bab i pendahuluan a. latar...
TRANSCRIPT

1
BAB I
PENDAHULUAN
A. Latar Belakang
Beberapa jenis rusa di Indonesia yaitu rusa Timor (Rusa timorensis
Blainville, 1822), rusa Sambar (Cervus unicolor), dan rusa totol (Axis axis). Rusa
di Indonesia mempunyai peluang untuk dibudidayakan. Rusa juga mempunyai
nilai estetika yang dapat dijadikan satwa peliharaan untuk kesenangan dan sebagai
satwa pajangan dalam taman, terutama rusa totol dan rusa Timor (Garsetiasih dan
Takandjadji, 2007).
Rusa Timor merupakan salah satu jenis rusa asli Indonesia. Populasi rusa
Timor di alam (in-situ) mulai berkurang karena perburuan dan perusakan habitat.
Secara tradisional, masyarakat memanfaatkan satwa rusa melalui perburuan
langsung dari habitat alam untuk dijadikan satwa peliharaan maupun diambil
daging dan hasil ikutannya (Bismark dkk., 2011).
Status rusa Timor dalam the International Union for Conservation of
Nature Redlist yaitu Vulnerable (rentan). Jumlah populasi rusa Timor di habitat
asli diperkirakan berjumlah kurang dari 10.000 individu dewasa. Dalam waktu
tiga generasi (diperkirakan minimal 15 tahun) diduga terjadi penurunan minimal
10% dalam waktu. Penurunan populasi ini sebagai akibat dari hilangnya habitat,
degradasi habitat, dan perburuan (Hedges dkk., 2008).
Peraturan Pemerintah (PP) Nomor 7 Tahun 1999 tentang Pengawetan
Jenis Tumbuhan dan Satwa Liar menyatakan bahwa seluruh jenis rusa diIndonesia
dilindungi, namun perburuan masih saja terjadi. Selain itu, kerusakan habitat, baik

2
disebabkan eksploitasi hutan oleh manusia maupun kerusakan karena bencana
alam, turut pula menyebabkan penurunan terhadap eksistensi jenis dan populasi
rusa. Upaya konservasi jenis dan populasi rusa perlu dilakukan baik secara in-situ
maupun ex-situ, sehingga pelestarian dan pemanfaatannya dapat tercapai (Bismark
dkk., 2011).
Upaya konservasi melalui penangkaran sudah banyak dilakukan.
Peningkatan manfaat dan penangkaran perlu dikembangkan di masyarakat
terutama generasi kedua (F2) yang termuat dalam PP Nomor 8 Tahun 1999
tentang Pemanfaatan Jenis Tumbuhan dan Satwa Liar. Manfaat rusa sebagai
sumber protein hewani serta hasil ikutan lainnya diperoleh melalui turunan kedua
(F2) dan seterusnya. Hasil penangkaran rusa tersebut memiliki prospek untuk
dikembangkan dalam skala budidaya komersial, sehingga asumsi hutan sebagai
sumber pangan dapat terpenuhi (Bismark dkk., 2011).
Sebuah pemahaman menyeluruh mengenai struktur genetik populasi yang
penting bagi manajemen spesies. Populasi yang terisolasi sering dikaitkan dengan
inbreeding sehingga memiliki keragaman genetik yang terbatas dan perfoma
reproduksi yang kurang baik. Populasi bottlenecks dapat memiliki efek keragaman
genetik dan mengakibatkan berkurangnya jumlah rata-rata alel dan
heterozigositasnya (Zidek dkk., 2008).
Daerah mitokondria dikenal mengalami perubahan cepat dalam evolusi.
Analisis restriksi dari DNA mitokondria (mtDNA) mamalia yang berhubungan
dekat menunjukkan bahwa genom ini mempunyai laju evolusi yang lebih tinggi
dari pada DNA inti (Wirdateti dkk., 2004). Di samping itu, mtDNA telah lama

3
digunakan sebagai penanda yang ideal karena kecocokan untuk rekonstruksi
silsilah gen dan sejarah populasi (Gissi dkk., 2000; Wirdateti dkk., 2004; Fonseca
dkk., 2008).
Identifikasi rusa Timor di penangkaran maupun di alam penting untuk
dilakukan karena introduce spesies ke suatu daerah yang tidak terkendali baik
oleh alam maupun manusia akan mempengaruhi karakter spesifik dari hewan
tersebut, terlebih apabila proses hibridisasi antar jenis dimungkinkan terjadi secara
alami. Penelitian tentang karakteristik genetik rusa Indonesia berdasarkan gen
12SrRNA mtDNA sudah pernah dilakukan oleh Wirdateti dkk. (2004).
Berdasarkan hal tersebut, penelitian menggunakan gen lain perlu dilakukan.
Dalam penelitian ini, penulis memilih gen penyandi Cytochrome Oxidase Sub-
unit 2 (COX-2).
B. Tujuan Penelitian
Tujuan penelitian ini adalah untuk mengkaji keragaman genetik rusa
Timor terhadap spesies-spesies rusa di Indonesia dan di dunia serta famili lain
yang endemik di Indonesia yaitu Bos javanicus berdasarkan gen penyandi
Cytochrome Oxidase Sub-unit 2 (COX-2), sehingga dapat diketahui kemungkinan
gen penyandi ini sebagai penanda genetik.
C. Manfaat Penelitian
Keberadaan informasi genetik rusa Timor dapat mempermudah proses
identifikasi dan klasifikasi rusa tersebut dalam upaya untuk pelestarian plasma
nutfah, membantu tujuan konservasi serta biodiversitas rusa asli asal Indonesia.

