copyright©aquasains 2018
TRANSCRIPT



COPYRIGHT©AQUASAINS 2018
Cover Desain : Tim Editorial
Photo Properties : Coastal of Sebesi Island, South Lampung

KATA PENGANTAR
Puji Syukur Kehadirat Allah SWT karena Penyusunan Jurnal “AQUASAINS” telah
selesai. Jurnal ini disusun untuk mengapresiasi dan mempublikasi hasil-hasil
penelitian, dan kajian ilmiah bidang perikanan dan sumberdaya perairan. Untuk
mendukung tujuan tersebut, jurnal ini mengkhususkan diri dengan materi-materi
dalam bidang perikanan dan sumberdaya perairan. Edisi ketujuh Nomor satu ini ini
memuat tujuh artikel yang diharapkan akan menambah wawasan dan pemahaman
di bidang perikanan dan sumberdaya perairan.
Pada kesempatan ini redaksi menyampaikan terima kasih kepada semua pihak yang
telah mengirimkan artikelnya-artikelnya. Redaksi akan membuka kesempatan
seluas-luasnya bagi seluruh kalangangan akademisi maupun praktisi baik dari
dalam lingkungan maupun diluar Universitas Lampung untuk mempublikasikan
hasil-hasil penelitiannya.
Akhir kata semoga jurnal ilmu perikanan dan sumberdaya perairan “AQUASAINS’
ini dapat memberi manfaat yang sebesar-besarnya.
Bandar Lampung, November 2018
Redaksi


pISSN : 2301 - 816X
eISSN : 2579 - 7638
Deny Sapto Chondro Utomo,. Wardiyanto, Triando Kurniawan
Survival Rate and Larvae Development Giant Prawn
(Macrobrachium Rosebergii De Man) GImacro II at Different
Salinity …………………………………….……………………………………….
621 - 628
Heru Pramono, Taruna Fernando Putra, Sri Utari, Nova Andika
Reduction of Pathogenic Bacteria During Fermentation of Masin by
Protease and Bacteriocin-Producing Lactic Acid Bacteria …… ….
629 - 636
Prastito, Pinandoyo, Ristiawan Agung Nugroho, Vivi Endar
Herawati
The Effect of Addition Curcuma’s (Curcuma anthorrhiza Roxb)
Extract to The Increase of Feed Consumption, Efficiency and The
Growth of Catfish (Pangasius) …………………………………….
637 - 646
647 - 656
Gloria Ika Satriani, Burhanuddin Ihsan, Yulma, Rukisah
Sunscreen Cream Based on Local Raw Materials of Sonneratia
Alba from Tarakan City…….............................................................
657 – 664
Dwi Arum Mufidah, Wardiyanto, Rara Diantari
Avicennia alba Fruit Extract as a Natural Antibacterial Treatment
of Vibrio parahaemolyticus Infection in Vaname Shrimp
(Litopenaeus vannamei Boone, 1931) ……………………………….
665 - 672
Yeni Hurriyani
Suitability and Carrying Capacity of Mangrove Ecotourism in
Mempawah District, West Kalimantan ……………………………
673 - 684
DAFTAR ISI Vol 7 No. 1
Esti Harpeni, Supono, Dwi Risca Septiani
The Growth Kinetics of Bacillus Sp. D2.2 at Different pH and
Salinity…………………….............................................………...
pISSN : 2301 - 816X
eISSN : 2579 - 7638

AQUASAINS(Jurnal Ilmu Perikanan dan Sumberdaya Perairan)(Vol 7 No. 1 Tahun 2018)
SUITABILITY AND CARRYING CAPACITY OFMANGROVE ECOTOURISM IN MEMPAWAHDISTRICT, WEST KALIMANTAN
Yeni Hurriyani1
Ringkasan Mangrove forests as naturalecosystems, have high ecological andeconomic value. Rapidly growing po-pulation, causing the needs of humanlife is increasing, as a consequence ofincreased development and settlement.This will create pressure on natural re-sources, where the utilization has notyet taken into consideration any los-ses that have ecological impacts. Im-plementation of ecotourism system inmangrove forest ecosystem is an app-roach in sustainable utilization of ma-ngrove forest ecosystem. The ecosys-tem suitability index for mangrove to-urism in Village Pasir Mempawah Hi-lir Mempawah District is classified in-to very suitable (SS) and conditional(SB) categories for ecotourism develo-pment. This shows that the mangrovearea in the Village Pasir MempawahHilir Mempawah District is suitable toserve as a tourist area.
Keywords Ecoutourism, Mangrove, Carr-ying Capacity, Suitability Index
1)Program Studi Manajemen Sumberdaya Perairan,Fakultas Pertanian, Universitas Tanjungpura, Jl. Prof.Dr. H. Hadari Nawawi Pontianak 78124 KalimantanBaratE-mail: [email protected]
Received : 27 September 2018
Accepted : 30 Oktober 2018
PENDAHULUAN
Hutan Mangrove merupakan ekosistemalami yang ditemukan baik di pantaitropis maupun subtropik. Kawasan ma-ngrove didominasi oleh beberapa jenispohon mangrove yang mampu tumbuhdan berkembang baik pada daerah pasang-surut maupun pantai berlumpur. Eko-sistem mangrove mempunyai sifat yangunik dan khas, serta memiliki fungsidan manfaat yang beraneka ragam mah-luk hidup. Di Indonesia, hutan mangro-ve tumbuh dan tersebar luas diseluruhNusantara, mulai dari Sumatera, Jawa,Kalimantan, Sulawesi sampai denganPapua (Pramudji, 2004; Bengen, 2001).Vegetasi hutan mangrove umumnya di-tumbuhi tanaman mangrove dari fami-ly Rhizoporaceae, Combretaceae, Me-liaceae, Sonneratiaceae, Euphorbiace-ae dan Sterculiaceae, selanjutnya ke arahdarat umumnya ditumbuhi oleh jenispaku-pakuan (Acrostichum aureum).
Hutan mangrove sebagai ekosistem ala-miah memiliki nilai ekologis dan eko-

674 Yeni Hurriyani1
nomis penting. Fungsi penting ekolo-gis hutan mangrove antara lain sebagaitempat mencari makan (feeding groun-ds), tempat pemijahan (spawning gro-unds), dan tempat pengasuhan (nurserygrounds) bagi berbagai biota laut. Se-dangkan dari aspek nilai ekonomis, hut-an mangrove menyediakan berbagai bah-an dasar baik untuk keperluan rumahtangga maupun industri, seperti kayubakar, arang, kertas dan rayon. Ekosis-tem hutan mangrove termasuk ke da-lam tipe sistem fragile sehingga sangatrentan terhadap perubahan lingkungan.Keberadaannya yang bersifat open accesspada kawasan tertentu semakin memi-cu peningkatan eksploitasi oleh manu-sia yang pada akhirnya memberikan dam-pak negatif terhadap kualitas dan ku-antitasnya.
Pada beberapa dekade terakhir ini, pe-manfaatan hutan mangrove terus me-ningkat, bukan saja dari segi pemanfa-atan lahannya, tetapi juga segi peman-faatan tumbuhan mangrovenya, baik se-cara tradisional maupun komersial. Per-tumbuhan penduduk yang terus mening-kat menyebabkan kebutuhan hidup ma-syarakat untuk pembangunan dan pe-mukiman juga semakin meningkat. Halini tentu saja akan menimbulkan tekan-an terhadap sumberdaya alam dikare-nakan pemanfaatannya yang kurang mem-perhitungkan dampaknya terhadap ling-kungan ekologis. Demikian juga hal-nya dengan pembangunan di wilayahpesisir sekitar kawasan hutan mangro-ve yang belum dilaksanakan secara bi-jaksana dan berwawasan lingkungan.Adanya masukan beban limbah baik yangberasal rumah tangga maupun insutrike dalam kawasan mangrove juga me-yebabkan semakin menurunnya kuali-tas dan kuantitas hutan mangrove.
Usaha untuk menekan dampak kerusak-an dan melestarikan fungsi biologis ser-ta ekologis ekosistem hutan mangro-ve perlu dilakukan perlu melalui suatupendekatan yang rasional. Dimana da-lam pemanfaatannya hendaknya meli-batkan masyarakat sekitar kawasan danmasyarakat yang memanfaatkan kawas-an hutan mangrove secara langsung. Eko-sistem mangrove dengan keunikannyamerupakan sumberdaya alam yang sa-ngat potensial untuk dikembangkan se-bagai tempat kunjungan wisata. Pene-rapan sistem ekowisata pada ekosistemhutan mangrove merupakan salah satupendekatan yang dapat dilakukan da-lam pemanfaatan ekosistem hutan ma-ngrove secara lestari.
Ekowisata pertama kali diperkenalkanpada tahun 1990 oleh organisasi Theecotourisma Society. Ekowisata dide-fensikan sebagai suatu kegiatan perja-lanan ke daerah-daerah yang masih ala-mi, dimana didalamnya terdapat kegi-atan mengkonservasi lingkungan sertamemelihara kesejahteraan masyarakatsetempat (Yulianda, 2007). Ekowisatadi daerah pesisir dan laut merupakanbentuk wisata yang mengarah ke me-tatourism. Artinya, kegiatan ekowisatadi daerah pesisir dan laut tidak menjualtujuan atau objek wisata namun lebihmenjual aspek filosofi dan rasa. Berda-sarkan pertimbangan inilah, usaha eko-wisata di daerah pesisir dan laut tidakmudah mengalami kejenuhan pasar (Tu-wo, 2011).
Ekowisata merupakan kegiatan perja-lanan wisata ke daerah-daerah yang ma-sih alami yang dilakukan secara ber-tanggungjawab dengan mengutamakanaspek pendidikan, aspek konservasi alam,serta aspek pemberdayaan ekonomi ma-syarakat lokal. Kegiatan ekowisata sa-

Suitability and Carrying Capcity Mangrove Ecotourism 675
ngat erat dengan prinsip konservasi (Wo-od, 2002). Bahkan dalam strategi pe-ngembangannya, juga menggunakan stra-tegi konservasi. Dengan demikian eko-wisata sangat tepat dan berdayaguna da-lam mempertahankan keutuhan dan ke-aslian ekosistem di areal yang masihalami. Bahkan dengan ekowisata peles-tarian alam dapat ditingkatkan kuali-tasnya karena desakan dan tuntutan da-ri para eco-traveler alam. Penerapan kon-sep ekowisata di kawasan ekosistem hut-an mangrove secara umum diharapk-an dapat mengurangi tingkat perusak-an kawasan tersebut oleh masyarakatdan berpengaruh pada peningkatan eko-nomi. Dengan adanya ekowisata akanmemberikan alternatif sumber penda-patan bagi masyarakat di sekitar kawas-an tersebut dan meningkatkan Penda-patan Asli Daerah (PAD), selanjutnyaberkontribusi pada peningkatan pertum-buhan ekonomi daerah.
Pemahaman mengenai daya dukung ka-wasan perlu diperhatikan dalam upayapengembangan wilayah hutan mangro-ve sebagai kawasan ekowisata. Perhi-tungan daya dukung kawasan dimak-sudkan sebagai upaya untuk mencegahpemanfaatan sumberdaya alam secaraberlebihan yang pada akhirnya akan me-nimbulkan kerusakan ekosistem pesi-sir. Konsep daya dukung merupakan stra-tegi yang berguna untuk mencapai ta-hap akhir yang diinginkan untuk pe-ngambilan keputusan dalam manajemen.Pengelolaan hutan mangrove di kawas-an hutan mangrove Desa Pasir, Keca-matan Mempawah Hilir, Kabupaten Mem-pawah, Kalimantan Barat belum dilak-sanakan secara optimal, sehingga perlupeningkatan pengelolaan dan peman-faatan hutan mangrove dengan poten-si yang terdapat di dalamnya. Oleh ka-
rena itu, kesesuaian dan daya dukunghutan mangrove untuk kegiatan ekowi-sata perlu dianalisis sebagai acuan da-lam rangka mewujudkan pengelolaanyang berkelanjutan.
MATERI DAN METODE
Penelitian dilaksanakan di Desa Pasir,Kecamatan Mempawah Hilir, Kabupa-ten Mempawah, Kalimantan Barat (Gam-bar 1). pada bulan April-September 2017.Penentuan stasiun pengamatan meng-gunakan dilakukan menggunakan me-tode purposive sampling dengan per-timbangan berdasarkan ketebalan arealmangrove, karena ketebalan mangro-ve merupakan parameter ekologis uta-ma yang digunakan untuk menganali-sis kesesuaian kawasan mangrove se-bagai objek wisata.
Alat yang digunakan dalam penelitianini antara lain GPS, kamera digital, alattulis, meteran gulung 100 m, tali rafia,parang, dan sepatu boot. Bahan yangdigunakan dalam penelitian yaitu da-ta sheet, daftar pertanyaan (kuisioner)dan buku identifikasi mangrove berupaBuku Panduan Pengenalan Mangrovedi Indonesia (Noor et al., 1999).
Jenis data dalam penelitian ini terdi-ri dari data primer dan data sekunder.Data primer yang dikumpulkan anta-ra lain berupa data pengamatan eko-sistem mangrove dan parameter sosialekonomi berupa data pemahaman danpersepsi responden tentang ekowisatamangrove. Sedangkan data sekunder yangdikumpulkan berupa parameter fisik ya-itu data gambaran lokasi penelitian. Pe-ngambilan sampel ekosistem mangro-ve dilakukan dengan menggunakan me-tode petak ganda. Sampling mangro-

676 Yeni Hurriyani1
Gambar 1 Peta lokasi penelitian
ve dilakukan dengan menggunakan ba-nyak petak contoh yang peletakannyadilakukan secara acak (simple randomsampling).
Jumlah responden (masyarakat dan pe-ngunjung) yang ditetapkan masing-masingadalah sebanyak 30 orang. Penentuanjumlah responden masyarakat dilakuk-an dengan metode accidential sampling,dimana jumlah responden tidak dapatditentukan dan tergantung dari sebera-pa banyak masyarakat Desa Pasir yangdapat ditemui dan diwawancarai olehpeneliti pada saat pengambilan data. Un-tuk penentuan jumlah responden pengun-jung, metode accidential sampling di-lakukan karena responden yang diwa-wancarai tergantung dari jumlah pengun-jung yang berkunjung ke kawasan ma-ngrove Desa Pasir saat penelitian di-laksanakan. Metode penentuan respon-den menggunakan metode purposive sam-pling, yaitu pengambilan sampel yangdilakukan secara sengaja atau tidak se-cara acak melainkan berdasarkan per-timbangan tertentu. Dalam hal ini yang
menjadi pertimbangan sebagai respon-den masyarakat adalah warga desa yangbertempat tinggal di sekitar lokasi pe-nelitian. Pertimbangan yang diambil un-tuk responden pengunjung adalah orang-orang yang mengunjungi kawasan ma-ngrove Desa Pasir baik yang berasaldari luar desa maupun dari luar Kabu-paten Mempawah, Kalimantan Barat.
Data yang dikumpulkan meliputi: da-ta mengenai jenis spesies, jumlah in-dividu dan diameter pohon. Data-datatersebut kemudian diolah untuk meng-etahui kerapatan setiap spesies dan ke-rapatan total semua spesies. Kerapatanspesies, Kerapatan total adalah jumlahindividu spesies i dalam suatu unit areayangdinyatakan dalam rumus:
K =ni
A(1)
KT =∑nA
(2)
dimana, K = kerapatan jenis i; ni = jum-lah total individu dari jenis i; KT = ke-rapatan total; Σn = jumlah total tegak-an seluruh jenis;dan A = luas total area

Suitability and Carrying Capcity Mangrove Ecotourism 677
pengambilan contoh (luas petak con-toh)
Kegiatan wisata yang akan dikembangk-an hendaknya disesuaikan dengan po-tensi sumberdaya dan peruntukannya.Setiap kegiatan wisata mempunyai per-syaratan sumberdaya dan lingkunganyang sesuai dengan objek wisata yangakan dikembangkan. Kategori wisata ma-ngrove mempertimbangkan 5 parame-ter dengan 4 klasifikasi penilaian (Ta-bel 1) (Yulianda, 2007).
IKW = ∑
(Ni
Nmax
)x100% (3)
dimana, IKW = Indeks kesesuaian wi-sata mangrove (sesuai: 83% - 100%;sesuai bersyarat: 50% - < 83%; tidaksesuai: < 50) Ni = Nilai parameter ke-i(Bobot x Skor). Nmaks = Penjumlah-an dari hasil pengalian nilai bobot de-ngan nilai skor dari kategori baik padamasing-masing parameter ekologi ma-ngrove.
Daya Dukung Kawasan merupakan jum-lah maksimum pengunjung yang dapatditampung pada suatu kawasan pada wak-tu tertentu tanpa menimbulkan gang-guan baik pada alam maupun manu-sia. Perhitungan Daya Dukung Kawas-an (DDK) (Yulianda, 2007).
DK = k xLp
Ltx
Wt
Wp(4)
dimana, DK : Daya Dukung Kawasan;k : Potensi ekologis pengunjung per sa-tuan unit area (orang); Lp : Luas areaatau panjang area yang dapat dimanfa-atkan (m atau m2); Lt : Unit area un-tuk kategori tertentu (m atau m2) Wt :Waktu yang disediakan oleh kawasanuntuk kegiatan wisata dalam satu hari(jam); dan Wp : Waktu yang dihabisk-an oleh pengunjung untuk setiap kegi-atan tertentu (jam)
HASIL DAN PEMBAHASAN
Kabupaten Mempawah merupakan sa-lah satu kabupaten yang ada PropinsiKalimantan Barat yang memiliki eko-sistem mangrove terbesar kedua sete-lah Kabupaten Kubu Raya. Secara ge-ografis, kabupaten Mempawah terletakdiantara 0°44’ Lintang Utara dan 0°00,4’Lintang Selatan serta 108°24’ Bujur Ti-mur dan 109°21,5’ Bujur Timur. Luaswilayah Kabupaten Mempawah (Luasdaratan dan perairan) menurut Permen-Dagri No. 06 Tahun 2008 tentang Ko-de Dan Data Wilayah Administrasi Pe-merintahan 2.797,88 km² atau sekitar1,90 % dari luas wilayah Provinsi Ka-limantan Barat, 146.807 km². Secaraadministratif, Kabupaten Mempawah ber-batasan dengan:
1. Sebelah Utara : berbatasan denganKabupaten Bengkayang
2. Sebelah Timur : berbatasan denganKab. Landak
3. Sebelah Selatan : berbatasan denganKabupaten Kubu Raya dan Kota Pon-tianak
4. Sebelah Barat : berbatasan denganLaut Natuna.
Kabupaten Mempawah terdiri dari sem-bilan kecamatan yaitu Siantan, Sege-dong, Sungai Pinyuh, Anjongan, Mem-pawah Hilir, Mempawah Timur, SungaiKunyit, Toho dan Sadaniang. Enam di-antaranya yaitu Kecamatan MempawahHilir, Mempawah Timur, Sungai Ku-nyit, Sungai Pinyuh, Segedong dan Si-antan merupakan kecamatan pesisir yangdisebelah barat berbatasan langsung de-ngan Laut Natuna.
Desa Pasir secara administratif terletakdi Kecamatan Mempawah Hilir, Kabu-paten Mempawah, Provinsi Kalimant-

678 Yeni Hurriyani1
Tabel 1 Matriks kesesuaian lahan untuk wisata pantai kategori wisata mangrove
Parameter BKategori
SKategori
SKategori Cukup
SKategori
Baik Cukup baik buruk Buruk
Ketebalan mangrove (m) 5 >500 3 >200 - 500 2 50 - 200 1 <50
Kerapatan mangrove (100 m2) 3 > 15 - 25 3 > 10 - 5 2 10 - 5 1 <5
Jenis mangrove 3 > 5 3 5 - 3 2 2 - 1 1 0
Pasang surut (m) 1 0 – 1 3 >1 - 2 2 >2 - 5 1 >5
Objek biota 1 Ikan, udang, kepiting, moluska, reptile, burung 3 Ikan, udang, kepiting, moluska 2 Ikan, moluska 1 Salah satu biota air
an Barat. Secara geografis, Desa Pasirberada pada 00o 20
′00
′′Lintang Uta-
ra sampai dengan 00o 30′′00
′′Lintang
Utara dan 108o 55′00
′′Bujur Timur sam-
pai dengan 109o 06′40
′′Bujur Timur
dengan luas wilayah kurang lebih 60Ha. Adapun batas-batas wilayah yangmelingkupi Desa Pasir adalah sebagaiberikut :
1. Sebelah Utara : berbatasan denganDesa Penibung
2. Sebelah Timur : berbatasan denganDesa Sekabuk
3. Sebelah Selatan : berbatasan denganKelurahan Terusan
4. Sebelah Barat : berbatasan denganLaut Natuna .
Saat ini ekosistem mangrove di Kabu-paten Mempawah (Tabel 2) semakin ter-desak sebagai akibat meningkatnya ke-butuhan penggunaan lahan untuk me-menuhi kebutuhan manusia, sehinggamengancam kelestariannya. Degradasimangrove di Kabupaten Mempawah di-sebabkan kurangnya mendapat perha-tian dalam perencanaan dan pengelo-laan, kurangnya kesepahaman dianta-ra para aktor yang terlibat merupakanhal yang mempercepat kerusakan padaekosistem mangrove.
Pasang surut merupakan gerakan ver-tikal seluruh partikel massa air laut da-ri permukaan sampai bagian terdalamdasar laut yang disebabkan oleh penga-ruh gaya tarik menarik antara bumi de-
Tabel 2 Perbandingan luas ekosistem mangrove diKabupaten Mempawah
Kecamatan Luas wilayah (ha)Luas kawasan mangrove (ha)
1989 2014
Mempawah Hilir 16.042,53 184,75 186,16
Mempawah Timur 11.253,87 30,5 21,6
Sungai Kunyit 19.487,1 90,91 147,38
Sungai Pinyuh 18.264,93 509,89 384,17
Total 65.048,43 816,05 739,31
ngan benda-benda angkasa terutama ma-tahari dan bulan (Wibisono, 2005). Ki-saran pasang surut di lokasi antara 0.6sampai dengan 1.2 meter. Pola arus pa-sang surut tipe ini merupakan meka-nisme penting dalam distribusi dan trans-portasi sedimen tersuspensi di sepan-jang dasar perairan baik di alur mau-pun pantai pesisir Kabupaten Mempa-wah sehingga sangat berpengaruh padahabitat mangrove. Tipe pasut ini jugaakan memberikan indikasi ke arah ma-na kecenderungan terjadinya sedimen-tasi/pendangkalan dimana pada perair-an yang mempunyai aruslambat dan te-nang akan memberikan kesempatan ke-pada material tersuspensi untuk meng-endap sehingga kawasan ini lebih do-minan ditumbuhi jenis Rhizophora sp.,sebaliknya pada arus yang cepat me-nyebabkan material tersuspensi akan te-tap bergerak bersama arus dan pada ka-wasan ini spesies mangrove yang do-minan adalah jenis Avicennia sp.
Ketebalan mangrove diperoleh berda-sarkan hasil pengukuran secara manu-al. Pengukuran dilakukan denggunak-an roll meter yang ditarik secara tegak

Suitability and Carrying Capcity Mangrove Ecotourism 679
lurus terhadap garis pantai mulai da-ri tumbuhan mangrove di bagian daratsampai dengan ujung tumbuhan mangro-ve di hidup pada batas laut. Hasil pe-ngukuran menunjukkan bahwa ketebal-an Mangrove pada Stasiun 1, 2, dan 3berturut-turut adalah 98 m, 93 m, dan90 m dengan rata-rata 93,7 m.
Secara umum ekosistem mangrove disepanjang pantai Desa Pasir Kecamat-an Mempawah Hilir Kabupaten Mem-pawah dapat dibagi ke dalam zona dep-an (dekat perairan), selanjutnya zonatengah, kemudian zona akhir (dekat da-ratan). Hasil pengamatan jenis mangro-ve (Gambar 2) di lokasi menunjukk-an bahwa terdapat beberapa jenis ma-ngrove antara lain Avicennia marina,Avicennia lanata, Rhizopora mucrona-ta, Rhizopora stylosa, Rhizopora api-culata, Sonneratia ovata, Bruguiera gym-noriza, Bruguiera cilindrica, Kandeliacandle, Excoecaria agallocha, Cerio-ps decandra, Xylocarpus garantum ,danNypa fruticans. Dari ketiga stasiun ter-sebut, jenis spesies mangrove paling ba-nyak ditemukan pada stasiun 1 diban-ding stasiun 2 dan 3. Hal ini disebabk-an karena perbedaan kondisi lingkung-an seperti substrat, pengenangan air, sa-linitas, serta intensitas cahaya.
Hasil pengamatan di lapangan menun-jukkan bahwa jenis A. marina, A. la-nata dan R. Mucronata merupakan je-nis yang paling mendominasi kawas-an mangrove Desa Pasir. Hal ini dika-renakan lokasi penelitian yang beradadi tepi pantai dengan kondisi substratberpasir. Jenis R. Mucronata diketahuimempunyai daya toleran yang baik un-tuk tumbuh pada substrat yang lebihkasar dan berpasir. Kerapatan jenis ve-getasi mangrove kategori pohon padastasiun 1 sebesar 960 ind/ha dengan ke-
Gambar 2 Jenis Mangrove Di Kawasan MangroveDesa Pasir
rapatan jenis tertinggi pada jenis R. Mu-cronata (840 ind/ha), dan kerapatan je-nis terendah pada jenis A. marina (120ind/ha). Stasiun 2 memiliki kerapatanjenis vegetasi mangrove sebesar 1420ind/ha, dengan kerapatan jenis terting-gi dijumpai pada jenis A. marina (1120ind/ha) serta kerapatan jenis terendahpada jenis R. Mucronata (140 ind/ha).Pada stasiun 3 kerapatan vegetasi ma-ngrove kategori pohon sebesar 960 ind/ha,dengan kerapatan jenis tertinggi dijum-pai pada jenis R. Mucronata (780 ind/ha).
Mangrove mempunyai fungsi ekologispenting sebagai habitat berbagai jenisfauna. Fauna yang terdapat pada eko-sistem mangrove merupakan perpadu-an antara fauna terrestrial dan fauna aku-atik. Adapun fauna terestrial umumnyamenepati bagian atas pohon mangro-ve dan tidak mempunyai sifat adapa-tasi khusus di dalam mangrove sepertiinsekta, ular, primata, serta burung. Se-dangkan kelompok fauna akuatik yanghidup pada ekosistem mangrove terdiridari dua tipe, yaitu: (a) kelompok orga-nisme yang hidup pada kolom air ter-utama dari jenis ikan dan udang (b) ke-

