parung Panjang 060.42 LS 1060.48 BT
128 0 Hutan tanaman industri dengan tanaman utama Acacia mangium disertai populasi C. odorata dominan
Setu 060.46 LS 1060.45 BT
128 6 Lahan terbuka dengan dominasi Melastoma affine dan C. odorata yang berbatasan dengan lahan tanaman budidaya (kacang tanah, kedelai, dan jagung)
Darmaga 060.55 LS 1060.72 BT
170 30 Tanaman sengon (Albizia falcataria) dengan dominasi C. odorata dan Widelia trilobata yang berbatasan dengan jalan raya
Gunung Bunder 060.66 LS 1060.69 BT
650 35 Lahan terbuka dengan dominasi Ageratum conyzoides dan Boreria alata yang berbatasan dengan lahan persawahan dan tanaman budidaya (jagung, talas, pisang, kopi, dan singkong)
1) LS = Lintang selatan, BT = Bujur timur, 2) Meter di atas permukaan laut, 3) Jarak lokasi dari titik pelepasan C. connexa

8
Metode Survei Keberadaan Lalat Puru C. connexa. Lokasi penelitian
ditentukan berdasarkan metode purposive sampling, yaitu lokasi yang dijumpai C.
odorata, dengan pertimbangan gradien jarak dari tempat pelepasan lalat puru C.
connexa, ketinggian tempat, serta adanya barier berupa pegunungan. Dari hasil
survei pendahuluan maka ditetapkan daerah yang terpilih sebagai tempat
pengambilan sampel, yaitu Hutan Tanaman Industri (HTI) Parung Panjang-
Jasinga, Bogor dan Pakuwon-Parung Kuda, Sukabumi sebagai tempat pelepasan
lalat puru C. connexa, Desa Setu-Jasinga, Kampus IPB Darmaga, Gunung
Bunder, Gunung Salak, Gunung Halimun, Ciawi, Cisarua, dan Cianjur (Tabel 2).
Tabel 2 Deskripsi lokasi yang dipilih berdasarkan metode survei keberadaan lalat puru C. connexa
Lokasi/Desa Koordinat (LS dan BT)2)
Ketinggian (mdpl)3)
Jarak (km)4)
Jarak (km)5)
Parung Panjang1) 060.42 LS, 1060.48 BT 128 0 55
Pakuwon1) 060.84 LS, 1060.74 BT 300 55 0
Setu 060.46 LS, 1060.45 BT 128 6 52
Darmaga 060.55 LS, 1060.72 BT 170 31 32
Gunung Bunder 060.66 LS, 1060.69 BT 650 35 21
Gunung Salak 060.78 LS, 1060.61 BT 700 42 17
Gunung Halimun 060.89 LS, 1060.51 BT 650 52 27
Ciawi 060.64 LS, 1060.84 BT 443 48 25
Cisarua 060.69 LS, 1060.93 BT 962 60 28
Cianjur 1 060.80 LS, 1070.09 BT 714 80 39
Cianjur 2 060.80 LS, 1070.11 BT 608 82 42 1) Lokasi pelepasan lalat puru C. connexa, 2) LS = Lintang selatan, BT = Bujur timur, 3) Meter di
atas permukaan laut, 4) Jarak lokasi dari titik pelepasan di Parung Panjang, 5) Jarak lokasi dari titik pelepasan di Pakuwon
Pengambilan Contoh Tumbuhan
Pengambilan contoh tumbuhan dilakukan dengan cara analisis vegetasi
menggunakan metode transek yang dikombinasikan dengan metode kuadrat (Cox
2002). Pada masing-masing lokasi dibuat 5 jalur transek sepanjang 30 m dengan
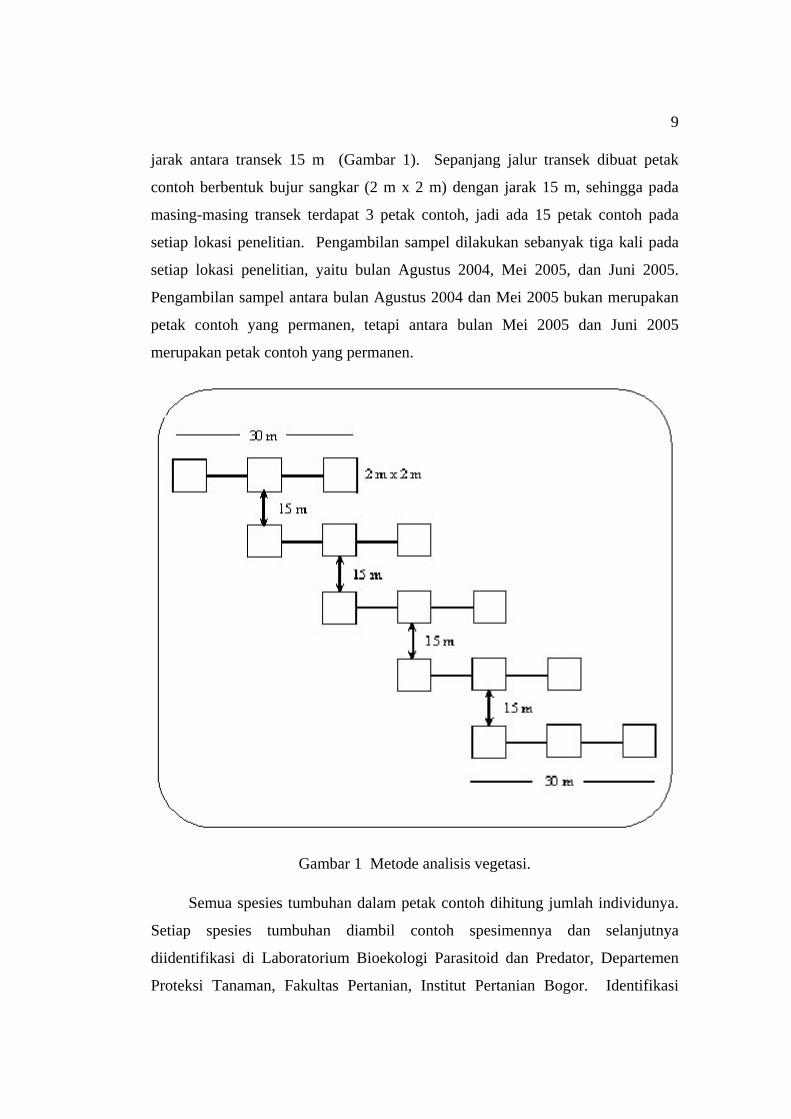
9
jarak antara transek 15 m (Gambar 1). Sepanjang jalur transek dibuat petak
contoh berbentuk bujur sangkar (2 m x 2 m) dengan jarak 15 m, sehingga pada
masing-masing transek terdapat 3 petak contoh, jadi ada 15 petak contoh pada
setiap lokasi penelitian. Pengambilan sampel dilakukan sebanyak tiga kali pada
setiap lokasi penelitian, yaitu bulan Agustus 2004, Mei 2005, dan Juni 2005.
Pengambilan sampel antara bulan Agustus 2004 dan Mei 2005 bukan merupakan
petak contoh yang permanen, tetapi antara bulan Mei 2005 dan Juni 2005
merupakan petak contoh yang permanen.
Gambar 1 Metode analisis vegetasi.
Semua spesies tumbuhan dalam petak contoh dihitung jumlah individunya.
Setiap spesies tumbuhan diambil contoh spesimennya dan selanjutnya
diidentifikasi di Laboratorium Bioekologi Parasitoid dan Predator, Departemen
Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor. Identifikasi

10
dilakukan dengan mengacu buku Kostermans et al. (1987) dan Laumonier et al.
(1987). Spesies tumbuhan yang tidak diketahui nama ilmiahnya diidentifikasi di
Herbarium SEAMEO-BIOTROP Bogor.
Estimasi Populasi C. odorata dan Lalat Puru C. connexa
Untuk mengestimasi populasi C. odorata, di dalam setiap petak contoh
jumlah pucuk C. odorata dihitung sehingga setiap batang C. odorata bisa terdiri
dari beberapa pucuk. Jumlah pucuk lebih akurat untuk mengestimasi populasi C.
odorata dibandingkan jumlah batang karena serangan lalat puru C. connexa
terdapat di pucuk tanaman yang merupakan tempat terbentuknya bunga dan buah.
Populasi lalat puru C. connexa diestimasi dengan menghitung kelimpahan jumlah
puru di dalam setiap petak contoh, sedangkan populasi C. odorata ditentukan
dengan menghitung jumlah pucuk pada setiap batang C. odorata dalam setiap
petak contoh tersebut.
Analisis Data
Data keseluruhan spesies tumbuhan yang diperoleh pada setiap lokasi dapat
diduga dengan menggunakan kurva akumulasi spesies yang dibuat dengan
program EstimateS 6.0b1 (Colwell 2000). Jumlah spesies tumbuhan yang
diperoleh pada setiap petak contoh diacak sebanyak 50 kali menggunakan
program tersebut. Prediksi kekayaan spesies tumbuhan diduga dengan
abundance-based coverage estimator (ACE) (Colwell & Coddington 1994).
Indeks keanekaragaman tumbuhan diukur berdasarkan Shannon-Wiener
(H’) = -Σ pi ln pi dimana pi = proporsi spesies ke-i terhadap total jumlah spesies,
Indeks kemerataan berdasarkan Shannon-Wiener (E) = H’/ln (S) dimana S = total
jumlah spesies yang diperoleh. Kemiripan komunitas tumbuhan antar lokasi
diukur dengan menggunakan Indeks Sorensen (Cs) = 2j / a+b dimana j adalah
jumlah spesies yang ditemukan di daerah a dan b, a = jumlah spesies yang
ditemukan di daerah a, b = jumlah spesies yang ditemukan di daerah b (Magurran
1988; Kreb 1998). Indeks tersebut dihitung dengan mengggunakan Biodiv97 yang
merupakan perangkat lunak macro pada Microsoft Excel. Matrik yang diperoleh

11
kemudian di analisis lanjut dengan menggunakan analisis kelompok (cluster
analysis) (Krebs 1998). Pengelompokan dalam bentuk dendogram menggunakan
Unweighted Pair-Group Average (UPGMA) dan jarak Euclidean yang dibuat
dengan perangkat lunak Statistica for Windows 6.0 (StatSoft 1995).
Analisis vegetasi dihitung dengan menentukan kerapatan, frekuensi, dan
indeks nilai penting (INP) menggunakan rumus Dumbois-Mueller (Cox 2002)
sebagai berikut:
Jumlah individu suatu spesies i Kerapatan Mutlak (i) = -------------------------------------- Jumlah total luas area petak sampel Kerapatan mutlak spesies i Kerapatan Relatif (i) = --------------------------------- Kerapatan total seluruh spesies Jumlah satuan petak sampel yang diduduki spesies i Frekuensi Mutlak (i) = --------------------------------------------------------------- Jumlah petak sampel yang dibuat dalam analisis Frekuensi mutlak spesies i Frekuensi Relatif (i) = -------------------------------- Frekuensi total seluruh spesies Indeks Nilai Penting (i) = Kerapatan relatif (i) + Frekuensi relatif (i)
HASIL DAN PEMBAHASAN
Distribusi dan Kelimpahan Populasi C. odorata
Tumbuhan eksotik invasif C. odorata merupakan tumbuhan asli dari
Amerika Selatan dan Amerika Tengah yang kemudian menyebar ke Asia
Tenggara (Anonim 1991). Hasil pengamatan pada semua lokasi penelitian di
Jawa Barat khususnya daerah Bogor, C. odorata cenderung menempati lahan
yang terbuka seperti di sisi jalan, tepian sawah yang kondisi tanahnya kering,
ladang, dan perkebunan. Selain itu, C. odorata juga dapat tumbuh di bawah
tegakan hutan yang terbuka tajuknya, bahkan masih bisa hidup pada daerah yang
didominasi oleh batu-batuan. Pada lahan yang dibudidayakan seperti sawah,

12
ladang, dan hutan tanaman industri, keberadaan C. odorata memiliki status
sebagai gulma penting yang merugikan karena populasinya sangat padat sehingga
mampu berkompetisi dengan tanaman budidaya dalam memperoleh unsur hara
yang dibutuhkan (Tjitrosemito 1998).
Persebaran C. odorata saat ini sudah sangat luas yang dapat tumbuh pada
berbagai tipe habitat dengan kelimpahan populasi yang berbeda, namun
kelimpahan populasinya cenderung menurun seiring dengan bertambahnya
ketinggian suatu lokasi. Binggeli (1997) melaporkan bahwa C. odorata hanya
dapat tumbuh pada ketinggian dibawah 1000 mdpl. Keadaan ini dibuktikan dari
hasil survei yang dilakukan di Gunung Bunder, dimana populasi C. odorata hanya
ditemukan sampai pada ketinggian 650 mdpl. Pada ketinggian lebih dari 650
mdpl C. odorata tidak dijumpai tetapi lebih didominasi oleh Austroeupatorium
inulaefolium (L.) (Asteraceae) dengan kelimpahan populasi yang sangat tinggi. A.
inulaefolium merupakan spesies tumbuhan yang secara taksonomi masih memiliki
hubungan kekerabatan dekat dengan C. odorata yang berada dalam satu famili
yaitu Asteraceae (McFadyen et al. 2003).
Untuk melihat kelimpahan populasi C. odorata pada berbagai tipe habitat
yang berbeda, dipilih empat lokasi yang mewakili habitat perkebunan (Parung
Panjang), hutan buatan (Darmaga), ladang dataran rendah (Setu), dan ladang
dataran tinggi (Gunung Bunder). Dari hasil observasi yang dilakukan pada empat
habitat C. odorata memperlihatkan bahwa kelimpahan populasi C. odorata
tertinggi diperoleh pada habitat C. odorata di Parung Panjang dibandingkan
dengan habitat C. odorata lainnya (Gambar 2). Habitat C. odorata di Parung
Panjang adalah perkebunan yang merupakan Hutan Tanaman Industri dengan
tanaman utama Acacia mangium.
Kelimpahan populasi C. odorata pada empat habitat tersebut meningkat
seiring dengan bertambahnya waktu pengambilan sampel yang dilakukan
sebanyak tiga kali kecuali pada habitat C. odorata di Darmaga (Gambar 2).
Kelimpahan populasi C. odorata antara bulan Agustus 2004 dan Mei 2005
mengalami peningkatan, tetapi bukan berasal dari populasi yang sama.
Sedangkan antara bulan Mei 2005 dan Juni 2005 kelimpahan populasi C. odorata

13
juga mengalami peningkatan yang berasal dari populasi waktu pengamatan
sebelumnya. Pada bulan Agustus 2004 kelimpahan populasi C. odorata mencapai
titik terendah kemudian meningkat pada bulan Mei 2005 kecuali pada habitat C.
odorata di Darmaga dan mencapai puncaknya pada bulan Juni 2005.
Menurunnya populasi C. odorata di Darmaga pada bulan Mei 2005 disebabkan
karena adanya pemangkasan di habitat tersebut, namun pada bulan berikutnya
yaitu Juni 2005 kelimpahan populasi C. odorata kembali mengalami peningkatan.
0
2
4
6
8
10
Agustus 2004 Mei 2005 Juni 2005
Waktu pengamatan
Jum
lah
pucu
k/
P. Panjang Darmaga
Setu Gn. Bunder
m2
Gambar 2 Kelimpahan populasi C. odorata pada empat lokasi penelitian selama tiga kali pengambilan sampel.
Rendahnya populasi pucuk C. odorata pada bulan Agustus disebabkan
karena kondisi iklim yang memasuki musim kemarau sehingga menyebabkan
batang C. odorata menjadi kering. Keringnya bagian batang dan daun C. odorata
tidak menyebabkan kematiannya, karena walaupun terlihat seperti mati tetapi akar
yang berada di dalam tanah tetap hidup. Bulan Mei batang kirinyuh tumbuh
kembali sehingga pucuk C. odorata bertambah banyak sampai menjelang waktu
berbunga (fase vegetatif). Kemudian pada bulan berikutnya yaitu bulan Juni
populasi C. odorata akan tumbuh secara serentak sehingga populasinya
meningkat dengan pesat dimana sebagian pucuk telah berubah menjadi bunga
(fase generatif) (Tjitrosemito 1999). Keadaan inilah yang menyebabkan kurang

14
efektifnya lalat puru C. connexa untuk menekan populasi C. odorata karena
kelimpahan populasi lalat puru tidak mampu mengimbangi populasi inangnya
yang tumbuh dengan populasi yang sangat berlimpah.
Distribusi dan Kelimpahan Populasi Lalat Puru C. connexa
Lalat puru C. connexa mulai dilepas pada tahun 1995 setelah memperoleh
surat izin pelepasan yang dikeluarkan oleh Menteri Pertanian. Di Jawa Barat ada
tiga titik pelepasan lalat puru C. connexa yang dilakukan di Parung Panjang-
Jasinga, Bogor pada bulan Desember tahun 1995, Pakuwon-Parung Kuda,
Sukabumi pada bulan Mei dan Juni tahun 1996, dan Cagar Alam Pangandaran,
Ciamis pada bulan Oktober tahun 1999 (Tjitrosemito 1998; Widayanti et al.
2001). Dalam penelitian ini, pengamatan terhadap distribusi lalat puru C. connexa
hanya difokuskan pada titik pelepasan di Parung Panjang-Jasinga (128 mdpl) dan
Pakuwon-Parung Kuda, Sukabumi (300 mdpl). Distribusi lalat puru C. connexa
berdasarkan lokasi penelitian disajikan pada gambar 3.
Gambar 3 Distribusi lalat puru C. connexa dari titik pelepasan Parung Panjang-
Jasinga, Bogor, dan Pakuwon-Parung Kuda, Sukabumi Jawa Barat.

15
Hasil observasi berdasarkan metode survei keberadaan lalat puru C.
connexa, memperlihatkan bahwa setelah 10 tahun dilepas lalat puru C. connexa
memiliki kemampuan menyebar cukup jauh dari titik pelepasannya baik dari
Parung Panjang-Jasinga, Bogor maupun dari Pakuwon-Parung Kuda, Sukabumi.
Secara spesifik, untuk mengetahui keberadaan lalat puru C. connexa pada suatu
lokasi berasal dari titik pelepasan di Parung Panjang-Jasinga, Bogor atau dari
Pakuwon-Parung Kuda, Sukabumi sulit untuk diketahui secara pasti. Walaupun
demikian, keberadaan lalat puru C. connexa tersebut pada suatu lokasi dipastikan
berasal dari titik pelepasan yang terdekat.
Keberadaan lalat puru C. connexa di daerah Setu (128 mdpl), Darmaga (170
mdpl), dan Gunung Bunder (650 mdpl) berasal dari titik pelepasan di Parung
Panjang-Jasinga, Bogor. Walaupun demikian, tidak menutup kemungkinan
bahwasanya keberadaan lalat puru C. connexa di daerah Darmaga dan Gunung
Bunder juga bisa berasal dari titik pelepasan di Pakuwon-Parung Kuda, Sukabumi
(Gambar 3). Di Gunung Bunder yang berjarak 35 km dari titik pelepasan di
Parung Panjang-Jasinga dan 21 km dari titik pelepasan di Pakuwon-Parung Kuda,
Sukabumi (Tabel 2), lalat puru C. connexa masih dijumpai walaupun dengan
kelimpahan populasi yang sangat rendah. Keberadaan lalat puru C. connexa di
Gunung Bunder lebih banyak berasal dari titik pelepasan di Pakuwon-Parung
Kuda, Sukabumi karena memiliki jarak yang lebih dekat dari titik pelepasan
tersebut dibandingkan dari titik pelepasan di Parung Panjang-Jasinga, Bogor.
Di Darmaga yang berjarak relatif hampir sama baik dari titik pelepasan di
Parung Panjang-Jasinga, Bogor (31 km) maupun dari titik pelepasan di Pakuwon-
Parung Kuda, Sukabumi (32 km) (Tabel 2), keberadaan lalat puru C. connexa
berasal dari kedua titik pelepasan tersebut. Keberadaan lalat puru C. connexa di
daerah Ciawi (443 mdpl) dan Cisarua (962 mdpl) berasal dari titik pelepasan yang
terdekat yaitu berasal dari Pakuwon-Parung Kuda, Sukabumi. Begitu juga
keberadaan lalat puru C. connexa yang dijumpai di Gunung Salak (700 mdpl)
yang merupakan distribusi dari titik pelepasan di Pakuwon-Parung Kuda,
Sukabumi.