4
BAB II
TINJAUAN PUSTAKA
A. Rusa Timor (Rusa timorensis)
1. Klasifikasi
Rusa Timor diklasifikasikan ke dalam Kingdom Animalia, Filum
Chordata, Kelas Mammalia, Ordo Cetartiodactyla, Famili Cervidae, Genus Rusa,
dan Spesies Rusa timorensis (de Blainville, 1822). Rusa Timor juga mempunyai
nama spesies lain selain Rusa timorensis, yaitu: Cervus celebensis (Rorig, 1896);
Cervus hippelaphus (Cuvier, 1825 [preoccupied]); Cervus lepidus (Sundevall,
1846); Cervus moluccensis (Quoy dan Gaimard, 1830); Cervus peronii (Cuvier,
1825); Cervus russa (Muller dan Schlegel, 1845); Cervus tavistocki (Lydekker,
1900); dan Cervus tunjuc (Horsfield, 1830 [nomen nudum]) (Hedges dkk., 2008).
2. Habitat
Habitat alami rusa adalah vegetasi dan hutan savana dimanfaatkan sebagai
sumber pakan. Vegetasi hutan yang tidak terlalu rapat untuk tempat bernaung
(istirahat), kawin, dan menghindarkan diri dari predator. Habitat yang paling
disukai oleh rusa Timor adalah hutan sampai ketinggian 2.600 meter di atas
permukaan laut dengan padang rumput (Santoso, 2011). Habitat alami rusa Timor
menurut Bismark dkk. (2011) adalah hutan tropis dan dataran rendah yang
bervegetasi savana, lontar, cemara, dan mangrove.

5
3. Morfologi
Morfologi rusa Timor menurut Bismark dkk. (2011) mempunyai ukuran
tubuh yang kecil, tungkai pendek, ekor panjang, dahi cekung, gigi seri relatif
besar, dan rambut berwarna coklat kekuning-kuningan. Secara umum rusa terlihat
pendek dan gemuk karena memiliki kaki yang relatif pendek.
Tinggi dan Berat Badan. Rusa Timor memiliki tinggi badan 91-102 cm.
Berat badannya berkisar antara 103-155 kg. Rusa jantan relatif lebih besar
dibandingkan dengan betinanya (Santoso, 2011).
Warna Rambut. Warna rambut yang dimiliki rusa Timor bervariasi
antara coklat kemerahan sampai abu-abu kecoklatan (Semiadi dan Nugraha,
2004), sedangkan menurut Santoso (2011) warna rambutnya coklat tetapi pada
bagian bawah perut dan ekor berwarna putih. Warna rambutnya pada musim
kemarau adalah merah kecoklatan, agak gelap pada bagian belakang dan lebih
terang pada bagian dada, sedangkan pada musim hujan bagian atasnya berwarna
keabu-abuan (Bismark dkk., 2011).
Ranggah. Rusa jantan memiliki ranggah yang akan tumbuh pertama kali
pada umur delapan bulan. Bila dewasa, ranggah rusa telah menjadi sempurna
yang ditandai dengan terdapatnya tiga ujung runcing (Santoso, 2011). Cabang
yang pertama mengarah ke depan, cabang belakang kedua terletak pada satu garis
dengan cabang belakang pertama, cabang belakang kedua lebih panjang cabang
depan kedua, cabang belakang kedua sisi kiri dan kanan terlihat sejajar (Bismark
dkk., 2011). Rusa jantan menjadi lebih agresif ketika ranggah keras menjadi

6
sempurna (Pattiselanno dkk., 2008). Gambar 1 menunjukkan ranggah Rusa
timorensis yang sempurna.
Gambar 1. Ranggah Rusa timorensis yang sempurna (Hedges dkk., 2008)
4. Sebaran
Rusa Timor mempunyai daya adaptasi yang tinggi, sehingga dengan
mudah menyesuaikan diri dan hidup di daerah basah, kering, berpasir maupun
pegunungan. Rusa Timor lebih mampu beradaptasi karena ketergantungannya
terhadap air lebih kecil (Bismark dkk., 2011).
Rusa Timor tersebar di Pulau Jawa, Sulawesi, Maluku, dan Nusa
Tenggara, sedangkan rusa Sambar (R. unicolor) tersebar di Pulau Sumatera dan
Kalimantan (Santoso, 2011). Penyebaran sub-spesies rusa di Indonesia disajikan
pada Gambar 2.

7
Gambar 2. Penyebaran sub-spesies rusa di Indonesia (Bismark dkk., 2011)
Keterangan:
1. R. u. equinus (Curier, 1823) 6. R. t. djonga (Bemmel, 1949)
2. R. u. brookei (Hose, 1893) 7. R. t. moluccensis (Muller, 1836)
3. R. u. russa (Muller dan Schi, 1844) 8. R. t. renschi (Sody, 1933)
4. R. t. floresiensis (Heude, 1896) 9. R. t. laronesiotes (Bemmel, 1949)
5. R. t. timorensis (Blainville, 1822) 10. R.t.macassaricus (Heude, 1896)
5. Tingkah Laku
Beberapa perilaku kegiatan harian rusa yang menonjol yaitu merumput
(31,17%), memamah (14,63%), dan berbaring/istirahat (13,54%). Waktu
merumput terbanyak dilakukan di pagi hari dan menjelang malam hari, sedangkan
memamah, istirahat dan kegiatan lainnya banyak dilakukan pada siang hari. Pakan
yang disukai rusa umumnya dari jenis rumput-rumputan, legum/kacang-kacangan
sedangkan pakan yang tidak disukai adalah daun tebal, pakan berasa pahit dan
berbau menusuk (Wirdateti dkk., 2005). Perilaku sosialnya hampir menyerupai
rusa Sambar, namun rusa ini cenderung tenang pembawaannya (Semiadi dan
Nugraha, 2004).

8
B. Deoxyribonucleic Acid (DNA)
Deoxyribonucleic acid (DNA) merupakan polimer yang terdiri atas
molekul-molekul deoksiribonukleotida yang terikat satu dengan lain, sehingga
membentuk rantai polinukleotida yang panjang. Basa purin yang terdapat pada
DNA ialah adenine dan guanine. Cytosine dan thymine adalah basa pirimidin yang
terdapat pada asam nukleat ini (Poedjiadi dan Supriyanti, 2005).
Setiap rantai DNA mempunyai polaritas yaitu dua ujung polimernya
berbeda. Pada salah satu ujung DNA (ujung 5’), atom karbon yang terdekat
dengan ujung polimer adalah karbon 5’ unit deoksiribosa yang terakhir. Pada
ujung lain DNA (3’), atom karbon yang terdekat dengan ujung polimer adalah
karbon 3’ (Colby, 1996). Gambaran bentuk skematik DNA heliks ganda
ditunjukkan pada Gambar 3.
Gambar 3. Bentuk skematik DNA heliks ganda (Fatchiyah dkk., 2011)
Kedua untai DNA bersifat antiparalel, yaitu satu untai DNA berjalan dari
arah 5’ ke 3’, sementara untai yang lain dari arah 3’ ke 5’. Ini analog dengan dua

