review jurnal imunostimulan alami pada udang vaname
DESCRIPTION
imunostimulan alamiTRANSCRIPT

BAB 1
PENDAHULUAN
1.1. Latar Belakang
Udang merupakan salah satu komoditas unggulan
revitalisasi perikanan di Indonesia. Kementrian
Kelautan dan Perikanan sendiri telah
menargetkan peningkatan produksi sebesar 74,75% yaitu dari
400 ribu ton menjadi 699 ribu ton pada tahun 2009 sampai
2014, yang terdiri atas udang vaname Litopenaeus
vannamei dan udang windu Penaeus monodon (KKP, 2010).
Keberhasilan produksi tersebut sangat
didukung oleh keberhasilan dalam budidaya. Banyak
kendala yang harus dihadapi dalam
berbudidaya udang, salah satunya adalah adanya serangan
penyakit.
Penyakit merupakan salah satu faktor pembatas dalam
budidaya udang vannamei (Litopennaeus vannamei). Tingginya
tingkat mortalitas udang budidaya diduga disebabkan oleh
infeksi virus maupun bakteri patogen. Nitimulyo et al. (2005)
menyatakan bahwa bakteri patogen yang umum menyerang
dalam budidaya perikanan adalah Vibrio alginolyticus, V.flufialis,
V. vulfinicus, dan V. ordalii. Epidemik yang banyak menyerang
budidaya udang adalah White Spot Syndrome Virus (WSSV),
Taura Syndrome Virus (TSV) dan Yellow Head Virus (YHV) (Smith
et al., 2003).
Salah satu upaya dalam penanggulangan dan
pencegahan penyakit udang adalah melalui peningkatan sistem
pertahanan tubuh udang, yaitu dengan menggunakan
imunostimulan, vitamin dan hormon (Johny et al., 2005). Udang
1

mempunyai daya tahan alami yang bersifat non spesifik
terhadap organisme patogen berupa pertahanan fisik (mekanik),
kimia, seluler dan humoral. Daya tahan alami ini dipengaruhi
oleh faktor genetik dan lingkungan, sehingga terdapat tingkatan
yang berbeda-beda. tergantung strain, lingkungan pemeliharaan,
spesies maupun famili (Bellanti, 1989).
Rumput laut merupakan alga multiselular yang
mengandung substansi yang aktif secara imunologi.
Pemanfaatan rumput laut selama ini masih terbatas pada produk
karagenan dan agar. Potensi rumput laut di bidang pengendalian
penyakit masih belum banyak di eskplorasi dan di eksploitasi.
Beberapa penelitian menunjukkan bahwa rumput laut
mempunyai prospek yang masih terbuka bagi
pengembangannya dalam bidang pengendalian penyakit. Ekstrak
rumput laut telah diketahui mempunyai aktivitas sebagai
antitumor, meningkatkan aktivitas kemotaksis macrophage,
menstimulasi aktivitas sekresi radikal oksigen dan fagositosis
pada peritonial and splenic murine macrophage (Castro et al.,
2004). Metabolit sekunder dari Halimeda macroloba memiliki
senyawa bioaktif anti jamur (Widiastuti, 2003). Rumput laut Ulva
sp., Dendrilla sp.,Spirulina sp., Enteromorpha sp., Dictyota sp.,
dan Porphira sp. telah terbukti mampu meningkatkan aktifitas
imunostimulan udang (Castro et al., 2004; Selvin et al., 2004).
1.2. Tujuan
Tujuan dari penulisan makalah mengenai penggunaan
teknologi bioaktif alami dalam meningkatkan imunitas budidaya
udang vanamei adalah untuk memberikan informasi kepada
civitas akademika dan masyarakat mengenai penggunaan
teknologi bioaktif alami dalam meningkatkan imunitas budidaya
2

udang vaname yang tepat guna dan dapat dikembangkan oleh
masyarakat.
BAB 2
METODE
2.1. Metode Penulisan
Makalah ini ditulis dengan menggunakan metode
penulisan deskriptif, dimana menurut Whitney dalam Nazir
(2003) metode deskriptif merupakan pencarian fakta dengan
interpretasi yang tepat. Tujuan dari metode ini adalah untuk
membuat deskripsi, gambaran atau lukisan secara sistematis,
faktual dan akurat mengenai fakta-fakta, Sifat-sifat serta
hubungan antar fenomena yang diselidiki. Penjelasan dari suatu
keadaan dan kejadian akan dapat semakin memperjelas obyek
yang diamati. Dalam penyusunan makalah ini akan lebih
ditekankan pada review beberapa jurnal yang saling terkait, yang
mana data dari jurnal tersebut dapat digunakan untuk studi
literatur dalam penyusunan penggunaan teknologi bioaktif alami
dalam meningkatkan imunitas budidaya udang vannamei.
2.2. Materi dan Metode
Obyek yang digunakan dalam penelitian adalah hewan uji
udang vannamei dan jenis imunostimulan alami yang digunakan
adalah rumput laut dan ganggang. Dalam hal ini, perlu diketahui
bahwa ada beberapa jenis rumput laut dan ganggang yang
3

digunakan dalam penelitian dalam rangka meningkatkan
imunostimulan udang vannamei diantaranya Kappaphycus
alvarezii, Dictyocta sp, Padina sp, Glacilaria sp, Sargassum sp
dan Spirulina sp, dalam makalah ini akan ditekankan pada
rumput laut dan ganggang tersebut yang digunakan sebagai
imunostimulan alami dalam meningkatkan imunitas udang
vannamei.
Pokok bahasan yang akan penulis coba paparkan adalah
mengenai Uji efektivitas aplikasi rumput laut laut dan ganggang
sebagai imunostimulan sistem pertahanan tubuh non spesifik
udang L. vannamei dapat dilakukan dengan pengamatan
terhadap sistem kekebalan tubuh nonspesifik berdasarkan
gambaran hematologinya, yaitu dengan menghitung jumlah
hemosit dan aktivitas fagositosis.
4

BAB 3
TINJAUAN PUSTAKA
3.1. Biologi Udang Vannamei
3.1.1 Taksonomi
Menurut Haliman dan Adijaya (2005), Udang vaname
digolongkan dalam genus Penaeid pada filum Arthropoda. Ada
ribuan spesies di filum ini. Namun, yang mendominasi perairan
berasal dari subfilum Crustacea. Ciri-ciri subfilum crustacea yaitu
memiliki 3 pasang kaki berjalan yang berfungsi untuk mencapit,
terutama dari ordo Decapoda, seperti Litopenaeus chinenses, L.
indicus, L. japonicus, L.monodon, L.stylirostris, dan L.vanname.
Berikut tata nama udang vaname menurut ilmu taksonomi :
Kingdom : Animalia
Subkingdom : Metazoa
Filum : Artrhopoda
Subfilum : Crustacea
Kelas : Malascostraca
Subkelas : Eumalacostraca
5

