hasil dan pembahasan - repository.ipb.ac.id · ca yanlappa-jasinga 1 petak 46 (hymaneae caubaril)...
TRANSCRIPT

HASIL DAN PEMBAHASAN
Pengambilan Sampel Rayap di Lapangan
Rayap yang dikoleksi dari lapangan ditemukan pada lokasi dengan ketinggian yang berbeda. Lokasi CA Yanlappa-Jasinga berada pada ketinggian 106 m dpl, sedangkan rayap yang berlokasi di Kampus IPB Dramaga berada pada ketinggian 205-213 m dpl (Tabel 3). Hal ini membuktikan bahwa rayap spesies M. gilvus dapat ditemukan pada lokasi dengan ketinggian yang berbeda.
Pohon yang menjadi naungan pada lokasi pengambilan sampel di CA Yanlappa-Jasinga pada petak 46, 29, dan 28 secara berurutan yaitu Hymaneae
caubaril, Schima walichi, dan Artocarpus altisima, sedangkan pohon yang menjadi naungan pada lokasi pengambilan sampel di Kampus IPB Dramaga yaitu Cyrtostachys sp., Ficus sp., Elaeis guineensis, Hevea brasiliensis (Tabel 3). Lokasi CA Yanlappa-Jasinga mempunyai naungan pohon-pohon yang besar, sehingga persentase naungannya lebih dari 90% dan sinar matahari yang masuk sangat sedikit ke dalam tanaman di bawahnya. Lokasi Kampus IPB Dramaga mempunyai naungan dengan persentase yang cukup tinggi juga yaitu sekitar 80% (Mubin 2013), sehingga sinar matahari yang masuk juga tidak terlalu banyak. Kanopi dengan jumlah yang semakin banyak akan menutupi permukaan tanah maka akan berpengaruh dengan kelimpahan dan keanekaragaman jenis rayap (Dibog et al.
1999).
Tabel 3 Lokasi pengambilan sampel rayap di lapangan Plot Lokasi petak
(tanaman pelindung) Kode isolat Koordinat Elevasi
(m dpl) CA Yanlappa-Jasinga 1 Petak 46
(Hymaneae caubaril)
Mg 3 JSG S: 060 24.951’ E: 1060 29.972’
106
2 Petak 29 (Schima walichi}
Mg 4 JSG S: 060 25.080’ E: 1060 30.013’
106
3 Petak 28
(Artocarpus altisima)
Mg 5 JSG S: 060 25.080’ E: 1060 30.013’
106
Kampus IPB-Dramaga 1 Perumahan Dosen IPB
(Cyrtostachys sp., Ficus sp.) Mg 2 DMG S: 06033.229’
E: 106043.428’ 209
2 Kebun Percobaan Kelapa Sawit Cikabayan (Elaeis guineensis)
Mg 4 DMG S: 06033.119’ E: 106043.997’
213
3 Kebun Percobaan Karet Rektorat (Hevea brasiliensis)
Mg 6 DMG S: 06033.640’ E: 106043.476’
205
Keterangan: CA: Cagar Alam, Mg: Macrotermes gilvus, JSG: Jasinga, DMG: Dramaga, S: South (Lintang Selatan), E: East (Lintang Timur), m dpl: meter di atas permukaan laut

16
Rayap yang diperoleh dari CA Yanlappa-Jasinga dan Kampus IPB Dr amaga mempunyai spesies yang sama yaitu M. gilvus. Spesies ini melimpah di Cagar Alam Yanlappa (Subekti 2010) dan di Kampus IPB Dramaga (Mubin 2013). Koloni rayap yang ditemukan di lapangan sebanyak 3 koloni pada setiap lokasi yaitu 3 koloni dari CA Yanlappa-Jasinga dengan kode koloni Mg 3 JSG, Mg 4 JSG, dan Mg 5 JSG, sedangkan koloni rayap dari Kampus IPB Dramaga yaitu Mg 2 DMG, Mg 4 DMG, dan Mg 6 DMG. Lokasi penemuan koloni rayap di CA Yanlappa-Jasinga mempunyai jarak yang berdekatan, begitu pula dengan koloni rayap yang ditemukan di Kampus IPB Dramaga (Tabel 3). Koordinat pada lokasi pengambilan sampel di CA Yanlappa-Jasinga dan Kampus IPB Dramga mempunyai perbedaan yaitu lokasi pengambilan sampel di CA Yanlappa-Jasinga hanya berbeda di bagian menit, sedangkan lokasi pengambilan sampel di Kampus IPB Dramaga hanya berbeda di bagian detik (Tabel 3). Akan tetapi, lokasi yang dijadikan pengambilan sampel di CA Yanlappa mempunyai elevasi yang sama (106 mdpl), sedangkan di Kampus IPB mempunyai elevasi yang berbeda (205, 209, dan 213 mdpl) (Tabel 3).
Rayap yang diambil dari CA Yanlappa-Jasinga maupun dari Kampus IPB Dramaga merupakan rayap yang berasal dari koloni yang berbeda. Berdasarkan koordinat dan elevasi, sampel rayap yang diambil mempunyai lokasi yang yang berbeda meskipun perbedaannya tidak terlalu signifikan. Koordinat yang telah tercatat hanya menunjukkan bahwa koloni rayap mempunyai perbedaan lokasi berdasarkan nilai menit dan detik, sehingga sampel rayap yang ditemukan pada lokasi pengambilan sampel tidak jauh berbeda dengan koloni yang lain. Sama halnya dengan nilai elevasi, sampel rayap yang diambil menunjukkan nilai elevasi yang mempunyai rentang yang dekat (Tabel 3).
Gambar 4 Rayap M. gilvus kasta prajurit mayor dan minor Keterangan: 1) prajurit mayor, 2) prajurit minor, a) CA Yanlappa- Jasinga, b) Kampus IPB Dramaga
1 2
(a) (b) 5 mm
(a) (b) 5 mm
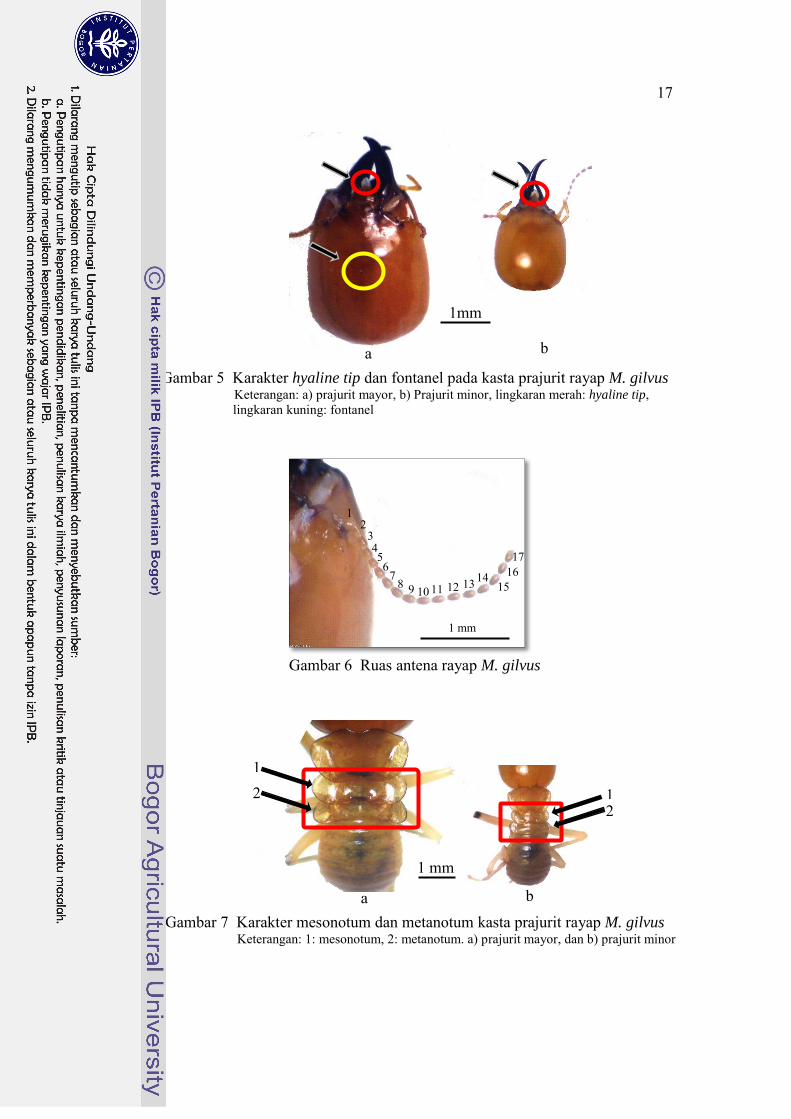
Gambar 5 Karakter hyaline tip dan fontanel pada kasta prajurit rayap M. gilvus Keterangan: a) prajurit mayor, b) Prajurit minor, lingkaran merah: hyaline tip, lingkaran kuning: fontanel
Gambar 6 Ruas antena rayap M. gilvus
Gambar 7 Karakter mesonotum dan metanotum kasta prajurit rayap M. gilvus Keterangan: 1: mesonotum, 2: metanotum. a) prajurit mayor, dan b) prajurit minor
1mm
a b
1 2
3 4 5 6
7 8 9 10 11 12 13 14
15 16 17
1 mm
1 2 1
2
a b
1 mm
17

18
Karakter kepala pada kasta prajurit mayor yaitu kepala berwarna coklat kemerahan, berambut jarang, mempunyai panjang (kepala+mandibel) 5.18-5.20 mm, dan lebar kepala 2.90-2.93 mm. Bagian tengah kepala (nampak dorsal) akan terlihat adanya fontanel dan hialin tip yang berada di ujung kepala (dekat mandibel) (Gambar 5). Mandibel pada rayap prajurit mayor terlihat kuat (strong) dengan warna hitam yang berfungsi untuk mencabik musuhnya. Ruas antena berjumlah 17 ruas (Gambar 6). Ukuran meso- dan metanotum hampir sama yaitu 2.21-2.24 mm (Tabel 4 dan Gambar 7).
