pola ikatan pembuluh bambu sebagai penduga … · belum diajukan dalam bentuk apapun kepada...
TRANSCRIPT

POLA IKATAN PEMBULUH BAMBU
SEBAGAI PENDUGA PEMANFAATAN BAMBU
NANI NURIYATIN
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2012

PERNYATAAN MENGENAI DISERTASI
DAN SUMBER INFORMASI
Saya menyatakan dengan sebenar-benarnya bahwa segala pernyataan dalam
disertasi saya yang berjudul Pola Ikatan Pembuluh Bambu sebagai Penduga
Pemanfaatan Bambu adalah hasil karya saya sendiri bersama pembimbing dan
belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber
informasi yang berasal dan dikutip dari karya yang diterbitkan maupun tidak
diterbitkan oleh penulis lain disebutkan dalam teks dan dicantumkan dalam Daftar
Pustaka di bagian akhir setiap bab disertasi ini.
Bogor, Januari 2012
Nani Nuriyatin
NRP E061050061

ABSTRACT
NANI NURIYATIN. Vascular bundle pattern as predictor of bamboo utilization.
Under direction of KURNIA SOFYAN, SURJONO SURJOKUSUMO, and
SUMINAR S ACHMADI.
Bamboo is a plant which is widely distributed but can not be optimally
utilized. This can be proved by the 143 species of bamboo in Indonesia, only 32
types among them have distinct purpose. Therefore, to optimize the bamboo
utilization is through evaluating anatomy, mechanical, and chemical properties so
that the results can be useful. The research method was approach by regression
with dummy variables, and description analyze. The result of anatomy study
showed that every bamboo species had specific vascular bundle pattern (type 1-4)
as well as the single patern and its combination. The difference of vascular bundle
pattern did not contributed to the physical and mechanical properties of bamboo
investigated, except for MOR. The difference species of bamboo and vertical
position of samples contributed to the different value of compressive strength
parallel to grain, whereas tension strength was only affected by bamboo species.
The results of chemical study showed that interaction between the species and
the vascular bundle patterns of bamboo influences extractive, ash, lignin, and
starch content except alpha cellulose content. The vascular bundle patterns have
important value for distinguishing species of bamboo and they can also be used
to help direction of the use of bamboo.
Keyword: bamboo properties, vascular bundle, density, fibre .

RINGKASAN
NANI NURIYATIN. Pola ikatan pembuluh bambu sebagai penduga pemanfaatan
bambu. Dibimbing oleh KURNIA SOFYAN, SURJONO SURJOKUSUMO, dan
SUMINAR S ACHMADI.
Bambu adalah tanaman yang tersedia melimpah di Indonesia. Tetapi dalam
pemanfaatannya belum optimal karena penggunaan bambu selama ini masih
mengandalkan cara tradisional secara turun-temurun. Tidak setiap jenis bambu
mempunyai penggunaan yang sama, terkait dengan perbedaan sifat yang dimiliki
bambu. Dengan demikian perlu penelitian dasar ilmiah yang hasilnya dapat
digunakan sebagai dasar acuan penggunaannya.
Beberapa sifat dasar bambu yang diteliti adalah sifat anatomi, fisik,
mekanik, dan sifat kimia. Analisis statistik yang digunakan adalah analisis
regresi dengan peubah boneka untuk aspek anatomi, fisik, dan mekanik.
Sedangkan pengolahan data sifat kimia menggunakan analisis deskripsi dan
keragaman.
Analisis deskripsi terhadap bambu yang diteliti sifat anatominya
menetapkan bahwa seluruh jenis bambu yang diteliti memiliki pola ikatan
pembuluh. Pola-pola tersebut dijumpai dalam bentuk pola tunggal maupun pola
gabungan. Pola tunggal dimiliki oleh bambu Arundinaria hundsii, Arundinaria
javonica, Melocanna baccifera, Cephalostachyum pergracile, Dendrocalamus
strictus dan Dendrocalamus giganteus. Pola gabungan dimiliki oleh bambu
Dendrocalamus asper, Gigantochloa apus dan Gigantochloa atroviolacea. Nilai
kerapatanikatan pembuluh bambu dipengaruhi oleh seluruh faktor yang diujikan,
yaitu jenis bambu, posisi horizontal, posisi vertikal dan pola ikatan pembuluh.
Sementara panjang serabut dipengaruhi oleh jenis bambu dan pola ikatan
pembuluh. Hanya persen serabut yang dipengaruhi oleh satu faktor saja yaitu
posisi horizontal penampang lintang batang.
Nilai keteguhan lentur patah (MOR) bambu-bambu yang diteliti dipengaruhi
oleh jenis bambu dan pola ikatan pembuluh bambu. Pola 3 memiliki nilai MOR
yang lebih tinggi dibandingkan pola 4. Dendrocalamus asper memiliki nilai
MOR yang paling tinggi dibandingkan bambu lain. Nilai kekakuan (MOE) tidak
dipengaruhi oleh faktor yang diujikan. Jenis bambu dan posisi vertikal batang
mempengaruhi nilai keteguhan tekan sejajar serat. Dendrocalamus asper
memiliki nilai keteguhan tekan paling tinggi dibandingkan bambu-bambu lain.
Hanya pada posisi pangkal, keteguhan tekan memiliki nilai tertinggi. Hasil
pengujian keteguhan tarik dipengaruhi oleh jenis bambu. Dendrocalamus asper
cenderung mempunyai nilai keteguhan tarik lebih tinggi dibandingkan jenis lain.
Hasil analisis deskripsi dan analisis keragaman menyatakan bahwa pada
umumnya sifat-sifat kimia bambu dipengaruhi oleh interaksi antara jenis bambu
dan pola yang dimilikinya. Beberapa sifat kimia yang dipengaruhi oleh interaksi
adalah kandungan ekstraktif, kadar abu, kadar lignin, dan kandungan pati. Ciri
yang menonjol yang dimiliki oleh pola 1 adalah mengandung ekstraktif, lignin
dan alfa selulosa yang tinggi tetapi kadar pati rendah. Pola 2 memiliki ciri kadar
ekstraktif dan pati rendah tetapi kadar abu tinggi. Pola 3 mempunyai ciri kadar

alfa selulosa dan kadar pati rendah. Nilai alfa selulosa yang paling rendah
dimiliki oleh bambu dengan pola 4. Sifat-sifat kimia yang dimiliki oleh bambu
membantu dalam mengarahkan penggunaan bambu sebagai bahan serat.
Pola ikatan pembuluh bambu adalah variabel sifat anatomi yang sebaiknya
dipertimbangkan dalam penggunaan bambu. Pola ikatan pembuluh pada bambu
memiliki ciri tertentu yang dapat membantu mengarahkan penggunaan bambu.
Bambu yang memiliki pola ikatan pembuluh 1 atau 2 dapat digunakan untuk
bahan non struktural sedangkan bambu dengan pola ikatan pembuluh 3 atau 4
dapat digunakan sebagai bahan struktural. Pola ikatan pembuluh bambu terkait
dengan sifat kimia yang penting peranannya dalam pendugaan pemanfaatan serat
serta pemanfaatan bambu di lapangan.
Kata kunci: bambu, pola ikatan pembuluh, anatomi, kimia, fisik mekanik bambu,
penggunaan bambu

© Hak cipta milik IPB, tahun 2012
Hak cipta dilindungi undang-undang
1. Dilarang mengutip sebagaian atau seluruh karya tulis ini tanpa
mencantumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan
karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu
masalah
b. Pengutipan tidak merugikan kepentingan yang wajar IPB
2. Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis
dalam bentuk apapun

POLA IKATAN PEMBULUH BAMBU
SEBAGAI PENDUGA PEMANFAATAN BAMBU
NANI NURIYATIN
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar
Doktor pada
Program Studi Ilmu Pengetahuan Kehutanan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2012

Penguji pada Ujian Tertutup:
1. Dr.Ir. Andi Gunawan, MSc
(Pengajar Departemen Arsitektur Landscap, Fakultas Pertanian, IPB)
2. Dr.Ir.Naresworo Nugroho, MS
(Ketua Program Studi IPK)
Penguji pada ujian terbuka:
1. Dr. Ir. Sandra Arifin Aziz, MS
(Pengajar Departemen Agronomi dan Holtikultur)
2. Dr. Krisdianto Sugiyanto, MSc
(Peneliti Hasil Hutan di Lembaga Penelitian Hasil Hutan, Bogor)

Judul : Pola Ikatan Pembuluh Bambu sebagai Penduga
Pemanfaatan Bambu
Nama Mahasiswa : Nani Nuriyatin
NRP : E 061050061
Disetujui
Komisi Pembimbing
Prof. Dr. Ir. Kurnia Sofyan
Ketua
Prof.Dr.Ir. Surjono Surjokusumo, MSF Prof.Dr.Ir. Suminar S Achmadi, MSc
Anggota Anggota
Ketua Program Studi Dekan Pascasarjana IPB
Ilmu Pengetahuan Kehutanan
Dr.Ir.Naresworo Nugroho,MS Dr.Ir.Dahrul Syah,MSc.Agr
Tanggal ujian: 27 Januari 2012 Tanggal lulus:

PRAKATA
Puji dan syukur penulis panjatkan kepada Allash SWT atas segala karunia-
Nya sehingga seluruh rangkaian penelitian/karya ilmiah berhasil diselesaikan.
Selain penelitian di berbagai laboratorium di lingkup Institut Pertanian Bogor,
penelitian juga dilakukan di laboratorium Pusat Penelitian dan Pengembangan
Kehutanan, Kementerian Kehutanan, Bogor.
Terima kasih penulis ucapkan kepada Bapak Prof.Dr.Ir.Kurnia Sofyan,
Bapak Prof.Dr.Ir.Surjono Surjokusumo, MSF dan Ibu Prof. Dr.Ir.Suminar
S.Achmadi,MSc selaku pembimbing yang telah memberikan arahan, bimbingan,
gagasan, dan motivasi selama proses studi doktor. Ucapan terima kasih juga
disampaikan kepada Rektor Unib Bengkulu yang telah memberikan ijin kepada
penulis untuk melanjutkan studi di sekolah Pascasarjana, Institut Pertanian Bogor.
Ucapan terima kasih kepada Dirjen Dikti atas beasiswa pendidikan BPPS yang
sangat membantu kelancaran studi di IPB, Sekolah Pascasarjana IPB, Rektor IPB,
Ketua Program Studi IPK yang telah memberikan kesempatan untuk menimba
ilmu. Penghargaan juga penulis sampaikan kepada penguji luar yaitu Ibu Dr. Ir.
Sandra Arifin Aziz, MS, Bapak Dr. Krisdianto Sugiyanto, MSc, Bapak Dr.Ir.Andi
Gunawan, MSc serta Dr.Ir.Naresworo Nugroho, MS yang telah memberi arahan
dan masukan dalam penulisan disertasi. Ucapan terima kasih yang tulus juga
disampaikan kepada Bapak dan mimih (alm.) serta Ama, teteh Ifa, teteh Angi,
Adam serta semua pihak yang telah membantu kelancaran seluruh pelaksanaan
studi.
Semoga karya ilmiah ini bermanfaat.
Bogor, Januari 2012
Nani Nuriyatin

RIWAYAT HIDUP
Penulis dilahirkan di Ciamis pada tanggal 5 Mei 1963 dari ayah H. Hardja
Sadeli dan ibu Hj.Sri Suweni (alm.) Penulis merupakan anak kelima dari
delapan bersaudara. Pendidikan sarjana ditempuh di jurusan Teknologi Hasil
Hutan, Fakultas Kehutanan Institut Pertanian Bogor, lulus pada tahun 1987. Pada
tahun 1996 penulis diterima di Progam Studi Ilmu Pengetahuan Kehutanan IPB
dan lulus pada tahun 2000. Kesempatan untuk melanjutkan studi ke program
doktor diperoleh pada tahun 2005 dengan beasiswa BPPS-Dikti.
Penulis bekerja sebagai dosen di Universitas Bengkulu sejak tahun 1994.
Bidang pengajaran dan penelitian yang ditekuni adalah struktur dan sifat kayu dan
berlanjut sampai saat ini.
Selama mengikuti program S-3, penulis menjadi anggota MAPEKI
(Masyarakat Peneliti Kayu Indonesia). Karya ilmiah berjudul Kemungkinan
Pemanfaatan Beberapa Jenis Bambu Tertentu, Berdasarkan Pola Penyusunan
Berkas Pembuluh, sebagai Bahan Baku Pulp dan Kertas diterbitkan di Jurnal
Penelitian Hasil Hutan, Vol. 29 No. 4 tahun 2011 halaman 287-300. Karya
tersebut merupakan bagian dari program S-3 penulis.

DAFTAR ISI
Halaman
DAFTAR TABEL …………… ………………………………………. . xii
DAFTAR GAMBAR ………………………………………………… . xiii
DAFTAR LAMPIRAN…………………………………………………. . xvii
I PENDAHULUAN UMUM
Latar Belakang………………………………………………….. 1
Tujuan Penelitian ………………………………………………. 8
Hipotesis ……………………………………………………….. 8
Manfaat Penelitian ……………………………………………… 8
Ruang Lingkup Penelitian ………..………………….………… 8
DAFTAR PUSTAKA………………………………………………. . 9
II ANATOMI PENAMPANG LINTANG BATANG 9 JENIS
BAMBU
Pendahuluan …………………………………………………… 12
Bahan dan Metode ……………………………………………. . 13
Hasil dan Pembahasan .……………………………………….. . 15
Simpulan dan Saran ……….…..………………….…………... . 49
DAFTAR PUSTAKA………………………………………………. . 51
III KORELASI POLA IKATAN PEMBULUH PADA SIFAT FISIK
DAN MEKANIK 3 JENIS BAMBU
Pendahuluan …………………………………..……………… . 53
Bahan dan Metode …………………………………………….. 55
Hasil dan Pembahasan .……………………………………….. . 60
Simpulan dan Saran ……….…..………………….…………... . 68
DAFTAR PUSTAKA……………………………………………… . 69
IV KORELASI POLA IKATAN PEMBULUH PADA
KANDUNGAN KIMIA 4 JENIS BAMBU
Pendahuluan …………………………………..……………… . 72
Bahan dan Metode ……………………………………………. . 73
Hasil dan Pembahasan .……………………………………….. . 73
Simpulan dan Saran ……….…..………………….……………. 87
DAFTAR PUSTAKA………………………………………………. . 88
V . PEMBAHASAN DAN SIMPULAN UMUM
Pembahasan Umum……………………………………………. . 90
Simpulan Umum……………..………………………………… . 96
DAFTAR PUSTAKA……………………………………………….. 98
LAMPIRAN……………………………………………………………… 99

DAFTAR TABEL
Halaman
1 Pola ikatan pembuluh pada 9 jenis bambu….....……….…… ........... 41
2 Hasil uji Duncan untuk persentase serabut……………………….. ... 50
3 Pola ikatan pembuluh pada bambu yang d iteliti………………….. 63
4 Persentase serabut pada 2 jenis bambu ….……………………….. 68
5 Standar pengujian untuk analisis sifat kimia bambu………………… 77
6 Rata-rata nilai kandungan kimia pada berbagai jenis dan pola
bambu…………………………………………………………........... 78
7 Hasil uji lanjut Duncan terhadap kadar ekstraktif pada jenis/pola
bambu………………………………………………………………... 80
8 Ketebalan dinding sel dan persen serabut D. asper, D. giganteus,
C.pergracile dan A.hundsii………………………………………….. 82
9 Hasil uji beda Duncan terhadap kandungan abu……………………... 83
10 Kerapatan ikatan pembuluh dan diameter metaxilem bambu………... 85
11 Hasil uji Duncan terhadap kadar pati bambu ....................................... 88
12 Kerapatan ikatan pembuluh pada berbagai jenis/pola bambu............... 90

DAFTAR GAMBAR
Halaman
1 Skema Kerangka Pemikiran ……………………….……………. 6
2 Alur penelitian................................................................................. 7
3 (a). Bentuk pohon bambu A. hundsii secara utuh, (b) Sayatan
mikro pada penampang lintang bambu (pembesaran 40x)……….. 18
4 Sketsa pola ikatan pembuluh tipe 1 pada A. hundsii.……..….... 18
5 (a). Bentuk pohon bambu A. javonica secara utuh, (b). Sayatan
pada penampang lintang bambu (pembesaran 40x)………………. 19
6 Sketsa pola ikatan pembuluh tipe 1 pada A. javonica……………. 19
7 Bentuk pohon bambu M. baccifera secara utuh ………………….. 19
8 (a). M. baccifera bagian pangkal dengan pembesaran 40x
(b). Sketsa pola ikatan pembuluh 2 pada M. baccifera..…………. 20
9 (a). M. baccifera bagian tengah (pembesaran 40x) (b) Sketsa
pola ikatan pembuluh 2 pada M. baccifera..…………………….. 20
10 (a) Penampang lintang M. baccifera bagian ujung
(pembesaran 40x), (b) Sketsa pola ikatan pembuluh 2 pada
M. baccifera………………………………………………………. 21
11 Bentuk pohon bambu C. pergracile ….………..….……………… 21
12 (a) Tampilan penampang lintang bambu C. pergracile bagian
pangkal dengan pembesaran 40x, (b) Sketsa 1 pola ikatan
pembuluh 2 pada C. pergracile…………………………………… 22
13 (a). Penampang lintang bambu C. pergracile bagian tengah
dengan pembesaran 40x (b) Sketsa pola ikatan pembuluh 2 pada
C. pergracile………………………………………………………. 23
14 (a). Penampang lintang bambu C. pergracile bagian ujung dengan
pembesaran 40x, (b) Sketsa pola ikatan pembuluh 2 pada
C. pergracile .................................................................................... 23
15 Tampilan utuh bambu D. giganteus................................................ 24

16 (a) Penampang lintang bambu D. giganteus bagian pangkal
dengan pembesaran 40x, (b) Sketsa pola ikatan pembuluh 3 pada
D. giganteus..................................................................................... 24
17 (a) Penampang lintang bambu D. giganteus bagian tengah
(pembesaran 40x) (b) Sketsa pola ikatan pembuluh 3 pada
D. giganteus.................................................................……............. 25
18 (a) Penampang lintang bambu D. giganteus bagian ujung dengan
pembesaran 40x, (b) Sketsa pola ikatan pembuluh 3 pada
D. giganteus...............................................................……………... 25
19 Bentuk pohon bambu D. strictus secara utuh ….………………… 26
20 (a). Penampang lintang Dendrocalamus strictus bagian pangkal
(pembesaran 40x) (b). Sketsa pola ikatan pembuluh 3 D. strictus... 27
21 (a) Penampang lintang D. strictus bagian tengah dengan
pembesaran 40x, (b) Sketsa pola ikatan pembuluh 3 pada
D. strictus…………………………………………………………. 27
22 (a) Penampang melintang D. strictus bagian ujung dengan
pembesaran 40x, (b) Sketsa pola ikatan pembuluh 3 pada
D. strictus…………………………………………..……………… 28
23 Bentuk pohon bambu D. asper…………………………………….. 28
24 (a) Penampang melintang bambu D. asper bagian pangkal dengan
pembesaran 40x, (b) Sketsa pola ikatan pembuluh 4……………... 29
25 (a) Penampang lintang D. asper bagian tengah dengan pembesaran
40x, (b) Sketsa pola ikatan pembuluh 3 pada D. asper…………… 30
26 (a) Penampang lintang D. asper bagian ujung dengan pembesaran
40x, (b) Sketsa 1 pola ikatan pembuluh 3 pada D. asper.………… 30
27 Tampilan utuh bambu G. apus…………………………………………. 31
28 (a). Penampang lintang G. apus bagian pangkal dengan
pembesaran 40x, (b) Sketsa pola ikatan pembuluh 4 pada
G. apus……………………………………………………………… 31
29 (a) Tampilan penampang lintang G. apus bagian tengah dengan
pembesaran 40x, (b) Sketsa pola ikatan pembuluh 4 pada
G. apus……………………………………………………………… 32
30 (a) Penampang lintang G. apus bagian ujung dengan pembesaran
40x, (b) Sketsa pola ikatan pembuluh 3 pada G. apus…………….. 32