680 Yeni Hurriyani1
lompok organisme yang hidup menem-pati baik pada substrat keras seperti akardan batang maupun substrat lunak se-perti lumpur. Fauna yang hidup di ha-bitat mangrove memainkan peran pen-ting dalam fungsi ekosistem sehinggadapat menjadi indikator yang bergunabagi kawasan mangrove (Bengen, 2002).
Zonasi dan distribusi spesies mangro-ve berhubungan dengan amplitude pa-sang surut. Pada zona intertidal ting-gi, krustasea berkaki sepuluh mendo-minasi, sedangkan pada zona intertidalrendah, fauna yang ada terdiri dari filter-feeders, seperti tiram dan teritip. Zona-si spesies juga tergantung pada topo-grafi pantai, variasi suhu dan distribusibahan organik, yang merupakan sum-ber makanan penting.
Hasil identifikasi kelompok fauna (Gam-bar 3) terrestrial yang ditemukan padaekosistem mangrove Desa Pasir Keca-matan Mempawah Hilir antara lain :(a) jenis burung seperti Burung Encit-encitan (Orthotomus sp.), Burung La-yang (Hirundo sp.), Burung Pipit (Lon-chura sp.) dan (b) jenis reptil sepertibiawak (Varanussalvator), ular Bakau(Chrysopelea sp), Cacing nypa (Nama-lycatis sp.). Sedangkan kelompok fau-na akuatik yang ditemukan antara la-in: (a) jenis ikan seperti ikan Temba-kol (Periopthalmus sp.), Belanak (Mu-gil sp.), (b) jenis krustasea seperti Udangputih (Penaeus marguensis) Udang kro-sok (P. semiculcatus), kepiting bakau(Scylla serrata), serta (c) jenis molus-ka seperti Kerang Bakau (Polymesodasp. ), Kerang Darah (Anadara grano-sa), Kerang Bulu (Anadra Antiquata),dan Siput mangrove (Littorina, sp).
Aksesibilitas merupakan salah satu fak-tor penting dalam mendukung keber-hasilan pengembangan suatu kawasan
Gambar 3 Jenis Fauna Di Kawasan Mangrove DesaPasir
ekowisata. Aksesibilitas yang baik da-pat dinilai ketersediaan sarana dan pra-sarana yang seperti jalan maupun ken-daraan yang layak menuju lokasi eko-wisata. Akses menuju kawasan ekowi-sata mangrove di Desa Pasir, Kecamat-an Mempawah Hilir, Kabupaten Mem-pawah dapat ditempuh menggunakankendaraan pribadi atau umum, baik da-ri jenis kendaraan roda dua ataupun ro-da empat. Lokasi ekowisata yang bera-da pada jalur utama jalan provinsi yangberaspal baik dan cukup lebar sangatmemudahkan pengunjung untuk men-capai lokasi ekowisata. Selain itu, ak-ses jalan menuju lokasi ekowisata ma-ngrove juga sudah di semen sehinggasangat memudahkan pengunjung. sedangk-an untuk sarana pendukung yang ter-dapat di lokasi ekowisata yaitu tersedi-anya dermaga perahu dan lokasi parkiryang dekat dengan lokasi dan sudah di-jaga oleh petugas parkir.
Pemahaman masyarakat mengenai eko-sistem mangrove dapat dikatakan cu-kup baik. Sebagian besar masyarakatsudah mengetahui peran dan fungsi eko-sistem mangrove, namun disisi lain ju-

Suitability and Carrying Capcity Mangrove Ecotourism 681
ga masih ditemukan kelompok masya-rakat yang belum mengetahui peran pen-ting dari ekosistem mangrove ini. Ke-rusakan mangrove di Kabupaten Mem-pawah lebih banyak disebabkan olehkegiatan perkebunan warga, pembangunatau pengembang untuk membuka lah-an perumahan dan masyarakat yang me-nebang mangrove untuk kayu dan arang.Hal ini merupakan konsekuensi bertam-bahnya jumlah penduduk, dimana lah-an yang ditumbuhi mangrove akan te-rus mendapat tekanan untuk dikonver-si menjadi lahan pertanian dan perke-bunan bahkan pemukiman. Potensi dankegunaan ekosistem mangrove yang be-gitu luas menyebabkan pengelolaannyamenjadi rumit karena akan mendapatk-an benturan kepentingan antar sektorbaik masyarakat bahkan antar lembagapemerintah.
Hasil penelitian menunjukkan pemaham-an masyarakat sekitar pesisir Desa Pa-sir, Kecamatan Mempawah Hilir, Ka-bupaten Mempawah terhadap kegiatanekowisata sudah baik. Pemahaman ma-syarakat ini sangat penting terkait pe-ngembangan kawasan sebagai lokasi eko-wisata. Suatu kawasan yang akan di-kembangkan menjadi lokasi ekowisa-ta, maka perlu diadakan sosialisasi pro-gram atau penyuluhan konservasi se-cara kontinyu kepada masyarakat seki-tar. Hal ini dilakukan agar masyarakatsekitar mengetahui dan dapat berpar-tisipasi dalam kegiatan pengembangankawasan ekowisata yang akan dilaksa-nakan. Selain itu, dengan adanya ke-giatan sosialisasi dan penyuluhan yangkontinu dapat meningkatkan pemaham-an masyarakat sekitar mengenai kon-sep konservasi mangrove.
Hal yang sama juga didapatkan padapengunjung, dimana mayoritas pengun-
jung sudah mengeal istilah ekowisata.Pemahaman pengunjung tentang eko-sistem mangrove dan ekowisata dapatdikatakan sudah cukup baik. Melaluikegiatan ekowisata diharapkan pema-haman pengunjung tentang ekosistemmangrove dapat meningkat. Pengunjungmenyatakan bahwa kondisi mangrovedi lokasi ekowisata berada dalam kea-daan baik.
Salah satu prinsip dasar dan tujuan daripengembangan kegiatan ekowisata ada-lah untuk meningkatkan kesejahteraanmasyarakat lokal. Keterlibatan masya-rakat lokal dalam kegiatan ekowisatasangat penting, mengingat peran sertamereka sebagai penyedia fasilitas, me-nyajikan atraksi budaya, sekaligus me-nentukan kualitas produk wisata. Ber-dasarkan hasil wawancara dengan ma-syarakat setempat (Gambar 4), sebanyak60% responden berkeinginan untuk ter-libat dalam kegiatan ekowisata, 30% res-ponden menyatakan tidak tahu dan ha-nya 10 % yang menyatakan tidak inginterlibat. Masyarakat yang menyatakankeinginan untuk terlibat dalam kegiat-an ekowisata ini bersedia menjadi pe-mandu wisata (guide), menyewakan ru-mahnya untuk dijadikan penginapan (gu-est house) dan sisanya menyatakan ber-keinginan untuk menjadi relawan (vo-lunteer).
Keberhasilan didalam pengelolaan eko-sistem mangrove sangat ditentukan olehketepatan strategi dalam melaksanak-an kebijakan yang dikembangkan de-ngan melibatkan dan memperhatikan ke-pentingan berbagai pihak (stakeholders)yang meliputi pemerintah baik pusat ma-upun daerah, nelayan, LSM, tokoh ma-syarakat dan peneliti di perguruan ting-gi. Hal ini sangat penting diperhatikanagar kebijakan yang dihasilkan dapat

682 Yeni Hurriyani1
Gambar 4 Keterlibatan masyarakat dalam kegiatanekowisata mangrove
diimplementasikan dengan baik kare-na telah disesuaikan dengan potensi, as-pirasi dan bersama masyarakat dan yangpaling terpenting adalah bagaimana me-lalui kebijakan ini dapat meningkatk-an kesejahteraan masyarakat di wila-yah pesisir khususnya.
Analisis daya dukung kawasan diper-lukan dalam pemanfaatan potensi sum-berdaya pesisir untuk pengembanganwisata bahari secara lestari. Mengingattingkat kerentanan dan keterbatasan ru-ang untuk pengunjung maka perlu di-tentukan daya dukung kawasan. Meto-de yang digunakan untuk menghitungdaya dukung pengembangan ekowisa-ta alam dikenal dengan konsep daya du-kung kawasan (DDK). Dengan adanyakonsep daya dukung ini diharapkan mam-pu meminimalkan atau mencegah ke-rusakan sumberdaya alam dan lingkung-an dari usaha pemanfaatan yang dila-kukan.
Usaha pengelolaan sumberdaya alamdan lingkungan secara lestari dapat ter-laksana dengan tetap memperhatikan ke-sejahteraan masyarakat pengguna sum-berdaya. Daya dukung kawasan diper-lukan dalam pengelolaan ekowisata ma-
ngrove agar tidak mengganggu potensiekologis ekosistem mangrove sehing-ga kelestariannya tetap terjaga dan ke-giatan ekowisata dapat berjalan seca-ra terus menerus tanpa merusak ling-kungan.
Nilai daya dukung kawasan mangroveDesa Pasir untuk kegiatan tracking ya-itu sebesar 44 orang/hari dengan pan-jang area yang dimanfaatkan sebesar550 meter. Kondisi nyata di lapangandimana waktu yang disediakan oleh lo-kasi wisata hutan mangrove adalah 12jam (waktu buka 06.00 dan waktu tu-tup 18.00) dengan rerata waktu kun-jungan setiap orang adalah 2 jam ma-ka perhitungan daya dukung kawasanuntuk wisatanya adalah 66 orang/hari.Kegiatan tracking yang dapat dilakuk-an oleh pengunjung antara lain untukmenikmati keindahan ekosistem mangro-ve dan juga dapat digunakan sebagaiwisata pendidikan (educational tour) meng-gunakan jembatan kayu (wooden tra-il). Selain itu, kegiatan penanaman bi-bit mangrove di lokasi wisata juga da-pat dilakukan sebagai sarana pendidik-an konservasi.
Eksplorasi potensi dan nilai kawasanmerupakan prioritas utama dalam pe-ngelolaan ekowisata kawasan mangro-ve. Tujuan dari kegiatan ini adalah un-tuk mengetahui seberapa besar poten-si dan nilai tersebut sehingga dapat di-manfaatkan untuk meningkatkan pen-dapatan daerah yang berdasarkan prinsip-prinsip keadilan dan kemandirian se-hingga pada akhirnya dapat meningkatk-an kesejahteraan masyarakat sekitar.
Keanekaragaman flora dan fauna me-rupakan modal dasar dalam pengem-bangan kegiatan ekowisata. Semakin ba-nyak potensi wisata alam yang ada pa-da suatu kawasan akan semakin me-

Suitability and Carrying Capcity Mangrove Ecotourism 683
narik minat wisatawan untuk berkun-jung. Kegiatan ekowisata yang akan di-kembangkan harus disesuaikan denganpotensi sumberdaya dan peruntukannya.Indeks kesesuaian ekologis dapat digu-nakan untuk mengidentifikasikan apa-kah suatu ekosistem Sangat Sesuai (SS),Sesuai (S), Sesuai Bersyarat (SB), atauTidak Sesuai (N) untuk pengembangansuatu kegiatan wisata. Secara umum,kesesuaian pengembangan usaha eko-wisata mangrove dilaksanakan denganmempertimbangkan 5 parameter dengan4 klasifikasi penilaian. Parameter - pa-rameter tersebut antara lain meliputi obyekbiota, pasag surut air laut, ketebalan,kerapatan, serta jenis mangrove, pasangsurut dan obyek biota. Nilai parameter-parameter ini didapatkan melalui hasilpengukuran dan pengamatan langsungdi lapangan.
Berdasarkan hasil perhitungan yang te-lah dilakukan diperoleh nilai indeks ke-sesuaian wisata (IKW) pada Stasiun 1menunjukkan nilai total skor 26 dariskor maksimum 39 dan nilai IKW nyaadalah 66,67% dengan kategori SesuaiBersyarat (SB). Selanjutnya pada sta-siun 2 didapatkan total skor 25 dari skormaksimum 39 dan nilai IKW nya ada-lah 64,10% dengan kategori Sesuai Ber-syarat (SB). Pada stasiun 3 didapatkantotal skor 25 dari skor maksimum 39dan nilai IKW nya adalah 64,10% de-ngan kategori Sesuai Bersyarat (SB).Perbedaan nilai indeks ini dikarenakanperbedaan kondisi ekologis pada seti-ap stasiun seperti ketebalan dan jenismangrove yang mempengaruhi keane-karagaman biota di lingkungan terse-but.
Secara keseluruhan nilai indeks kese-suaian wisata (IKW) pada kawasan hut-an mangrove Desa Pasir, Kabupaten Mem-
pawah berada pada kategori Sesuai Ber-syarat (SB). Hal ini menunjukkan bah-wa perlu dilakukan tindakan pengelo-laan lebih lanjut dan intensif untuk men-jadikan hutan mangrove tersebut seba-gai kawasan ekowisata seperti melakuk-an kegiatan penanaman bibit mangro-ve (replantasi) sehingga ketebalan are-al mangrove bertambah, selanjutnya meng-embangkan potensi rekreasi seperti ke-giatan lintas alam, memancing, berla-yar, pengamatan jenis burung dan atrak-si satwa liar, fotografi, pendidikan, pe-nelitian, piknik dan berkemah.
SIMPULAN
Indeks kesesuaian ekosistem mangro-ve di Desa Pasir Kecamatan Mempa-wah Hilir Kabupaten Mempawah un-tuk kegiatan wisata tergolong ke dalamkategori Sesuai Bersyarat (SB) untukdikembangkan sebagai kawasan ekowi-sata. Hal ini menunjukan bahwa kawas-an mangrove di Desa Pasir mempunyaipotensi untuk dikembangkan sebagai ka-wasan ekowisata.
Acknowledgements Universitas Tanjungpu-ra atas bantuan pembiyaan penelitianmelalui Dana DIPA Untan tahun 2017dan semua pihak yang telah membantukelancaran penelitian ini.
Pustaka
Bengen, D. G. (2001). Pengenalandan pengelolaan ekosistem mangro-ve: pedoman teknis. Pusat KajianSumberdaya Pesisir dan Lautan, In-stitut Pertanian Bogor.
Bengen, D. G. (2002). Sinopsis eko-sistem dan sumberdaya alam pesisir

684 Yeni Hurriyani1
dan laut serta prinsip pengelolaan-nya. Pusat Kajian Sumberdaya Pe-sisir dan Lautan IPB: Bogor, 63.
Noor, Y. R., Khazali, M., and I NN,S. (1999). Panduan pengenalanmangrove di Indonesia. PKA/WI-IP(Wetlands International-IndonesiaProgramme).
Pramudji (2004). Penanganan hutanmangrove di kawasan pesisir indo-nesia: Suatu program yang sangatmendesak. Oseana, 1:19–26.
Tuwo, A. (2011). Pengelolaan eko-wisata pesisir dan laut: pendekatanekologi, sosial-ekonomi, kelembaga-an, dan sarana wilayah. Brilian in-ternasional.
Wibisono, M. (2005). Pengantar ilmukelautan. Grasindo. Jakarta, 226.
Wood, M. (2002). Ecotourism: Princi-ples, practices and policies for sus-tainability. UNEP.
Yulianda, F. (2007). Ekowisata baharisebagai alternatif pemanfaatan sum-ber daya pesisir berbasis konservasi.makalah disampaikan pada seminarsains 21 februari 2007. DepartemenMSP. FPIK. IPB. Bogor, 19.

AQUASAINS(Jurnal Ilmu Perikanan dan Sumberdaya Perairan)(Vol 7 No. 1 Tahun 2018)
DEVELOPMENT AND SURVIVAL RATE OF GIANTSHRIMP LARVA (Macrobrachium rosebergii de Man)GIMacro II AT DIFFERENT SALINITIES
Deny Sapto Chondro Utomo1 · Wardiyanto1 · TriandoKurniawan1
Ringkasan GIMacro II prawn larvae isable to grow well at a salinity of 8-15 ppt. This condition can be impro-ved by improving the methods of adap-tation to changes in salinity prawn la-rvae production activities, by determi-ning the pattern of changes in salini-ty are right. Salinity media through os-motic pressure affect the physiologicalactivity, where the cells in body organsprawns should be in liquid media withionic composition and concentration ofthe same with the environment. Havingobtained the optimum salinity on la-rval rearing prawns GIMacro II in di-fferent salinity media is expected to pro-duce a population of prawns GIMacroII with superior durability specific toenvironmental conditions, which can thenbe directed to improve the utilization ofmarine resources to the salinity of thebest. This research aims were to studythe growth and survival of larvae pra-wns GIMacro II reared on media of di-fferent salinities. The study used com-pletely randomized design with threetreatments and three replications. Thetreatments were larval rearing prawns
1)Department of Aquaculture University of LampungE-mail: [email protected]
GIMacro II at different salinities (10ppt, 12 ppt, and 14 ppt). The resultsshowed that the culture of prawn la-rvae GIMacro II at different salinity affe-ct significantly on growth and survivalof larvae prawns GIMacro II. The hi-ghest development of GIMacro II pra-wn larvae obtained from 12 ppt sali-nity treatment (7.13 ± 0.03%) and thehighest survival rate was obtained alsofrom 12 ppt salinity treatment (67.67 ±4.51%)
Keywords Macrobrachium rosebergii,larvae, salinity
Received : 07 Juli 2018
Accepted : 13 Agustus 2018
PENDAHULUAN
Udang Galah (Macrobrachium rosenber-gii de Man) merupakan salah satu ko-moditas perikanan yang bernilai eko-nomis tinggi. Selain mempunyai ukur-an terbesar diban- dingkan dengan udangair tawar lainnya, udang galah juga sa-ngat di- gemari konsumen baik di da-lam maupun di luar negeri terutama diJepang dan beberapa Negara Eropa (Pri-yono et al., 2011).

622 Deny Sapto Chondro Utomo1 et al.
Dalam kegiatan budidaya udang galah,faktor kualitas air, termasuk di dalam-nya salinitas media, merupakan faktoryang sangat mempengaruhi kehidupanudang galah. Salinitas media, melaluitekanan osmotiknya, mempengaruhi ak-tivitas fisiologis, baik pada osmoregu-lasi maupun bioenergetik. Apabila te-kanan osmotik media (salinitas) berbe-da jauh dengan tekanan osmotik cairantubuh (kondisi tidak ideal) maka per-bedaan tekanan osmotik akan menjadibeban bagi udang sehingga dibutuhkanenergi yang relatif besar untuk mem-pertahankan osmotik tubuhnya agar te-tap pada keadaan yang ideal.
Salinitas termasuk ke dalam kelompokmasking factor yaitu faktor-faktor yangdapat memodifikasi pengaruh faktor ling-kungan lain menjadi satu kesatuan pe-ngaruh osmotik melalui suatu mekanis-me pengaturan tubuh organisme. Sali-nitas media melalui tekanan osmotik-nya mempengaruhi aktivitas fisiologis,salah satunya osmoregulasi, dimana sel-sel pada organ tubuh harus berada da-lam cairan media dengan konsentrasiionik yang sama dengan lingkungan-nya. Oleh karena itu diperlukan penga-turan (osmoregulasi) agar tercipta kon-sentrasi ionik cairan dalam sel dengancairan luar sel yang hampir sama (Nu-grahaningsih, 2008). Oleh karena itu,salinitas media akan mempengaruhi peng-gunaan energi untuk osmoregulasi (Aliand Waluyo, 2015).
Penggunaan energi yang besar dalamproses osmoregulasi pada udang galahmenyebabkan terhambatnya perkembang-an dan tingginya mortalitas udang ga-lah. Sehingga perlu dilakukan peneli-tian untuk mencari pola-pola perubah-an kebutuhan salinitas dari penetasanhingga post larva, untuk mendapatkan
salinitas optimum yang menghasilkankelangsungan hidup dan pertumbuhanlarva udang galah GIMacro II yang ter-baik.
MATERI DAN METODE
Penelitian ini dilaksanakan pada bulanOktober - November 2016 bertempatdi Balai Penelitan dan Pemuliaan Ikan,Sukamandi, Subang, Jawa Barat. Alat-alat yang digunakan antara lain: bak fi-ber kerucut, akuarium ukuran 0,3x0,3x0,3m, blower, refraktometer, termometer,DO meter, pH meter, timbangan digi-tal, plankton net, alat tulis, mangkokplastik, penggaris, kertas label. Sedangk-an bahan yang digunakan adalah larvaudang galah GIMacro II, air tawar, danair laut.
Rancangan yang digunakan dalam pe-nelitian ini adalah rancangan acak leng-kap (RAL) yang terdiri atas 3 perla-kuan dengan 3 kali ulangan. Perlakuanyang digunakan adalah sebagai berikut:
1. Perlakuan A = Pemeliharaan larvaudang galah GIMacro II dengan sa-linitas 10 ppt.
2. Perlakuan B = Pemeliharaan larvaudang galah GIMacro II dengan sa-linitas 12 ppt.
3. Perlakuan C = Pemeliharaan larvaudang galah GIMacro II dengan sa-linitas 14 ppt.
Larva udang galah yang didapatkan da-ri hasil pemijahan dipindahkan ke masing-masing media pemeliharaan sesuai de-ngan perlakuan yang diberikan. Masing-masing ulangan diisi udang galah se-banyak 300 ekor. Selama 21 hari pe-meliharaan, larva udang galah diberik-an pakan Artemia sp. dengan frekuensi

Development and survival rate giant prawn at different salinities 623
2 kali sehari sebanyak 20 – 80 indivi-du/larva/hari.
Parameter yang diamati adalah kelang-sungan hidup, laju perkembangan la-rva, dan kualitas air. Parameter kelang-sungan hidup dan laju perkembanganlarva dianalisis menggunakan uji Ano-va dengan tingkat kepercayaan 95% danjika terdapat perbedaan nyata antar per-lakuan, diuji lanjut menggunakan ujiDuncan.
HASIL DAN PEMBAHASAN
Tahap perkembangan larva diamati se-telah larva berumur 1 hari dan dima-sukkan ke dalam wadah pemeliharaan.Perkembangan larva udang galah da-pat dilihat pada Gambar 1. Perkembang-an tertinggi terjadi pada pelakuan B ya-itu sebesar 7,13 ± 0,03 %, kemudiandiikuti dengan perlakuan A sebesar 6,97± 0,06 % dan perlakuan C sebesar 6,86± 0,06 % (Gambar 2).
Berdasarkan analisis statistik (Anova)dengan tingkat kepercayaan 95%, per-bedaan salinitas berpengaruh nyata (P<0,05)terhadap perkembangan larva udang ga-lah GIMacro II stadia 1 hingga post la-rvae. Rata-rata perkembangan perlaku-an B dengan nilai osmolaritas pada me-dia sebesar 323 mmol/kg lebih baik di-bandingkan perlakuan A dengan nilaiosmolaritas yang ada pada media se-besar 311 mmol/kg dan C dengan nilaiosmolaritas media sebesar 335 mmol/kg.Ali and Waluyo (2015) menyatakan bah-wa larva akan hidup dengan baik pa-da media dengan kisaran salinitas 3 –12 ppt. Salinitas juga berhubungan eratdengan osmoregulasi hewan air. Perbe-daan perkembangan stadia yang dilaluitiap-tiap perlakuan terjadi dikarenakan
Gambar 1 Perkembangan larva udang galah (Macro-brachium rosenbergii) GIMacro II