Karakter kepala pada kasta prajurit minor yaitu kepala berwarna coklat terang, mempunyai panjang (kepala+mandibel) 3.16-3.18 mm, dan lebar kepala 1.11 mm. Bagian tengah kepala (nampak dorsal) akan terlihat adanya hialin tip yang berada di dekat mandibel (Gambar 5). Mandibel pada rayap prajurit minor tidak sekuat prajurit mayor tetapi mempunyai fungsi yang sama yaitu mencabik musuhnya. Ruas antena berjumlah 17 ruas (Gambar 6). Ukuran meso- dan metanotum tidak sama, ukuran mesonotum lebih kecil (1.10-1.12 mm) sedangkan ukuran metanotum lebih lebar (1.23 mm) (Tabel 4 dan Gambar 7).
Tabel 4 Ukuran tubuh rayap kasta prajurit M. gilvus dari CA Yanlappa-Jasinga dan Kampus IPB Dramaga
Karakter Ukuran morfometrik (mm) JSG DMG Ahmad* Panjang kepala + mandibel
PM : 5.20 Pm : 3.18
PM : 5.18 Pm : 3.16
PM : 4.80-5.48 Pm : 3.07-3.43
Lebar kepala PM : 2.93 Pm : 1.11
PM : 2.90 Pm : 1.11
PM : 2.88-3.17 Pm : 1.14-1.35
Mesonotum PM : 2.23 Pm : 1.10
PM : 2.21 Pm : 1.12
PM : 2.10-2.30 Pm : 1.09-1.14
Metanotum PM : 2.24 Pm : 1.23
PM : 2.21 Pm : 1.23
PM : 2.20-2.40 Pm : 1.19-1.25
Keterangan: JSG: CA Alam Yanlappa-Jasinga, DMG: Kampus IPB Dramaga, PM: Prajurit mayor, Pm: Prajurit minor, *Ahmad (1958)
Rayap M. gilvus diidentifikasi berdasarkan kasta prajurit yang mempunyai karakter morfologi yang mudah dibedakan dibandingkan kasta pekerja. Kasta prajurit mempunyai ukuran dan bentuk dari kepala dan mandibel yang mudah diamati, dibandingkan dengan kasta pekerja yang mempunyai struktur mandibel yang cukup rumit sehingga jarang dilakukan identifikasi. Ukuran kepala dan mandibel pada kasta prajurit digunakan sebagai karakter khusus dalam identifikasi. Kasta prajurit mayor dan minor yang ditemukan di CA Yanlappa-Jasinga dan Kampus IPB mempunyai ukuran kepala dan mandibel yang relatif sama pada kedua lokasi, sehingga identifikasi berdasarkan Ahmad (1958) dan Tho (1992) sudah tepat.

Analisis Kekerabatan Rayap Tanah M. gilvus dengan Pendekatan Perilaku
Uji Agonistik
Pengujian rayap di dalam arena dengan jarak + 5 cm memperlihatkan perilaku yang beragam. Hasil pengujian agonistik diperoleh hasil bahwa rayap dari koloni yang berasal dari lokasi yang sama (CA Yanlappa-Jasinga atau Kampus IPB Dramaga) tidak terlihat adanya perilaku agresif dengan ditandai rayap yang diujikan tidak saling menyerang (Tabel 5). Akan tetapi, rayap yang diuji dari lokasi yang berbeda (rayap dari CA Yanlappa-Jasinga vs Kampus IPB Dramaga) terlihat adanya perilaku saling menyerang yaitu kedua rayap saling menggigit.
Rayap pekerja yang diujikan memperlihatkan respon perilaku yang unik. Beberapa respon yang diperlihatkan yaitu pekerja langsung menjauh, pekerja mengetuk-ketukkan antena, dan ada yang langsung menyerang pekerja lain. Rayap dari lokasi yang sama (Dramaga vs Dramaga) memberikan respon yang langsung karena isyarat yang dikeluarkan sama dan respon yang dihasilkan yaitu rayap akan mengetuk-ketukkan antena kemudian berputar berkeliling. Berbeda halnya rayap dari lokasi yang berbeda (Dramaga vs CA Yanlappa) akan memberikan respon yang yang terlebih dahulu berputar-putar mengelilingi pekerja lain dan berusaha menyerang saat bertemu dengan rayap pekerja dari koloni yang lain tersebut. Jarak yang digunakan saat pengujian tidak menjadi pengaruh dalam respon pengenalan rayap pekerja lain. Rayap pekerja mayor dan prajurit minor antara M. subhyalinus
dan M. bellicosus diujikan dan hasil yang diperoleh menunjukkan bahwa tidak terlihat adanya korelasi jarak spasial dan indeks agresi (Jmhasly dan Leuthold 1999).
Tabel 5 Hasil uji agonistik rayap M. gilvus dari CA Yanlappa-Jasinga dan Kampus IPB Dramaga
1 2 3 4 5 6 [1] - - √ √ √ [2] - - √ √ √ [3] - - √ √ √ [4] √ √ √ - - [5] √ √ √ - - [6] √ √ √ - -
Keterangan: 1-3: koloni dari CA Yanlappa, 4-6: koloni dari Kampus IPB Dramaga, Mg: M. gilvus,
JSG: CA Yanlappa-Jasinga, DMG: Kampus IPB-Dramaga, (√): menunjukkan perilaku agresif, (-): tidak menunjukkan perilaku agresif
Koloni rayap yang berasal dari lokasi yang sama tidak menunjukkan adanya
perilaku agresif. Rayap dari koloni CA Yanlappa-Jasinga maupun Kampus IPB Dramaga terlihat adanya perilaku saling mendekat kemudian mengetuk-ketukkan antena ke individu yang lain. Hal ini menunjukkan bahwa kedua individu rayap yang diujikan melakukan pengenalan isyarat satu sama lain. Rayap pekerja yang terlihat saling menyerang mengindikasikan bahwa rayap mempunyai perilaku yang agresif pada saat proses pengenalan (Delphia et al. 2003). Individu rayap langsung
19

20
mengenali isyarat yang dikeluarkan dari individu yang lain dan secara langsung menyerang dan berusaha menggigit individu rayap tersebut.