31 Bentuk pohon bambu G. atroviolecea ………………………….… 33
32 (a) Penampang lintang G. atroviolacea bagian pangkal dengan
pembesaran 40x, (b) Sketsa pola ikatan pembuluh 4 pada
G. atroviolacea…………………………………….………………. 33
.
33 (a) Penampang lintang bambu Gigantochloa atroviolacea bagian
tengah dengan pembesaran 40x, (b) Sketsa pola ikatan pembuluh
4 pada G. atroviolacea……………………………………………... 34
34 (a) Penampang lintang G. atroviolacea bagian ujung dengan
pembesaran 40x, (b) Sketsa pola ikatan pembuluh 3 pada
G. atroviolacea……………………………………………………... 34
35 Sketsa pola ikatan pembuluh 1 (a) dan 2 (b)…………...…………… 35
36 Sketsa pola ikatan pembuluh 3 (c) dan 4 (d)……………………….. 36
37 Pola ikatan pembuluh pada (a) A. javonica (pembesaran 40x),
(a1) Sketsa pola ikatan pembuluh A. javonica, (b) Pola ikatan
pembuluh A. hundsii (pembesaran 40x), (b1) Sketsa pola ikatan
pembuluh 1 pada A. hundsii........………………………………….. 37
38 Pola ikatan pembuluh pada (a) C. pergracile (pembesaran 40x),
(a1) Sketsa pola ikatan pembuluh 2 pada C. pergracile,
(b) M. baccifera dengan pembesaran 40x, (b1) Sketsa pola
ikatan pembuluh 2 pada M. baccifera …………………………….. 38
39 Pola ikatan pembuluh pada (a) D. strictus (pembesaran 40x),
(a1) Sketsa pola ikatan pembuluh 3 pada D. strictus, (b) Pola
ikatan pembuluh D. giganteus (pembesaran 40x), (b1) Sketsa
pola ikatan pembuluh 3 pada D. giganteus …………………………….. 39
40 Bentuk rantai serabut pada bagian ujung (a) D. asper,
(b) G. atroviolacea dan (c) G. apus ……………………………...... 40
41 Kerapatan ikatan pembuluh (/mm2) pada 8 jenis bambu…………... 42
42 Kerapatan ikatan pembuluh pada posisi pangkal, tengah dan ujung... 43
43 Kerapatan ikatan pembuluh pada posisi horizontal…………………. 44
44 Kerapatan ikatan pembuluh pada berbagai pola……………………... 45
45 Posisi panjang serabut (µm) berbagai jenis bambu………………….. 47
46 Posisi panjang serabut (µm) pada 4 pola bambu…………………….. 47

47 Persentase serabut penampang lintang bambu……………………… 51
48 Hubungan antara kerapatan ikatan pembuluh dan persentase
serabut……………………………………………………………….. 52
49 Bentuk contoh uji bobot jenis (BJ)...................................................... 59
50 Bentuk contoh uji tekan sejajar serat................................................... 59
51 Bentuk contoh uji lentur…………………………………………….. 60
52 Bentuk contoh uji tarik sejajar serat………………………………… 60
53 Posisi nilai MOR untuk 3 jenis bambu……………………………… 64
54 Posisi nilai MOR (kg/cm2) untuk bambu dengan pola 3 dan 4…….. 65
55 Posisi nilai keteguhan tekan pada 3 jenis bambu…………………... 67
56 Posisi nilai keteguhan tekan sejajar serat (kg/cm2) pada bagian
batang bambu ……………………………………………………….. 69
57 Posisi jenis bambu berdasar nilai keteguhan tarik (kg/cm2)…………. 70
58 Kadar ekstraktif (%) pada 4 jenis/pola bambu………………………. 79
59 Kadar abu (%) pada 4 jenis /pola bambu …………………….............. 83
60 Kadar lignin (%) pada 4 jenis/pola bambu…………………………... 86
61 Kadar pati (%) 4 jenis /pola bambu…………………………………... 88
62 Kadar alfa selulosa pada 4 jenis/pola bambu…………………………. 91
63 Pemanfaatan bambu berdasarkan pola………………………………... 101

DAFTAR LAMPIRAN
Halaman
1 Panduan penetapan pola ikatan pembuluh berdasar Grosser dan .............
Liese (1971) .............................................................................................. 105
2 Kerapatan ikatan pembuluh pada penampang lintang 8 jenis bambu ....... 106
3 Analisis keragaman pengaruh jenis bambu, posisi vertikal batang
dan pola pada kerapatan pola pembuluh…………… ………………… . 107
4 Panjang serabut 8 jenis bambu .................................................................. 108
5 Analisis keragaman panjang serabut………………………………… ..... 109
6 Analisis keragaman persentase serabut ……………… ............................ 109
7 Nilai Bobot Jenis (BJ) ............................................................................... 110
8 Nilai MOE dan MOR (kg/cm2) pada beberapa jenis bambu..................... 111
9 a. Analisis keragaman MOR …………………………………… ............ 112
b. Analisis keragaman MOE…. ................................................................ 112
10 Nilai keteguhan keteguhan tekan sejajar serat dan keteguhan tarik .......... 113
11 a. Analisis keragaman keteguhan tekan………………………………... 114
b. Analisis keragaman keteguhan tarik………………………………… 114
12 Klasifikasi kelas kuat pada kayu...................................................... 115
13 Persyaratan kayu sebagai bahan baku pulp dan kertas........................ ...... 110
14 Penetapan nilai serat bambu berdasar dimensi serat dan nilai
turunan 8 jenis bambu dikelompokkan menjadi 4 macam pola
mengacu kepada standar kriteria………………………………………. 106

I. PENDAHULUAN UMUM
Latar Belakang
Pemanfaatan hutan sebagai modal pembangunan ekonomi nasional telah
melebihi kemampuannya sebagai sumber daya alam yang dapat diperbaharui.
Peran hutan selama ini baru terfokus pada sisi produksi kayu, sementara hasil
hutan nonkayu yang telah diusahakan oleh masyarakat secara tradisional dan jasa
lingkungan dari ekosistem hutan belum dimanfaatkan secara optimum. Berbagai
kebijakan yang telah dilaksanakan masih belum mampu menyelesaikan
permasalahan di bidang kehutanan. Penerapan kebijakan soft landing hingga kini
berdampak pada kesenjangan bahan baku yang diperkirakan mencapai sekitar 26
juta m3 per tahun. Sementara itu, nilai tambah dari produk hutan nonkayu seperti
air, udara bersih, keanekaragaman hayati, dan keindahan alam belum berkembang
seperti yang diharapkan untuk mendukung sektor ekonomi (Bappenas 2011).
Saat ini kebutuhan bahan baku kayu di tingkat nasional semakin meningkat
sedangkan kemampuan pasokan kayu dari hutan alam terus menurun karena
tingkat deforestasi yang tinggi dari tahun ke tahun (Forest Watch Indonesia 2011).
Hal ini akan mengakibatkan berkurangnya persediaan bahan baku. Keadaan ini
tentu saja kurang menguntungkan bagi keberadaan dunia usaha perkayuan. Upaya
yang dapat dilakukan agar usaha tetap berlangsung adalah dengan mencari
material yang setara dengan kayu, salah satu yang ditawarkan adalah bambu
yang memiliki sifat seperti kayu dan potensinya cukup besar.
Bambu merupakan tanaman monokotil yang memiliki anatomi sederhana
karena pertumbuhan dan diferensiasi selnya terjadi sangat cepat (Liese 2006).
Struktur jaringan pada bambu tersusun dalam bentuk pola ikatan pembuluh dan
terletak terpencar pada jaringan dasar parenkim. Bambu sebagai bahan substitusi
kayu memiliki keunggulan, yaitu sebagai tanaman yang cepat tumbuh, dapat
dipanen pada umur 4 tahun (memiliki rotasi tebang pendek) sehingga untuk satuan
waktu tertentu mempunyai tingkat produktivitas yang tinggi. Ditinjau dari sisi
potensi, negara Indonesia memiliki 143 jenis dari keseluruhan keanekaragaman
bambu di dunia yang berjumlah 1200–1300 jenis (Widjaja 2001) juga 50%
bambu-bambu unggul di dunia ada di Indonesia (Darupratomo 2008). Bambu

juga dapat ditemukan hampir di setiap pulau di Indonesia (FAO 2002) sehingga
masih banyak peluang untuk memanfaatkan bambu sebagai bahan baku, apalagi
baru 32 jenis bambu saja yang memiliki kegunaan yang jelas (Widjaja et al.
2004). Dengan kelebihan ini diharapkan kebutuhan bahan baku teratasi,
walaupun bambu memiliki kelemahan, yaitu pada umumnya memiliki daya tahan
yang rendah terhadap jamur dan serangga (Barly 1999).
Di antara negara penghasil bambu, China adalah salah satu penghasil
bambu terbesar di dunia (Jifan 1985). Sebagai bahan yang sangat penting, bambu
di negara China digunakan untuk industri perikanan, konstruksi, pulp dan kertas,
bahan kerajinan dan dalam kehidupan sehari-hari. Penggunaan bambu di
Indonesia mengacu pada hasil penelusuran Nuriyatin (2000), ternyata memiliki
20 jenis kegunaan di masyarakat. Penggunaan bambu bisa berkembang lebih
banyak lagi baik melalui eksplorasi jenis-jenis bambu maupun eksplorasi
penggunaan lain dengan serangkaian kegiatan penelitian. Mengingat keadaan ini,
perlu dilakukan berbagai pendukung penggunaan bambu secara lebih luas
termasuk penggunaan bukan hanya secara tradisional. Dengan demikian terbuka
lebar peluang untuk memanfaatkan bambu secara tepat baik yang menyangkut
mutu maupun ragam penggunaan.
Ada hal yang menjadi pertanyaan, yaitu kenapa beberapa jenis bambu
mempunyai kegunaan tertentu sedangkan jenis bambu yang lain tidak dapat
dipergunakan untuk keperluan itu. Hal ini berarti bahwa suatu jenis bambu
mempunyai ciri tertentu sehingga akan sesuai jika digunakan untuk tujuan
tertentu saja. Pendekatan yang dilakukan untuk menjawab pertanyaan tersebut
adalah melalui serangkaian kegiatan penelitian sifat-sifat dasar terutama dimulai
dari penelusuran sifat anatomi. Penelitian sifat anatomi mempunyai peranan yang
sangat penting karena dapat menentukan sifat-sifat bambu dan penggunaannya
(Liese 2003).
Penelitian anatomi bambu diawali oleh Ota (1951) yang meneliti pengaruh
persentase elemen struktur pada bobot jenis (BJ) dan kekuatan bambu.
Perkembangan selanjutnya adalah munculnya penelitian Liese (1986) yang
mempelajari tentang karakter dan penggunaan bambu. Setahun kemudian, Liese
(1987) meneliti sifat-sifat dasar bambu yang menginformasikan sifat-sifat

anatomi, kimia, fisik, dan mekanik bambu, Mohmod et al. (1990) mendalami
tampilan anatomi dan sifat-sifat mekanik dari 3 jenis bambu Malaysia.
Selanjutnya Liese (1992) meneliti struktur bambu dan hubungannya dengan sifat-
sifat dan penggunaan di masyarakat. Penelitian ini lebih diperdalam lagi oleh
Liese (2003) melalui pengamatan pengaruh struktur bambu terhadap
pemanfaatannya. Penelitian yang lebih terperinci tentang karakterisasi anatomi
bambu telah dilakukan oleh Londono et al. (2002) yang mencoba menganalisis
karakter anatomi bambu Guadua angustifolia. Penelitian yang sama telah
dilakukan juga oleh Nuriyatin (2000), yaitu tentang sifat-sifat dasar bambu pada
beberapa tujuan penggunaan namun belum sampai pada tahap analisis hubungan
antara pola struktur dan tujuan penggunaan. Dengan demikian kajian lebih
mendalam perlu dikembangkan mengingat manfaatnya bagi perkembangan ilmu
pengetahuan dan penggunaannya di masyarakat.
Batang bambu tersusun selain atas 50% parenkim, 40% serabut, juga oleh
10% sel-sel penyalur (Dransfield dan Widjaja 1995). Grosser dan Liese (l973)
mengemukakan bahwa struktur anatomi batang-batang bambu terutama
ditentukan oleh ikatan pembuluh yang tertanam dalam jaringan parenkim. Lebih
jauh dinyatakan pula oleh Lwin et al. (2007) bahwa struktur anatomi penampang
melintang ruas bambu ditentukan oleh bentuk, ukuran, susunan dan jumlah ikatan
pembuluh. Ikatan pembuluh itu sendiri merupakan susunan pola pembuluh yang
terdiri atas 2 pembuluh metaxilem dengan 1 atau 2 elemen protoxilem yang kecil
dan floem (Liese 1980). Ikatan pembuluh merupakan karakteristik anatomi yang
stabil karena tidak dipengaruhi umur (Londono et al. 2002). Dengan demikian
penelitian pada anatomi bambu selalu mempertimbangkan tampilan ikatan
pembuluh sebagai suatu hal yang penting (Liese 2006).
Grosser dan Liese (l971) menyatakan bahwa pola ikatan pembuluh yang
terdapat pada bambu terdiri atas 4 jenis yaitu tipe 1, 2, 3, dan 4 yang menjadi
pembeda antargenus dan antarspesies. Perkembangan lebih lanjut, pola ikatan
pembuluh menjadi 5 jenis dengan jenis yang ke-5 merupakan pengembangan pola
1 (Liese 1985, Taihui dan Wenwei 1985). Liese (1998) menemukan
pengembangan pola ke-2 sehingga secara keseluruhan ada 6 jenis pola ikatan
pembuluh.

Pola ikatan pembuluh merupakan susunan pola pembuluh yang didukung oleh
serabut baik berada dalam bentuk selubung sklerenkim (pola 1 dan 2) maupun
berbentuk rantai serabut (pola 3 dan 4). Keberadaan serabut pada setiap pola
cukup penting karena memberikan kontribusi 60-70% terhadap bobot total
jaringan batang (Lwin et al. 2007). Tempat beradanya dan distribusi serabut akan
mempengaruhi sifat-sifat tertentu seperti misalnya mempengaruhi BJ dan sifat
kekuatan. Kandungan serabut yang tinggi juga berpengaruh pada produksi pulp
(Liese 1992). Demikian pula panjang serabut mempengaruhi sifat-sifat kekuatan.
Struktur lamela serabut pada bagian pinggir tepi batang berpengaruh pada sifat-
sifat mekanik. Kandungan dan juga penyebaran serabut pada setiap pola akan
memberikan kontribusi pada tujuan pemakaian akhir bambu di mana di antara ke-
4 pola ikatan yang ada masing-masing memiliki karakteristik yang berbeda dan
perbedaan ini signifikan diantara spesies dan genus (Londono et al. 2002, Grosser
dan Liese 1971).
Dalam penggunaan bambu sebagai bahan kostruksi ternyata bambu
memiliki nilai kekuatan yang cenderung meningkat dari bagian pangkal ke bagian
ujung (Nuriyatin 2000). Penelitian Nuriyatin (2001) telah menganalisis sifat dasar
bambu dikaitkan dengan tujuan penggunaan, melalui penelitian tersebut
terungkap bahwa dari 5 jenis bambu yang diteliti yaitu seluruhnya layak untuk
dijadikan bahan baku pulp dan kertas. Hasil pengujian mutu suara menyatakan
bahwa Gigantochloa atroviolacea (bambu hitam) memiliki mutu suara terbaik
karena selain memiliki BJ cukup tinggi juga ikatan pembuluh dalam batang
menyebar secara merata. Penelitian ini belum sempurna karena tidak sampai pada
keterkaitan langsung antara pola ikatan pembuluh dengan tujuan penggunaan.
Berdasarkan analisa data dan hasil penelitian sebelumnya diduga bahwa pola
ikatan pembuluh berpengaruh pada pola penggunaan.
Struktur kimia bambu memiliki peranan penting karena mempunyai banyak
hubungan dengan penggunaan (Liese l992, Liese 2006) diantaranya bahwa
kandungan holoselolosa dari bahan tanaman adalah penting untuk industri seperti
pulp dan kertas (Youdi et al. 1985). Demikian pula kandungan ekstraktif suatu
jenis bambu akan memberikan informasi terhadap penggunaan karena dapat
mengontrol keawetan juga bertanggung jawab terhadap peningkatan BJ dan

penurunan keseimbangan kadar air (Liese 2006). Dengan demikian sifat kimia
merupakan sifat yang selalu diikutsertakan dalam studi sifat dasar.
Berdasar uraian tersebut diduga terdapat hubungan antara pola ikatan
pembuluh dengan keragaman sifat dasar. Hal ini berarti juga diduga ada
keterkaitan antara pola ikatan pembuluh dengan penggunaan. Sehubungan
dengan hal itu maka perlu dikembangkan penelitian sehingga dapat diperoleh
dasar yang kuat untuk membentuk pola pendugaan pemanfaatan bambu. Untuk
lebih jelasnya alur kerangka pemikiran dalam penelitian ini dapat diamati pada
Gambar 1 sedangkan kerangka penelitian terdapat pada Gambar 2.
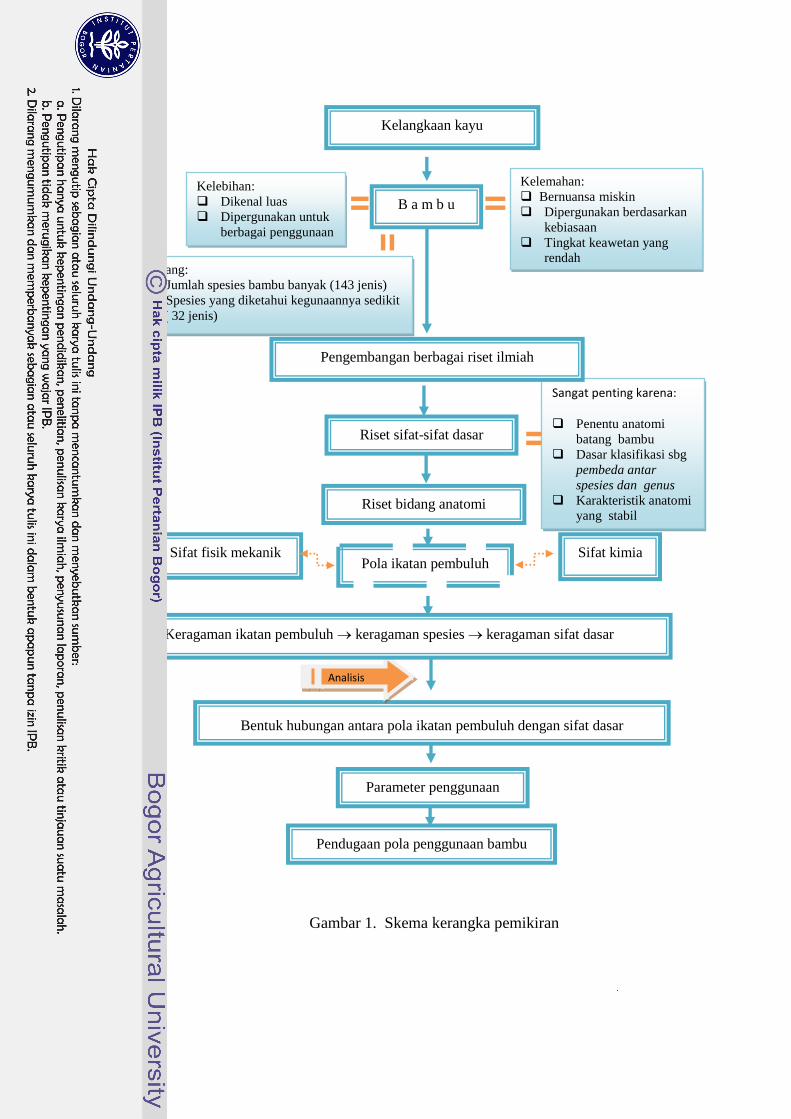
Gambar 1. Skema kerangka pemikiran
Bentuk hubungan antara pola ikatan pembuluh dengan sifat dasar
Analisis
data
Pendugaan pola penggunaan bambu
Keragaman ikatan pembuluh keragaman spesies keragaman sifat dasar
Riset sifat-sifat dasar
Pola ikatan pembuluh
Kelemahan:
Bernuansa miskin
Dipergunakan berdasarkan
kebiasaan
Tingkat keawetan yang
rendah
Sangat penting karena:
Penentu anatomi
batang bambu
Dasar klasifikasi sbg
pembeda antar
spesies dan genus
Karakteristik anatomi
yang stabil
Kelebihan:
Dikenal luas
Dipergunakan untuk
berbagai penggunaan
Peluang:
Jumlah spesies bambu banyak (143 jenis)
Spesies yang diketahui kegunaannya sedikit
( 32 jenis)
Riset bidang anatomi
Sifat fisik mekanik Sifat kimia
Kelangkaan kayu
B a m b u
Pengembangan berbagai riset ilmiah
Parameter penggunaan
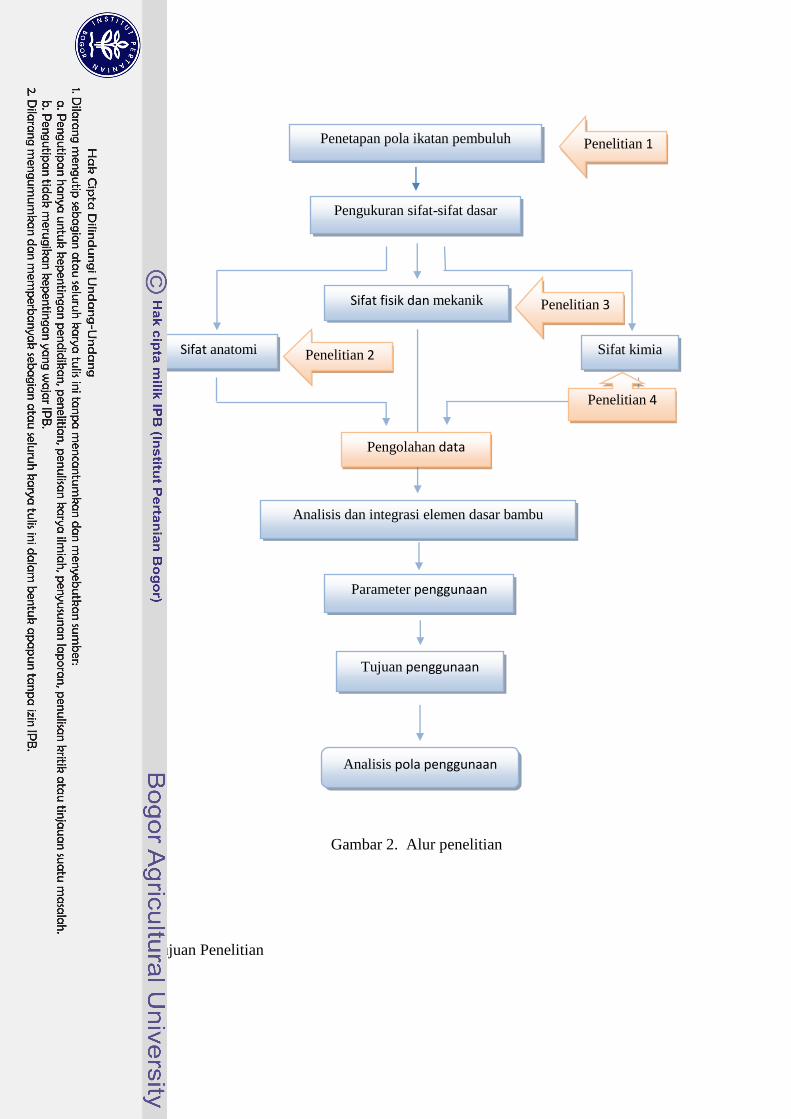
Tujuan Penelitian
Penelitian 3
Penelitian 2
Sifat fisik dan mekanik
Sifat anatomi
Pengolahan data
Analisis dan integrasi elemen dasar bambu
Tujuan penggunaan
Analisis pola penggunaan
Parameter penggunaan
Penelitian 4
Gambar 2. Alur penelitian
Penelitian 1
Pengukuran sifat-sifat dasar
Penetapan pola ikatan pembuluh
Sifat kimia