624 Deny Sapto Chondro Utomo1 et al.
Gambar 2 Indeks perkembangan stadia larva udanggalah
penerapan salinitas yang berbeda-bedamenuntut larva untuk beradaptasi de-ngan osmoregulasi, kondisi ini diyaki-ni mempengaruhi beban kerja osmotiklarva (Zikri et al., 2014). Hal ini sesuaidengan pernyataan Hana (2007) yangmenyatakan bahwa perbedaan perkem-bangan ini diduga akibat perbedaan ke-mampuan pengaturan osmotik dan io-nik, yang secara fisiologis harus terjadiuntuk mempertahankan keadaan stea-dy state dan aktivitas ini memerlukanenergi metabolisme, sehingga akan ter-jadi pula perbedaan proses-proses pem-bentukan jaringan tubuh. Semakin be-sar beban osmotik, maka semakin la-ma waktu yang dibutuhkan larva udanggalah untuk berubah stadia (Syafei, 2006).
Pada saat larva udang sedang dalam ma-sa perkembangan stadia, larva meng-alami masa kritis yang cukup tinggi.Terlebih lagi saat peralihan ke post la-rvae. Masa kritis larva udang juga dia-lami saat terjadinya moulting (pergan-tian kulit) (Mukti and Satyantini, 2016).Perkembangan larva udang galah GI-Macro II selama 21 hari pemelihara-an pada penelitian ini dapat dikatakantinggi jika dibandingkan pada pembe-
nihan umumnya yang biasanya menca-pai 25 – 30 hari. Hal tersebut didugakarena larva udang galah GIMacro IIdapat mempertahankan tekanan osmo-tik yang ada pada lingkungan. Supono(2017) menyatakan bahwa jika terja-di perbedaan tekanan osmotik pada he-molim udang dan air kolam yang besarmenyebabkan udang akan banyak ke-hilangan energi untuk adaptasi sehing-ga perkembangan menjadi lambat. Ku-alitas air yang baik akan mendukungperkembangan yang optimal. Sebalik-nya, kualitas air yang jelek akan menu-runkan nafsu makan udang yang bera-kibat terhambatnya perkembangan la-rva udang galah GIMacro II. Perubah-an kualitas air yang terlalu signifikanakan menyebabkan stres pada udang bahk-an akan menyebabkan mortalitas padaudang yang pada akhirnya dapat me-nurunkan biomasa udang yang dipeli-hara (Supono, 2017).
Kelangsungan hidup merupakan para-meter yang dapat menunjukkan presen-tase organisme yang mampu bertahanhidup pada akhir penelitian. Berdasark-an hasil penelitian selama 21 hari, ting-kat kelangsungan hidup udang galah GI-Macro II menunjukkan hasil yang ber-beda untuk setiap perlakuan. Tingkatkelangsungan hidup tertinggi terjadi pa-da pelakuan B yaitu sebesar 67,67 ±4,51 %, kemudian diikuti dengan per-lakuan C sebesar 56,00 ± 3,61 %, danperlakuan A sebesar 50,67 ± 4,51 (Gam-bar 3).
Berdasarkan analisis uji Anova dengantingkat kepercayaan 95%, perbedaan sa-linitas berpengaruh nyata (P<0,05) ter-hadap kelangsungan hidup larva udanggalah GIMacro II stadia 1 hingga postlarvae. Rata-rata kelangsungan hidupperlakuan B lebih baik dibandingkan

Development and survival rate giant prawn at different salinities 625
Gambar 3 Tingkat kelangsungan hidup larva udanggalah
perlakuan A dan C. Pada perlakuan Bdengan nilai osmolaritas media sebe-sar 311 mmol/kg, udang galah GIMa-cro II melakukan kerja mendekati isos-motik yang artinya osmolaritas haemo-lymp hampir sama dengan osmolari-tas medianya, hal ini menunjukkan ter-bukti dengan tingkat kerja osmotik yangpaling kecil dibandingkan dengan sali-nitas 10 ppt dan 14 ppt, sehingga hasilanlisis statistik (Anova) dengan tingkatkepercayaan 95% menunjukkan perbe-daan yang nyata antar perlakuan. Per-nyataan tersebut sesuai dengan Ali andWaluyo (2015) yang menyatakan bah-wa tingkat kelangsungan hidup udanggalah tertinggi diperoleh pada mediadengan kisaran salinitas 3 – 12 ppt. Ha-sil uji lanjut Duncan pada taraf keper-cayaan 95% menunjukkan bahwa per-bedaan salinitas memberikan pengaruhnyata terhadap tingkat kelangsungan hi-dup larva udang galah GIMacro II yangmana pada perlakuan B berbeda nya-ta terhadap perlakuan A dan perlaku-an C. Pada perlakuan A menunjukkantingkat kelangsungan hidup yang lebihrendah dari perlakuan lainnya disebabk-an perkembangan yang tidak seragamantar individu sehingga individu yang
tumbuh dan berkembang dengan cepatakan menguasai wilayah dan persaing-an dalam memperebutkan makanan. La-rva udang yang lemah cenderung akansusah mendapatkan makanan dan mu-dah stress dan terserang penyakit, se-hingga kanibalisme pun tidak dapat di-hindari (Ali and Waluyo, 2015). Sedangk-an pada perlakuan B, tingkat kelang-sungan hidup yang didapat lebih tinggidari perlakuan lainnya disebabkan ka-rena larva udang sudah mampu bera-daptasi dengan baik pada lingkungan-nya sehingga larva udang mampu men-dapatkan makanan yang diberikan de-ngan mudah.
Kehidupan organisme akuatik, terma-suk udang galah, sangat ditentukan olehdaya dukung lingkungan, yang salah sa-tunya adalah parameter kualitas air pe-meliharaan (Khasani et al., 2018). Se-lama penelitian dilakukan pengamatankualitas air yang meliputi suhu air, sa-linitas, oksigen terlarut, dan pH. Ha-sil pengukuran parameter kualitas airselama 21 hari pemeliharaan menun-jukkan parameter kualitas air dalam ba-tas optimal bagi kehidupan larva udanggalah GIMacro II (Tabel 1).
Suhu selama penelitian berkisar antara28 – 29 oC. Kisaran suhu tersebut ma-sih dalam kisaran toleransi udang ga-lah GIMacro II. Jika suhu air lebih dariangka tersebut maka metabolisme da-lam tubuh udang akan berlangsung ce-pat. Suhu dapat mempengaruhi berba-gai fungsi metabolisme dari organisasiakuatik seperti pertumbuhan dan ting-kat konsumsi pakan (Serang et al., 2006).Semakin tinggi suhu maka akan mem-percepat proses metabolisme dan me-ningkatkan konsumsi pakan pada udangsehingga mempercepat pula proses mo-

626 Deny Sapto Chondro Utomo1 et al.
Tabel 1 Kualitas air media pemeliharaan udang galah selama penelitian
Parameter Kisaran Optimum* A B CSuhu (°C) 27 – 31 28 – 29 28 – 29 28 – 29
pH 7 – 8,5 7-7,78 7,73 – 7,83 7,57 – 7,81DO (mg/L) 1,90 – 6,98 4,4 – 5,3 4,8 – 5,8 4,7 – 5,5
Salinitas (ppt) 8 – 16 10 12 14
ulting yang akan mempengaruhi per-tumbuhan panjang dan beratnya.
Udang galah GIMacro II memiliki to-leransi luas terhadap salinitas sehing-ga udang galah GIMacro II dapat ber-adaptasi dalam keadaan salinitas yangrendah. Hal ini sesuai dengan pernya-taan Ali and Waluyo (2015) bahwa per-kembangan dan kelangsungan hidup udanggalah yang baik diperoleh pada mediadengan kisaran salinitas 3 – 12 ppt.
Salinitas adalah tingkat kadar garam ter-larut dalam air yang merupakan salahsatu aspek kualitas air yang memegangperanan penting karena mempengaru-hi pertumbuhan udang. Salinitas ada-lah konsentrasi semua ion-ion terlarutdalam air (klorida, karbonat, bikarbo-nat, sulfat, natrium, kalsium dan mag-nesium). Konsentrasi salinitas sangat ber-pengaruh terhadap proses osmoregola-si yaitu upaya hewan air untuk meng-ontrol keseimbangan air dan ion antaratubuh dan lingkungannya. Jika kondi-si salinitas berfluktuasi maka semakinbanyak energi yang dibutuhkan untukmetabolisme. Metabolisme yang dila-kukan merupakan bentuk adaptasi (Fu-jaya, 2004).
Kandungan oksigen terlarut dalam airmerupakan faktor kritis bagi kehidup-an udang. Oksigen terlarut berperan da-lam perkembangan dan kelangsunganhidup organisme akuatik. Kandunganoksigen terlarut juga sangat penting ba-gi udang galah GIMacro II karena ok-sigen dibutuhkan untuk proses meta-bolisme. Pergantian air pada media pe-
meliharaan juga membantu meningkatk-an oksigen terlarut.
Pada penelitian ini didapatkan nilai ka-dar oksigen terlarut yang berada padakisaran 4,48 - 5,58 mg/L. Kisaran yangdihasilkan saat pengukuran kualitas airselama pemeliharaan ini masih dapatditoleransi bagi kelangsungan hidup udanggalah GIMacro II. Semakin besar udanggalah GIMacro II maka konsumsi ok-sigen yang dibutuhkan semakin besar,namun oksigen di dalam media peme-liharaan berkurang. Hal ini dikarenak-an aerasi di dalam media pemelihara-an dari awal sampai akhir pemelihara-an udang tetap sama, tetapi kebutuhanoksigen udang galah semakin mening-kat seiring dengan peningkatan ukurantubuhnya. Kadar oksigen terlarut opti-mum > 3 mg/L dengan toleransi 2 mg/L.Kandungan oksigen terlarut yang ren-dah di bawah 1,5 mg/L akan bersifatlethal bagi udang (Maimunah and Ki-lawati, 2015).
Hasil pengukuran pH selama peneliti-an berkisar antara 7 – 7,83. pH perair-an 6,5 - 8,5 merupakan batas optimumyang memungkinkan udang dapat hi-dup dan berkembang. Nilai pH di ba-wah 4,5 atau di atas 9,0 akan mengaki-batkan udang mudah sakit, lemah, dannafsu makan menurun, bahkan cang-kang udang galah cenderung keroposdan berlumut.

Development and survival rate giant prawn at different salinities 627
SIMPULAN
Perbedaan salinitas memberi-kan penga-ruh yang nyata terhadap perkembang-an dan kelangsungan hidup larva udanggalah GIMacro II. Hasil penelitian me-nunjukkan bahwa salinitas 12 ppt me-rupakan salinitas terbaik dengan indeksperkembangan larva sebesar 7,13 ± 0,03% dan kelangsungan hidup sebesar 67,67± 4,51 %. Berdasarkan penelitian yangtelah dilakukan maka penulis menya-rankan dalam kegiatan pembenihan la-rva udang galah GIMacro II menggu-nakan salinitas 12 ppt.
Pustaka
Ali, F. and Waluyo, A. (2015). Tingkatkelangsungan hidup dan pertumbuh-an udang galah (macrobrachium ro-senbergii de man) pada media bersa-linitas. LIMNOTEK-Perairan DaratTropis di Indonesia, 22(1).
Fujaya, Y. (2004). Fisiologi ikan da-sar pengembangan teknik perikanan.Rineka Cipta. Jakarta, 179.
Hana, G. C. (2007). Respon udangvanname (litopenaeus vannamei)terhadap media bersalinitas rendah.
Khasani, I., Imron, I., Suprapto, R.,and Himawan, Y. (2018). Evalu-asi keragaan persilangan udang ga-lah (macrobrachium rosenbergii) da-ri beberapa sumber populasi. In Pro-siding FORUM INOVASI TEKNO-LOGI AKUAKULTUR, pages 581–590.
Maimunah, Y. and Kilawati, Y. (2015).Kualitas lingkungan tambak insentiflitapenaeus vannamei dalam kaitan-nya dengan prevalensi penyakit whi-te spot syndrome virus. Research Jo-urnal of Life Science, 2(1):50–59.
Mukti, A. T. and Satyantini, W. H.(2016). Peranan l-carnitine pada per-kembangan dan pertumbuhan larvaudang galah, macrobrachium rose-nbergii de man. Jurnal Ilmu-IlmuPerairan dan Perikanan Indonesia,12(1):23–26.
Nugrahaningsih, K. A. (2008). Penga-ruh tekanan osmotik media terhadaptingkat kelangsungan hidup dan per-tumbuhan benih ikan patin (pangasi-us sp.) pada salinitas 5 ppt.
Priyono, S. B., Sukardi, S., and Hari-anja, B. S. (2011). Pengaruh shelterterhadap perilaku dan pertumbuhanudang galah (macrobrachium rose-nbergii). Jurnal Perikanan Univer-sitas Gadjah Mada, 13(2):78–85.
Serang, M., Suprayudi, M., Jusadi, D.,and Mokoginta, I. (2006). Pengaruhkadar protein dan rasio energi pro-tein pakan berbeda terhadap kinerjapertumbuhan benih rajungan (portu-nus pelagicus). Bogor: Tesis. InstitutPertanian Bogor.
Supono, S. (2017). Studi keraga-an udang windu (penaeusmonodon)dan udang putih (litopenaeusvanna-mei) yang dipelihara pada tambaksemi plastik. In Prosiding SeminarNasional Teknologi Pertanian.
Syafei, L. S. (2006). Pengaruh bebankerja osmotik terhadap kelangsung-an hidup, lama waktu perkembanganlarva dan potensi tumbuh pascalarvaudang galah.
Zikri, O., Hukama Taqwa, F., et al.(2014). Penentuan pola perubahansalinitas pada penetasan dan pemeli-haraan larva udang galah (macrobra-chium rosenbergii) asal sumatera se-latan. Jurnal Akuakultur Rawa Indo-nesia, 1(1):46–56.

628 Deny Sapto Chondro Utomo1et al.

AQUASAINS(Jurnal Ilmu Perikanan dan Sumberdaya Perairan)(Vol 7 No. 1 Tahun 2018)
REDUCTION OF PATHOGENIC BACTERIA DURINGFERMENTATION OF MASIN BY PROTEASEAND BACTERIOCIN-PRODUCING LACTIC ACIDBACTERIA
Heru Pramono1 · Taruna Fernando Putra2 · Sri Utari2 · NoviAndika2
Ringkasan Contamination of pathogenicand spoilage bacteria on fermentationprocess of seafood and fisheries pro-duct is a major concern on food safe-ty. The aims of this study were isola-ting and applying the bacteriocin- andprotease-producing lactic acid bacte-ria from the gastrointestinal tract of mudcrab for starter culture of masin, a tra-ditional fermented shrimp from East Ja-va. This study consisted of characteri-zation of lactic acid bacteria, the ap-plication on the fermentation processand microbial analysis. Ninety-four iso-lates were isolated from mud crab wasscreened for the bacteriocin and prote-ase producing as well as characterizedby pH, salinity and biochemical. Iso-late IKP-29 was exhibited strong pro-tease and bacteriocin activity. Appli-cation of Isolate IKP-29 on masin fer-mentation showed that sharp reductionof Escherichia coli, Vibrio sp. and la-ctic acid bacteria counted. This studysuggested that the application of lactic
1)Departemen Kelautan, Fakultas Perikanan dan Ke-lautan Universitas Airlangga, Kampus C UNAIR, Jl.Mulyorejo, Surabaya, 60115, Indonesia; 2)ProgramStudi Akuakultur, Fakultas Perikanan dan KelautanUniversitas Airlangga, Kampus C UNAIR, Jl. Mulyo-rejo, Surabaya, 60115, Indonesia.E-mail: [email protected]
acid bacteria which producing bacte-riocin and protease improved the foodsafety of traditional fermented fish.
Keywords masin, lactic acid bacteria,fermentation, bacteriocin, protease
Received : 27 Juli 2018
Accepted : 23 Agustus 2018
PENDAHULUAN
Salah satu permasalahan yang dihada-pi terkait keamanan pangan ialah ada-nya food-borne pathogen seperti Sal-monella thypii penyebab demam tifus,Eschericia coli penyebab gangguan pen-cernaan, maupun Vibrio cholera penye-bab kolera. Berbagai upaya untuk meng-atasi permasalahan tersebut telah dila-kukan, baik berupa preservasi dengantermal maupun non termal. Preserva-si pangan tersebut dengan termal dapatberupa pengeringan, pemanasan dan pe-masakan maupun dengan non termalseperti ultrasonikasi, tekanan hiperba-rik, ionisasi, dan aplikasi biopreserva-si. Salah satu metode biopreservasi ia-lah dengan aplikasi bakteri asam lak-tat (BAL) penghasil bakteriosin seba-gai starter (Hwanhlem et al., 2011) atau-

630 Heru Pramono1 et al.
pun aplikasi ekstrak bakteriosin padapangan (Udhayashree et al., 2012).
BAL tersebar luas di alam, termasukdi dalam saluran pencernaan. Saluranpencernaan ikan merupakan ekosistemkompleks yang dihuni oleh mikrobio-ta, baik mikrobia aerob, fakultatif ana-erob maupun obligat anaerob (Talpuret al., 2012). Hal tersebut menyebabk-an saluran pencernaan dapat menyimp-an potensi kolonisasi bakteri patogenmaupun penghasil antibakteri (Sugitaet al., 2002).
Isolasi BAL penghasil bakteriosin pa-da produk fermentasi telah banyak di-lakukan (Kopermsub and Yunchalard,2010; Moroni et al., 2011Moraes et al.,2010Pramono et al., 2015). Talpur et al.(2012) telah melakukan isolasi BAL pa-da saluran pencernaan rajungan untuktujuan probiotik. Namun masih sedikitinformasi terkait BAL penghasil bak-teriosin pada saluran pencernaan kepi-ting yang diaplikasikan sebagai kandi-dat starter untuk memperbaiki mutu ser-ta keamanan pangan produk fermenta-si.
Scylla sp. merupakan salah satu jeniskepiting yang banyak ditemukan padaekosistem mangrove Samudra Hindia,Laut Merah, Samudra Pasifik dan ter-masuk spesies yang paling banyak ter-sebar di dunia (Shelley and Lovatelli,2011). Genus Scylla memiliki ciri-cirisebagai berikut: panjang pasangan ka-ki jalan lebih pendek daripada capit,pasangan kaki terakhir berbentuk da-yung, karapas berbentuk lebar, dileng-kapi dengan 3–9 buah gigi anterolate-ral, ruas dasar dari antena biasanya le-bar, sudut anteroexternal seringkali ber-lobi, flagel kadang-kadang berada pa-da orbital mata (Fielder and Heasman,1978).
Kepiting pada ekosistem mangrove me-miliki potensi besar untuk menjadi sum-ber isolasi BAL penghasil bakteriocin-likeuntuk aplikasi pangan sebagai agenbiopreservatif sekaligus starter perba-ikan mutu produk. Hal ini disebabkankepiting mampu hidup pada salinitasyang tinggi dan mengkonsumsi detri-tus sehingga dimungkinkan adanya ko-lonisasi bakteri pemecah protein yangkuat. Oleh karena itu penelitian ini pen-ting dilakukan untuk mendapatkan kan-didat BAL penghasil bakteriosin kuatpada saluran pencernaan kepiting un-tuk kemudian dimanfaatkan sebagai star-ter pada produksi fermentasi ikan tra-disional.
Tujuan penelitian adalahmendapatkanisolat starter bakteri asam laktat peng-hasil bacteriocin-like untuk memperba-iki produk fermentasi tradisional danmendapatkan data karakteristik produkdan keamanan produk fermentasi ikanyang dihasilkan dengan aplikasi star-ter.
MATERI DAN METODE
Isolat Bakteri Asam Laktat
Isolat bakteri asam laktat (BAL) dipe-roleh dari hasil penelitian sebelumnya.Sebanyak 94 isolat BAL hasil isolasisaluran pencernaan kepiting bakau te-lah disimpan pada gliserol steril dandisimpan pada suhu -10oC hingga pe-nelitian dilakukan (Pramono et al.., 2015).Se-belum uji dilakukan sebanyak 10 µLisolat BAL-gliserol telah dikultur pada1 mL de Mann, Rogosa, Sharpe (MRS)broth (Merck, Darmstadt, Jerman) pa-da suhu 30oC selama 48 jam.Bakteripatogen indikator yang digunakan me-rupakan Vibrio sp. hasil isolasi dari sa-te kerang (Pramono et al.., 2015)dan E.

Reducing of Pathogenic Bacteria 631
coli ATCC 2595. Bakteri Vibrio sp. di-kultur pada medium saline tryptic soyabroth (3% NaCl), sedangkan E. coli di-kultur pada medium tryptic soya broth(Merck, Darmstadt, Jerman). Uji Pro-duksi Enzim Protease
Isolat BAL kandidat starter diuji ke-mampuan menghasilkan senyawa enzimproteolitik dengan menggunakan me-dia skim milk agar (SMA) (skim milk2%, agar 1,5%) untuk mengetahui ak-tivitas penghasilan enzim proteolitik-nya. Dua puluh mikroliter kultur bro-th MRS isolat kandidat starter BAL di-teteskan pada permukaan media SMA.Plate kemudian diinkubasi pada suhu37oC selama 24 jam. Isolat yang mam-pu mendegradasi protein susu menun-jukkan zona jernih di sekeliling kolo-ni. Tiga isolat dengan diameter zonajernih terbesar akan dikarakterisasi le-bih lanjut untuk mendapatkan satu iso-lat kandidat starter fermentasi ikan
Karakterisasi Bakteri Asam Laktat
- Pengecatan Gram
Pengecatan Gram dilakukan dengan meng-ambil satu ose larutan garam fisiolo-gis dan diteteskan di atas gelas prepa-rat kemudian satu ose biakan BAL di-ratakan diatasnya kemudian difiksasi diatas api bunsen. Setelah terfiksasi ber-ikan Gram A (kristal violet) selama sa-tu menit, bilas dengan air mengalir. Te-teskan Gram B (Kalium Iodida) sela-ma satu menit, bilas dengan air meng-alir. Setelah itu teteskan Gram C (eta-nol) selama 30 detik dan bilas denganair mengalir baru kemudian berikan GramD (safranin) selama dua menit sebelumdiamati di bawah mikroskop. Morfolo-gi sel diamati di bawah lensa mikros-kop untuk mengetahui bentuk, ukuran,maupun konformasi sel BAL.
- Uji Oksidasi Fermentasi
Uji fermentasi dilakukan dengan meng-gunakan medium O/F. Satu ose kulturBAL diinokulasikan pada medium OFsecara aseptis, satu tabung ditutup de-ngan parafin sedangkan satu tabung di-tutup dengan kapas steril. Bakteri yangmampu melakukan oksidasi ditunjukk-an dengan warna kuning pada medi-um OF tanpa parafin sedangkan bak-teri yang mampu melakukan fermenta-si ditunjukkan oleh warna kuning padamedium OF berparafin.
- Uji Tipe Fermentasi
Uji tipe fermentasi dilakukan degan de-ngan cara menginokulasikan 50 µL BALke dalam medium MRS cair dalam ta-bung reaksi yang telah dimasukkan ta-bung durham dan diinkubasi selama 24jam pada suhu 37oC. bakteri yang meng-hasilkan gas dinyatakan heterofermen-tatif sedangkan yang tidak menghasilk-an gas dinyatakan homofermentatif.
– Uji Produksi H2S
Produksi H2S dilakukan dengan meng-gunakan medium triple sugar iron agar(TSIA). Sebanyak satu ose kultur BALditusukkan pada ujung medium TSIAmiring dan digoreskan dengan meng-gunakan jarum inokulasi. Hasil streakdiinkubasi pada suhu 30oC selama 24jam dan dilakukan pengamatan. Bakte-ri yang mampu menghasilkan H2S ak-an menunjukkan warna hitam pada me-dia TSIA.
- Uji ketahanan Terhadap pH
Uji fisiologi yang dilakukan adalah ujiproduksi gas dan uji pertumbuhan pa-da pH 1, 3, 5, 7 dan 9. Medium MRSbroth diatur pH-nya dengan menggu-nakan 1N HCl atau 1N NaOH untukmendapatkan pH yang sesuai. Sebanyak

632 Heru Pramono1 et al.
50 µL kultur BAL diinokulasikan pa-da mikrotube yang telah diisi denganMRS dengan pH yang telah disesuaik-an, yaitu 1,3,5,7, dan 9 kemudian te-lah dilakukan inkubasi pada suhu 30oCdan dilakukan pengamatan. BAL yangtumbuh menimbulkan media MRS bro-th menjadi keruh dan setelah diplate pa-da medium MRS agar menunjukkan tum-buhnya koloni.
- Uji Ketahanan Terhadap Salinitas
Uji ketahanan terhadap salinitas dila-kukan dengan menggunakan mediumMRS. Sebanyak 50 µL kultur BAL di-inokulasikan pada medium MRS bro-th dengan konsentrasi NaCl yang ber-beda (1%, 2%, 3%, 4%, dan 5%) dandiinkubasi pada suhu 30oC selama 24jam.BAL yang tumbuh menunjukkankekeruhan pada media dan setelah di-plate pada medium MRS agar menun-jukkan tumbuhnya koloni merupakanbakteri yang tahan terhadap salinitas yangdigunakan.
Aplikasi Bakteri Asam Laktat Starterpada Produksi Masin
- Preparasi masin dan inokulasi star-ter
Masin diproduksi dengan mencampurk-an 400 g udang dengan nasi kering yangtelah disangrai sebanyak 120 gram di-tambah dengan 400 gram garam dapur,dan 100 mL air keran. Setelah diadukmerata, bakteri E.coli ATCC 2595, Vi-brio sp. dan BAL starter dimasukkandalam adonan dengan kepadatan 6 logCFU/g untuk campuran dari ketiga iso-lat kuat (IKP-29, IKP-30, dan IKP-94).Proses fermentasi dilakukan pada suhuruang dan dilakukan pengamatan ber-upa analisis mikrobiologi dan pengu-kuran pH pada jam ke-0, 24, 48, 72,dan 96.
Tabel 1 Diameter zona hidrolisis kasein susu tiga iso-lat terkuat
Isolat Zona hidrolisis (mm)24 jam 48 jam 72 jam
IKP29 3 12 12IKP30 5 6 6IKP94 5 7 9
- Analisis mikrobiologi
Analisis mikrobiologi terdiri atas per-hitungan jumlah E. coli, Vibrio sp., danBAL dengan metode TPC pada medi-um eosin methylene blue agar (EMBA),thiosulfate citrate bile salts sucrose (TCBS),dan de Man, Rogosa dan Sharpe (MRS)agar, secara berurutan.
HASIL DAN PEMBAHASAN
Uji Produksi Protease
Sebanyak 94 isolat bakteri asam laktat(BAL) penghasil bakteriosin-like yangtelah diuji dengan netralisasi pH danuji sumuran terhadap bakteri indikatortelah diperoleh dari saluran pencerna-an kepiting bakau (Pramono et al., 2015)kemudian telah diuji aktivitas produksienzim protease. Berdasarkan hasil ujiaktivitas proteolitik, tiga isolat yang mam-pu menghasilkan protease kuat (>6 mm)pada medium skim milk adalah isolaIKP29, IKP30, dan IKP94. Kemampu-an bakteri dalam menghasilkan prote-ase ekstraseluler menyebabkan terjadi-nya lisis pada kasein susu pada medi-um skim milk agar sehingga terdapatzona jernih disekeliling koloni (Nespo-lo and Brandelli, 2010). Semakin be-sar zona yang dihasilkan maka sema-kin besar aktivitas proteolitik isolat yangdiuji. Hasil penelitian menunjukkan bah-wa terdapat 3 isolat BAL yang mampumenghasilkan enzim protease > 6 mmpada jam ke 72 (Tabel 1).