Rayap yang terlihat adanya perilaku agresif mengindikasikan bahwa rayap masih mampu mengenali isyarat yang dikeluarkan oleh individu yang lain. Isyarat yang dikeluarkan dari individu rayap pekerja kemudian dikenali oleh individu rayap pekerja yang lain merupakan sifat dari perilaku pengenalan rayap di dalam koloninya (nestmate). Isyarat yang dikeluarkan berasal dari induk betina, sehingga rayap yang berperilaku saling mengenali satu sama lain merupakan rayap yang masih berasal dari tetua yang sama. Rayap pekerja mayor dan prajurit minor dari M. subhyalinus dan M. bellicosus diuji dan diperoleh hasil bahwa rayap pekerja mayor M. subhyalinus mempunyai kemampuan membedakan ekspresi dari variasi tingkat agresi terhadap pekerja mayor dari rayap M. bellicosus. Rayap prajurit minor (M. subhyalinus maupun M. bellicosus) juga bereaksi terhadap koloni asing dengan tingkat agresi yang berbeda (Jmhasly dan Leuthold 1999).
Rayap yang saling yang saling mendekat kemudian melakukan respon untuk menyerang atau menggigit mengindikasikan bahwa proses pengenalan isyarat dari individu satu dengan individu yang lain berhasil. Isyarat yang dikeluarkan dari individu satu kemudian diterima oleh individu rayap yang lain. Akan tetapi isyarat yang dikeluarkan tidak dikenali, sehingga menyebabkan rayap saling menyerang. Rayap menganggap bahwa proses pengenalan dari koloninya tidak berhasil karena isyarat yang dikeluarkan berbeda dengan yang dikeluarkannya. Rayap yang menganggap isyarat yang dikeluarkan tidak sama dengan koloninya, menganggap bahwa rayap lain tersebut sebagai musuh (Shelton dan Grace 1996). Koloni yang dulunya berasal dari tetua yang sama, tetapi karena sudah terlalu lama terpisah dapat menyebabkan pengenalan (recognize) dari koloni sebelumnya menjadi semakin luntur. Pengenalan dari koloni yang semakin melemah terhadap pengenalan dari koloni sebelumnya menyebabkan kedua koloni yang terpisah tidak saling mengenal. Koloni yang tidak saling mengenal terhadap isyarat dari koloni yang lain menyebabkan perilaku agresif pada rayap (Andrews 1911).
Analisis Kekerabatan Rayap Tanah M. gilvus dengan Teknik Molekuler
Hasil amplifikasi gen COI pada rayap. Primer forward dan reverse mampu mengamplifikasi gen COI dengan baik, terlihat pita DNA 1 sampai 4 merupakan hasil amplifikasi gen COI dari rayap. Pita DNA 1 dan 2 berasal dari DNA total rayap pekerja dari CA Yanlappa-Jasinga, sedangkan pita DNA 3 dan 4 berasal dari DNA total rayap pekerja Kampus IPB. Pita yang terlihat sesuai dengan target yang diinginkan yaitu berukuran sekitar 750pb (Gambar 8).
Analisis homologi sekuen DNA gen COI pada rayap. Keempat sekuen DNA rayap M. gilvus dari Bogor (CA Yanlappa-Jasinga dan Kampus IPB Dramaga) mempunyai homologi dengan rayap M. gilvus dari Laos dengan nilai query cover, e-value, dan max-identity berturut-turut adalah 100%, 0.0, dan 98% (Tabel 6). Rayap yang telah diidentifikasi secara morfologi dan berdasarkan hasil BLAST-N merujuk pada spesies yang sama yaitu M. gilvus.
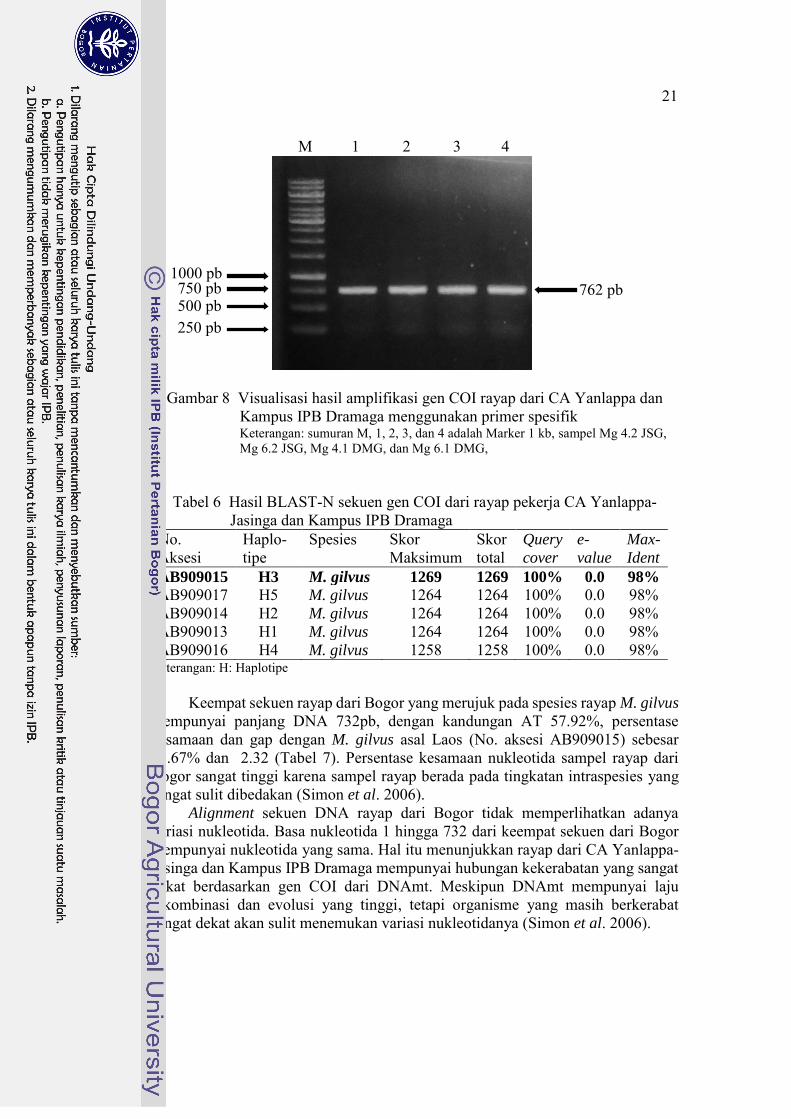
Gambar 8 Visualisasi hasil amplifikasi gen COI rayap dari CA Yanlappa dan Kampus IPB Dramaga menggunakan primer spesifik
Keterangan: sumuran M, 1, 2, 3, dan 4 adalah Marker 1 kb, sampel Mg 4.2 JSG, Mg 6.2 JSG, Mg 4.1 DMG, dan Mg 6.1 DMG,
Tabel 6 Hasil BLAST-N sekuen gen COI dari rayap pekerja CA Yanlappa- Jasinga dan Kampus IPB Dramaga
No. Aksesi
Haplo- tipe
Spesies Skor Maksimum
Skor total
Query
cover
e-
value
Max-
Ident
AB909015 H3 M. gilvus 1269 1269 100% 0.0 98%
AB909017 H5 M. gilvus 1264 1264 100% 0.0 98% AB909014 H2 M. gilvus 1264 1264 100% 0.0 98% AB909013 H1 M. gilvus 1264 1264 100% 0.0 98% AB909016 H4 M. gilvus 1258 1258 100% 0.0 98%
Keterangan: H: Haplotipe
Keempat sekuen rayap dari Bogor yang merujuk pada spesies rayap M. gilvus
mempunyai panjang DNA 732pb, dengan kandungan AT 57.92%, persentase kesamaan dan gap dengan M. gilvus asal Laos (No. aksesi AB909015) sebesar 97.67% dan 2.32 (Tabel 7). Persentase kesamaan nukleotida sampel rayap dari Bogor sangat tinggi karena sampel rayap berada pada tingkatan intraspesies yang sangat sulit dibedakan (Simon et al. 2006).