Tujuan penelitian ini ialah menentukan penggunaan bambu berdasarkan
karakter yang dimiliki oleh pola ikatan pembuluh.
Hipotesis
Hipotesis dalam penelitian ini ialah bahwa pola ikatan pembuluh pada
penampang lintang bambu akan memberikan kontribusi terhadap karakter
bambu, dan pola ikatan pembuluh akan membantu dalam mengarahkan pola
penggunaan bambu
Manfaat Penelitian
Bagi peneliti diharapkan pola ikatan pembuluh memberikan kontribusi
dalam pemanfaatan bambu secara optimum dengan melibatkan secara langsung
sebagai faktor tunggal ataupun bagian dari variabel yang menentukan pola
pemanfaatan bambu. Bagi masyarakat/kalangan industriawan diharapkan hasil
penelitian dapat diaplikasikan dalam pendugaan pemanfaatan bambu. Hal ini
didukung dengan cara penetapan pola ikatan pembuluh yang relatif mudah.
Dengan demikian pemanfaatan yang sesuai dengan karakter bambu diharapkan
akan berguna dalam penggunaan bambu sehingga proses dan produk yang
dihasilkan akan optimum. Bagi dunia keilmuan diharapkan hasil penelitian akan
memberikan kontribusi dalam pengembangan ilmu terutama karena peranannya
dalam menemukan hal baru sehingga dapat memanfaatkan sumber daya bambu
secara efisien.
Ruang Lingkup Penelitian
Penelitian dilakukan pada jenis-jenis bambu yang mewakili 4 pola ikatan
pembuluh terutama yang diambil dari koleksi bambu yang tumbuh di Kebun Raya
Bogor dan dari lokasi arboretum Fakultas Kehutanan di Kampus Institut Pertanian
Bogor, Darmaga, Bogor. Pengujian sifat anatomi, fisik, mekanik dan kimia
dilakukan pada posisi pangkal, tengah dan ujung batang khususnya pada bagian
ruas. Penetapan pola ikatan pembuluh dilakukan pada 9 jenis bambu dan
pengolahan data sifat anatomi hanya pada 8 jenis bambu. Pengujian sifat fisik
mekanik pada 3 jenis bambu (terkait dengan persyaratan sampel uji). Pengujian

sifat kimia hanya pada 4 jenis bambu yang mewakili setiap pola ikatan pembuluh.
Pengolahan data menggunakan analisis deskripsi, analisis keragaman dan analisis
regresi dengan peubah boneka (dummy).

DAFTAR PUSTAKA
Barly. 1999. Pengawetan bambu untuk bahan konstruksi bangunan dan mebel
(petunjuk teknis). Bogor: Badan Penelitian dan Pengembangan Kehutanan
dan Perkebunan Pusat Penelitian dan Pengembangan Hasil Hutan dan Sosial
Ekonomi Kehutanan.
[Bappenas] Badan perancang pembangunan nasional. 2011. Perbaikan
pengelolaan sumber daya alam dan pelestarian lingkungan hidup.
www.bappenas.go.id/get-file-server/node/1134/ [ 25 D 2011].
Darupratomo. 2008. Pengaruh proses pengawetan bambu terhadap karakteristik
bambu sebagai bahan bangunan. Prospect,tahun 4, no. 6, hal: 7-20.
Dransfield, Widjaja. E. 1995. Bamboos. Indonesia: Prosea Bogor.
Forest Watch Indonesia. 2011. Potret keadaan hutan Indonesia periode 2000-
2009. www.fwi.or.id [25 Desember 2011].
[FAO] Food and Agricultural Organizatin. 2002. Non-wood forest products in
15 countries of tropical Asia : A regional and national overview. Vantomme
P, Markkula A, Leslie RN, editor.
Grosser D, Liese W. 1971. On the anatomy of Asian bamboos, with spesial
reference to their vaskular bundles. Wood Sci and Tech 5: 290-312.
Grosser D, Liese W. l973. Present status and problems of bamboo
classification. J Arn Abor 54 (2): 293-308
Jifan Z. 1985. Bamboo development in China. Dalam Rao AN, Dhanarajan G,
Sastry CB, editor. Proceedings of the International Bamboo Workshop;
Hangzhou, People’s Republic of China, October 6-14, 1985.
Lwin KM, Han YY, Maung W, Moe AKZ, Than SBM. 2007. An investigation on
morphology, anatomy and chemical properties of some Myanmar bamboos.
http://www.myanmar.gov.mm/Ag/Jur/ProcFo01.10.[3 Nov. 2007]
Liese W. 1980. Anatomy of Bamboo. Di dalam: Lessard G dan Chounard A,
editor. Proceedings of a Workshop. Singapore, 28-30 Mei 1980.
Liese W. 1985. Anatomy and properties of bamboo. Di dalam : Rao, A.N.,
Dhanarajan, G. dan Sastry, C.B, editor. Recent Research on Bamboo.
Proceedings of the International Bamboo Workshop, Hangzholu, People's
Republic of China, 6-14 Oktober, 1985. Academy of Forestry, People's
Republic of China & International ,Development Research Centre, Canada,
hal : 196-208.

Liese W. 1986. Characterization and utilization of bamboo. In: Bamboo
production and utilization. Proceedings VIII IUFRO World Congress,
Ljubljana, Yugoslavia, September 7-21: 11-16.
Liese W. 1987. Anatomy and properties of bamboo. Di dalam : Rao, A.N.,
Dhanarajan, G. dan Sastry, C.B, editor. Recent Research on Bamboo.
Proceedings of the International Bamboo Workshop, Hangzholu, People's
Republic of China, 6-14 Oktober, 1985. Academy of Forestry, People's
Republic of China & International ,Development Research Centre, Canada,
hal: 196-208.
Liese W. 1992. The structure of bamboo in relation to its properties and
utilization. Dalam Bamboo And Its Use, International Symposium On
Industrial Use Of Bamboo. Beijing, China, 7-11 Desember 1992, hal: 1 – 6.
Liese W. 1998. The anatomy of bamboo culms. INBAR Technology Report No
18.
Liese W. 2003. Structures of bamboo culm affecting its utilization. Di dalam
Xuhe C, Yiping L, Ying H, editor. Proceedings of International Workshop
on Bamboo Industrial Utilization. Hubei dan Xianning, Oktober 2003. hlm
6 – 10.
Liese W. 2006. The Anatomy of bamboo culms. Http://www.inbar.int/
publication/txt/tr18/default2.htm [24 Desember 2006].
Londono X, Camayo GC, Riano NM, Lopez Y. 2002. Characterization of the
anatomy of Guadua angustifolia (Poaceae: Bambusoideae) culms. J Am
Bam Soc 16 (1): 18-31.
Nuriyatin N. 2000. Studi analisa sifat-sifat dasar bambu pada beberapa tujuan
penggunaan [tesis]. Bogor: Jurusan Ilmu Pengetahuan Kehutanan, Institut
Pertanian Bogor.
Nuriyatin N. 2001. Studi analisa sifat dasar bambu sebagai bahan baku kertas. J
Ilm Pert Ind 3: 56-61.
Ota M. 1951. The influence of the percentage of structural elements on the
specific gravity and compressive strength of bamboo splint. J Jappan
Forest Soc 19: 25-47.
Taihui W, Wenwei C. 1985. A Study on the anatomy of the vascular bundles of
bamboos from China. Di dalam : Rao, A.N., Dhanarajan, G. dan Sastry,
C.B, editor. Recent Research on Bamboo. Proceedings of the International
Bamboo Workshop, Hangzholu, People's Republic of China, 6-14 Oktober,
1985. Academy of Forestry, People's Republic of China & International
,Development Research Centre, Canada.

Widjaja, EA. 2001. Identifikasi Jenis-jenis Bambu di Jawa. Bogor : LIPI–seri
panduan lapangan.
Widjaja EA, Utami NW, Saefudin. 2004. Panduan Membudidyakan Bambu.
Bogor : LIPI.
Youdi C, Wenlong Q, Xiuling L, Jianping G, Nimanna. 1985. The Chemical
Composition of Ten Bamboo Spesies. Di dalam : Rao, A.N., Dhanarajan,
G. dan Sastry, C.B, editor. Recent Research on Bamboo. Proceedings of the
International Bamboo Workshop, Hangzholu, People's Republic of China, 6-
14 Oktober, 1985. Academy of Forestry, People's Republic of China &
International Development Research Centre, Canada.

II. ANATOMI PENAMPANG LINTANG BATANG
9 JENIS BAMBU
Abstrak
Ketersediaan bambu yang melimpah serta keterbatasan pemanfaatan bambu
mendorong dilakukannya penelitian dasar di bidang anatomi untuk memperoleh
landasan ilmiah mengenai karakter bambu yang dapat digunakan untuk
mengarahkan pemanfaatan bambu secara optimum. Metode analisis adalah
pendekatan regresi dengan peubah boneka dengan melibatkan faktor jenis, posisi
bambu secara vertikal (pangkal, tengah, dan ujung), posisi bambu secara
horizontal (tepi, tengah, pusat dan dalam), serta pola ikatan pembuluh bambu
(pola 1, 2, 3, dan 4). Hasil penelitian ini menetapkan bahwa setiap spesies bambu
memiliki pola ikatan pembuluh dari pola 1 sampai pola 4, baik itu pola tunggal
ataupun pola kombinasi. Dendrocalamus strictus, bagian ujung batang, bagian
tepi penampang lintang, dan pola ikatan pembuluh 1 memiliki nilai kerapatan
ikatan pembuluh tertinggi dan berbeda nyata jika dibandingkan dengan lainnya.
Bambu Cephalostachyum pergracile memiliki nilai panjang serabut terendah
dibandingkan dengan spesies bambu lain. Panjang serabut tertinggi dimiliki oleh
bambu-bambu yang memiliki pola 4. Persen serabut tertinggi pada penampang
lintang batang ada pada bagian tepi. Kesimpulan penelitian ini adalah bahwa pola
ikatan pembuluh mempunyai nilai penting untuk membedakan jenis bambu juga
dapat digunakan untuk menentukan arah penggunaan bambu.
Kata kunci: kerapatan ikatan pembuluh, pola, persentase serabut, panjang serabut,
penampang lintang
Abstrak
Abundant availability of bamboo and limitations in the use of bamboo
encourage basic research in the field of anatomy. The study is expected to
provide a scientific basic for the character of bamboo that can be used to direct the
optimum utilization of bamboo. The research method is the approach of
regression with dummy variables which involved factors of bamboo species,
position of bamboo vertical (base, middle, and top), bamboo horizontal position
(edge, middle, center, inner) and bamboo patterns (1, 2, 3 and 4 ). The result of
this study is that every bamboo species has vascular bundle pattern from 1 to 4 as
well as the single patern and the combination pattern. Dendrocalamus strictus
species, the culm top, edges of the cross section, vascular bundle pattern 1 have
the highest vascular bundle density values and significantly different with others.
Cephalostachyum pergracile fiber length has the lowest compared to other
species of bamboo but pattern 4 has the highest fiber length. The highest percent
of fibers in the cross section of the stem is at the edge. The conclusion of this
study is the vascular bundle patterns have important value for distinguishing
species of bamboo and they can also be used to determine the direction of
bamboo’s utilization.
Key word : vascular bundle density, pattern, fibre percentage, fibre length, cross
section

Pendahuluan
Bambu merupakan tanaman monokotil yang tersedia melimpah di Indonesia
bahkan di dunia, yang digunakan untuk berbagai tujuan walaupun dalam hal ini
tidak semua spesies sesuai untuk tujuan tertentu. Bambu mempunyai sifat-sifat
fisik dan mekanik yang berbeda sehingga menghasilkan produk dengan mutu
yang berbeda pula. Pengetahuan mengenai komponen anatomi bambu memegang
peranan penting bahkan diperlukan dalam penemuan suatu produk baru. Dengan
demikian, riset dasar sangat penting untuk mendapatkan suatu karakterisasi
bambu. Menurut American Bamboo Society (1999), riset pada sifat-sifat dasar
akan membawa kepada penggunaan yang lebih baik dan peningkatan nilai tambah
produk. Untuk penggunaan yang lebih optimum diperlukan kriteria tertentu yang
sesuai dan sering terkait dengan struktur sel (anatomi) dan sifat-sifat pada bambu
(Liese 1987). Gritsch dan Murphy (2005) menyatakan bahwa struktur anatomi
bambu menentukan sifat dasar terutama sifat fisik dan mekanik.
Bambu dikenal sebagai salah satu tanaman cepat tumbuh sehingga dapat
menjadi alternatif terbaik pengganti kayu di masa datang. Tidak seperti kayu,
bambu hanya memerlukan 3-4 tahun untuk siap tebang dan digunakan (Wahab et
al. 2009). Jenis-jenis bambu yang tumbuh di Indonesia sangat banyak dan belum
dimanfaatkan secara optimum. Dengan demikian, terbuka peluang untuk
memanfaatkan lebih banyak jenis-jenis bambu yang ada.
Untuk mengenal seluruh jenis bambu relatif sulit karena banyak bambu
yang belum dikenal di masyarakat. Perlu dicari upaya lain agar dapat mengenal
dalam konteks menggunakan bambu secara tepat dengan lebih mudah. Penelitian
ini mencoba untuk mengamati secara mendalam sifat anatomi penampang lintang
bambu dengan memperhatikan berbagai informasi yang muncul dari penampang
lintang bambu-bambu yang diujikan. Penelitian pada bidang anatomi sebaiknya
dikembangkan untuk menggali potensi yang dimiliki bambu sehingga hasilnya
dapat dipergunakan terutama dalam memanfaatkan bambu secara optimum (Lwin
et al. 2007).
Seperti halnya tanaman monokotil lain, anatomi batang bambu tersusun
selain oleh parenkim sebagai jaringan dasar juga oleh ikatan pembuluh yang
tertanam dalam parenkim (Londono et al. 2002). Keragaman di antara genus dan

spesies bambu terkait dengan jenis pola ikatan pembuluh (Grosser dan Liese
1971). Tampilan pola ikatan pembuluh itu sendiri akan dapat dilihat dengan jelas
pada penampang melintang bambu (Lwin et al. 2007). Bambu memiliki 4 pola
ikatan pembuluh yaitu tipe 1 yang terdapat pada genus Leptomorph seperti
Arundinaria, tipe 2 terdapat pada genus Melocanna dan Cephalostachyum,
sedangkan tipe 3 dan 4 muncul pada genus Dendrocalamus dan Gigantochloa.
Perbedaan struktur anatomi yang mendasar antara keempat pola mempengaruhi
sifat-sifat kerapatan, kekuatan, dan kelenturan (Grosser dan Liese 1971). Dengan
demikian, melalui penelitian sifat anatomi diharapkan akan dapat diketahui sifat-
sifat struktural dan hubungan dengan sifat dasar lainnya secara lengkap.
Penetapan pola ikatan pembuluh dilakukan pada 9 jenis bambu. Pada
proses selanjutnya sampel bambu yang diambil hanya 8 jenis bambu (tanpa
Gigantochloa atroviolacea) karena setiap pola cukup terwakili oleh 2 jenis
bambu. Diharapkan melalui kegiatan penelitian ini akan diperoleh informasi
secara lengkap mengenai sifat anatomi penampang lintang batang bambu.
Bahan dan Metode
Bahan
Bahan penelitian adalah 9 jenis bambu yang telah berumur 3-4 tahun serta
memiliki pola ikatan pembuluh 1-4 yang ditentukan berdasarkan panduan
penetapan pola ikatan pembuluh bambu oleh Grosser dan Liese (1971) (Lampiran
1) dengan ulangan 3 kali. Bambu-bambu tersebut terdiri atas Arundinaria
hundsii Munro (Ah), Arundinaria javonica (Aj), Melocanna baccifera (Mb),
Cephalostahyum pergracile (Munro) (Cp), Dendrocalamus giganteus (Wallich ex
Munro (Dg/sembilang), Dendrocalamus strictus (Roxb.) Nees (Ds),
Dendrocalamus asper (Schultes f.) (Da/betung/petung), Gigantochloa
atroviolacea (Widjaja) (Gat/hitam atau pring wulung), dan Gigantochloa apus
(J.A. & J.H. Schultes) Kurz (Ga/tali) dengan lokasi pengambilan 6 jenis bambu
pertama di Kebun Raya Bogor, sedangkan lokasi pengambilan 3 jenis bambu yang
terakhir dari daerah di Fakultas Kehutanan, Institut Pertanian Bogor.