16
Saat ini lalat puru C. connexa telah mapan dan menyebar secara alami.
Adanya barier seperti pegunungan merupakan faktor pembatas terhadap distribusi
lalat puru C. connexa. Keadaan ini dibuktikan dengan tidak dijumpainya lalat
puru C. connexa tersebut di daerah Cianjur 1 (714 mdpl) dan Cianjur 2 (608
mdpl), serta di Gunung Halimun (650 mdpl), namun keberadaan inangnya C.
odorata pada tiga daerah tersebut masih bisa ditemukan. Adanya barier Gunung
Halimun bagian selatan merupakan faktor pembatas terhadap distribusi lalat puru
C. connexa sehingga tidak ditemukannya lalat puru tersebut di daerah Gunung
Halimun. Di daerah Cianjur, lalat puru C. connexa juga tidak dijumpai karena
adanya barier berupa Gunung Gede dan Gunung Pangranggo sehingga
menyebabkan terbatasnya distribusi lalat puru tersebut.
Kelimpahan populasi lalat puru C. connexa dipelajari secara spesifik pada 4
lokasi penelitian berdasarkan perbedaan habitat dan ketinggian lokasi penelitian.
Empat lokasi tersebut mewakili habitat perkebunan (Parung Panjang), hutan
buatan (Darmaga), ladang dataran rendah (Setu), dan ladang dataran tinggi
(Gunung Bunder). Hasil observasi berdasarkan metode pengukuran populasi pada
empat lokasi penelitian tersebut memperlihatkan bahwa keberadaan C. odorata
diikuti juga dengan keberadaan musuh alaminya yaitu lalat puru C. connexa.
Lalat puru mampu menyebar secara alami dan beradapatasi pada setiap habitat
tetapi memiliki kelimpahan berbeda yang dipengaruhi oleh ketersediaan inangnya,
kondisi habitat, dan keberadaan musuh alami.
Berdasarkan hasil survei, kelimpahan populasi lalat puru C. connexa
meningkat seiring dengan bertambahnya waktu pengambilan sampel kecuali
kelimpahan lalat puru di Gunung Bunder yang relatif hampir sama selama tiga
kali pengambilan sampel (Gambar 4). Kelimpahan populasi lalat puru C. connexa
antara bulan Agustus 2004 dan Mei 2005 mengalami peningkatan, tetapi bukan
berasal dari populasi yang sama. Sedangkan antara bulan Mei 2005 dan Juni 2005
kelimpahan populasi lalat puru C. connexa juga mengalami peningkatan yang
berasal dari populasi waktu pengamatan sebelumnya. Pada bulan Agustus 2004,
kelimpahan populasi lalat puru C. connexa di Parung Panjang, Darmaga dan Setu
memiliki kelimpahan yang terendah, kemudian meningkat pada bulan Mei 2005
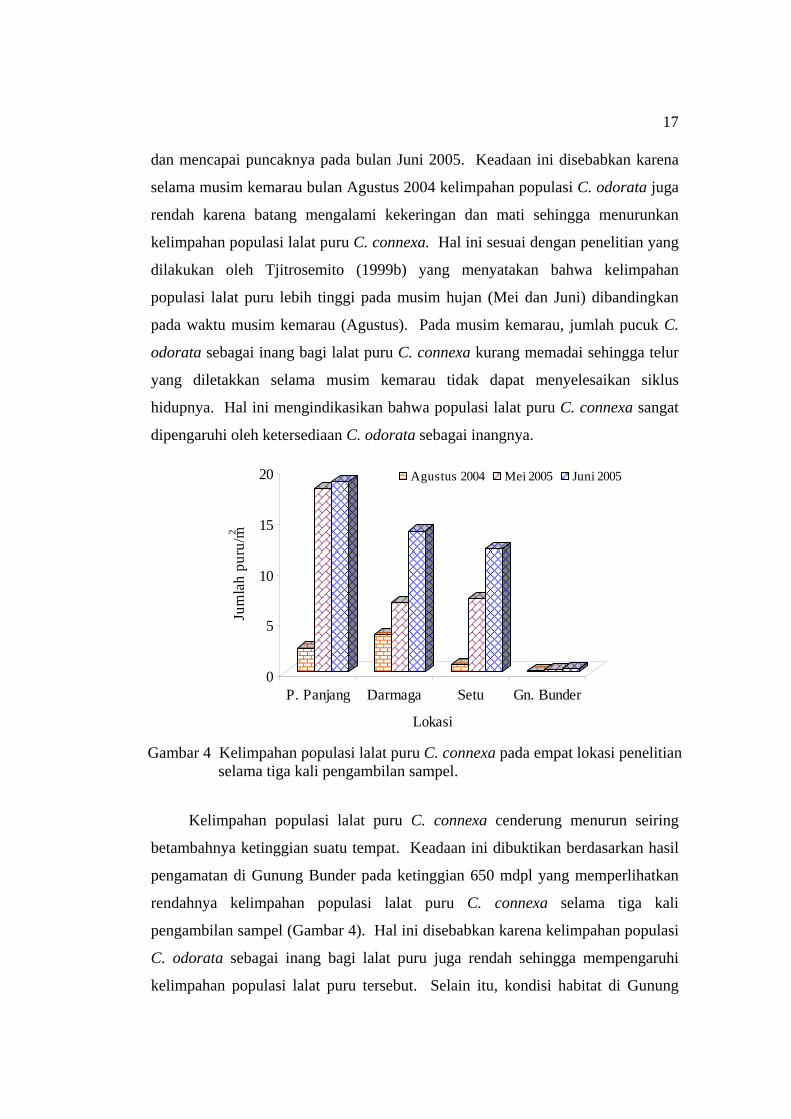
17
dan mencapai puncaknya pada bulan Juni 2005. Keadaan ini disebabkan karena
selama musim kemarau bulan Agustus 2004 kelimpahan populasi C. odorata juga
rendah karena batang mengalami kekeringan dan mati sehingga menurunkan
kelimpahan populasi lalat puru C. connexa. Hal ini sesuai dengan penelitian yang
dilakukan oleh Tjitrosemito (1999b) yang menyatakan bahwa kelimpahan
populasi lalat puru lebih tinggi pada musim hujan (Mei dan Juni) dibandingkan
pada waktu musim kemarau (Agustus). Pada musim kemarau, jumlah pucuk C.
odorata sebagai inang bagi lalat puru C. connexa kurang memadai sehingga telur
yang diletakkan selama musim kemarau tidak dapat menyelesaikan siklus
hidupnya. Hal ini mengindikasikan bahwa populasi lalat puru C. connexa sangat
dipengaruhi oleh ketersediaan C. odorata sebagai inangnya.
Kelimpahan populasi lalat puru C. connexa cenderung menurun seiring
betambahnya ketinggian suatu tempat. Keadaan ini dibuktikan berdasarkan hasil
pengamatan di Gunung Bunder pada ketinggian 650 mdpl yang memperlihatkan
rendahnya kelimpahan populasi lalat puru C. connexa selama tiga kali
pengambilan sampel (Gambar 4). Hal ini disebabkan karena kelimpahan populasi
C. odorata sebagai inang bagi lalat puru juga rendah sehingga mempengaruhi
kelimpahan populasi lalat puru tersebut. Selain itu, kondisi habitat di Gunung
0
5
10
15
20
Jum
lah
puru
/
Agustus 2004 Mei 2005 Juni 2005
m2
P. Panjang Darmaga Setu Gn. Bunder
Lokasi
Gambar 4 Kelimpahan populasi lalat puru C. connexa pada empat lokasi penelitian selama tiga kali pengambilan sampel.

18
Bunder berupa pegunungan yang memiliki perbedaan suhu dan kelembaban
dengan dataran rendah menyebabkan terbatasnya distribusi lalat puru yang
berdampak terhadap rendahnya kelimpahan populasi lalat puru C. connexa di
lokasi tersebut.
Terbentuknya puru pada batang dapat mengurangi pertumbuhan batang,
produksi biji, dan penyimpanan karbohidrat untuk cadangan makanan (Erasmus et
al. 1992). Jika terdapat dalam jumlah banyak, puru batang dapat menghambat
pertumbuhan bahkan mematikan inangnya (Ehler et al. 1984 dalam McFadyen et
al. 2003). Adanya lalat dalam puru batang C. odorata diharapkan mampu untuk
menurunkan populasinya. Berdasarkan pengamatan secara visual di lapangan
memperlihatkan bahwa intensitas serangan lalat puru sangat tinggi terhadap setiap
batang dan pucuk C. odorata, namun keberadaan puru tersebut tidak mematikan
C. odorata. Terbentuknya puru pada batang dan pucuk hanya bisa menghambat
pertumbuhan C. odorata saja. Walaupun lalat puru C. connexa tidak mampu
menurunkan populasi inangnya secara nyata, C. odorata yang telah terinfestasi
oleh puru memiliki produksi biji yang rendah dan proses perkecambahan juga
menurun terutama bila seluruh pucuk terinfestasi oleh puru. Fenomena lain yang
dijumpai adalah banyaknya C. odorata dengan tinggi kurang dari 100 cm, selain
itu terlihat secara umum diameter batang lebih kecil bila dibandingkan dengan
diameter batang yang normal. Chenon et al. 2002 melaporkan bahwa tinggi
batang C. odorata yang terinfestasi oleh lalat puru C. connexa memiliki kisaran
antara 70-85 cm sedangkan panjang batang normal berkisar antara 224-244 cm.
Kekayaan Spesies Tumbuhan di Habitat C. odorata
Dengan menggunakan metode transek yang dikombinasikan dengan metode
kuadrat, diharapkan dapat memberikan gambaran keseluruhan spesies tumbuhan
yang ditemukan pada setiap habitat C. odorata. Keanekaragaman spesies
tumbuhan pada empat habitat C. odorata berdasarkan metode analisis vegetasi
dan jumlah plot yang dibuat memperlihatkan bahwa pengambilan contoh spesies
tumbuhan sudah dapat menggambarkan keseluruhan spesies tumbuhan yang ada
pada masing-masing habitat C. odorata. Hal ini ditunjukkan dengan landainya

19
kurva akumulasi spesies hasil observasi di lapangan pada keempat habitat C.
odorata tersebut (Gambar 5).
Keanekaragaman spesies tumbuhan pada empat habitat C. odorata
memperlihatkan bahwa kekayaan spesies (species richness) tertinggi diperoleh di
Darmaga dan terendah di Parung Panjang (Gambar 5). Hal ini mengindikasikan
bahwa habitat C. odorata di Darmaga memiliki keanekaragaman tumbuhan yang
lebih beragam dibandingkan habitat C. odorata di daerah lain. Berdasarkan nilai
penduga ACE, jumlah spesies tumbuhan yang dikoleksi tertinggi diperoleh di
Darmaga, Gunung Bunder, dan Setu (ACE 100%), sedangkan terendah diperoleh
di Parung Panjang (ACE 95.6%) dari total jumlah spesies yang ada.
5
10
15
20
25
30
35
1 5 9 13 17 21 25 29 33 37 41 45
Jumlah petak contoh
Jum
lah
spes
ies
Darmaga (ACE 100%) Setu (ACE 100%) Gn. Bunder (ACE 100%) P. Panjang (ACE 95.6)
Gambar 5 Kurva akumulasi spesies tumbuhan pada empat habitat C. odorata.
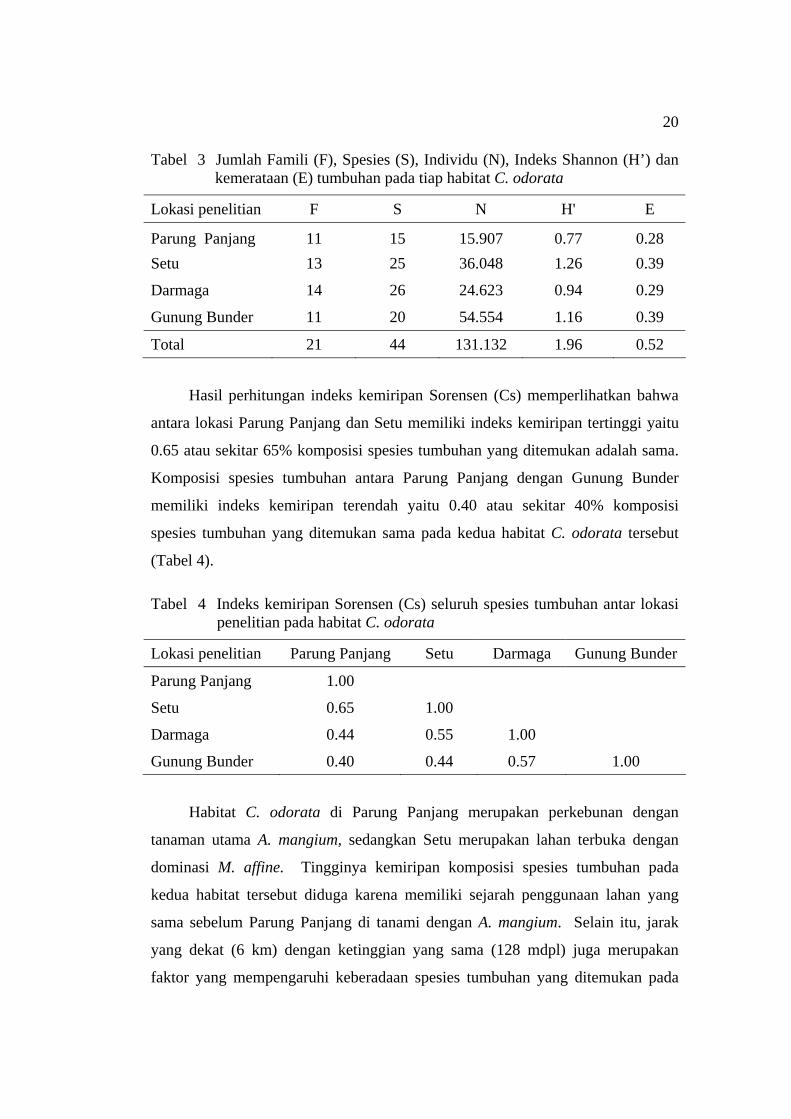
Hasil analisis vegetasi pada empat habitat C. odorata ditemukan sebanyak
131.132 individu tumbuhan yang termasuk ke dalam 21 famili dan 44 spesies
(Tabel 3). Kelimpahan individu (species abudance) tumbuhan terbesar diperoleh
dari habitat C. odorata di Gunung Bunder yaitu sebesar 54.554 individu.
Kekayaan spesies dan jumlah famili tertinggi di dapat di Darmaga yaitu 26 spesies
dan 14 famili. Berdasarkan perhitungan nilai indeks keanekaragaman Shannon
dan kemerataan (evenness), spesies tumbuhan tertinggi diperoleh pada habitat C.
odorata di Setu masing-masing sebesar 1.26 dan 0.39.
20
Tabel 3 Jumlah Famili (F), Spesies (S), Individu (N), Indeks Shannon (H’) dan kemerataan (E) tumbuhan pada tiap habitat C. odorata
Lokasi penelitian F S N H' E
Parung Panjang 11 15 15.907 0.77 0.28 Setu 13 25 36.048 1.26 0.39
Darmaga 14 26 24.623 0.94 0.29
Gunung Bunder 11 20 54.554 1.16 0.39
Total 21 44 131.132 1.96 0.52
Hasil perhitungan indeks kemiripan Sorensen (Cs) memperlihatkan bahwa
antara lokasi Parung Panjang dan Setu memiliki indeks kemiripan tertinggi yaitu
0.65 atau sekitar 65% komposisi spesies tumbuhan yang ditemukan adalah sama.
Komposisi spesies tumbuhan antara Parung Panjang dengan Gunung Bunder
memiliki indeks kemiripan terendah yaitu 0.40 atau sekitar 40% komposisi
spesies tumbuhan yang ditemukan sama pada kedua habitat C. odorata tersebut
(Tabel 4).
Tabel 4 Indeks kemiripan Sorensen (Cs) seluruh spesies tumbuhan antar lokasi penelitian pada habitat C. odorata
Lokasi penelitian Parung Panjang Setu Darmaga Gunung Bunder
Parung Panjang 1.00
Setu 0.65 1.00
Darmaga 0.44 0.55 1.00
Gunung Bunder 0.40 0.44 0.57 1.00
Habitat C. odorata di Parung Panjang merupakan perkebunan dengan
tanaman utama A. mangium, sedangkan Setu merupakan lahan terbuka dengan
dominasi M. affine. Tingginya kemiripan komposisi spesies tumbuhan pada
kedua habitat tersebut diduga karena memiliki sejarah penggunaan lahan yang
sama sebelum Parung Panjang di tanami dengan A. mangium. Selain itu, jarak
yang dekat (6 km) dengan ketinggian yang sama (128 mdpl) juga merupakan
faktor yang mempengaruhi keberadaan spesies tumbuhan yang ditemukan pada
21
kedua lokasi tersebut. Kondisi habitat C. odorata di Gunung Bunder yang
berbatasan langsung dengan lahan persawahan dan tanaman jagung serta berada
pada ketinggian 650 mdpl, diduga menjadi penyebab perbedaan komposisi spesies
tumbuhan dengan habitat C. odorata di Parung Panjang.
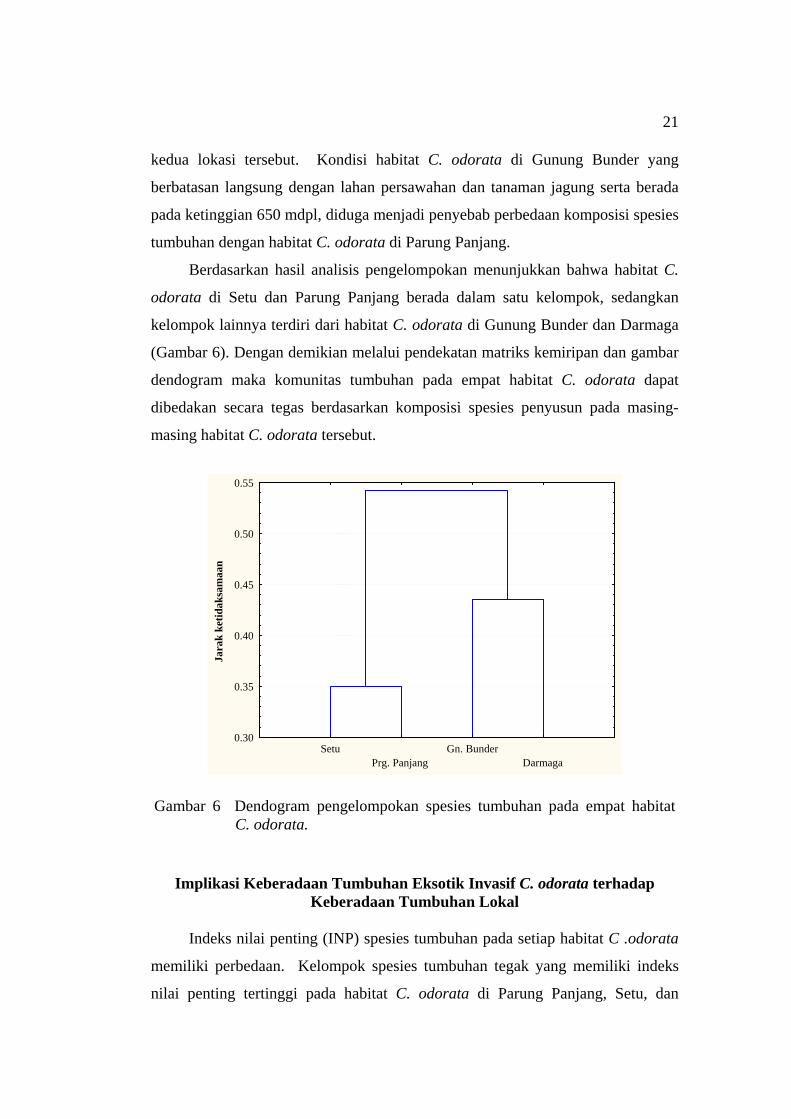
Berdasarkan hasil analisis pengelompokan menunjukkan bahwa habitat C.
odorata di Setu dan Parung Panjang berada dalam satu kelompok, sedangkan
kelompok lainnya terdiri dari habitat C. odorata di Gunung Bunder dan Darmaga
(Gambar 6). Dengan demikian melalui pendekatan matriks kemiripan dan gambar
dendogram maka komunitas tumbuhan pada empat habitat C. odorata dapat
dibedakan secara tegas berdasarkan komposisi spesies penyusun pada masing-
masing habitat C. odorata tersebut.
SetuPrg. Panjang
Gn. BunderDarmaga
0.30
0.35
0.40
0.45
0.50
0.55
Jara
k ke
tidak
sam
aan
Gambar 6 Dendogram pengelompokan spesies tumbuhan pada empat habitat C. odorata.
Implikasi Keberadaan Tumbuhan Eksotik Invasif C. odorata terhadap
Keberadaan Tumbuhan Lokal
Indeks nilai penting (INP) spesies tumbuhan pada setiap habitat C .odorata
memiliki perbedaan. Kelompok spesies tumbuhan tegak yang memiliki indeks
nilai penting tertinggi pada habitat C. odorata di Parung Panjang, Setu, dan