Alignment sekuen DNA rayap dari Bogor tidak memperlihatkan adanya variasi nukleotida. Basa nukleotida 1 hingga 732 dari keempat sekuen dari Bogor mempunyai nukleotida yang sama. Hal itu menunjukkan rayap dari CA Yanlappa-Jasinga dan Kampus IPB Dramaga mempunyai hubungan kekerabatan yang sangat dekat berdasarkan gen COI dari DNAmt. Meskipun DNAmt mempunyai laju rekombinasi dan evolusi yang tinggi, tetapi organisme yang masih berkerabat sangat dekat akan sulit menemukan variasi nukleotidanya (Simon et al. 2006).
M 1 2 3 4
250 pb
762 pb 500 pb 750 pb
1000 pb
21

22
Tabel 7 BLAST-N gen COI M. gilvus haplotipe baru menggunakan program BLAST-N (www.ncbi.nlm.nih.gov)
Karakter Gen COI M. gilvus Mg 4 DMG Mg 6 DMG Mg 4 JSG Mg 5 JSG
Panjang DNA (pb) 732 732 732 732 Kandungan AT (%) 57.92 57.92 57.92 57.92 Spesies lain M. gilvus
(Mg.15.H3) M. gilvus
(Mg.15.H3) M. gilvus
(Mg.15.H3) M. gilvus
(Mg.15.H3) No. Aksesi AB909015 AB909015 AB909015 AB909015 E-value 0.0 0.0 0.0 0.0 Persentase kesamaan (%)
97.67 97.67 97.67 97.67
Gap 2.32 (17/732)
2.32 (17/732)
2.32 (17/732)
2.32 (17/732)
Tabel 8 Posisi nukleotida gen COI M. gilvus dengan database sekuen nukleotida berdasarkan GenBank yang menunjukkan variasi haplotipe
Sampel Nukleotida ke- 12/90/384/399 414/639 33/264 39/174/341
/534/611 200 258 408 706
Mg 4 DMG C G T G T A T Mg 6 DMG . . . . . . . Mg 4 JSG . . . . . . . Mg 5 JSG . . . . . . . Mg 13 LAO T A C C C G . Mg 14 LAO T A C C C G . Mg 15 LAO T A C C . G . Mg 16 LAO T A C C C G G Mg 17 LAO T A C C . G G
Keterangan: Mg: M. gilvus, JSG: CA Yanlappa-Jasinga, DMG: Kampus IPB-Dramaga, LAO: Laos (asal spesimen rayap)
Sekuen DNA rayap dari Bogor di-alignment dengan sekuen DNA rayap M.
gilvus gen COI dari Laos dengan nomor aksesi secara berurutan AB909013, AB909014, AB909015, AB909016, AB909017 (secara berurutan Mg.13.H1, Mg.14.H2, Mg.15.H3, Mg.16.H4, dan Mg.17.H5). Sekuen-sekuen DNA rayap M.
gilvus dari Bogor tidak ada perbedaannya setelah di-alignment. Akan tetapi, sekuen-sekuen DNA rayap dari Bogor yang dibandingkan dengan sekuen DNA rayap dari Laos terdapat beberapa perbedaan sekuen nukleotida.
Sebanyak 17 basa nukleotida pada sekuen DNA rayap dari Bogor yang dibandingkan dengan sekuen DNA rayap haplotipe dari Laos (Tabel 8 dan Lampiran 6). Urutan basa ke-12, 90, 384, 399, 414, dan 639 pada sekuen rayap M.
gilvus dari Bogor menunjukkan basa “Cytosin (C)”, sedangkan pada haplotipe M.
gilvus dari Laos menunjukkan basa “Timin (T)”. Urutan basa ke-33 dan 264 dari rayap sekuens rayap Bogor menunjukkan basa “Guanin (G)”, sedangkan pada haplotipe dari Laos menunjukkan basa Adenin “A”. Sekuen DNA rayap dari Bogor

pada basa ke-200 yang mempunyai basa “G” yang sama dengan ke-33 dan 264 menunjukkan basa yang berbeda pada haplotipe dari Laos yaitu basa “C”. Sekuens rayap dari Bogor pada basa ke 408 menunjukkan basa “A”, sedangkan pada sekuen haplotipe dari Laos menunjukkan basa “G” (Tabel 8).
Gen COI pada penelitian ini merupakan gen yang digunakan untuk analisis dengan DNA Barkode. DNA barkode merupakan suatu cara untuk mengidentifikasi spesies secara cepat dan akurat menggunakan DNA pendek dari mitokondria yang distandardisasi (Hebert et al. 2003). Gen COI dipilih karena gen COI yang digunakan relatif pendek, bersifat conserve, relatif stabil tidak mudah mengalami perubahan-perubahan dengan gen-gen mitokondria yang lainnya, dan gen ini terdapat dalam jumlah kopi yang banyak (Hebert et al. 2003).
Beberapa penelitian lain menunjukan bahwa gen COI dapat mengidentifikasi spesies kriptik. Rayap yang telah diidentifikasi berdasarkan morfologi sebanyak 31 spesies, akan tetapi setelah dilihat kembali menggunakan penanda genetik gen COI ada tiga spesies kriptik dari 31 spesies tersebut (Bourguignon et al. (2013). Nukleotida yang berbeda pada sekuen dari Bogor dan haplotipe dari Laos telah berevolusi dan beradaptasi terhadap seleksi alam dari tempat asalnya (Asia Tenggara) ke Indonesia. Rayap M. gilvus mempunyai persebaran yang sangat luas dan hampir ditemukan di semua Asia Tenggara. Strategi dalam cara bertahan hidup menjadi salah satu faktor yang sangat penting bagi persebaran M. gilvus di berbagai Asia Tenggara. Hibernasi dan migrasi adalah dua strategi adaptasi utama untuk kelangsungan hidup serangga di musim dingin (Tauber et al. 1986). Strategi ini akan berpengaruh terhadap peningkatan proses distribusi M. gilvus.
Rayap Coptotermes formosanus Shiraki (famili Rhinotermtidae) dianalisis menggunakan gen COII dari DNAmt. Hasil yang diperoleh menunjukkan bahwa rayap C. formosanus yang menginfestasi dan merusak di Hawai berasal dari tetua yang sama yaitu berasal dari Asia dan Amerika utara. Hal ini menunjukkan bahwa rayap C. formosanus yang menginfestasi daerah Hawai merupakan satu tetua yang sama kemudian tersebar dan bertahan hidup. Hubungan evolusi dapat dilihat dari pohon filogenetik yang menunjukkan bahwa rayap C. formosanus yang berasal dari Jepang, China, dan Amerika membentuk cabang parafiletik (Gentz et al. 2008).
Analisis filogeni dan jarak genetik rayap. Analisis filogeni 732 basa dari
gen COI rayap diperoleh bahwa sebanyak 715 basa konstan dari 732, basa variable sebanyak 17 (2.32%), dan parsimony-informatif sebanyak 17 (2.32%). Jumlah basa konstan yang diperoleh tergolong sangat tinggi dan basa variable dan basa parsimony-informatif mempunyai nilai yang sangat rendah persentasenya karena rayap berada dalam tingkat intraspesies.