Penentuan sifat anatomi
Variabel pengamatan adalah tipe dan kerapatan ikatan pembuluh, panjang
serabut dan persentase serabut. Sampel uji ditetapkan pada penampang lintang
ruas tengah bagian pangkal, tengah, dan ujung batang bambu, sedangkan pada
posisi horizontal (penampang lintang batang), sampel uji ditetapkan pada bagian
tepi, tengah, pusat, dan dalam. Maserasi menggunakan metode Schultze,
sedangkan pembuatan preparat sayatan mengacu kepada Sass (1951).
Kerapatan ikatan pembuluh ditentukan berdasarkan perhitungan jumlah pola
ikatan pembuluh untuk setiap luasan tertentu. Persentase serabut ditentukan
dengan menghitung luasan serabut untuk setiap luasan tertentu dalam satuan
persen. Kerapatan ikatan pembuluh dan persentase serabut diukur dengan alat
stereo discovery V8 merk Zeiss dengan kamera Axio Cam M Rc 5 yang
dihubungkan komputer dengan perangkat lunak Axio Vision Rel. 4.6. Sementara
pengukuran panjang serabut dilakukan dengan mikroskop. Dokumentasi foto
mikro bambu dilakukan dengan bantuan mikroskop dengan pembesaran 40 kali.
Analisis data
Data dianalisis dengan pendekatan regresi dengan peubah boneka. Peubah
boneka dalam analisis ini adalah 8 (delapan) jenis bambu yang terwakili dalam
peubah X1-X7; 3 (tiga) posisi vertikal yaitu pangkal, tengah, dan ujung yang
terwakili dalam peubah X8 dan X9; 4 (empat) posisi horizontal yaitu tepi, tengah,
pusat, dan dalam yang terwakili dalam X10-X12; 4 (empat) pola bambu yang
terwakili dalam X13- X15. Dalam penentuan panjang serabut hanya ada 12 peubah
boneka (dummy) karena tidak ada peubah untuk posisi horizontal. Kontribusi
keseluruhan peubah akan dianalisis dalam persamaan regresi.
Hasil dan Pembahasan
Deskripsi pola ikatan pembuluh bambu
Pengamatan pola ikatan pembuluh pada 9 jenis bambu memberikan hasil
berupa penetapan pola ikatan pembuluh. Penetapan pola ikatan pembuluh

dilakukan dengan pengamatan bentuk pola pada bagian pusat (setelah bagian
tengah) penampang lintang batang karena bentuk dan ukuran pola telah mencapai
diferensiasi optimum (Londono et al. 2002). Pada bagian tengah batang bambu,
Arundinaria hundsii dan Arundinaria javanica memiliki ikatan pembuluh pola 1,
sedangkan Melocanna baccifera dan Cephalostahyum pergracile baik bagian
pangkal, tengah, dan ujung memiliki pola 2. Demikian pula Dendrocalamus
giganteus dan Dendrocalamus strictus memiliki pola 3 pada keseluruhan bagian
batang (pangkal, tengah, dan ujung). Namun, pola pada bambu Dendrocalamus
asper berbeda dengan spesies bambu sebelumnya, yaitu pada bagian pangkal
memiliki pola 4, sedangkan pada bagian tengah dan ujung mempunyai pola 3.
Bambu Gigantochloa atroviolacea dan Gigantochloa apus pada bagian pangkal
dan tengah mempunyai pola 4, sedangkan pada bagian ujung mempunyai pola 3.
Selanjutnya, akan diuraikan deskripsi pola ikatan pembuluh untuk setiap jenis
bambu.
Arundinaria hundsii
A. hundsii adalah bambu yang termasuk kelompok bambu monopodial.
Umumnya, ketinggian batang 3 m. Bambu ini memiliki dinding batang tipis
dibandingkan dengan spesies bambu lain, yaitu tebal dinding batang bagian
pangkal rata-rata 0.6 cm, sedangkan bagian ujung rata-rata 0.4 cm. Bentuk pola
pada penampang lintang bambu dapat diamati secara jelas pada bagian tengah dan
bagian dalam yang berbentuk pola 1. Bentuk pola pada bagian tepi belum terlihat
jelas karena rantai pembuluh pusat masih bergabung dengan rantai serabut.
Sementara itu, bentuk pola pada bagian tengah dan dalam terlihat jelas. Selubung
sklerenkim pada bagian tengah lebih tebal dibandingkan dengan bagian dalam.
Adapun gambaran tanaman bambu secara utuh dan sayatan pola ikatan pembuluh
pada penampang lintang bambu A. hundsii selengkapnya pada Gambar 3 dan
sketsa pola ikatan pembuluh pada tipe 1 pada Gambar 4.

Arundinaria javonica
Seperti halnya spesies bambu sebelumnya, bambu A. javonica adalah bambu
yang termasuk kelompok monopodial yang berasal dari negara Jepang (Botanic
Garden of Indonesia 2001). Ketinggian batang bambu ini rata-rata 4 m dengan
tebal dinding batang bagian pangkal rata-rata adalah 0.6 cm dan bagian ujung
adalah 0.4 cm. Umumnya, dinding batang hanya terbagi atas 3 bagian saja yaitu
bagian tepi, tengah, dan bagian dalam. Bentuk pola ikatan pembuluh antara bagian
tengah dan dalam hampir sama, tetapi ada sedikit perbedaan dalam selubung
serabut, yaitu pada bagian dalam ukurannya lebih kecil. Bentuk pola terlihat jelas
pada bagian tengah yaitu membentuk pola 1. Tampilan bambu secara utuh dan
sayatan penampang lintang A. javonica dapat dilihat pada Gambar 5, sedangkan
sketsa pola ikatan pembuluh tercantum pada Gambar 6.
Gambar 3. (a). Bentuk pohon bambu A. hundsii secara utuh, (b). Sayatan
mikro pada penampang lintang bambu (pembesaran 40x)
parenkim
metaxilem
ruang antar sel
sklerenkim
jar. floem
Gambar 4. Sketsa pola ikatan pembuluh tipe 1 pada A. hundsii
kulit
dalam
(a)
(b)

Melocanna baccifera
Bambu M. baccifera berasal dari negara Banglades, Burma, dan India, tetapi
telah menyebar luas ke seluruh dunia ( Dransfield dan Widjaja 1995). Bambu ini
termasuk kelompok simpodial. Di Kebun Raya Bogor, bambu ini tumbuh dengan
tinggi batang rata-rata 8 m dan tebal dinding batang pangkal adalah 0.7 cm dan
bagian ujung 0.3 cm. Tampilan secara utuh pohon bambu Melocanna baccifera
terlihat pada Gambar 7. Uraian sifat anatomi berdasarkan bagian pada batang,
yaitu pada pangkal, tengah, dan ujung.
Gambar 7. Bentuk pohon bambu M. baccifera secara utuh
Gambar 5. (a). Bentuk pohon bambu A. javonica secara utuh, (b). Sayatan pada
penampang lintang bambu (pembesaran 40x)
parenkim
metaxilem
ruang antar sel
sklerenkim
jar. floem
Gambar 6. Sketsa pola ikatan pembuluh tipe 1 pada A. javonica
kulit
dalam
(a)
(b)

Pangkal
Pola ikatan pembuluh pada bambu M. baccifera merupakan pola 2 yang
terlihat jelas pada bagian tengah penampang lintang batang. Pada bagian tepi
penampang lintang, selubung serabut masih menyatu dengan rantai pembuluh
pusat. Sementara itu, pada bagian tengah penampang lintang batang, bentuk pola
sudah jelas terlihat. Pada bagian dalam, gambaran pola tidak sempurna karena
perluasan selubung sklerenkim pada ruang antar sel sangat sedikit (Gambar 8).
Tengah
Seperti pada bagian pangkal, pola ikatan pembuluh bambu M. baccifera
bagian tepi penampang lintang batang belum memberikan gambaran pola yang
jelas. Pada bagian tengah penampang lintang batang, gambaran pola sudah
terbaca berupa pola 2 dan bentuk pola yang tidak sempurna muncul pada bagian
dalam seperti yang tertera pada Gambar 9.
Gambar 9. (a) M. baccifera bagian tengah (pembesaran 40x) (b) sketsa pola
ikatan pembuluh 2 pada M. baccifera
parenkim
metaxilem
jar. floem
sklerenkim
ruang antar
sel
parenkim
metaxilem
ruang antar sel
sklerenkim
jar. floem
Gambar 8. (a) M. baccifera bagian pangkal dengan pembesaran 40x
(b). Sketsa pola ikatan pembuluh 2 pada M. baccifera
kulit
dalam
kulit
dalam
(a) (b)
(a) (b)

Ujung
Pola ikatan pembuluh pada bagian ujung sama seperti pada bagian pangkal
dan tengah, tetapi ukurannya lebih kecil. Pada bagian tepi penampang lintang
batang, gambaran pola tidak jelas terlihat. Pola ikatan pembuluh terlihat jelas
hanya pada bagian tengah. Bagian dalam disusun oleh pola-pola yang tidak
sempurna (Gambar 10).
Cephalostachyum pergracile
Bambu C.pergracile adalah kelompok bambu simpodial yang berasal dari
India, Nepal, Burma, Thailand, dan China ( Dransfield dan Widjaja 1995).
Bambu ini tumbuh di Kebun Raya Bogor dengan tinggi batang rata-rata 14 m.
Ketebalan dinding batang pada bagian pangkal rata-rata 1.4 cm, sedangkan bagian
ujung adalah 0.6 cm. Tampilan pohon bambu secara utuh dapat diamati pada
Gambar 11. Pembahasan selanjutnya adalah uraian sifat anatomi berdasarkan
bagian batang, yakni pangkal, tengah, dan ujung.
Gambar 10. (a). Penampang lintang M. baccifera bagian ujung (pembesaran 40x),
(b). Sketsa pola ikatan pembuluh 2 pada M. baccifera
(pola 2)
kulit
dalam
parenkim
metaxilem
ruang antar sel
sklerenkim
jar. floem (b)
(a)

Pangkal
Pada bagian tepi penampang lintang batang, rantai pembuluh pusat
dikelilingi oleh selubung serabut sehingga belum dapat ditentukan bentuk
polanya. Pada bagian tengah penampang lintang terlihat perluasan selubung
serabut dekat ruang antar sel sehingga bakal pembentukan pola 2 sudah mulai
terlihat. Demikian pula pada bagian pusat, bentuk pola ikatan hampir sama
dengan bagian tengah, tetapi perluasan selubung serabutnya agak memanjang.
Namun, pada bagian dalam, perluasan selubung serabut pada ruang antar sel tidak
seluas seperti pada bagian tepi, tengah, dan pusat (Gambar 12).
Gambar 12. (a) Tampilan penampang lintang bambu C. pergracile bagian
pangkal l dengan pembesaran 40x, (b) Sketsa 1 pola ikatan
pembuluh 2 pada C. pergracile
Tengah
Pada bagian tepi penampang lintang batang, selubung serabut mengelilingi
seluruh rantai pembuluh pusat. Bagian tengah penampang lintang berbeda dengan
bagian tepi karena perluasan selubung sudah mulai terlihat. Hal yang sama terjadi
pada bagian dalam penampang lintang batang. Dengan demikian, pada bagian
tengah dan bagian dalam, bentuk pola ikatan pembuluh sudah terlihat jelas, yaitu
pola 2 (Gambar 13).
Gambar 11. Bentuk pohon bambu C. pergracile
parenkim
metaxilem
ruang antar sel
sklerenkim
jar. floem
kulit
dalam
(a)
(b)
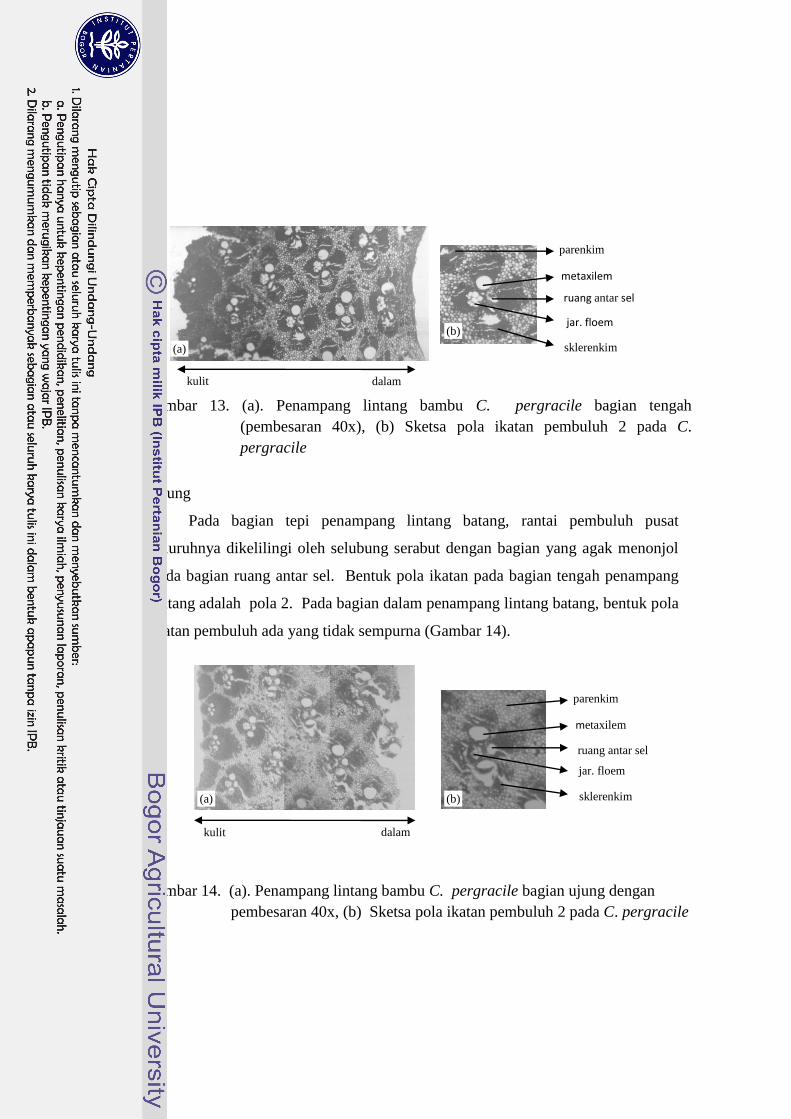
Ujung
Pada bagian tepi penampang lintang batang, rantai pembuluh pusat
seluruhnya dikelilingi oleh selubung serabut dengan bagian yang agak menonjol
pada bagian ruang antar sel. Bentuk pola ikatan pada bagian tengah penampang
lintang adalah pola 2. Pada bagian dalam penampang lintang batang, bentuk pola
ikatan pembuluh ada yang tidak sempurna (Gambar 14).
Gambar 13. (a). Penampang lintang bambu C. pergracile bagian tengah
(pembesaran 40x), (b) Sketsa pola ikatan pembuluh 2 pada C.
pergracile
Gambar 14. (a). Penampang lintang bambu C. pergracile bagian ujung dengan
pembesaran 40x, (b) Sketsa pola ikatan pembuluh 2 pada C. pergracile
parenkim
metaxilem
ruang antar sel
sklerenkim
jar. floem
parenkim
metaxilem
ruang antar sel
sklerenkim
jar. floem
kulit
dalam
kulit
dalam
(a)
(b)
(a) (b)

Pengamatan pada susunan pola ikatan pembuluh pada posisi vertikal, pada
bagian pangkal, jarak antara ikatan pembuluh agak renggang dan selubung
serabut agak memanjang ke arah radial. Pada bagian tengah, selubung serabut
relatif agak pendek. Pada bagian ujung, selubung serabut lebih tipis dengan tetap
menunjukkan kekhasan pola 2-nya.
Dendrocalamus giganteus
D. giganteus adalah bambu simpodial dengan asal yang belum diketahui
secara pasti, tetapi kemungkinan berasal dari Burma dan Thailand (Dransfield dan
Widjaja 1995). Di Kebun Raya Bogor, tinggi batang bambu ini rata-rata 24 m.
Rata-rata tebal dinding batang bagian pangkal adalah 2.4 cm, sedangkan pada
bagian ujung adalah 0.6 cm. Adanya perbedaan yang mencolok dari ketebalan
dinding batang juga terlihat dari perbedaan diameter batang antara bagian pangkal
dan bagian ujung (Gambar 15). Selanjutnya, diuraikan sifat anatomi pada bagian
pangkal, tengah, dan ujung bambu.
Pangkal
Pada bagian tepi penampang lintang batang, sebagian besar sudah terlihat
cikal bakal munculnya pola 3, yaitu dengan adanya rantai serabut, sedangkan pada
bagian tengah penampang lintang sudah memunculkan bentuk pola 3 dengan
bentuk rantai serabut yang membulat. Bentuk pola yang hampir sama juga ada
pada bagian pusat dengan rantai serabut yang lebih pipih dan memanjang. Pada
bagian dalam, bentuk pola ikatan pembuluhnya tidak sempurna.
Gambar 15. Tampilan utuh bambu D. giganteus
rantai serabut
parenkim
metaxilem
ruang antar sel
sklerenkim
jar. floem

Tengah
Pada bagian tepi penampang lintang batang, bentuk pola tidak terlihat karena
pembuluh pusat dan rantai serabut masih menyatu. Pada bagian tengah
penampang lintang, bentuk pola sudah terlihat jelas dengan rantai serabut yang
bulat dan agak runcing pada bagian ujungnya. Pada bagian pusat, bentuk pola
hampir sama dengan tengah penampang lintang, tetapi bentuk rantai serabut
ramping. Bentuk yang hampir sama ditemukan pada bagian dalam, tetapi
dikombinasikan juga dengan adanya pola yang tidak sempurna karena tidak
mengandung rantai serabut (Gambar 17).
Ujung
Bentuk pola pada seluruh bagian penampang lintang bambu bagian ujung
hampir sama dengan bagian tengah dengan ukuran yang lebih kecil. Pada bagian
tepi penampang lintang batang, bentuk pola belum terlihat. Pada bagian tengah,
bentuk pola sudah terlihat, yaitu pola 3. Pada bagian dalam penampang lintang,
bentuk pola tidak sempurna ( Gambar 18).
Gambar 16. (a) Penampang lintang bambu D. giganteus bagian pangkal
(pembesaran 40x), (b) Sketsa pola ikatan pembuluh 3 pada
D. giganteus
Gambar 17. (a) Penampang lintang bambu D. giganteus bagian tengah
(pembesaran 40x), (b) Sketsa pola ikatan pembuluh 3 pada
D. giganteus
rantai serabut
parenkim
metaxilem
ruang antar sel
sklerenkim
jar. floem
rantai serabut
parenkim
metaxilem
ruang antar sel
sklerenkim
jar. floem
kulit
dalam
kulit
dalam
(a) (b)
(a) (b)