22
Darmaga adalah C. odorata dengan nilai penting masing-masing sebesar 47%,
28.20%, dan 49.57%, sedangkan di Gunung Bunder Boreria alata memiliki
indeks nilai penting tertinggi yaitu sebesar 73.38% (Tabel 5). Untuk kelompok
spesies tumbuhan merambat, indeks nilai penting tertinggi di Parung Panjang dan
Setu adalah Cyrtococcum oxyphyllum dengan nilai penting masing-masing sebesar
131.8% dan 89.97%, di Darmaga indeks nilai penting tertinggi adalah Panicum
repens sebesar 98.34%, sedangkan di Gunung Bunder adalah Axonopus
compressus dengan nilai penting sebesar 72.85% (Tabel 6). Dengan demikian
spesies-spesies yang memiliki indeks nilai penting tertinggi merupakan spesies
yang mempunyai kemampuan adaptasi dan toleransi yang lebih baik dibandingkan
dengan spesies yang lainnya, baik bila dihubungkan dengan pengaruh keberadaan
C. odorata maupun dalam kaitannya dengan kompetisi dengan spesies lain
sehingga spesies-spesies tumbuhan tersebut mendominasi pada setiap habitat C.
odorata tersebut.
Tabel 5 Indeks Nilai Penting (INP) spesies tumbuhan tegak pada empat habitat C. odorata
Lokasi penelitian / INP (%) No. Spesies
P. Panjang Setu Darmaga Gn. Bunder 1 Chromolaena odorata 47.00* 28.20* 49.57* 23.29 2 Boreria leavis 46.18 - 16.26 - 3 Corchorus aestuans 27.56 - - - 4 Ageratum conyzoides - - - 48.28 5 Imperata cylindrica - 25.31 - - 6 Mimosa pudica - - 17.76 - 7 Urena lobata - 25.04 - - 8 Boreria alata - - - 73.38* 9 Spesies lain# 58.96 69.82 95.76 53.49
Total 200 200 200 200 *) INP tertinggi pada setiap habitat C. odorata #) Tetracera scandens, Breynia racemosa, Melastoma affine, Lantana camara, Hydrocotyle sibthorpioides, Phyllanthus
urinaria, Scleria ciliaris, Clidemia hirta, Stachytarpheta jamaicensis, Solanum involucratum, Mimosa invisa, Pennisetum polystachyon, Panicum maximum, Mimosa pigra, Amaranthus spinosus, Nephrolepis bisserata, Cyclosorus aridus, Bergia capendis, Pityrogramma tartara

23
Tabel 6 Indeks Nilai Penting (INP) spesies tumbuhan merambat pada empat habitat C. odorata
Lokasi penelitian / INP (%) No. Spesies P. Panjang Setu Darmaga Gn. Bunder
1 Cyrtococcum oxyphyllum 131.8* 89.97* - - 2 Axonopus compressus 48.60 37.11 19.13 72.85* 3 Lygodium microphyllum 19.55 - - - 4 Operculina turpethum - 28.60 - - 5 Ipomoea triloba - - 20.89 - 6 Panicum repens - - 98.34* - 7 Ischaemum timorense - - - 50.93 8 Rostellularia sundana - - - 18.36 9 Spesies lain 0 44.3 61.6 57.8
Total 200 200 200 200 *) INP tertinggi pada setiap habitat C. odorata #) Chrysopogon aciculatus, Mikania micrantha, Centrosema pubescen, Widelia trilobata, Eleusine indica, Commelina
diffusa, Setaria palmifolia, Cyperus kyllingia, Eragrostis unioloides
Hasil perhitungan indeks nilai penting untuk spesies tumbuhan eksotik
invasif C. odorata di Parung Panjang, Setu, dan Darmaga memperlihatkan bahwa
kelimpahan populasi tumbuhan ini mendominasi pada tiga habitat tersebut kecuali
di Gunung Buder (Tabel 5). Keadaan ini mengindikasikan bahwa kehadiran
spesies tumbuhan eksotik invasif C. odorata pada tiga habitat tersebut telah
menyebabkan terjadinya pengambilalihan atau pergantian skala ruang, yaitu
tempat atau ruang yang seharusnya ditempati oleh spesies-spesies tumbuhan lokal
kemudian diambil alih keberadaannya oleh tumbuhan eksotik invasif C. odorata.
Selain itu, kehadiran tumbuhan eksotik invasif C. odorata pada empat lokasi
penelitian menyebabkan terjadinya penurunan keanekaragaman spesies tumbuhan
yang berada di sekitarnya. Hal ini diperkuat berdasarkan hasil perhitungan indeks
keanekaragaman Shannon-Wiener yang memperlihatkan rendahnya nilai indeks
keanekaragaman tumbuhan pada keempat habitat C. odorata tersebut (Tabel 3).
Pada 8 minggu pertama setelah tumbuh dari biji, C. odorata
mengalokasikan sebagian besar hasil fotosintesis atau biomassanya untuk
pembentukan daun. Setelah 8 minggu pertama, hasil fotosintesis atau
biomassanya diarahkan untuk pembentukan batang sehingga C. odorata
membentuk tumbuhan yang banyak daunnya dan terlihat rindang, padat serta rapat

24
jumlah daunnya. Keadaan seperti ini menyebabkan C. odorata bersifat agresif
karena akan segera menutupi pertumbuhan kecambah spesies tumbuhan lain yang
kurang cepat pertumbuhannya sehingga akan menghambat pertumbuhan
kecambah spesies tumbuhan tersebut dan bahkan dapat menyebabkan kematian
kecambah (Tjitrosemito 1997).
Keberadaan tumbuhan eksotik invasif C. odorata ini pertumbuhannya akan
berkompetisi dengan spesies-spesies tumbuhan lain yang berada dibawahnya baik
dari kelompok tumbuhan tegak maupun yang merambat. Apabila C. odorata ini
menaungi spesies-spesies tumbuhan yang berada di bawahnya secara terus
menerus dalam jangka waktu yang lama, maka dapat menyebabkan hilangnya
spesies-spesies tumbuhan yang berada di bawah naungan C. odorata tersebut
(Tjitrosemito 22 Agustus 2006, komunikasi pribadi).
KESIMPULAN
Persebaran spesies tumbuhan eksotik invasif C. odorata di Jawa Barat
khususnya daerah Bogor sudah sangat luas. C. odorata dapat tumbuh pada
berbagai tipe habitat meliputi habitat perkebunan, hutan buatan, ladang dataran
rendah, maupun ladang dataran tinggi dengan kelimpahan yang berbeda.
Kelimpahan populasinya cenderung menurun seiring dengan bertambahnya
ketinggian suatu lokasi.
Keberadaan C. odorata diikuti pula dengan keberadaan musuh alaminya
yaitu lalat puru C. connexa pada semua lokasi pengamatan, kecuali di Gunung
Halimun dan Cianjur. Ada dua titik pelepasan lalat puru C. connexa yaitu di
Parung Panjang-Jasinga, Bogor dan Pakuwon-Parung Kuda, Sukabumi. Lalat
puru C. connexa saat ini telah mapan keberadaannya dan menyebar secara alami
serta memiliki kemampuan persebaran yang cukup jauh baik dari titik
pelepasannya di Parung Panjang-Jasinga, Bogor maupun dari titik pelepasan di
Pakuwon-Parung Kuda, Sukabumi. Lalat puru C. connexa mampu hidup pada
berbagai ketinggian tempat yang berbeda, namun kelimpahan populasinya
menurun seiring dengan bertambahnya ketinggian suatu tempat. Adanya barier

25
berupa pegunungan merupakan faktor pembatas terhadap distribusi lalat puru C.
connexa. Kelimpahan populasi lalat puru C. connexa sangat dipengaruhi oleh
ketersediaan inangnya yaitu C. odorata.
Komunitas tumbuhan yang diperoleh pada habitat C. odorata di Parung
Panjang, Setu, Darmaga, dan Gunung Bunder secara keseluruhan terdiri dari
131.132 individu tumbuhan yang termasuk ke dalam 21 famili dan 44 spesies.
Tingkat kemiripan komposisi spesies tumbuhan tertinggi terdapat pada habitat C.
odorata antara Parung Panjang dan Setu yaitu 65%, sedangkan yang terendah
diperoleh pada habitat C. odorata antara Parung Panjang dengan Gunung Bunder
sebesar 40%.
Kelimpahan populasi C. odorata pada lokasi penelitian di Parung Panjang,
Setu, dan Darmaga mendominasi berdasarkan perhitungan indeks nilai penting
kecuali di Gunung Bunder. Implikasi dari keberadaan spesies tumbuhan eksotik
invasif C. odorata pada tiga habitat tersebut telah menyebabkan terjadinya
pengambilalihan atau pergantian skala ruang, yaitu tempat atau ruang yang
seharusnya ditempati oleh spesies-spesies tumbuhan lokal kemudian diambil alih
keberadaannya oleh tumbuhan eksotik invasif C. odorata. Selain itu, kehadiran
tumbuhan eksotik invasif C. odorata pada empat lokasi penelitian menyebabkan
terjadinya penurunan keanekaragaman spesies tumbuhan yang berada di
sekitarnya.

STRUKTUR KOMUNITAS SERANGGA PADA HABITAT TUMBUHAN EKSOTIK INVASIF: STUDI KASUS PADA HABITAT Chromolaena odorata (L.) KING & ROBINSON
(ASTERACEAE) DI BOGOR, JAWA BARAT
PENDAHULUAN
Keberadaan spesies eksotik invasif merupakan suatu ancaman yang serius
terhadap ekosistem dan keanekaragaman hayati (Wittenberg & Cock 2003).
Spesies eksotik invasif dapat menyebabkan homogenisasi biotik, merubah suatu
komunitas, dan kepunahan spesies lokal (Olden et al. 2004; Untung 2005).
Sebagai contoh, kirinyuh atau Chromolaena odorata (Asteraceae) adalah spesies
tumbuhan eksotik invasif. Kemampuan adaptasinya dapat mendominasi suatu
habitat, seperti di hutan lindung Panunjang, Jawa Barat dan Taman Nasional
Baluran, Jawa Timur (Tjitrosemito 1998).
Pengendalian kirinyuh dengan memanfaatkan agens hayatinya telah
dilakukan dengan mengintroduksi lalat pembentuk puru Cecidochares connexa
Macquart (Diptera: Tephritidae) pada tahun 1993 dari Columbia (McFadyen et al.
2003). Di Jawa Barat lalat puru C. connexa mulai dilepas pada tahun 1995 di
Hutan Tanaman Industri Parung Panjang-Jasinga, Bogor (Tjitrosemito 1998).
Sampai saat ini lalat puru C. connexa telah mapan keberadaannya dan menyebar
secara alami.
Keberadaan spesies eksotik dapat dikendalikan dengan teknik pengendalian
hayati, seperti C. odorata yang dikendalikan dengan lalat puru C. connexa.
Namun tidak semua pengendalian spesies eksotik berhasil menurunkan populasi
inangnya. Kemapanan populasi agens hayati yang tidak mampu menurunkan
populasi inangnya dapat memfasilitasi efek bottom-up yang menghubungkan
gulma sasaran dengan organisme lokal lainnya (Pearson & Callaway 2003).
Pengendalian hayati merupakan teknik pengendalian yang penting dalam
menghadapi invasi spesies eksotik, namun agens hayati tersebut dapat berdampak
merugikan pada spesies lokal. Dari beberapa hasil penelitian memperlihatkan
bahwa agens hayati yang inangnya spesifik dapat menimbulkan efek tidak

27
langsung (indirect effects) melalui interaksi dan subsidi jaring-jaring makanan.
Efek tidak langsung dapat terjadi melalui penggantian ekosistem ketika agens
hayati secara fisik dan fungsional menggantikan spesies lokal (Pearson &
Callaway 2003).
Sampai saat ini, studi ekologi mengenai dampak keberadaan spesies ekostik
invasif C. odorata belum pernah dilakukan, seperti penelitian bagaimana dampak
keberadaan C. odorata dan agens hayati C. connexa terhadap struktur komunitas
serangga lokal. Berdasarkan permasalahan di atas, maka diperlukan suatu
penelitian untuk mengetahui struktur komunitas serangga pada habitat C. odorata.
Tujuan penelitian ini adalah untuk mempelajari struktur komunitas serangga di
daerah yang telah diinvasi oleh C. odorata dengan mengambil studi kasus di
daerah Bogor, Jawa Barat.
BAHAN DAN METODE
Lokasi dan Waktu Penelitian
Lokasi penelitian ditentukan berdasarkan metode purposive sampling, yaitu
lokasi yang dijumpai C. odorata, dengan pertimbangan tipe habitat dan ketinggian
tempat. Dari hasil survei pendahuluan maka ditetapkan daerah yang terpilih
sebagai tempat pengambilan sampel: (1) Hutan Tanaman Industri (HTI) Parung
Panjang-Jasinga mewakili habitat perkebunan dan tempat pelepasan lalat puru
C. connexa, (2) Desa Setu-Jasinga mewakili habitat ladang dataran rendah, (3)
Kampus IPB Darmaga mewakili habitat hutan buatan, dan (4) Gunung Bunder
mewakili habitat ladang dataran tinggi (Tabel 7). Penelitian dilakukan dari bulan
Agustus 2004 hingga Juni 2005.
Pengambilan Contoh Serangga
Pada masing-masing lokasi dibuat 5 jalur transek sepanjang 30 m dengan
jarak antara transek 15 m (Gambar 1). Sepanjang jalur transek dibuat petak
contoh pengambilan sampel berbentuk bujur sangkar (2 m x 2 m) dengan jarak 15
m, sehingga pada masing-masing transek terdapat 3 petak contoh, jadi ada 15
28
petak contoh pada setiap lokasi penelitian. Pengambilan sampel dilakukan
sebanyak 3 kali pada setiap lokasi penelitian, yaitu bulan Agustus 2004, Mei
2005, dan Juni 2005.
Tabel 7 Deskripsi lokasi yang dipilih untuk penelitian
Lokasi/Desa Koordinat (LS dan BT)1)
Ketinggian (mdpl)2)
Jarak (km)3) Kondisi Habitat
Parung Panjang 060.42 LS 1060.48 BT
128 0 Hutan tanaman industri dengan tanaman utama Acacia mangium disertai populasi C. odorata dominan
Setu 060.46 LS 1060.45 BT
128 6 Lahan terbuka dengan dominasi Melastoma affine dan C. odorata yang berbatasan dengan lahan tanaman budidaya (kacang tanah, kedelai, dan jagung)
Darmaga 060.55 LS 1060.72 BT
170 30 Tanaman sengon (Albizia falcataria) dengan dominasi C. odorata dan Widelia trilobata yang berbatasan dengan jalan raya
Gunung Bunder 060.66 LS 1060.69 BT
650 35 Lahan terbuka dengan dominasi Ageratum conyzoides dan Boreria alata yang berbatasan dengan lahan persawahan dan tanaman budidaya (jagung, talas, pisang, kopi, dan singkong)
1) LS = Lintang selatan, BT = Bujur timur, 2) Meter di atas permukaan laut, 3) Jarak lokasi dari titik pelepasan C. connexa
Pengambilan sampel serangga pada setiap petak contoh menggunakan
perangkap jebak (pitfall trap), perangkap nampan kuning (yellow pan trap), dan
perangkap malaise (malaise trap) (Gambar 7). Perangkap jebak terbuat dari gelas
plastik bekas air mineral volume 220 ml, diameter mulut gelas 7 cm dan tinggi 10
cm. Gelas diisi 110 ml larutan air sabun dan garam secukupnya untuk
mengurangi tegangan permukaan, sehingga serangga yang terperangkap
tenggelam dan mati Perangkap jebak ditempatkan sebanyak 4 buah pada setiap
petak contoh sehingga terdapat 60 buah perangkap jebak yang dipasang selama 24
jam pada setiap lokasi penelitian.

29
Gambar 7 Perangkap serangga yang digunakan; (a) yellow pan trap (b) pitfall trap (c) malaise trap.
a b c
Nampan kuning yang digunakan adalah wadah plastik berukuran 15 cm x 25
cm x 7 cm. Nampan kuning diisi dengan air sabun dan garam secukupnya untuk
mengurangi tegangan permukaan, sehingga serangga yang terperangkap
tenggelam dan mati. Selanjutnya nampan kuning diletakkan pada permukaan
tanah sebanyak 2 buah dalam setiap petak contoh, sehingga terdapat 30 buah
nampan kuning yang dipasang selama 24 jam pada setiap lokasi penelitian.
Perangkap malaise terbuat dari jaring yang berbentuk seperti tenda
(berbentuk prisma). Pada bagian puncaknya dipasang botol plastik yang berfungsi
sebagai perangkap. Perangkap malaise efektif untuk serangga yang aktif terbang,
serangga terbang akan menabrak jaring kemudian serangga akan bergerak ke atas
mengikuti pola jaring menuju botol perangkap. Perangkap ditempatkan secara
diagonal masing-masing diantara transek 2-3 dan transek 4-5, sehingga terdapat 2
buah perangkap malaise yang dipasang selama 24 jam. Serangga yang tertangkap
dengan perangkap jebak, nampan kuning, dan malaise dibersihkan dari kotoran.
Selanjutnya disimpan dalam tabung film berisi alkohol 70% untuk diidentifikasi
di laboratorium.
Sortasi dan identifikasi sampel serangga yang dikoleksi dari lapangan
dilakukan di Laboratorium Bioekologi Parasitoid dan Predator, Departemen
Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor. Semua serangga
yang diperoleh dipisahkan berdasarkan ordonya. Selanjutnya identifikasi
dilanjutkan sampai morfospesies tingkat famili (hanya diberi kode). Identifikasi
serangga untuk tingkat famili mengacu pada kunci identifikasi yang tersedia.