Reducing of Pathogenic Bacteria 633
Enzim protease dihasilkan mikrobia un-tuk mendapatkan asam amino dan pep-tida dari hidrolisis protein. Asam ami-no tersebut digunakan oleh bakteri se-bagai bahan penyusun protein seluler,baik fungsional maupun struktural, atausebagai sumber nitrogen pada sel bak-teri (Setyati and Subagiyo, 2012; Re-migio et al., 2012).
Karakterisasi Bakteri Asam Laktat Peng-hasil Protease
Identifikasi BAL didasarkan pada duametode umum yaitu metode morfologidan fisiologidan dengan menggunakanmetode molekuler (Mohammed et al.,2009) yaitu dengan sekuensing gen 16SrDNA. Identifikasi BAL dengan meto-de morfologi dan fisiologi dilakukan de-ngan tahapan: pengecatan Gram, ben-tuk sel, uji motilitas, uji fisiologi danbiokimia (uji katalase, tipe fermentasi,dan pertumbuhan sel pada pH terten-tu).
Hasil pengecatan Gram dan morfolo-gi (Tabel 2) menunjukkan bahwa keti-ga isolat merupakan Gram positif. Iso-lat IKP29 dan IKP94 memiliki bentukbulat sedangkan isolat IKP30 berben-tuk batang pendek. Bakteri asam lak-tat merupakan kelompok bakteri Grampositif dengan bentuk batang ataupunbulat (Speranza et al., 2017). Bebera-pa spesies bakteri asam laktat mampumenghasilkan senyawa antimikrobia se-perti bakteriosin, hidrogen peroksida,maupun asam organik(Putra et al., 2018).
Hasil uji TSIA menunjukkan bahwa BALtersebut tidak menghasilkan hidrogendisulfida. Ketiga isolat mampu mela-kukan fermentasi glukosa dan terma-suk dalam katagori BAL homofermen-tatif yang melakukan fermentasi glu-kosa menjadi asam laktat.Menurut Sai-thong et al. (2010)salah satu parameter
BAL dapat dimanfaatkan sebagai kul-tur starter fermentasi ikan ialah terma-suk dalam jenis homofermentatif. Ha-sil uji oksidase dan katalase menunjukk-an ketiga isolat tidak menghasilkan enzimoksidase dan katalase. BAL termasukdalam ordo Lactobacillales (Ludwig et al.,2015)) yang memiliki ciri: Gram po-sitif, berbentuk batang atau kokus, ti-dak berspora, non motil, katalase nega-tif dan oksidase negatif. BAL menggu-nakan karbohidrat sebagai sumber ener-gi dan menghasilkan asam laktat se-bagai produk utama atau satu-satunyafermentasi dan merupakan golongan bak-teri fakultatif anaerob (Willey et al., 2009).Bak-teri asam laktat termasuk penghasil bak-teriosin yang masuk dalam golongangenerally recognized as safe (GRAS)oleh WHO sehingga aman digunakandalam produk pangan.Hasil uji ketahan-an terhadap pH dan salinitas ditunjukk-an oleh Tabel 3. Ketahanan terhadapsalinitas dan pH penting kaitannya de-ngan kemampuan bakteri starter dalammatrik pangan. Berdasarkan hasil ujiketahanan terhadap pH, dapat diketa-hui bahwa ketiga isolat memiliki ren-tang ketahanan yang baik pada salini-tas dan pH yang bervariasi. Meskipundemikian, isolat IKP29 memiliki ren-tang ketahanan yang terbaik terhadapsalinitas dan pH sehingga isolat IKP29dapat digunakan sebagai kultur starteryang potensial pada produk dengan ka-dar garam yang tinggi dan pH yang lu-as. Hasil identifikasi isolat IKP29 me-nunjukkan bahwa isolat termasuk da-lam Genus Pediococcus.
Aplikasi Bakteri Asam Laktat Starterpada Produksi Masin Analisis mikro-biologi dan pH
Hasil analisis menunjukkan bahwa ter-jadi penurunan jumlah bakteri yang sig-
634 Heru Pramono1 et al.
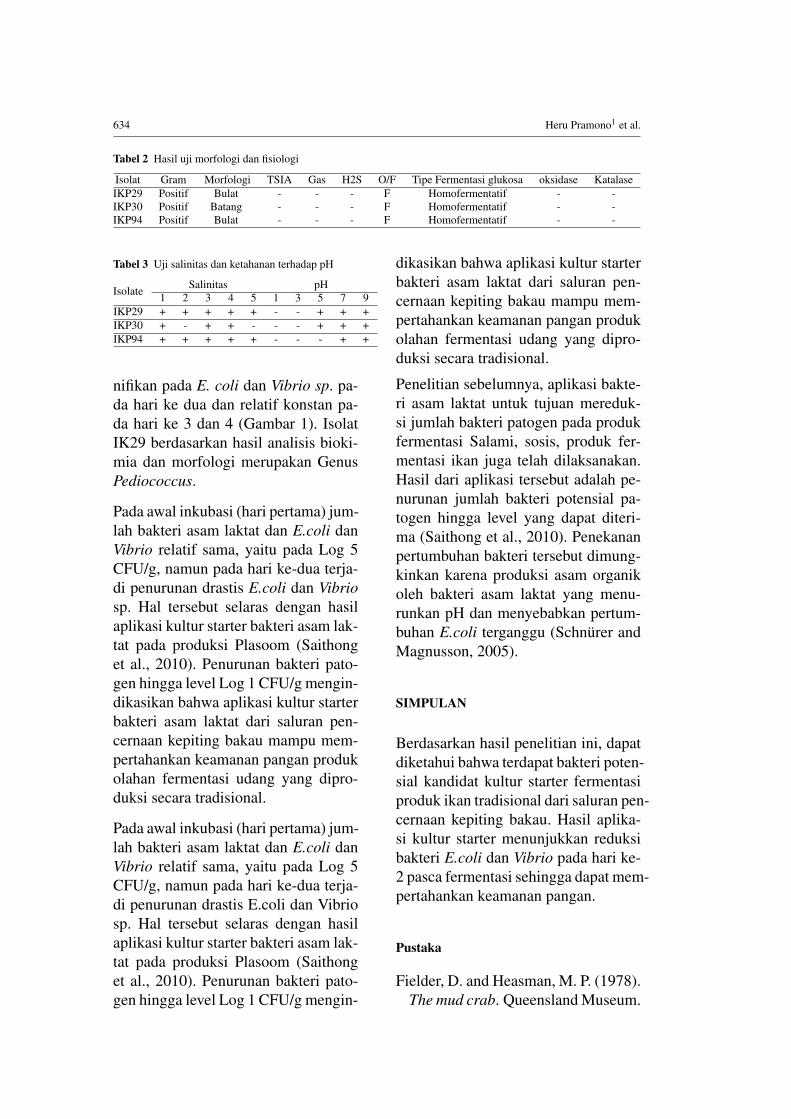
Tabel 2 Hasil uji morfologi dan fisiologi
Isolat Gram Morfologi TSIA Gas H2S O/F Tipe Fermentasi glukosa oksidase KatalaseIKP29 Positif Bulat - - - F Homofermentatif - -IKP30 Positif Batang - - - F Homofermentatif - -IKP94 Positif Bulat - - - F Homofermentatif - -
Tabel 3 Uji salinitas dan ketahanan terhadap pH
Isolate Salinitas pH1 2 3 4 5 1 3 5 7 9
IKP29 + + + + + - - + + +IKP30 + - + + - - - + + +IKP94 + + + + + - - - + +
nifikan pada E. coli dan Vibrio sp. pa-da hari ke dua dan relatif konstan pa-da hari ke 3 dan 4 (Gambar 1). IsolatIK29 berdasarkan hasil analisis bioki-mia dan morfologi merupakan GenusPediococcus.
Pada awal inkubasi (hari pertama) jum-lah bakteri asam laktat dan E.coli danVibrio relatif sama, yaitu pada Log 5CFU/g, namun pada hari ke-dua terja-di penurunan drastis E.coli dan Vibriosp. Hal tersebut selaras dengan hasilaplikasi kultur starter bakteri asam lak-tat pada produksi Plasoom (Saithonget al., 2010). Penurunan bakteri pato-gen hingga level Log 1 CFU/g mengin-dikasikan bahwa aplikasi kultur starterbakteri asam laktat dari saluran pen-cernaan kepiting bakau mampu mem-pertahankan keamanan pangan produkolahan fermentasi udang yang dipro-duksi secara tradisional.
Pada awal inkubasi (hari pertama) jum-lah bakteri asam laktat dan E.coli danVibrio relatif sama, yaitu pada Log 5CFU/g, namun pada hari ke-dua terja-di penurunan drastis E.coli dan Vibriosp. Hal tersebut selaras dengan hasilaplikasi kultur starter bakteri asam lak-tat pada produksi Plasoom (Saithonget al., 2010). Penurunan bakteri pato-gen hingga level Log 1 CFU/g mengin-
dikasikan bahwa aplikasi kultur starterbakteri asam laktat dari saluran pen-cernaan kepiting bakau mampu mem-pertahankan keamanan pangan produkolahan fermentasi udang yang dipro-duksi secara tradisional.
Penelitian sebelumnya, aplikasi bakte-ri asam laktat untuk tujuan mereduk-si jumlah bakteri patogen pada produkfermentasi Salami, sosis, produk fer-mentasi ikan juga telah dilaksanakan.Hasil dari aplikasi tersebut adalah pe-nurunan jumlah bakteri potensial pa-togen hingga level yang dapat diteri-ma (Saithong et al., 2010). Penekananpertumbuhan bakteri tersebut dimung-kinkan karena produksi asam organikoleh bakteri asam laktat yang menu-runkan pH dan menyebabkan pertum-buhan E.coli terganggu (Schnürer andMagnusson, 2005).
SIMPULAN
Berdasarkan hasil penelitian ini, dapatdiketahui bahwa terdapat bakteri poten-sial kandidat kultur starter fermentasiproduk ikan tradisional dari saluran pen-cernaan kepiting bakau. Hasil aplika-si kultur starter menunjukkan reduksibakteri E.coli dan Vibrio pada hari ke-2 pasca fermentasi sehingga dapat mem-pertahankan keamanan pangan.
Pustaka
Fielder, D. and Heasman, M. P. (1978).The mud crab. Queensland Museum.

Reducing of Pathogenic Bacteria 635
Gambar 1 Hasil perhitungan TPC, (a) Escherichia coli ATCC 2595 dan Vibrio sp.; (b). Bakteri asam laktat
Hwanhlem, N., Buradaleng, S., Watta-nachant, S., Benjakul, S., Tani, A.,and Maneerat, S. (2011). Isolationand screening of lactic acid bacteriafrom thai traditional fermented fish(plasom) and production of plasomfrom selected strains. Food Control,22(3-4):401–407.
Kopermsub, P. and Yunchalard, S.(2010). Identification of lactic acidbacteria associated with the produ-ction of plaa-som, a traditional fer-mented fish product of thailand. In-ternational journal of food microbi-ology, 138(3):200–204.
Ludwig, W., Schleifer, K.-H., andWhitman, W. B. (2015). Revisedroad map to the phylum firmicutes.Bergey’s Manual of Systematics ofArchaea and Bacteria, pages 1–16.
Mohammed, M., El-Aziz, H. A., Omr-an, N., Anwar, S., Awad, S., andEl-Soda, M. (2009). Rep-pcr cha-racterization and biochemical sele-ction of lactic acid bacteria isolatedfrom the delta area of egypt. Interna-tional journal of food microbiology,128(3):417–423.
Moraes, P. M., Perin, L. M., Ortola-ni, M. B. T., Yamazi, A. K., Viçosa,
G. N., and Nero, L. A. (2010). Pro-tocols for the isolation and detectionof lactic acid bacteria with bacterio-cinogenic potential. LWT-Food Sci-ence and Technology, 43(9):1320–1324.
Moroni, A. V., Arendt, E. K., andDal Bello, F. (2011). Biodiversi-ty of lactic acid bacteria and yeas-ts in spontaneously-fermented buc-kwheat and teff sourdoughs. FoodMicrobiology, 28(3):497–502.
Nespolo, C. R. and Brandelli, A.(2010). Production of bacteriocin-like substances by lactic acid bacte-ria isolated from regional ovine che-ese. Brazilian Journal of Microbio-logy, 41(4):1009–1018.
Pramono, H., Suciati, P., and Andri-yono, S. (2015). Isolation of lacticacid bacteria that produce proteaseand bacteriocin-like substance frommud crab (scylla sp.) digestive tra-ct (isolasi bakteri asam laktat yangmenghasilkan protease dan senyawabacteriocin-like dari saluran pencer-naan kepiting. ILMU KELAUTAN:Indonesian Journal of Marine Scien-ces, 20(1):33–37.

636 Heru Pramono1 et al.
Putra, T., Suprapto, H., Tjahjaningsih,W., and Pramono, H. (2018). The an-tagonistic activity of lactic acid ba-cteria isolated from peda, an indo-nesian traditional fermented fish. InIOP Conference Series: Earth andEnvironmental Science, volume 137,page 012060. IOP Publishing.
Remigio, Z., William, M., Olle, H.,and Wilson, P. (2012). Iso-lation and characterization of aprotease-producing thermophilic ba-cterium from an african hot spring.African Journal of Biotechnology,11(62):12571–12578.
Saithong, P., Panthavee, W., Boonya-ratanakornkit, M., and Sikkhamon-dhol, C. (2010). Use of a starter cul-ture of lactic acid bacteria in plaa-som, a thai fermented fish. Jour-nal of bioscience and bioenginee-ring, 110(5):553–557.
Schnürer, J. and Magnusson, J. (2005).Antifungal lactic acid bacteria as bi-opreservatives. Trends in Food Sci-ence & Technology, 16(1-3):70–78.
Setyati, W. A. and Subagiyo, S.(2012). Isolasi dan seleksi bakteripenghasil enzim ekstraseluler (pro-teolitik, amilolitik, lipolitik dan se-lulolitik) yang berasal dari sedimenkawasan mangrove (isolation and se-lection of extracellular enzyme pro-ducing bacteria originating from ma-ngrove sedimen. ILMU KELAUT-AN: Indonesian Journal of MarineSciences, 17(3):164–169.
Shelley, C. and Lovatelli, A. (2011).Mud crab aquaculture: a practicalmanual. FAO Fisheries and aquacul-ture technical paper, (567):I.
Speranza, B., Racioppo, A., Beneduce,L., Bevilacqua, A., Sinigaglia, M.,and Corbo, M. R. (2017). Auto-chthonous lactic acid bacteria with
probiotic aptitudes as starter culturesfor fish-based products. Food micro-biology, 65:244–253.
Sugita, H., Okano, R., Suzuki, Y., Iwai,D., Mizukami, M., Akiyama, N., andMatsuura, S. (2002). Antibacterialabilities of intestinal bacteria fromlarval and juvenile japanese floun-der against fish pathogens. FisheriesScience, 68(5):1004–1011.
Talpur, A., Memon, A., Khan, M., Ikh-wanuddin, M., Daniel, M. D., andAbol-Munafi, A. (2012). Isolationand screening of lactic acid bacteriafrom the gut of blue swimming crab,p. pelagicus, an in vitro inhibitionassay and small scale in vivo modelfor validation of isolates as probioti-cs. Journal of Fisheries and AquaticScience, 7(1):1–28.
Udhayashree, N., Senbagam, D., Sen-thilkumar, B., Nithya, K., and Guru-samy, R. (2012). Production of ba-cteriocin and their application in fo-od products. Asian Pacific Journalof Tropical Biomedicine, 2(1):S406–S410.
Willey, J., Sherwood, L., and Woolver-ton, C. (2009). Prescotts principlesof microbiology2009.

AQUASAINS(Jurnal Ilmu Perikanan dan Sumberdaya Perairan)(Vol 7 No. 1 Tahun 2018)
THE EFFECT OF ADDITION CURCUMA’S (CurcumaXanthorrhiza Roxb) EXTRACT TO THE INCREASEOF FEED CONSUMPTION, EFFICIENCY AND THEGROWTH OF CATFISH (Pangasius)
Prastito1 · Pinandoyo1 · Ristiawan Agung Nugroho1 · ViviEndar Herawati1
Ringkasan Catfish (Pangasius sp.) Is oneof the most cultivated freshwater fish,because it is one of the fish that has ahigh economic value. However, the useof phytopharmaca is used to improvefeed efficiency and fish growth. One ofthe phytopharmaca which can be usedas an antimicrobial is temulawak (Cur-cuma xanthorrhiza Roxb). This studyaims to determine the optimal dose ofthe effect of temulawak extract on arti-ficial feed in increasing feed consump-tion, efficiency of feed utilization, andgrowth rate of catfish (Pangasius sp.).The test fish is maintained with a stoc-king density of 1 tail / l with a mainte-nance time of 42 days. This study usedan experimental method with 4 trea-tments and 3 replications. The treatmen-ts in this study were Treatment A (addi-tion of 0 ml ginger extract), B (additionof 6 ml temulawak extract), C (addi-tion of 12 ml curcuma extract), and D(addition of 18 ml ginger extract). Thedata observed included absolute wei-ght, crime scene, EPP, SGR, REP, SR
1)Departemen Akuakultur Jurusan Perikanan FakultasPerikanan dan Ilmu Kelautan, Universitas Diponego-ro Jl. Prof. Soedarto, SH, Tembalang, Semarang, JawaTengah – 50275, Telp/Fax. +6224 7474698E-mail: [email protected]
and water quality. The results showedthat the addition of temulawak extractto artificial feed had a very significanteffect on absolute weights, crime sce-nes, EPP, SGR, REP but did not differsignificantly for survival. The optimumdose of addition of curcuma extract tototal feed consumption is 11 ml capa-ble of producing a maximum crime sce-ne of 168.1 g, efficiency of feed utiliza-tion is 10.5 ml capable of producing amaximum EPP of 70.3% and specificgrowth rate is 10.8 ml resulting in ma-ximum SGR 2.46% / day.
Keywords masin, lactic acid bacteria,fermentation, bacteriocin, protease
Received : 24 Agustus 2018
Accepted : 23 September 2018
PENDAHULUAN
Ikan patin adalah salah satu ikan air ta-war yang paling banyak dibudidayak-an, karena merupakan salah satu ikanyang memiliki nilai ekonomis yang cu-kup tinggi. Di samping itu, patin meng-andung protein yang tinggi dan koles-terol yang rendah. Budidaya ikan patin

638 Prastito1 et al.
tidak terlalu sulit karena ikan patin to-leran dangan kandungan oksigen yangrelatif rendah dan merupakan pemak-an segala atau omnivora (Ananda et al.,2015).
Beberapa upaya yang dilakukan untukmengefisienkan pemberian pakan danmeningkatkan pertumbuhan salah sa-tunya dengan menambahkan suplementpada pakan. Manfaat yang diperoleh de-ngan pemberian suplemen diantaranyameningkatkan daya tahan tubuh terha-dap serangan penyakit, melancarkan sis-tem pencernaan, menghemat dalam peng-gunaan pakan dan meningkatkan nafsumakan ikan (Puspitasari, 2018).
Salah satu fitofarmaka yang bisa dija-dikan sebagai antimikrobial adalah te-mulawak (C. xanthorrhiza Roxb). Rim-pang temulawak mengandung zat ber-warna kuning (kurkumin), serat, pati,kalium oksalat, minyak atsiri, dan fla-vonida, zat-zat tersebut berfungsi seba-gai antimikroba/antibakteri, mencegahpenggumpalan darah, anti peradangan,melancarkan metabolisme dan fungsiorgan tubuh (Ditjen, 2000).
Tujuan dari penelitian ini adalah un-tuk mengetahui dosis yang optimal pe-ngaruh ekstrak temulawak pada pak-an buatan dalam peningkatan konsum-si pakan, efisiensi pemanfaatan pakan,dan laju pertumbuhan ikan patin . Ha-sil penelitian ini diharapkan dapat mem-berikan informasi dan gambaran meng-enai salah satu alternatif untuk mening-katkan konsumsi pakan ikan patin me-lalui penambahan ekstrak temulawak.Penelitian ini dilaksanakan pada bulanMei hingga Juli 2017 yang bertempatdi UPTD Balai Benih Ikan Cangkiran,Semarang, Jawa Tengah
MATERI DAN METODE
Ikan yang digunakan dalam penelitianini adalah ikan patin Siam yang bobotawal rata-rata 4,36±0,19 gram/ekor pa-da stadia benih. Padat tebar setiap wa-dah 1 ekor / L dilakukan berdasarkanpenelitian Ananda et al.,(2015). Wadahpemeliharaan yang digunakan dalam pe-nelitian ini adalah bak plastik denganukuran 24 L. Sebanyak 12 buah bakplastik sebagai wadah pemeliharaan dandiisi air sebanyak 15 L. Bak tersebutditutup dengan waring supaya ikan ujitidak keluar. Penelitian ini menggunak-an metode eksperimental Rancangan AcakLengkap (RAL). Penelitian ini meng-gunakan 4 perlakuan dan 3 kali pengu-langan. Perlakuan dalam penelitian iniadalah kombinasi penambahan ekstraktemulawak pada pakan dengan dosis yangberbeda yaitu: perlakuan A (pakan ujidengan penambahan ekstrak temulawakdengan dosis 0 ml), perlakuan B (pak-an uji dengan penambahan ekstrak te-mulawak dengan dosis 6 ml), perlaku-an C (pakan uji dengan penambahanekstrak temulawak dengan dosis 12 ml),dan perlakuan D (pakan uji dengan pe-nambahan ekstrak temulawak dengandosis 18 ml). Pemeliharaan Ikan Patindilakukan selama 42 hari.
Tahapan yang dilakukan sebelum me-nambahkan ekstrak temulawak ke pak-an buatan yaitu pembuatan larutan eks-trak temulawak. Temulawak yang di-gunakan untuk pembuatan ekstrak te-mulawak yaitu sebanyak 250 gram. Pro-ses pembuatan ekstrak temulawak meng-gunakan metode maserasi atau peren-daman selama 2 x 24 jam yang dicam-pur dengan etanol 70%. Proses pencam-puran ekstrak temulawak dengan pak-an menggunakan metode penyemprot-

Effect Addition of Curcuma’s Extract to Pangasius 639
an atau sprayer. Ekstrak temulawak di-campur terlebih dahulu dengan aqua-dest. Setelah dilakukan pencampuranekstrak temulawak dengan aquadest, pak-an uji langsung disemprot dengan la-rutan ekstrak temulawak hingga mera-ta dan di angin-anginkan agar pakantidak berjamur. Pemberian pakan dila-kukan sebanyak 3 kali sehari yakni pa-da pukul 08.00 WIB, 12.00 WIB, dan16.00 WIB. Jumlah pakan yang dibe-rikan per wadah yaitu 5% dari bobotikan.
Perhitungan pertumbuhan bobot mut-lak dihitung dengan menggunakan ru-mus Takeuchi (1988), sebagai berikut:
W =Wt − W0 (1)
dimana W adalah pertambahan bobotmutlak (g); W0 adalah bobot awal ikanuji (gram); dan Wt adalah bobot akhirikan uji (gram)
Perhitungan total konsumsi pakan di-gunakan rumus berdasarkan Pereira et al.(2007), sebagai berikut :
T KP = F1 − F2 (2)
dimana, (TKP) Total konsumsi pakan,(gram); (F1) Jumlah pakan awal (gram);(F2)Jumlah pakan akhir (gram)
Perhitungan nilai efisiensi pemanfaat-an pakan dihitung dengan menggunak-an rumus Tacon (1987), sebagai beri-kut :
EPP =Wt −W0
Fx100% (3)
dimana, (EPP) Efisiensi pemanfaatanpakan (%); (Wt) Bobot total hewan uji
pada akhir penelitian (gram); (Wo) Bo-bot total hewan uji pada awal peneliti-an (gram); dan (F) Jumlah pakan yangdikonsumsi selama penelitian (gram).
Menurut Takeuchi (1988), laju pertum-buhan harian (Survival Growth Rate)ikan dihitung dengan menggunakan ru-mus:
SGR =Wt −W0
W0x tx100% (4)
dimana SGR (Laju pertumbuhan Hari-an, % per hari) ; Wt(Bobot total ikanpada akhir pemeliharaan, gram); Wo (Bo-bot total ikan pada awal pemeliharaan,gram); dan t (Waktu pemeliharaan, ha-ri)
Perhitungan rasio efisiensi protein meng-gunakan rumus Tacon (1987)):
GR =Wt −W0
Pix100% (5)
dimana PER (Rasio Efisiensi Protein,%); Wt (Biomassa ikan uji pada akhirpenelitian, gram); W0 (Biomassa ikanuji pada awal penelitian, gram); dan Pi(Berat pakan yang dikonsumsi x % pro-tein pakan).
HASIL DAN PEMBAHASAN
Hasil penelitian pengaruh penambahanekstrak temulawak pada pakan terha-dap peningkatan konsumsi pakan, efi-siensi pemanfaatan pakan, dan pertum-buhan meliputi nilai W, TKP, EPP, SGR,REP; dan SR. Berdasarkan data per-tumbuhan bobot mutlak total konsum-si pakan, efisiensi pemanfaatan pakan,laju pertumbuhan spesifik, protein efi-siensi rasio dan kelulushidupan padaikan patin selama pemeliharaan dapatdibuat tabel 1.