Susunan pola pada berbagai posisi vertikal terlihat berbeda. Pada bagian
pangkal bentuk rantai serabut ramping agak melengkung ke arah tangensial.
Sementara pada bagian tengah rantai serabut membulat agak melengkung
walaupun sebagian kecil berbentuk lurus (tidak melengkung). Pada bagian ujung
bentuk rantai serabut membulat dengan ukuran yang lebih kecil dibandingkan
bagian pangkal dan tengah.
Dendrocalamus strictus
Bambu D. strictus termasuk ke dalam bambu simpodial yang berasal dari
negara India, Nepal, Banglades, Burma dan Thailand (Dransfield danWidjaja
1995). Di kebun raya Bogor, bambu ini tumbuh dengan rata-rata tinggi batang
adalah 9 m dengan rata-rata tebal dinding batang bagian pangkal adalah 1.5 cm
(Gambar 19). Pada bagian ujung, penampang lintang batang berbentuk solid
(tidak berongga). Tahapan selanjutnya adalah uraian sifat anatomi pada pangkal,
tengah dan ujung batang.
Pangkal
Gambar 18. (a) Penampang lintang bambu D. giganteus bagian ujung (pembesaran
40x), (b) Sketsa pola ikatan pembuluh 3 pada D. giganteus
Gambar 19. Bentuk pohon bambu D. strictus secara utuh
kulit
dalam
(a) (b)
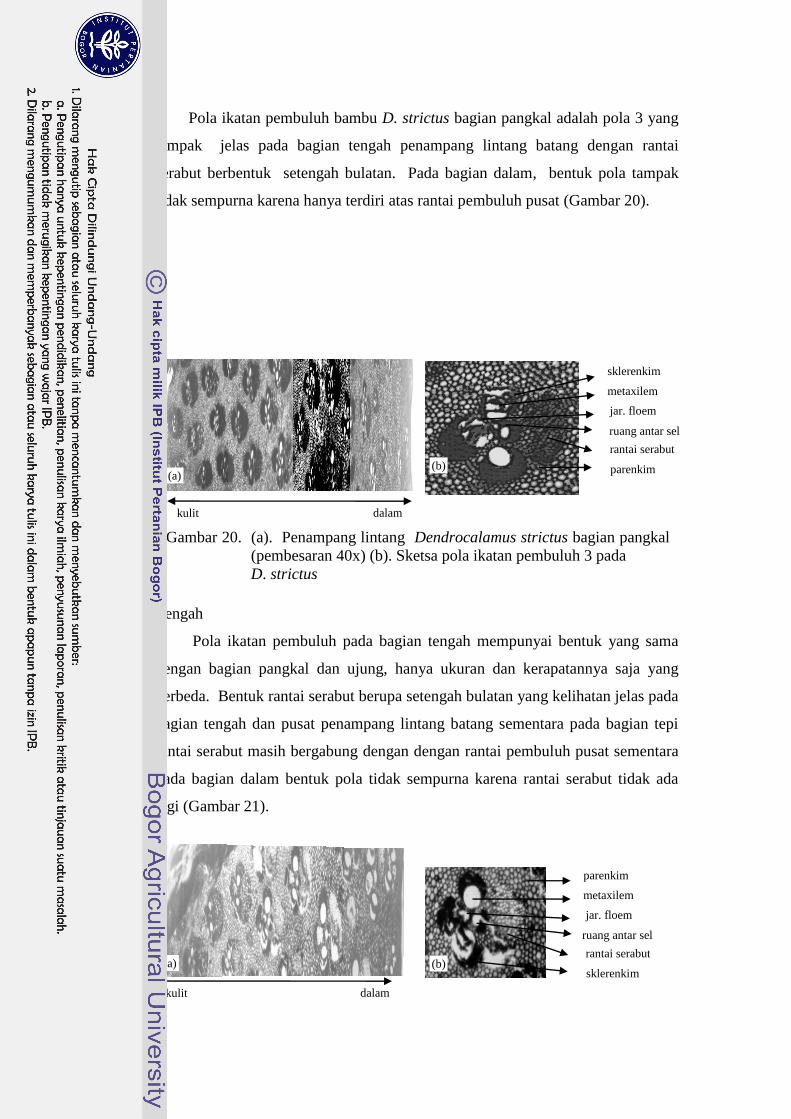
Pola ikatan pembuluh bambu D. strictus bagian pangkal adalah pola 3 yang
tampak jelas pada bagian tengah penampang lintang batang dengan rantai
serabut berbentuk setengah bulatan. Pada bagian dalam, bentuk pola tampak
tidak sempurna karena hanya terdiri atas rantai pembuluh pusat (Gambar 20).
Tengah
Pola ikatan pembuluh pada bagian tengah mempunyai bentuk yang sama
dengan bagian pangkal dan ujung, hanya ukuran dan kerapatannya saja yang
berbeda. Bentuk rantai serabut berupa setengah bulatan yang kelihatan jelas pada
bagian tengah dan pusat penampang lintang batang sementara pada bagian tepi
rantai serabut masih bergabung dengan dengan rantai pembuluh pusat sementara
pada bagian dalam bentuk pola tidak sempurna karena rantai serabut tidak ada
lagi (Gambar 21).
Gambar 20. (a). Penampang lintang Dendrocalamus strictus bagian pangkal
(pembesaran 40x) (b). Sketsa pola ikatan pembuluh 3 pada
D. strictus
rantai serabut
sklerenkim
metaxilem
ruang antar sel
parenkim
jar. floem
rantai serabut
parenkim
metaxilem
ruang antar sel
sklerenkim
jar. floem
kulit
dalam
kulit
dalam
(a) (b)
(a) (b)

Ujung
Bentuk pola ikatan pembuluh pada bagian ujung agak berbeda
dibandingkan dengan bagian pangkal dan tengah karena bentuknya ramping.
Rantai serabut setengah bulatan yang lonjong yang terlihat jelas polanya pada
bagian tengah. Perubahan bentuk pola yang sangat mendadak terlihat antara
bagian tepi dengan tengah yang diduga terjadi karena dinding batang yang sempit
(Gambar 22).
Dendrocalamus asper
Bambu D. asper adalah bambu kelompok simpodial yang berasal dari Jawa
dan Malaysia (Botanic Garden of Indonesia 2001). Tingggi batang bambu rata-
rata 21 m dengan diameter batang pada bagian pangkal sebesar 1.8 cm sedangkan
ketebalan batang pada bagian ujung adalah 0.8 cm. Tampilan bambu D. asper
secara utuh dapat diamati pada Gambar 23. Uraian sifat anatomi pada berbagai
posisi vertikal dijelaskan pada uraian selanjutnya.
Gambar 21. (a) Penampang lintang D. strictus bagian tengah dengan
pembesaran 40x, (b) Sketsa pola ikatan pembuluh 3 pada
D. strictus
Gambar 22. (a) Penampang melintang D. strictus bagian ujung dengan
pembesaran 40x, (b) Sketsa pola ikatan pembuluh 3 pada D.
strictus
rantai serabut
sklerenkim
metaxilem
ruang antar sel
parenkim
jar. floem
kulit
dalam
(a) (b)

Pangkal
Pada bagian tepi penampang lintang batang, bentuk pola ikatan pembuluh
tampak terdiri atas 2 bagian yaitu rantai pembuluh pusat dan rantai serabut.
Ukuran rantai serabut lebih besar dibandingkan rantai pembuluh pusat. Pada
bagian tengah penampang lintang, sudah terlihat bentuk pola yaitu pola 4 dengan
bentuk rantai serabut agak membulat dan tebal. Demikian pula pada bagian pusat
bentuk pola 4 terlihat jelas dengan sedikit perbedaan dalam bentuk rantai
serabutnya yang lebih memanjang. Sementara pada bagian dalam bentuk pola
mirip dengan bagian pusat namun dengan rantai serabut yang memanjang tipis
atau bahkan berbentuk tidak sempurna (Gambar 24).
Tengah
Pada bagian tepi penampang lintang batang, kedua rantai serabut saling
bersambungan sehingga rantai pembuluh pusat seperti berada di tengah-tengah
serabut. Pada bagian tengah penampang lintang batang, bentuk pola ikatan
pembuluh terlihat jelas membentuk pola ikatan pembuluh 4 dan sebagian kecil
muncul pola ikatan pembuluh 3 (Gambar 25). Hal sama juga terlihat pada bagian
pusat penampang lintang, hanya bentuk kedua rantai serabut relatif memanjang
Gambar 23. Bentuk pohon bambu D. asper
Gambar 24. (a) Penampang lintang bambu D. asper bagian pangkal
dengan pembesaran 40x, (b) Sketsa pola ikatan pembuluh 4
rantai serabut
parenkim
metaxilem
ruang antar sel
sklerenkim
jar. floem
kulit
dalam
rantai serabut
parenkim
metaxilem
ruang antar sel
sklerenkim
jar. floem
(a) (b)

dan pada bagian ini terbentuk pola 3 dan 4. Pada bagian dalam bentuk pola ikatan
pembuluh ada yang sempurna dan tidak sempurna.
Ujung
Pada bagian tepi penampang lintang batang, bentuk pola ikatan pembuluh
belum terlihat karena rantai serabut masih menyatu. Sedangkan pada bagian
tengah penampang lintang sudah terlihat pemisahan rantai serabut membentuk
pola 3. Pada bagian pusat bentuk kedua rantai serabut terlihat memanjang dan
menjadi tidak sempurna pada bagian dalam (Gambar 26).
Gambar 25. (a) Penampang lintang D. asper bagian tengah dengan
pembesaran 40x, (b) Sketsa pola ikatan pembuluh 3 pada
D. asper
rantai serabut
sklerenkim
metaxilem
ruang antar sel
parenkim
jar. floem
kulit
dalam
kulit
dalam
rantai serabut
sklerenkim
metaxilem
ruang antar sel
parenkim
jar. floem
(a)
(b)
(a) (b)

Gigantochloa apus
G. apus adalah bambu yang berasal dari Burma dan Thailand dan termasuk
ke dalam bambu simpodial (Dransfield dan Widjaja 1995). Tinggi batang rata-
rata 17 m dengan diameter batang bagian pangkal adalah 7.2 cm. Pada bagian
ujung batang, diameternya adalah 5.2 cm. Ketebalan dinding batang berturut-
turut pada bagian pangkal dan ujung adalah 1 cm dan 0.5 cm (Gambar 27).
Selanjutnya adalah uraian sifat anatomi bambu pada bagian pangkal, tengah, dan
ujung batang.
Pangkal
Pola ikatan pembuluh pada bambu G.apus adalah pola 4 yang kelihatan
jelas pada bagian tengah dan pusat penampang lintang batang (Gambar 28).
Rantai serabut yang muncul mendahului rantai pembuluh pusat berbentuk
setengah bulatan dengan ukuran lebih kecil dibandingkan rantai serabut
pasangannya. Pada bagian tepi penampang lintang, kedua rantai serabut masih
menyatu dengan rantai pembuluh pusat sehingga bentuk pola tidak terlihat jelas.
Hal yang agak berbeda terlihat di bagian dalam yaitu bentuk pola tampak tidak
sempurna karena ada sebagian rantai serabut yang hilang.
Gambar 26. Penampang lintang D. asper bagian ujung dengan pembesaran
40x, (b) Sketsa 1 pola ikatan pembuluh 3 pada D. asper
rantai serabut
parenkim
metaxilem
ruang antar sel
sklerenkim
jar. floem
Gambar 27. Tampilan utuh bambu G. apus

Tengah
Pola ikatan pembuluh pada bagian tengah batang adalah pola 4 yang jelas
terlihat pada bagian tengah dan pusat penampang lintang batang. Pada bagian tepi
penampang lintang, rantai serabut sebagian masih bergabung dengan rantai
pembuluh pusat sedangkan yang lainnya terlihat sudah terpisah. Dua rantai
serabut yang bergabung dengan rantai pembuluh pusat memiliki ukuran yang
berbeda (Gambar 29).
Ujung
Pola ikatan pembuluh pada bagian ujung batang adalah pola 3. Rantai
serabut pada bagian ujung berbentuk memanjang ramping di bagian pusat
penampang lintang dan agak membulat pada bagian tengah penampang lintang.
Bentuk pola yang lengkap juga sebagian muncul pada bagian dalam. Tampilan
lengkap tersaji pada Gambar 30.
Gambar 28. (a). Penampang lintang G. apus bagian pangkal (pembesaran 40x),
(b) Sketsa pola ikatan pembuluh 4 pada G. apus
Gambar 29. (a) Tampilan penampang lintang G. apus bagian tengah
(pembesaran 40x, (b) Sketsa pola ikatan pembuluh 4 pada G. apus
rantai serabut
parenkim
metaxilem
ruang antar sel
sklerenkim
jar. floem
rantai serabut
parenkim
metaxilem
ruang antar sel
sklerenkim
jar. floem
kulit
dalam
kulit
dalam
(a) (b)
(a) (b)

Gigantochloa atroviolacea
G. atrovilacea adalah bambu yang berasal dari negara Indonesia dan
tumbuh secara simpodial (Dransfield dan Widjaja 1995). Bambu ini tumbuh di
daerah Dermaga, Bogor (Fakultas Kehutanan) dengan tinggi batang rata-rata 19
m. Diameter batang bagian pangkal adalah 6.2 cm dengan ketebalan dinding
batang 1.1 cm. Diameter batang bagian ujung adalah 5.1 cm dengan ketebalan
dinding batang sebesar 0.6 cm. Gambaran lengkap bambu G. atroviolacea dapat
diamati pada Gambar 31. Adapun uraian sifat anatomi batang pada bagian
pangkal, tengah dan ujung diuraikan dalam tulisan selanjutnya.
Pangkal
Pola ikatan pembuluh bambu Gigantochloa atroviolacea adalah pola 4
yang terlihat sangat jelas pada bagian tengah dan pusat penampang lintang
batang. Rantai serabut pada bagian tepi penampang lintang masih bergabung
dengan rantai pembuluh pusat sehingga bentuk pola belum terlihat. Pada bagian
dalam sebagian pola tersusun sempurna dan sebagian lagi tidak sempurna.
Bentuk rantai serabut sangat unik dibandingkan dengan pola pada spesies bambu
lain yaitu seperti bulan sabit yang melengkung tajam. Memang ada 2 rantai
serabut yang menyusun pola tetapi ukuran salah satu rantai serabut yang satu
Gambar 31. Bentuk pohon bambu G. atroviolecea
Gambar 30. (a) Penampang lintang G. apus bagian ujung ( pembesaran 40x),
(b) Sketsa pola ikatan pembuluh 3 pada G. apus
kulit
dalam
(a) (b)

lebih kecil dibanding pasangannya (Gambar 32). Sifat anatomi batang bagian
pangkal, tengah, dan ujung batang akan diuraikan pada penjelasan selanjutnya.
Tengah
Pada posisi ini, bambu G. atroviolacea memiliki pola 4 yang terlihat sangat
jelas baik pada bagian tengah maupun pusat penampang lintang bambu (Gambar
33). Rantai serabut membentuk setengah bulatan tipis yang melengkung tajam.
Ukuran rantai serabut yang membentuk satu pola ikatan tidak sama. Pola yang
sangat jelas terlihat pada penampang lintang bagian tengah dan pusat sementara
pola ikatan pembuluh pada bagian dalam tidak sempurna.
Ujung
Bagian ujung batang bambu G. atroviolacea memiliki pola 3. Pada bagian
ujung ini rantai serabut berbentuk setengah bulatan yang lebih tebal dibandingkan
dengan bagian pangkal dan tengah. Dinding batang yang relatif tipis membagi
penampang lintang batang hanya terdiri atas tepi, tengah dan dalam (Gambar 34).
Gambar 32. (a). Penampang lintang G. atroviolacea bagian pangkal (pembesaran
40x), (b) Sketsa pola ikatan pembuluh 4 pada G. atroviolacea
rantai serabut
parenkim
metaxilem
jar. floem
sklerenkim
ruang antar sel
Gambar 33. (a) Penampang lintang bambu G. atroviolacea bagian tengah
(pembesaran 40x), (b) Sketsa pola ikatan pembuluh 4 pada
G. atroviolacea
ruang antar sel
parenkim
metaxilem
jar. floem
sklerenkim
rantai serabut
rantai serabut
parenkim
metaxilem
ruang antar sel
sklerenkim
jar. floem
kulit
dalam
kulit
dalam
(a) (b)
(a)
(b)

Hasil penelitian peneliti sebelumnya menyatakan bahwa genus Arundinaria
memiliki pola 1 (Grosser dan Liese 1971). Dalam hal ini khusus untuk bambu A.
hundsii dan A. javonica, pembuatan contoh uji dan pengamatan tidak dilakukan
pada bagian pangkal dan ujung. Hal ini terjadi karena ukuran bambu sangat kecil
(diameter 1-2 cm) sedangkan penelitian yang dilakukan membutuhkan bahan
yang sangat banyak karena selain untuk bahan penelitian anatomi juga diperlukan
juga sebagai bahan dalam penelitian kimia, fisik, dan mekanik. Demikian pula
dinyatakan bahwa bambu genus Cephalostachyum hanya memiliki satu pola,
yaitu pola 2 untuk semua bagian batang (pangkal, tengah, dan ujung) sedangkan
untuk genus Melocanna pada ruas bawah terdapat gabungan pola 2 dan 3. Pada
Dendrocalamus dan Gigantochloa pun pada ruas bawah umumnya merupakan
gabungan antara pola 3 dan 4.
Penelitian yang telah dilakukan memberikan hasil yang berbeda seperti
yang ditunjukkan oleh spesies Gigantochloa apus dan G atroviolacea. Pada
kedua bambu ini pada ruas bawah tidak terdapat pola gabungan, namun pada D.
asper khususnya pada ruas bawah murni terdiri atas pola 4 sedangkan pola
gabungan hanya ada pada bagian tengah yaitu gabungan pola 3 dan 4 yang
didominasi oleh pola 3. Sebagai pembanding penelitian ini, sketsa 4 macam pola
ikatan pembuluh hasil penelitian Grosser dan Liese (1971) disajikan pada Gambar
35 dan 36.
metaxilem
Selubung
sklerenkim
Selubung pada
ruang antar sel
Gambar 34. (a) Penampang lintang G. atroviolacea bagian ujung
(pembesaran 40x, (b) Sketsa pola ikatan pembuluh 3 pada
G. atroviolacea
kulit
dalam
(a) (b)

Sumber : Grosser dan Liese (1971)
Gambar 35. Sketsa pola ikatan pembuluh 1 (a) dan 2 (b)
Sumber : Grosser dan Liese (1971)
Gambar 36. Sketsa pola ikatan pembuluh 3 (c) dan 4 (d)
Penjelasan dari sketsa pola ikatan pembuluh menurut Grosser and Liese
(1971) bahwa pola 1 tersusun atas rantai pembuluh pusat yang terdiri atas
metaxilem, floem, dan ruang antar sel sedangkan pada pola 2 hanya ada tambahan
berupa selubung pada ruang antar sel yang lebih besar dibanding selubung yang
lain. Sementara pola 3 selain memiliki rantai pembuluh pusat juga didukung oleh
1 rantai serabut dan pola 4 memiliki susunan yang hampir sama dengan pola 3 ,
yaitu hanya memiliki 2 rantai serabut.
Hasil penelitian menunjukkan bahwa pola ikatan pembuluh pada bambu A.
javonica dan A. hundsii yang sama-sama memiliki pola 1 ternyata memiliki
beberapa perbedaan (Gambar 37). Perubahan bentuk pola secara bertahap terjadi
pada bambu A. javonica sedangkan pada bambu A. hundsii perubahan bentuk pola
Rantai
serabut
Selubung
sklerenkim
metaxilem
(a) (b)
(c) (d)

tidak secara bertahap. Hal ini diduga karena adanya perbedaan ketebalan dinding
batang. Bambu A. javonica memiliki dinding batang tipis dan secara umum
dinding batang hanya terbagi atas 3 bagian saja yaitu bagian tepi, tengah, dan
bagian dalam. Bentuk pola ikatan pembuluh antara bagian tengah dan dalam
hampir sama tetapi ada sedikit perbedaan dalam selubung sklerenkim dimana
pada bagian dalam ukurannya lebih kecil. Bentuk pola terlihat jelas pada bagian
tengah yaitu membentuk pola 1.
Gambar 37. (a) Pola ikatan pembuluh pada A. javonica (pembesaran 40x),
(a1) Sketsa pola ikatan pembuluh A. javonica, (b) Pola ikatan
pembuluh A. hundsii (pembesaran 40x), (b1) Sketsa pola
ikatan pembuluh 1 pada A. hundsii.
Bambu A. hundsii juga memiliki dinding tipis sehingga penampang batang
terbagi atas bagian tepi, tengah, dan bagian dalam. Bentuk pola 1 yang sudah
jelas tampak pada bagian dalam. Hal ini dimungkinkan karena bambu A. hundsii
memiliki dinding batang tipis sehingga perubahan bentuk pola yang cukup jelas
baru muncul di bagian dalam batang.
Meta
xilem
Meta
xilem
Selubung
sklerenkim
Selubung
sklerenkim
(a)
(b)
(a1)
(b1)