30
Analisis Data
Data keseluruhan spesies serangga yang diperoleh pada setiap lokasi dapat
diduga dengan menggunakan kurva akumulasi spesies yang dibuat dengan
program EstimateS 6.0b1 (Colwell 2000). Jumlah spesies serangga yang
diperoleh pada setiap petak contoh diacak sebanyak 50 kali menggunakan
program tersebut. Prediksi kekayaan spesies serangga diduga dengan abundance-
based coverage estimator (ACE) (Colwell & Coddington 1994).
Indeks keanekaragaman serangga diukur berdasarkan Shannon-Wiener (H’)
= -Σ pi ln pi dimana pi = proporsi spesies ke-i terhadap total jumlah spesies,
Indeks kemerataan berdasarkan Shannon-Wiener (E) = H’/ln (S) dimana S = total
jumlah spesies yang diperoleh. Kemiripan komunitas serangga antar lokasi diukur
dengan menggunakan Indeks Sorensen (Cs) = 2j / a+b dimana j adalah jumlah
spesies yang ditemukan di daerah a dan b, a = jumlah spesies yang ditemukan di
daerah a, b = jumlah spesies yang ditemukan di daerah b (Magurran 1988; Kreb
1998). Indeks tersebut dihitung dengan mengggunakan Biodiv97 yang merupakan
perangkat lunak macro pada Microsoft Excel. Matrik yang diperoleh kemudian
dianalisis lanjut dengan menggunakan analisis kelompok (cluster analysis) (Krebs
1998). Pengelompokan dalam bentuk dendogram menggunakan Unweighted
Pair-Group Average (UPGMA) dan jarak Euclidean yang dibuat dengan
perangkat lunak Statistica for Windows 6.0 (StatSoft 1995).
HASIL DAN PEMBAHASAN
Kekayaan Spesies Serangga di Habitat C. odorata
Keanekaragaman spesies serangga pada empat habitat C. odorata
berdasarkan perangkap yang digunakan memperlihatkan bahwa kekayaan spesies
(species richness) tertinggi diperoleh di Darmaga dan terendah di Parung Panjang
(Gambar 8). Keadaan ini mengindikasikan bahwa habitat C. odorata di Darmaga
memiliki keanekaragaman serangga yang lebih beragam dibandingkan habitat C.
odorata di daerah lain. Berdasarkan nilai penduga ACE, jumlah spesies serangga
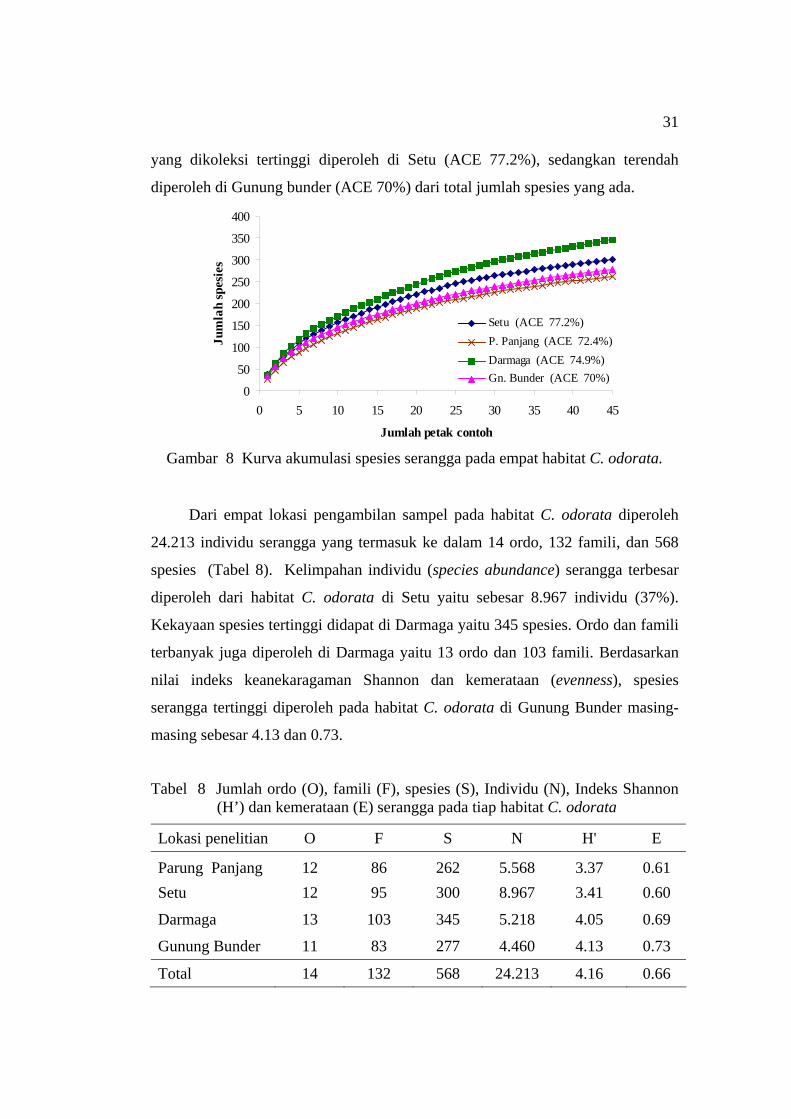
31
yang dikoleksi tertinggi diperoleh di Setu (ACE 77.2%), sedangkan terendah
diperoleh di Gunung bunder (ACE 70%) dari total jumlah spesies yang ada.
Gambar 8 Kurva akumulasi spesies serangga pada empat habitat C. odorata.
0
50
100
150
200
250
300
350
400
0 5 10 15 20 25 30 35 40 45
Jumlah petak contoh
Jum
lah
spes
ies
Setu (ACE 77.2%)P. Panjang (ACE 72.4%)Darmaga (ACE 74.9%)Gn. Bunder (ACE 70%)
Dari empat lokasi pengambilan sampel pada habitat C. odorata diperoleh
24.213 individu serangga yang termasuk ke dalam 14 ordo, 132 famili, dan 568
spesies (Tabel 8). Kelimpahan individu (species abundance) serangga terbesar
diperoleh dari habitat C. odorata di Setu yaitu sebesar 8.967 individu (37%).
Kekayaan spesies tertinggi didapat di Darmaga yaitu 345 spesies. Ordo dan famili
terbanyak juga diperoleh di Darmaga yaitu 13 ordo dan 103 famili. Berdasarkan
nilai indeks keanekaragaman Shannon dan kemerataan (evenness), spesies
serangga tertinggi diperoleh pada habitat C. odorata di Gunung Bunder masing-
masing sebesar 4.13 dan 0.73.
Tabel 8 Jumlah ordo (O), famili (F), spesies (S), Individu (N), Indeks Shannon (H’) dan kemerataan (E) serangga pada tiap habitat C. odorata
Lokasi penelitian O F S N H' E
Parung Panjang 12 86 262 5.568 3.37 0.61 Setu 12 95 300 8.967 3.41 0.60
Darmaga 13 103 345 5.218 4.05 0.69
Gunung Bunder 11 83 277 4.460 4.13 0.73
Total 14 132 568 24.213 4.16 0.66
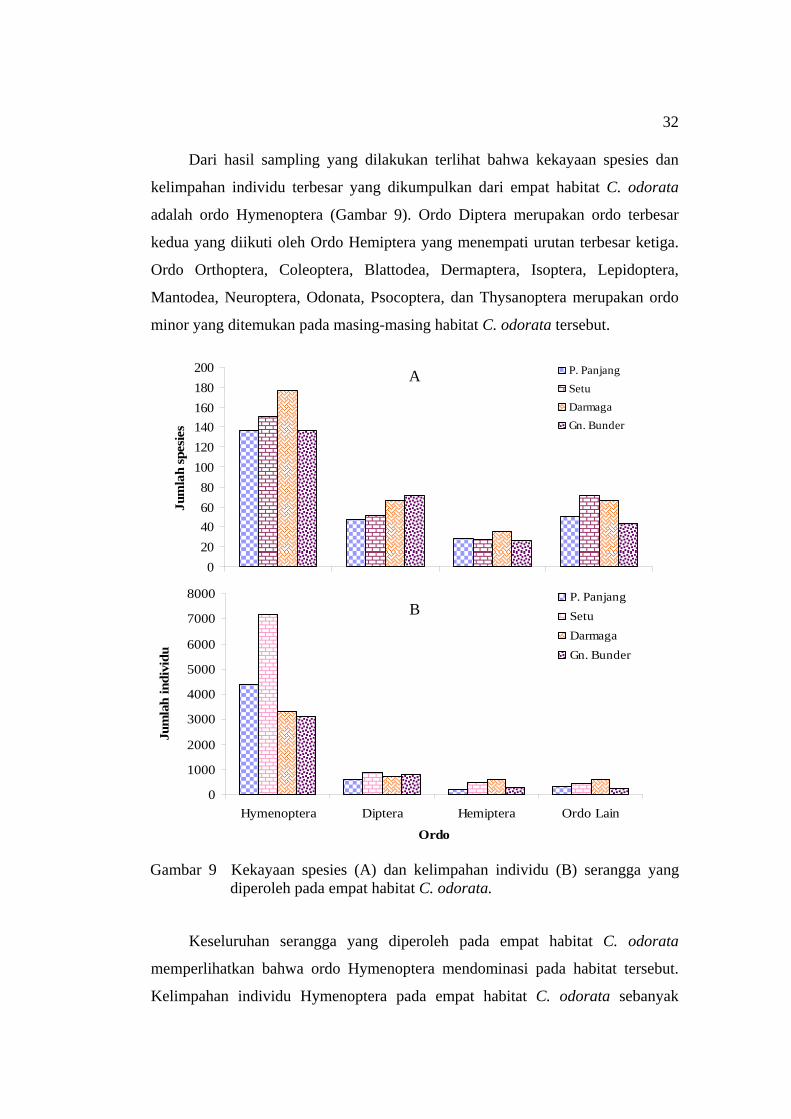
32
Dari hasil sampling yang dilakukan terlihat bahwa kekayaan spesies dan
kelimpahan individu terbesar yang dikumpulkan dari empat habitat C. odorata
adalah ordo Hymenoptera (Gambar 9). Ordo Diptera merupakan ordo terbesar
kedua yang diikuti oleh Ordo Hemiptera yang menempati urutan terbesar ketiga.
Ordo Orthoptera, Coleoptera, Blattodea, Dermaptera, Isoptera, Lepidoptera,
Mantodea, Neuroptera, Odonata, Psocoptera, dan Thysanoptera merupakan ordo
minor yang ditemukan pada masing-masing habitat C. odorata tersebut.
020406080
Keseluruhan serangga yang diperoleh pada empat habitat C. odorata
memperlihatkan bahwa ordo Hymenoptera mendominasi pada habitat tersebut.
Kelimpahan individu Hymenoptera pada empat habitat C. odorata sebanyak
100120
200
Hymenoptera Diptera Hemiptera Ordo Lain
Ordo
Jum
lah
spes
ies
P. PanjangA 180 Setu
160 DarmagaGn. Bunder140
0
1000
2000
3000
4000
5000
6000
7000
8000
Hymenoptera Diptera Hemiptera Ordo Lain
Ordo
Jum
lah
indi
vid
P. PanjangB Setu
Darmaga
u Gn. Bunder
Gambar 9 Kekayaan spesies (A) dan kelimpahan individu (B) serangga yang diperoleh pada empat habitat C. odorata.
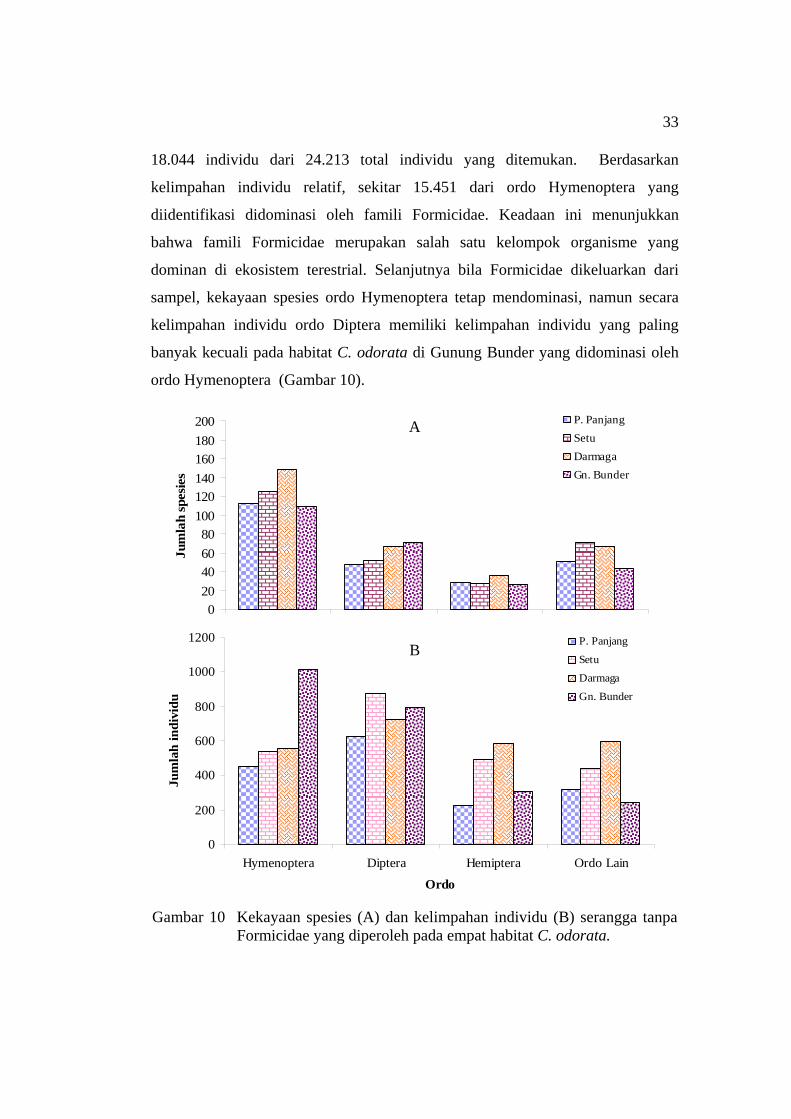
33
18.044 individu dari 24.213 total individu yang ditemukan. Berdasarkan
kelimpahan individu relatif, sekitar 15.451 dari ordo Hymenoptera yang
diidentifikasi didominasi oleh famili Formicidae. Keadaan ini menunjukkan
bahwa famili Formicidae merupakan salah satu kelompok organisme yang
dominan di ekosistem terestrial. Selanjutnya bila Formicidae dikeluarkan dari
sampel, kekayaan spesies ordo Hymenoptera tetap mendominasi, namun secara
kelimpahan individu ordo Diptera memiliki kelimpahan individu yang paling
banyak kecuali pada habitat C. odorata di Gunung Bunder yang didominasi oleh
ordo Hymenoptera (Gambar 10).
020406080
100120140160180200
Hymenoptera Diptera Hemiptera Ordo Lain
Ordo
Jum
lah
spes
ies
P. PanjangA SetuDarmagaGn. Bunder
0
200
400
600
800
1000
1200
Hymenoptera Diptera Hemiptera Ordo Lain
Ordo
Jum
lah
indi
vidu
P. Panjang
Setu
Darmaga Gn. Bunder
B
Gambar 10 Kekayaan spesies (A) dan kelimpahan individu (B) serangga tanpa Formicidae yang diperoleh pada empat habitat C. odorata.
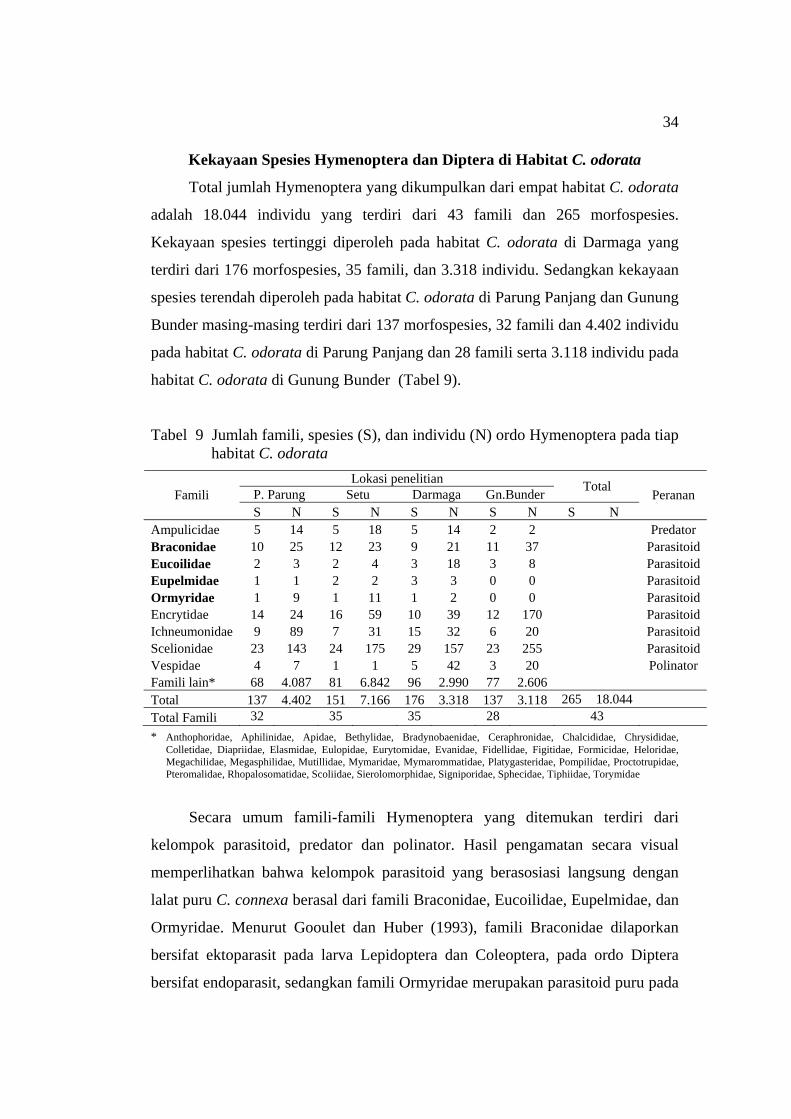
34
Kekayaan Spesies Hymenoptera dan Diptera di Habitat C. odorata
Total jumlah Hymenoptera yang dikumpulkan dari empat habitat C. odorata
adalah 18.044 individu yang terdiri dari 43 famili dan 265 morfospesies.
Kekayaan spesies tertinggi diperoleh pada habitat C. odorata di Darmaga yang
terdiri dari 176 morfospesies, 35 famili, dan 3.318 individu. Sedangkan kekayaan
spesies terendah diperoleh pada habitat C. odorata di Parung Panjang dan Gunung
Bunder masing-masing terdiri dari 137 morfospesies, 32 famili dan 4.402 individu
pada habitat C. odorata di Parung Panjang dan 28 famili serta 3.118 individu pada
habitat C. odorata di Gunung Bunder (Tabel 9).
Tabel 9 Jumlah famili, spesies (S), dan individu (N) ordo Hymenoptera pada tiap habitat C. odorata
Lokasi penelitian P. Parung Setu Darmaga Gn.Bunder Total
Famili S N S N S N S N S N
Peranan
Ampulicidae 5 14 5 18 5 14 2 2 Predator Braconidae 10 25 12 23 9 21 11 37 Parasitoid Eucoilidae 2 3 2 4 3 18 3 8 Parasitoid Eupelmidae 1 1 2 2 3 3 0 0 Parasitoid Ormyridae 1 9 1 11 1 2 0 0 Parasitoid Encrytidae 14 24 16 59 10 39 12 170 Parasitoid Ichneumonidae 9 89 7 31 15 32 6 20 Parasitoid Scelionidae 23 143 24 175 29 157 23 255 Parasitoid Vespidae 4 7 1 1 5 42 3 20 Polinator Famili lain* 68 4.087 81 6.842 96 2.990 77 2.606 Total 137 4.402 151 7.166 176 3.318 137 3.118 265 18.044 Total Famili 32 35 35 28 43 * Anthophoridae, Aphilinidae, Apidae, Bethylidae, Bradynobaenidae, Ceraphronidae, Chalcididae, Chrysididae,
Colletidae, Diapriidae, Elasmidae, Eulopidae, Eurytomidae, Evanidae, Fidellidae, Figitidae, Formicidae, Heloridae, Megachilidae, Megasphilidae, Mutillidae, Mymaridae, Mymarommatidae, Platygasteridae, Pompilidae, Proctotrupidae, Pteromalidae, Rhopalosomatidae, Scoliidae, Sierolomorphidae, Signiporidae, Sphecidae, Tiphiidae, Torymidae
Secara umum famili-famili Hymenoptera yang ditemukan terdiri dari
kelompok parasitoid, predator dan polinator. Hasil pengamatan secara visual
memperlihatkan bahwa kelompok parasitoid yang berasosiasi langsung dengan
lalat puru C. connexa berasal dari famili Braconidae, Eucoilidae, Eupelmidae, dan
Ormyridae. Menurut Gooulet dan Huber (1993), famili Braconidae dilaporkan
bersifat ektoparasit pada larva Lepidoptera dan Coleoptera, pada ordo Diptera
bersifat endoparasit, sedangkan famili Ormyridae merupakan parasitoid puru pada

35
lalat famili Cecidomyiidae dan Tephritidae (Ordo Diptera). Famili Eucoilidae
merupakan parasitoid pupa pada ordo Diptera (Boror et al. 1981), sedangkan
famili Eupelmidae bersifat parasitoid pada Ordo Diptera (famili Cecidomyiidae)
(Nauman 1991). Famili Braconidae dan Eucoilidae masing-masing terdiri dari 22
morfospesies dan 3 morfospesies yang ditemukan pada empat habitat C. odorata.
Famili Eupelmidae terdiri dari 3 morfospesies sedangkan famili Ormyridae hanya
terdiri dari 1 morfospesies yang dijumpai pada habitat C. odorata di Parung
Panjang, Setu, dan Darmaga, sedangkan di Gunung Bunder tidak ditemukan.
Hasil penelitian ini menunjukkan bahwa Hymenoptera dari kelompok
parasitoid memiliki kekayaan spesies dan kelimpahan individu yang lebih tinggi
dibandingkan Hymenoptera dari kelompok polinator maupun predator (Tabel 9).
Famili Braconidae, Encyrtidae, Ichneumonidae, dan Scelionidae memiliki jumlah
spesies dan kelimpahan individu yang paling tinggi pada habitat C. odorata di
Parung Panjang, Setu, Darmaga, dan Gunung Bunder.
Tingginya kekayaan spesies dan kelimpahan individu kelompok
Hymenoptera parasitoid pada empat habitat C. odorata tidak terlepas dari
ketersediaan tumbuhan berbunga pada masing-masing habitat tersebut. Terdapat
lima spesies tumbuhan yang diperoleh pada empat habitat C. odorata, yaitu
Ageratum conyzoides, Melastoma affine, Mimosa pudica, Phyllanthus urinaria,
dan Axonopus compressus (Tabel Lampiran 1 sampai 8). Vegetasi ini merupakan
tumbuhan liar yang umum dijumpai di sekitar habitat ladang, sawah, dan
perkebunan. Tumbuhan liar merupakan tempat mengungsi, berlindung, dan
bertahan hidup bagi Hymenoptera dari kelompok parasitoid. Altieri dan Nicholls
(2004) melaporkan bahwa tumbuhan liar dapat berperan sebagai tempat
berlindung atau pengungsian musuh alami apabila kondisi pertanaman tidak
sesuai seperti ketika tanaman tidak ada (saat panen), pemberaan, dan perlakuan
insektisida. Selain itu, hasil penelitian Kartosuwondo (2001) melaporkan bahwa
ketersediaan tumbuhan berbunga pada suatu ekosistem tidak hanya mampu
meningkatkan lama hidup dan keperidian parasitoid, tetapi juga meningkatkan
keanekaragaman spesies parasitoid.