9
jalan sejajar yang masing-masing berjalan menuju satu tempat, tetapi membawa
arus kendaraan dalam arah yang berlawanan (Murray dkk., 2003).
Molekul DNA bentuk heliks ganda sangat stabil karena adanya tiga ikatan
kimia. Ikatan hidrogen terbentuk pasangan-pasangan basa antara masing-masing
benang. Ikatan van der Waals terbentuk antara pasangan basa yang satu dengan
pasangan basa di atas dan di bawahnya sepanjang sumbu utama heliks. Ikatan
fosfodiester pada bagian luar dari kerangka DNA, menyebabkan molekul tersebut
bersifat asam (Subaron, 1983).
Mitokondria dan kloroplas pada sel eukariotik juga mengandung DNA.
Molekul DNA mitokondria (mtDNA) jauh lebih kecil daripada kromosom inti.
Pada sel hewan, mtDNA mengandung kurang dari 20.000 bp (16,569 bp mtDNA
manusia) dan merupakan dupleks melingkar. Setiap mitokondria biasanya
memiliki 2-10 salinan molekul mtDNA ini, dan jumlahnya dapat meningkat
sampai ratusan dalam sel tertentu ketika embrio mengalami diferensiasi sel
(Nelson dan Cox, 2004).
Kode DNA mitokondria untuk tRNA mitokondria dan rRNA dan untuk
beberapa protein mitokondria. Lebih dari 95% protein mitokondria dikode oleh
DNA nuklear. Mitokondria dan kloroplas membagi ketika sel membelah. Molekul
DNA direplikasi sebelum dan selama pembelahan, dan molekul DNA baru masuk
ke dalam organel sel (Nelson dan Cox, 2004).
1. Sintesis DNA
Sintesis DNA pada sel binatang, termasuk sel manusia terjadi pada fase S
atau fase sintesis. Selama fase S, DNA polimerase sel mamalia berada dalam

10
jumlah yang lebih besar dibandingkan dengan fase nonsintesis.
Deoksiribonukleosida trifosfat yaitu enzim-enzim yang bertanggung jawab atas
pembentukan substrat bagi sintesis DNA, sehingga mengalami peningkatan
aktivitas, dan aktivitas ini akan menurun sesudah fase sintesis sampai sinyal bagi
sintesis DNA baru muncul kembali (Murray dkk., 2003).
2. Inti Sel
Di dalam inti sel terdapat kromatin atau kromosom yang terdiri atas serat-
serat DNA yang bergabung dengan histon. Inti sel juga mengandung beberapa
enzim antara lain DNA polymerase, RNA polymerase dan enzim yang digunakan
dalam proses glikolisis maupun dalam siklus asam sitrat. Di dalam anak inti
terdapat enzim-enzim RNA polymerase, RNAase, NADP pirofosforilase, ATPase,
tetapi tidak terdapat DNA polymerase (Poedjiadi, 1994).
3. Mitokondria
Mitokondria adalah organela yang terletak di dalam sitoplasma sel
eukariota. Struktur organela ini berupa kantung yang diselaputi oleh dua membran
yaitu membran luar dan membran dalam. Mitokondria memiliki dua kompartemen
yaitu matriks mitokondria yang diselimuti langsung oleh membran dalam dan
ruang antar membran (Susmiarsih, 2010). Struktur mitokondria ditunjukkan pada
Gambar 4.
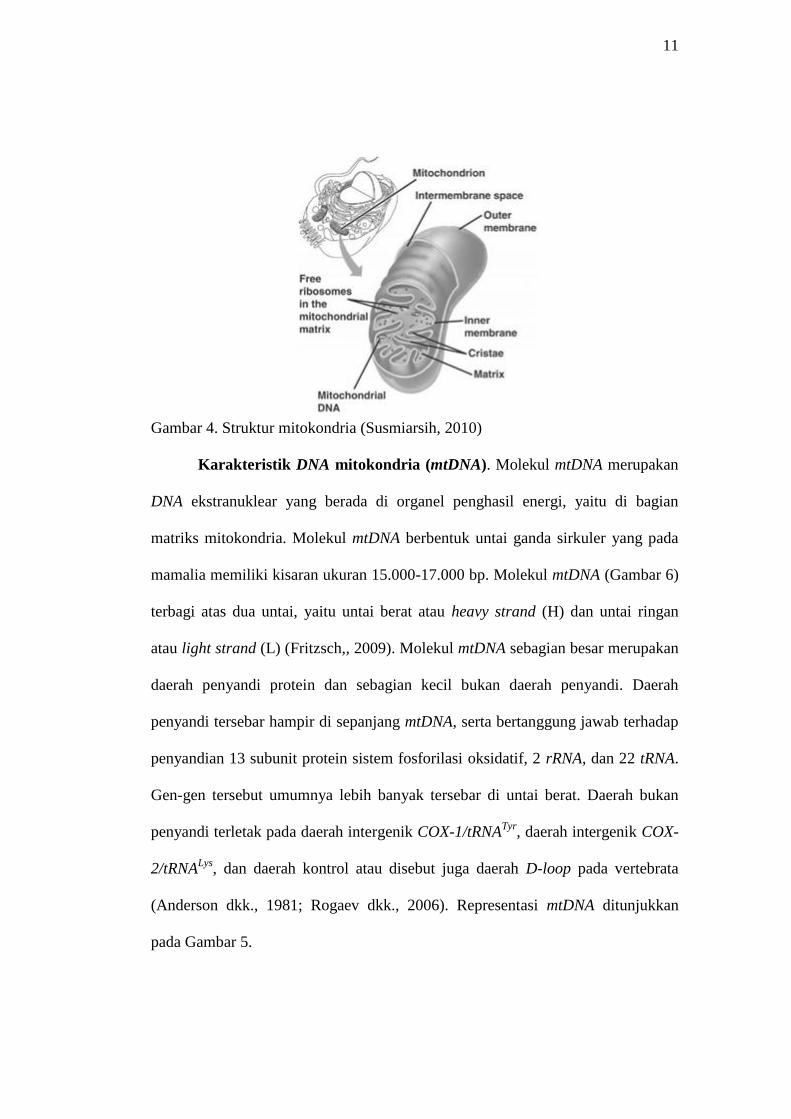
11
Gambar 4. Struktur mitokondria (Susmiarsih, 2010)
Karakteristik DNA mitokondria (mtDNA). Molekul mtDNA merupakan
DNA ekstranuklear yang berada di organel penghasil energi, yaitu di bagian
matriks mitokondria. Molekul mtDNA berbentuk untai ganda sirkuler yang pada
mamalia memiliki kisaran ukuran 15.000-17.000 bp. Molekul mtDNA (Gambar 6)
terbagi atas dua untai, yaitu untai berat atau heavy strand (H) dan untai ringan
atau light strand (L) (Fritzsch,, 2009). Molekul mtDNA sebagian besar merupakan
daerah penyandi protein dan sebagian kecil bukan daerah penyandi. Daerah
penyandi tersebar hampir di sepanjang mtDNA, serta bertanggung jawab terhadap
penyandian 13 subunit protein sistem fosforilasi oksidatif, 2 rRNA, dan 22 tRNA.
Gen-gen tersebut umumnya lebih banyak tersebar di untai berat. Daerah bukan
penyandi terletak pada daerah intergenik COX-1/tRNATyr
, daerah intergenik COX-
2/tRNALys
, dan daerah kontrol atau disebut juga daerah D-loop pada vertebrata
(Anderson dkk., 1981; Rogaev dkk., 2006). Representasi mtDNA ditunjukkan
pada Gambar 5.