Bambu C. pergracile dan M. baccifera memiliki pola 2 baik pada bagian
pangkal, tengah maupun ujung. Antara kedua bambu tersebut selain berbeda
dalam ketebalan dinding sel di mana bambu C.pergracile lebih tebal dibandingkan
dengan bambu M. baccifera juga selubung sklerenkim pada C. pergracile lebih
lebar baik yang mengelilingi metaxilem maupun yang mengelilingi ruang antar
sel. Untuk lebih jelasnya Gambar 38 menampilkan pola ikatan pembuluh 2
bagian tengah.
Gambar 38. Pola ikatan pembuluh pada (a) C. pergracil (pembesaran 40x), (a1)
Sketsa pola ikatan pembuluh 2 pada C. pergracile, (b) M. baccifera
dengan pembesaran 40x, (b1) Sketsa pola ikatan pembuluh 2 pada
M. baccifera
Bambu-bambu yang memiliki pola 3 dalam penelitian ini adalah D. strictus
dan D. giganteus. Bentuk pola bagian pangkal yang dimiliki oleh D. strictus
lebih membulat dan berukuran lebih kecil tapi pada D. giganteus bentuk rantai
serabutnya memanjang. Demikian pula bentuk rantai serabut kedua bambu pada
Selubung
sklerenkim
Selubung sklerenkim
pada ruang antar sel
Selubung
sklerenkim
(a)
(b)
(a1)
)
(b1)
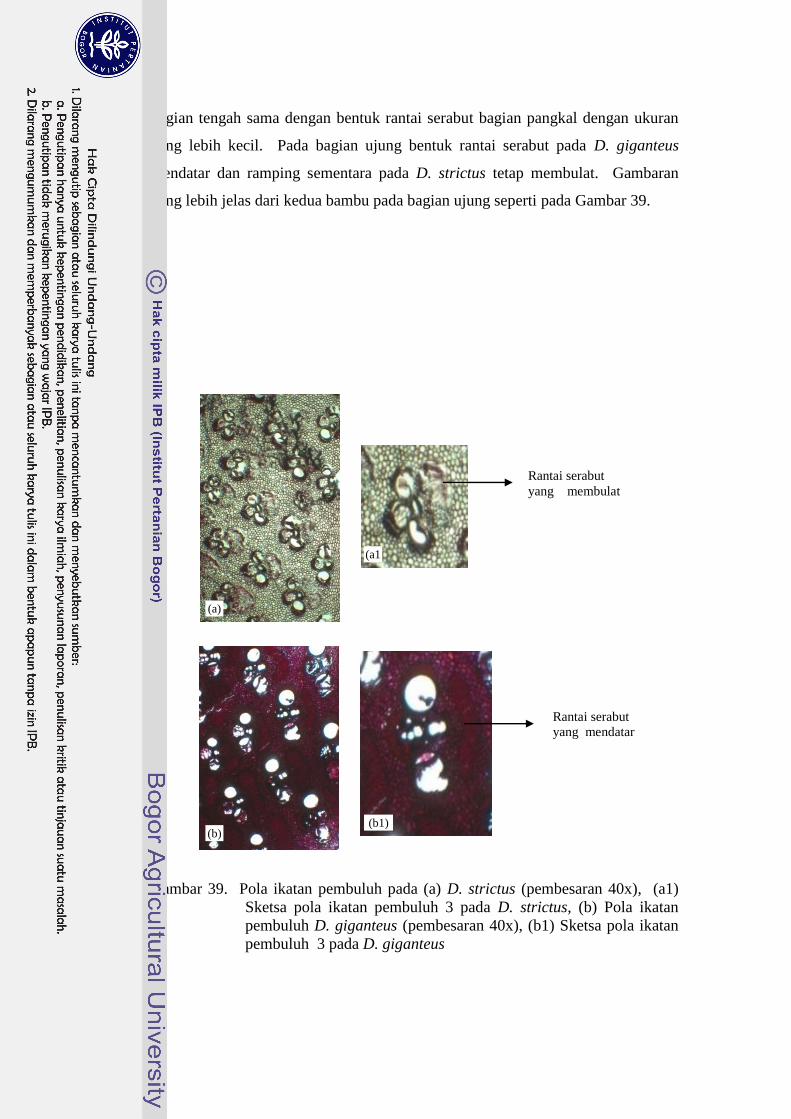
bagian tengah sama dengan bentuk rantai serabut bagian pangkal dengan ukuran
yang lebih kecil. Pada bagian ujung bentuk rantai serabut pada D. giganteus
mendatar dan ramping sementara pada D. strictus tetap membulat. Gambaran
yang lebih jelas dari kedua bambu pada bagian ujung seperti pada Gambar 39.
Gambar 39. Pola ikatan pembuluh pada (a) D. strictus (pembesaran 40x), (a1)
Sketsa pola ikatan pembuluh 3 pada D. strictus, (b) Pola ikatan
pembuluh D. giganteus (pembesaran 40x), (b1) Sketsa pola ikatan
pembuluh 3 pada D. giganteus
Rantai serabut
yang membulat
Rantai serabut
yang mendatar
(a)
(b)
(a1
)
(b1)

Beberapa jenis bambu ternyata dalam tampilan batangnya mempunyai 2
jenis pola. Perbedaan pola antar bagian batang pada tingkatan jenis/spesies
memang tidak diungkapkan dalam penelitian Grosser dan Liese (1971) tetapi
dinyatakan bahwa perbedaan pola dalam satu batang bambu jenis tertentu
kemungkinan ada terutama dengan munculnya pola gabungan. Dalam penelitian
ini ternyata ditemukan ada 3 jenis bambu yang memiliki perbedaan pola antar
bagian batangnya, misal untuk bambu G. apus dan G. atroviolacea memiliki tren
pola yang sama yaitu pada bagian pangkal dan tengah mempunyai pola 4
sedangkan pada bagian ujung mempunyai pola 3. Hal ini mudah untuk
dimengerti karena pola 4 merupakan pola yang memerlukan ruangan yang relatif
lebih luas sehingga muncul hanya pada bagian batang dengan dinding yang relatif
lebih tebal sedangkan pola 3 merupakan pola yang lebih sederhana dan
memerlukan ruangan lebih sempit sehingga memungkinkan muncul pada bagian
ujung. Hal yang sama berlaku pula untuk bambu D. asper namun perbedaan pola
terjadi antar bagian pangkal dengan bagian tengah/ujung yaitu pola 4 berada pada
bagian pangkal sedangkan bagian tengah dan ujung memiliki pola 3.
Bambu D. asper terutama pada bagian pangkal dan tengah memiliki bentuk
rantai serabut yang lancip hampir seperti setengah lingkaran dengan jarak antar
pola yang relatif renggang jika dibandingkan dengan pola ikatan pembuluh pada
bambu G atroviolacea dan G. apus (Gambar 40). Pada G. atroviolacea bentuk
rantai serabut lancip tajam dan jarak antar pola relatif rapat sementara pada G.
apus bentuk rantai serabut membulat dan kecil dengan jarak antar pola relatif
renggang. Pada bagian tengah batang bentuk rantai serabut sama dengan bagian
pangkal dengan ukuran yang lebih kecil. Bentuk rantai serabut bagian ujung dari
bambu D. asper adalah membulat ramping sampai datar, juga terdapat pada G.
atroviolacea tetapi bentuk yang agak berbeda dimiliki oleh bambu G. apus
dengan bentuk rantai serabut mulai dari yang mendatar hingga membulat dengan
salah satu sisi melengkung atau meruncing.
Rantai serabut
membulat ramping
Rantai serabut mendatar
Rantai serabut
membulat Rantai serabut
membulat dengan satu
sisi melengkung
(a) (b) (c)
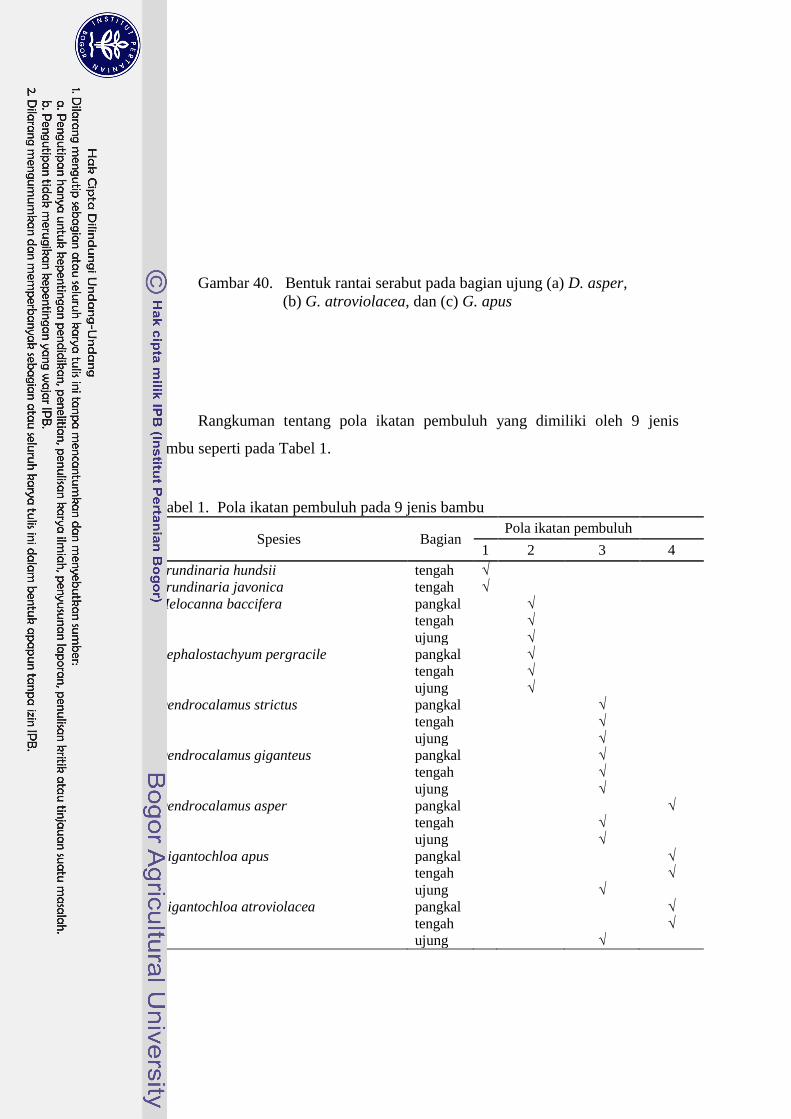
Gambar 40. Bentuk rantai serabut pada bagian ujung (a) D. asper,
(b) G. atroviolacea, dan (c) G. apus
Rangkuman tentang pola ikatan pembuluh yang dimiliki oleh 9 jenis
bambu seperti pada Tabel 1.
Tabel 1. Pola ikatan pembuluh pada 9 jenis bambu
Spesies Bagian Pola ikatan pembuluh
1 2 3 4
Arundinaria hundsii tengah √
Arundinaria javonica tengah √
Melocanna baccifera pangkal
√
tengah
√
ujung
√
Cephalostachyum pergracile pangkal
√
tengah
√
ujung
√
Dendrocalamus strictus pangkal
√
tengah
√
ujung
√
Dendrocalamus giganteus pangkal
√
tengah
√
ujung
√
Dendrocalamus asper pangkal
√
tengah
√
ujung
√
Gigantochloa apus pangkal
√
tengah
√
ujung
√
Gigantochloa atroviolacea pangkal
√
tengah
√
ujung √

Kerapatan ikatan pembuluh
Nilai kerapatan ikatan pembuluh pada penampang lintang batang 8 jenis
bambu dapat diamati pada Lampiran 2. Analisis keragaman terhadap nilai
kerapatan ikatan pembuluh sebagai variabel y dan faktor jenis, posisi vertikal,
posisi horizontal, dan pola sebagai variabel x menghasilkan persamaan regresi (1)
bersifat nyata (Lampiran 3).
Dari berbagai variabel yang diujikan ternyata yang mempengaruhi nilai
kerapatan adalah semua faktor yang terdiri atas faktor jenis, posisi vertikal, posisi
horizontal dan pola. Apabila dari berbagai faktor tersebut dibandingkan nilai
signifikannya maka faktor pola menduduki urutan pertama dalam tingkat
kesignifikannya. Hal ini berarti pola paling mempengaruhi nilai kerapatan
bambu. Uji lanjutan Duncan untuk melihat jenis-jenis bambu yang berpengaruh
nyata terlihat bahwa jenis bambu A. hundsii (Ah), G. apus (Ga), A. javonica (Aj),
D. asper (Da), M. baccifera (Mb), C. pergracile (Cp), dan D. giganteus (Dg)
memiliki nilai kerapatan ikatan pembuluh yang sama sementara antara jenis-jenis
bambu yang sudah diuraikan sebelumnya dengan bambu D. strictus memiliki nilai
kerapatan ikatan pembuluh yang berbeda nyata. Hal ini terjadi karena kerapatan
ikatan pembuluh bambu D. strictus bagian ujung batang terutama pada bagian tepi
penampang lintang relatif paling tinggi jika dibandingkan dengan bambu-bambu
lain (Lampiran 2). Nilai kerapatan ikatan pembuluh untuk 8 jenis bambu
tercantum pada Gambar 41.
Y = 3.4 + 0.4X2 + 0.6X3 + 0.6X4 - 2.7X5 +2.1X6 + 1.6X7 + 1.1X8 + 0.5X9 + 0.5X10
- 0.4X11 - 1.2X12 + 0.9X13 - 1.2X14 - 2.4X15…………………………………(1)
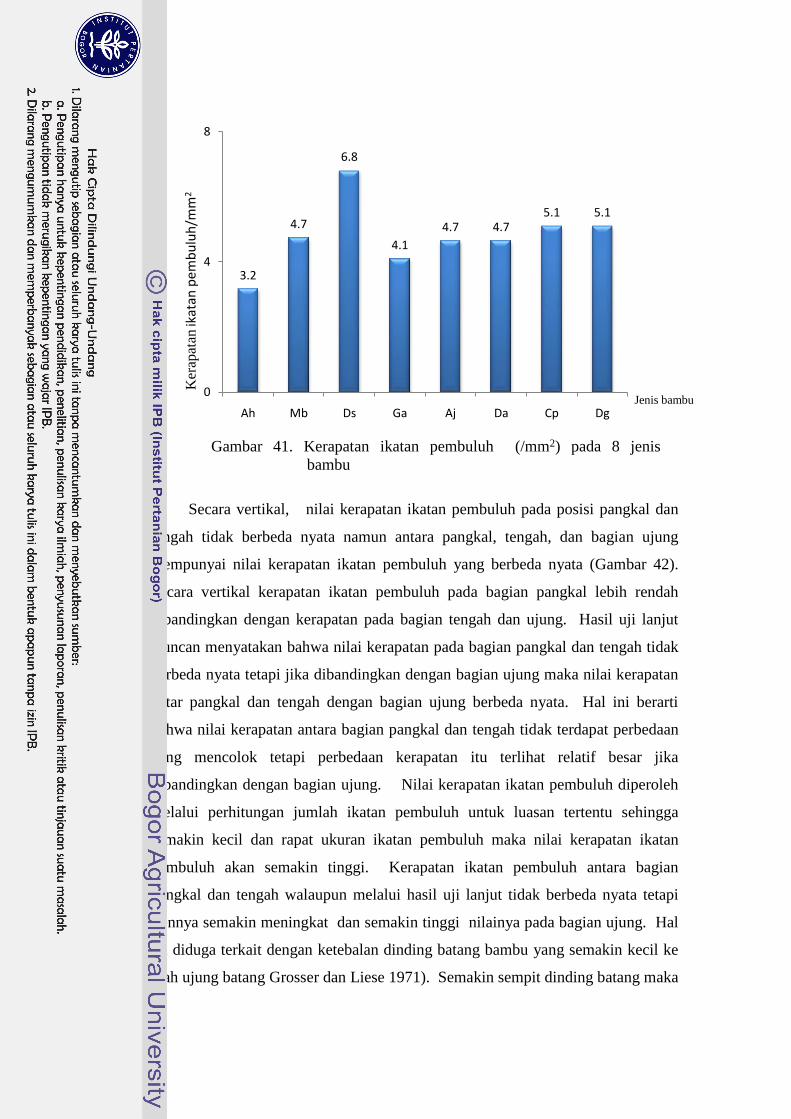
Secara vertikal, nilai kerapatan ikatan pembuluh pada posisi pangkal dan
tengah tidak berbeda nyata namun antara pangkal, tengah, dan bagian ujung
mempunyai nilai kerapatan ikatan pembuluh yang berbeda nyata (Gambar 42).
Secara vertikal kerapatan ikatan pembuluh pada bagian pangkal lebih rendah
dibandingkan dengan kerapatan pada bagian tengah dan ujung. Hasil uji lanjut
Duncan menyatakan bahwa nilai kerapatan pada bagian pangkal dan tengah tidak
berbeda nyata tetapi jika dibandingkan dengan bagian ujung maka nilai kerapatan
antar pangkal dan tengah dengan bagian ujung berbeda nyata. Hal ini berarti
bahwa nilai kerapatan antara bagian pangkal dan tengah tidak terdapat perbedaan
yang mencolok tetapi perbedaan kerapatan itu terlihat relatif besar jika
dibandingkan dengan bagian ujung. Nilai kerapatan ikatan pembuluh diperoleh
melalui perhitungan jumlah ikatan pembuluh untuk luasan tertentu sehingga
semakin kecil dan rapat ukuran ikatan pembuluh maka nilai kerapatan ikatan
pembuluh akan semakin tinggi. Kerapatan ikatan pembuluh antara bagian
pangkal dan tengah walaupun melalui hasil uji lanjut tidak berbeda nyata tetapi
trennya semakin meningkat dan semakin tinggi nilainya pada bagian ujung. Hal
ini diduga terkait dengan ketebalan dinding batang bambu yang semakin kecil ke
arah ujung batang Grosser dan Liese 1971). Semakin sempit dinding batang maka
3.2
4.7
6.8
4.1
4.7 4.75.1 5.1
0
4
8
Ah Mb Ds Ga Aj Da Cp Dg
Gambar 41. Kerapatan ikatan pembuluh (/mm2) pada 8 jenis
bambu
Jenis bambu
Ker
apat
anik
atan
pem
bu
luh
/mm
2
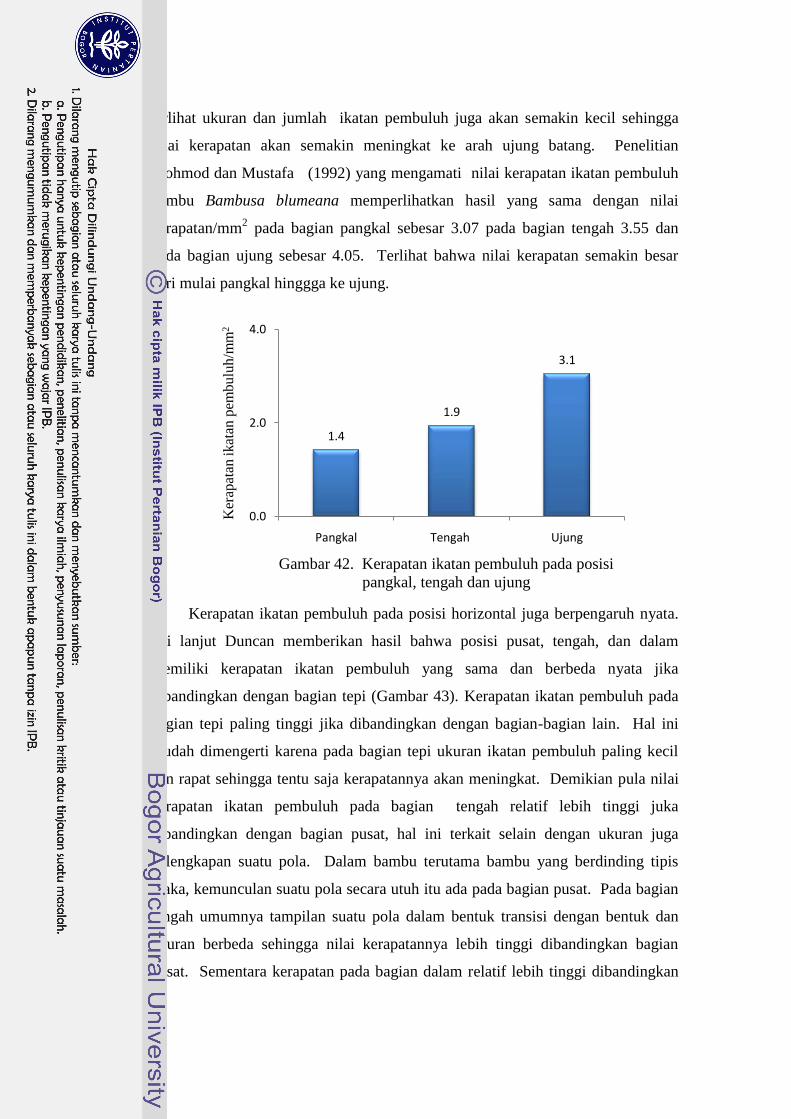
terlihat ukuran dan jumlah ikatan pembuluh juga akan semakin kecil sehingga
nilai kerapatan akan semakin meningkat ke arah ujung batang. Penelitian
Mohmod dan Mustafa (1992) yang mengamati nilai kerapatan ikatan pembuluh
bambu Bambusa blumeana memperlihatkan hasil yang sama dengan nilai
kerapatan/mm2 pada bagian pangkal sebesar 3.07 pada bagian tengah 3.55 dan
pada bagian ujung sebesar 4.05. Terlihat bahwa nilai kerapatan semakin besar
dari mulai pangkal hinggga ke ujung.
Kerapatan ikatan pembuluh pada posisi horizontal juga berpengaruh nyata.
Uji lanjut Duncan memberikan hasil bahwa posisi pusat, tengah, dan dalam
memiliki kerapatan ikatan pembuluh yang sama dan berbeda nyata jika
dibandingkan dengan bagian tepi (Gambar 43). Kerapatan ikatan pembuluh pada
bagian tepi paling tinggi jika dibandingkan dengan bagian-bagian lain. Hal ini
mudah dimengerti karena pada bagian tepi ukuran ikatan pembuluh paling kecil
dan rapat sehingga tentu saja kerapatannya akan meningkat. Demikian pula nilai
kerapatan ikatan pembuluh pada bagian tengah relatif lebih tinggi juka
dibandingkan dengan bagian pusat, hal ini terkait selain dengan ukuran juga
kelengkapan suatu pola. Dalam bambu terutama bambu yang berdinding tipis
maka, kemunculan suatu pola secara utuh itu ada pada bagian pusat. Pada bagian
tengah umumnya tampilan suatu pola dalam bentuk transisi dengan bentuk dan
ukuran berbeda sehingga nilai kerapatannya lebih tinggi dibandingkan bagian
pusat. Sementara kerapatan pada bagian dalam relatif lebih tinggi dibandingkan
1.4
1.9
3.1
0.0
2.0
4.0
Pangkal Tengah Ujung
Gambar 42. Kerapatan ikatan pembuluh pada posisi
pangkal, tengah dan ujung
Ker
apat
an ikat
an p
emb
ulu
h/m
m2
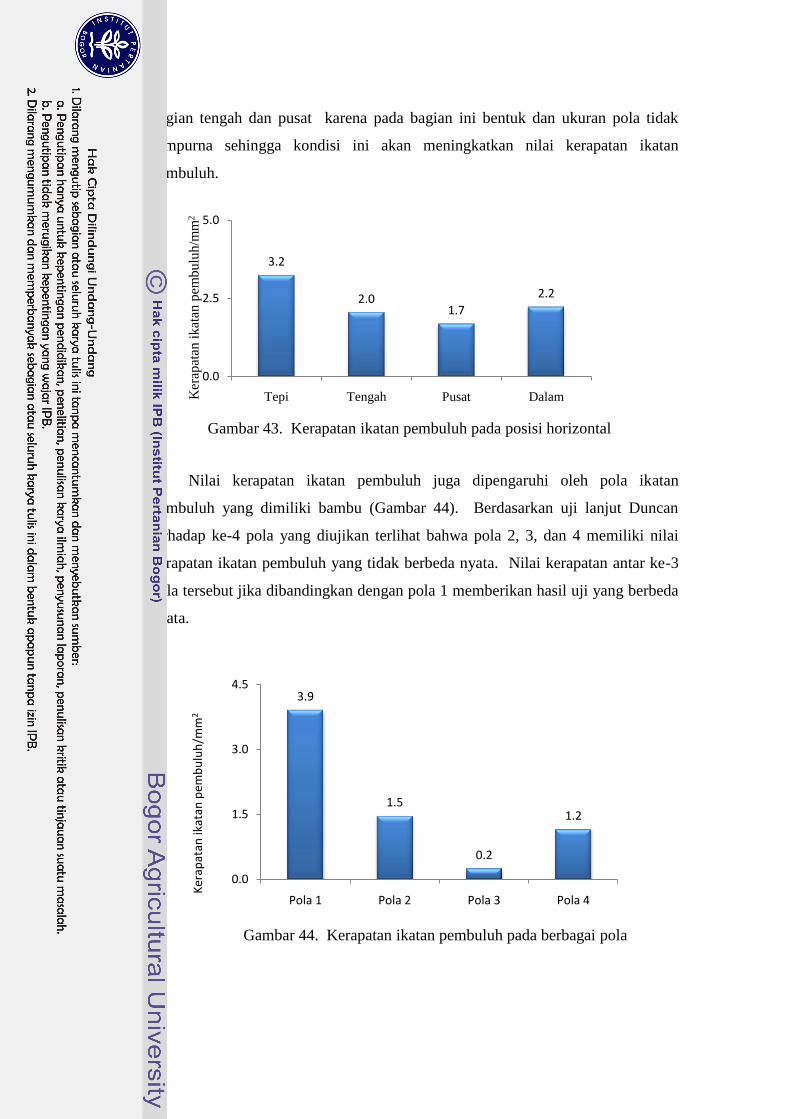
bagian tengah dan pusat karena pada bagian ini bentuk dan ukuran pola tidak
sempurna sehingga kondisi ini akan meningkatkan nilai kerapatan ikatan
pembuluh.
Nilai kerapatan ikatan pembuluh juga dipengaruhi oleh pola ikatan
pembuluh yang dimiliki bambu (Gambar 44). Berdasarkan uji lanjut Duncan
terhadap ke-4 pola yang diujikan terlihat bahwa pola 2, 3, dan 4 memiliki nilai
kerapatan ikatan pembuluh yang tidak berbeda nyata. Nilai kerapatan antar ke-3
pola tersebut jika dibandingkan dengan pola 1 memberikan hasil uji yang berbeda
nyata.
3.2
2.01.7
2.2
0.0
2.5
5.0
Tepi Tengah Pusat Dalam
Gambar 43. Kerapatan ikatan pembuluh pada posisi horizontal
Ker
apat
an i
kat
an p
emb
ulu
h/m
m2
3.9
1.5
0.2
1.2
0.0
1.5
3.0
4.5
Pola 1 Pola 2 Pola 3 Pola 4
Gambar 44. Kerapatan ikatan pembuluh pada berbagai pola
Ker
apat
an ik
atan
pem
bu
luh
/mm
2