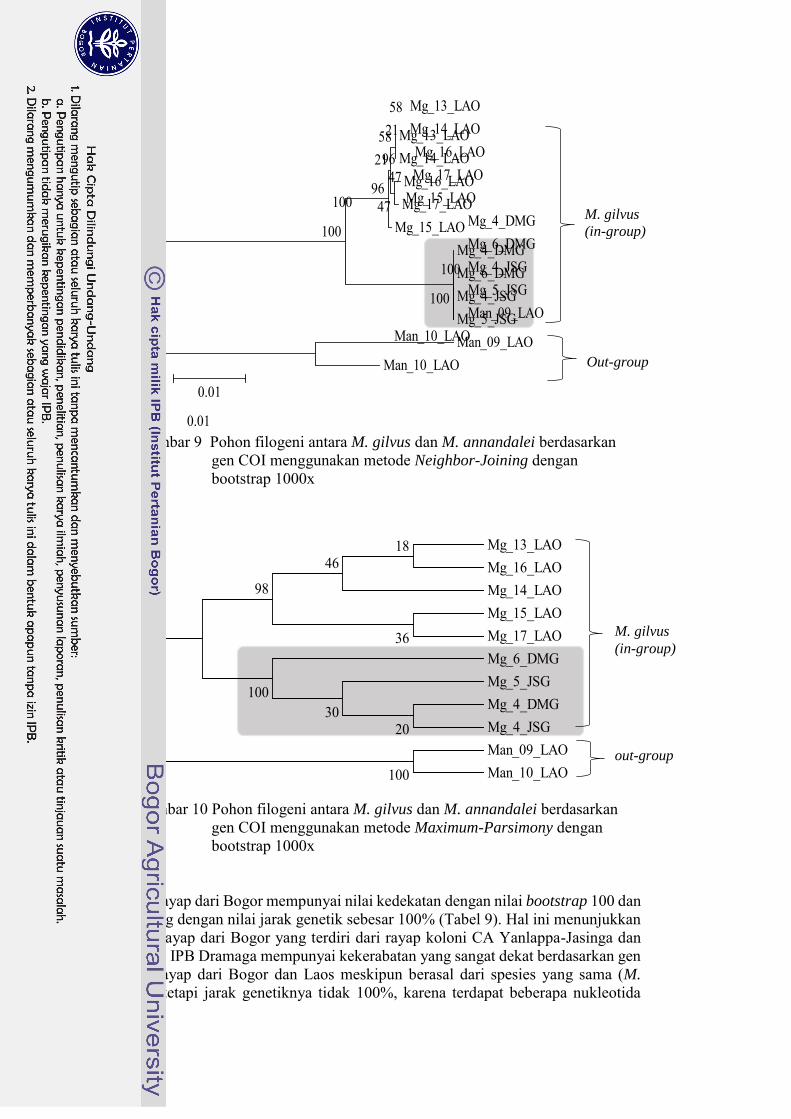
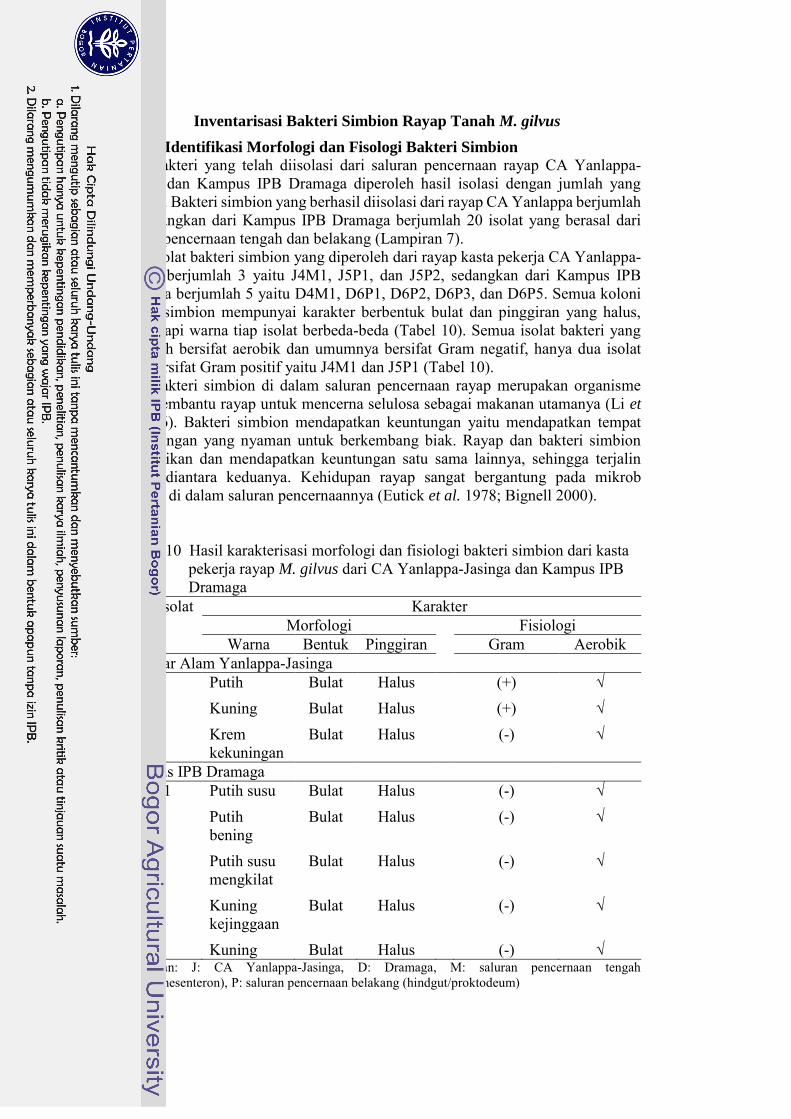
Konstruksi pohon filogeni dengan metode Neighbor-Joining (NJ) dan Maximum-Parsimony (MP) menunjukkan sedikit perbedaan pada percabangan antara rayap-rayap di Bogor. Rayap dari Bogor pada pohon filogeni NJ tidak membentuk percabangan (Gambar 9), sedangkan pada pohon filogeni MP membentuk percabang (Gambar 10), walaupun dengan nilai kepercayaan (bootstrap) yang rendah yaitu 30 dan 20%. Rayap M. annandalei membentuk cabang yang berbeda karena termasuk out-group.
23
24
Gambar 9 Pohon filogeni antara M. gilvus dan M. annandalei berdasarkan gen COI menggunakan metode Neighbor-Joining dengan bootstrap 1000x
Gambar 10 Pohon filogeni antara M. gilvus dan M. annandalei berdasarkan
gen COI menggunakan metode Maximum-Parsimony dengan bootstrap 1000x
Rayap dari Bogor mempunyai nilai kedekatan dengan nilai bootstrap 100 dan didukung dengan nilai jarak genetik sebesar 100% (Tabel 9). Hal ini menunjukkan bahwa rayap dari Bogor yang terdiri dari rayap koloni CA Yanlappa-Jasinga dan Kampus IPB Dramaga mempunyai kekerabatan yang sangat dekat berdasarkan gen COI. Rayap dari Bogor dan Laos meskipun berasal dari spesies yang sama (M.
gilvus) tetapi jarak genetiknya tidak 100%, karena terdapat beberapa nukleotida
Mg_13_LAO Mg_16_LAO Mg_14_LAO Mg_15_LAO Mg_17_LAO Mg_6_DMG Mg_5_JSG Mg_4_DMG Mg_4_JSG Man_09_LAO Man_10_LAO100
2030
1846
36
98
100
M. gilvus
(in-group)
out-group
Mg_13_LAO Mg_14_LAO Mg_16_LAO Mg_17_LAO
Mg_15_LAO Mg_4_DMG Mg_6_DMG Mg_4_JSG Mg_5_JSG Man_09_LAO
Man_10_LAO
100
100
58
47
21
96
0.01
Mg_13_LAO Mg_14_LAO Mg_16_LAO Mg_17_LAO
Mg_15_LAO Mg_4_DMG Mg_6_DMG Mg_4_JSG Mg_5_JSG Man_09_LAO
Man_10_LAO
100
100
58
47
21
96
0.01
Out-group
M. gilvus
(in-group)

yang berbeda (tabel 8). Rayap M. gilvus dari Bogor yang dibandingkan dengan out-
group (M. annandalei) mempunyai jarak genetik sebesar 91-92% karena rayap dibandingkan pada tingkat intraspesies.