36
Ordo Diptera yang berhasil dikumpulkan dari empat habitat C. odorata
sebanyak 3.015 individu yang terdiri dari 35 famili dan 120 morfospesies.
Kekayaan spesies tertinggi diperoleh pada habitat C. odorata di Gunung Bunder
yang terdiri dari 71 morfospesies, 25 famili, dan 795 individu. Sedangkan
kekayaan spesies terendah diperoleh pada habitat C. odorata di Parung Panjang
yang terdiri dari 47 morfospesies, 22 famili dan 627 individu (Tabel 10). Famili-
famili Diptera yang ditemukan terdiri dari kelompok herbivor, predator, saprofag,
dan polinator.
Tabel 10 Jumlah famili, spesies (S), dan individu (N) ordo Diptera pada tiap habitat C. odorata
Lokasi penelitian P. Parung Setu Darmaga Gn.Bunder Total
Famili S N S N S N S N S N
Peranan
Cecidomyiidae 2 4 2 46 1 5 2 25 Herbivor Dolicophodidae 6 214 7 273 6 193 8 95 Predator Phoridae 4 123 4 260 8 155 5 229 Saprofag Tabanidae 2 38 2 2 1 3 1 1 Polinator Tephritidae 1 5 2 5 1 2 2 3 Herbivor Famili lain 30 238 32 284 49 360 50 437 Total 47 627 51 872 67 721 71 795 120 3.015 Total Famili* 22 23 29 25 35
* Anthomyiidae, Asteiidae, Calliphoridae, Carnidae, Ceratopogonidae, Chamaemyiidae, Chloropidae, Culicidae, Curtonotidae, Diastatidae, Diopsidae, Drosophilidae, Ephydridae, Lauxanidae, Micropezidae, Muscidae, Mycetophilidae, Otitidae, Pipunculidae, Sarcophagidae, Scathophagidae, Scatopsidae, Sciaridae, Sphaeroceridae, Stratiomyidae, Syrphidae, Tachinidae, Tethinidae, Therevidae, Tipulidae
Kelompok herbivor terdiri dari 2 famili yaitu famili Cecidomyiidae (3
morfospesies) dan Tephritidae (3 morfospesies selain C. connexa). Menurut
Boror et al. (1981) famili Cecidomyiidae merupakan kelompok pembentuk puru
pada batang dan daun tanaman, sedangkan famili Tephritidae selain menyebabkan
terbentuknya puru pada tanaman juga menyebabkan kerusakan pada buah.
Keberadaan spesies dari famili Cecidomyiidae dan Tephritidae (selain C.
connexa) yang berperan sebagai herbivor pada habitat C. odorata diduga memiliki
potensi untuk berkompetisi dengan lalat puru C. connexa. Walalupun belum
diketahui secara pasti spesies-spesies tersebut berinteraksi dengan C. odorata,
namun spesies tersebut dikhawatirkan berpotensi untuk menyebabkan
terbentuknya puru pada tumbuhan eksotik invasif C. odorata. Waterhouse (1994)

37
melaporkan bahwa di Amerika Selatan ada 8 spesies dari famili Cecidomyiidae
dan 2 spesies dari famili Tephritidae yang berpotensi sebagai agens hayati untuk
mengendalikan C. odorata.
Famili Dolicophodidae dan Phoridae memiliki kekayaan spesies dan
kelimpahan individu tertinggi (Tabel 10). Menurut Boror et al. (1981) Famili
Dolicophodidae merupakan kelompok predator yang jumlah individunya
berlimpah di hutan dan padang rumput. Melimpahnya kekayaan spesies dan
kelimpahan individu Famili Dolicophodidae diduga terkait dengan banyaknya
mangsa yang berada disekitar vegetasi pada keempat habitat C. odorata tersebut.
Famili Phoridae termasuk dalam kelompok serangga saprofag (Boror et al. 1981).
Adanya kotoran dan daun membusuk yang berasal dari vegetasi di sekitar habitat
C. odorata merupakan makanan bagi kelompok serangga saprofag, sehingga
menyebabkan tingginya kekayaan spesies dan kelimpahan individu serangga dari
famili tersebut. Serangga saprofag sangat berguna dalam proses jaring makanan
karena membantu menguraikan bahan organik yang ada.
Dari hasil perangkap yang digunakan berupa pitfall trap, yellow pan trap,
dan malaise trap tidak banyak imago lalat puru C. connexa yang dijumpai pada
habitat C. odorata. Keadaan ini dibuktikan pada habitat C. odorata di Parung
Panjang, Setu, dan Darmaga secara berturut-turut hanya ditemukan sebanyak 5, 3,
dan 2 individu lalat puru C. connexa, sedangkan di Gunung Bunder tidak dijumpai
sama sekali. Untuk mengukur populasi lalat puru C. connexa di lapangan tidak
dapat diukur dengan menggunakan metode berupa perangkap pitfaall trap, yellow
pan trap, dan malaise trap, tetapi dengan metode koleksi puru langsung dari
lapangan atau dengan menggunakan staner trap yang diberi metyl eugenol.
Hubungan Keanekaragaman Serangga dengan Kondisi Habitat
Kondisi masing-masing habitat C. odorata memiliki perbedaan (Tabel 7).
Habitat C. odorata di Darmaga (hutan buatan dengan tanaman utama A.
falcataria) memiliki kekayaan spesies serangga tertinggi yang terdiri dari 345
spesies diikuti oleh Setu dan Gunung Bunder masing-masing 300 spesies dan 277
spesies. Keadaan ini berbeda dengan Parung Panjang (perkebunan dengan

38
tanaman utama A. mangium) yang memiliki kekayaan spesies serangga terendah
yaitu 262 spesies (Tabel 8). Tingginya kekayaan spesies serangga pada habitat C.
odorata di Darmaga diduga karena selain adanya tanaman utama juga terdapat
komposisi vegetasi tumbuhan yang lebih beragam sehingga mempengaruhi
kekayaan spesies serangga di sekitarnya dibandingkan dengan habitat C. odorata
di Parung Panjang, Setu, dan Gunung Bunder yang memiliki komposisi vegetasi
tumbuhan yang cenderung lebih sedikit di banding Darmaga.
Kemiripan komposisi spesies serangga yang diperoleh antar lokasi dengan
menggunakan indeks kemiripan Sorensen (Cs) memperlihatkan bahwa antara
Parung Panjang dan Setu memiliki indeks kemiripan tertinggi yaitu 0.63 atau
sekitar 63% dari spesies serangga yang ditemukan adalah sama. Komposisi
spesies serangga yang ditemukan di Gunung Bunder dengan Parung Panjang
memiliki indeks terendah yaitu 0.49 yang mengindikasikan bahwa spesies
serangga yang ditemukan memiliki kemiripan sekitar 49% (Tabel 11).
Tabel 11 Indeks kemiripan Sorensen (Cs) seluruh spesies serangga antar lokasi penelitian pada habitat C. odorata
Lokasi penelitian Parung Panjang Setu Darmaga Gunung Bunder
Parung Panjang 1.00 Setu 0.63 1.00
Darmaga 0.54 0.55 1.00
Gunung Bunder 0.49 0.54 0.57 1.00
Habitat C. odorata di Parung Panjang merupakan perkebunan dengan
tanaman utama A. mangium, sedangkan Setu merupakan lahan terbuka dengan
dominasi M. malabathrichum. Dari tipe penggunaan lahan antara keduanya
memiliki perbedaaan, tetapi memiliki komposisi kemiripan spesies serangga yang
cukup tinggi. Diduga banyaknya kesamaan ini karena pengaruh faktor vegetasi
pada kedua habitat C. odorata tersebut memiliki kesamaan sehingga
mempengaruhi keberadaan serangga di sekitarnya. Selain itu, jarak yang dekat (6
km) dengan ketinggian yang sama (128 mdpl) juga merupakan faktor yang
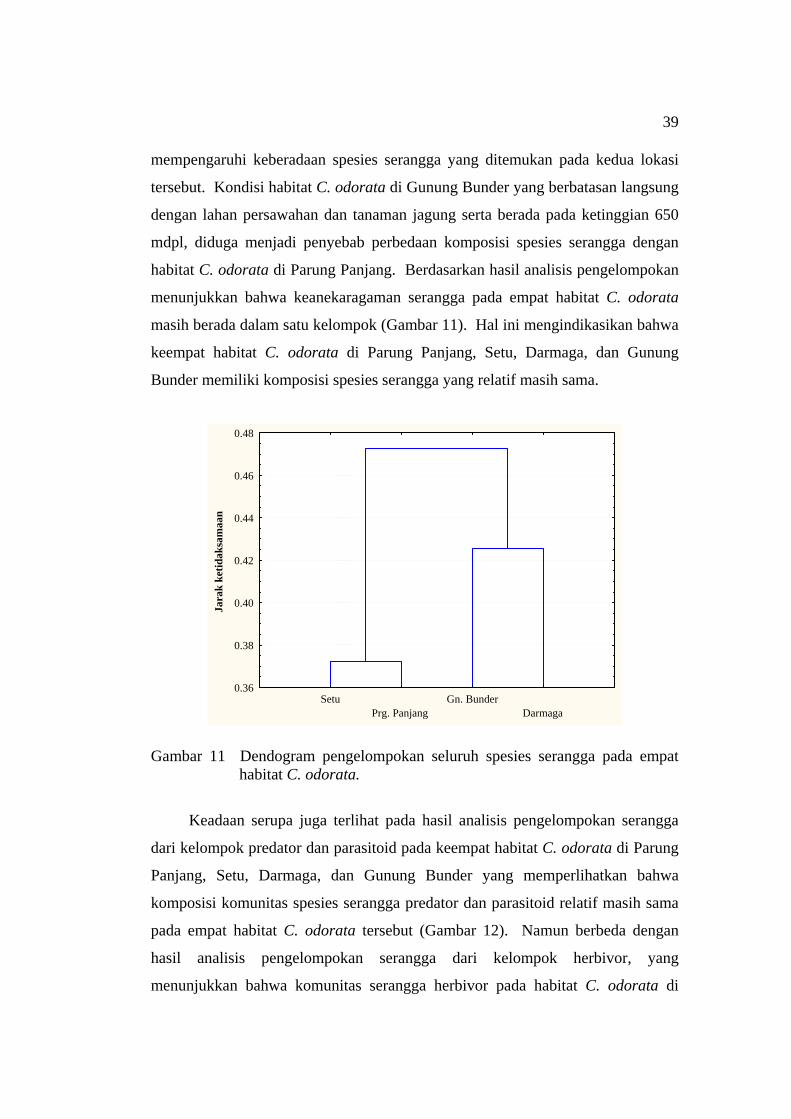
39
mempengaruhi keberadaan spesies serangga yang ditemukan pada kedua lokasi
tersebut. Kondisi habitat C. odorata di Gunung Bunder yang berbatasan langsung
dengan lahan persawahan dan tanaman jagung serta berada pada ketinggian 650
mdpl, diduga menjadi penyebab perbedaan komposisi spesies serangga dengan
habitat C. odorata di Parung Panjang. Berdasarkan hasil analisis pengelompokan
menunjukkan bahwa keanekaragaman serangga pada empat habitat C. odorata
masih berada dalam satu kelompok (Gambar 11). Hal ini mengindikasikan bahwa
keempat habitat C. odorata di Parung Panjang, Setu, Darmaga, dan Gunung
Bunder memiliki komposisi spesies serangga yang relatif masih sama.
SetuPrg. Panjang
Gn. BunderDarmaga
0.36
0.38
0.40
0.42
0.44
0.46
0.48
Jara
k ke
tidak
sam
aan
Gambar 11 Dendogram pengelompokan seluruh spesies serangga pada empat
habitat C. odorata.
Keadaan serupa juga terlihat pada hasil analisis pengelompokan serangga
dari kelompok predator dan parasitoid pada keempat habitat C. odorata di Parung
Panjang, Setu, Darmaga, dan Gunung Bunder yang memperlihatkan bahwa
komposisi komunitas spesies serangga predator dan parasitoid relatif masih sama
pada empat habitat C. odorata tersebut (Gambar 12). Namun berbeda dengan
hasil analisis pengelompokan serangga dari kelompok herbivor, yang
menunjukkan bahwa komunitas serangga herbivor pada habitat C. odorata di
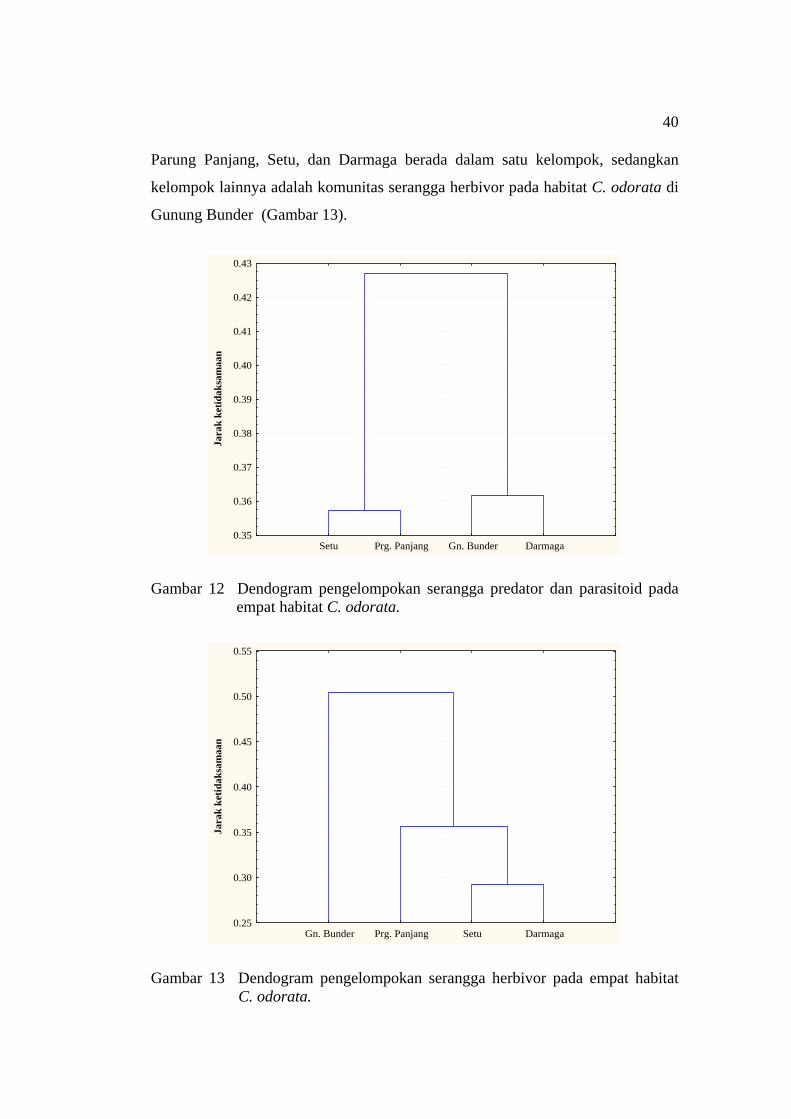
40
Parung Panjang, Setu, dan Darmaga berada dalam satu kelompok, sedangkan
kelompok lainnya adalah komunitas serangga herbivor pada habitat C. odorata di
Gunung Bunder (Gambar 13).
Setu Prg. Panjang Gn. Bunder Darmaga0.35
0.36
0.37
0.38
0.39
0.40
0.41
0.42
0.43
Jara
k ke
tidak
sam
aan
Gambar 12 Dendogram pengelompokan serangga predator dan parasitoid pada
empat habitat C. odorata.
Gn. Bunder Prg. Panjang Setu Darmaga0.25
0.30
0.35
0.40
0.45
0.50
0.55
Jara
k ke
tidak
sam
aan
Gambar 13 Dendogram pengelompokan serangga herbivor pada empat habitat C. odorata.

41
Asosiasi Serangga Lokal dengan Tumbuhan Eksotik Invasif C. odorata dan Agens Hayatinya C. connexa
Fenomena ekologi introduksi spesies tumbuhan eksotik invasif dapat
menyebabkan terjadinya asosiasi dalam struktur komunitas baru. Hasil
pengamatan memperlihatkan ada spesies serangga yang berasosiasi dengan
tumbuhan eksotik invasif C. odorata yaitu kutu aphid (Hemiptera: Aphididae).
Kutu aphid membentuk koloni pada bagian batang dan pucuk C. odorata yang
masih muda (Gambar 14). Klingauf (1987) melaporkan bahwa bagian pucuk
tanaman yang aktif tumbuh dan berkembang biasanya dipilih oleh kutu aphid
karena aktifitas pertumbuhan dan proses metabolismenya tinggi. Kutu aphid
menghisap jaringan floem pada lamina daun sehingga jaringan lamina daun mati
yang menyebabkan daun berkeriting. Keberadaan kutu aphid pada pucuk muda
C. odorata menyebabkan kehadiran koloni semut (Hymenoptera: Formicidae).
Asosiasi semut dengan kutu aphid karena adanya eksresi embun madu (honey)
kutu aphid yang dimanfaatkan oleh koloni semut. Ditemukan dua spesies semut
yang berasosiasi dengan kutu aphid yaitu Anoplolepis gracilipes dan
Crematogaster sp.01 (Gambar 14).
Gambar 14 Asosiasi serangga pada tumbuhan eksotik invasif C. odorata.
Keberadaan serangga herbivor eksotik juga akan berasosiasi dengan
serangga lokal di habitat baru. Asosiasi serangga lokal terhadap lalat puru C.
connexa yang terjadi diantaranya adalah musuh alami. Musuh alami ini terdiri
atas predator dan parasitoid dari ordo Hymenoptera, Coleoptera, Hemiptera, dan

42
Mantodea. Semut merupakan kelompok predator dari ordo Hymenoptera yang
banyak dijumpai pada bagian batang dan pucuk C. odorata. Terbentuknya koloni
semut yang banyak dijumpai disekitar batang dan pucuk muda C. odorata selain
berasosiasi dengan kutu aphid, semut juga memangsa kelompok telur yang
diletakkan oleh imago betina C. connexa pada bagian pucuk terminal dan lateral.
Selain itu semut mampu menembus puru yang berjendela dan memangsa larva
atau pupa yang berada di dalamnya. McFadyen et al. (2003) melaporkan bahwa
spesies semut Tetraponera sp. (Pseudomyrmecinae) mampu menembus puru yang
telah berjendela dan memangsa larva dan pupa didalamnya.
Kumbah kubah (Coleoptera: Coccinellidae) dijumpai pada pucuk-pucuk C.
odorata (Gambar 15). Sebagian besar famili kumbang kubah bersifat predator
yang memangsa hama pada fase telur sampai dewasa dan lainnya bertindak
sebagai hama tanaman (Boror et al. 1981). Karena sifatnya sebagai predator,
kumbang ini diduga selain memangsa kutu aphid juga memangsa telur dan larva
C. connexa instar awal yang baru menetas. Belalang sembah (Mantodea:
Mantidae) memangsa pupa C. connexa dalam puru yang berjendela. Perilaku
makan serangga ini menyebabkan puru menjadi terkoyak (Gambar 15). Predator
Sycanus sp. (Hemiptera: Reduviidae) memangsa larva instar akhir atau pupa yang
berada di dalam puru. Perilaku memangsanya dengan cara menusukkan stilet ke
dalam puru yang berjendela kemudian menghisap cairan larva dan pupa sehingga
meninggalkan eksuvianya saja.
Gambar 15 Serangga predator yang berasosiasi dengan lalat puru C. connexa.

43
Berdasarkan pengamatan secara visual kelompok parasitoid yang dijumpai
berasosiasi langsung dengan lalat puru C. connexa berasal dari ordo Hymenoptera
famili Braconidae, Eucoilidae, Eupelmidae, dan Ormyridae. Famili Braconidae
dilaporkan bersifat ektoparasit pada larva Lepidoptera dan Coleoptera, pada ordo
Diptera bersifat endoparasit, sedangkan famili Ormyridae merupakan parasit puru
pada lalat famili Cecidomyiidae dan Tephritidae (Ordo Diptera) (Gooulet &
Huber 1993). Famili Eucoilidae merupakan parasit pupa pada ordo Diptera
(Boror et al. 1981), sedangkan famili Eupelmidae bersifat parasit pada Ordo
Diptera (famili Cecidomyiidae) (Nauman 1991).
KESIMPULAN
Komunitas serangga yang diperoleh pada habitat C. odorata di Parung
Panjang, Setu, Darmaga, dan Gunung Bunder secara keseluruhan terdiri dari
24.213 individu serangga yang termasuk ke dalam 14 ordo, 132 famili, dan 568
spesies. Ordo Hymenoptera, Diptera, dan Hemiptera merupakan tiga ordo
terbesar yang ditemukan dengan kelimpahan individu (species abundance) dan
kekayaan spesies (species richness) paling tinggi. Kemiripan komposisi spesies
serangga pada empat habitat C. odorata di Parung Panjang, Setu, Darmaga, dan
Gunung Bunder masih relatif sama. Introduksi tumbuhan eksotik invasif C.
odorata dan agens hayatinya C. connexa telah menyebabkan terjadinya asosiasi
dengan serangga-serangga lokal.