640 Prastito1 et al.
Tabel 1 Hasil analisis data selama penelitian
Variabel Data PerlakuanA B C D
Bobot Mutlak (g) 60,57±2,89d 116,12±2,82a 107,33±2,32b 93,29±2,18c
TKP (g) 131,39±2,49d 166,67±0,93a 162,06±1,79b 155,94±1,40c
EPP (%) 46,14±3,08c 69,97±1,54a 66,23±0,69a 59,83±1,66b
SGR (%/hari) 1,56±0,06d 2,43±0,04a 2,31±0,04b 2,12±0,03c
PER (%) 1,09±0,05d 2,06±0,05a 1,90±0,04b 1,68±0,04c
SR (%) 88,89±3,85a 97,78±3,85a 95,56±7,70a 91,11±3,85a
keterangan : Nilai dengan superscript yang sama pada baris yang sama menunjukan tidak adanya perbedaan yangnyata
Berdasarkan hasil analisis ragam yangtelah dilakukan menunjukkan bahwa pe-nambahan ekstrak temulawak pada pak-an buatan memberikan pengaruh nyata(P<0,05) terhadap pertumbuhan bobotmutlak dan laju pertumbuhan spesifikikan patin. Pertumbuhan bobot mutlakdisebabkan karena temulawak memi-liki kandungan antibakteri yang dapatmelisiskan racun yang menempel pa-da dinding usus, sehingga penyerapanzat nutrisi menjadi lebih baik dan da-pat memicu pertumbuhan.
Hasil analisis uji duncan didapatkan per-lakuan tertinggi pada pertumbuhan bo-bot mutlak adalah perlakuan dengan pe-nambahan ekstrak temulawak 6 ml/kgpakan, yang kedua perlakuan C denganpenambahan ekstrak temulawak 12 ml/kgpakan sebesar, perlakuan D dengan pe-nambahan ekstrak temulawak 18 ml/kgpakan sebesar dan terendah adalah per-lakuan A dengan penambahan ekstraktemulawak 0 ml/kg pakan sebesar. Per-lakuan B mendapatkan nilai pertumbuh-an bobot mutlak yang tertinggi yaitusebesar 8,03 ± 0,35 g. Hal ini didu-ga karena temulawak memiliki fung-si sebagai imunostimulan secara tidaklangsung yang dimana prosesnya ber-asal dari pemberian pakan yang telahdicampur dengan ekstrak temulawak.Pertumbuhan bobot mutlak terendah pa-da perlakuan 12 ml dan 18 ml dipero-leh nilai rata-rata 7,25±0,53 g dan 6,77
± 0,05 g. Hal ini diduga karena dosisekstrak temulawak yang diberikan cu-kup berlebihan sehingga menyebabkanpertumbuhan bobot yang kurang mak-simal. Menurut Insana and Wahyu (2015)bah-wa semakin besar dosis temulawak yangdiberikan maka akan menyebabkan per-tumbuhan yang tidak stabil.
Penambahan ekstrak temulawak padapakan buatan mempunyai berbagai man-faat bagi tubuh ikan terutama untuk imu-nostimulan dan pertumbuhan bobot mut-lak. Hal ini diduga karena temulawakmengandung senyawa kurkumin. Fung-si dari kurkumin yaitu sebagai pema-cu pertumbuhan yang ditandai denganmeningkatkanya nafsu makan pada ik-an. Menurut (Purwati and Fitriliyani,2016), meningkatnya pertumbuhan di-dukung dengan kesehatan yang baik pa-da ikan dan akan meningkatkan efisi-ensi penyerapan zat makanan untuk me-menuhi kebutuhan hidup dan produksiyang ditunjukkan dengan pertambah-an bobot. Penambahan ekstrak temu-lawak pada pakan buatan mempunyaiberbagai manfaat bagi tubuh ikan ter-utama untuk imunostimulan dan per-tumbuhan bobot mutlak.
Dosis optimum ekstrak temulawak pa-da pertumbuhan bobot mutlak ikan pa-tin diketahui dengan cara melakukanuji Polinomial Orthogonal. Uji Polino-mial Orthogonal pada pertumbuhan bo-

Effect Addition of Curcuma’s Extract to Pangasius 641
bot mutlak diperoleh hubungan yangberpola kuadratik (y = -0,4833x2 + 10,19x+ 63,521) dan R2 = 0,89. Titik opti-mum pada perlakuan B (pemberian eks-trak temulawak 6 ml) didapatkan dosisekstrak temulawak optimal yang dida-pat dari persamaan tersebut yaitu 10,5ml mampu menghasilkan pertumbuh-an bobot mutlak maksimal 117,23 g.Nilai R
2menunjukkan bahwa 89% per-
tumbuhan bobot mutlak dipengaruhi olehpemberian ekstrak temulawak dalam pak-an buatan ikan patin .
Berdasarkan hasil analisa ragam me-nunjukkan bahwa perbedaan pemberi-an frekuensi pakan itu berpengaruh nya-ta (P < 0,05) terhadap total konsumsipakan ikan patin . Total konsumsi pak-an yang tertinggi adalah perlakuan de-ngan dosis 6 ml (Perlakuan B) meng-hasilkan nilai total konsumsi pakan ting-gi dikarenakan frekuensi pakan yangdihasilkan oleh perlakuan dosis 6 ml(perlakuan B) lebih tinggi dibandingk-an dengan perlakuan tanpa penambah-an ekstrak temulawak, perlakuan dosis12 ml (perlakuan C) dan perlakuan do-sis 18 ml (perlakuan D). Hal ini didu-ga temulawak memiliki fungsi untukmeningkatkan nafsu makan pada ikanpatin. Menurut Noviana et al. (2014),bahwa proses makan pada ikan dimu-lai dari tingkat konsumsi nafsu mak-an, kemudian dilanjutkan dengan res-pon terhadap rangsangan dan pencari-an sumber rangsangan, menentukan lo-kasi, jenis pakan dan penangkapan pak-an.
Perlakuan dosis 6 ml (perlakuan B) me-rupakan perlakuan yang menghasilkannilai total konsumsi pakan tertinggi ya-itu 166,67±0,93 gr. Berdasarkan darihasil tersebut maka banyaknya pembe-rian pakan untuk ikan patin dapat mem-
pengaruhi kebutuhan pakan ikan danlaju pertumbuhan pada ikan patin. Pe-ngaruh penambahan ekstrak temulawakpada pakan mampu memberikan penga-ruh nyata terhadap total konsumsi pak-an. Hal tersebut diduga karena ekstraktemulawak memiliki kandungan kurku-min yang fungsinya sebagai imunosti-mulan. Menurut Manoppo and Kolopi-ta (2016), imunostimulan memiliki per-an sebagai penjaga sistem kekebalan tu-buh sehingga dapat memacu peningkat-an konsumsi pakan pada ikan.
Dosis optimum ekstrak temulawak pa-da total konsumsi pakan ikan patin di-ketahui dengan cara melakukan uji Po-linomial Orthogonal. Uji Polinomial Or-thogonal pada pertumbuhan bobot mut-lak diperoleh hubungan yang berpolakuadratik (y = -0,2875x2 + 6,3251x +133,31) dan R2 = 0,89. Titik optimumpada perlakuan B (pemberian ekstraktemulawak 6 ml) didapatkan dosis eks-trak temulawak optimal yang didapatdari persamaan tersebut yaitu 11% mam-pu menghasilkan total konsumsi pak-an maksimal 168,1 g. Nilai R² menun-jukkan bahwa 89% total konsumsi pak-an dipengaruhi oleh pemberian ekstraktemulawak dalam pakan buatan ikan pa-tin .
Berdasarkan hasil penelitian menunjukk-an bahwa penambahan ekstrak temu-lawak pada pakan buatan terdapat in-teraksi (P<0,05) terhadap efisiensi pe-manfaatan pakan (EPP) ikan patin. Ha-sil tersebut menunjukkan bahwa efisi-ensi pemanfaatan pakan pada ikan pa-tin dipengaruhi oleh adanya kandung-an kurkumin yang memiliki pengaruhnyata dalam memanfaatkan pakan. Se-telah dilakukan uji Duncan bahwa per-lakuan yang tertinggi pada efisiensi pe-manfaatan pakan (EPP) (P<0,05) ada-

642 Prastito1 et al.
lah perlakuan B dengan penambahanekstrak temulawak sebanyak 6 ml/kgpakan sebesar 69,67±1,54%. Nilai efi-siensi pemanfaatan pakan (EPP) yangterendah pada perlakuan A dengan pe-nambahan ekstrak temulawak sebanyak0% pakan sebesar 46,14±3,08%. Haltersebut diduga karena pengaruh zat imu-nostimulan yang terdapat pada temu-lawak berperan dalam meminimalisirstress lingkungan pada ikan patin se-hingga dapat berpengaruh terhadap pe-manfatan pakannnya. Hal ini sesuai de-ngan penelitian Wong et al. (2013) yangmembuktikan bahwa pemberian pakanbuatan dengan tambahan Sargassum cris-taefokium 2 g/kg pakan pada ikan kera-pu muara (E. coides) selama 9 hari da-pat meningkatkan sistem kekebalan tu-buh terhadap bakteri Streptococcus sp.dan nilai efisiensi pemanfaatan pakan.
Dosis optimum ekstrak temulawak pa-da efisiensi pemanfaatan pakan ikan pa-tin diketahui dengan cara melakukanuji Polinomial Orthogonal. Uji Polino-mial Orthogonal pada pertumbuhan bo-bot mutlak diperoleh hubungan yangberpola kuadratik (y = -0,2079x2 + 4,3702x+ 47,335) dan R2 = 0,88. Titik opti-mum pada perlakuan B (pemberian eks-trak temulawak 6 ml) didapatkan dosisekstrak temulawak optimal yang dida-pat dari persamaan tersebut yaitu 10,5%mampu menghasilkan efisiensi peman-faatan pakan maksimal 70,30%. NilaiR2 menunjukkan bahwa 88% efisien-si pemanfaatan pakan dipengaruhi olehpemberian ekstrak temulawak dalam pak-an buatan ikan patin .
Ikan patin yang dipelihara selama pe-nelitian mengalami pertambahan bobotdan panjang. Laju pertumbuhan spesi-fik berfungsi untuk menghitung persen-tase pertumbuhan berat ikan per hari.
Berdasarkan dari hasil analisis ragam(P > 0,05) nilai SGR menunjukkan bah-wa pemberian pakan campuran ekstraktemulawak dengan dosis yang berbe-da memberikan pengaruh yang sangatnyata pada ikan patin . Pengaruh ter-sebut diduga karena kandungan nutri-si di dalam pakan digunakan sebagi-an besar untuk pertumbuhan ikan patin. Menurut Insana and Wahyu (2015),bahwa perbedaan nilai laju pertumbuh-an disebabkan oleh kandungan jumlahdosis temulawak yang diberikan. Per-bedaan dosis temulawak yang diberik-an berpengaruh terhadap peningkatankonsumsi pakan dan efisiensi peman-faatan pakan.
Perlakuan dosis 6 ml merupakan per-lakuan yang menghasilkan nilai SGRyaitu 2,43±0,04. Perlakuan dosis 12 mlmenghasilkan nilai SGR yaitu 2,31±0,04.Perlakuan dosis 18 ml menghasilkannilai SGR yaitu 2,12±0,03. Perlakuantanpa dosis menghasilkan nilai SGR ter-endah yaitu 1,56±0,06. Nilai SGR ter-tinggi yaitu pada perlakuan dosis 6 ml,hal ini menunjukkan bahwa pertumbuh-an berkaitan dengan frekuensi pakanyang diberikan. Nilai SGR tertinggi ke-dua dan ketiga yaitu pada perlakuan do-sis 12 dan 18 ml, hal ini diduga ka-rena pengaruh dosis temulawak yangterlalu tinggi yang menyebabkan nutri-si pada pakan mudah hilang sehinggadapat mengganggu organ pencernaanpada ikan. Sedangkan nilai SGR ter-endah yaitu pada perlakuan tanpa do-sis temulawak, hal ini diduga karenapakan yang diberikan pada perlakuantanpa dosis temulawak tidak terdapatekstrak temulawak yang menyebabkanefisiensi pemanfaatan pakan dan nafsumakan ikan menurun. Menurut Mulya-di et al. (2010), bahwa Kekurangan ma-

Effect Addition of Curcuma’s Extract to Pangasius 643
kanan dan energi yang dibutuhkan da-pat mengakibatkan kekurangan pertum-buhan karena energi digunakan untukmemelihara fungsi tubuh dan pergerak-an.
Pengaruh Penambahan ekstrak temula-wak pada pakan buatan mampu mem-berikan pengaruh yang nyata terhadaplaju pertumbuhan spesifik. Hal ini di-duga karena pengaruh kandungan kur-kumin dari temulawak. Kandungan kur-kumin pada temulawak selain berfung-si sebagai imunostimulan, juga untukmeningkatkan daya cerna dan sebagaipenunjang pertumbuhan. Menurut Pur-wati and Fitriliyani (2016), laju pertum-buhan dipengaruhi oleh makanan, ke-padatan, keturunan dan kecepatan per-tumbuhan bobot mutlak.
Dosis optimum ekstrak temulawak pa-da laju pertumbuhan spesifik ikan pa-tin diketahui dengan cara melakukanuji Polinomial Orthogonal. Uji Polino-mial Orthogonal pada pertumbuhan bo-bot mutlak diperoleh hubungan yangberpola kuadratik (y = -0,0073x2 + 0,1582x+1,6069) dan R2 = 0,89. Titik optimumpada perlakuan B (pemberian ekstraktemulawak 6 ml) didapatkan dosis eks-trak temulawak optimal yang didapatdari persamaan tersebut yaitu 10,8 mlmampu menghasilkan laju pertumbuh-an spesifik maksimal 2,46%/hari. Ni-lai R2 menunjukkan bahwa 89% lajupertumbuhan spesifik dipengaruhi olehpemberian ekstrak temulawak dalam pak-an buatan ikan patin .
Berdasarkan hasil penelitian menunjukk-an bahwa pemberian ekstrak temula-wak pada pakan buatan terdapat inte-raksi (P<0,05) terhadap rasio efisien-si protein (REP) ikan patin . Hal ter-sebut menunjukkan bahwa rasio efisi-ensi protein pada ikan patin dapat di-
pengaruhi adanya perbedaan taraf pa-da setiap dosis faktor. Setelah dilakuk-an uji Duncan perlakuan yang tertinggipada REP adalah perlakuan B denganpenambahan ekstrak temulawak.
Rasio efisiensi protein (REP) terbaikyaitu pada perlakuan B hal tersebut di-duga menjadi salah satu dosis yang te-pat untuk ikan patin. Hal ini diduga do-sis temulawak yang diberikan mampumengefisienkan pakan khususnya pro-tein cell dengan enzim protease. Sedangk-an nilai rasio efisiensi protein pada per-lakuan C dan D di nilai kurang baik.Hal ini dikarenakan dosis dari ekstraktemulawak tidak mampu mengefisienk-an pakan khususnya dari protein celldengan enzim protease. Menurut Ma-rzuqi et al. (2012), pemberian pakanyang memiliki kadar protein lebih ting-gi,semakin banyak protein pada pak-an yang dipergunakan oleh ikan untukpertumbuhannya. Akibatnya pertambah-an berat tubuh ikan semakin tinggi.b)sebanyak 6 ml/kg pakan sebesar 2,06±0,05dibandingkan dengan perlakuan lain-nya. Menurut Pratama et al. (2015), bah-wa pakan yang berkualitas adalah pak-an yang memiliki kandungan nutrisi yanglengkap dan seimbang sesuai dengankebutuhan ikan, serta memiliki kandung-an energi total yang optimum untuk me-nunjang pertumbuhan yang maksimumpada ikan tersebut.
Dosis optimum ekstrak temulawak pa-da rasio efisiensi protein ikan patin di-ketahui dengan cara melakukan uji Po-linomial Orthogonal. Uji Polinomial Or-thogonal pada pertumbuhan bobot mut-lak diperoleh hubungan yang berpolakuadratik (y = -0,0083x2 + 0,1767x +1,1446)dan R2 = 0,88. Titik optimum pada per-lakuan B (pemberian ekstrak temula-wak 6 ml) didapatkan dosis ekstrak te-

644 Prastito1 et al.
mulawak optimal yang didapat dari per-samaan tersebut yaitu 10,6 ml mam-pu menghasilkan rasio efisiensi prote-in maksimal 2,08%. Nilai R2 menun-jukkan bahwa 88% rasio efisiensi pro-tein dipengaruhi oleh pemberian eks-trak temulawak dalam pakan buatan ik-an patin
Berdasarkan hasil analisis ragam me-nunjukkan bahwa penambahan ekstraktemulawak pada pakan buatan ikan pa-tin tidak berpengaruh nyata (P>0,05)terhadap tingkat kelulushidupan. Nilairata-rata kelulushidupan tertinggi ada-lah perlakuan B sebesar 97,78±3,85%dan adapun nilai terendah adalah per-lakuan A 88,89±3,85%. Secara statis-tik nilai kelulushidupan dari perlakuanA, perlakuan B, perlakuan C dan perla-kuan D tidak ada yang lebih baik. Me-nurut Handayani et al. (2015), bahwatingginya kelulushidupan ikan uji dise-babkan karena kualitas air yang digu-nakan masih dalam batas toleransi se-hingga mendukung untuk pemelihara-an ikan.
SIMPULAN
Kesimpulan yang didapat dari peneliti-an “Pengaruh Penambahan Ekstrak Te-mulawak Terhadap Peningkatan Kon-sumsi Pakan, Efisiensi Pemanfaatan Pak-an, dan Pertumbuhan Ikan Patin ” ada-lah Dosis optimum dari penambahanekstrak temulawak terhadap total kon-sumsi pakan adalah 11 ml mampu meng-hasilkan TKP maksimal 168,1 g, efsi-ensi pemanfaatan pakan adalah 10,5 mlmampu menghasilkan EPP maksimal70,3% dan laju pertumbuhan spesifikadalah 10,8 ml menghasilkan SGR mak-simal 2,46%/hari.
Acknowledgements Terima kasih penulis ucapkan ke-pada kepala UPTD Cangkiran Semarang yang telahmenyediakan tempat dan fasilitas untuk pelaksanaanpenelitian ini dan semua pihak yang telah membantukelancaran penelitian ini.
Pustaka
Ananda, T., Rachmawati, D., and Sa-midjan, I. (2015). Pengaruh pa-pain pada pakan buatan terhadappertumbuhan ikan patin (pangasiushypopthalmus). Journal of Aqua-culture Management and Technolo-gy, 4(1):47–53.
Ditjen, P. (2000). Parameter standarumum larutan tumbuhan obat. cetak-an pertama. Depkes RI. Jakarta. Hal,pages 13–31.
Handayani, I., Nofyan, E., and Wija-yanti, M. (2015). Optimasi ting-kat pemberian pakan buatan terha-dap pertumbuhan dan kelangsunganhidup ikan patin jambal (pangasiusdjambal). Jurnal Akuakultur RawaIndonesia, 2(2):175–187.
Insana, N. and Wahyu, F. (2015).Subtitusi tepung temulawak (curcu-ma xanthorhiza sp) pada pakan de-ngan dosis berbeda terhadap pertum-buhan dan sintasan benih ikan ni-la (oreochromis niloticus). OCTO-PUS: JURNAL ILMU PERIKANAN,4(2):381–391.
Manoppo, H. and Kolopita, M. E.(2016). Penggunaan ragi roti(saccharomyces cerevisiae) sebagaiimunostimulan untuk meningkatkanresistensi ikan mas (cyprinus carpiol) terhadap infeksi bakteri aeromo-nas hydrophila. e-Journal BUDIDA-YA PERAIRAN, 4(3).
Marzuqi, M., Astuti, N. W. W., andSuwirya, K. (2012). Pengaruh ka-dar protein dan rasio pemberian pak-an terhadap pertumbuhan ikan kera-

Effect Addition of Curcuma’s Extract to Pangasius 645
pu macan (epinephelus fuscogutta-tus). Jurnal Ilmu dan Teknologi Ke-lautan Tropis, 4(1):55–65.
Mulyadi, Usman, and Suryani (2010).Pengaruh frekuensi pemberian pak-an yang berbeda terhadap pertum-buhan dan kelulushidupan benih ik-an silais (ompok hypophthalmus).berkala perikanan terubuk. BerkalaPerikanan Terubuk, 38(2):21–40.
Noviana, P. et al. (2014). Penga-ruh pemberian probiotik dalam pak-an buatan terhadap tingkat konsumsipakan dan pertumbuhan benih ikannila (oreochromis niloticus). Jour-nal of Aquaculture Management andTechnology, 3(4):183–190.
Pereira, L., Riquelme, T., and Hosoka-wa, H. (2007). Effect of three pho-toperiod regimes on the growth andmortality of the japanese abalone ha-liotis discus hannai ino. Journal ofShellfish Research, 26(3):763–768.
Pratama, M. A. et al. (2015). Penga-ruh berbagai rasio e/p pakan berka-dar protein 30% terhadap efisiensipemanfaatan pakan dan pertumbuh-an ikan mas (cyprinus carpio). Jour-nal of Aquaculture Management andTechnology, 4(4):74–81.
Purwati, H. H. and Fitriliyani, H.(2016). Pengaruh penambahan vi-tamin c dan ekstrak temulawak pa-da pakan komersil terhadap pertum-buhan post larva ikan papuyu (ana-bas testudineus bloch). Fish scienti-ae (Jurnal Ilmu-Ilmu Perikanan danKelautan), pages 60–72.
Puspitasari, D. (2018). Efektivitas su-plemen herbal terhadap pertumbuh-an dan kululushidupan benih ikan le-le (clarias sp.). Jurnal Ilman: JurnalIlmu Manajemen, 5(1).
Tacon, A. G. (1987). The nutrition andfeeding of farmed fish and shrimp; a
training manual. 1: The essential nu-trients.
Takeuchi, T. (1988). Laboratory work-chemical evaluation of dietary nutri-ents. Fish nutrition and mariculture,pages 179–226.
Wong, S.-L., Gao, L.-H., Chang, C.-C., and Cheng, W. (2013). Theeffect of hot-water extract of sargas-sum cristaefolium on growth, innateimmune responses and resistance ofgrouper, epinephelus coiodes. J. Fi-sh. Soc, 40(1):11–26.