Tabel 9 Jarak genetik M. gilvus gen COI dengan sampel dari GenBank 1 2 3 4 5 6 7 8 9 10 11 [1] [2] 100 [3] 100 100 [4] 100 100 100 [5] 97.8 97.8 97.8 97.8 [6] 97.8 97.8 97.8 97.8 100 [7] 97.9 97.9 97.9 97.9 99.8 99.8 [8] 97.6 97.6 97.6 97.6 99.8 99.8 99.7 [9] 97.8 97.8 97.8 97.8 99.7 99.7 99.8 99.8 [10] 91.5 91.5 91.5 91.5 92.3 92.3 92.3 92.3 92.3 [11] 92.4 92.4 92.4 92.4 93 93 93 93 93 97.2
Keterangan: 1: Mg_4_DMG, 2: Mg_6_DMG, 3: Mg_4_JSG, 4: Mg_5_JSG, 5: Mg_13_LAO, 6: Mg_14_LAO, 7: Mg_15_LAO, 8: Mg_16_LAO, 9: Mg_17_LAO, 10: Man_09_LAO, 11: Man_10_LAO
Analisis Kekerabatan Rayap Tanah M. gilvus
Keempat sekuen rayap dari Bogor (CA Yanlappa-Jasinga dan Kampus IPB Dramaga) yang telah dianalisis homologinya memberikan hasil yang sama dengan identifikasi rayap secara morfologi yaitu merujuk pada spesies rayap M. gilvus. Analisis homologi dari sekuen basa nukleotida dari keempat rayap dari Bogor mempunyai basa nukleotida yang sama.
DNA mitokondria merupakan DNA yang diturunkan secara murni dari induk betina (maternal) dan mempunyai tingkat evolusi yang tinggi (Simon et al. 2006). Analisis kekerabatan menggunakan pendekatan perilaku menunjukkan bahwa rayap dari CA Yanlappa-Jasinga dan Kampus IPB Dramaga diduga berasal dari tetua yang berbeda. Rayap pekerja yang terlihat saling agresif ini mengindikasikan bahwa rayap berasal dari induk yang berbeda, karena isyarat yang dikeluarkan rayap pekerja dari masing-masing lokasi dikenali bukan sebagai anggota koloninya (nestmate) tetapi dikenali sebagi musuh (Shelton & Grace 1996).
Hasil yang diperoleh antara pendekatan perilaku dan molekuler menunjukkan hasil yang bertolak belakang. Analisis menggunakan pendekatan perilaku menunjukkan hasil bahwa rayap M. gilvus berasal dari tetua yang berbeda. Berdasarkan pendekatan molekuler menunjukkan hasil bahwa rayap mempunyai kedekatan yang tinggi berdasarkan gen COI.
25
26
Inventarisasi Bakteri Simbion Rayap Tanah M. gilvus
Isolasi, Identifikasi Morfologi dan Fisologi Bakteri Simbion
Bakteri yang telah diisolasi dari saluran pencernaan rayap CA Yanlappa-Jasinga dan Kampus IPB Dramaga diperoleh hasil isolasi dengan jumlah yang berbeda. Bakteri simbion yang berhasil diisolasi dari rayap CA Yanlappa berjumlah 17, sedangkan dari Kampus IPB Dramaga berjumlah 20 isolat yang berasal dari saluran pencernaan tengah dan belakang (Lampiran 7).
Isolat bakteri simbion yang diperoleh dari rayap kasta pekerja CA Yanlappa-Jasinga berjumlah 3 yaitu J4M1, J5P1, dan J5P2, sedangkan dari Kampus IPB Dramaga berjumlah 5 yaitu D4M1, D6P1, D6P2, D6P3, dan D6P5. Semua koloni bakteri simbion mempunyai karakter berbentuk bulat dan pinggiran yang halus, akan tetapi warna tiap isolat berbeda-beda (Tabel 10). Semua isolat bakteri yang diperoleh bersifat aerobik dan umumnya bersifat Gram negatif, hanya dua isolat yang bersifat Gram positif yaitu J4M1 dan J5P1 (Tabel 10).
Bakteri simbion di dalam saluran pencernaan rayap merupakan organisme yang membantu rayap untuk mencerna selulosa sebagai makanan utamanya (Li et
al. 2006). Bakteri simbion mendapatkan keuntungan yaitu mendapatkan tempat perlindungan yang nyaman untuk berkembang biak. Rayap dan bakteri simbion memberikan dan mendapatkan keuntungan satu sama lainnya, sehingga terjalin sinergi diantara keduanya. Kehidupan rayap sangat bergantung pada mikrob simbion di dalam saluran pencernaannya (Eutick et al. 1978; Bignell 2000).
Tabel 10 Hasil karakterisasi morfologi dan fisiologi bakteri simbion dari kasta pekerja rayap M. gilvus dari CA Yanlappa-Jasinga dan Kampus IPB Dramaga
Kode isolat Karakter Morfologi Fisiologi
Warna Bentuk Pinggiran Gram Aerobik Cagar Alam Yanlappa-Jasinga
J4M1
Putih Bulat Halus (+) √ J5P1
Kuning Bulat Halus (+) √ J5P2 Krem
kekuningan Bulat Halus (-) √
Kampus IPB Dramaga D4M1
Putih susu Bulat Halus (-) √ D6P1
Putih bening
Bulat Halus (-) √
D6P2 Putih susu mengkilat
Bulat Halus (-) √
D6P3 Kuning kejinggaan
Bulat Halus (-) √
D6P5 Kuning Bulat Halus (-) √ Keterangan: J: CA Yanlappa-Jasinga, D: Dramaga, M: saluran pencernaan tengah (midgut/mesenteron), P: saluran pencernaan belakang (hindgut/proktodeum)
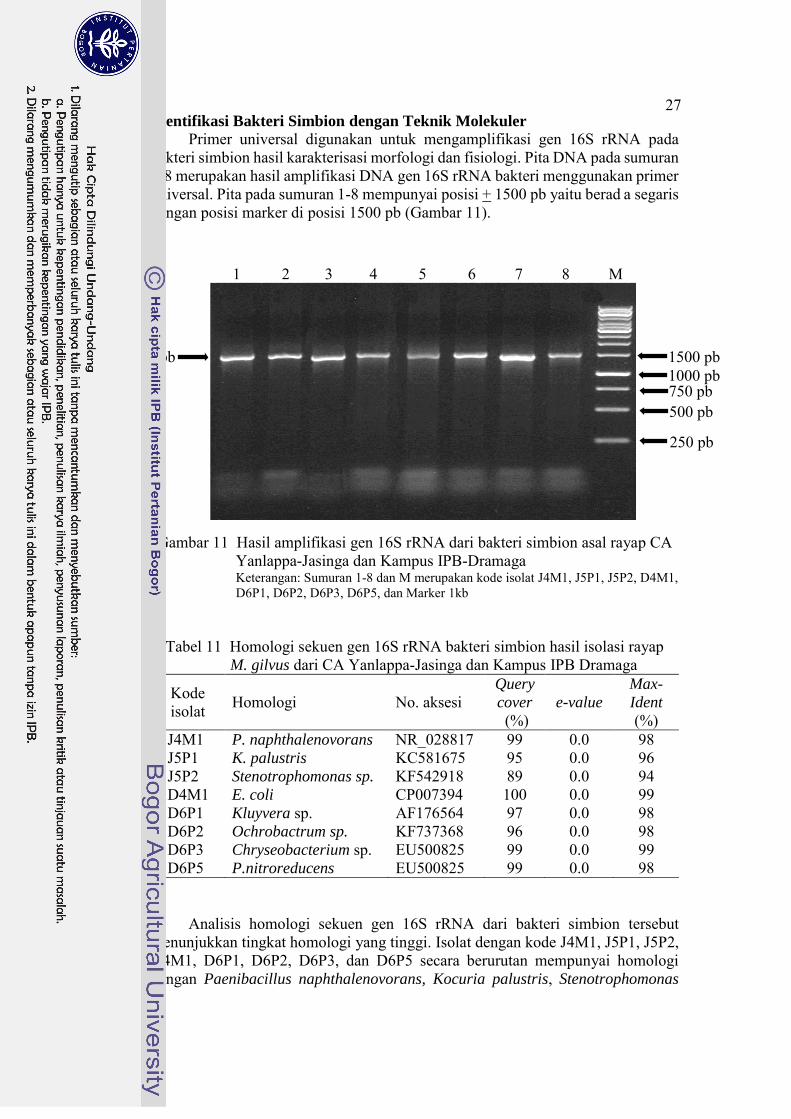
Identifikasi Bakteri Simbion dengan Teknik Molekuler
Primer universal digunakan untuk mengamplifikasi gen 16S rRNA pada bakteri simbion hasil karakterisasi morfologi dan fisiologi. Pita DNA pada sumuran 1-8 merupakan hasil amplifikasi DNA gen 16S rRNA bakteri menggunakan primer universal. Pita pada sumuran 1-8 mempunyai posisi + 1500 pb yaitu berad a segaris dengan posisi marker di posisi 1500 pb (Gambar 11).