Superordo : Eucarida
Ordo : Decapoda
Subordo : Dendrobrachiata
Famili : Penaeidae
Genus : Litopenaeus
Spesies : Litopenaeus vannamei
3.1.2 Morfologi
Menurut Haliman dan Adijaya (2005), tubuh udang
vaname dibentuk oleh dua cabang (biramous), yaitu exopodite
dan endopodite. Vaname memiliki tubuh berbuku-buku dan
aktifitas berganti kulit luar atau eksoskeleton secara periodik
(moulting).
Farchan (2006) menerangkan bahwa, warna tubuh udang
vaname secara keseluruhan putih agak mengkilap dengan titik-
titik warna hitam yang menyebar di sepanjang tubuhnya. Bagian
tubuh udang vaname dibagi menjadi 2 bagian, yang terdiri dari
kepala dan dada (cephalothorax) serta perut (abdomen).
1. Kepala (Thorax)
Cephalotorax disusun oleh kulit yang keras dan tebal
dengan kandungan utamanya chitin yang disebut carapace.
Bagian ujungnya terdapat antena sebanyak dua buah dan
rostrum yang bergerigi. Belakang rostrum terdapat sepasang
mata yang bertangkai berada di kanan dan kiri rostrum. Pada
bagian badan kepala bawah terdapat kaki jalan (pereopoda)
sebanyak 5 pasang, 2 pasang maxillae yang sudah mengalami
modifikasi dan berfungsi sebagai organ untuk makan (Farchan,
2006).
6

2. Perut (Abdomen)
Risaldi (2013) menerangkan bahwa, Abdomen terdiri dari
dari 6 ruas. Pada bagian abdomen terdapat 5 pasang kaki renang
dan sepasang uropods (mirip ekor) yang membentuk kipas
bersama-sama telson. Vaname memiliki tubuh yanng berbuku-
buku dan aktivitas berganti kulit luar atau eksoskelaton secara
priodik (moulting).
Untuk lebih jelas tentang morfologi udang vaname dapat
dilihat pada Gambar 1.
Gambar 1. Morfologi Udang vaname (Risaldi, 2013)
3.1.3 Habitat dan Siklus Hidup
Habitat udang berbeda-beda tergantung dari jenis dan
persyaratan hidup dari setiap fase dalam daur hidupnya. Pada
umumnya udang bersifat bentis dan hidup pada permukaan
dasar laut. Habitat yang disukai oleh udang adalah dasar laut
yang lembut (soft) yang biasanya campuran lumpur dan pasir.
Secara alami udang vaname termasuk jenis katadromus, yaitu
udang dewasa hidup di laut terbuka dan udang muda migrasi ke
arah pantai. Di habitat aslinya, udang matang gonad (matur),
7

kawin (mating) dan bertelur (spawning) berada pada perairan
dengan kedalaman sekitar 70 meter di Amerika Selatan, Tengah
dan Utara, dengan suhu 26 - 28°C dan salinitas sekitar 35 gr/l.
Telur menetas dan larva berkembang di laut dalam sebagai
tempat berkembangnya zooplankton. Post larva udang vaname
bergerak mendekati pantai dan menetap di dasar estuari
/muara. Di estuari, tersedia nutrien, air laut dengan salinitas dan
suhu yang bervariasi dari pada di laut terbuka. Setelah beberapa
bulan di estuari, udang muda kembali ke lingkungan laut
menjauhi pantai, dimana aktivitas matur, mating dan spawning
terjadi (Wahyudi, 2010).
Menurut Haliman dan Adijaya (2005), udang vaname
bersifat nokturnal, yaitu melakukan aktifitas pada malam hari.
Siklus hidup udang vaname sebelum ditebar di tambak yaitu :
a. Stadia Naupli
Pada stadia ini larva berukuran 0,32-0,58mm. Sistem
pencernaannya belum sempurna dan masih memiliki cadangan
makanan berupa kuning telur, sehingga pada stadia ini benih
udang vaname belum membutuhkan makanan dari luar.
b. Stadia Zoea
Stadia zoea terjadi setelah naupli ditebar pada bak
pemeliharaan sekitar 15-24 jam. Larva sudah berukuran 1,05-
3,30 mm. Pada stadia ini, benih udang mengalami moulting
sebanyak tiga kali, yaitu stadia zoea1, zoea2, zoea3. Lama waktu
proses pergantian kulit sebelum memasuki stadia berikutnya
(mysis) sekitar 4-5 hari. Pada stadia ini, larva sudah dapat diberi
pakan alami seperti artemia.
c. Stadia Mysis
Pada stadia ini, benih sudah menyerupai bentuk udang
yang dicirikan dengan sudah terlihat ekor kipas (urupods) dan
8

ekor (telson). Benih pada stadia ini sudah mampu memakan
fitoplankton dan zooplankton. Ukuran benih berkisar antara 3,50-
4,80 mm. Stadia ini memiliki 3 substadia, yaitu mysis1, mysis2,
mysis3 yang berlangsung selama 3-4 hari, sebelum memasuki
stadia post larva (PL).
d. Stadia post larva (PL)
Pada stadia ini, benih udang vaname sudah tampak
seperti udang dewasa. Hitungan stadia yang digunakan sudah
berdasarkan hari. Misalnya, PL 1 berarti postlarva berumur 1hari.
Pada stadia ini udang sudah mulai aktif bergerak lurus ke depan.
3.1.4 Kebiasaan Makan
Menurut Haliman dan Adijaya (2005), udang termasuk
dalam golongan omnivora atau bersifat pemakan segala.
Beberapa sumber pakan udang antara lain udang kecil (rebon),
fitoplankton, cocepoda, polyhaeta, larva kerang dan lumut.
Udang vaname merupakan hewan nocturnal sehingga
sepanjang hari hewan ini tinggal pada substrat dan tidak mencari
makan. Kegiatan makan dilakukan malam hari atau ketika
suasana redup. Kandungan protein yang dibutuhkan dalam
pakan lebih rendah dibandingkan dengan udang windu yaitu
35%, sedangkan untuk udang windu, pakan paling tidak harus
memiliki kandungan protein 45% (Wyban dan Sweeney, 1991).
3.1.5 Kebiasaan dan Tingkah Laku
Menurut Edhy, dkk., (2010), menyatakan bahwa udang
vaname memiliki sifat yang berbeda dengan udang windu. Kalau
udang windu cenderung berada di dasar perairan tambak,
9

