sejarah evolusi suatu organisme tersimpan di dalam genomnya
DESCRIPTION
evolusiTRANSCRIPT

1
Sejarah Evolusi Suatu Organisme Tersimpan di dalam Genomnya
Famili Gen Sox (Sry-like HMG box) Pada Berbagai Organisme
ARTIKEL
Disusun untuk memenuhi tugas mata kuliah Konsep dan Teori Evolusi
Kode Mata Kuliah: BI5201
Dosen Pengampu: Prof. Intan Ahmad, Ph.D.
Oleh: Rini Agustin (20613011)
Mahasiswa Program Studi Biologi Jurusan Biologi Sekolah Ilmu Teknologi Hayati ITB
Pendahuluan
Salah satu bukti evolusi adalah berasal dari analisis kesamaan antara organisme yang berbeda.
Oleh karena evolusi adalah merupakan proses penurunan dengan modifikasi, dimana karakteristik
yang ada pada organisme nenek moyang berubah (melalui seleksi alam) pada keturunannya seiring
waktu ketika organisme berhadapan dengan kondisi lingkungan yang berbeda-beda. Akibatnya,
spesies yang berkerabat dapat memiliki karakteristik dengan kesamaan yang mendasar. Kesamaan
yang berasal dari nenek moyang yang sama dikenal sebagai homologi. Ahli biologi juga mengamati
kemiripan organisme selain dari morfoloi, juga pada tingkat molekuler. Semua bentuk kehidupan
menggunakan bahasa genetika yang sama yakni DNA dan RNA. Sehingga kemungkinannya bahwa
semua spesies merupakan keturunan dari nenek moyang bersama yang menggunakan kode ini
(Campbell, et al., 2010).
Salah satu bukti sejarah evolusi suatu organisme adalah tersimpan di dalam genomnya.
Pembahasan artikel ini selanjutnya akan melihat analisis famili gen Sox pada berbagai organisme.
Analisis molekular Sox bermula pada analisis genom yang berperan dalam determinasi sex yang
teridentifikasi pada DNA sekuens gen Sry yang terletak pada kromosom Y ditemukan pada tahun
1990 oleh Sinclair et.al. dan Gubbay et al. Perkembangan pengetahuan molekular sampai pada
kesimpulan bahwa selain gen Sry juga ditemukan sekuens lain dengan konservasi sekuens yang
memiliki kemiripan dengan gen sebelumnya. Pada penelitian lebih lanjut sampailah pada
perkembangan penemuan berbagai sekuens gen serupa dengan gen Sry dan Sox ini yang kemudiaan
teridentifikasi sebagai Sox1, Sox2, Sox3, Sox4, Sox5, Sox6, dan seterusnya. Analisis kesamaan pola
pengulangan sekuens yang menyatakan adanya kesamaan terkonservasinya suatu sekuens DNA pada
gen tertentu dikenal sebagai famili gen. Selanjutnya Sry dan Sox ini termasuk ke dalam family gen
Sox dengan superfamili gen HMGbox (Soullier et al., 1999).
Famili gen Sox memiliki peran yang sangat penting dalam pembentukan suatu jaringan dan
organ pada proses perkembangan awal suatu organisme pada saat embrio. Gen Sox juga memiliki
peran dalam menjaga fungsi normalitas dari sel tertentu setelah proses kelahiran. Dengan demikian,
dalam perannya tersebut, gen-gen yang termasuk ke dalam famili gen Sox berperan untuk
memberikan perintah dan mempengaruhi suatu protein untuk menempel dan bekerja pada wilayah
tertentu DNA. Dengan penempelan suatu protein pada wilayah spesifik DNA yang memiliki jarak
yang dekat dengan gen, protein Sox dapat membantu mengontrol aktifitas gen tersebut dalam bekerja.
Pada kondisi ini, protein Sox ini juga seringkali disebut sebagai sebuah protein faktor transkripsi
(USA gov., 2013).
High Mobility Group (HMG) box didefinisikan sebagai super famili dari protein yang dapat
berikatan pada DNA organisme eukariotik sebagai pusat terpenting regulasi gen pada mamalia (Ner,
1992). Anggota famili gen Sox memiliki DNA-binding HMG domain yang memiliki 240 sekuens
nukleotida yang mengkode sekitar 79 asam amino. Gen HMG-box umumnya merupakan faktor

2
transkripsi regulasi ekspresi gen selama proses differensiasi sel dalam penentuan peta nasib embrio
(Wang, et al., 2009). Protein ekspresi HMG-box domain ini memiliki bentuk dan struktur seperti
huruf L yang mengandung 3 α-helix dan ujung-N β-strand.