Gambar 11 Hasil amplifikasi gen 16S rRNA dari bakteri simbion asal rayap CA Yanlappa-Jasinga dan Kampus IPB-Dramaga Keterangan: Sumuran 1-8 dan M merupakan kode isolat J4M1, J5P1, J5P2, D4M1, D6P1, D6P2, D6P3, D6P5, dan Marker 1kb
Tabel 11 Homologi sekuen gen 16S rRNA bakteri simbion hasil isolasi rayap M. gilvus dari CA Yanlappa-Jasinga dan Kampus IPB Dramaga
Kode isolat Homologi No. aksesi
Query
cover
(%) e-value
Max-
Ident
(%) J4M1 P. naphthalenovorans NR_028817 99 0.0 98 J5P1 K. palustris KC581675 95 0.0 96 J5P2 Stenotrophomonas sp. KF542918 89 0.0 94 D4M1 E. coli CP007394 100 0.0 99 D6P1 Kluyvera sp. AF176564 97 0.0 98 D6P2 Ochrobactrum sp. KF737368 96 0.0 98 D6P3 Chryseobacterium sp. EU500825 99 0.0 99 D6P5 P.nitroreducens EU500825 99 0.0 98
Analisis homologi sekuen gen 16S rRNA dari bakteri simbion tersebut menunjukkan tingkat homologi yang tinggi. Isolat dengan kode J4M1, J5P1, J5P2, D4M1, D6P1, D6P2, D6P3, dan D6P5 secara berurutan mempunyai homologi dengan Paenibacillus naphthalenovorans, Kocuria palustris, Stenotrophomonas
1500 pb 1500 pb 1000 pb 750 pb 500 pb
250 pb
1 2 3 4 5 6 7 8 M
27

28
sp., Escherichia coli, Kluyvera sp., Ochrobactrum sp., Chryseobacterium sp., dan Pseudomonas nitroreducens (Tabel 11).
Bakteri yang ditemukan di rayap penumbuh jamur (fungus garden) dari subfamili Macrotermitidae tidak terlihatnya ada kemiripan jenis. Zhu et al. (2012) telah menemukan 105 klon bakteri dari M. barneyi yaitu Bacteroidetes sebesar 47.6%, Firmicutes sebesar 28.6%, Proteobacteria sebesar 14.3%, Deferribacteres sebesar 4.8%, Actinobacteria sebesar 2.8%, dan Planctomycetes sebesar 1.9%. Bakteri pendegradasi Rhodococcus opacus strain RW juga ditemukan di rayap M.
michaelseni (Ngugi et al. 2005). Rayap Odontotermes sp. ditemukan bakteri Wolbachia sp. (Salunke et al. 2010) dan kelompok Firmicutes, Bacteriodates/ Chlorobi, Proteobacteria, dan Actinobacteria dari domain Bacteria yang ditemukan di saluran pencernaan rayap (Shinzato et al. 2007).
Analisis Bakteri Simbion di dalam Saluran Pencernaan
Rayap Tanah M. gilvus
Paenibacillus naphthalenovorans
Isolat J4M1 merupakan isolat bakteri yang diisolasi dari saluran pencernaan tengah (mesenteron) rayap pekerja dari CA Yanlappa. Isolat Y3M1 mempunyai homologi dengan Paenibacillus naphthalenovorans strain PR-N1 (No. aksesi NR_028817). Isolat bakteri J4M1 mempunyai nilai query-cover, e-value, dan maximum identity sebesar 99%, 0.0, dan 98% (Tabel 11).
Paenibacillus naphthalenovorans berasal dari kata naph.tha.le.no dan vo.rans yaitu naphthalene dari bahasa Persia, sedangkan vo.rans atau vorare
menjadi devour yang berarti menelan/melahap, sehingga bakteri tersebut berarti bahwa mampu menelan senyawa kimia naphthalen. Bakteri Paenibacillus sp. merupakan golongan Proteobacteria yang bersifat gram positif. Jenis bakteri ini mampu mendegradasi polisakarida dan senyawa aromatik (Konig 2005). Bakteri ini ditemukan di saluran pencernaan rayap Mastotermes darwinensis, R.
santonensis, dan Nasutitermes nigriceps yang mampu mendegradasi monomer lignin dan senyawa aromatik (Kuhnigk et al. 1994). Bakteri jenis ini juga mampu mendegradasi media CMC dengan potensi yang kuat yang ditemukan pada rayap Zootermopsis angusticollis (Wenzel et al. 2002).
Bakteri jenis ini ditemukan dimana-mana seperti di lingkungan dengan tekanan yang tinggi seperti di kontaminasi perminyakan hidrokarbon (Claus dan Berkeley 1986) dan rizosfer dari tanaman rawa (Daane et al. 2002). Spesis bakteri Paenibacillus pernah diisolasi dari beberapa sumber termasuk tanah, rizosfer tanaman, makanan, feses, larva yang terserang penyakit (Alexander dan Priest 1989; Claus dan Berkeley 1986; Kanzawa et al. 1995; Shida et al. 1997b; Yoon et
al. 1998).
Kocuria palustris
Isolat J5P1 merupakan isolat bakteri yang diisolasi dari saluran pecernaan bagian belakang (proktodeum) rayap pekerja dari CA Yanlappa. Isolat J5P1 mempunyai homologi dengan Kocuria palustris strain IARI-ABL-32 (No. aksesi KC581675). Isolat bakteri J5P1 mempunyai nilai query-cover, e-value, dan maximum identity sebesar 95%, 0.0, dan 96% (Tabel 11).

Deskripsi dari nama K. palustris adalah kelompok bakteri Kocuria yang ditemukan di rawa-rawa (palustris/swamphy) (Kovacs et al. 1999). Koloni bakteri K. palustris berwarna kuning, berbentuk bulat dengan pinggiran yang halus. Bakteri ini bersifat gram positif (+). Bakteri K. palustris termasuk ke dalam famili Micrococcaceae. Bakteri Kocuria sp. juga ditemukan oleh Wenzel et al. (2002) pada rayap Z. angusticollis.
Stenotromonas sp.
Isolat J5P2 merupakan isolat bakteri yang diisolasi dari saluran pencernaan belakang (proktodeum) rayap pekerja dari CA Yanlappa. Isolat J5P2 mempunyai homologi dengan Stenotrophomonas sp. S13 (No. aksesi KF542918). Isolat bakteri J5P2 mempunyai nilai query-cover, e-value, dan maximum identity sebesar 89%, 0.0, dan 94% (Tabel 11).
Bakteri Stenotrophomonas sp. merupakan famili dari Xanthomonadaceae. Bakteri ini berwarna krem kekuningan, berbentuk bulat dengan pinggiran yang halus. Bakteri ini bersifat gram negatif (-). Bakteri jenis ini mampu mendegradasi kitin (Konig 2005), tetapi tidak mempunyai aktivitas oksidase dan katalase (Rossing dan Hietpas 2007). Bakteri S. maltophilia sudah pernah diisolasi dari dalam saluran pencernaan pada siput (Gastropoda) (Rossing & Hietpas 2007).