sedangkan vaname berada di kolom air dan sangat aktif
berenang kesana kemari.
Berikut ini adalah sifat-sifat udang vaname :
Nocturnal : aktif pada malam hari
Diurnal : aktif pada siang hari
Omnivora : memakan segala jenis makanan
Forager : sangat aktif kesana kemari mencari makanan
Detritivora: makan detritus, yaitu bahan organik yang
dikerubuti oleh bakteri
3.1.6 Respon Imunitas
Respon imunitas dibentuk oleh jaringan limfoid. Pada
udang, jaringan limfoid menyatu dengan jaringan mieloid,
sehingga dikenal sebagai jaringan limfomieloid (Corbel 1975;
Itami 1994). Produk jaringan limfomieloid adalah sel-sel darah
dan respon imunitas baik seluler maupun humoral.
Pada udang, organ limfoidnya disebut sebagai organ oka,
yang mirip dengan sel dentritik retikulum pada folikel mamalia
(Itami 1994). Organ oka ini terdiri dari 2 lobus, terletak di dorso-
anterior hepatopankreas dan ventro-lateral lambung anterior dan
posterior; secara histologis, anastomosa tubul organ limfoid
mengandung massa basofilik (Bell dan Lightner 1988).
Respon humoral pada udang dimungkinkan oleh adanya
multivalen sugar binding agglutinin, disebut sebagai lektin atau
hemagglutinin dan monovalen sugar binding residue, disebut
beta glukan binding protein (BGBP). Selain itu, monomerik
glikoprotein merupakan faktor humoral yang berperan dalam
respon humoral. Molekul ini dengan berat molekul 76 kDA dan
titik isoelektriknya sebesar 7,2 berperan sebagai faktor pelekat
sel hemosit pada permukaan benda asing dan berkaitan dengan
sistem proPO, enkapsulasi. Secara in vitro sistem memacu proses
10

degranulasi dengan menghambat sintesis protein dan aggregasi
sel hemosit.
3.2. Imunostimulasi
Mori (1990) mengemukakan, bahwa respon imunitas
pada hewan merupakan upaya proteksi terhadap infeksi maupun
preservasi fisiologik homeostasi. Respon imunitas hewan akuatik
terdiri dari respon non spesifik dan spesifik baik pada ikan
(Corbel 1975) maupun pada udang (Itami 1994; Bechère 2000).
Karenanya, memori, spesifitas dan pengenalan zat asing
merupakan dasar mekanisme respon imunitas baik pada ikan
maupun udang.
3.2.1. Vaksinasi
Vaksinasi merupakan suatu upaya untuk menimbulkan
ketahanan tubuh yang bersifat spesifik melalui pemberian
vaksin. Secara umum aktivitas ini dikenal sebagai imunisasi aktif
dan pasif. Imunisasi pasif diperoleh dengan pemberian serum
kebal maupun dengan cara diturunkan oleh induk ikan yang
dikenal sebagai imunitas maternal; sedangkan imunisasi aktif
dilakukan melalui tindak vaksinasi. Induk-induk ikan yang
divaksini dapat menurunkan respon imunitas tersebut pada
turunannya. Ellis (1988) telah menguraikan tentang vaksinasi
terutama untuk ikan.
Tujuan spesifik vaksinasi adalah untuk memperoleh
ketahanan terhadap suatu infeksi tertentu, sehingga diperoleh
sintasan hidup yang tinggi akibat proteksi imunologik tersebut.
Secara umum, manfaat vaksinasi antara lain dalam hal:
peningkatan daya tahan ikan, pencegahan efek samping
kemoterapeutika, proteksi terhadap serangan penyakit infeksi
tertentu, keamanan lingkungan budidaya dari pencemaran
11

bahan kemoterapeutik dan keamanan konsumen dari residu
antibiotik.
Secara umum terdapat 2 jenis vaksin yakni vaksin
konvensional dan vaksin moderen. Penjenisan ini semata-mata
didasarkan atas teknologi produksi vaksin yang digunakan.
Produk vaksin dengan teknologi tinggi (hi-tech) dikenal sebagai
vaksin moderen; sedangkan vaksin konvensional diproduksi
dengan teknologi sederhana. Vaksin konvensional dibedakan
atas vaksin mati dan vaksin hidup. Vaksin mati berasal dari
patogen yang dimatikan, ekstrak atau bagian-bagian tertentu
dari patogen; sedang vaksin hidup berasal dari patogen yang
dilemahkan atau diatenuasi. Vaksin yang termasuk kelompok
vaksin moderen atau vaksin biotek adalah vaksin rekombinan,
vaksin monoklonal, protein engineering vaccine dan genetic
attenuation vaccine.
Vaksinasi yang merupakan tindakan memasukkan antigen
ke dalam tubuh akan memacu terbentuknya ketahanan spesifik.
Proses pembentukan respon ini dipengaruhi oleh faktor kualitas
vaksin, ikan dan lingkungan media budidaya. Kualitas vaksin
dipengaruhi oleh keasingan struktur molekuler vaksin, mudah
dikenali oleh limfosit dan kekuatannya berikatan dengan
antibodi. Faktor ikan meliputi antara lain, umur, jenis dan kondisi
fisiologis. Salah satu faktor lingkungan budidaya yang sangat
berpengaruh terhadap vaksinasi adalah suhu. Suhu media
budidaya harus optimal bagi proses pembentukan respon
imunitas spesifik. Respon spesifik yang terbentuk yakni ini
respon yang sangat bergantung kepada suhu (temperature
dependent). Karena itu, suhu media budidaya harus diatur
sedemikian rupa berkisar 20-25 C, agar respon spesifik dapat
terbentuk optimum dalam waktu 1-2 minggu.
12