PEMBAHASAN UMUM
Penelitian ini bertujuan untuk mempelajari distribusi dan kelimpahan
populasi C. odorata dan agens hayatinya C. connexa, serta mempelajari struktur
komunitas serangga dan tumbuhan di daerah yang telah diinvasi oleh C. odorata
dengan mengambil studi kasus di daerah Bogor, Jawa Barat. Diharapkan hasil
penelitian yang diperoleh dapat memberikan informasi mengenai distribusi dan
kelimpahan populasi C. odorata dan agens hayatinya C. connexa, memberikan
informasi struktur komunitas serangga dan tumbuhan di daerah yang telah
diinvasi oleh C. odorata, serta memberikan gambaran fenomena ekologi
introduksi spesies eksotik dapat menyebabkan terjadinya asosiasi antara spesies
eksotik tersebut dengan serangga lokal dalam struktur komunitas baru, sehingga
menjadi rekomendasi bagi pemerintah bahwa perlunya upaya perhatian dan
penyaringan yang ketat terhadap spesies eksotik yang didatangkan dari luar.
Hasil pengamatan pada semua lokasi penelitian di Jawa Barat khususnya
daerah Bogor, C. odorata cenderung menempati lahan yang terbuka seperti di sisi
jalan, tepian sawah yang kondisi tanahnya kering, ladang, dan perkebunan. Selain
itu, C. odorata juga dapat tumbuh di bawah tegakan hutan yang terbuka tajuknya,
bahkan masih bisa hidup pada daerah yang didominasi oleh batu-batuan. Pada
lahan yang dibudidayakan seperti sawah, ladang, dan hutan tanaman industri,
keberadaan C. odorata memiliki status sebagai gulma penting yang merugikan
karena populasinya sangat padat sehingga mampu berkompetisi dengan tanaman
budidaya dalam memperoleh unsur hara yang dibutuhkan (Tjitrosemito 1998).
Keberadaan C. odorata diikuti juga dengan keberadaan musuh alaminya
yaitu serangga herbivor berupa lalat pembentuk puru C. connexa pada semua
lokasi pengamatan, kecuali di Gunung Halimun dan Cianjur. Setelah dilepas di
Parung Panjang-Jasinga, Bogor pada tahun 1995 dan di Parung Kuda-Sukabumi
pada tahun 1996, lalat puru C. connexa mampu menyebar secara alami dan saat
ini telah mapan keberadaannya. Lalat puru C. connexa mampu hidup pada
berbagai ketinggian tempat yang berbeda, namun kelimpahan populasinya
menurun seiring dengan bertambahnya ketinggian suatu tempat. Distribusi lalat

45
puru C. connexa ternyata dibatasi oleh adanya barier seperti pegunungan. Hal ini
dibuktikan dengan tidak dijumpainya lalat puru tersebut di daerah Gunung
Halimun bagian selatan dan Cianjur. Keberadaan lalat puru C. connexa pada
suatu lokasi pengamatan berasal dari titik pelepasan yang terdekat. Sebagai
contoh, keberadaan lalat puru C. connexa di daerah Ciawi, Cisarua, dan Gunung
Salak berasal dari titik pelepasan di Parung Kuda-Sukabumi. Begitu juga
keberadaan lalat puru C. connexa ini di daerah Setu, Darmaga, dan Gunung
Bunder berasal dari titik pelepasan di Parung Panjang-Jasinga, Bogor. Namun
tidak menutup kemungkinan keberadaan lalat puru C. connexa di daerah Darmaga
dan Gunung Bunder juga bisa berasal dari titik pelepasan di Pakuwon-Parung
Kuda, Sukabumi.
Implikasi dari keberadaan spesies tumbuhan eksotik invasif C. odorata dan
agens hayatinya C. connexa terhadap struktur komunitas serangga dan tumbuhan
lokal telah dipelajari secara spesifik. Keberadaan C. odorata dan agens hayatinya
C. connexa berpengaruh terhadap keberadaan serangga dan tumbuhan lokal yang
berada di sekitarnya. Kehadiran tumbuhan eksotik invasif C. odorata
berhubungan erat dengan keseimbangan komunitas dalam ekosistem tersebut.
Implikasi dari keberadaan C. odorata terhadap struktur komunitas tumbuhan lokal
telah menyebabkan terjadinya pengambilalihan atau pergantian skala ruang, yaitu
tempat atau ruang yang seharusnya ditempati oleh spesies-spesies tumbuhan lokal
kemudian diambil alih keberadaannya oleh tumbuhan eksotik invasif C. odorata.
Selain itu, kehadiran tumbuhan eksotik invasif C. odorata pada empat lokasi
penelitian menyebabkan terjadinya penurunan keanekaragaman spesies tumbuhan
yang berada disekitarnya.
Implikasi dari keberadaan C. odorata terhadap serangga-serangga lokal
yang berada disekitarnya tidak dipelajari secara spesifik, tetapi berdasarkan hasil
observasi di lapangan memperlihatkan bahwa telah terjadi asosiasi serangga lokal
terhadap spesies tumbuhan eksotik invasif C. odorata. Seperti asosiasi kutu aphid
(Hemiptera: Aphididae) dengan C. odorata yang membentuk koloni pada bagian
batang dan pucuk C. odorata yang masih muda sehingga menyebabkan gejala
keriting pada pucuk muda C. odorata tersebut.

46
Keberadaan serangga herbivor eksotik berupa lalat puru C. connexa juga
akan berasosiasi dengan serangga lokal atau bahkan dengan organisme lain di
habitat baru. Asosiasi serangga lokal terhadap lalat puru C. connexa yang terjadi
diantaranya adalah musuh alami. Musuh alami ini terdiri atas predator dan
parasitoid dari ordo Hymenoptera, Coleoptera, Hemiptera, dan Mantodea. Musuh
alami ini cenderung mempengaruhi populasi serangga herbivor lalat puru C.
connexa, dan pada akhirnya akan berdampak terhadap populasi tumbuhan eksotik
invasif C. odorata yang merupakan inang dari lalat puru tersebut.

KESIMPULAN DAN SARAN
Kesimpulan
Kesimpulan yang bisa ditarik dari penelitian ini adalah :
1. Persebaran spesies tumbuhan eksotik invasif C. odorata di Jawa Barat
khususnya daerah Bogor sudah sangat luas. C. odorata dapat tumbuh pada
berbagai tipe habitat meliputi habitat perkebunan, hutan buatan, ladang
dataran rendah, maupun ladang dataran tinggi dengan kelimpahan yang
berbeda. Kelimpahan populasinya cenderung menurun seiring dengan
bertambahnya ketinggian suatu lokasi.
2. Keberadaan C. odorata diikuti pula dengan keberadaan musuh alaminya yaitu
lalat puru C. connexa pada semua lokasi pengamatan, kecuali di Gunung
Halimun dan Cianjur. Ada dua titik pelepasan lalat puru C. connexa yaitu di
Parung Panjang-Jasinga, Bogor dan Pakuwon-Parung Kuda, Sukabumi. Lalat
puru C. connexa saat ini telah mapan keberadaannya dan menyebar secara
alami serta memiliki kemampuan persebaran yang cukup jauh baik dari titik
pelepasannya di Parung Panjang-Jasinga, Bogor maupun dari titik pelepasan
di Pakuwon-Parung Kuda, Sukabumi.
3. Lalat puru C. connexa mampu hidup pada berbagai ketinggian tempat yang
berbeda, namun kelimpahan populasinya menurun seiring dengan
bertambahnya ketinggian suatu tempat. Distribusi lalat puru C. connexa
dibatasi oleh adanya barier berupa pegunungan. Kelimpahan populasinya
sangat dipengaruhi oleh ketersediaan inangnya yaitu C. odorata.
4. Komunitas serangga yang diperoleh pada habitat C. odorata di Parung
Panjang, Setu, Darmaga, dan Gunung Bunder secara keseluruhan terdiri dari
24.213 individu serangga yang termasuk ke dalam 14 ordo, 132 famili, dan
568 spesies. Ordo Hymenoptera, Diptera, dan Hemiptera merupakan tiga
ordo terbesar yang ditemukan dengan kelimpahan individu (species
abundance) dan kekayaan spesies (species richness) paling tinggi.
5. Introduksi tumbuhan eksotik invasif C. odorata dan agens hayatinya C.
connexa telah menyebabkan terjadinya asosiasi dengan serangga-serangga
lokal.

48
6. Komunitas tumbuhan yang diperoleh pada habitat C. odorata di Parung
Panjang, Setu, Darmaga, dan Gunung Bunder secara keseluruhan terdiri dari
131.132 individu tumbuhan yang termasuk ke dalam 21 famili dan 44 spesies.
7. Kelimpahan populasi C. odorata pada lokasi penelitian di Parung Panjang,
Setu, dan Darmaga mendominasi berdasarkan perhitungan Indeks Nilai
Penting (INP) kecuali di Gunung Bunder. Implikasi dari keberadaan spesies
tumbuhan eksotik invasif C. odorata pada tiga habitat tersebut telah
menyebabkan terjadinya pengambilalihan atau pergantian skala ruang, yaitu
tempat atau ruang yang seharusnya ditempati oleh spesies-spesies tumbuhan
lokal kemudian diambil alih keberadaannya oleh tumbuhan eksotik invasif C.
odorata. Selain itu, kehadiran tumbuhan eksotik invasif C. odorata pada
empat lokasi penelitian menyebabkan terjadinya penurunan keanekaragaman
spesies tumbuhan yang berada disekitarnya.
Saran
Hasil penelitian ini dapat dijadikan rujukan untuk penelitian selanjutnya.
Diantara hal-hal yang dapat disarankan adalah :
1. Ditemukannya famili Tephritidae dan Cecidomyiidae yang menarik untuk
dipelajari lebih lanjut, mengingat bahwa spesies-spesies dari famili
Tephritidae dan Cecidomyiidae sebagian besar anggotanya juga bisa
menyebabkan terbentuknya puru pada tanaman.
2. Asosiasi antara lalat puru C. connexa dengan parasitoid dari ordo
Hymenoptera famili Braconidae, Eucoilidae, Eupelmidae, dan Ormyridae
dapat dipelajari lebih mendalam pada penelitian selanjutnya seperti uji
biologi parasitoid tersebut serta identifikasi sampai pada tingkat spesies.

DAFTAR PUSTAKA
Altieri MA, Nicholl CI. 2004. Biodiversity and Pest Management in
Agroecosystem. Ed ke-2. New York: Food Product Press. [Anonim]. 1991. Chromolaena odorata (L.) R.M. King & H. Robinson. Weed
Info Sheet 5:1-4. Binggeli P. 1997. Chromolaena odorata (L.) King & Robinson (Asteraceae).
http://members.lycos.co.uk/WoodyPlantEcology/docs/web-sp4.htm. [13 Oktober 2003].
Borror DJ, De Long DM, Triplehorn CA. 1981 An Introduction to the Study of
Insect. New York: Saunders College Publishing. Chenon RD, Sipayung A, Sudharto P. 2002. A decade of Biological control
against Chromolaena odorata at the Indonesian Oil Palm Research Institute in Marihat. Di dalam: Zachariades C, Muniapan R, Strathie LW, editor. Proceedings of the Fifth International Workshop on Biological Control and Management of Chromolaena odorata, Durban, South Africa, 23-25 October 2000. ARC-PPRI. Hlm 46-52.
Colwell RK, Coddington JA. 1994. Estimating terrestrial biodiversity through
extrapolation. Philosophical Transactions: Biol Sci 345:101-118. Colwell RK. 2000. EstimateS: Statistical estimation of species richness and
shared species from samples. Version 6.0d1. [Serial online]. http://www.viceroy.eeb.uconn.edu/estimates. [16 Desember2003].
Cox GW. 2002. Laboratory Manual of General Ecology. Ed ke-8. USA. The
McGraw-Hill Companies. Erasmus DJ, Bennet PH, Van Standen J. 1992. The effect of galls induced by the
gall fly Procecidochares utilis on vegetative growth and reproductive potential of crofton weed, Ageratina adenophora. Annal of Appl Biol 120: 173-181.
Goulet H, Huber JT, Editor. 1993. Hymenoptera of the World: An Identification
Guide to Families. Canada: Canada Communication Group. Kartosuwondo U. 2001. Peranan tumbuhan bukan budi daya dalam pengendalian
hayati serangga hama. Hayati 8: 55-87.
Kasno, Putri ASR, Widayanti S, Sunjaya. 2001. Establishment of Neochetina sp.: Their pattern of local dispersal and age structure at release site. Biotropia 13:18-29.

50
Klingauf FA. 1987. Feeding, adaptation and excretion. Didalam: Minus AK, Harrewijn P, editor. Aphid: Their Biology, Natural Enemies and Control. Amsterdam: Elsevier. Hlm 225-253.
Kostermans AJGH, Wirdjahardja S, Dekker RJ. 1987. The weed: description,
ecology and control. Di dalam: Soerjani M, Kostermans AJGH, Tjitrosoepomo G, editor. Weed of rice in Indonesia. Jakarta: Balai Pustaka. hlm 24-566.
Krebs CJ. 1998. Ecological Methodology. New York. Harper Collins. Laumonier EKW, Megia R, Veenstra H. 1987. The Seedling. Di dalam: Soerjani
M, Kostermans AJGH, Tjitrosoepomo G, editor. Weed of Rice in Indonesia. Jakarta: Balai Pustaka. hlm 567-686.
Magguran AE. 1988. Ecological Diversity and Its Measurement. London. Chapman & Hall.
McFadyen REC, Chenon RD, Sipayung A. 2003. Biology and host Specificity
of the Chromolaena stem gall fly, Cecidochares connexa (Macquart) (Diptera: Tephritidae). Aust J of Entomol 42:294-297.
Mooney HA, Cleland EE. 2001. The Evolutionary impact of invasive species.
Proc Nat and Acad Sci 98(10):5446-5451. Nauman ID. 1991. The Insect of Australia: Textbook for Student and Research
Workers 2th eds. Melbourne: Melbourne University Press. Olden JD, Poff NL, Douglas MR, Douglas ME, Faucsh KD. 2004. Ecological
and evolutionary consequences of biotic homogenization. Trends in Ecol and Evol 19(1):18-24.
Pearson DE, Callaway RM. 2003. Indirect effects of host-specific biological
control agents. Trends in Ecol and Evol 18(9):456-461. Schoonhoven LM, Jermy T, van Loon JJA. 1996. Insect-Plant Biology: From
Physiology to Evolution. London. Chapman & Hall. Simberloff D. 1996. How risky of biological control?. Ecol 77(7):1965-1974. Sipayung A, Chenon RD. 1995. Procecidochares connexa untuk pengendalian
gulma Chromolaena odorata [makalah]. Di dalam: Prosiding Seminar Pelepasan Procecidochares connexa; Medan, 17 Juli 1995. Medan: Pusat Penelitian Kelapa Sawit. hlm 1-14.
StatSoft 1995. Statistica for Windows Release 5.0. Tulsa: StatSoft.

51
Tjitrosemito S. 1997. Pengendalian Chromolaena odorata di lahan petani [makalah]. Di dalam: Seminar Nasional Pengendalian Chromolaena odorata; Kupang, 14-15 April 1997. Bogor: Biotrop. hlm 1-18.
Tjitrosemito S. 1998. Integrated management of Chromolaena odorata:
emphasizing the classical biological control. Biotropia 11:9-21. Tjitrosemito S. 1999. The establishment of Procecidochares connexa in West
Java, Indonesia: a biological control agent of Chromolaena odorata. Biotropia 12:19-24.
Tjitrosemito S. 1999b. Dinamika populasi Procecidochares connexa (Diptera:
Tephritidae) di Jawa Barat dan Jawa Timur [makalah]. Di dalam: Seminar Nasional Pengendalian Hayati; Yogyakarta, 12-13 Juli 1999. Bogor: Biotrop. hlm 1-7.
Tjitrosemito S. 2000. Procecidochares connexa Macquart (Diptera: Tephritidae):
A biological control agent of siam weed (Chromolaena odorata (L.) R. King & Robinson) [makalah]. Di dalam: International Warkshop on Biological Control and Management of Chromolaena odorata; Durban, South Africa, 23-25 Oktober 2000. Bogor: Biotrop. hlm 1-16.
Untung S. 2005. Pencegahan dan pengendalian spesies asing di Indonesia.
http://kasumbogo.staff.ugm.ac.id/detailarticle.php?mesid=38&kata_kunci= spesies%20asing,%20alien%20Invasive%20species. [4 Maret 2006].
Waterhouse DF. 1994. Biological Control of Weeds: Southeast Asian Prospects.
Canberra: Australia. Wittenberg R, Cock MJW. 2003. Invasive Alien Spesies: A Toolkit of Best
Prevention and Management Practices. CAB International. Widayanti S, Tjitrosemito S, Kasno. 1999. Biologi dan neraca kehidupan lalat
Argentina Procecidochares connexa (Diptera: Tephritidae) agens pengendali hayati kirinyuh (Chromolaena odorata) [makalah]. Di dalam: Prosiding Seminar Nasional Pengendalian Hayati; Yogyakarta, 12-13 Juli 1999. hlm 1-10
Widayanti S, Tjitrosemito S, Muhammad A. 2001. Pengendalian hayati
Chromolaena odorata dengan menggunakan lalat puru Procecidochares connexa di Cagar Alam Pangandaran, Jawa Barat [makalah]. Di dalam: Konferensi Nasional HIGI XV; Universitas Sebelas Maret, Surakarta, 17-19 Juli 2001. Bogor; Biotrop. hlm 1-7.