646 Prastito1et al.

AQUASAINS(Jurnal Ilmu Perikanan dan Sumberdaya Perairan)(Vol 7 No. 1 Tahun 2018)
THE GROWTH KINETICS OF Bacillus sp. D2.2 ATDIFFERENT pH AND SALINITY
Esti Harpeni1 · Supono2 · Dwi Risca Septiani2
Ringkasan Isolate D2.2 is a bacterialisolate with 97% homology level withBacillus sp. This isolate is a biocontrolbacterium capable of inhibiting the grow-th of Gram positive bacteria and Gramnegative bacteria. The use of biocon-trol bacteria is one solution of diseaseproblem in cultivation organism. Priorto being applied to the field, D2.2 iso-lates need to go through a series of tes-ts, one of which is growth kinetics tes-ting under various environmental con-ditions, such as different degrees of pHand salinity. This is because environmen-tal factors can affect the rapid growthof bacteria. The purpose of this studywas to study the bacterial growth kine-tics of D2.2 at different pH and sali-nity. The growth kinetics was observedby measuring optical density (OD) thro-ugh a method of turbidimetry using aspectrophotometer at a wavelength of625 nm to the stage of death. The resul-ts showed that the fastest growth ratewas found at 20 ppt salinity with 0.179h-1 and generation time of 5,588 ho-
1)Marine Science Study Program, University of Lam-pung, Bandar Lampung, Indonesia; 2)Aquaculture Stu-dy Program, University of Lampung, Bandar Lam-pung, Indonesia.E-mail: [email protected]
urs. While in all pH treatments, gene-ration time and growth rates achievedall treatments have the same pattern.
Keywords growth kinetics, Bacillus sp. D2.2,pH, salinity
Received : 12 September 2018
Accepted : 23 Oktober 2018
PENDAHULUAN
The environment acts as an indicatorof the state of the organism in it. If theenvironmental conditions are bad thenthe organisms inside it become stres-sed and decrease in the body’s resis-tance. As a result, pathogenic diseasessuch as bacteria can easily attack aqua-culture organisms and cause infectiousdiseases. In a shrimp pond, Kharismaand Manan (2019) stated that the abun-dance of Vibrio sp. can be used as anearly detection of Vibriosis disease. V.harveyi, V. alginolyticus and V. para-haemolitycus are the types of bacteriathat most commonly cause Vibriosis inshrimp. Furthermore, bacteria can alsocause infection in fish, such as Gram-positive bacteria Staphylococcus aure-us which is a cause of food-poisoning

648 Esti Harpeni1 et al.
due to production of enterotoxins the-refore the high population of this ba-cteria indicates the general quality offish (Ali, 2014), while from Gram ne-gative bacteria such as Aeromonas hydro-phila can result in a significant redu-ction in fisheries production due to hi-gh mortality (Pachanawan et al., 2008).
Infectious diseases should be treated asearly as possible with appropriate tre-atment. Prevention and treatment aremostly done by giving antibiotics andother chemicals, but the use of antibi-otics and chemical ingredients to era-dicate pathogenic bacteria in the longterm can have a negative effect on thesurrounding environment and resistan-ce to pathogenic bacteria. Some weak-nesses in the use of antibiotic ingredi-ents include, can make the bacteria re-sistant to antibiotics, can cause residu-es in the body of cultivating organisms,damage the environment and can enda-nger human health to consume aqua-culture organisms that have been givenantibiotic substances(Kümmerer, 2004;Kesarcodi-Watson et al., 2008). This iswhat causes developed countries to pro-hibit the use of antibiotics and fisheryproducts that contain antibiotic residu-es (Carlet et al., 2012).
The use of biocontrol bacteria is onesolution to solving disease problems inaquaculture organisms. In addition, theuse of biocontrol bacteria is also envi-ronmentally friendly and can suppressthe growth of pathogenic bacteria. Bio-control bacteria utilize the antagonisticrelationship between organisms to getspace for movement and food. Biocon-trols of pathogens have been reportedpotential in aquaculture such as bio-control of vibriosis in shrimp (Chytha-
nya et al., 2002; Widanarni et al., 2003;Ravi et al., 2007).
Bacillus sp. D2.2 is isolated from tra-ditional shrimp ponds in Mulyosari Vi-llage, Pasir Sakti District, East Lam-pung Regency and able to inhibit the invitro growth of Vibrio harveyi (Setya-wan et al., 2014). Moreover, this iso-late was proven to be non-pathogenicbacteria (Hardiyani et al., 2016). Be-fore being applied to the field, isola-te Bacillus sp. D2.2 needs to be testedfor the activity of antimicrobial com-pounds which are influenced by seve-ral factors such as pH, salinity, stabili-ty of antibacterial compounds, tempe-rature, inoculums of incubation micro-organisms, and metabolic activity (Iri-anto, 2006).
Kinetics of bacterial growth is one wayto determine the speed of cell biomassproduction and the influence of the en-vironment (nutrient restricted) on thespeed of cell growth (Schaechter, 2009).Kinetics of bacterial growth is influen-ced by environmental factors such astemperature, pH and NaCl (Sutherlandet al., 1994). The aim of this study wasto observe the growth of Bacillus sp.D2.2 in media using various ranges ofpH and salinity as controlling factors.
MATERI DAN METODE
Bacterial strains and media
Bacillus sp. D2.2 isolated from tradi-tional tiger shrimp in East Lampung(Setyawan et al., 2014) was inocula-ted on seawater complete-agar (SWC,5 g bacto peptone (Oxoid, England),1 g yeast extract (Oxoid, England), 3ml glycerol, 15 g bacto agar (Oxoid,England), 750 ml seawater, and 250 ml

The Growth Kinetisc of Bacillus D2.2 649
distilled water) and then transferred toSWC-broth (without agar) with rotaryshaker at 140 rpm for 24 h at 30°C (Har-peni et al., 2018). V. alginolyticus, A.hydrophila, and S. aureus were bacte-rial collections of the Laboratory of Fi-sheries Culture, Department of Aqua-culture, University of Lampung and cul-tured on trypticase soy agar (TSA) me-dium (Oxoid, England).
Bacterial growth stage
Isolates of Bacillus sp. D2.2 were cul-tured on SWC broth media at salini-ty 0, 10, 20 and 30 ppt; pH 6, 7 and8 and incubated at 300 C. Cell densi-ty measurement (optical density) wascarried out every 3 hours using a spe-ctrophotometer at 625 nm wavelengthto the death stage. Calculations of Ba-cillus sp. D2.2 growth rate and genera-tion time were calculated based on thefollowing formulas (Augustin et al., 1999):
µ =log10Xt − log10X0
0.301t(1)
where: µ = Growth rate; Xt= Total den-sity at exponential time; X0 = Numberof initial densities at exponential time;and t = Exponential growth time
n =logN − logN0
0.301(2)
where: n = Number of Generations; LogN = End cell number; and Log N0 =Number of initial cells
Generation time = t/n (3)
where: t = Exponential growth time;and n = Number of Generations
Production of antibacterial substances
Antibacterial extraction was a broth ofSWC fermentation medium and incu-bated with agitation at 150 rpm until
the death stage at 300 C. After incu-bation, it was centrifuged at 5000 rpmfor 25 minutes to separate the superna-tant and bacterial cell pellet (Isnanse-tyo et al., 2009). Bacterial cell pelletwas used in this study based on Se-tyawan et al. (2014). Bacterial cell pe-llet was washed using phosphate bu-ffer saline (PBS) and 15 ml of the sa-me buffer was added. The suspensionof bacterial cell was then broken downusing an ultrasonicator for 5 minutes.The bacterial cell suspension was cen-trifuged at a speed of 5000 rpm for 25minutes to obtain the supernatant. Thesupernatant was divided into two por-tions. The first portion was added wi-th 50 ml distilled water and then extra-cted with ethyl acetate twice. The ethylacetate layer was evaporated using arotary evaporator at 50ºC, while the se-cond portion was saturated with am-monium sulfate with the same treatmentas the first portion.
Antagonistic spectrum activity of Baci-llus sp. D2.2
Antagonistic spectrum activity of Ba-cillus sp. D2.2 in different range of sa-linity and pH was carried out using di-ffusion method. About 24 h culturesof V. alginolyticus, A. hydrophila, andS. aureus in 10 ml trypticase soy bro-th (TSB) media (Oxoid, England) we-re inoculated respectively into a petridish containing TSA media. Then twopaper discs with a diameter of 6 mmwere impregnated with the extractionof ethyl acetate cells and saturation ofammonium sulfate cells respectively. Twen-ty five ppm of Amoxicillin (PT IndoFarma, Bekasi) was used as positive con-trol while distilled water was used asnegative control. Those paper discs we-re placed on agar plate of TSA medi-

650 Esti Harpeni1 et al.
um and then incubated at 300 C for 24h. After the incubation period, the di-ameter of the inhibition zone formedaround the paper disc was measured.
HASIL DAN PEMBAHASAN
Growth Stages in Different Salinity Tre-atments
Different salinity treatments showed thatthese bacteria had fast growth at 30 pptsalinity and had slow growth at 0 pptsalinity. From the graph, it can be se-en that the lag stage in all treatmentsexcept 0 ppt salinity had the same ti-me period that occured until the 9 hourthen entered the exponential stage. Theexponential stage in salinity treatmentof 10, 20 and 30 ppt showed the si-milar pattern until around 30 hours. Incontrast to the treatment of 0 ppt sali-nity, the exponential stage occured lo-nger until the 94 hours. There was nostationary stage at 0 ppt salinity reco-rded, since bacterial cells directly en-countered the death stage. Furthermo-re, salinity 20 and 30 ppt faced the sta-tionary stage at 36 hours while salini-ty 10 entered the stationary stage in 60hours then met the death stage (Figu-re 1). The salinity treatment of 20 ppthas a long stationary stage from 30-54hours (18 hours stationary stage).
Bacillus sp. D2.2 had different grow-th stages in reaching the death stage.Zwietering et al. (1991)and Buchananet al. (1997) stated that the growth ra-te of bacteria shows several stages orphases, a lag stage that is followed byan exponential stage, and finally it sho-ws a decreasing growth rate down tozero stationary stage. If linked betweenpopulation growth acceleration and a
Gambar 1 The growth of Bacillus sp. D2.2 in diffe-rent range of salinity
certain time will form a growth curve.Pelczar et al. (1981) states that the lagstage (adaptation) is a stage where the-re is no population growth, cells onlyexperience changes in chemical com-position, increase in size and increasein intracellular substances. The adap-tation process includes the synthesis ofnew enzymes that are appropriate to themedia and recovery of toxic metaboli-tes such as acids, alcohols and bases(Purwoko, 2009).
Among the treatments carried out at 0ppt salinity the lag stage had remai-ned until the 27th hour then enteredthe exponential stage until the 94th ho-ur. Different from 30 ppt salinity trea-tment, where the stage lag occurs only6 hours, then went to the exponenti-al stage faster until approximately the40th hour. While in salinity of 10 pptand 20 ppt the growth stage was notmuch different, the lag stage occurs inthe 12th hour. Bacillus sp. D2.2 rea-ched exponential stage faster in sali-nity 30 ppt than presumably becausethe isolate had been taken from tradi-tional shrimp ponds in East Lampungwith salinity ranged from 25-30 ppt.

The Growth Kinetisc of Bacillus D2.2 651
Gambar 2 The growth of Bacillus sp. D2.2 in diffe-rent range of pH
So that, Bacillus sp. D2.2 did not ne-ed longer time to adapt to the salinityculture condition.
Growth Stages in Different pH Trea-tments
Similar with those in salinity treatmen-ts, the lag stage in the treatment of pH6, 7 and 8 occured until the 9th ho-ur then entered the exponential stage.While the stationary stage the pH tre-atment began at the 54th hour beforeexperiencing a decrease to the death sta-ge (Figure 2). Based on the graph ofdensity comparison on pH treatment sho-ws, the highest to lowest density in arow are treatment pH 6, pH 8 and pH7. The treatment of pH 7 shows the lo-west density of 1.23 x 109 CFU/ml.
The growth of Bacillus sp. D2.2 in pHtreatments exhibited similar pattern. Thebacterial exponential stage D2.2 had occu-red from the 6th hour until the 48th ho-ur. This showed that all three pH trea-tments did not significantly affect thebacterial growth. It was indicated thatthe growth of Bacillus sp. D2.2 werestable in pH range 6-8.
Generation Time, Growth Rate and Ha-rvest Time of Bacillus sp. D2.2
Tabel 1 Generation time and growth rate of Bacillussp. D2.2
TreatmentsGeneration time Growth rate Harvest time
(hours) (hour-1) (hours)0 ppt 14.783 0.069 9510 ppt 7.613 0.134 6520 ppt 5.588 0.179 6030 ppt 5.976 0.173 57pH 6 7.311 0.136 66pH 7 7.395 0.136 66pH 8 7.048 0.156 66
The treatment of 0 ppt salinity the lo-west bacterial growth rate reached 0.069hour-1 with the longest generation ti-me reaching 14.78 hours, while incre-asing salinity up to 30 ppt had shor-ten generation time more than half andhad increased generation rate. Harve-sing times of Bacillus sp. D2.2 isolateswere usually produced as cultures rea-ch the end of stationary stage or befo-re the death stage. They also had simi-lar trend to the generation time and thegrowth rate. They had been harvestedfaster in higher salinity, range from 57to 95 hours (Table 1). Compare to sa-linity treatments, generation time andgrowth rate of Bacillus sp. D2.2 amongall pH treatments were alike, just about7 hours and 0.136 hour-1 sequentially,so that the isolate could be harvestedtogether in 66 hours (Table 1).
The exponential stage of Bacillus sp.D2.2 in this study was the longest stagewhich required one until three days be-fore entering the stationary stage. Ba-sed on Pelczar et al. (1981), during theexponential stage, bacterial cells willdivide at a constant rate, bacteria dou-ble at the same rate, metabolite activityis also constant and the state of growthis balanced.
Entering the static stage, bacteria willproduce secondary compounds such asantibiotics. Before entering the death

652 Esti Harpeni1 et al.
stage some bacteria are able to survivethe static stage from hours to severalyears before finally entering the stageof death (Purwoko, 2009). Bacillus sp.D2.2 had a static stage that lasts for se-veral hours before finally entering thedeath stage (Figure 1).
Entering the stage of death, all cellswill die faster in a few days to severalmonths (Pelczar et al., 1981). Bacillussp. D2.2 utilized glycerol as the ma-in carbon source and other sources inthe media used. The provision of car-bon sources with different concentra-tions can also improve the life stage ofbacteria (Devianto and Kardena, 2010).
Growth is an increase in mass or totalcell number (for example in a culture),the rate of growth is the increase in thenumber of cells per unit of time, whilethe generation time is the time intervalneeded by a cell to divide. The resul-ts showed that salinity treatment of 20and 30 ppt had the fastest growth ra-te (Table 1), while all of pH treatmen-ts exhibited the average growth rate of0.136 hour-1. This study showed thatthe higher the salinity accelerated thegrowth rate and generation time whiledifferent range of pH did not result di-fferent value of either the growth rateor generation time. Arellano-Carbajaland Olmos-Soto (2002) reported, theirisolate, Bacillus LMM-12 had the grow-th rate 0.75 h-1.
Observation of bacterial growth stageD2.2 aimed to determine harvest timeto produce antibacterial substances. Inthis study the production time of an-tibacterial compounds was carried outat the stationary stage entering the de-ath stage where the time of each tre-atment was different, at 0 ppt salini-ty treatment at the 95th hour, 10 ppt
salinity at 65 hours, 20 ppt salinity inthe 60th hour and 30 ppt salinity in the57th hour. While at the pH treatmentthe production time is at the same timewhich was at 66th hour. The predictionof the harvest time was suspected atthe time that Bacillus sp. D2.2 had pro-duced secondary metabolites or anti-bacterial compounds. Isnansetyo et al.(2009) stated in their study that bacte-ria from the genus Pseudoalteromonasfrom S2V2 strain produced the highestantimicrobial compounds at 96th houror in the stationary stage to the deathstage. In this stage the highest activi-ty of the bacteria more likely releasedsecondary or antimicrobial metabolites(Tsai and Su, 1999; Arellano-Carbajaland Olmos-Soto, 2002; Liu et al., 2006).
Antagonistic spectrum activity of Baci-llus sp. D2.2
Antibacterial substance produced by Ba-cillus sp. D2.2 was present in the ethylacetate extract of bacterial cells. Baci-llus sp. D2.2 inhibited the growth ofV. alginolyticus, A. hydrophila, and S.aureus in various salinities. The inhibi-tion diameter of Bacillus sp. D2.2 aga-inst V. alginolyticus ranged from 6-14mm, against A. hydrophila ranged from2-7 mm, and against S. aureus rangedfrom 2-4 mm. On the other hand, theinhibition diameter that had been for-med by Bacillus sp. D2.2 against thepathogenis bacteria was relatively sma-ller in different pH, ranged from 2-9mm (Table 2).
Although smaller, antibacterial substan-ce produced by Bacillus sp. D2.2 wasalso present in the ammonium sulfateprecipitate. Mostly the inhibition zo-ne was produced by Bacillus sp. D2.2against V. alginolyticus and A. hydro-phila from different salinities, range from

The Growth Kinetisc of Bacillus D2.2 653
Tabel 2 Inhibition diameter (mm) of Bacillus sp. D2.2 in different salinity and pH
Pathogenic bacteria Salinity/pH Extraction of ethyl acetate Saturation of ammonium sulfate Positive control (antibiotic) Negatif control (aquadest)
V. alginolyticus
0 ppt 8 6 20 -
10 ppt 14 - 18 -
20 ppt 6 6 23 -
30 ppt 10 3 20 -
pH 6 4 - 20 -
pH 7 9 4 19 -
pH 8 5 3 23 -
A. hydrophila
0 ppt 4 - 17 -
10 ppt 3 4 17 -
20 ppt 2 3 19 -
30 ppt 7 4 18 -
pH 6 - - 17 -
pH 7 2 - 17 -
pH 8 3 - 19 -
S. aureus
0 ppt 3 - 19 -
10 ppt 2 - 17 -
20 ppt 4 - 19 -
30 ppt 2 2 24 -
pH 6 - - 19 -
pH 7 3 - 17 -
pH 8 2 3 19 -
3-6 mm, but was barely detected fromdifferent range of pH (Table 2).
This study indicated that antibacteri-al substance produced by Bacillus sp.D2.2 mostly from the ethyl acetate extra-ct. Abraham (2004) in his research onthe antibacterial activity of Alteromo-nas P7 stain also showed the similarresults, since it was suspected that an-tibacterial compounds were still storedin the cell wall of the bacteria. Ethylacetate is a specific solvent that can dis-solve water up to 3% and solubility ofup to 8% in water at room tempera-ture. Ethyl acetate is a medium polarsolvent (semi polar) as well as organiccompounds which are esters of ethanoland acetic acid (Daluningrum, 2009).Ethyl acetate was used to obtain secon-dary metabolites in Bacillus sp. D2.2which function as antibacterial compo-unds while ammonium sulfate was usedto obsrve enzyme activity. The produ-ction of antibacterial substances obta-ined by extraction and cell saturationshowed the presence of clear zones in
the diffusion test using bacteria V. algi-nilyticus, A. hidrophyla and S. aureus.This proved that the Bacillus sp. D2.2had antibacterial compounds that cou-ld inhibit the growth of the pathogenicbacteria in the diffusion test. The resul-ts showed that the best inhibition zonewas found in cell extraction tested onV. alginolyticus. This is consistent withresearch conducted by Setyawan et al.(2014) where cell extracts tested on V.alginolyticus obtained the highest inhi-bition zone.
The study showed that Bacillus sp. D2.2was able to inhibit the growth of patho-genic bacteria from both Gram-positiveand Gram-negative, but when compa-red with the positive control, there wasstill not enough potential as an antiba-cterial compound producer. Neverthe-less, Pan et al. (2009) scored bacteriathat had diameter of the zone between0 and 3 mm as weak, between 3 and6 mm as good, and more than 6 mmas a strong inhibition. The best inhibi-tion activity of on types of Bacillus sp.

654 Esti Harpeni1 et al.
D2.2 was present against Gram-negativebacteria, V. alginolytcus. In conclusion,salinities created different growth pat-tern of Bacillus sp. D2.2. The isolatereached the fast growth rate in salinity20 and 30 ppt with 0.179 and 0.173 h-1 and generation time of 5.59 and 5.97hours respectively. In contrast, the iso-late grew in similar pattern when thepH treatments were applied. The highestantibacterial activity (14 mm inhibitionzone) of Bacillus sp. D2.2 was foundin the antagonistic test against V. algi-nolyticus with ethyl acetate extract.
SIMPULAN
In conclusion, salinities created diffe-rent growth pattern of Bacillus sp. D2.2.The isolate reached the fast growth ra-te in salinity 20 and 30 ppt with 0.179and 0.173 h-1 and generation time of5.59 and 5.97 hours respectively. In con-trast, the isolate grew in similar pat-tern when the pH treatments were ap-plied. The highest antibacterial activi-ty (14 mm inhibition zone) of Bacillussp. D2.2 was found in the antagonistictest against V. alginolyticus with ethylacetate extract.
Pustaka
Abraham, T. (2004). Antibacterial ma-rine bacterium deter luminous vi-briosis in shrimp larvae. NAGA,WorldFish Center Quarterly, 27(3-4):28–31.
Ali, H. H. (2014). Isolation and iden-tification of staphylococcus bacteriafrom fish of fresh water and its anti-biotics sensitivity in mosul city. Bas-rah Journal of Veterinary Research.,13(1):33–42.
Arellano-Carbajal, F. and Olmos-Soto,J. (2002). Thermostable α-1, 4-andα-1, 6-glucosidase enzymes frombacillus sp. isolated from a mari-ne environment. World Journalof Microbiology and Biotechnology,18(8):791–795.
Augustin, J.-C., Rosso, L., and Carlier,V. (1999). Estimation of temperatu-re dependent growth rate and lag ti-me of listeria monocytogenes by op-tical density measurements. Journalof microbiological methods, 38(1-2):137–146.
Buchanan, R., Whiting, R., and Da-mert, W. (1997). When is simplegood enough: a comparison of thegompertz, baranyi, and three-phaselinear models for fitting bacterialgrowth curves. Food microbiology,14(4):313–326.
Carlet, J., Jarlier, V., Harbarth, S.,Voss, A., Goossens, H., and Pittet,D. (2012). Ready for a world witho-ut antibiotics? the pensières antibio-tic resistance call to action.
Chythanya, R., Karunasagar, I., andKarunasagar, I. (2002). Inhibition ofshrimp pathogenic vibrios by a ma-rine pseudomonas i-2 strain. Aqua-culture, 208(1-2):1–10.
Daluningrum, I. (2009). Penapisanawal komponen bioaktif dari kerangdarah (anadara granosa) sebagai se-nyawa antibakteri. Skripsi ProgramStudi Teknologi Hasil Perikanan Fa-kultas Perikanan dan Ilmu KelautanInstitut Pertanian Bogor.
Devianto, L. and Kardena, E. (2010).Pengaruh glukosa terhadap produk-si biosurfaktan oleh azotobacter vi-nelandii dan pengaruh biosurfaktanterhadap biodegradasi tph oleh kon-sorsium bakteri petrofilik. Biotech-nology, 8(1):1–10.

The Growth Kinetisc of Bacillus D2.2 655
Hardiyani, S., Harpeni, E., Setyawan,A., and Supono, S. (2016). Pathoge-nicity and in vivo study of local iso-late bacillus sp. d2. 2 at the vanna-mei culture (litopenaeus vannamei).AQUASAINS, 5(1):441–446.
Harpeni, E., Santoso, L., Supono, S.,Wardiyanto, W., Widodo, A., andYolanda, L. (2018). Effects of di-etary probiotic bacillus sp. d2. 2and prebiotic sweet potato extracton growth performance and resistan-ce to vibrio harveyi in pacific whiteshrimp, litopenaeus vannamei. Aqu-acultura Indonesiana, 18(2):55–61.
Irianto, K. (2006). Mikrobiolo-gi menguak dunia mikroorganisme.Yrama Widya. Bandung, 1:164–172.
Isnansetyo, A., Istiqomah, I., Sinansa-ri, S., Hernawan, R. K., Widada, J.,et al. (2009). A potential bacterialbiocontrol agent, strain s2v2 againstpathogenic marine vibrio in aquacul-ture. World Journal of Microbiolo-gy and Biotechnology, 25(6):1103–1113.
Kesarcodi-Watson, A., Kaspar, H., La-tegan, M. J., and Gibson, L. (2008).Probiotics in aquaculture: the need,principles and mechanisms of actionand screening processes. Aquacultu-re, 274(1):1–14.
Kharisma, A. and Manan, A. (2019).Kelimpahan bakteri vibrio sp. padaair pembesaran udang vannamei (li-topenaeus vannamei) sebagai detek-si dini serangan penyakit vibriosis[the abundance of vibrio sp. bacteriaon enlargement water of litopenaeusvannamei as the early detection ofvibriosis]. Jurnal Ilmiah Perikanandan Kelautan, 4(2):128–134.
Kümmerer, K. (2004). Resistance inthe environment. Journal of Antimi-crobial Chemotherapy, 54(2):311–
320.Liu, N., Chen, X.-G., Park, H.-J., Liu,
C.-G., Liu, C.-S., Meng, X.-H., andYu, L.-J. (2006). Effect of mw andconcentration of chitosan on antiba-cterial activity of escherichia coli.Carbohydrate polymers, 64(1):60–65.
Pachanawan, A., Phumkhachorn, P.,and Rattanachaikunsopon, P. (2008).Potential of psidium guajava supple-mented fish diets in controlling ae-romonas hydrophila infection in ti-lapia (oreochromis niloticus). Jo-urnal of bioscience and bioenginee-ring, 106(5):419–424.
Pan, X., Chen, F., Wu, T., Tang, H., andZhao, Z. (2009). The acid, bile tole-rance and antimicrobial property oflactobacillus acidophilus nit. FoodControl, 20(6):598–602.
Pelczar, M. J., Chan, E. C. S., and Pe-lczar, M. F. (1981). Elements of mi-crobiology. McGraw-Hill Interna-tional Book Company.
Purwoko, T. (2009). Fisiologi mikro-ba. Bumi Aksara.
Ravi, A., Musthafa, K., Jegathammbal,G., Kathiresan, K., and Pandian, S.(2007). Screening and evaluationof probiotics as a biocontrol agentagainst pathogenic vibrios in marineaquaculture. Letters in Applied Mi-crobiology, 45(2):219–223.
Schaechter, M. (2009). Encyclopediaof microbiology. Academic Press.
Setyawan, A., Harpeni, E., Ali, M.,Mariska, D., and Aji, M. (2014). Po-tensi agen bakteri biokontrol indige-nous tambak tradisional udang win-du (penaeus monodon) di lampungtimur strain d. 2.2, terhadap bakteripatogen pada udang dan ikan. Pro-siding Pertemuan Ahli Kesehatan Ik-an, pages 11–13.