3.2.2. Imunostimulasi dengan Imunostimulan
Imunostimulan merupakan senyawa kimia, obat atau
bahan lainnya yang mampu meningkatkan mekanisme respon
imunitas ikan (Anderson 1992), baik seluler maupun humoral
(Alifuddin 1999). Galleotti (1998) dan Anderson (1992) telah
mengungkap jenis, berbagai aspek dan aplikasi imunostimulan
berkaitan dengan budidaya perikanan. Lipopolisakarida (LPS)
merupakan salah satu imunostimulan yang digunakan untuk
stimulasi sel B. Kajita et al. (1990) telah mengevaluasi efek
levamisole terhadap peningkatan aktivitas fagositik ikan rainbow
trout (Onchorhynchus mykiss). Anderson & Rumsey (1995)
mengemukakan, bahwa Candida utilis dan Saccharomyces
cerevisiae dapat meningkatkan produksi radikal oksidatif,
aktivitas fagositik, produksi mieloperoksidase dan imunoglobulin
plasma ikan rainbow trout.
Berbeda dengan vaksin, imunostimulan tidak direspon
ikan dengan mensintesis antibodi, melainkan peningkatan
aktivitas dan reaktivitas sel pertahanan seluler ataupun humoral.
Secara in vitro peningkatan respon seluler ditujukkan oleh
aktivitas fagositik yang diukur melalui uji nitro blue tetrazolium
(NBT) (Anderson dan Siwicki 1993). Peningkatan ini didasarkan
atas kemampuan imunostimulan menginduksi berlangsungnya
transformasi limfoblastik yang ditunjukkan dengan memakai
isotop tritium (H3) (Alifuddin 1989). Aktivitas fagositik ini
merupakan manifestasi peningkatan respon seluler dan pada
akhirnya akan meningkatkan respon humoral. Imunotimulan
yang sering dipakai untuk imunostimulasi adalah LPS
(lipopolisakarida), dan β1,3 glukan yang diperoleh dari
Saccharaomyces cerevisiae, dan Levamisol. Beberapa vitamin
seperti vitamin A, B dan vitamin C juga dapat digunakan sebagai
13

imunostimulan (Sohne et al. 2000; Galeotti 1998). Seperti halnya
dengan vaksin, imunostimulan dapat diberikan melalui injeksi,
bersama pakan (per oral) dan perendaman (Anderson 1992).
Dosis imunostimulan yang digunakan sebesar 100-200 ppm.
Imunostimulan ini dapat diberikan secara terus menerus selama
1 minggu kepada larva ikan ketika masih dalam hapa
pendederan; kemudian dihentikan pemberiannya, diberikan
kembali pada minggu ke 3 selama satu minggu. Karena itu, pada
tahap awal, imunostimulan diberikan melalui perendaman, dan
pada pemberian selanjutnya dapat diberikan bersama pakan.
Pemilihan cara aplikasi imunostimulan didasarkan atas
kepraktisan dan efisiensi dalam kegiatan budidaya. Mengingat
keragaman patogen yang ada dalam media budidaya ikan,
imunostimulan merupakan alternatif upaya pengendalian
penyakit infeksi yang harus dilakukan bersama dengan
vakinansi. Pemanfaatannya dalam kegiatan budidaya dapat
mengoptimalkan produksi budidaya melalui peningkatan
ketahanan tubuh ikan atau udang windu terhadap penyakit
infeksi (Pujiharto 1998; Alifuddin 1999; Bagni et al. 2000; Sohne
et al. 2000).
3.3. Imunostimulan Alami
Imunostimulasi biasa dilakukan dengan pemberian
komponen mikrobia seperti β-glukan dan lipopolisakarida (LPS)
atau sel bakteri yang telah dimatikan (Smith et al., 2003).
Kelemahan dari imunostimulan ini adalah harganya relatif mahal,
sehingga diperlukan usaha pencarian sumber alternatif
imunostimulan yang murah dan mudah penanganannya, salah
satunya adalah dari rumput laut dan ganggang.
3.3.1 Rumput Laut
14

Penggunaan vaksin dan kemoterapi telah dilaporkan tidak
efektif untuk penyakit ini (OIE, 2009). Rumput laut merupakan
salah satu bahan yang dapat digunakan sebagai imunostimulan
karena merupakan sumber senyawa bioaktif, yang telah
terdeteksi dalam alga hijau, alga coklat dan alga merah yang
memproduksi berbagai karekteristik metabolit sekunder dengan
spektrum aktifitas yang luas. Dinding sel dari alga laut kaya akan
polisakarida sulfat (SPs) seperti karagenan yang terkandung
dalam alga merah, dan memiliki banyak senyawa bioaktif
menguntungkan sebagai anti koagulan, antiviral, antioksidan,
antikanker serta aktifasi modulasi imun (Wijesekara et al, 2011).
Rumput laut merupakan alga multiselular yang
mengandung substansi yang aktif secara imunologi.
Pemanfaatan rumput laut selama ini masih terbatas pada produk
karagenan dan agar. Potensi rumput laut di bidang pengendalian
penyakit masih belum banyak di eskplorasi dan di eksploitasi.
Beberapa penelitian menunjukkan bahwa rumput laut
mempunyai prospek yang masih terbuka bagi
pengembangannya dalam bidang pengendalian penyakit. Ekstrak
rumput laut telah diketahui mempunyai aktivitas sebagai
antitumor, meningkatkan aktivitas kemotaksis macrophage,
menstimulasi aktivitas sekresi radikal oksigen dan fagositosis
pada peritonial and splenic murine macrophage (Castro et al.,
2004). Metabolit sekunder dari Halimeda macroloba memiliki
senyawa bioaktif anti jamur (Widiastuti, 2003). Rumput laut Ulva
sp., Dendrilla sp.,Spirulina sp., Enteromorpha sp., Dictyota sp.,
dan Porphira sp. telah terbukti mampu meningkatkan aktifitas
imunostimulan udang (Castro et al., 2004; Selvin et al., 2004).
3.3.2 Spirulina sp
15

Spirulina sp. mengandung protein 60% yang terdiri dari
12 asam amino esensial, 10 vitamin, dan juga sifat terapi seperti
pigmen fikosianin yang bersifat antioksidan dan anti-inflamatori,
polisakarida yang memiliki efek antitumor dan antiviral, dan γ-
asam linoleat (GLA) yang berfungsi dalam penurun kolesterol
(Desmorieux and Decaen, 2005). Menurut Boajiang (1994),
polisakarida Spirulina sp. dapat memperbaiki fungsi imunitas
seluler non-spesifik dan fungsi humoral spesifik, termasuk pula
hemosit dan sel-sel fagositosis.
3.4. Hemosit
Hemosit merupakan faktor yang sangat penting dalam
sistem pertahanan seluler yang bersifat non spesifik.
Kemampuan hemosit dalam aktivitas fagositosis yang dapat
meningkat pada kejadian infeksi, menunjukkan pertahanan
tubuh yang bersifat seluler.
Hemosit berperan dalam proses fagositosis, enkapsulasi,
degranulasi, dan agregasi nodular terhadap patogen maupun
partikel asing serta produksi dan pelepasan prophenoloxidase
(proPO) dalam sistem imun krustasea (Sahoo et al., 2008).
Jumlah total hemosit pada krustasea sangat penting dalam
menjaga resistensi terhadap patogen. Apabila kondisi
penurunan total hemosit terjadi, maka hal tersebut dapat
mengakibatkan infeksi akut yang mematikan (Rodriguez & Le
Moullac, 2000).
3.5. Aktifitas Fagositosis
Meningkatnya ketahanan tubuh udang dapat diketahui
dari meningkatnya aktivitas fagositosis sel-sel hemosit.
Fagositosis merupakan mekanisme pertahanan non spesifik yang
16