Gambar 1. Ilustrasi model pita struktur 3D HMG-box
(Sumber: Weiss, 2001)
Protein famili gen Sox merupakan faktor transkripsi hanya memiliki satu HMG-box domain
dan berikatan dengan sekuens DNA dengan konsensus DNA pada sekuens 5’-
(A/T)(A/T)CAA(A/T)G-3’, dan diklasifikasikan ke dalam kelompok HMGB (Mertin, et al., 1999).
Protein famili gen Sox dapat berfungsi sebagai aktivator transkripsi, sebagai repressor, dan sebagai
lacks transactivation domain. Anggota famili gen Sox menunjukkan homologi sekuensing asam
amino > 50% dengan sekuens asam amino pada HMG-box domain Sry. Menurut Chew & Gallo
(2009) famili gen Sox memiliki anggota kurang lebih sebanyak 30 gen (20 gen pada manusia dan
mencit, dan delapan pada Drosophila). Berdasarkan homologi sekuens DNA HMG-box domain,
struktur motif bagian luar HMG-box domain, dan karakteristik fungsionalnya famili gen Sox
diklasifikasikan menjadi 10 sub kelompok famili gen yang berbeda (Kelompok A-J) (Bowles, et al.,
2000). Protein Sox yang diklasifikasikan ke dalam satu kelompok memiliki homologi yang identik
pada sekuensing HMG-box domain dan struktur motif luar berkisar 70-90% (Lefebvre, et al., 2007).
Identifikasi famili gen Sox dilakukan dengan menggunakan alignment sekuensing HMG-box
domain pada anggota famili gen Sox. HMG-box domain dari protein Sox memiliki kesamaan
sedikitnya 50% dengan HMG-box domain dari gen Sry. Diluar sekuens HMG-box domain tersebut,
sekuens gen Sox sangat bervariasi, sehingga walaupun domain nonbox dapat diidentifikasi dari
sejumlah protein dan sekuens gen Sox, HMG-box domain ini telah menunjukkan kesamaan yang
menunjukkan kesamaan gen nenek moyang (Bowles, et al., 2000). Diantara spesies-spesies
vertebrata, gen ortolog (yang merepresentatifkan gen yang sama pada spesies yang berbeda) secara
umum memiliki kesamaan sekuensing berdasarkan sekuens HMG-box domain homologi maupun
konservasi struktural nonbox. Identifikasi sekuens HMG-box domain pada keseluruhan sekuens DNA
beberapa spesies terus dilakukan untuk melihat kesamaan dan fungsi famili gen Sox pada berbagai
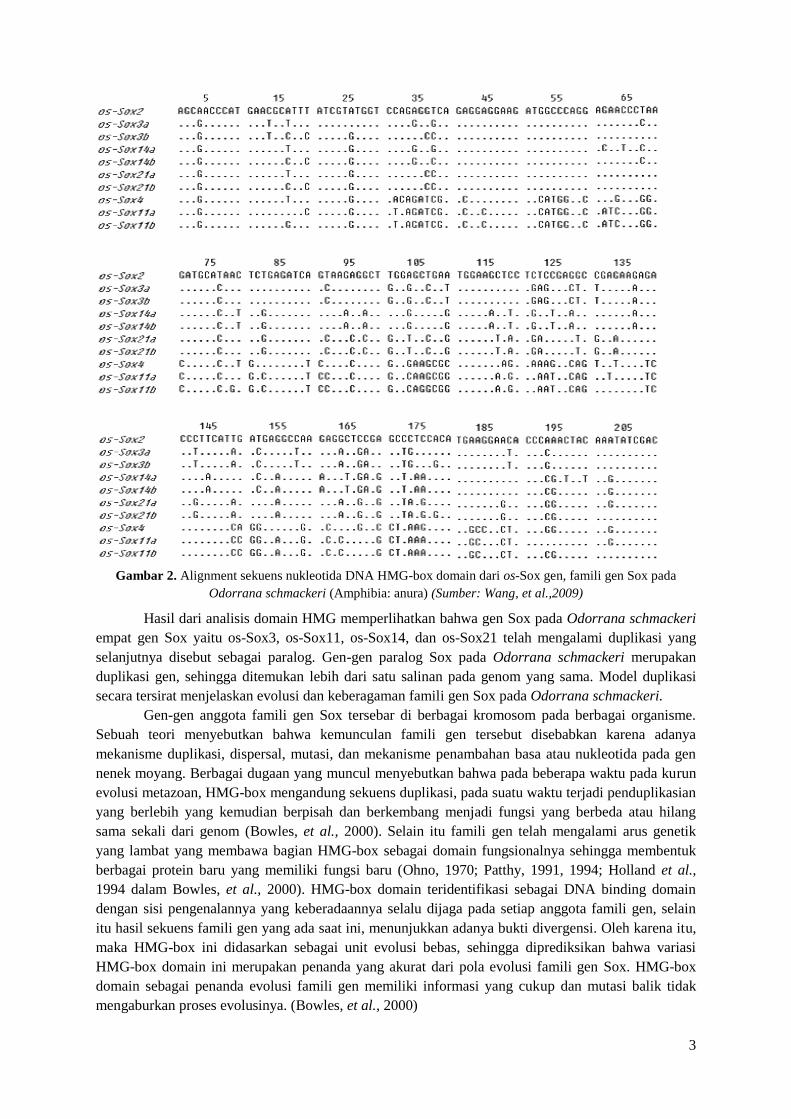
spesies. Pada gambar 2 berikut adalah sekuens HMG-box domain anggota famili gen Sox pada