656 Esti Harpeni1 et al.
Sutherland, J., Bayliss, A., and Rober-ts, T. (1994). Predictive modellingof growth of staphylococcus aureus:the effects of temperature, ph and so-dium chloride. International jour-nal of food microbiology, 21(3):217–236.
Tsai, G.-J. and Su, W.-H. (1999). Anti-bacterial activity of shrimp chitosanagainst escherichia coli. Journal offood protection, 62(3):239–243.
Widanarni, W., Suwanto, A., Sukenda,S., and Lay, B. (2003). Potency ofvibrio isolates for biocontrol of vi-briosis in tiger shrimp (penaeus mo-nodon) larvae. BIOTROPIA-The So-utheast Asian Journal of Tropical Bi-ology, (20).
Zwietering, M., De Koos, J., Hase-nack, B., De Witt, J., and Van’t Ri-et, K. (1991). Modeling of bacte-rial growth as a function of tempe-rature. Appl. Environ. Microbiol.,57(4):1094–1101.

AQUASAINS(Jurnal Ilmu Perikanan dan Sumberdaya Perairan)(Vol 7 No. 1 Tahun 2018)
SUNSCREEN CREAM BASED ON LOCAL RAWMATERIALS OF Sonneratia alba FROM TARAKANCITY
Gloria Ika Satriani1 · Burhanuddin Ihsan1 · Yulma2 ·Rukisah1
Ringkasan Traditional products derivedfrom the local wisdom of the TarakanCity coastal communities, namely in theform of cold protective skin powder fromthe sun which is made from mangro-ve raw Sonneratia alba. This producthas potential to be developed to impro-ve the economic value of local commu-nities. They can compete with productsother sunscreen products produced byindustry (commercial) through creampreparation technology (sunscreen).Thisresearch was carried out for seven mon-ths (February-August 2018) in the En-vironmental Laboratory of FPIK UBT,Samarinda Pharmacy laboratory (AKFAR-SAM), and the UNMUL Mathematicsand Biochemistry laboratory. The re-search procedure used maserasi methodwith 70% ethanol solvent followed byconcentrating the extract using a ro-tary evaporator to obtain extracts ofactive ingredients and carried out qua-litative phytochemical testing. The ob-tained active ingredient was is then for-mulated into a sunscreen cream prepa-ration. The level of protection of mixedsunscreen cold powder and commer-1)Jurusan Akuakultur FPIK UBT 2)Jurusan Manaje-men Sumberdaya Perairan FPIK UBTE-mail: [email protected]
cial cream were tested by using UV-VIS spectrophotometer to see the va-lue of Sun Protection Factor (SPF) atabsorbance of 290-320 nm. Qualitati-ve phytochemical test showed that onS.alba contains of positive alkaloids,phenols, flavonoids, steroids, and tan-nins, and negative saponins. ANOVAstatistical analysis (95%) based on thecalculated F value <F table so that H1is rejected and H0 is accepted, ie thereis no difference in SPF values amongsunscreen products tested in this stu-dy. Based on the category of sunscreenability, it is known that the SPF valueof the three different products tested inthis study ranged from maximum to ul-tra categories with pH values rangingfrom 6.5-7.5.
Keywords Cold protective skin powder,phytochemical, sun protection factor (SPF)
Received : 15 September 2018
Accepted : 20 Oktober 2018
PENDAHULUAN
Sudah sejak lama, secara turun temu-run masyarakat nelayan di Kota Tarak-an, khususnya kaum ibu-ibu (istri ne-

658 Gloria Ika Satriani1 et al.
layan lokal) senantiasa menggunakanpupur/bedak dingin ketika beraktivitassiang hari (outdoor) terutamasaat sinarmatahari terik dan udara sangat kering.Pupur/bedak dingin tersebut umumnyadibalurkan ke seluruh wajah sehinggaketika mengering akan menghasilkantampilan wajah seperti menggunakantopeng putih tebal. Khasiat pengguna-an pupur/bedak dingin yang dipercayaoleh ibu-ibu nelayan lokal setempat ia-lah mampu membuat kulit wajah me-reka tetap sejuk, lembab, dan mengu-rangi dampak buruk sinar matahari yangdapat menyebabkan kulit berubah men-jadi semakin coklat atau menghitam (go-song). Pupur/bedak dingin tersebut di-buat sendiri secara tradisional denganmenggunakan racikan yang terdiri da-ri bahan baku lokal diantaranya bahanmangrove tanaman S. alba (baik darikulit, batang, bunga, buah, daun, danbiji) yang dipercaya dapat memberik-an khasiat untuk mengurangi dampakburuk paparan sinar matahari siang yangterik. Masyarakat lokal Indonesia umum-nya memiliki kekayaan sumberdaya ala-mi yang telah digunakan secara turun-temurun khususnya tumbuhan kosme-tik tradisional yang memiliki potensibahan aktif kosmetik (Darmakusuma,2014).
Kearifan lokal yang dimiliki masyara-kat setempat (ibu-ibu nelayan Suku As-li Tidung) perlu dilestarikan dan dikem-bangkan serta didukung dengan kajian-kajian ilmiah penelitian agar manfaat-nya dapat terus dirasakan, dan memili-ki nilai tambah (ekonomi). Produk tra-disional berupa bedak/pupur dingin iniberpotensi untuk dikembangkan gunameningkatkan nilai ekonomi masyara-kat lokal, dan dapat bersaing denganproduk-produk tabir surya lainnya yang
dihasilkan oleh produksi industri (pa-brik). Menurut keputusan Kepala BPOMRI (No: HK. 00. 05. 42. 1018) menya-takan bahwa pengertian bahan tabir sur-ya ialah bahan yang digunakan untukmelindungi kulit dari radiasi sinar UVdengan cara menyerap, memancarkan,dan menghamburkan.
Bahan baku yang relatif ramah ling-kungan sangat mudah diperoleh di Ko-ta Tarakan, yakni tanaman S. alba men-jadi nilai tambah tersendiri bagi pengem-bangan bedak/pupur dingin menjadi se-diaan krim tabir surya. Sebagian besaraktivitas farmakologis suatu tumbuhanberkaitan erat dengan kandungan me-tabolit sekundernya. Metabolit sekun-der adalah kelompok senyawa yang di-hasilkan tumbuhan dengan fungsi yanghingga saat ini belum diketahui denganjelas, namun diduga kuat berkaitan de-ngan kemampuan tumbuhan untuk mem-pertahankan diri dari serangan hama danpenyakit (Yadav and Agarwala, 2011).Pe-nelitian terbaru menemukan bahwa kan-ker tenggorokan dapat disembuhkan me-lalui aktivitas berkumur menggunakanekstrakyang berasal dari kulit mangro-ve (Revathi et al., 2013). Hingga de-ngan saat ini, masih terus dilakukan pem-buktian ilmiah tentang manfaat dari ak-tivitas metabolit sekunder yang diha-silkan oleh tanaman mangrove khusus-nya buah S. alba (apple mangrove) ser-ta pembuatan produk turunannya. Pe-nelitian ini merupakan produk tradisio-nal hasil dari kearifan lokal yang ra-mah lingkungan karena menggunakanbahan alami (buah mangrove) dan me-lalui penelitian ini diharapkan dapat me-ningkatkan pemanfaatan, daya simpan,nilai tambah estetika, dan potensi eko-nomi, serta mampu meningkatkan pe-manfaatan “Buah Pedada (S. alba)” yang

Sunscreen Cream Based Sonneratia 659
selama ini belum termanfaatkan seca-ra optimal, sehingga menciptakan pe-luang bagi masyarakat untuk menda-patkan penghasilan dari menanam danmemelihara pohon mangrove jenis pe-dadadisamping ikut serta juga(secara ti-dak langsung) memelihara kawasan hut-an mangrove yang memiliki nilai eko-logis penting bagi ekosistem pesisir danlaut tropis khususnya wilayah TarakanKalimantan Utara.
MATERI DAN METODE
Ruang lingkup dalam penelitian ini ada-lah metode maserasi menggunakan pe-larut etanol 70% dilanjutkan dengan ro-tary evaporasi. Pengujian fisik ekstrakS. alba (kadar air, kadar abu dan rende-men), pengujian fitokimia secara kua-litatif (alkaloid, fenol, flavonoid, sapo-nin, steroid, dan tanin) dan pembuatanracikan sediaan krim tabir surya berda-sarkan IMO (2004) yaitu menimbang,mencampur, dan penapisan bahan sun-screen menjadi krim sediaan setengahpadat berupa emulsi. Tahapan peneli-tian terdiri dari preparasi sampel, eks-traksi, uji fitokimia, peracikan sediaankrim dan penentuan nilai Sun Prote-ction Factor (SPF). Penelitian ini di-laksanakan selama tujuh bulan (Febuari-Agustus 2018) di laboratorium Ling-kungan FPIK UBT untuk melakukanekstraksi dan pengujian kualitatif fito-kimia, laboratorium Farmasi Samarin-da (AKFARSAM) untuk kegiatan me-racik sediaan krim, dan laboratoriumBiokimia MIPA UNMUL untuk mela-kukan pengujian nilai SPF.
Penentuan Nilai Sun Protection Factor(SPF) menurut Pissavini et al, (2003)adalah dengan menimbang sebanyak 1
Tabel 1 Kostanta Spektrum
Panjang gelombang (nm) (EE) x (I)290 0,015295 0,0817300 0,2874305 0,3278310 0,1864315 0,0839320 0,018
Total 1
gram sampel dan dipindahkan ke da-lam labu ukur dan ditambah etanol sam-pai 100 ml, dihomogenkan dan kemu-dian disaring menggunakan kertas sa-ring. Sebanyak 5 mL larutan dipindahk-an ke dalam labu ukur dan ditambahetanol sampai 50 mL. Sebanyak 5 mLlarutan dipindahkan ke dalam labu ukurdan ditambah etanol sampai 25 mL. Spek-trum absorbansi ditentukan dalam ki-saran gelombang 290-320 nm denganinterval 5 nm dan menggunakan etanolsebagai blanko. Data absorbansi dipe-roleh dimasukan dalam Persamaan 1berikut ini:
SPFSpek =FK x329
∑290
EE(ג)xI(ג)xAbs(ג)(1)
dimana, FK/Faktor Koreksi (10); Abs/AbsorbansiSampel; EE(ג)/Spektrum Efek Eritema;I(ג)/Spektrum Cahaya Matahari
Nilai EE(ג) x I(ג) adalah Konstanta yangditetapkan oleh Sayre et al. (1979) yak-ni (Tabel 1)
Efektifitas tabir surya dinyatakan olehSun Protection Factor (SPF) yang di-definisikan sebagai perbandingan Do-sis Eritema Minimum (DEM) pada ku-lit manusia terlindungi tabir surya de-ngan DEM tanpa perlindungan. NilaiSPF ditentukan melalui perbandinganenergi dari sinar yang dipaparkan un-tuk dapat menimbulkan eritema (efekkemerahan pada kulit manusia) dan da-

660 Gloria Ika Satriani1 et al.
pat juga melalui waktu yang diperluk-an sampai timbul eritema. Kategori ke-mampuan tabir surya menurut Damo-galad et al. (2013)adalah minimal (2-4), sedang (4-6), ekstra (6-8), maksi-mal (8-15), dan ultra (>15). Hasil Per-hitungan SPF pada penelitian ini meng-gunakan Spektrofotometer dengan meng-amati tiga perlakuan (Pupur/Bedak Di-ngin, Sediaan Krim, dan Tabir SuryaKomersil) yang masing-masing pengu-jian (Tabel 4) diulang sebanyak tiga ka-li menurut percobaan Rancangan AcakLengkap (RAL) sebagai berikut ini (Per-samaan 2):
Yi j = µi + Ti + εi j (2)
dimana, Yij = Pengamatan ke j dari per-lakuan ke i; µi = Nilai tengah umum;Ti = Pengaruh dari perlakuan ke i εi j= Galat percobaan dari perlakuan ke ipada pengamatan ke j; i = 1, 2, 3, .... tadalah jumlah perlakuan; j = 1, 2, 3, ....r adalah jumlah ulangan.
HASIL DAN PEMBAHASAN
Pada umumnya pelarut yang sering di-gunakan pada produk keamanan panganadalah etanol pro analysis (etil alkoholC2H5OH) karena etanol memiliki po-laritas yang tinggi sehingga dapat meng-ekstrak bahan lebih banyak dibandingk-an pelarut organik yang lain. Etanol iniadalah suatu jenis etanol dengan kadar96,0-96,5% yang umum digunakan un-tuk industri farmasi dan kosmetik ser-ta untuk minuman beralkohol. Etanolmempunyai titik didih yang rendah dancenderung aman, tidak beracun dan ber-bahaya, namun larut dalam air dan me-larutkan komponen senyawa organik se-perti karbohidrat, resin dan gum (FDCA,
2002). Pelarut yang berbeda akan mem-berikan nilai yang berbeda karena pe-larut akan berdifusi ke dalam sampel,serta pada proses tersebut pelarut ter-difusi pada komponen yang memilikitingkat kepolaran yang sama. Hal ter-sebut juga diutarakan oleh Sayre et al.(1979) bahwa faktor-faktor yang dapatmempengaruhi nilai rendemen yang di-hasilkan yaitu metode ekstraksi yangdigunakan, ukuran partikel sampel, kon-disi dan waktu penyimpanan, lama wak-tu ekstraksi, perbandingan jumlah sam-pel terhadap jumlah pelarut yang digu-nakan, dan jenis pelarut yang digunak-an.
Pemekatan ekstrak dilakukan denganmetode penguapan atau rotary evapo-rasi, yakni instrumen yang menggunak-an prinsip destilasi (pemisahan) di ma-na terjadi proses perubahan molekul da-lam fasa cair menjadi gas sehingga di-hasilkan pemekatan ekstrak (Harvey, 2000).Prinsip utama ini menggunakan penu-runan tekanan pada labu alas bulat danpemutaran labu alas bulat sehingga pe-larut dapat menguap lebih cepat di ba-wah titik didihnya. Ekstrak tersebut se-lanjutnya dikeringkan dengan oven (su-hu konstan 40˚C) hingga menghasilk-an rendemen ekstrak pasta kental.
Penapisan fitokimia ekstrak buah pe-dada (S.alba) dilakukan dengan uji al-kaloid, fenol, flavonoid, saponin, ste-roid, tanin dan kadar total fenol (pa-da Tabel 2). Menurut Lim et al. (2002)bahwa ada hubungan antara total fenoldan aktivitas antioksidan dimana jikadi dalam suatu bahan memiliki konsen-trasi senyawa fenol yang tinggi makaaktivitas antioksidan dalam bahan ter-sebut juga tinggi. Andayani et al. (2008)menyatakan bahwa senyawa fenol danaktivitas antioksidan memiliki gugus –OH

Sunscreen Cream Based Sonneratia 661
dan –OR seperti flavonoid dan asamfenolat. Menurut Svobodová et al. (2003),senyawa fenolat berperan dalam me-nurunkan sinyal redoks-sensitif untukmenghambat kerusakan DNA.
Berdasarkan penelitian Herawati et al.(2011) ekstrak S. alba memiliki kan-dungan senyawa alkaloid, flavonoid, tri-penoid, steroid, fenol hidrokuinon, sa-ponin dan tanin. Berbeda dengan pe-nelitian ini, bahwa ekstrak S.alba yanghidup di kawasan pesisir Pantai AmalKota Tarakan pada pengujian kualita-tif fitokimia menghasilkan nilai negatifyaitu tidak mengandung saponin. Per-bedaan variasi genetik, geografi, iklim,teknik budidaya dan pemanenan, wak-tu penyimpanan, dan teknik persiapanekstrak merupakan faktor penyebab per-bedaan komposisi senyawa, khususnyametabolit sekunder, sehingga mempe-ngaruhi kualitas senyawa ekstrak (Wuet al., 2009). Penelitian lain terhadaphasil analisis proksimat buah Sonnera-tia sp melaporkan bahwa buah tersebutmemiliki kadar air (bb) 84,76%, kadarabu (bk) 8,4%, kadar lemak (bk) 4,82%,kadar protein (bk) 9,21%, dan kadarkarbohidrat (bk) 77,57% (Manalu, 2011).
Berdasarkan pengujian kadar air, ka-dar abu, dan rendemen terhadap sam-pel S.alba diketahui nilainya Tabel 3.Pasta kental ini selanjutnya diuji seca-ra fisika terhadap kandungan kadar airdan kadar abu berdasarkan metode de-ngan tujuan untuk memberikan batas-an minimal atau rentang besarnya kan-dungan air dan kadar abu dalam sua-tu bahan. Semakin kecil kandungan airdalam suatu bahan, maka akan sangatberguna untuk memperpanjang daya simp-an pasta kental hasil ekstraksi selamapenyimpanan. Sedangkan, penetapan ka-dar abu pasta kental dilakukan untuk
memberikan gambaran kandungan se-nyawa anorganik yang terkandung da-lam bahan, baik yang berasal dari ta-naman secara alami maupun kontamin-an selama proses pembuatan ekstrak ber-upa pasta kental. Berdasarkan pengu-jian kadar air dan kadar abu terhadapekstrak pasta kental pada penelitian inidiketahui bahwa bahan tersebut memi-liki potensi masa simpan yang panjangserta memiliki kandungan garam anor-ganik dan mineral penting, dan tidakterjadi kontaminasi kotoran selama pe-ngerjaan proses ekstraksi. Nilai kadarair dan kadar abu akan mempengaru-hi nilai rendemen dan ekstrak yang di-hasilkan oleh sample. Menurut Ahmedet al. (2010) kandungan air dan kadarabu turut menentukan nilai kompenenlain yang turut terkandung dalam ren-demen.
Indonesia memiliki kekayaan sumberbahan kosmetik alami, khususnya tum-buhan kosmetik tradisional yang me-miliki peluang untuk dikembangkan se-bagai bahan aktif tabir surya (sunscre-en). Penentuan khasiat dan mekanismetabir surya berbahan aktif S. alba asalKota Tarakan pada Tabel. 5 telah ter-uji bahwa berpotensi menjadi produksunscreen yang efektif menangkal ra-diasi sinar UV. Pengujian SPF terdi-ri atas tahapan: (i) Penapisan kandidatekstrak bahan aktif (ii) Penentuan nilaiSPF berdasarkan kategori tabir suryamenurut Damogalad et al. (2013); dimana pada pengujian SPF pada pene-litian ini untuk bahan uji bedak dingin,sediaan krim tabir surya, dan krim sun-screen komersial (Tabel 4).
Analisis statistik ANOVA (95%) ber-dasarkan nilai F hitung < F tabel se-hingga H1 ditolak dan H0 diterima yak-ni tidak ada perbedaan nilai SPF di an-

662 Gloria Ika Satriani1 et al.
Tabel 2 Hasil Uji Fitokimia
Parameter Penapisan Ekstrak (Pelarut Etanol 70% Indikator
Alkaloid + Terbentuknya endapan jingga merah hingga coklat
Fenol + Terbentuk endapan hijau hingga hijau biru
Flavonoid + Terbentuk warna kuning jelas dahulu kemudian larutan kembali seperti semula
Saponin - Terbentuk busa stabil yang melimpah ketika dikocok
Steroid + Apabila berwarna merah ungu maka mengandung triterpenoid, dan positif steroid apabila berubah menjadi hijau biru.
Tanin + Terbentuk warna kuning gelap hingga hitam biru.
Keterangan: + (Positif atau mengandung senyawa yang diuji); dan – (negatif atau tidak mengandung senyawa yangdiuji)
Tabel 3 Hasil Pengujian Fisik
Sampel Kadar Air (%) Kadar Abu (%) Rendemen (%)Simplisia kering 29.21 ± 3.03 17.99 ± 2.65 -
Ekstrak pasta (70%) 0.21 ± 0.02 7.90 ± 0.01 9.38
Tabel 4 Nilai SPF Ekstrak S. alba
Bahan Uji Nilai SPF Kategori
Bedak Dingin12,29 Maksimal16,41 Ultra8,20 Maksimal
Sediaan Krim15,31 Ultra21,86 Ultra10,93 Maksimal
Krim komersil11,48 Maksimal16,99 Ultra16,99 Ultra
tara produk tabir surya yang diujikanpada penelitian ini. Berdasarkan kata-gori kemampuan tabir surya diketahuinilai SPF ketiga produk berbeda yangdiujikan pada penelitian ini berkisar dikategori maksimal hingga ultra. Ken-dali mutu untuk mengkarakterisasi bah-an aktif dan pengujian nilai SPF me-rupakan tahap penting bagi penelitiandan pengembangan produk kosmetik ber-bahan alami (Darmakusuma, 2014). Pe-nilaian mutu karakteristik pH pada pe-nelitian ini berkisar antara 6,5-7,5 ser-ta masih layak untuk diaplikasikan pa-da kulit manusia (Masnec and Pujube,2008).
SIMPULAN
Performa sediaan krim tabir surya ber-basis bahan baku lokal hutan mangro-
ve (S. alba) Kota Tarakan memiliki ni-lai SPF yang tidak berbeda dengan pro-duk tabir surya lainnya (pupur/bedakdingin dan tabir surya komersil) sehing-ga berpotensi untuk dikembangkan gu-na meningkatkan nilai ekonomi masya-rakat lokal. Disarankan untuk peneli-tian selanjutnya dilakukan juga anali-sis spektroskopi massa dengan meng-gunakan peralatan GCMS terhadap kan-didat ekstrak bahan aktif S. alba.
Acknowledgements Terima kasih disampa-ikan kepada Direktorat Riset dan Pe-ngabdian Masyarakat, Ditjen Penguat-an Riset dan Pengembangan, Kemenris-tekdikti.
Pustaka
Ahmed, R., Moushumi, S. J., Ahmed,H., Ali, M., Haq, W. M., Jahan, R.,and Rahmatullah, M. (2010). Serumglucose and lipid profiles in rats fo-llowing administration of sonneratiacaseolaris (l.) engl.(sonneratiaceae)leaf powder in diet. Advancesin Natural and Applied Sciences,4(2):171–174.
Andayani, R., Maimunah, M., and Yo-vita, L. (2008). Penentuan aktivitasantioksidan, kadar fenolat total danlikopen pada buah tomat (solanumlycopersicum l). Jurnal Sains danTeknologi Farmasi.

Sunscreen Cream Based Sonneratia 663
Damogalad, V., Edy, H. J., and Supri-ati, H. S. (2013). Formulasi krim ta-bir surya ekstrak kulit nanas (ananascomosus l merr) dan uji in vitro nilaisun protecting factor (spf). PHAR-MACON, 2(2).
Darmakusuma, D. (2014). Pengem-bangan Kandidat Bahan Aktif Anti-photoaging dari Beberapa Tumbuh-an Kosmetik Tradisional Indonesia.PhD thesis, Institut Pertanian Bogor.
FDCA (2002). Liquid liquid extractionand solid-liquid extraction, instituteof thermal process and environmen-tal engineering.
Harvey, D. (2000). Modern analyti-cal chemistry. Boston: McGraw-HillCompanies, Inc.
Herawati, N., Daha, J. L., and Zenta, F.(2011). Potensi antioksidan ekstrakmetanol kulit batang tumbuhan ma-ngrove s. alba. J Chemical, 12(1):9–13.
IMO (2004). Handbook of Pharma-ceutical Manufacturing FormulationSemisolid Products. CRC Press.
Lim, S., Cheung, P., Ooi, V., and Ang,P. (2002). Evaluation of antioxi-dative activity of extracts from abrown seaweed, sargassum siliquas-trum. Journal of Agricultural andFood Chemistry, 50(13):3862–3866.
Manalu, R. D. E. (2011). Kadar bebe-rapa vitamin pada buah pedada (son-neratia caseolaris) dan hasil olahan-nya. Skripsi. Departemen Teknolo-gi Hasil Perairan Fakultas Perikan-an dan Ilmu Kelautan Institut Perta-nian Bogor. Bogor.
Masnec, I. S. and Pujube, S. (2008).Photoaging. Collagen Antropology,2:177–180.
Revathi, P., Thirumalaikolundusubra-manian, P., Prabhu, N., et al. (2013).Medicinal properties of mangrove
plants–an overview. InternationalJournal of Bioassays, 2(12):1597–1600.
Sayre, R. M., Agin, P. P., LeVee,G. J., and Marlowe, E. (1979). Acomparison of in vivo and in vi-tro testing of sunscreening formu-las. Photochemistry and Photobio-logy, 29(3):559–566.
Svobodová, A., Psotová, J., Walterová,D., et al. (2003). Natural phenolicsin the prevention of uv-induced skindamage. a review. Biomed Pap MedFac Univ Palacky Olomouc CzechRepub, 147(2):137–145.
Wu, S.-B., Wen, Y., Li, X.-W., Zhao,Y., Zhao, Z., and Hu, J.-F. (2009).Chemical constituents from the fru-its of sonneratia caseolaris and son-neratia ovata (sonneratiaceae). Bi-ochemical Systematics and Ecology,37(1):1–5.
Yadav, R. and Agarwala, M. (2011).Phytochemical analysis of some me-dicinal plants. Journal of phytology.