L A M P I R A N
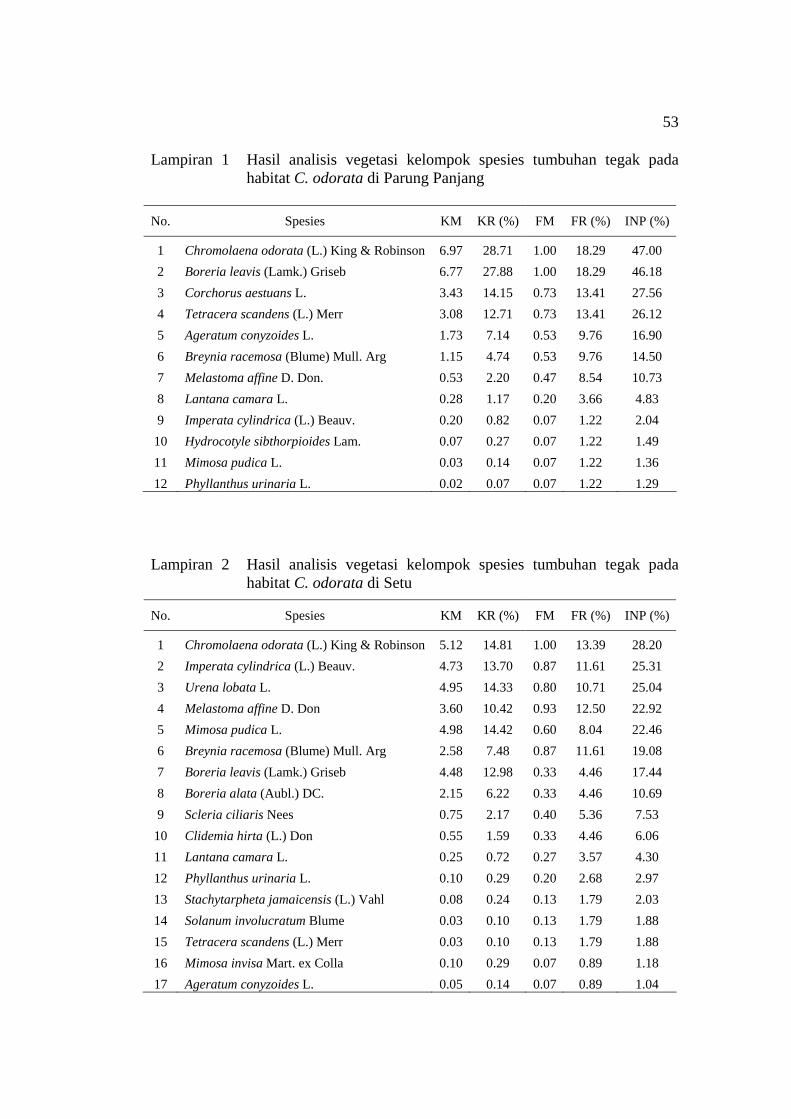
53
Lampiran 1 Hasil analisis vegetasi kelompok spesies tumbuhan tegak pada habitat C. odorata di Parung Panjang
No. Spesies KM KR (%) FM FR (%) INP (%)
1 Chromolaena odorata (L.) King & Robinson 6.97 28.71 1.00 18.29 47.00 2 Boreria leavis (Lamk.) Griseb 6.77 27.88 1.00 18.29 46.18 3 Corchorus aestuans L. 3.43 14.15 0.73 13.41 27.56 4 Tetracera scandens (L.) Merr 3.08 12.71 0.73 13.41 26.12 5 Ageratum conyzoides L. 1.73 7.14 0.53 9.76 16.90 6 Breynia racemosa (Blume) Mull. Arg 1.15 4.74 0.53 9.76 14.50 7 Melastoma affine D. Don. 0.53 2.20 0.47 8.54 10.73 8 Lantana camara L. 0.28 1.17 0.20 3.66 4.83 9 Imperata cylindrica (L.) Beauv. 0.20 0.82 0.07 1.22 2.04
10 Hydrocotyle sibthorpioides Lam. 0.07 0.27 0.07 1.22 1.49 11 Mimosa pudica L. 0.03 0.14 0.07 1.22 1.36 12 Phyllanthus urinaria L. 0.02 0.07 0.07 1.22 1.29
Lampiran 2 Hasil analisis vegetasi kelompok spesies tumbuhan tegak pada habitat C. odorata di Setu
No. Spesies KM KR (%) FM FR (%) INP (%)
1 Chromolaena odorata (L.) King & Robinson 5.12 14.81 1.00 13.39 28.20 2 Imperata cylindrica (L.) Beauv. 4.73 13.70 0.87 11.61 25.31 3 Urena lobata L. 4.95 14.33 0.80 10.71 25.04 4 Melastoma affine D. Don 3.60 10.42 0.93 12.50 22.92 5 Mimosa pudica L. 4.98 14.42 0.60 8.04 22.46 6 Breynia racemosa (Blume) Mull. Arg 2.58 7.48 0.87 11.61 19.08 7 Boreria leavis (Lamk.) Griseb 4.48 12.98 0.33 4.46 17.44 8 Boreria alata (Aubl.) DC. 2.15 6.22 0.33 4.46 10.69 9 Scleria ciliaris Nees 0.75 2.17 0.40 5.36 7.53
10 Clidemia hirta (L.) Don 0.55 1.59 0.33 4.46 6.06 11 Lantana camara L. 0.25 0.72 0.27 3.57 4.30 12 Phyllanthus urinaria L. 0.10 0.29 0.20 2.68 2.97 13 Stachytarpheta jamaicensis (L.) Vahl 0.08 0.24 0.13 1.79 2.03 14 Solanum involucratum Blume 0.03 0.10 0.13 1.79 1.88 15 Tetracera scandens (L.) Merr 0.03 0.10 0.13 1.79 1.88 16 Mimosa invisa Mart. ex Colla 0.10 0.29 0.07 0.89 1.18 17 Ageratum conyzoides L. 0.05 0.14 0.07 0.89 1.04
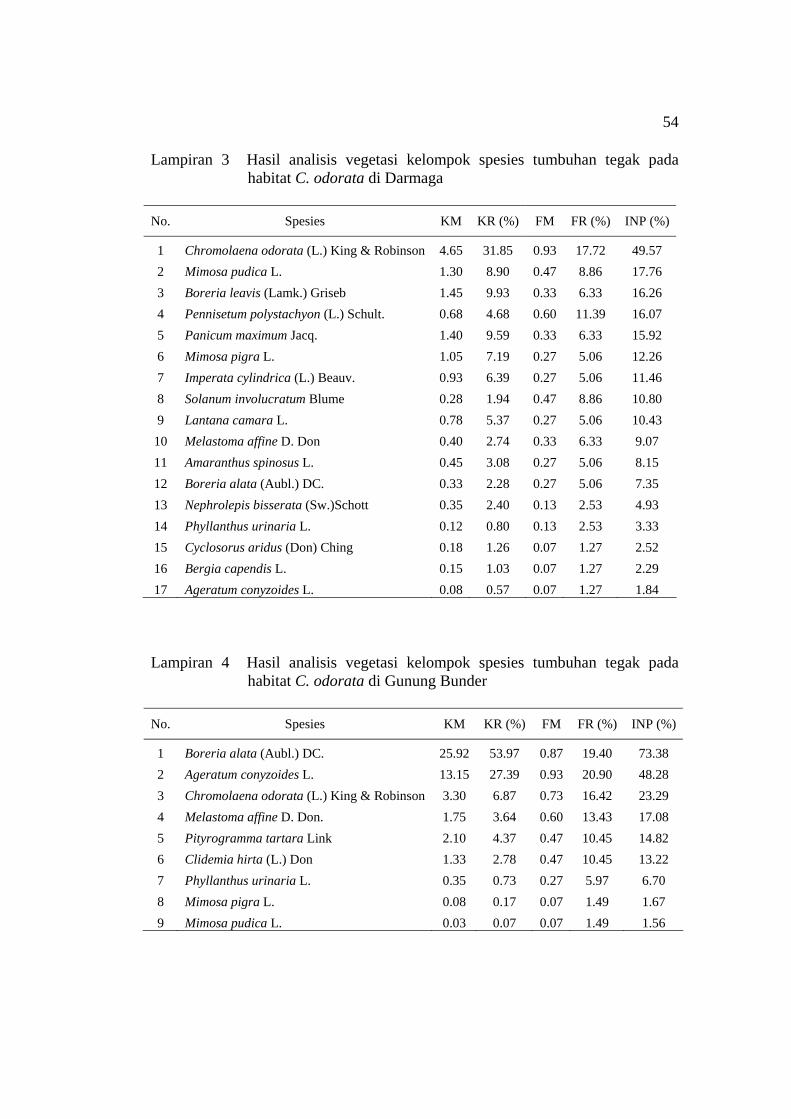
54
Lampiran 3 Hasil analisis vegetasi kelompok spesies tumbuhan tegak pada habitat C. odorata di Darmaga
No. Spesies KM KR (%) FM FR (%) INP (%)
1 Chromolaena odorata (L.) King & Robinson 4.65 31.85 0.93 17.72 49.57 2 Mimosa pudica L. 1.30 8.90 0.47 8.86 17.76 3 Boreria leavis (Lamk.) Griseb 1.45 9.93 0.33 6.33 16.26 4 Pennisetum polystachyon (L.) Schult. 0.68 4.68 0.60 11.39 16.07 5 Panicum maximum Jacq. 1.40 9.59 0.33 6.33 15.92 6 Mimosa pigra L. 1.05 7.19 0.27 5.06 12.26 7 Imperata cylindrica (L.) Beauv. 0.93 6.39 0.27 5.06 11.46 8 Solanum involucratum Blume 0.28 1.94 0.47 8.86 10.80 9 Lantana camara L. 0.78 5.37 0.27 5.06 10.43
10 Melastoma affine D. Don 0.40 2.74 0.33 6.33 9.07 11 Amaranthus spinosus L. 0.45 3.08 0.27 5.06 8.15 12 Boreria alata (Aubl.) DC. 0.33 2.28 0.27 5.06 7.35 13 Nephrolepis bisserata (Sw.)Schott 0.35 2.40 0.13 2.53 4.93 14 Phyllanthus urinaria L. 0.12 0.80 0.13 2.53 3.33 15 Cyclosorus aridus (Don) Ching 0.18 1.26 0.07 1.27 2.52 16 Bergia capendis L. 0.15 1.03 0.07 1.27 2.29 17 Ageratum conyzoides L. 0.08 0.57 0.07 1.27 1.84 Lampiran 4 Hasil analisis vegetasi kelompok spesies tumbuhan tegak pada
habitat C. odorata di Gunung Bunder No. Spesies KM KR (%) FM FR (%) INP (%)
1 Boreria alata (Aubl.) DC. 25.92 53.97 0.87 19.40 73.38 2 Ageratum conyzoides L. 13.15 27.39 0.93 20.90 48.28 3 Chromolaena odorata (L.) King & Robinson 3.30 6.87 0.73 16.42 23.29 4 Melastoma affine D. Don. 1.75 3.64 0.60 13.43 17.08 5 Pityrogramma tartara Link 2.10 4.37 0.47 10.45 14.82 6 Clidemia hirta (L.) Don 1.33 2.78 0.47 10.45 13.22 7 Phyllanthus urinaria L. 0.35 0.73 0.27 5.97 6.70 8 Mimosa pigra L. 0.08 0.17 0.07 1.49 1.67 9 Mimosa pudica L. 0.03 0.07 0.07 1.49 1.56
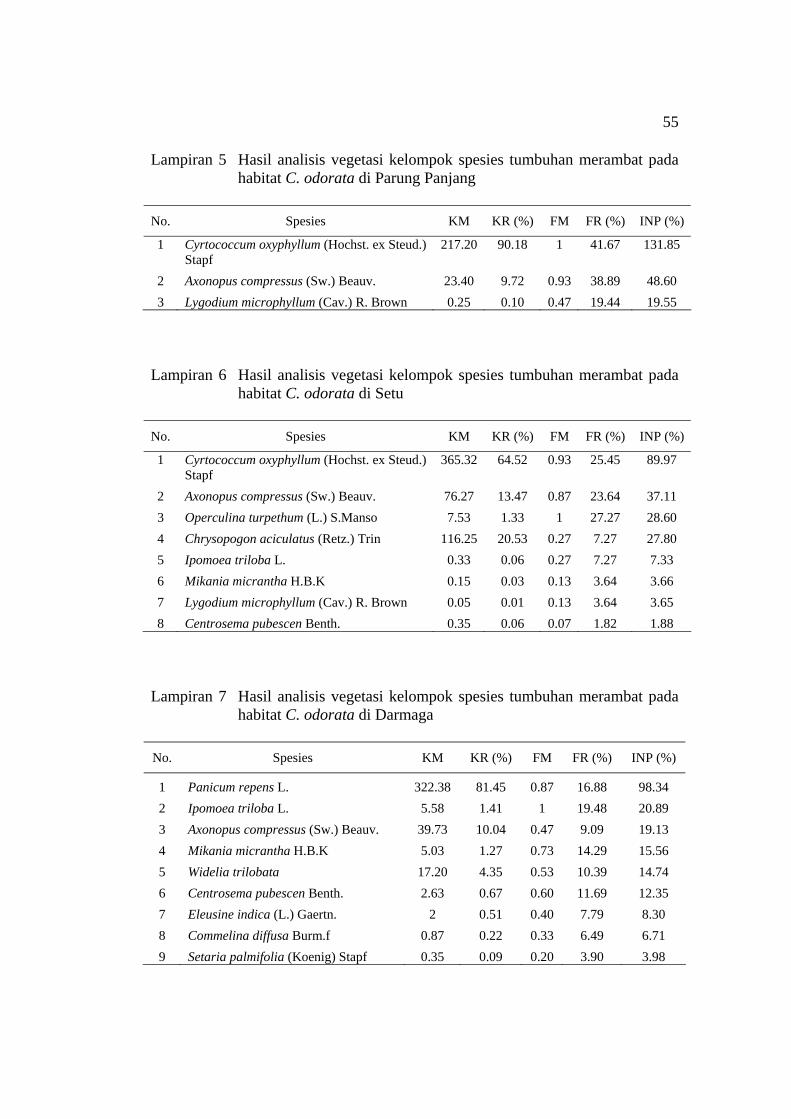
55
Lampiran 5 Hasil analisis vegetasi kelompok spesies tumbuhan merambat pada habitat C. odorata di Parung Panjang
No. Spesies KM KR (%) FM FR (%) INP (%)
1 Cyrtococcum oxyphyllum (Hochst. ex Steud.) Stapf
217.20 90.18 1 41.67 131.85
2 Axonopus compressus (Sw.) Beauv. 23.40 9.72 0.93 38.89 48.60 3 Lygodium microphyllum (Cav.) R. Brown 0.25 0.10 0.47 19.44 19.55
Lampiran 6 Hasil analisis vegetasi kelompok spesies tumbuhan merambat pada
habitat C. odorata di Setu No. Spesies KM KR (%) FM FR (%) INP (%)
1 Cyrtococcum oxyphyllum (Hochst. ex Steud.) Stapf
365.32 64.52 0.93 25.45 89.97
2 Axonopus compressus (Sw.) Beauv. 76.27 13.47 0.87 23.64 37.11 3 Operculina turpethum (L.) S.Manso 7.53 1.33 1 27.27 28.60 4 Chrysopogon aciculatus (Retz.) Trin 116.25 20.53 0.27 7.27 27.80 5 Ipomoea triloba L. 0.33 0.06 0.27 7.27 7.33 6 Mikania micrantha H.B.K 0.15 0.03 0.13 3.64 3.66 7 Lygodium microphyllum (Cav.) R. Brown 0.05 0.01 0.13 3.64 3.65 8 Centrosema pubescen Benth. 0.35 0.06 0.07 1.82 1.88
Lampiran 7 Hasil analisis vegetasi kelompok spesies tumbuhan merambat pada
habitat C. odorata di Darmaga No. Spesies KM KR (%) FM FR (%) INP (%)
1 Panicum repens L. 322.38 81.45 0.87 16.88 98.34 2 Ipomoea triloba L. 5.58 1.41 1 19.48 20.89 3 Axonopus compressus (Sw.) Beauv. 39.73 10.04 0.47 9.09 19.13 4 Mikania micrantha H.B.K 5.03 1.27 0.73 14.29 15.56 5 Widelia trilobata 17.20 4.35 0.53 10.39 14.74 6 Centrosema pubescen Benth. 2.63 0.67 0.60 11.69 12.35 7 Eleusine indica (L.) Gaertn. 2 0.51 0.40 7.79 8.30 8 Commelina diffusa Burm.f 0.87 0.22 0.33 6.49 6.71 9 Setaria palmifolia (Koenig) Stapf 0.35 0.09 0.20 3.90 3.98