spesies Odorrana schackeri (Amphibia: anura) telah menunjukkan kesamaan sekuensing DNA pada
kromosom.
3
Gambar 2. Alignment sekuens nukleotida DNA HMG-box domain dari os-Sox gen, famili gen Sox pada
Odorrana schmackeri (Amphibia: anura) (Sumber: Wang, et al.,2009)
Hasil dari analisis domain HMG memperlihatkan bahwa gen Sox pada Odorrana schmackeri
empat gen Sox yaitu os-Sox3, os-Sox11, os-Sox14, dan os-Sox21 telah mengalami duplikasi yang
selanjutnya disebut sebagai paralog. Gen-gen paralog Sox pada Odorrana schmackeri merupakan
duplikasi gen, sehingga ditemukan lebih dari satu salinan pada genom yang sama. Model duplikasi
secara tersirat menjelaskan evolusi dan keberagaman famili gen Sox pada Odorrana schmackeri.
Gen-gen anggota famili gen Sox tersebar di berbagai kromosom pada berbagai organisme.
Sebuah teori menyebutkan bahwa kemunculan famili gen tersebut disebabkan karena adanya
mekanisme duplikasi, dispersal, mutasi, dan mekanisme penambahan basa atau nukleotida pada gen
nenek moyang. Berbagai dugaan yang muncul menyebutkan bahwa pada beberapa waktu pada kurun
evolusi metazoan, HMG-box mengandung sekuens duplikasi, pada suatu waktu terjadi penduplikasian
yang berlebih yang kemudian berpisah dan berkembang menjadi fungsi yang berbeda atau hilang
sama sekali dari genom (Bowles, et al., 2000). Selain itu famili gen telah mengalami arus genetik
yang lambat yang membawa bagian HMG-box sebagai domain fungsionalnya sehingga membentuk
berbagai protein baru yang memiliki fungsi baru (Ohno, 1970; Patthy, 1991, 1994; Holland et al.,
1994 dalam Bowles, et al., 2000). HMG-box domain teridentifikasi sebagai DNA binding domain
dengan sisi pengenalannya yang keberadaannya selalu dijaga pada setiap anggota famili gen, selain
itu hasil sekuens famili gen yang ada saat ini, menunjukkan adanya bukti divergensi. Oleh karena itu,
maka HMG-box ini didasarkan sebagai unit evolusi bebas, sehingga diprediksikan bahwa variasi
HMG-box domain ini merupakan penanda yang akurat dari pola evolusi famili gen Sox. HMG-box
domain sebagai penanda evolusi famili gen memiliki informasi yang cukup dan mutasi balik tidak
mengaburkan proses evolusinya. (Bowles, et al., 2000)

4
Gambar 3. Alignment sekuensing asam amino dari protein Sox domain HMG-box.