12
Gambar 5. Representasi mtDNA (Krause dkk., 2006)
Molekul mtDNA memiliki beberapa karakteristik yang unik dibanding
DNA inti. Molekul mtDNA memiliki banyak salinan sehingga mtDNA lebih
mudah diisolasi dengan jumlah sampel yang sedikit (Castro dkk., 1998; Morin
dkk., 2001; Hoong dan Lex, 2005; Pakendorf dan Stoneking, 2005).
Molekul mtDNA diwariskan secara maternal, kecuali pada beberapa
spesies seperti remis, lalat buah, tikus, dan burung hibrida (hasil hibridasi dari dua
spesies burung yang berbeda). Pola pewarisan maternal disebabkan mitokondria
sel sperma tidak ikut menembus sel telur pada saat fertilisasi (Ankel-simons dan
Cummins, 1996). Molekul mtDNA yang diwariskan bukan merupakan hasil
rekombinasi. Pewarisan uniparental yang demikian akan memudahkan
pengungkapan silsilah kekerabatan di masa lalu berdasarkan garis keturunan
maternal, tanpa harus dibaurkan dengan pengaruh yang muncul akibat
rekombinasi dan pewarisan biparental (Pakendorf dan Stoneking, 2005; Galtier
dkk., 2009).

13
Molekul mtDNA memiliki laju mutasi yang lebih tinggi (10-8
substitusi/tahun) dibanding DNA inti (10-9
substitusi/tahun), khususnya pada
daerah D-loop (Pakendorf dan Stoneking, 2005). Mutasi yang terjadi umumnya
berupa mutasi titik, namun dapat pula berupa delesi atau insersi. Laju mutasi yang
tinggi disebabkan mtDNA rentan terhadap peristiwa mutagenik serta produksi
radical oxydative species (ROS) dari proses fosforilasi oksidatif di organel
tersebut (Rose dkk., 2007). Molekul mtDNA tidak memiliki protein histon serta
enzim perbaikan untuk kesalahan replikasi atau kerusakan DNA, sehingga mutasi
cenderung diteruskan dari waktu ke waktu. Laju mutasi yang tinggi
mengakibatkan mtDNA mampu mengakumulasi polimorfisme 10-17 kali lebih
cepat dari DNA inti, sehingga dapat menunjukkan variasi yang tinggi pada
berbagai level, baik antar individu maupun populasi (Castro dkk., 1998).
C. Gen Cytochrome Oxidase Sub-unit 2 (COX-2)
Cytochrome Oxidase (COX) merupakan komponen rantai respirasi
terutama dalam proses reduksi oksigen menjadi air. Cytochrome Oxidase
memiliki tiga sub-unit, yaitu COX-1, COX-2, dan COX-3. Ketiga sub-unit ini
disandi dalam genom mitokondria dan paling sedikit ada empat sub-unit yang
disandi dalam nukleus (Adkins dan Honeycutt, 1994).
Gen COX-1 dan COX-2 merupakan enzim pengaturan utama yang terlibat
dalam produksi prostaglandin dan prostanoids lainnya. Prostaglandin merupakan
mediator penting dalam berbagai proses biologis normal, termasuk fungsi
kekebalan tubuh dan biologi reproduksi. Gen COX-1 konstitutif dinyatakan dalam
sebagian besar jaringan (Harper dan Tyson-Capper, 2008). Gen COX-3 berperan

14
dalam perakitan dan stabilisasi seluruh kompleks enzim dan terlibat dalam
translokasi proton atau dalam konversi energi oleh cytochrome oxidase (You dkk.,
2002).
Ekspresi gen COX-2 diatur di berbagai tingkatan level. Aktivasi transkripsi
pada 5’ untranslated region (UTR) dari gen COX-2 melibatkan cross-talk antara
beberapa jalur sinyal yang bergantung pada jenis sel dan stimulus. Peristiwa
pemrosesan RNA yang melibatkan interaksi kompleks dari ss-RNA non-coding
pendek yang mengatur regulasi gen pasca-trankripsi dengan mempengaruhi
stabilitas messenger RNA (mRNA) dan / atau represi translasi target binding
protein mengikat trans-acting yang spesifik, dan miRNAs (miRNAs merupakan
RNA non-coding pendek yang mengatur regulasi gen pasca-transkripsi dengan
mempengaruhi stabilitas mRNA dan / atau represi translasi target mRNA), dengan
unsur-unsur cis-acting yang berbeda dalam COX-2 3’UTR menambahkan lapisan
tambahan kompleksitas regulasi gen COX-2 (Harper dan Tyson-Capper, 2008).
Tingkat pergantian asam amino pada gen COX-2 dapat dilihat di sepanjang
garis keturunan primata, sedangkan pada artiodactyl dan rodensia tidak ada
heterogenitas tingkat pergantian asam amino. Hubungan filogenik pada tingkat
molekuler dari artiodactyl belum sepenuhnya dipelajari dibandingkan dengan
primata (Adkins dkk., 1996).