Pola 1 memiliki nilai kerapatan yang paling tinggi dibandingkan dengan
pola-pola lain, demikian juga pola 2 memiliki nilai kerapatan ikatan pembuluh
lebih tinggi dibandingkan pola 3 dan 4. Pola 3 memiliki nilai yang terendah
dibandingkan dengan pola-pola lain. Pola 4 mempunyai kerapatan yang
cenderung lebih tinggi jika dibandingkan pola 3. Pengamatan pada ukuran ikatan
pembuluh yang dimiliki oleh bambu dengan pola 1 ternyata ukuran ikatan pada
pola 1 paling kecil dibandingkan dengan pola-pola lain, demikian pula ukuran
pola ikatan pembuluh pada pola 2 lebih kecil dibandingkan dengan pola 3 dan
pola 4. Bentuk ikatan pembuluh antar pola 1 dan 2 tidak terlalu berbeda yaitu
sama-sama terdiri atas rantai pembuluh pusat dan didukung jaringan selubung
sklerenkim, hanya memang untuk pola 2 ada tambahannya yang berbentuk
selubung sklerenkim pada ruang interseluler. Demikian juga bentuk ikatan
pembuluh antara pola 3 dan 4 menunjukkan adanya kesamaan yaitu keduanya
terdiri atas selain rantai pembuluh pusat juga hadirnya rantai serabut, hanya untuk
pola 4 jumlah rantai serabut adalah 2 buah. Dengan demikian jumlah ikatan
pembuluh/satuan luas paling tinggi pada pola 1 karena ukurannya paling kecil,
demikian juga dengan ukuran ikatan pembuluh pada pola 4 paling besar
dibandingkan dengan pola-pola lain sehingga seharusnya memiliki nilai kerapatan
yang paling rendah. Akan tetapi, dalam uji ini ternyata nilai kerapatan pola 3
lebih rendah dibandingkan dengan nilai kerapatan ikatan pembuluh pada pola 4,
hal ini diduga karena jarak antar ikatan pembuluh yang relatif renggang
dibandingkan pada pola 4. Dalam penelitian ini jarak antar ikatan tidak diukur.
Dalam pengamatan visual terhadap pola 3 dan 4 terlihat bahwa jarak antar pola
relatif jarang dibandingkan dengan pola 1 dan 2. Diduga kondisi ini turut
mempengaruhi nilai kerapatan ikatan pembuluh yang relatif kecil. .
Hasil penelitian yang telah dilakukan oleh Mohmod & Mustafa (1992)
terhadap Bambusa blumeana dan B vulgaris serta Gigantochloa scortechinii
dengan rata-rata kerapatan ikatan pembuluh /mm2 adalah berturut-turut 2.9, 2.5
serta 1.8 . Sedangkan hasil penelitian Wahab et al. (2009) pada bambu Bambusa
vulgaris ternyata nilai rata-rata kerapatan ikatan pembuluh/mm2 adalah 2.6 ± 0.19.
Sementara nilai kerapatan ikatan pembuluh dalam penelitian ini adalah 0.2/mm2
untuk pola 3 dan 1.2/mm2 untuk pola 4.
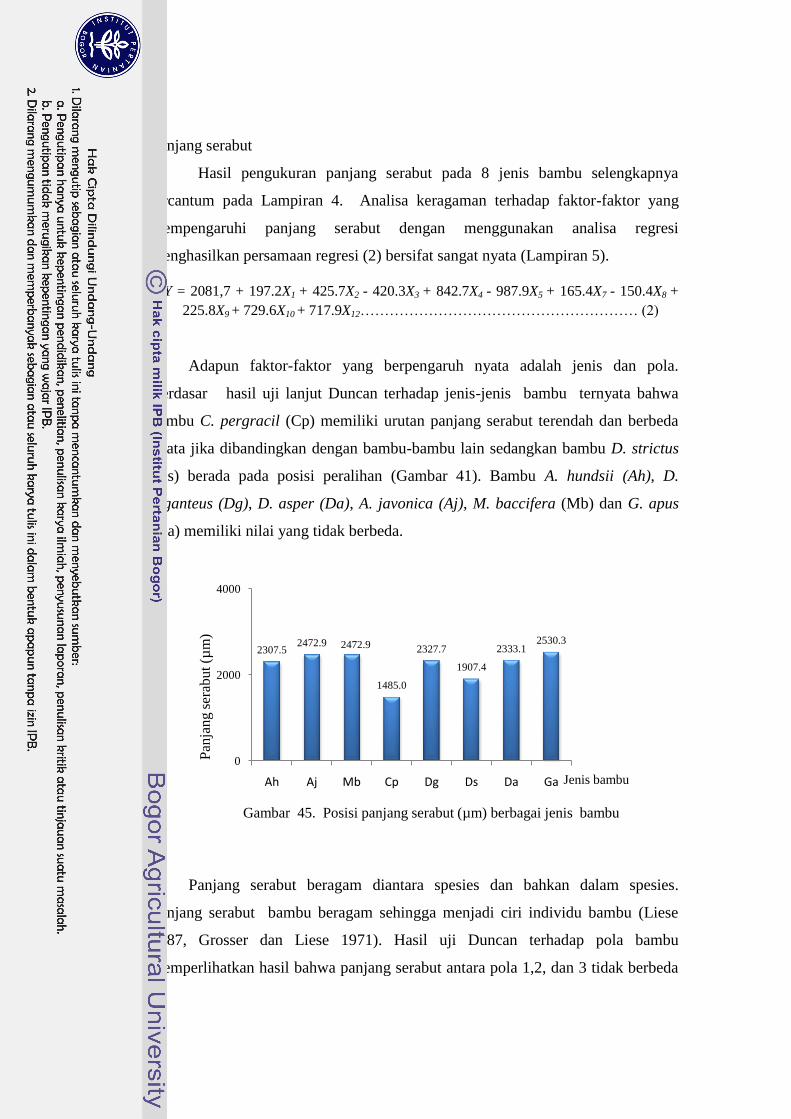
Panjang serabut
Hasil pengukuran panjang serabut pada 8 jenis bambu selengkapnya
tercantum pada Lampiran 4. Analisa keragaman terhadap faktor-faktor yang
mempengaruhi panjang serabut dengan menggunakan analisa regresi
menghasilkan persamaan regresi (2) bersifat sangat nyata (Lampiran 5).
Adapun faktor-faktor yang berpengaruh nyata adalah jenis dan pola.
Berdasar hasil uji lanjut Duncan terhadap jenis-jenis bambu ternyata bahwa
bambu C. pergracil (Cp) memiliki urutan panjang serabut terendah dan berbeda
nyata jika dibandingkan dengan bambu-bambu lain sedangkan bambu D. strictus
(Ds) berada pada posisi peralihan (Gambar 41). Bambu A. hundsii (Ah), D.
giganteus (Dg), D. asper (Da), A. javonica (Aj), M. baccifera (Mb) dan G. apus
(Ga) memiliki nilai yang tidak berbeda.
Panjang serabut beragam diantara spesies dan bahkan dalam spesies.
Panjang serabut bambu beragam sehingga menjadi ciri individu bambu (Liese
1987, Grosser dan Liese 1971). Hasil uji Duncan terhadap pola bambu
memperlihatkan hasil bahwa panjang serabut antara pola 1,2, dan 3 tidak berbeda
Y = 2081,7 + 197.2X1 + 425.7X2 - 420.3X3 + 842.7X4 - 987.9X5 + 165.4X7 - 150.4X8 +
225.8X9 + 729.6X10 + 717.9X12………………………………………………… (2)
2307.52472.9 2472.9
1485.0
2327.7
1907.4
2333.12530.3
0
2000
4000
Ah Aj Mb Cp Dg Ds Da Ga
Gambar 45. Posisi panjang serabut (µm) berbagai jenis bambu
Jenis bambu
Pan
jang s
erab
ut
(µm
)
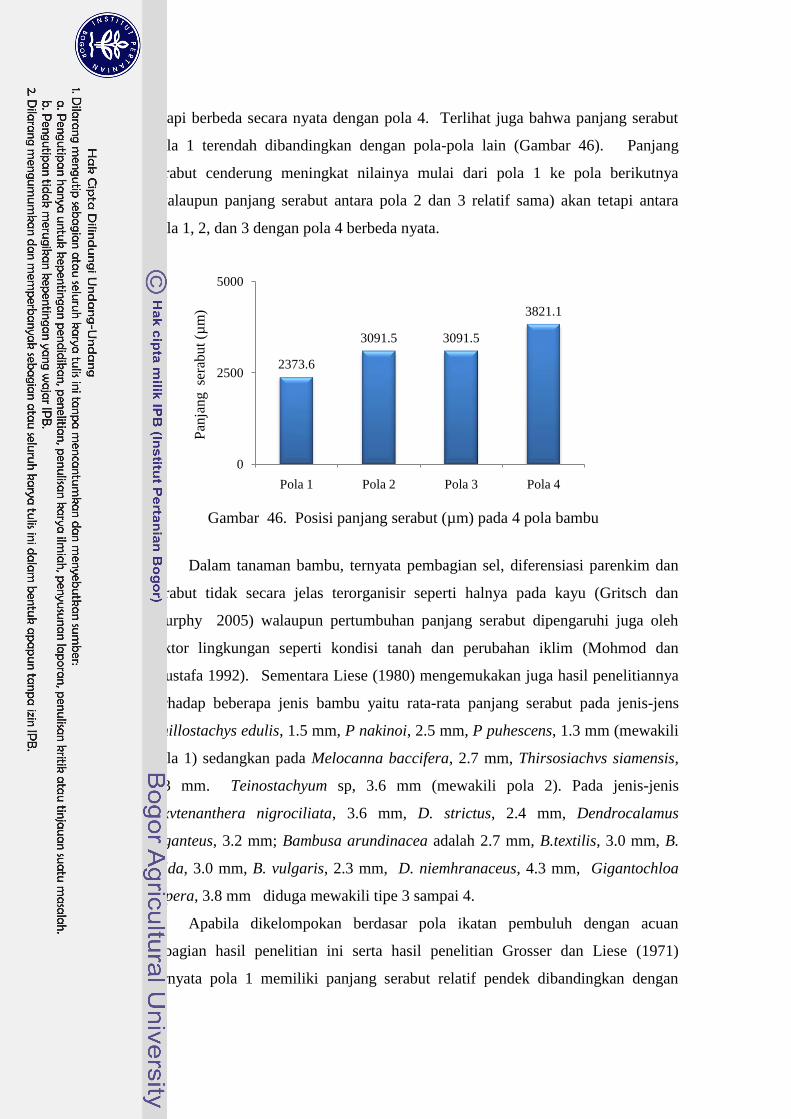
tetapi berbeda secara nyata dengan pola 4. Terlihat juga bahwa panjang serabut
pola 1 terendah dibandingkan dengan pola-pola lain (Gambar 46). Panjang
serabut cenderung meningkat nilainya mulai dari pola 1 ke pola berikutnya
(walaupun panjang serabut antara pola 2 dan 3 relatif sama) akan tetapi antara
pola 1, 2, dan 3 dengan pola 4 berbeda nyata.
Dalam tanaman bambu, ternyata pembagian sel, diferensiasi parenkim dan
serabut tidak secara jelas terorganisir seperti halnya pada kayu (Gritsch dan
Murphy 2005) walaupun pertumbuhan panjang serabut dipengaruhi juga oleh
faktor lingkungan seperti kondisi tanah dan perubahan iklim (Mohmod dan
Mustafa 1992). Sementara Liese (1980) mengemukakan juga hasil penelitiannya
terhadap beberapa jenis bambu yaitu rata-rata panjang serabut pada jenis-jens
Phillostachys edulis, 1.5 mm, P nakinoi, 2.5 mm, P puhescens, 1.3 mm (mewakili
pola 1) sedangkan pada Melocanna baccifera, 2.7 mm, Thirsosiachvs siamensis,
2.3 mm. Teinostachyum sp, 3.6 mm (mewakili pola 2). Pada jenis-jenis
Oxvtenanthera nigrociliata, 3.6 mm, D. strictus, 2.4 mm, Dendrocalamus
giganteus, 3.2 mm; Bambusa arundinacea adalah 2.7 mm, B.textilis, 3.0 mm, B.
tulda, 3.0 mm, B. vulgaris, 2.3 mm, D. niemhranaceus, 4.3 mm, Gigantochloa
aspera, 3.8 mm diduga mewakili tipe 3 sampai 4.
Apabila dikelompokan berdasar pola ikatan pembuluh dengan acuan
sebagian hasil penelitian ini serta hasil penelitian Grosser dan Liese (1971)
ternyata pola 1 memiliki panjang serabut relatif pendek dibandingkan dengan
2373.6
3091.5 3091.5
3821.1
0
2500
5000
Pola 1 Pola 2 Pola 3 Pola 4
Gambar 46. Posisi panjang serabut (µm) pada 4 pola bambu
Pan
jan
g se
rab
ut
(µm
)

pola-pola lain. Hal ini didukung pula oleh hasil penelitian Liese (1992)
terhadap bambu-bambu dengan panjang serabut yang cukup pendek yaitu
Phyllostachys edulis (1.5 mm), Ph. pubescens (1.3 mm), sedangkan bambu yang
memiliki panjang serabut yang cukup panjang yaitu Dendrocalamus giganteus
(3.2 mm), Oxytenanthera nigrocilliata (3.6 mm), D. membranaceus (4.3 mm).
Serabut-serabut itu sendiri tersusun dalam jaringan sklerenkim yang
terdapat sebagai selubung ikatan pembuluh atau rantai isolasi (Mohmod dan
Mustafa 1992). Sel serabut merupakan sel yang panjang dan meruncing pada
bagian ujungnya. Serabut dalam pola 1 terdapat dalam selubung sklerenkim
sedangkan serabut pada pola 2 selain ada pada selubung sklerenkim juga terdapat
perluasan serabut pada ruang antar sel. Serabut dalam pola 3 dan 4 selain dalam
selubung juga terdapat dalam bentuk rantai serabut. Posisi serabut dalam pola
ikatan pembuluh kemungkinan ikut mempengaruhi ukuran/panjang serabut. Pada
pola 4, sebagian besar serabut berada di dalam rantai serabut. Rantai serabut
akan membatasi perkembangan ke arah diameter dibanding ke arah panjang. Hal
ini terjadi terutama pada tahap awal diferensiasi jaringan di mana serabut yang
ada pada bagian dalam rantai terhalang berkembang ke arah diameter (Murphy
dan Alvin 1997).
Pada penampang melintang batang bambu, panjang serabut meningkat dari
bagian tepi sehingga mencapai puncaknya pada bagian tengah dan menurun ke
bagian dalam (Liese 1980). Terkait dengan hal tersebut dapat dijelaskan melalui
ketebalan masing-masing pola bambu. Ketebalan dinding batang pola 1dan 2
pada umumnya dibawah 1 cm (kecuali bagian pangkal bambu Cephalostachyum
pergracil memiliki ketebalan dinding batang paling tinggi (1.4 cm). Bambu-
bambu dengan pola 3 maksimum memiliki rata-rata ketebalan dinding batang
bagian pangkal sebesar 1.4 cm (kecuali bagian pangkal bambu Dendrocalamus
giganteus sebesar 2.40 cm dengan ketebalan bagian ujung rata-rata 0.5 cm).
Bambu dengan pola 4 umumnya pada bagian ujung memiliki pola 3 dengan
ketebalan dinding pangkal rata-rata dari 1 cm (Gigantochloa apus) sampai 1.8 cm
(Dendrocalamus asper) dengan ketebalan dinding batang bagian ujung adalah 0.5
cm (G. apus) dan 0.8 cm (D. asper). Adanya perbedaan nilai panjang serabut
antara pola 1, 2 dan 3 dengan pola 4 apabila dikaitkan dengan ketebalan dinding