(Sumber: Bowles, et al., 2000)

5
Ket: Sekuensing domain HMG-box dari berbagai spesies diberikan, disusun berdasarkan kelompok terkait
sebagai dasar dugaan kelompok berdasarkan kesamaan sekuensing. Tanda strip (-) menunjukkan kesamaan
asam amino dengan sekuens konsensus pada baris teratas. Simbol (|) menunjukkan posisi box intron. al,
alligator, Alligator mississippiensis; ce, nematode, Caenorhabditis elegans; ch, chicken, Gallus gallus; dr, fruit-
fly, Drosophila melanogaster; du or d, dunnart (marsupial), Sminthopsis macroura; fu, fungi, Saccharomyces
cerevisiae; hu atau h, human, Homo sapiens; mo atau m, mouse, Mus musculus; or, orangutan, Pongo
pygmaeus; pi atau p, pig, Sus scrofa; ra atau r, rat, Rattus norvegicus; tw, tammar wallaby (marsupial),
Macropus eugenii; sh atau s, sheep, Ovis aries; tr, rainbow trout, Oncorhynchus mykiss; se, sea urchin,
Strongylocentrotus purpuratus; xe, frog, Xenopus laevis; zf, zebrafish, Danio rerio.
Rekonstruksi evolusioner famili gen Sox dilakukan dengan menggunakan sekuensing HMG-
box domain dari anggota-anggota famili gen dari berbagai organisme untuk melihat kecenderungan
kesamaan dan mengetahui proses evolusi yang terjadi. Hasil alignment sekuensing HMG-box domain
pada gen Sox menunjukkan tetap terjaganya sekuens tersebut, pada alignment tidak ditemukan adanya
gap atau insersi. Hasil alignment sekuens menunjukkan adanya intron pada kelompok invertebrata
yang ditemukan pada Drosophila dan C. elegans yang merupakan anggota kelompok B dan C. Hal
tersebut telah sedikitnya menunjukkan adanya konservasi posisi dan dipertahankannya keberadaan
intron pada organisme invertebrata dan menunjukkan bahwa intron telah hilang pada keturunan
kelompok deuterostome. HMG-box intron juga ditemukan pada anggota kelompok D, E, dan F. Posisi
Sekuens intron HMG-box kelompok D dan F dipertahankan pada kedua kelompok tersebut, hal ini
menunjukkan hubungan kekerabatan invertebrata yang lebih dekat dengan nenek moyang (Bowles, et
al., 2000).
Perbedaan Sry dan Sox Lainnya yang Merupakan Evolusi Spesifik pada Spesies
Sry merupakan anggota Sox yang memiliki banyak perbedaan dengan Sox lainnya, Sry juga
memiliki homologi yang berbeda di luar daerah sekuens domain HMG-box. Keadaan ini merupakan
hasil lokalisasi keberadaan Sry pada kromosom Y. Hal ini menunjukkan bukti bahwa sekuens domain
non-HMG telah menunjukkan mekanisme fungsi yang berbeda pada berbagai spesies dan oleh karena
itu hal inilah yang memungkinkan terjadinya divergensi dan mekanisme fungsi yang berbeda pada
berbagai spesies (Bowles, et al., 2000).
Gambar 5 Skenario evolusi spesiasi dari sejarah duplikasi gen yang terdapat pada famili gen.