15
D. Teknik Biologi Molekuler
1. Isolasi DNA
Pada organisme tingkat tinggi seperti pada manusia, hewan, dan tumbuhan
DNA terdapat di dalam inti sel, dan beberapa organ lain di dalam sel seperti
mitokondria dan kloroplast. Prinsip dasar isolasi DNA adalah serangkaian proses
untuk memisahkan DNA dari komponen-komponen sel lainnya. Isolasi DNA dari
organisme eukariotik (manusia, hewan, dan tumbuhan) dilakukan melalui proses
penghancuran dinding sel (lysis of cell walls), penghilangan protein dan RNA (cell
digestion), pengendapan DNA (prepcipitation of DNA) dan pemanenan (Sulandari
dan Zein, 2003).
2. Polymerase Chain Reaction (PCR)
Reaksi polimerase berantai atau dikenal sebagai polymerase chain
reaction (PCR) merupakan suatu proses sintesis enzimatik untuk mengamplifikasi
nukleotida secara in vitro (Fatchiyah dkk., 2011). Polymerase chain reaction
merupakan suatu teknik perbanyakan molekul DNA dengan ukuran tertentu secara
enzimatik melalui mekanisme perubahan suhu (Sulandari dan Zein, 2003).
Metode PCR dapat meningkatkan jumlah urutan DNA sebanyak ribuan
bahkan jutaan kali dari jumlah semula, sekitar106
- 107 kali. Setiap urutan basa
nukleotida yang diamplifikasi akan menjadi dua kali jumlahnya. Pada setiap n
siklus PCR akan diperoleh akan diperoleh 2n kali banyaknya DNA target.
Metode ini digunakan untuk identifikasi penyakit genetik, infeksi oleh
virus, diagnosis dini penyakit seperti Acquired Immune Deficiency Syndrome
(AIDS), profil genetik dalam forensik, aplikasi dalam biodiversitas, evolusi

16
biologi, mutasi gen secara langsung dan mengukur kuantifikasi ekspresi mRNA di
dalam sel atau jaringan (Fatchiyah dkk., 2011).
Komponen PCR yang harus dipersiapkan untuk reaksi PCR yaitu:
a. Cetakan DNA
Ukuran target amplifikasi biasanya kurang dari 1000 pasangan basa (bp)
atau 1kB. Hasil amplifikasi yang efisien adalah antara 100-400 bp. Hasil
amplifikasi lebih dari 1 kb memiliki proses yang kurang efisien karena produk
yang panjang akan rentan terhadap inhibitor yang mempengaruhi kerja enzim
DNA polymerase dan waktu yang diperlukan lebih lama (Fatchiyah dkk., 2011).
b. Primer
Primer disusun dari urutan oligonukleotida sepanjang 15-32 bp pada
ujung-5’ pita DNA cetakan maupun komplemennya (Fatchiyah dkk., 2011). Suhu
annealing sangat tergantung pada primer dengan melting temperature (Tm)
tertentu (Sulandari dan Zein, 2003; Fatchiyah dkk., 2011).
c. Taq DNA Polymerase
Enzim ini bersifat termostabil dan diisolasi dari Thermus aquaticus.
Aktivitas polimerasi DNA dari ujung 5’ ke 3’, dan aktivitas enzimatik ini
mempunyai waktu paruh sekitar 40 menit pada suhu 95ºC. Penggunaan enzim ini
harus memperhatikan proses penyimpanan (selalu di freezer pada suhu -20ºC) dan
pada saat pengambilan tidak boleh terlalu lama di suhu ruang, diusahakan selalu
dalam kotak berisi water-ice (potongan es diberi air sedikit agar suhu tetap 4 ºC).
Hal ini dilakukan untuk meminimalkan kerusakan enzim yang mungkin terjadi
akibat pengaruh perubahan suhu (Fatchiyah dkk., 2011).

17
d. Buffer PCR dan konsentrasi Mg2+
Buffer standar untuk PCR tersusun atas 50 mM KCl, 10 mM Tris-Cl (pH
8,3), dan 1,5 mM MgCl2. Buffer ini akan bekerja dengan baik untuk cetakan DNA
dan primer dengan kondisi tertentu, tetapi mungkin tidak optimal dengan
kombinasi yang lain. Konsentrasi ion magnesium dalam buffer PCR merupakan
faktor yang sangat kritis, karena kemungkinan dapat mempengaruhi proses
annealing primer, suhu disosiasi untai cetakan DNA, dan produk PCR (Sulandari
dan Zein, 2003; Fatchiyah dkk., 2011).
e. Nukleotida (dNTP)
Konsentrasi setiap dNTP 50 mM pada titik estimasi Km harus selalu diatur
pH 7,0. Konsentrasi yang tinggi akan menimbulkan ketidakseimbangan dengan
enzim polimerase, sedangkan pada konsentrasi rendah akan memberikan
ketepatan dan spesifitas yang tinggi tanpa mereduksi hasil akhir (Sulandari dan
Zein, 2003; Fatchiyah dkk., 2011).
f. Thermo cycler
Alat ini secara tepat meregulasi suhu dan siklus waktu yang dibutuhkan
untuk reprodusibilitas dan keakuratan reaksi amplifikasi. Siklus PCR terbagi atas
tiga langkah utama yaitu denaturasi DNA (92-95ºC, selama 30-60 detik), primer
annealing (50-62ºC, selama 30-60 detik), dan ekstensi atau elongasi (70-72ºC,
selama 30-120 detik). Siklus ini berulang 30-35 kali. Siklus utama ini diawali
terlebih dahulu dengan denaturasi awal, misalnya 94ºC selama 5 menit. Langkah
ini dilakukan untuk memaksimalkan proses denaturasi cetakan DNA. Bila
denaturasi tidak sempurna akan menyebabkan kegagalan proses PCR. Setelah