secara umum dapat melindungi adanya serangan pathogen
(Fontaine and Lightner, 1974).
Aktifitas fagositosis merupakan salah satu cara yang
sangat penting dalam mengendalikan dan menghancurkan
partikel asing. Proses pertahanan melalui fagositosis ini dibagi
menjadi beberapa proses yaitu : kemotaksis, recognition, dan
internalization (Bachere, 1995).
17

BAB 4
HASIL DAN PEMBAHASAN
4.1 Spirulina sp Sebagai Imunostimulan
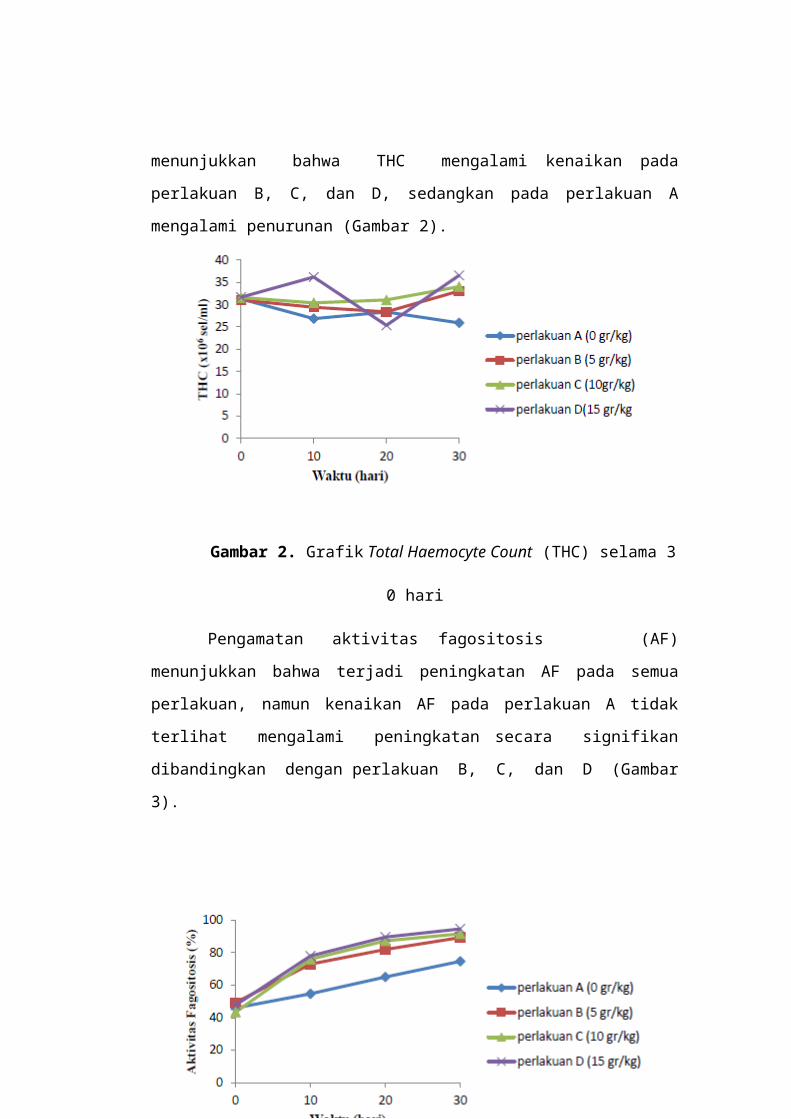
Berdasarkan hasil pengamatan terhadap jumlah
total hemosit (THC) diperoleh bahwa, THC pada hari ke-10 pada
perlakuan A, B, dan C mengalami penurunan dibandingkan pada
hari ke-0. Berbeda dengan perlakuan A, B, dan C yang
mengalami penurunan, pada perlakuan D terjadi kenaikan THC
pada hari ke-10. Selanjutnya pada hari ke-20, THC mengalami
kenaikan pada perlakuan A, B, dan C, sedangkan pada perlakuan
D mengalami penurunan. Pengamatan pada hari ke-30
menunjukkan bahwa THC mengalami kenaikan pada perlakuan
B, C, dan D, sedangkan pada perlakuan A mengalami penurunan
(Gambar 2).
Gambar 2. Grafik Total Haemocyte Count (THC) selama 3
0 hari
18

Pengamatan aktivitas fagositosis (AF) menunjukkan
bahwa terjadi peningkatan AF pada semua perlakuan, namun
kenaikan AF pada perlakuan A tidak terlihat mengalami
peningkatan secara signifikan dibandingkan dengan perlakuan
B, C, dan D (Gambar 3).
Gambar 3. Grafik aktivitas fagositosis (AF) selama 30
hari
Hasil pengamatan AF menunjukkan bahwa
penambahan Spirulina sp. Pada pakan berpengaruh nyata
(P<0,05) terhadap AF pada hari ke-10, sedangkan penambahan
Spirulina sp. pada pakan berpengaruh sangat nyata
(P<0,01) terhadap AF pada hari ke-20 dan 30.
Hasilpengamatan aktivitas fagositosis juga
menunjukkan bahwa, aplikasi penambahan Spirulina sp. Pada
perlakuan C (10 gr/kg) merupakan dosis terbaik dibandingkan
dengan perlakuan D (15 gr/kg), B (5 gr/kg), dan A (tanpa
pemberian Spirulina sp.)
4.2 Kappa-Karagenan Sebagai Imunostimulan
19