Escherichia coli
Isolat D4M1 merupakan isolat bakteri yang diisolasi dari saluran pencernaan tengah (mesenteron) rayap pekerja dari Kampus IPB Dramaga. Isolat D4M1 mempunyai homologi dengan Escherichia coli strain ST2747 (CP007394). Isolat bakteri D4M1 mempunyai nilai query-cover, e-value, dan maximum identity
sebesar 100%, 0.0, dan 99% (Tabel 11). Bakteri E. coli pernah diisolasi dari saluran pencernaan rayap
Anacanthotermes ahngerianus (famili Hodotermitidae) (Krasil’nikov dan Satdykov 1969). Bakteri ini juga ditemukan saluran pencernaan Milipede (Konig 2005). E.
coli merupakan famili dari Enterobacteriaceae. Bakteri ini mempunyai ciri koloni berwarna putih susu, berbentuk bulat dengan pinggiran yang halus. Bakteri ini bersifat gram negatif, tidak mempunyai aktivitas kitinolitik, dan bersifat aerobik.
Kluyvera sp.
Isolat D6P1 merupakan isolat yang diisolasi dari saluran pencernaan belakang (proktodeum) rayap pekerja dari Kampus IPB Dramaga. Isolat D6P1 mempunyai homologi dengan Kluyvera sp. (No. aksesi AF176564). Isolat bakteri D6P1 mempunyai nilai query-cover, e-value, dan maximum identity sebesar 97%, 0.0, dan 98% (Tabel 11).
Kluyvera sp. termasuk famili dari Enterobacteriaceae yang bersifat gram negatif (-), berbentuk batang. Bakteri ini pernah diisolasi dari rayap Coptotermes
formosanus (Husseneder et al. 2007) dan di saluran pencernaan siput dan bekicot (Rossing dan Hietpas 2007).
Ochrobactrum sp.
Isolat D6P2 merupakan isolat yang diisolasi dari saluran pencernaan belakang (proktodeum) rayap pekerja dari Kampus IPB Dramaga. Isolat D6P2 mempunyai homologi dengan Ochrobactrum sp. QW21 (No. aksesi KF737368). Isolat bakteri
29

30
D6P2 mempunyai nilai query-cover, e-value, dan maximum identity sebesar 96%, 0.0, dan 98% (Tabel 11). Bakteri Ochrobactrum sp. termasuk ke dalam famili Rhizobiaceae yang pernah diisolasi dari saluran pencernaan rayap Z. angusticollis (Wenzel et al. 2002). Chryseobacterium sp.
Isolat D6P3 merupakan isolat bakteri yang diisolasi dari saluran pencernaan belakang (proktodeum) rayap pekerja dari Kampus IPB Dramaga. Isolat D6P3 mempunyai homologi dengan Chryseobacterium sp. MH (No. aksesi AB179867). Isolat bakteri D6P3 mempunyai nilai query-cover, e-value, dan maximum identity
sebesar 99%, 0.0, dan 99% (Tabel 11). Chryseobacterium sp. termasuk ke dalam famili Flavobacteriaceae yang
mempunyai koloni berwarna kuning dan mirip dengan ciri dari bakteri C.
kwangyangense strain Cb (No. aksesi EU169201) pada rayap Coptotermes
curvignathus (Ramin 2008). Bakteri ini juga dikelompokkan ke dalam bakteri pada rayap yang mempunyai 2 aktivitas lignoselulolitik yaitu kelompok hidrolitik and fermentatif (Konig 2006). bakteri Chryseobacterium sp. pernah diisolasi dari saluran pencernaan kecoa Amerika (Dugas et al. 2001).
Pseudomonas nitroreducens
Isolat D6P5 merupakan isolat bakteri yang diisolasi dari saluran pencernaan belakang (proktodeum) rayap pekerja dari Kampus IPB Dramaga. Isolat D6P5 mempunyai homologi dengan P. nitroreducens strain SPQ03 (No. aksesi EU500825). Hasil BLAST-N pada isolat D6P5 mempunyai query-cover, e-value, dan maximum identity sebesar 99%, 0.0, dan 98% (Tabel 11).
Pseudomonas sp. merupakan famili dari Flavobacteriaceae. Bakteri ini mempunyai ciri koloni berwarna kuning, berbentuk bulat dengan pinggiran yang halus. Bakteri ini bersifat gram negatif, mempunyai aktivitas kitinolitik, dan bersifat aerobik. Bakteri Pseudomonas sp. pernah ditemukan di saluran pencernaan rayap Z. angusticollis (Wenzel et al. 2002; Rossing & Hietpas 2007).
Isolat bakteri yang telah diidentifikasi berdasarkan morfologi, fisiologi, dan molekuler sebanyak delapan isolat. Tujuh isolat diantaranya pernah ditemukan di saluran pencernaan rayap jenis lain. Isolat P. naphthalenovorans, K. palustris,
Ochrobactrum sp. dan P. nitroreducens pernah diisolasi dari rayap Mastotermes
darwinensis, R. santonensis, Nasutitermes nigriceps, dan Z. angusticollis (Kuhnigk et al. 1994; Wenzel et al. 2002), isolat E. coli pernah ditemukan pada rayap Anacanthotermes ahngerianus (Krasil’nikov dan Satdykov 1969), isolat Kluyvera sp. pernah diisolasi dari C. formosanus (Husseneder et al. 2007) dan Chryseobacterium sp. dari rayap C. curvignathus (Ramin 2008). Isolat Stenotrophomonas sp. belum pernah ditemukan dari saluran pencernaan rayap, tetapi sudah pernah ditemukan terisolasi dari siput (Rossing dan Hietpas 2007) dan Milipede (Konig 2005) (Tabel 12).
Hal ini menunjukkan bahwa isolat bakteri yang berhasil diisolasi merupakan data terbaru. Informasi baru tersebut adalah isolat–isolat bakteri yang sebelumnya ditemukan dari saluran pencernaan rayap tingkat rendah (M. darwinensis, R.
santonensis, Z. angusticollis, C. formosanus, dan C. curvignathus) dan tingkat tinggi (N. nigriceps). Rayap M. darwinensis (F: Mastotermitidae), R. santonensis, Z. angusticollis (F: Termopsidae), C. formosanus, dan C. curvignath us (F:

Rhinotermitidae), Anacanthotermes ahngerianus (F: Hodotermitidae) merupakan kelompok rayap tingkat rendah yang didominasi dengan simbion berupa protozoa, tetapi juga ditemukan rayap di dalam saluran pencernaannya. Rayap N. nigriceps dan M. gilvus (F: Termitidae) merupakan kelompok rayap tingkat tinggi yang didominasi dengan simbion berupa bakteri (Noirot dan Noirot-Timothee 1969; Zhang dan Leadbetter 2012).
Tabel 12 Isolat bakteri yang pernah diisolasi dari saluran pencernaan rayap Isolat Ditemukan di
Kode isolat Rayap Arthropoda lain
J4M1 P.naphthalenovorans Z. angusticollis
(Wenzel et al. 2002)
M. darwinensis, R.
santonensis, dan N.
nigriceps (Kuhnigk et
al. 1994)
-
J5P1 K. palustris Z. angusticollis
(Wenzel et al. 2002)
-
J5P2 Stenotrophomonas
sp.
- siput (Gastropoda) (Rossing & Hietpas (2007)
D4M1 E. coli Anacanthotermes
ahngerianus
(Krasil’nikov dan Satdykov 1969
Milipede (Konig 2005)
D6P1 Kluyvera sp. C. formosanus
(Husseneder et al.
2007)
Siput (Gastropoda) (Rossing & Hietpas 2007)
D6P2 Ochrobactrum sp. Z. angusticollis
(Wenzel et al. 2002)
-
D6P3 Chryseobacterium sp.
C. curvignathus (Ramin 2008)
Kecoa (Dugas et al. 2001)
D6P5 P. nitroreducens Z. angusticollis (Wenzel et al. 2002)
Siput (Rossing & Hietpas 2007)
31