18
siklus utama berakhir, maka ditambah program ekstensi final dengan suhu 70-
72ºC selama 7-10 menit (Fatchiyah dkk., 2011).
3. Elektroforesis DNA
Elektroforesis merupakan proses bergeraknya molekul bermuatan pada
suatu medan listrik (Fatchiyah dkk., 2011). Elektroforesis adalah proses migrasi
dari fragmen DNA di dalam gel yang direndam dalam larutan penyangga.
Fragmen DNA yang lebih kecil berat molekulnya akan berjalan lebih cepat dari
molekul DNA yang lebih besar (Sulandari dan Zein, 2003). Perjalanan molekul
DNA di dalam gel mengikuti arus listrik dari kutub negatif menuju kutub positif
(Sulandari dan Zein, 2003; Fatchiyah dkk., 2011). Semakin besar tegangan arus
listrik, perjalanan molekul DNA akan semakin cepat, demikian pula sebaliknya
(Sulandari dan Zein, 2003).
Elektroforesis untuk makromolekul memerlukan matriks penyangga untuk
mencegah terjadinya difusi karena timbulnya panas dari arus listrik yang
digunakan. Gel poliakrilamida dan agarosa merupakan matriks penyangga yang
banyak dipakai untuk pemisahan protein dan asam nukleat (Fatchiyah dkk., 2011).
Elektroforesis gel agarosa merupakan metode standar yang digunakan
untuk identifikasi, pemisahan, dan purifikasi fragmen DNA. Migrasi elektroforesis
DNA melalui gel agarosa dipengaruhi oleh faktor ukuran dan konformasi molekul
DNA, konsentrasi agarosa, arus listrik, dan suhu. Pewarna etidium bromide (EtBr)
digunakan untuk alat idenifikasi dan mengukur semikualitatif fragmen DNA yang
terpisah dalam gel. Pewarna ini akan terikat di antara dua untai ganda DNA,
sehingga pita DNA dalam gel agarosa akan berpendar karena pewarna ini

19
mengandung zat fluoresen. Ikatan DNA-EtBr ini akan terekspos sinar ultraviolet
(UV) level medium, sekitar panjang gelombang λ 300 nm. Etidium bromide dapat
diberikan pada setiap sampel yang akan dimasukkan ke sumur gel atau
dicampurkan ke gel agarosa sebelum gel dicetak dalam cetakan gel (Fatchiyah
dkk., 2011).
Beberapa faktor yang mempengaruhi proses migrasi DNA atau RNA yaitu
konsentrasi agarosa, ukura molekul, voltase dan suhu. Molekul besar seperti
genom utuh, dielektroforesis dengan agarosa berkonsentrasi 0,8%, sedangkan
hasil amplifikasi DNA dielektroforesis dengan konsentrasi yang lebih tinggi yaitu
1,5-2%. Ukuran dan struktur molekul DNA/RNA mempengaruhi mobilitas saat
dielektroforesis. Secara berurutan mobilitas molekul berbeda dalam hal kecepatan:
sirkuler lebih cepat dibanding linear, fragmen DNA lebih cepat dibanding genom
utuh. Molekul DNA/RNA bermuatan negatif. Voltase 100 Volt biasa digunakan
untuk analisis rutin, sedangkan bila diperlukan pemisahan yang sempurna, maka
digunakan arus listrik 50 Volt. Pada voltase ini, arus lebih lambat, tetapi hasil
pemisahannya lebih maksimal. Molekul DNA akan cepat terurai pada suhu tinggi
dan akan kembali menyatu bila suhu mendingin (Fatchiyah dkk., 2011).
4. Sekuensing DNA
Prinsip penentuan urutan (sekuensing) basa DNA melibatkan produksi
seperangkat molekul/fragmen DNA yang berbeda-beda ukurannya tetapi salah
satu ujungnya selalu sama. Selanjutnya, fragmen-fragmen ini
dimigrasikan/dipisahkan menggunakan elektroforesis gel poliakrilamid atau

20
polyacrylamide gel electrophoresis (PAGE) agar pembacaan sekuens dapat
dilakukan (Susanto, 2008).
Saat ini sekuensing dapat dilakukan dengan otomatis menggunakan mesin
sequensing. Metode ini berdasarkan teknik yang disebut dengan
dideoxyribonucleotide chain termination method. Metode ini merupakan
pengembangan dari metode Sanger. Metode ini dikembangkan oleh British
biochemist, Frederick Sanger (Campbell dkk., 2009).
Metode Sanger merupakan metode sekuensing yang ditemukan oleh
Frederick Sanger pada tahun 1977. Prinsip kerjanya yaitu terminasi sintesis DNA
oleh dideoksinukleotida yang ditempatkan pada empat tabung yang berbeda.
Terminasi sintesis DNA akan menghasilkan chain terminating dideoxynucleotide,
sehingga terbentuk fragmen-fragmen dengan ukuran yang beragam. Pembacaan
fragmen-fragmen tersebut dilakukan melalui elektroforesis dengan
mengidentifikasi jenis dideoksinukleosida yang digunakan untuk terminasi
(Susanto, 2008).
5. Analisis Data
Program MEGA 6.06. Program Molecular Evolutionary Genetics
Analysis (MEGA) merupakan perangkat lunak untuk menjelajahi, menemukan,
dan menganalisis urutan DNA dan protein dari perspektif evolusi. Program
MEGA 6.06 memungkinkan untuk mengalokasikan dua kali lebih banyak memori
pada sistem 64-bit seperti pada MEGA 5. Metode waktunya yang ditambahkan
dalam MEGA 5 telah digantikan oleh sistem RealTime berbasis yang akurat (atau

21
lebih baik dari) metodologi kontemporer, tetapi dengan kecepatan lebih dari 1.000
kali lebih cepat (Tamura dkk., 2013).