56
Lampiran 8 Hasil analisis vegetasi kelompok spesies tumbuhan merambat pada habitat C. odorata di Gunung Bunder
No. Spesies KM KR (%) FM FR (%) INP (%)
1 Axonopus compressus (Sw.) Beauv. 490.90 57.00 0.87 15.85 72.85 2 Ischaemum timorense Kunth. 323.12 37.52 0.73 13.41 50.93 3 Rostellularia sundana Bremek 21.62 2.51 0.87 15.85 18.36 4 Cyperus kyllingia Endl. 10.97 1.27 0.93 17.07 18.35 5 Eragrostis unioloides (Retz.) Nees ex Steud. 5.67 0.66 0.60 10.98 11.63 6 Mikania micrantha H.B.K 2.62 0.30 0.40 7.32 7.62 7 Commelina diffusa Burm.f 1 0.12 0.40 7.32 7.43 8 Setaria palmifolia (Koenig) Stapf 4.47 0.52 0.27 4.88 5.40 9 Widelia trilobata 0.62 0.07 0.13 2.44 2.51
10 Lygodium microphyllum (Cav.) R. Brown 0.13 0.02 0.13 2.44 2.45 11 Eleusine indica (L.) Gaertn. 0.12 0.01 0.13 2.44 2.45
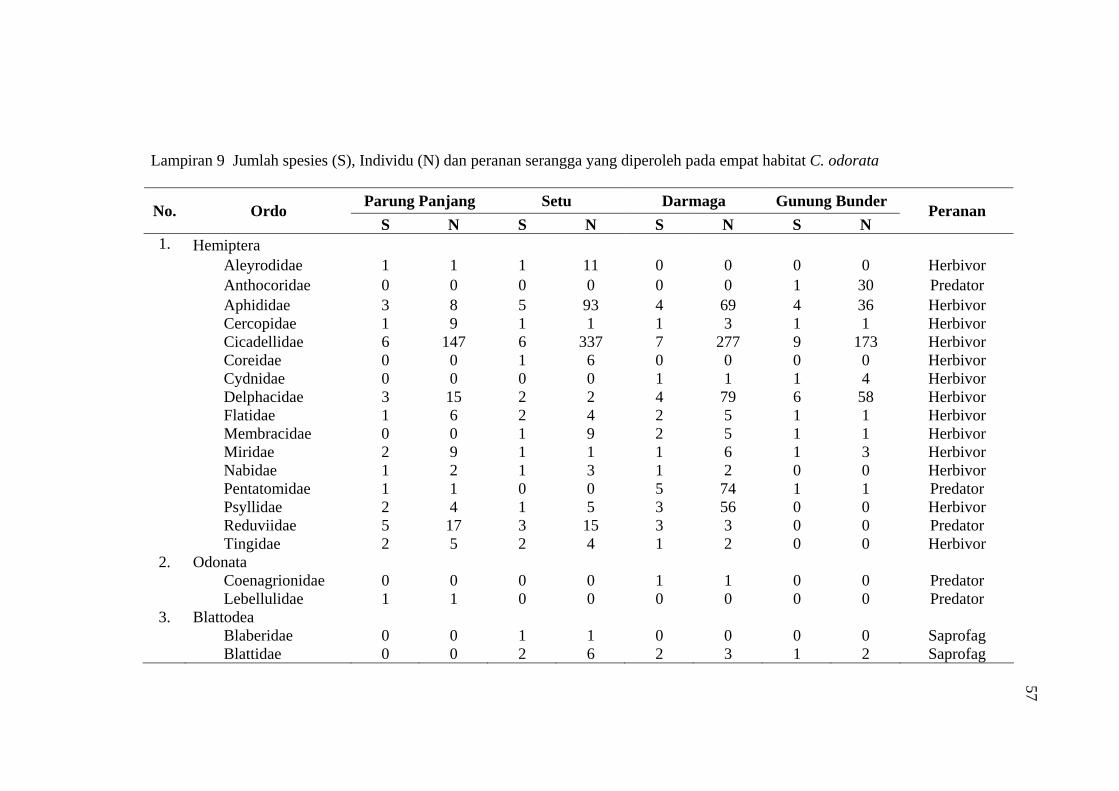
Lampiran 9 Jumlah spesies (S), Individu (N) dan peranan serangga yang diperoleh pada empat habitat C. odorata
Parung Panjang Setu Darmaga Gunung Bunder No. Ordo S N S N S N S N
Peranan
1. Hemiptera Aleyrodidae 1 1 1 11 0 0 0 0 Herbivor Anthocoridae 0 0 0 0 0 0 1 30 Predator Aphididae 3 8 5 93 4 69 4 36 Herbivor Cercopidae 1 9 1 1 1 3 1 1 Herbivor Cicadellidae 6 147 6 337 7 277 9 173 Herbivor Coreidae 0 0 1 6 0 0 0 0 Herbivor Cydnidae 0 0 0 0 1 1 1 4 Herbivor Delphacidae 3 15 2 2 4 79 6 58 Herbivor Flatidae 1 6 2 4 2 5 1 1 Herbivor Membracidae 0 0 1 9 2 5 1 1 Herbivor Miridae 2 9 1 1 1 6 1 3 Herbivor Nabidae 1 2 1 3 1 2 0 0 Herbivor Pentatomidae 1 1 0 0 5 74 1 1 Predator Psyllidae 2 4 1 5 3 56 0 0 Herbivor Reduviidae 5 17 3 15 3 3 0 0 Predator Tingidae 2 5 2 4 1 2 0 0 Herbivor
2. Odonata Coenagrionidae 0 0 0 0 1 1 0 0 Predator Lebellulidae 1 1 0 0 0 0 0 0 Predator
3. Blattodea Blaberidae 0 0 1 1 0 0 0 0 Saprofag Blattidae 0 0 2 6 2 3 1 2 Saprofag
57
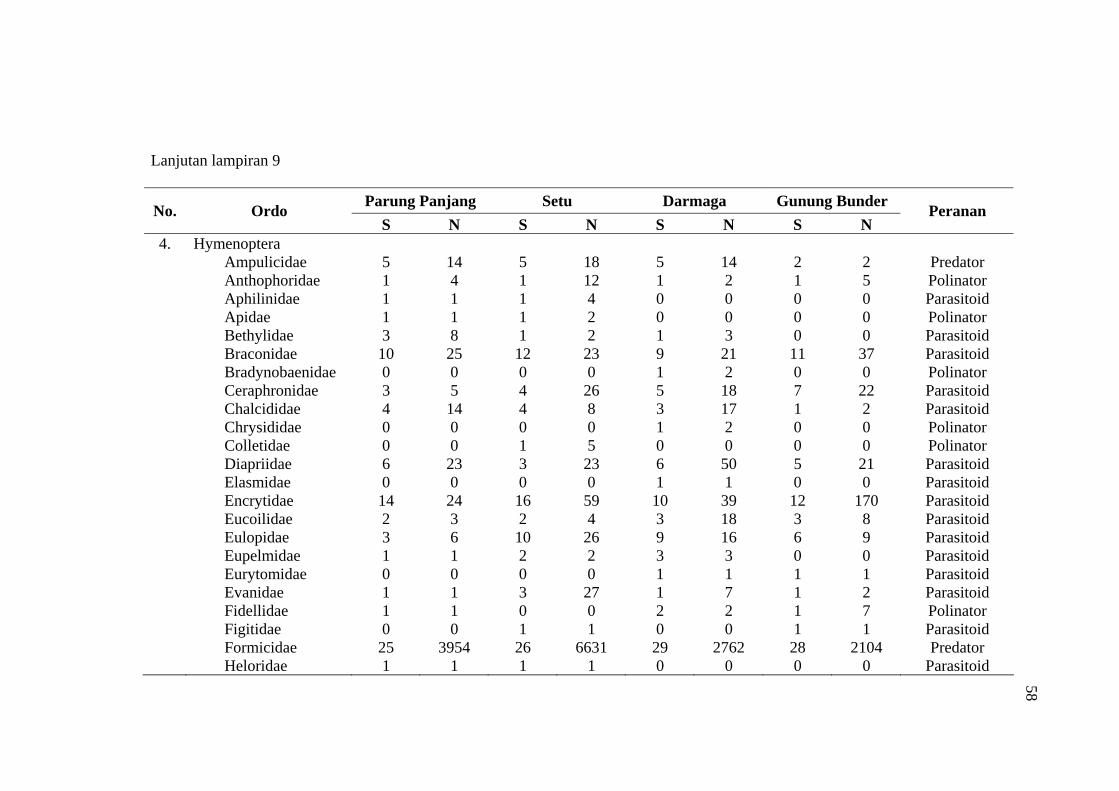
Lanjutan lampiran 9
Parung Panjang Setu Darmaga Gunung Bunder No. Ordo S N S N S N S N
Peranan
4. Hymenoptera Ampulicidae 5 14 5 18 5 14 2 2 Predator Anthophoridae 1 4 1 12 1 2 1 5 Polinator Aphilinidae 1 1 1 4 0 0 0 0 Parasitoid Apidae 1 1 1 2 0 0 0 0 Polinator Bethylidae 3 8 1 2 1 3 0 0 Parasitoid Braconidae 10 25 12 23 9 21 11 37 Parasitoid Bradynobaenidae 0 0 0 0 1 2 0 0 Polinator Ceraphronidae 3 5 4 26 5 18 7 22 Parasitoid Chalcididae 4 14 4 8 3 17 1 2 Parasitoid Chrysididae 0 0 0 0 1 2 0 0 Polinator Colletidae 0 0 1 5 0 0 0 0 Polinator Diapriidae 6 23 3 23 6 50 5 21 Parasitoid Elasmidae 0 0 0 0 1 1 0 0 Parasitoid Encrytidae 14 24 16 59 10 39 12 170 Parasitoid Eucoilidae 2 3 2 4 3 18 3 8 Parasitoid Eulopidae 3 6 10 26 9 16 6 9 Parasitoid Eupelmidae 1 1 2 2 3 3 0 0 Parasitoid Eurytomidae 0 0 0 0 1 1 1 1 Parasitoid Evanidae 1 1 3 27 1 7 1 2 Parasitoid Fidellidae 1 1 0 0 2 2 1 7 Polinator Figitidae 0 0 1 1 0 0 1 1 Parasitoid Formicidae 25 3954 26 6631 29 2762 28 2104 Predator Heloridae 1 1 1 1 0 0 0 0 Parasitoid
58
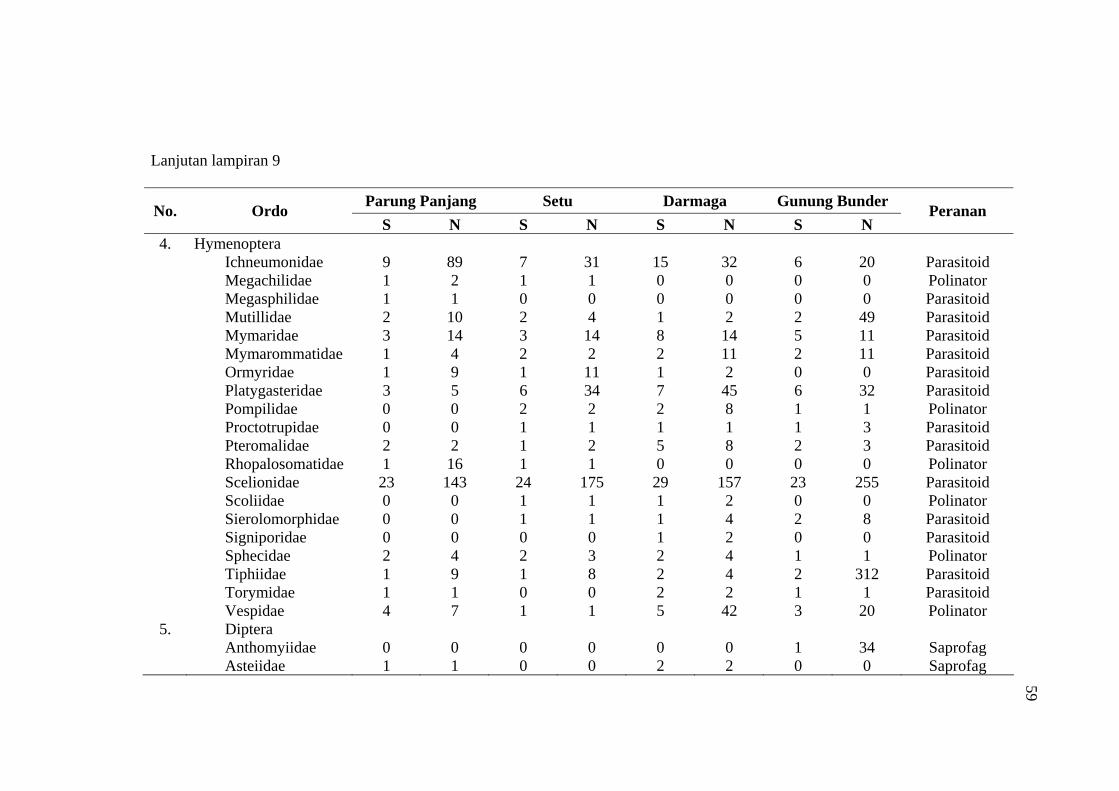
Lanjutan lampiran 9
Parung Panjang Setu Darmaga Gunung Bunder No. Ordo S N S N S N S N
Peranan
4. Hymenoptera Ichneumonidae 9 89 7 31 15 32 6 20 Parasitoid Megachilidae 1 2 1 1 0 0 0 0 Polinator Megasphilidae 1 1 0 0 0 0 0 0 Parasitoid Mutillidae 2 10 2 4 1 2 2 49 Parasitoid Mymaridae 3 14 3 14 8 14 5 11 Parasitoid Mymarommatidae 1 4 2 2 2 11 2 11 Parasitoid Ormyridae 1 9 1 11 1 2 0 0 Parasitoid Platygasteridae 3 5 6 34 7 45 6 32 Parasitoid Pompilidae 0 0 2 2 2 8 1 1 Polinator Proctotrupidae 0 0 1 1 1 1 1 3 Parasitoid Pteromalidae 2 2 1 2 5 8 2 3 Parasitoid Rhopalosomatidae 1 16 1 1 0 0 0 0 Polinator Scelionidae 23 143 24 175 29 157 23 255 Parasitoid Scoliidae 0 0 1 1 1 2 0 0 Polinator Sierolomorphidae 0 0 1 1 1 4 2 8 Parasitoid Signiporidae 0 0 0 0 1 2 0 0 Parasitoid Sphecidae 2 4 2 3 2 4 1 1 Polinator Tiphiidae 1 9 1 8 2 4 2 312 Parasitoid Torymidae 1 1 0 0 2 2 1 1 Parasitoid Vespidae 4 7 1 1 5 42 3 20 Polinator
5. Diptera Anthomyiidae 0 0 0 0 0 0 1 34 Saprofag Asteiidae 1 1 0 0 2 2 0 0 Saprofag
59
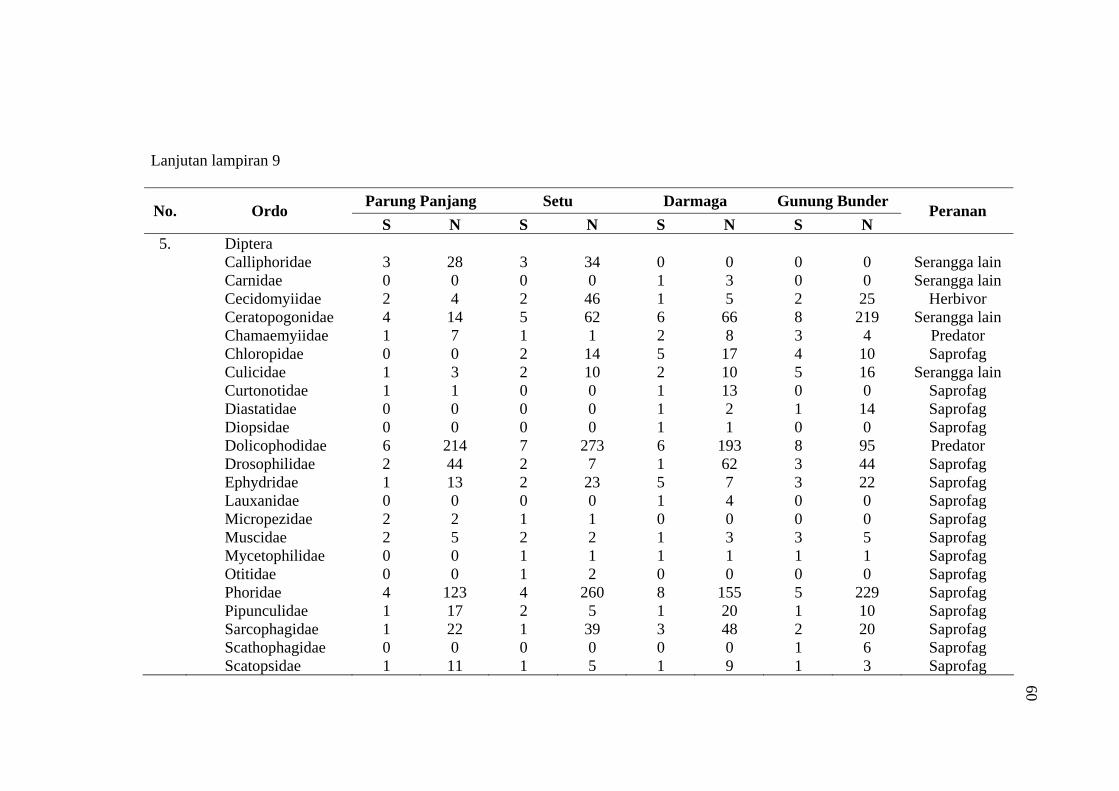
Lanjutan lampiran 9
Parung Panjang Setu Darmaga Gunung Bunder No. Ordo S N S N S N S N
Peranan
5. Diptera Calliphoridae 3 28 3 34 0 0 0 0 Serangga lain Carnidae 0 0 0 0 1 3 0 0 Serangga lain Cecidomyiidae 2 4 2 46 1 5 2 25 Herbivor Ceratopogonidae 4 14 5 62 6 66 8 219 Serangga lain Chamaemyiidae 1 7 1 1 2 8 3 4 Predator Chloropidae 0 0 2 14 5 17 4 10 Saprofag Culicidae 1 3 2 10 2 10 5 16 Serangga lain Curtonotidae 1 1 0 0 1 13 0 0 Saprofag Diastatidae 0 0 0 0 1 2 1 14 Saprofag Diopsidae 0 0 0 0 1 1 0 0 Saprofag Dolicophodidae 6 214 7 273 6 193 8 95 Predator Drosophilidae 2 44 2 7 1 62 3 44 Saprofag Ephydridae 1 13 2 23 5 7 3 22 Saprofag Lauxanidae 0 0 0 0 1 4 0 0 Saprofag Micropezidae 2 2 1 1 0 0 0 0 Saprofag Muscidae 2 5 2 2 1 3 3 5 Saprofag Mycetophilidae 0 0 1 1 1 1 1 1 Saprofag Otitidae 0 0 1 2 0 0 0 0 Saprofag Phoridae 4 123 4 260 8 155 5 229 Saprofag Pipunculidae 1 17 2 5 1 20 1 10 Saprofag Sarcophagidae 1 22 1 39 3 48 2 20 Saprofag Scathophagidae 0 0 0 0 0 0 1 6 Saprofag Scatopsidae 1 11 1 5 1 9 1 3 Saprofag
60
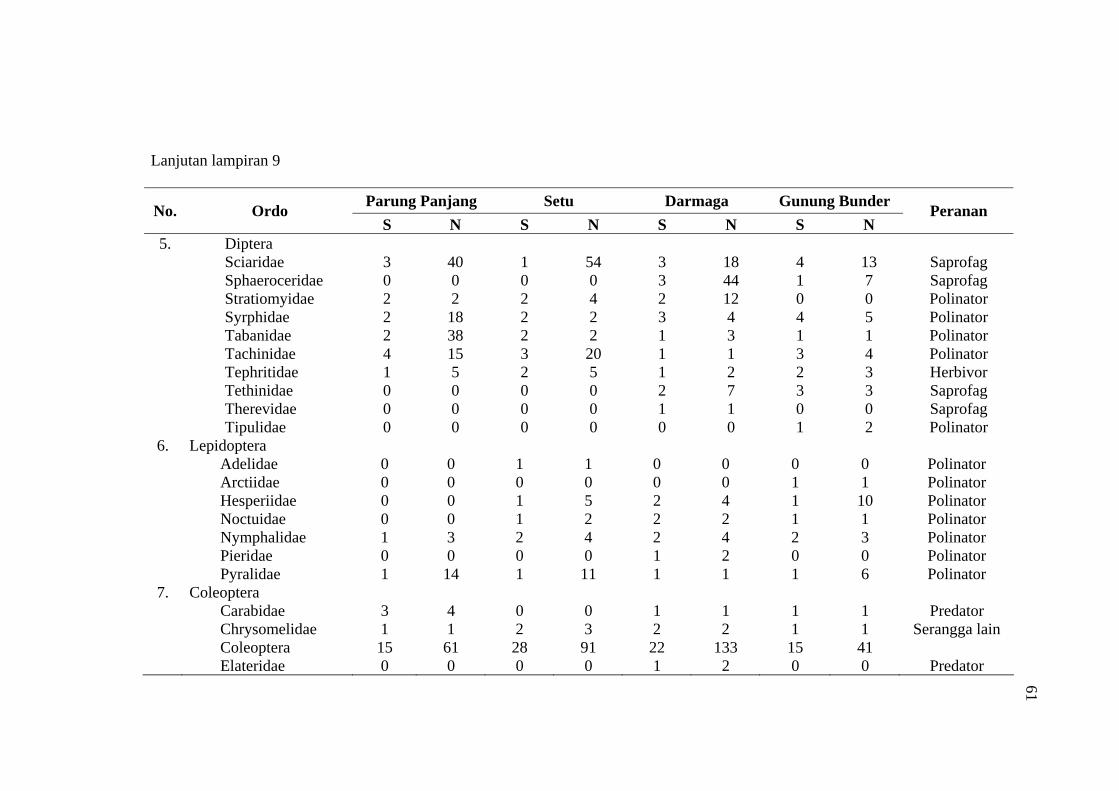
Lanjutan lampiran 9
Parung Panjang Setu Darmaga Gunung Bunder No. Ordo S N S N S N S N
Peranan
5. Diptera Sciaridae 3 40 1 54 3 18 4 13 Saprofag Sphaeroceridae 0 0 0 0 3 44 1 7 Saprofag Stratiomyidae 2 2 2 4 2 12 0 0 Polinator Syrphidae 2 18 2 2 3 4 4 5 Polinator Tabanidae 2 38 2 2 1 3 1 1 Polinator Tachinidae 4 15 3 20 1 1 3 4 Polinator Tephritidae 1 5 2 5 1 2 2 3 Herbivor Tethinidae 0 0 0 0 2 7 3 3 Saprofag Therevidae 0 0 0 0 1 1 0 0 Saprofag Tipulidae 0 0 0 0 0 0 1 2 Polinator
6. Lepidoptera Adelidae 0 0 1 1 0 0 0 0 Polinator Arctiidae 0 0 0 0 0 0 1 1 Polinator Hesperiidae 0 0 1 5 2 4 1 10 Polinator Noctuidae 0 0 1 2 2 2 1 1 Polinator Nymphalidae 1 3 2 4 2 4 2 3 Polinator Pieridae 0 0 0 0 1 2 0 0 Polinator Pyralidae 1 14 1 11 1 1 1 6 Polinator
7. Coleoptera Carabidae 3 4 0 0 1 1 1 1 Predator Chrysomelidae 1 1 2 3 2 2 1 1 Serangga lain Coleoptera 15 61 28 91 22 133 15 41 Elateridae 0 0 0 0 1 2 0 0 Predator
61
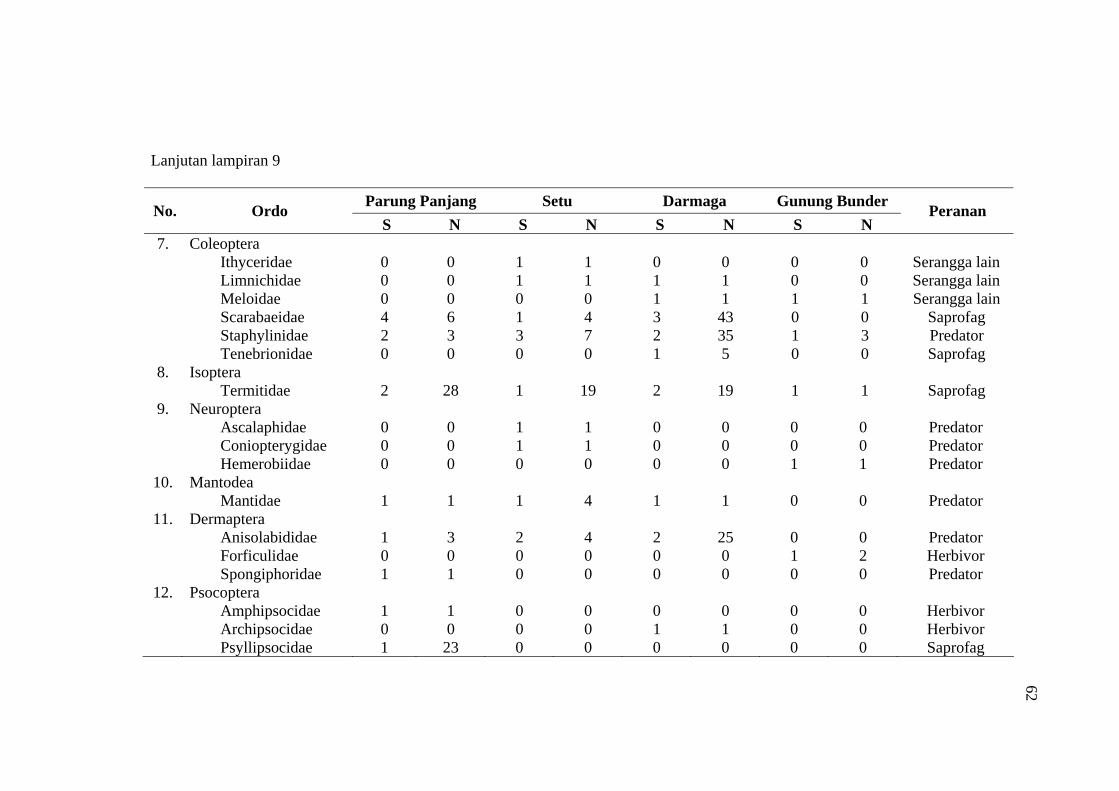
Lanjutan lampiran 9
Parung Panjang Setu Darmaga Gunung Bunder No. Ordo S N S N S N S N
Peranan
7. Coleoptera Ithyceridae 0 0 1 1 0 0 0 0 Serangga lain Limnichidae 0 0 1 1 1 1 0 0 Serangga lain Meloidae 0 0 0 0 1 1 1 1 Serangga lain Scarabaeidae 4 6 1 4 3 43 0 0 Saprofag Staphylinidae 2 3 3 7 2 35 1 3 Predator Tenebrionidae 0 0 0 0 1 5 0 0 Saprofag
8. Isoptera Termitidae 2 28 1 19 2 19 1 1 Saprofag
9. Neuroptera Ascalaphidae 0 0 1 1 0 0 0 0 Predator Coniopterygidae 0 0 1 1 0 0 0 0 Predator Hemerobiidae 0 0 0 0 0 0 1 1 Predator
10. Mantodea Mantidae 1 1 1 4 1 1 0 0 Predator
11. Dermaptera Anisolabididae 1 3 2 4 2 25 0 0 Predator Forficulidae 0 0 0 0 0 0 1 2 Herbivor Spongiphoridae 1 1 0 0 0 0 0 0 Predator
12. Psocoptera Amphipsocidae 1 1 0 0 0 0 0 0 Herbivor Archipsocidae 0 0 0 0 1 1 0 0 Herbivor Psyllipsocidae 1 23 0 0 0 0 0 0 Saprofag
62
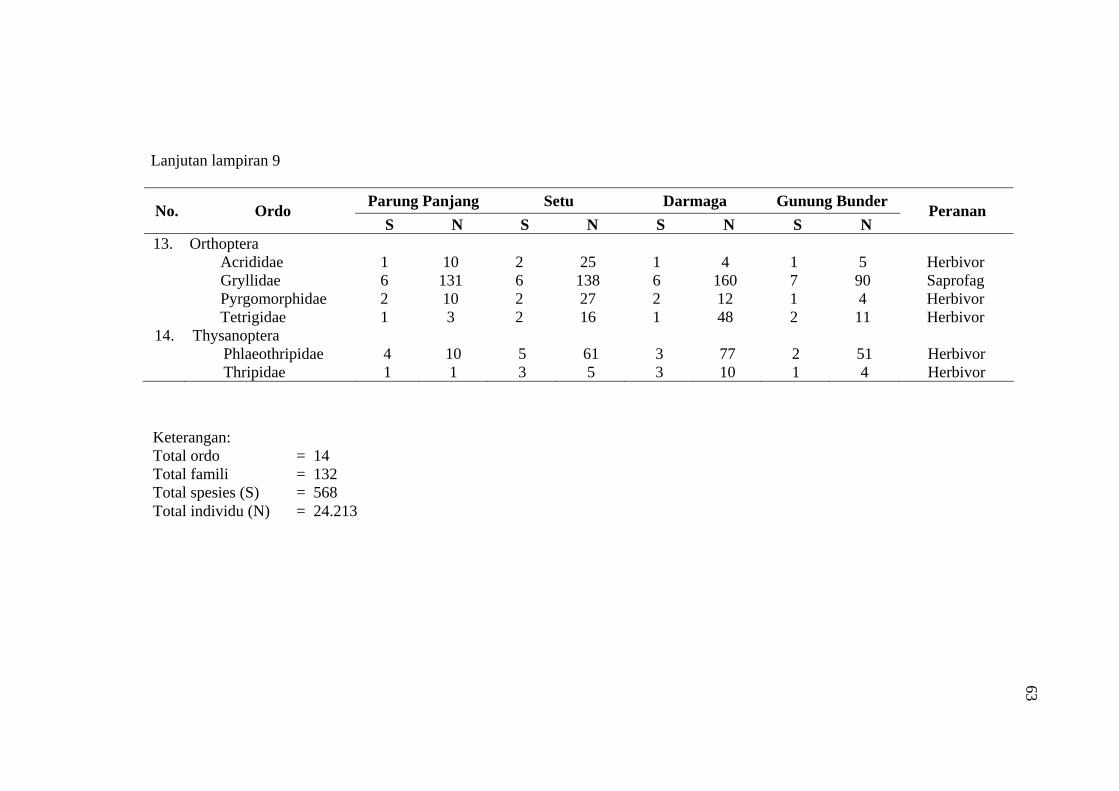
Lanjutan lampiran 9
Parung Panjang Setu Darmaga Gunung Bunder No. Ordo S N S N S N S N
Peranan
13. Orthoptera Acrididae 1 10 2 25 1 4 1 5 Herbivor Gryllidae 6 131 6 138 6 160 7 90 Saprofag Pyrgomorphidae 2 10 2 27 2 12 1 4 Herbivor Tetrigidae 1 3 2 16 1 48 2 11 Herbivor
14. Thysanoptera Phlaeothripidae 4 10 5 61 3 77 2 51 Herbivor Thripidae 1 1 3 5 3 10 1 4 Herbivor
Keterangan: Total ordo = 14 Total famili = 132 Total spesies (S) = 568 Total individu (N) = 24.213
63