Pemberian k-karagenan pada pakan udang vaname dalam
penelitian ini, dapat meningkatkan parameter imun nonspesifik
yang tercermin dari meningkatnya Jangka Waktu Total hemosit,
aktivitas fagositosis dan phenoloxidase selama Waktu
pengamatan (Gambar 1). Hasil analisis Ragam dan uji lanjut
Duncan menunjukkan total hemosit berbeda Nyata (p <0,05)
antar masing-masing perlakuan Dan juga dengan Kontrol.
Pemberian k-karagenan 15 g/kg pakan (C) memperlihatkan nilai
total hemosit lebih Tinggi dibandingkan perlakuan sebelumnya,
yaitu dengan kisaran nilai (10,23±0,23-12,00±0,72)x106 sel/mL
sedangkan Kisaran nilai perlakuan lainnya hanya sebesar
(5,47±0,15-9,57±0,15)×106 sel/mL, Pembongkaran pãda
ditunjukkan Gambar 4.
Meningkatnya sistem imun pada udang dapat dilihat dari
meningkatnya jumlah hemosit. Hemosit berperan dalam proses
fagositosis, enkapulasi, degranulasi dan agregasi nodular
terhadap patogen maupun partikel asing serta produksi dan
pelepasan prophenoloxidase (proPO) dalam system imun
krustasea (Sahoo et al., 2008). Jumlah total hemosit pada
krustasea sangat penting dalam menjaga resistensi terhadap
patogen. Apabila kondisi penurunan total hemosit
terjadi, maka hal tersebut dapat mengakibatkan
infeksi akut yang mematikan (Rodriguez & Le Moulla ,2000).
Meningkatnya total hemosit akan meningkatkan kemampuan
untuk memfagositosis. Meningkatnya total hemosit juga
meningkatkan sel granular yang dapat merangsang aktivasi
ProPO untuk menghasilkan aktifitas phenoloxidase, sehingga
mampu bertahan terhadap serangan
patogen (Yudiana, 2009). Dalam penelitian ini, aktivitas
fagositosis dan phenoloxidase berbeda nyata (p<0,05)
20

antara perlakuan dan control. Nilai tertinggi aktifitas fagositosis
dan phenoloxidase terjadi pada pemberian 15 g/kg pakan
(C), masing-masing sebesar 34,67±0,58% (minggu 4) dan
0,511±0,1 (minggu 3), diperlihatkan pada Gambar 4.
Gambar 4. Total hemosit (A), aktifitas fagositosis (B), dan Phenoloxidase (C) udang vaname Litopenaeus vannamei yang diberi k-karagenan 0 (K), 5 (A), 10 (B), dan 15 (C) g/kg pakan selama empat minggu pemeliharaan. Huruf yang berbeda di atas balok pada diagram batang dengan waktu pengamatan yang sama
21

menunjukkan adanya pengaruh yang berbeda nyata akibat adanya perlakuan (p<0,05).
Mekanisme k-karagenan dalam meningkatkan
sistem imun dalam tubuh udang masih terus dipelajari.
Penelitian Yeh & Chen (2008) menyatakan bahwa aktivitas
fagositosis dan respiratory burst meningkat pada L.
vannamei yang diberi perlakuan karagenan, hal
tersebut mengindikasikan adanya peranan reseptor
karagenan pada makrofag dan hemosit. Pada udang
karang P.leniusculus, β-glucan dan β-glucan binding
protein (βGBP) komplek dapat berikatan dengan
permukaan hemosit-granular melalui motif arginyl-glysyl-
aspartic acid (RGD) yang menunjukkan ikatan
integrin-like protein dan memastikan degranulasi
hemosit sehingga dapat mengaktifasi sistem imun. Yeh
& Chen (2008) menduga ada kesamaan mekanisme
karagenan dengan β-glucan dan βGBP komplek dalam
berikatan dengan permukaan hemosit-granular melalui
motif RGD.
4.3 Dictyota sp, Glacillaria sp, Padina sp dan
Sargassum sp Sebagai Imunostimulan
Hasil penelitian menunjukkan bahwa jumlah hemosit dan
aktivitas fagositosis bervariasi, tergantung pada jenis rumput
laut yang digunakan. Pemberian ekstrak rumput laut cenderung
meningkatkan jumlah total hemosit, terutama pada hari ke - 8
dan hari ke - 12. Jumlah total hemosit tertinggi dicapai oleh
Sargassum sp. pada hari ke-12 (1,127 x 107 sel/L ± 0,260)
(Tabel 1). Pada hari ke-8 pemberian ekstrak Gracilaria sp.
menunjukkan nilai tertinggi dibandingkan rumput laut jenis lain,
sedangkan pada hari ke-12, pemberian ekstrak Dictyota sp.,
22
Padina sp. dan Sargassum sp. memberikan hasil yang lebih tinggi
dibanding kontrol. Hal ini menunjukkan bahwa pemberian
ekstrak rumput laut mampu menstimulasi peningkatkan jumlah
total hemosit udang L. vannamei. Menurut Johansson et al.
(2000), hematosit udang memegang peranan penting dalam
respon imun diantaranya melalui recognition, phagocytosis,
melanization, cytotoxicity dan komunikasi antar sel.
Tabel 1. Pengaruh Pemberian Ekstrak Rumput Laut Terhadap Jumlah Total Hemosit (THC) Udang L. vannamei
Hasil penelitian menunjukkan bahwa pemberian
ekstrak rumput laut Sargassum sp., Dictyota sp. dan
Padina sp., mampu meningkatkan aktivitas fagositosis
udang L. vannamei terutama pada hari ke-8 (Lihat Tabel 2).
Hal ini disebabkan dalam ketiga jenis rumput laut tersebut
mengandung senyawa polisakarida alginat. Menurut Cheng
et al. (2005), sodium alginat dapat meningkatkan aktifitas
fagositosis udang. Disamping itu, Subagiyo (2009)
menyatakan bahwa ekstrak rumput laut jenis Halimeda sp.
dapat meningkatkan aktifitas fagositosis pada hari ke-12
sebesar 76,78%
Tabel 2. Pengaruh Pemberian Ekstrak Rumput Laut Terhadap Aktivitas Fagositosis Udang L. vannamei
23