22
BAB III
MATERI DAN METODE
A. Tempat dan Waktu
Penelitian ini dilaksanakan di Laboratorium Biokimia, Fakultas
Kedokteran Hewan, Universitas Gadjah Mada, Yogyakarta pada bulan Agustus
2013 sampai Januari 2015.
B. Materi Penelitian
1. Alat Penelitian
Alat yang digunakan dalam penelitian ini antara lain mikropipet (Gilson)
dengan ukuran 1-10 µl, 20-100 µl, dan 200-1000 µl, waterbath shaker (Eyla, Uni
Thermo Shaker NTS 3000), mini sentrifuge 12 (Hanil 5804R), sentrifuge (Model
DSC 158), freezer, thermocycler (Infigen Biotech, Inc.), seperangkat alat
elektroforesis (Mupid-Exu), microwave oven (Sanyo model EM-ST28),
seperangkat pencetak agar (plat dan sisir pencetak sumuran), timbangan digital
(Mettler Toledo PB303-S), magnetic stirer, vortex mixer (Maxi Mix II), gelas
ukur, tabung erlenmeyer, eppendorf, rak tabung, spin down, dan ultraviolet
transluminator (UVP® 95-0225-02).
2. Bahan Penelitian
Bahan yang digunakan dalam penelitian ini adalah darah rusa Timor yang
diambil di Taman Rusa, Universitas Gadjah Mada, Yogyakarta dengan kode

23
APRTUGM574, APRTUGM583, APRTUGM573, APRTUGM584 dan
APRTUGM364. Isolasi DNA menggunakan Genomic DNA Mini Kit (Geneaid)
yang berisi RBC lysis buffer, GB buffer, ethanol absolut, wash buffer, elution
buffer, GD column, dan collection tube. Bahan yang digunakan dalam PCR yaitu
readymix Taq DNA Polymerase (Kappa Taq DNA Polymerase), primer, sampel
DNA, dan ddH2O. Primer yang digunakan adalah primer forward berkode
APRA8C2F (10 pmol), primer reverse berkode APRA8C2R (10 pmol). Bahan
yang digunakan dalam elektroforesis antara lain agarose, marker DNA 1 kb
(Geneaid), Florosafe DNA Stain (1st BASE), buffer elektroforesis Tris-Borat
EDTA (TBE)1X yang diencerkan dari TBE 5X (1,625 gram Tris base, 13,75
gram asam borat, 1,875 gram EDTA (pH 8,0) dalam 500 ml H2O), dan loading
dye gliserin bromphenol blue (GBB).
C. Metode Penelitian
1. Bagan Penelitian
Koleksi sampel darah
Isolasi DNA Total
Amplifikasi Gen COX-2 dengan Teknik PCR
Elektroforesis Gel Agarose
Sekuensing DNA
Analisis data (Mega 6.06)

24
2. Koleksi Sampel Darah
Bahan utama penelitian yaitu lima sampel darah rusa Timor yang diambil
dari vena jugularis. Darah rusa tersebut dimasukkan ke dalam tabung berisi EDTA
dan tabung digoyang-goyang membentuk angka delapan agar darah tidak
menjendal dan rusak, kemudian darah disimpan dalam refrigerator suhu 4ºC.
3. Isolasi DNA Total
Isolasi DNA total menggunakan kit Genomic DNA Mini Kit (Geneaid).
Sebelum dilakukan isolasi DNA, masing-masing sampel darah disentrifugasi
selama lima menit dengan kecepatan 8000 rpm. Buffy coat dipisahkan dari
komponen darah yang lain dan dimasukkan ke dalam eppendorf yang berbeda.
Setiap eppendorf tersebut diberi label sesuai dengan kode sampel dan disimpan
dalam freezer pada suhu -20oC.
Buffy coat sebanyak 200 µl dimasukkan ke dalam eppendorf, kemudian
ditambahkan 600 µl RBC lysis buffer dan diinkubasi selama 10 menit pada suhu
ruang, diinversi setiap tiga menit. Selanjutnya campuran disentrifugasi 13.500
rpm selama tiga menit. Setelah terlihat batas supernatan (bagian atas) dan pelet
(bagian bawah), maka supernatan dibuang. Pelet ditambahkan 500 µl RBC lysis
buffer dan diresuspensi. Campuran tersebut ditambahkan 250 µl GB buffer dan di-
vortex dengan kecepatan tinggi selama satu menit. Campuran tersebut
diinkubasikan pada waterbath-shaker dengan kecepatan 25 rpm selama satu jam
pada suhu 60ºC. Dalam waktu dan suhu yang sama, 400 µl elution buffer
diinkubasikan juga.

25
Setelah diinkubasikan, campuran ditambahkan 250 µl ethanol absolut dan
segera di-vortex selama 10 detik. Bila ada endapan, maka larutan diresuspensi.
Campuran dimasukkan ke dalam collection tube baru melalui GD column
(terdapat filter di bawahnya) yang telah dipasang sebelumnya, kemudian
disentrifugasi dengan kecepatan 13500 rpm selama lima menit dan GD column
dipasang pada collection tube yang baru. Wash buffer 400 µl ditambahkan melalui
GD column dan disentrifugasi dengan kecepatan 13.500 rpm selama satu menit,
kemudian cairan yang ada dalam collection tube dibuang, GD column dipasang
kembali di atas collection tube tersebut. Wash-buffer 600 µl ditambahkan melalui
GD column dan disentrifugasi dengan kecepatan 13.500 rpm selama satu menit.
Cairan yang ada dalam collection tube dibuang, kemudian GD column dipasang
kembali di atas collection tube tersebut. Dengan kecepatan yang sama dengan
sebelumnya yaitu 13.500 rpm collection tube disentrifugasi kembali selama tiga
menit. Langkah selanjutnya adalah GD column dipindah di atas eppendorf steril
yang baru, kemudian ditambahkan 100 µl elution buffer (yang sudah
diinkubasikan sebelumnya pada suhu 60oC) melalui GD column). Beker glass
yang telah diisi air panas dari waterbath disiapkan dan ditambahkan air kran
sampai suhu menjadi 37oC (diukur memakai termometer). Setelah itu, GD column
beserta eppendorf diinkubasikan ke dalam beker glass tersebut dengan suhu 37oC
selama 10 menit. Setelah diangkat dari beker glass, dilakukan sentrifugasi dengan
kecepatan 13.500 rpm selama satu menit. Elution buffer 100µl ditambahkan
kembali dan diinkubasikan selama 10 menit pada suhu 37ºC dalam beker glass,
sehingga diperoleh DNA yang terlarut dalam 200 µl elution buffer.