(Sumber: Woelk, et al. 2007)
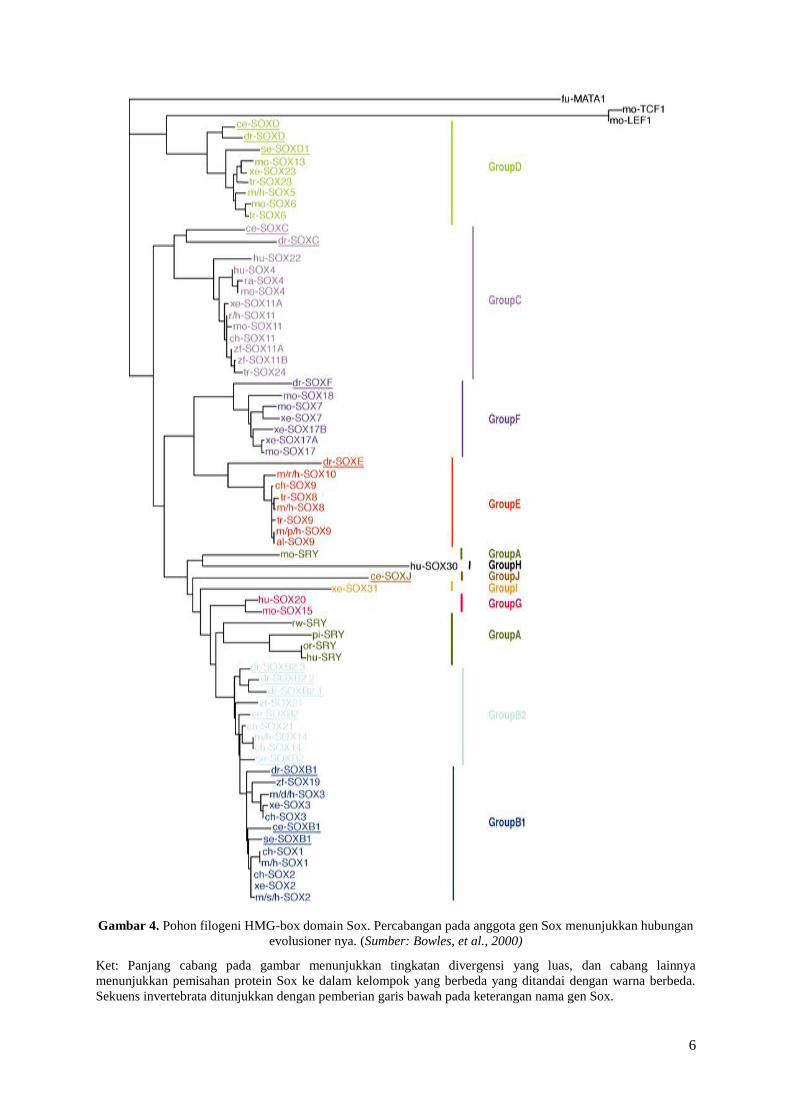
6
Gambar 4. Pohon filogeni HMG-box domain Sox. Percabangan pada anggota gen Sox menunjukkan hubungan
evolusioner nya. (Sumber: Bowles, et al., 2000)
Ket: Panjang cabang pada gambar menunjukkan tingkatan divergensi yang luas, dan cabang lainnya
menunjukkan pemisahan protein Sox ke dalam kelompok yang berbeda yang ditandai dengan warna berbeda.
Sekuens invertebrata ditunjukkan dengan pemberian garis bawah pada keterangan nama gen Sox.

7
Salah satu mekanisme Sry dan Sox sebagai faktor penentu jenis kelamin
Sebagai faktor penentu jenis kelamin, Sry sangat diperlukan pada saat perkembangan testis,
ketiadaan Sry pada organisme XY menyebabkan perkembangan genitalia betina. Sebaliknya keadaan
overekspresi gen Sry dapat menginduksi perkembangan testis pada organisme XX transgenik.
Kemampuan Sry dalam menginduksi perkembangan testis dimulai dari penginduksian diferensiasi
dari sel sertoli. Selain itu, Sox9 juga memiliki fungsi yang sama seperti Sry yang berperan penting
dalam perkembangan dan penentuan jenis kelamin organisme yang aktif setelah fungsi Sry.
Gambar 5. Hipotesis determinasi seks yang dikontrol oleh interaksi gen Sry dan Sox
(Sumber: Graves, 1998)
Hipotesis determinasi seks ini melibatkan interaksi gen Sry, Sox3, dan Sox9. Dalam hipotesis
ini peran Sry dan Sox3 yang saling berinteraksi dapat meregulasi peran Sox9 sebagai pemicu
determinasi testis. Sry memiliki kemampuan penghambatan secara tidak langsung pada
perkembangan testis. Gen antara dalam interaksi gen Sry dan Sox9 adalah Sox3 yang dikonservasi
dan berkaitan erat dengan sekuens kromosom X. Menurut hipotesis Graves adanya ekspresi gen Sox3
dapat menghambat aktivitas Sox9. Pada embrio organisme XX, ekspresi gen Sox3 dapat berikatan
dengan Sox9 dan menghambat proses transkripsinya. Sedangkan pada embrio XY, ekspresi protein
Sry dapat berkompetisi dengan daerah pengikatan pada Sox3 dan kemudian Sox3 menjadi tidak dapat
berikatan dengan Sox9 sehingga mempersilahkan Sox9 melakukan transkripsi dan melakukan inisiasi
diferensiasi pembentukan testis (Graves, 1998).