Berdasarkan hasil penelitian ini, pemberian pakan
dengan suplementasi ekstrak rumput laut guna
meningkatkan jumlah hemosit udang L. Vanname dapat
diberikan sampai dengan hari ke-12, sedangkan untuk
meningkatkan aktivitas fagositosis diberikan sampai
dengan hari ke-8. Hasil terbaik jumlah hemosit dan
aktivitas fagositosis dicapai oleh Sargassum sp. masing-
masing pada hari ke 12 dan pada hari 8.
4.4 Glacilaria tenuistipitata sebagai imunostimulan
THC L. vannamei yang menerima ekstrak air panas dari G.
tenuistipitata pada 4 dan 6 μg g-1 secara signifikan lebih tinggi
dibandingkan dengan udang menerima saline dan udang kontrol
setelah 1 hari. THC L. vannamei yang menerima ekstrak air
panas dari G. tenuistipitata pada 4 dan 6 μg g-1 secara signifikan
lebih tinggi dibandingkan dengan kontrol udang setelah 2 hari.
Namun, tidak ada perbedaan yang signifikan dalam THC yang
diamati antara udang yang menerima ekstrak air panas dari G.
tenuistipitata pada 4 dan 6 μg g-1, udang yang menerima garam,
dan udang kontrol setelah hari 1-6 perawatan (Gbr. 5).
24

Gambar. 5. Rata-rata (GSE) THC dari Litopenaeus vannamei yang menerima ekstrak air panas dari Gracilaria tenuistipitata pada 6 μg g-1, 4 μg g-1, menerima garam dan udang kontrol. Setiap balok menunjukkan nilai rata-rata dari delapan penentuan dengan standard error. Data pada saat pemaparan yang sama dengan huruf yang berbeda berbeda secara signifikan (p¿ 0,05)
Aktivitas fagositosis secara signifikan lebih tinggi
untuk udang yang menerima ekstrak air panas dari
G.tenuistipitata pada 6 μg g-1 dibandingkan dengan udang
menerima garam dan udang kontrol setelah 1 hari.
Aktivitas fagositosis adalah 56%, 53%, 43% dan 41% untuk
6 μg g-1, 4 μg g-1, garam dan kelompok kontrol, masing-
masing setelah 1 hari. Namun, tidak ada perbedaan yang
signifikan dalam aktivitas fagositosis diamati antara empat
perlakuan setelah hari 2-6 (Gbr. 6).
25

Gambar. 6. Rata-rata (GSE) aktivitas fagositosis dari Litopenaeus vannamei yang menerima ekstrak air panas dari Gracilaria tenuistipitata pada 6 μg g-1, 4 mg g-1, menerima garam dan udang kontrol. Setiap balok menunjukkan nilai rata-rata dari delapan penentuan dengan standard error.
4.5 Sargassum duplicatum sebagai imunostimulan
THC L. vannamei yang direndam dalam ekstrak air panas
dari S. Duplicatum pada 500 mg 1-1 dan 300 mg 1-1 secara
signifikan lebih tinggi dibandingkan dengan kontrol udang
setelah 1, 3 dan 4 jam (Gbr. 7 .
26

Gambar. 7. Rata-rata (GSE) THC (A) dari Litopenaeus vannamei direndam dalam air laut yang mengandung ekstrak air panas dari Sargassum Duplicatum di 500, 300 dan 100 μg 1-1, dan udang kontrol, dan mean (GSE) THC (B) udang yang disuntik dengan ekstrak air panas dari S. Duplicatum pada 20, 10, 6, dan 2 μg g-1, udang disuntik dengan garam, dan udang kontrol. Setiap balok menunjukkan nilai rata-rata dari delapan udang dengan standard error. Data pada saat pemaparan yang sama dengan huruf yang berbeda berbeda secara signifikan (p¿0,05).
THC L. vannamei yang disuntik dengan ekstrak air
panas dari S. Duplicatum pada 20, 10 dan 6 μg g-1 secara
signifikan lebih tinggi dari udang yang disuntik dengan
ekstrak air panas pada 2 μg g-1, udang yang disuntik
dengan garam serta udang kontrol setelah 1 dan 2 hari.
THC L. vannamei yang disuntik dengan dosis 20 μg g-1 dan
10 μg g-1 secara signifikan lebih tinggi dari udang yang
disuntik dengan 6 μg g-1 dan 2 μg g-1, udang yang
disuntikkan dengan garam serta udang kontrol setelah 4
dan 6 hari (Gbr. 1B).
Aktivitas fagositosis secara signifikan lebih tinggi
untuk udang yang menerima ekstrak air panas dari S.
Duplicatum pada 20 mg g-1 dibandingkan dengan udang
yang menerima garam dan udang kontrol setelah 1-4 hari.
Aktivitas fagositosis adalah 35%, 31%, 28%, 21%, 13% dan
13% untuk udang yang menerima air panas ekstrak pada
20, 10, 6, 2 μg g-1, untuk udang yang menerima garam dan
udang kontrol, masing-masing setelah 2 hari. Aktivitas
fagositosis udang yang menerima ekstrak air panas pada
20 μg g-1 dan 10 mg g-1 setelah 6 hari, masih menjaga nilai-
nilai secara signifikan lebih tinggi (26% dan 22%)
27

dibandingkan dengan udang yang menerima ekstrak air
panas di 2 μg g-1, udang yang menerima garam, serta
udang kontrol. Tidak ada perbedaan yang signifikan dalam
aktivitas fagositosis diamati antara udang yang menerima
ekstrak air panas pada 2 μg g-1, udang yang menerima
garam, dan udang kontrol setelah 4 dan 6 hari (Gbr. 8).
Gambar. 8. Rata-rata (GSE) aktivitas fagositosis dari Litopenaeus vannamei disuntik dengan ekstrak air panas dari Sargassum duplicatum pada 20, 10, 6, dan 2 μg g-1, udang disuntik dengan garam, dan kontrol udang. Setiap balok menunjukkan nilai rata-rata dari delapan udang dengan standard error.
BAB 5
KESIMPULAN
Berdasarkan hasil dari tiap penelitian yang telah
dilakukan, maka dapat diambil kesimpulan sebagai berikut:
28