664 Gloria Ika Satriani1et al.

AQUASAINS(Jurnal Ilmu Perikanan dan Sumberdaya Perairan)(Vol 7 No. 1 Tahun 2018)
Avicennia alba FRUIT EXTRACT AS A NATURALANTIBACTERIAL TREATMENT OF Vibrioparahaemolyticus INFECTION IN VANAME SHRIMP(Litopenaeus vannamei Boone, 1931)
Dwi Arum Mufidah1 · Wardiyanto1 · Rara Diantari2
Ringkasan Vaname shrimp (L. vannamei)has high economical value as an exportcommodity However, there are obsta-cles that cause a decline in the level ofshrimp exports in the world. One of theobstacle is shrimp disease which is tre-ated using antibiotics. This method mi-ght caused pathogenic resistance andbecome a residue when consumed byhumans. A. alba fruit extracthas bene-fits as natural antibacterial ingredientthat are safe to treat the shrimp whi-ch is infected by the V. parahaemolyti-cus. In this study, shrimp is infected wi-th Vibrio parahaemo-lyticus immersedusing A. alba with a concentration of300 mg L-1, 350 mg L-1, and 400 mgL-1 for 21 days plus the control tre-atment. The concentration of 400 mgL-1 showed better results compare toother concentration on all observed pa-rameters; faster recovery time, highersurvival rate and relative percent su-rvival (RPS), also mild damage on he-patopancreas test.
1)Program Studi Budidaya Perairan, Jurusan Perikan-an dan Kelautan, Fakultas Pertanian, Universitas Lam-pung 2)Program Studi Sumberdaya Akuatik, JurusanPerikanan dan Kelautan, Fakultas Pertanian, Universi-tas LampungE-mail: [email protected]
Keywords Litopenaeus vannamei, anti-biotics, Avicennia alba, fruit extract Vibrioparahaemolyticus
Received : 20 September 2018
Accepted : 23 Oktober 2018
PENDAHULUAN
Udang vaname merupakan udang in-troduksi yang bernilai ekonomis tinggisebagai komoditi ekspor karena dimi-nati oleh pasar dunia. Hal tersebut di-buktikan dengan tingginya jumlah eks-por ke beberapa Negara pada periode2012-2017 yang selalu mengalami ke-naikan tiap tahunnya sebesar 10,40%dan volume ekspor udang mencapai 23.620ton pada tahun 2016. Untuk memenuhitingginya permintaan tersebut, produk-si udang vaname di wilayah Indonesiaterus dilakukan. Lampung menjadi pe-nyumbang terbesar produksi udang va-name nasional tahun 2013 yaitu sebe-sar 72.051 ton (KKP, 2013).
Alasan petambak menggunakan jenisudang vaname ini adalah karena per-tumbuhannya cepat, nilai konsumsi pak-an atau Food Consumption Rate (FCR)yang rendah, mampu beradaptasi ter-hadap kisaran salinitas yang tinggi dan

666 Dwi Arum Mufidah1 et al.
dapat dipelihara pada padat tebar yangtinggi (Nur’aini et al., 2007). Akan te-tapi, terdapat beberapa kendala dalamkegiatan budidaya udang vaname, sa-lah satunya adalah penyakit. Penyakityang mudah timbul umumnya disebabk-an oleh bakteri Vibrio sp. (Ruangpanand Kitao, 1991). Fluktuasi pH, ting-kat oksigen terlarut yang rendah, tem-peratur, salinitas lebih dari 50 ppt, ka-dar amonia, dan sulfat, serta bahan-bahanorganik yang lain dapat menjadi penye-bab stress pada udang dan memicu ter-jadinya penyakit. Namun, peningkatanjumlah bakteri Vibrio tetap menjadi pe-nyebab timbulnya penyakit pada pem-besaran udang (Lightner, 1996).
Upaya pengobatan penyakit dapat dila-kukan dengan menggunakan antibiotik(Sipayung et al., 2015), namun dalamjangka waktu yang lama dapat menim-bulkan resistensi patogen. Selain itu, be-berapa negara maju mulai menerapkanpersyaratan mutu yang lebih ketat ter-hadap udang yang diimpor dari nega-ra berkembang seperti Indonesia. Vo-lume ekspor udang di Indonesia mero-sot sekitar 64% sejak diberlakukannyazero tolerance terhadap residu antibio-tik pada tahun 2001 (Putro, 2008).
Pencarian sumber antibakteri alami sa-ngat dibutuhkan sebagai pengganti per-an dari antibakteri sintetik. A. alba me-rupakan salah satu tumbuhan yang ba-nyak tersebar di Indonesia serta mem-berikan berbagai manfaat salah satunyaanti bakterial (Subashree et al., 2010).Untuk meminimalisir kerugian yang di-sebabkan oleh bakteri V. parahaemo-lyticus dapat dilakukan pengobatan udangyang terserang bakteri dengan meman-faatkan bahan antibakteri alami dari eks-trak buah mangrove jenis A. alba da-lam membunuh dan menghambat per-
tumbuhan bakteri Vibrio sp. (Manilalet al., 2009). Penggunaan bahan anti-bakteri alami yang berasal dari tum-buhan A. alba sangat dibutuhkan da-lam kasus ini. Selain murah dan lebihmudah didapat, bahan anti-bakteri ala-mi aman untuk digunakan dan tidak me-nyebabkan resistensi terhadap bakteri
MATERI DAN METODE
Uji Pendahuluan
Terdapat beberapa tahap dalam mela-kukan uji pendahuluan antara lain pre-parasi sampel buah A. alba. yang di-dapatkan dari Pulau Pasaran, Kota Ka-rang, Teluk Betung Timur, Kota Ban-dar Lampung, Lampung. Buah A. albadimaserasi menggunakan pelarut me-tanol yang selanjutnya dilakukan eva-porasi pada filtrat yang didapatkan meng-gunakan rotary evaporator dengan su-hu 37 oC (Handayani and Nurjanah, 2013).Ekstrak cair yang didapatkan kemudi-an diuji kualitatif fitokimia untuk meng-etahui kandungan senyawa aktif yangterdapat dalam buah A. alba. Dilakuk-an uji aktivitas antibakteri pada ekstrakyang telah didapat terhadap bakteri V.parahaemolyticus dengan konsentrasi50, 100, 200, 400 mg/l serta kontrolpositif yang mengandung 50 mg/l klo-ramfenikol dan kontrol negatif yang meng-andung pelarut metanol.
Uji Toksisitas Brine Shrimp Lethal To-xicity (BSLT)
Uji BSLT ini dilakukan dengan meng-gunakan larva Artemia salina sebanyak10 ekor yang dimasukkan ke dalam ta-bung reaksi berisi 3 ml air laut dan te-lah diberi larutan ekstrak buah mangro-ve dengan konsentrasi 0 (kontrol), 300

Extract Avicennia Fruit for Vibrio Infection in Vannamei 667
mg/l, 350 mg/l, dan 400 mg/l sebanyak2 kali ulangan.
Persiapan Media Pemeliharaan dan He-wan Uji
Wadah yang digunakan pada peneliti-an ini sebanyak 12 akuarium yang ber-warna gelap. Sebelum digunakan, aku-arium disterilisasi secara kimiawi de-ngan cara dicuci dan didesinfeksi meng-guanakan klorin 30 mg/l, kemudian di-netralkan dengan natrium thiosulfat 15mg/l (Widanarni et al., 2014).
Pemeliharaan Hewan Uji
Udang vaname diberi pakan komersildengan frekuensi pemberian pakan 4kali sehari yaitu pukul 07.00, 11.00, 15.00.dan 19.00 WIB secara ad libitum. Pe-ngelolaan kualitas air dilakukan denganpenyiponan dan pergantian air setiappagi hari sebanyak 10% serta dilakuk-an pengukuran kualitas air seperti su-hu, DO, pH, dan salinitas pada awaldan akhir penelitian.
Uji Kohabitasi
Bakteri V. parahaemolyticus dikultur pa-da media cair NB dan diinkubasi pa-da orbital shaker hingga mencapai ke-padatan 107 CFU/ml. Udang diinjeksibakteri dengan kepadatan 107 CFU/mldan dilakukan pengamatan setelah udangmenunjukkan gejala abnormal. Infek-si vibriosis diisolasi dengan mengam-bil bagian abnormal pada udang meng-gunakan jarum ose dan ditanam padamedia TCBS (Hardiyani et al., 2016).Bakteri yang telah tumbuh pada me-dia TCBS diambil koloni tunggal un-tuk dilakukan re-injeksi V. parahaemo-lyticus dengan kepadatan 107CFU/mlpada udang baru dengan masa peme-liharaan lebih cepat dari injeksi sebe-lumnya. Pemeliharaan dilakukan hing-ga udang memperlihatkan gejala klinis
abnormal yang berarti bakteri V. para-haemolyticus telah aktif. Kemudian di-lakukan isolasi kembali pada bagian ab-normal udang yang ditanam pada me-dia TCBS. Bakteri yang tumbuh padamedia TCBS ini digunakan sebagai ujitantang.
Uji Tantang
Uji tantang dilakukan menggunakan bak-teri V. parahaemolyticus dengan kepa-datan 107 CFU/ml sebanyak 0,1 ml/ekor.Injeksi dilakukan pada bagian dekat in-sang dengan menggunakan alat suntik.Gejala klinis udang pasca uji tantangdiamati selama 3 hari dan dilakukanperendaman menggunakan ekstrak bu-ah A. alba dengan konsentrasi 300 mg/l,350 mg/l dan 400 mg/l dan dilakukanpengamatan gejala klinis kembali sela-ma 21 hari.
Analisis Data
Data kelangsungan hidup dan kerusak-an jaringan pada hewan uji dianalisissecara statistik kemudian diuji norma-litas serta homogenitas. Apabila datatelah homogen, diolah dengan analisissidik ragam (ANNOVA) dengan ting-kat kepercayaan 95% untuk mengeta-hui pengaruh pemberian ekstrak buahA. alba dengan konsentrasi berbeda se-bagai pengobatan penyakit yang dise-babkan oleh bakteri V. parahaemolyti-cus pada udang vaname. Apabila ber-beda nyata antar perlakuan, maka diujilanjut menggunakan uji Duncan padatingkat kepercayaan 95%. Sedangkandata RPS (Relative Percent Survival)dianalisis menggunakan uji T dan datakualitas air serta gejala klinis dianali-sis secara deskriptif.

668 Dwi Arum Mufidah1 et al.
HASIL DAN PEMBAHASAN
Hasil pengamatan gejala klinis menun-jukkan perubahan morfologi udang va-name terjadi pada hari ke-2 pasca in-jeksi V. parahaemolyticus yaitu warnatubuh, ekor, dan pleopod memerah ser-ta hepatopankreas berwarna cokelat. Pa-da hari ke-3 udang mengalami nekro-sis pada karapas dan ekor geripis. Pe-ngamatan gejala klinis pasca perendam-an ekstrak buah A. alba menunjukkanbahwa semua perlakuan belum menga-lami adanya pemulihan hingga hari ke-5. Perlakuan D (400 mg/l) yang me-rupakan dosis tertinggi membutuhkanwaktu pemulihan lebih cepat dibandingk-an perlakuan A, B, dan C. Udang va-name pada perlakuan A (0 mg/l) tidakmengalami pemulihan tubuh hingga ha-ri ke-21, hal tersebut dikarenakan tidakterdapat kandungan bahan aktif padaperlakuan A yang membantu tubuh udangdalam menghambat infeksi bakteri. Haltersebut sesuai dengan pernyataan Ro-sidah and Afizia (2012) bahwa sema-kin tinggi konsentrasi ekstrak maka ke-mampuan anti-bakterinya juga semakinbesar. Pemulihan kondisi morfologi udangpasca perendaman ekstrak buah A. al-ba dikarenakan adanya kandungan se-nyawa aktif pada buah A. alba sepertiflavonoid dan tanin yang berfungsi se-bagai antibakteri. Pemulihan morfolo-gi udang vaname ditandai dengan me-mudarnya warna merah pada kaki danekor serta hilangnya nekrosis pada ka-rapas udang. Terjadinya pemulihan mor-fologi udang membuktikan bahwa se-nyawa aktif pada buah A. alba berper-an sebagai antibakteri pada udang va-name pasca injeksi V. parahaemolyti-cus.
Gambar 1 Kelangsungan hidup udang vaname
Hasil analisis ragam (ANNOVA) me-nunjukkan perendaman udang vanameyang terinfeksi V. parahaemolyticus meng-gunakan ekstrak buah A. alba berpe-ngaruh terhadap kelangsungan hidup udangvaname. Hasil analisis diuji lanjut meng-gunakan uji Duncan yang menunjukk-an bahwa perlakuan A berbeda nyatadengan perlakuan B, C, dan D (P < 0,05)(Gambar 1).
Kelangsungan hidup udang vaname ber-kaitan dengan kandungan bahan aktifdalam buah A. alba yang terdiri dariflavonoid dan tanin. Flavonoid berfung-si sebagai antibakteri dengan memben-tuk senyawa kompleks terhadap prote-in extraseluler yang mengganggu keu-tuhan membran sel bakteri. Mekanis-me kerja flavonoid yaitu dengan caramendenaturasi protein sel bakteri danmerusak membran sel sehingga tidakdapat diperbaiki lagi. Tanin memilikiaktivitas antibakteri yang dapat meng-erutkan membran sel sehingga meng-ganggu permeabilitas sel itu sendiri. Aki-bat terganggunya permeabilitas, sel ti-dak dapat melakukan aktivitas hidup se-hingga partumbuhannya terhambat danmati.Nilai kelangsungan hidup udangvaname juga berkaitan dengan patoge-

Extract Avicennia Fruit for Vibrio Infection in Vannamei 669
Gambar 2 Relative percent survival
nisitas V. parahaemolitycus yang ber-hubungan dengan kondisi udang. Kon-disi air selama pemeliharaan udang va-name berada dalam kisaran optimumyang mempengaruhi udang tetap dalamkondisi yang baik, dimana V. paraha-emolyticus bersifat oportunistik yaitudapat menjadi lebih patogen apabila kon-disi inang kurang optimal (Nasi et al.,2012).
Hasil analisis uji t menunjukkan bah-wa penggunaan ekstrak buah A. albasebagai pengobatan udang vaname yangterinfeksi V. parahaemolyticus tidak ber-pengaruh terhadap relative percent su-rvival (RPS) udang vaname (P > 0,05)Gambar 2.
Pemberian ekstrak buah A. alba seba-gai pengobatan udang vaname yang ter-infeksi V. parahaemolyticus memberik-an perlindungan yang cukup baik mes-kipun tidak berpengaruh nyata terha-dap relative percent survival udang. Halini sesuai dengan pernyataan Alifuddin(2007) secara umum efektivitas vaksindianggap baik apabila nilai RPS ≥ 50%.Ketahanan udang pada masing-masingperlakuan dapat dilihat dari pengamat-an langsung berupa perubahan tingkah
laku udang selama masa pemeliharanpasca perendaman.
Kualitas air merupakan faktor pentingyang sangat mempengaruhi kehidupanorganisme akuatik seperti reproduksi,pertumbuhan, dan kelangsungan hidup.Parameter kualitas air yang diamati da-lam penelitian ini seperti suhu, DO, sa-linitas, dan pH berada dalam kisaranoptimal.
Hepatopankreas merupakan organ tar-get infeksi bakteri dan merupakan org-an terpenting pada udang karena me-miliki beberapa fungsi seperti detoksi-fikasi, memproduksi enzim-enzim pen-cernaan, menyimpan hasil-hasil pencer-naan termasuk mineral dan bahan or-ganik, eksresi, metabolisme lemak dankarbohidrat sebagai energi, dan juga me-nyebarkan nutrisi ke berbagai bagiantubuh untuk fungsi fisiologis, khusus-nya saat pembentukan kembali kulit udangsaat periode moulting (Musallamah andAbdulgani, 2010). Apabila hepatopank-reas mengalami kerusakan, maka akanberpengaruh terhadap sistem metabo-lisme dalam tubuh udang.
Uji histopatologi dilakukan pada hepa-topankreas udang sehat, udang terin-feksi V. parahaemolyticus, dan udangpasca perendaman ekstrak buah A. al-ba untuk melihat perbedaan organ he-patopankreas udang vaname. Dari ha-sil uji didapatkan semua sampel meng-alami kerusakan jaringan berupa nek-rosis (N), vakuolasi (V), dan degene-rasi (D) dengan nilai kerusakan rata-rata < 25% yang menyebabkan hepato-pankreas mengalami kerusakan ringanakibat infeksi bakteri V. parahaemolyti-cyus.
Tingkat kerusakan yang terjadi pada ti-ap perlakuan mendapatkan persentase

670 Dwi Arum Mufidah1 et al.
Tabel 1 Rata-rata tingkat kerusakan hepatopankreasudang vaname
Tingkat kerusakanKerusakan jaringan
Nekrosis Degenerasi VakuolasiA (0 mg/l) ++ + ++
B (300 mg/l) + + +C (350 mg/l) + + +D (400 mg/l) + + +
Keterrangan: + = Kerusakan ringan ++ = Kerusakansedang
yang berbeda. Perbedaan tingkat keru-sakan yang terjadi disajikan pada (Ta-bel 1).
Tingkat kerusakan jaringan pada perla-kuan A (0 mg/l) mengalami kerusakanlebih parah dibandingkan dengan per-lakuan B (300 mg/l), C (350 mg/l), danD (400 mg/l). Hal tersebut dikarenak-an perlakuan A tidak dilakukan peren-daman dengan ekstrak buah A. alba Haltersebut sesuai dengan pernyataan Ri-dlo and Pramesti (2012) bahwa immu-nostimulan bergantung pada tinggi ren-dahnya konsentrasi ekstrak yang dibe-rikan. Konsentrasi dibawah nilai mini-mal untuk terjadinya respon imun ti-dak akan memberikan pengaruh padapeningkatan imun, sedangkan konsen-trasi yang terlalu tinggi juga tidak da-pat memberikan efek sebagai inhibitor.
Kerusakan hepatopankreas seperti nek-rosis, degenerasi, dan vakuolasi pernahdilaporkan oleh Pratama et al. (2014)bahwa adanya kerusakan pada hepato-pankreas berupa nekrosis, lesi, bolitasparah pada udang yang terinfeksi vi-briosis. Vakuolasi merupakan pemben-tukan ruang di dalam sel yang berisi le-mak akibat dari degenerasi sel yang di-tandai dengan munculnya vakuola pa-da tubulus hepatopankreas (Zhahrah et al.,2016). Kerusakan jaringan yang lain ter-lihat adanya bolitas dan nekrosis ser-ta sel epitel yang bengkak dan menun-jukkan sitoplasma menjadi padat.
Ambipillai et al. (2003)menyatakan bah-wa hepatopankreas udang yang terin-feksi vibriosis menunjukkan adanya nek-rosis parah, kehilangan struktur jaring-an, atropi sel epitel tubulus serta pem-bulatan vakuola dan pegelupasan sel-sel ke dalam lumen. Nekrosis merupak-an kematian sel atau jaringan yang meng-akibatkan jaringan tidak utuh lagi atautidak normal. Nekrosis disebabkan olehagen-agen biologis seperti bakteri se-hingga terjadi perubahan sel (Austin andZhang, 2006). Karunasagar et al. (1994)melaporkan terjadinya penyakit bolitasdapat menyebabkan degenerasi hepa-topankreas serta diperkuat oleh Aus-tin and Zhang (2006) yang menyatak-an bahwa bolitas menyebabkan terha-langinya kelenjar pencernaan akibat pem-bengkakan jaringan yang berbentuk se-perti bola.
SIMPULAN
Ekstrak buah A. alba berpengaruh ter-hadap tingkat kelangsungan hidup dankondisi hepatopankreas udang vanameyang terinfeksi V. parahaemolyticus de-ngan nilai tertinggi pada konsentrasi 400mg/l. Penggunaan ekstrak buah A. albatidak berpengaruh terhadap nilai rela-tive percent survival udang vaname.
Pustaka
Alifuddin, M. (2007). Immunostimul-an on aquatic organisms. JurnalAkuakultur Indonesia, 1(2):87–92.
Ambipillai, L., Sobhana, K., George,K., and Sanil, N. (2003). Histopa-thological survey of cultured shrim-ps in cochin, kerala. Journal of theMarine Biological Association of In-dia, 45(2):178–185.

Extract Avicennia Fruit for Vibrio Infection in Vannamei 671
Austin, B. and Zhang, X.-H. (2006).Vibrio harveyi: a significant patho-gen of marine vertebrates and inver-tebrates. Letters in applied microbi-ology, 43(2):119–124.
Handayani, S. and Nurjanah, S. R.(2013). Kandungan flavonoid kulitbatang dan daun pohon api-api (avi-cennia marina (forks.) vierh.) seba-gai senyawa aktif antioksidan. Bo-gor. Fakultas Perikanan dan IlmuKelautan Institut Pertanian Bogor.Skripsi, pages 35–37.
Hardiyani, S., Harpeni, E., Setyawan,A., and Supono, S. (2016). Pathoge-nicity and in vivo study of local iso-late bacillus sp. d2. 2 at the vanna-mei culture (litopenaeus vannamei).AQUASAINS, 5(1):441–446.
Karunasagar, I., Pai, R., Malathi, G.,and Karunasagar, I. (1994). Massmortality of penaeus monodon la-rvae due to antibiotic-resistant vi-brio harveyi infection. Aquaculture,128(3-4):203–209.
KKP (2013). Kelautan dan perikanandalam angka 2013.
Lightner, D. V. (1996). A handbookof shrimp pathology and diagnosticprocedures for diseases of culturedpenaeid shrimp.
Manilal, A., Sujith, S., Kiran, G. S.,Selvin, J., and Shakir, C. (2009).Biopotentials of mangroves colle-cted from the southwest coast of in-dia. Global J Biotechnol Biochem,4(1):59–65.
Musallamah, A. and Abdulgani, N.(2010). Pengaruh paparan tim-bal (pb) terhadap perubahan histo-patologi hepatopankreas udang ga-lah (macrobrachium rosenbegii demann).[skripsi]. ITS. Surabaya.
Nasi, L., Prayitno, S. B., and Sarji-to (2012). Kajian bakteri penye-
bab vibriosis pada udang secara bi-omolekuler. Master’s thesis, Magis-ter Manajemen Sumberdaya Pantai.Universitas Diponegoro, Semarang.
Nur’aini, Y., Bambang, H., Subyakto,S., and Gemi, T. (2007). Active su-rveilance of infectious myonecrosisvirus (imnv) in pond-cultured whiteshrimp (lithopenaeus vannamei) ineast java and bali. Jurnal PerikananUGM. IX (1), pages 25–31.
Pratama, P. N., Prayitno, S. B., et al.(2014). Pemanfaatan ekstrak daunbinahong (anredera cordifolia) un-tuk penanggulangan penyakit bakte-ri (vibrio harveyi) pada udang win-du. Journal of Aquaculture Manage-ment and Technology, 3(4):281–288.
Putro, S. (2008). Peran mutu dalammenunjang ekspor udang nasional.Squalen Bulletin of Marine and Fi-sheries Postharvest and Biotechno-logy, 3(1):1–6.
Ridlo, A. and Pramesti, R. (2012).Aplikasi ekstrak rumput laut sebagaiagen imunostimulan sistem perta-hanan non spesifik pada udang (lito-pennaeus vannamei). ILMU KELA-UTAN: Indonesian Journal of Mari-ne Sciences, 14(3):133–137.
Rosidah and Afizia, W. M. (2012). Po-tensi ekstrak daun jambu biji sebagaiantibakterial untuk menanggulangiserangan bakteri aeromonas hydro-phila pada ikan gurame (osphrone-mus gouramy lacepede). Jurnal Aku-atika, 3(1):19–27.
Sipayung, B. S., Ma’ruf, W. F., and De-wi, E. N. (2015). Pengaruh senya-wa bioaktif buah mangrove avicen-nia marina terhadap tingkat oksida-si fillet ikan nila merah o. niloticusselama penyimpanan dingin. JurnalPengolahan dan Bioteknologi HasilPerikanan, 4(2):115–123.

672 Dwi Arum Mufidah1 et al.
Subashree, M., Mala, P., Umanrnaha-sewari, M., Jayakumari, M., Mahes-wari, K., Sevanthi, T., and Mani-kandan, T. (2010). Screening of theantibacterial properties of avicenniamarina from pichavaram mangrove.Asian Journal of Science and Tech-nology, 1:16–19.
Widanarni, Noermala, J. I., and Su-kenda (2014). Pemberian prebiotik,probiotik, dan sinbiotik untuk pe-ngendalian ko-infeksi vibrio harve-yi dan infectious myonecrosis viruspada udang vaname litopenaeus van-namei. Jurnal Akuakultur Indonesia,13(1):11–20.
Zhahrah, Z., Nur, I., and Sabilu, K.(2016). Kerusakan jaringan hepa-topankreas pada udang vaname (li-topenaeus vannamei) akibat paparanlogam berat nikel (ni) secara buatan.Jurnal Media Akuatika, 1(2).