Evolusi aturan determinasi seks oleh gen Sry pada mamalia bermula pada ekspresi spesifikasi
gonad pada ayam seperti hal nya pada mencit disebabkan oleh ekspresi gen Sox9 sebagai salah satu
gen penting dalam pathway determinasi seks vertebrata sebelumnya. Determinasi seks ini lebih dulu
terjadi, dan kemudian divergensi menjadi garis keturunan spesifik menjadi determinasi seks dengan
berbagai sistem penamaan seperti XX/XY atau ZW/ZZ yang telah terjadi lebih dari 350 miliyar tahun
lalu (Graves, 1998). Sox3 diduga merupakan gen ancestor dari Sry dan memiliki banyak kesamaan
fungsi termasuk dalam diferensiasi sel sertoli (Graves, 1998). Gen Sry hanya ditemukan pada
mamalia dan berlokasi pada kromosom Y. Sehingga pada organisme yang tidak memiliki kromosom
Y, peran Sox9 yang diregulasi oleh sejumlah gen regulator dengan melibatkan gen Sox3 atau
beberapa gen lain atau bahkan hanya melibatkan gen Sox9 saja dapat membuat regulasi fungsi
determinasi seks pada organisme (Graves, 1998). Pada burung dan vertebrata lain selain mamalia
yang tidak memiliki gen Sry, kemungkinan Sox9 pada kromosom Z atau W diregulasi dengan
menggunakan mekanisme yang berbeda dengan atau tanpa adanya keterlibatan gen lain dalam
mekanismenya. Akan tetapi alur tahapan perkembangan dari diferensiasi gonad ini identik.
Mekanisme determinasi seks pada mamalia diduga melalui tahapan evolusi determinasi seks dengan
regulasi Sox3, Sox9 dan pengontrolan determinasi perkembangan testis.

8
Gambar 6. Evolusi determinasi seks mamalia melalui tahapan regulasi determinasi seks oleh Sox3
(Sumber: Graves, 1998)
Ket: Pada garis keturunan nenek moyang reptil, burung, dan mamalia sistem determinasi berdasarkan atas
kromosom lain atau determinasi seks oleh lingkungan. Sox3 pada pasangan autosom nenek moyang
berkembang menjadi kromosom X dan Y yang berinteraksi dengan gen Sox9 untuk melakukan diferensiasi sel
gonad jantan atau betina. Pada tahapan intermediet, aktifitas Sox9 diregulasi dengan adanya kadar Sox3 pada
jumlah tertentu. Diikuti dengan mutasi pada alel Sox3 untuk menon aktifkan peran menginisiasi atau
menghasilkan bentuk awal degradasi kromosom-Y. Evolusi pada sistem dominan Sry diikuti mutasi alel Sox3,
Sry berperan menghambat peran Sox3 sebagai penghambat Sox9. Sehingga dengan adanya Sry, dapat
menghambat Sox3, sehingga memperbolehkan Sox9 untuk melakukan diferensiasi.
Hubungan Filogeni Terhadap Fungsi Protein Sox
Pada beberapa kelompok famili gen Sox, konservasi sekuens HMG-box domain dan struktur
ternyata memiliki hubungan yang erat terkait dengan kesamaan fungsinya. Diantaranya pada
kelompok gen B1 menunjukan kesamaan fungsional diantaranya adalah sebagai aktivator transkripsi
perkembangan sistem saraf pusat, serta regulasi fenotip pada neuron (Collignon, et al., 1996).
Kelompok gen B2 sama-sama berperan dalam sistem saraf pusat, hanya saja perannya kali ini bukan
lagi sebagai aktivator transkripsi, melainkan sebagai repressor (Uchikawa, et al., 1999). Kelompok
gen D memiliki fungsi yang berkaitan dengan proses spermatogenesis dan chondrogenesis (Lefebvre,
et al., 1998). Serta kelompok gen E yang memiliki kesamaan dalam struktur, akan tetapi memiliki
fungsi yang berkaitan pada rentang penyakit yang luas pada manusia mengindikasikan bahwa gen-gen
tersebut telah memiliki pertimbangan terkait fungsi yang berbeda (Bowles, et al., 2000).