1. Pada penelitian pertama penambahan Spirulina sp. dalam
pakan udang vaname memberikan pengaruh sangat
nyata (P<0,01) terhadap aktivitas fagositosis sebesar
94,66% pada hari ke-30, akan tetapi tidak berpengaruh
nyata terhadap jumlah total hemosit udang vaname
(P>0,05). Perlakuan C (10 gr/kg) merupakan dosis terbaik
bagi peningkatan jumlah total hemosit dan peningkatan
aktivitas fagositosis hemolim udang vaname pada aplikasi
pemberian selama 30 hari.
2. Pemberian k-karagenan sebagai imunostimulan melalui
pakan mampu meningkatkan respons imun,
pertumbuhan dan resistensi udang vaname,
terhadap infeksi IMNV. Pemberian k-karagenan dosis 15
g/kg pakan dengan frekuensi pemberian selama 14
hari secara berulang, dengan interval tujuh hari
memberikan hasil terbaik dengan pertumbuhan bobot
relatif 88,57% dan kelangsungan hidup setelah
diinfeksi dengan IMNV sebesar 90% .
3. Suplementasi ekstrak rumput laut Dictyota sp., Gracilaria
sp., Padina sp. dan Sargassum sp. Pada dosis 10 g/ kg
pakan mampu meningkatkan jumlah total hemosit dan
aktivitas fagositosis udang L. Vannamei
4. L. vannamei yang menerima ekstrak air panas dari G.
tenuistipitata pada 4 dan 6 μg g-1 ditingkatkan kekebalan
dengan THC-nya, meningkatkan aktivitas phenoloxidase,
dan mempercepat pernapasan. L. vannamei yang
menerima air ekstrak G. tenuistipitata dengan dosis 6 μg
g-1 atau kurang peningkatan resistensi terhadap V.
alginolyticus dengan meningkatkan aktivitas dan
pembersihan fagositosis yang efisien. Ekstrak air panas
29

dari G. tenuistipitata dapat digunakan sebagai
imunostimulan untuk L. vannamei.
5. Pemberian ekstrak air panas dari S. Duplicatum melalui
perendaman atau injeksi meningkatkan kemampuan
kekebalan L. vannamei dengan meningkatkan THC,
aktivitas phenoloxidase, percepatan respirasi, dan
perlawanan terhadap V. alginolyticus.
DAFTAR PUSTAKA
30

Alifuddin, M. 2002. Imunostimulasi pada hewan akuatik. Jurnal akuakultur Indonesia.
Febriani, D, Sukenda dan Nuryati, S. 2013. Kappa-karagenan sebagai Imunostimulan untuk pengendalian penyakit infectious myonecrosis (IMN) pada udang vaname Litopenaeus vannamei. Jurnal Akuakultur Indonesia.
Putri, F,M, Sarjito, Suminto. 2013. Pengaruh Penambahan Spirulina sp. dalam Pakan Buatan Terhadap Jumlah Total Hemosit dan Aktivitas Fagositosis Udang Vaname (Litopenaeus vannamei). Fakultas Perikanan dan Ilmu Kelautan, Universitas Diponogoro. Semarang.
Ridlo, A dan Pramesti, R. 2009. Aplikasi Ekstrak Rumput Laut Sebagai Agen Imunostimulasi Sistem Pertahanan Non Spesifik Pada Udang (Litopennaeus Vannamei). Jurusan Ilmu Kelautan, Fakultas Perikanan dan Ilmu Kelautan. Semarang.
Yeh, S, T, Lee, C, S and Chen J, C. 2005. Administration of hot-water extract of brown seaweed Sargassum duplicatum via immersion and injection enhances the immune resistance of white shrimp Litopenaeus vannamei. Department of Aquaculture, College of Life and Resource Sciences, National Taiwan Ocean University, Keelung 202. Taiwan.
Hou, W, Y and Chen J, C. 2004. The immunostimulatory effect of hot-water extract of Gracilaria tenuistipitata on the white shrimp Litopenaeus vannamei and its resistance against Vibrio alginolyticus. Department of Aquaculture, College of Life and Resource Sciences, National Taiwan Ocean University. Taiwan, ROC.
31

DAFTAR ISI
KATA PENGANTAR................................................. ii
DAFTAR ISI............................................................ iii
1. PENDAHULUAN ......................................................... 1
1.1 Latar Belakang ................................................ 11.2 Tujuan ............................................................. 2
2. METODE .................................................................... 3
2.1 Metode penulisan............................................. 32.2 Materi dan metode........................................... 3
3. TINJAUAN PUSTAKA.................................................... 5
3.1 Biologi Udang Vannamei.................................. 5
3.1.1 Taksonomi............................................... 53.1.2 Morfologi................................................. 63.1.3 Habitat dan Siklus Hidup......................... 73.1.4 Kebiasaan Makan.................................... 93.1.5 Kebiasaan dan tingkah laku.................... 93.1.6. Respon Imunitas.................................... 9
3.2 Imunostimulasi................................................. 10
3.2.1 Vaksinasi................................................. 103.2.2 Imunostimulasi dengan Imunostimulan. . 12
3.3 Imunostimulan Alami....................................... 13
3.3.1 Rumput Laut........................................... 143.3.2 Spirulina sp............................................. 14
3.4 Hemosit............................................................ 15
3.5 Aktifitas Fagositosis......................................... 15
4. HASIL DAN PEMBAHASAN ......................................... 17
4.1 Spirulina sp Sebagai Imunostimulan................ 174.2 Kappa-Karagenan Sebagai Imunostimulan...... 184.3 Dictyota sp, Glacillaria sp, Padina sp dan
Sargassum sp Sebagai Imunostimulan............ 214.4 Glacilaria tenuistipitata sebagai imunostimulan
.........................................................................224.5 Sargassum duplicatum sebagai imunostimulan
.........................................................................245. KESIMPULAN ............................................................. 27
DAFTAR PUSTAKA .......................................................... 29
32iii

KATA PENGANTAR
Puji syukur Alhamdulillah, penulis panjatkan Kehadirat Allah SWT
karena atas limpahan Karunia-Nya sehingga penulis dapat
menyelesaikan makalah mengenai “Penggunaan Teknologi
Bioaktif Alami dalam Meningkatkan Imunitas Budidaya
Udang Vanamei“ dalam rangka memenuhi tugas Mata Kuliah
Pengembangan Budidaya Prof. Dr. Ir. Arief Prajitno, MS
Penulis menyadari sepenuhnya bahwa penulisan makalah ini
masih jauh dari kata sempurna dengan segala kekurangan dan
keterbatasan yang dimiliki penulis, sehingga kritik dan saran
yang membangun sangat diharapkan dari semua pihak agar
makalah ini dapat memberikan manfaat yang lebih kepada
civitas akademika yang membutuhkan
Malang, November
2014
Penulis
33

TUGAS TERSTRUKTUR MATA KULIAH PENGEMBANGAN BUDIDAYA
Prof. Dr. Ir. Arief Prajitno, MS
“ Penggunaan Teknologi Bioaktif Alami dalam Meningkatkan Imunitas Budidaya Udang Vanamei“
KELAS B
Oleh :Muhammad Ikhwan I146080100111021
PROGRAM MAGISTER BUDIDAYA PERAIRANFAKULTAS PERIKANAN DAN ILMU KELAUTAN
UNIVERSITAS BRAWIJAYAMALANG
2014
34
ii

35