26
Kualitas DNA hasil isolasi dilihat dengan dielektroforesis pada gel agarose
1%. Loading sampel ke dalam sumuran dilakukan dengan komposisi 8 µl sampel
ditambah dengan 2 µl GBB. Elektroforesis dilakukan dengan arus listrik 90 Volt
selama 30 menit.
4. Amplifikasi Gen COX-2 dengan Teknik PCR
Hasil isolasi DNA digunakan sebagai cetakan atau template DNA dalam
proses amplifikasi dengan teknik PCR. Amplifikasi gen COX-2 menggunakan
primer APRA8C2F dan APRA8C2R. Susunan basa primer dan melting
temperature (Tm) disajikan dalam Tabel 1.
Tabel 1. Urutan basa primer serta melting dan annealing temperature Primer Susunan basa Tm (ºC) Ta (
oC)
APRA8C2F 5’ – GCT GGT TTC AAG CCA ACA TC – 3’ 54,8 52
APRA8C2R 5’ – TGT TGG AGG GAA ATA AGG CG – 3’ 55,1 52
Komposisi campuran pereaksi PCR DNA untuk satu reaksi 50 µl adalah
25 µl Kappa Taq DNA Polymerase readymix, 2 µl 10 pmol untuk masing-masing
primer APRA8C2F dan APRA8C2R, 16 µl DNA total dari Rusa timorensis dan
ditambahkan ddH2O 5 µl. Kondisi thermocycler untuk amplifikasi DNA adalah
pre-denaturasi pada suhu 94ºC selama 5 menit, denaturasi pada 94ºC selama satu
menit, annealing atau proses penempelan pada 52ºC selama 45 detik, elongation
atau proses pemanjangan pada suhu 72ºC selama satu menit, dan post-elongation
pada suhu 72ºC selama lima menit. Reaksi dilakukan sebanyak 35 siklus.
5. Elektroforesis Gel Agarose
Proses elektroforesis dengan gel agarose dilakukan untuk melihat hasil
isolasi DNA dan PCR. Proses yang dilakukan pertama kali adalah pembuatan gel.
Konsentrasi gel yang digunakan adalah 1% yaitu 0,25 gram agarose dilarutkan

27
dengan 25 ml TBE 1X atau 0,5 gram agarose dalam 50 ml TBE 1X. Volume 25
ml mencukupi untuk membuat gel agarose dengan delapan sumuran, sedangkan
volume 50 ml mencukupi untuk membuat gel agarose dengan 17 sumuran.
Pelarutan agarose dalam TBE 1X dengan cara dipanaskan di dalam microwave
oven selama 45 detik. Setelah itu, dalam gel cair ditambahkan Florosafe DNA
sebagai pewarna sebanyak 1-2 µl untuk volume gel 25 ml dan 3 µl untuk volume
gel 50 ml. Cetakan yang terbuka ditutup dengan isolasi dan diletakkan di dudukan
cetakan, kemudian sisir diletakkan di atasnya untuk menciptakan sumuran pada
gel. Jika gel cair sudah tidak terlalu panas (suhu sekitar 55ºC), gel cair dituangkan
ke dalam plat pencetak agar hingga mengeras.
Setelah gel mengeras, plat yang berisi gel agarose diletakkan dalam bak
elektroforesis yang berisi larutan buffer sesuai dengan buffer yang digunakan
untuk membuat gel agarose yaitu TBE 1X. Sampel hasil PCR sebanyak 3 µl
diambil dengan mikropipet kemudian dicampur dengan gliserin bromphenol blue
(GBB) sebanyak 1 µl dan dimasukkan ke dalam sumuran agarose. Elektroforesis
dilakukan dengan arus listrik 90 Volt dan ditunggu hingga proses selesai sekitar ±
30 menit. Pita molekul DNA diamati dengan bantuan ultraviolet transluminator
(λ=260 nm).
6. Sekuensing DNA
Sekuensing DNA untuk menentukan runutan nukleotida dilakukan oleh
First Base International di Singapura menggunakan metode Sanger (metode
terminasi rantai). Sekuensing DNA dilakukan sebanyak dua kali reaksi untuk
masing-masing sampel, yaitu menggunakan primer APRA8C2F dan APRA8C2R

28
masing-masing dengan konsentrasi 10 pmol. Hasil PCR digunakan sebagai
cetakan dalam reaksi sekuensing. Komponen lain yang digunakan selain hasil
PCR dan primer adalah enzim Taq Polymerase, dNTP dan ddNTP.
7. Analisis Data
Analisis data hasil sekuensing menggunakan program MEGA versi 6.06.
Hasil sekuensing gen dilakukan penjajaran berganda dengan program Clustal W
(Thompson dkk., 1994). Berdasarkan hasil sekuensing (perunutan nukleotida),
analisis gen dilakukan berdasarkan urutan nukleotida dan asam amino. Urutan
asam amino diolah dari basa yang diterjemahkan mengikuti vertebrate
mitochondrial translation code. Untuk penjajaran urutan nukleotida dari 765
sampai 2.000 bp diperlukan MEGA versi 6.06 yang sangat efisien baik dari segi
kinerja maupun memori yang dibutuhkan (Tamura dkk., 2013).
Sekuen gen penyandi COX-2 dari spesies rusa lain dan dari famili lain
yang digunakan sebagai pembanding diambil dari Genbank, antara lain Rusa
timorensis (NC_020745.1); spesies-spesies dalam rumpun Cervini yaitu yaitu
Rusa unicolor swinhoei (NC_008414.3), Rusa alfredi (NC_020744.1), Cervus
elaphus (AB245427.2), Cervus nippon yesoensis (AB210267.2) dan Axis axis
(NC_020680.1); spesies dalam rumpun Muntiacini yaitu Muntiacus muntjak
(NC_004563.1); dan spesies dalam famili lain (Bovidae) yaitu Bos javanicus
(NC_012706.1).