Walaupun telah sebelumnya teridentifikasi setiap gen mengkodekan protein yang memiliki
fungsi spesifik pada perkembangan spesifik, namun menurut Kiefer (2007) secara umum prinsip kerja
protein famili gen Sox diantaranya adalah:
1. Gen Sox meregulasi spesifikasi dan diferensiasi dari berbagai tipe sel,
2. Gen dengan kelompok gen Sox yang sama umumnya memiliki peran yang sama,
3. Gen suatu kelompok dapat menghilangkan pengaruh fungsi dari gen pada kelompok lain,
4. Gen yang sama memfasilitasi tahapan yang berbeda dari sebuah tahapan perkembangan sel
yang sama ataupun yang lebih dari satu tipe sel yang sama.

9
Referensi
Bowles, J., Schepers, G., Koopman, P. 2000. Phylogeny
of the SOX Family of Developmental
Transcription Factors Based on Sekuens and
Structural Indicators. Developmental Biology
227: 239–255
Campbell, N.A., Reece, J.B., Urry, L.A., Cain, M.L.,
Wasserman, S.A., Minorsky, P.V., Jackson,
R.B., 2010. Biologi. Eight Edition. Jakarta:
Erlangga
Collignon J, Sockanathan, Hacker AS, Cohen-Tannoudji
M, Norris D, Rastan S, et al. 1996. A
comparison of the properties of Sox-3, with
Sry and two related genes, Sox-1 and Sox-2.
Development. 122 : 509–20
Graves, J.A.M. 1998. Interactions between SRY and
SOX genes in mammalian sex determination.
BioEssays 20:264-269
Gubbay J, Collignon J, Koopman P, Capel B, Economou
A, Munsterberg A, et al. 1990. A gene
mapping to the sex-determining region of the
mouse Y chromosome is a member of a new
family of embryonically expressed genes.
Nature. 346 : 245–50
Kiefer, J.C. 2007. Back to Basics: Sox Genes.
Developmental Dynamic. 236 : 2356-2366
Lefebvre, V., Dumitriu, B., M´endez, A.P., Han, Y.,
Pallavi, B. 2007. Control of cell fate and
differentiation by Sry-related high-mobility-
group box (Sox) transcription factors. The
International Journal of Biochemistry & Cell
Biology. 39 : 2195–2214
Lefebvre, V., Li, P., de Crombrugghe, B. 1998. A new
long form of Sox5 (L-Sox5), Sox6 and Sox9
are coexpressed in chondrogenesis and
cooperatively activate the type II collagen
gene. EMBO J 17 : 5718–5733
Mertin, S., McDowall, S. G., & Harley, V. R. (1999).
The DNAbinding specificity of SOX9 and
other SOXproteins. Nucleic Acids Research,
27 : 1359–1364
Ner, S. S. 1992. HMGs everywhere. A new and diverse
class of DNA-binding proteins is identified
by a common sequence motif.Curr Biol
2:208–210
Uchikawa, M., Kamachi, Y., and Kondoh, H. 1999. Two
distinct subgroups of Group B Sox genes for
transcriptional activators and repressors:
Their expression during embryonic
organogenesis of the chicken. Mech. Dev. 84
:103–120
USA Government. 2010. What is SOX gene family?
[online. Tersedia: http://ghr.nlm.nih.gov/
gene/SOX ]. Oktober. 2013
Wang et al 2009. Isolation and Sequencing of The HMG
Domain of Ten SOX Gene from Odorrana
Schmackeri. Zoologia. 26 : 109–117
Soullier, S., Jay, P., Vanacker, J-M., Berta, P., Laudet,
V. 1999. Diversification of the HMG and
SOX family member during evolution.
J.Mol.Evol. 48: 517-527
Woelk, C.H., Frost, S.D.W., Richman, D.D., Higley,
P.E., Kosakovsky Pond, S.L. 2007. Evolution
of the interferon alpha gene family in
eutherian mammals. Gene 397: 38–50