batang terutama yang untuk jenis bambu yang memiliki 2 pola terlihat bahwa pola
4 berada pada dinding batang yang relatif lebih tebal dibandingkan dengan pola 3.
Apabila hal ini dikaitkan dengan pernyataan Liese (1980) yang mengungkapkan
bahwa panjang serabut pada bagian tengah batang adalah maksimum maka
dengan proporsi bagian tengah pada pola 4 lebih besar dibandingkan dengan pola
3 maka panjang serabut pada pola 4 lebih menonjol.
Persentase serabut
Berdasarkan hasil analisa keragaman terhadap faktor-faktor yang diduga
mempengaruhi nilai persentase serabut ternyata menghasilkan persamaan regresi
(3) yang bersifat sangat nyata (Lampiran 6).
Di antara faktor yang diujikan ternyata hanya posisi horizontal saja yang
berpengaruh sangat nyata sedangkan faktor jenis, posisi vertikal, dan pola tidak
berpengaruh nyata. Melalui uji lanjut Duncan ternyata nilai persentase serabut
pada posisi bagian dalam berbeda nyata dengan persentase serabut pada bagian
pusat. Demikian pula nilai persentase serabut antara bagian dalam, pusat dan tepi
berbeda nyata, hanya pada bagian tengah nilai persentase serabut berada diantara
bagian dalam dan pusat. Berikut hasil ujinya dalam Tabel 2.
Tabel 2. Hasil uji Duncan untuk persentase serabut
Posisi
Horizontal Persen Serabut Kode
Dalam 24.63 a
Tengah 31.97 ab
Pusat 33.17 b
Tepi 53.49 c
Pada bagian dalam penampang lintang batang, bentuk pola tidak sempurna
sehingga wilayah serabut menjadi berkurang. Dengan demikian bagian dalam
akan memiliki persentase serabut yang rendah. Persentase serabut pada bagian
Y= 49.5 – 2.1X2 + 1.5X4 - 7.9X5 + 10.4X6 - 7.4X7 + 1.6X8 + 5.6X9 - 8.5X10 +
1.2X11 - 21.5X12 + 4.6X13 - 8.7X14 + 7.8X15 ………………………………(3)
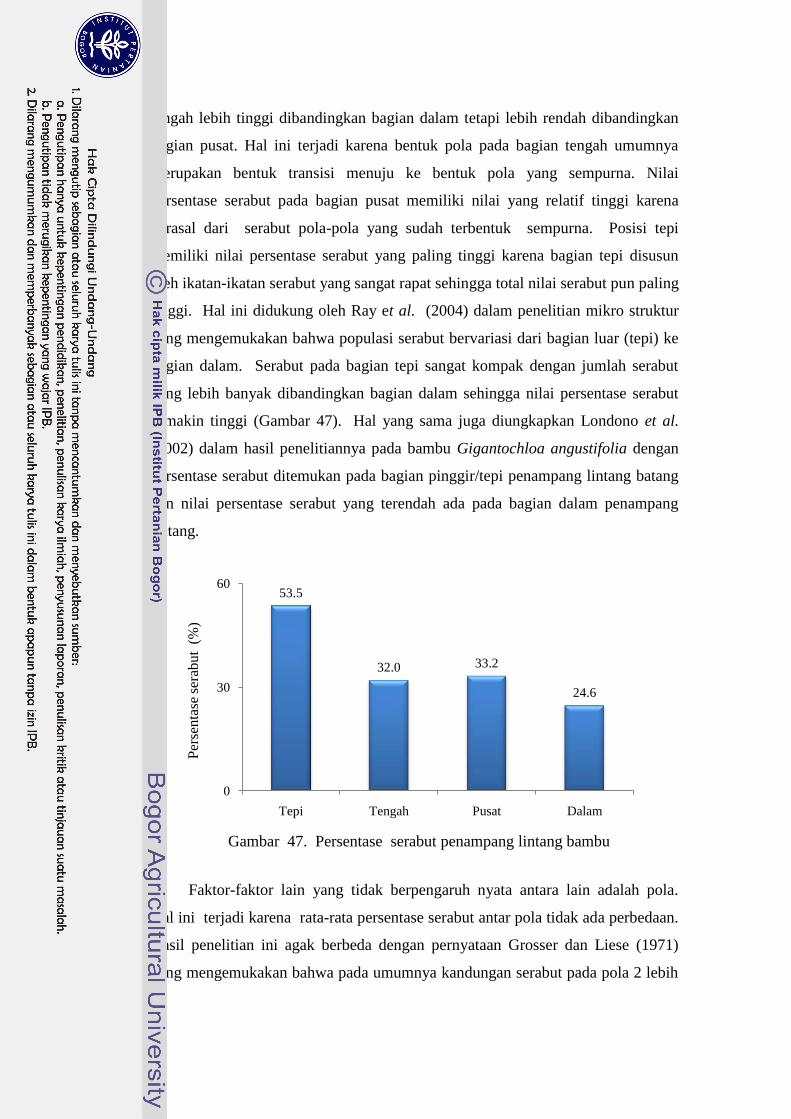
tengah lebih tinggi dibandingkan bagian dalam tetapi lebih rendah dibandingkan
bagian pusat. Hal ini terjadi karena bentuk pola pada bagian tengah umumnya
merupakan bentuk transisi menuju ke bentuk pola yang sempurna. Nilai
persentase serabut pada bagian pusat memiliki nilai yang relatif tinggi karena
berasal dari serabut pola-pola yang sudah terbentuk sempurna. Posisi tepi
memiliki nilai persentase serabut yang paling tinggi karena bagian tepi disusun
oleh ikatan-ikatan serabut yang sangat rapat sehingga total nilai serabut pun paling
tinggi. Hal ini didukung oleh Ray et al. (2004) dalam penelitian mikro struktur
yang mengemukakan bahwa populasi serabut bervariasi dari bagian luar (tepi) ke
bagian dalam. Serabut pada bagian tepi sangat kompak dengan jumlah serabut
yang lebih banyak dibandingkan bagian dalam sehingga nilai persentase serabut
semakin tinggi (Gambar 47). Hal yang sama juga diungkapkan Londono et al.
(2002) dalam hasil penelitiannya pada bambu Gigantochloa angustifolia dengan
persentase serabut ditemukan pada bagian pinggir/tepi penampang lintang batang
dan nilai persentase serabut yang terendah ada pada bagian dalam penampang
lintang.
Faktor-faktor lain yang tidak berpengaruh nyata antara lain adalah pola.
Hal ini terjadi karena rata-rata persentase serabut antar pola tidak ada perbedaan.
Hasil penelitian ini agak berbeda dengan pernyataan Grosser dan Liese (1971)
yang mengemukakan bahwa pada umumnya kandungan serabut pada pola 2 lebih
53.5
32.0 33.2
24.6
0
30
60
Tepi Tengah Pusat Dalam
Gambar 47. Persentase serabut penampang lintang bambu
Per
senta
se s
erab
ut
(%
)

banyak dibandingkan dengan pola 1 demikian pula dengan kandungan serabut
pada pola 3 lebih banyak dibandingkan dengan pola 2 dan seterusnya. Pernyataan
yang dikemukakan oleh Grosser dan Liese tentang persentase serabut akan
terbukti jika jarak antar pola ikatan cukup rapat. Dalam pengamatan visual
terhadap pola 3 dan 4 terlihat bahwa jarak antar pola relatif jarang dibandingkan
dengan pola 1 dan 2. Diduga kondisi ini turut mempengaruhi nilai persentase
serabut.
Hubungan antara kerapatan ikatan pembuluh dengan persentase serabut
menghasilkan persamaan Y = 0.572 + 0.045X dengan R2=0.159 yang berarti
bahwa terdapat hubungan yang positif antara kerapatan ikatan pembuluh dengan
persentase serabut yang bermakna bahwa kerapatan ikatan pembuluh akan
semakin tinggi nilainya apabila nilai persentase serabut semakin besar. Hal ini
terjadi karena dengan nilai persentase serabut yang tinggi maka semakin tinggi
peluang untuk memperoleh kerapatan yang tinggi walaupun koefisien korelasinya
kecil (39.87%), hal ini terutama berlaku untuk pola-pola yang sederhana (1 dan 2).
Susunan pola ikatan pembuluh pada pola yang sederhana terdiri atas 1 rantai
pembuluh pusat yang dikelilingi selubung serabut (sklerenkim) dan biasanya jarak
antar pola relatif dekat. Susunan pola ikatan pembuluh pada pola yang kompleks
(pola 3 dan 4) selain terdiri atas unsur-unsur yang ada pada pola 1 dan 2 juga
ditambah dengan 1 atau 2 rantai serabut dengan jarak tertentu. Dengan kondisi
tersebut akan memunculkan nilai koefisien korelasi yang kecil (Gambar 48).
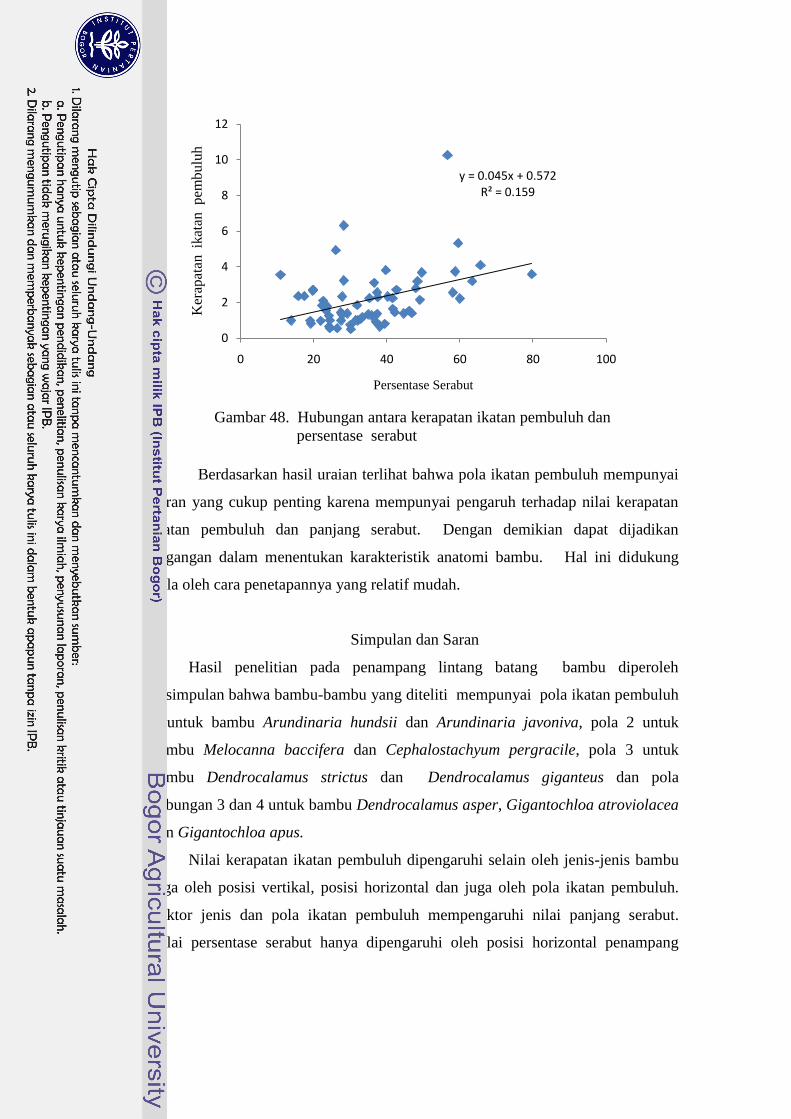
Berdasarkan hasil uraian terlihat bahwa pola ikatan pembuluh mempunyai
peran yang cukup penting karena mempunyai pengaruh terhadap nilai kerapatan
ikatan pembuluh dan panjang serabut. Dengan demikian dapat dijadikan
pegangan dalam menentukan karakteristik anatomi bambu. Hal ini didukung
pula oleh cara penetapannya yang relatif mudah.
Simpulan dan Saran
Hasil penelitian pada penampang lintang batang bambu diperoleh
kesimpulan bahwa bambu-bambu yang diteliti mempunyai pola ikatan pembuluh
1 untuk bambu Arundinaria hundsii dan Arundinaria javoniva, pola 2 untuk
bambu Melocanna baccifera dan Cephalostachyum pergracile, pola 3 untuk
bambu Dendrocalamus strictus dan Dendrocalamus giganteus dan pola
gabungan 3 dan 4 untuk bambu Dendrocalamus asper, Gigantochloa atroviolacea
dan Gigantochloa apus.
Nilai kerapatan ikatan pembuluh dipengaruhi selain oleh jenis-jenis bambu
juga oleh posisi vertikal, posisi horizontal dan juga oleh pola ikatan pembuluh.
Faktor jenis dan pola ikatan pembuluh mempengaruhi nilai panjang serabut.
Nilai persentase serabut hanya dipengaruhi oleh posisi horizontal penampang
y = 0.045x + 0.572R² = 0.159
0
2
4
6
8
10
12
0 20 40 60 80 100
Ker
apat
an i
kat
an p
emb
ulu
h
Persentase Serabut
Gambar 48. Hubungan antara kerapatan ikatan pembuluh dan
persentase serabut

lintang bambu. Persentase serabut yang semakin tinggi akan memberikan nilai
kerapatan ikatan pembuluh yang tinggi.
Pola ikatan pembuluh pada bambu berhubungan erat dengan struktur
penyusun ikatan pembuluh dan bersifat konsisten pada genus dan spesies sehingga
dapat digunakan untuk identifikasi bamboo. Karakteristik struktur penyusun pola
ikatan pembuluh tersebut merupakan dasar bagi arah pemanfaatan bambu.
Pola ikatan pembuluh perlu dipertimbangkan dalam pengenalan jenis bambu
dan perencanaan pemanfaatan bambu sesuai dengan karakteristik yang dimiliki
setiap jenis bambu. Untuk itu perlu pendalaman lebih lanjut mengenai sifat
struktural dan teknologinya.
DAFTAR PUSTAKA
[ABS] American Bamboo Society. 1999. Bamboo: Past-present-future.
Newsletter 20 (1).

Botanic Gardens of Indonesia. 2001. An Alphabetical List of Plants Spesies
Cultivated in the Bogor Botanical Garden. Indonesian Institute of Sciences.
Dransfield S, Widjaja EA. 1995. Bamboos. Indonesia: Prosea Bogor.
Draper N, Smith H. 1992. Analisis Regresi Terapan. Ed ke-2. Jakarta: PT
Gramedia Pusta Utama.
Gritsch CS, Murphy RJ. 2005. Ultrastructure of fibre and parenchyma cell walls
during early stages of culm development in Dendrocalamus asper. Annals
of Botany 95: 619–629.
Grosser D, Liese W. 1971. On the anatomy of Asian bamboos, with spesial
reference to their vascular bundles. Wood Sci and Tech 5: 290-312
Liese W. 1980. Anatomy of Bamboo. Di dalam: Lessard G dan Chounard A,
editor. Proceedings of a Workshop. Singapore, 28-30 Mei 1980.
Lwin KM, Han YY, Maung W, Moe AKZ, Than SBM. 2007. An investigation on
morphology, anatomy and chemical properties of some Myanmar bamboos.
http://www.myanmar.gov.mm/Ag/Jur/ProcFo01.10.[3 Nov. 2007].
Liese W. 1987. Anatomy and properties of bamboo. Di dalam: Rao, A.N.,
Dhanarajan, G. dan Sastry, C.B, editor. Recent Research on Bamboo.
Proceedings of the International Bamboo Workshop, Hangzholu, People's
Republic of China, 6-14 Oktober, 1985. Academy of Forestry, People's
Republic of China & International ,Development Research Centre, Canada,
hal: 196-208.
Liese W. 1992. The structure of bamboo in relation to its properties and
utilization. Dalam Bamboo And Its Use, International Symposium On
Industrial Use Of Bamboo. Beijing, China, 7-11 Desember 1992. hlm 1 - 6
Londono X, Camayo GC, Riano NM, Lopez Y. 2002. Characterization of the
anatomy of Guadua angustifolia (Poaceae: Bambusoideae) culms. J Am
Bam Soc 16 (1): 18–31.
Mohmod AL, Mustafa MT. 1992. Variasi in anatomical properties of three
Malaysian bamboos from natural stands. J Tropic For Sci 5(1): 90-96
Murphy RJ , Alvin KL. 1997. Fibre maturation in the bamboo Gigantochloa
scortechinii. IAWA J, Vol. 18 (2): 147-156.
Ray AK, Das SK, Mondal S. 2004. Microstructural characterization of bamboo.
J Mater Sci 39: 1055-1060.
Sass JE. 1951. Botanical Microtechnique . Iowa: The Iowa State College Press.

Wahab R, Mohammed A, Mustafa MT, Hassan A. 2009. Physical characteristics
and anatomical properties of cultivated bamboo Bambusa vulgaris (Schrad.)
culms. J Biol Sci 9: 753-759.
Widjaja, EA. 2001. Identifikasi Jenis-jenis Bambu di Jawa. Bogor : LIPI – seri
panduan lapangan.
III. KORELASI POLA IKATAN PEMBULUH PADA SIFAT FISIK
DAN MEKANIK TIGA JENIS BAMBU

Abstrak
Sifat fisik dan mekanik 3 jenis bambu, yaitu Dendrocalamus giganteus,
Dendrocalamus asper, dan Gigantochloa apus diteliti dalam hubungannya dengan
pola ikatan pembuluh. Sifat fisik dan mekanik seperti berat jenis, keteguhan
lentur patah (MOR), modulus elastisitas (MOE), keteguhan tekan sejajar serat,
dan keteguhan tarik sejajar serat ditentukan, dan pola ikatan pembuluh dianalisis
dengan metode berdasarkan panduan dari Grosser dan Liese (1971). Hubungan
diantara sifat fisik dan mekanik dengan pola ikatan pembuluh dianalisis regresi
dengan variabel dummy. Kombinasi pola ikatan pembuluh ditemukan pada G.
apus dan D. asper, sedangkan D. giganteus mempunyai pola ikatan pembuluh
tunggal. Perbedaan pola ikatan pembuluh tidak memberikan kontribusi pada sifat
fisik dan mekanik bambu yang diteliti kecuali pada MOR. Perbedaan jenis bambu
dan posisi vertikal batang memberikan kontribusi pada perbedaan nilai keteguhan
tekan sejajar serat, sedangkan keteguhan tarik hanya dipengaruhi oleh jenis
bambu.
Kata kunci: pola ikatan pembuluh, sifat fisik, sifat mekanik
Abstract
The physical and mechanical properties of three species of bamboo, namely
Dendrocalamus giganteus, Dendrocalamus asper, and Gigantochloa apus were
investigated in relation to its vascular bundle pattern. As physical and
mechanical properties, specific gravity, modulus of rupture (MOR), modulus of
elasticity (MOE), compressive strength parallel to grain and tension strength
parallel to grain were determined, and the vascular bundle pattern was evaluated
by method according to Grosser and Liese (1971). The relationship between
physical and mechanical properties with the vascularl bundle pattern was
analyzed by regression with dummy variables. Pattern combination of vascular
bundle was found on G. apus and D. asper, while D. giganteus has a single
pattern of vascular bundle type. The difference of vascular bundle pattern did not
contributed to the physical and mechanical properties of bamboo investigated,
except for MOR. The difference species of bamboo and vertical position of
samples contributed to the different value of compressive strength parallel to
grain, whereas tension strength was only affected by bamboo species.
Key words: vascularl bundle pattern, physical properties, mechanical properties.
Pendahuluan

Tanaman bambu merupakan tanaman serba guna bagi masyarakat
Indonesia. Pentingnya tanaman bambu dalam berbagai penggunaan telah
diperkenalkan di berbagai negara mulai dari makanan, kerajinan, mebel, sampai
berbagai produk industri (Erakhrumen dan Ogunsanwo 2009). Hal ini didukung
juga oleh kondisi semakin menurunnya ketersediaan kayu sehingga bambu
berpeluang tinggi sebagai bahan yang diharapkan setara dengan kayu. Selama
ini kayu merupakan bahan bangunan yang dominan dipergunakan sebagai
konstruksi rangka struktur (rangka lantai, rangka dinding dll) dan yang bersifat
nonstruktur (penutup lantai, penutup dinding, penutup langit-langit, dll). Bambu
dapat menggantikan kayu (Purwito 2008) karena bambu memiliki keunggulan
sebagai bahan bangunan dan merupakan salah satu material yang sangat potensial
untuk pemenuhan kebutuhan perumahan, serta telah diakui masyarakat dunia
dengan terbitnya standar internasional (ISO) yang masih perlu diadaptasi untuk
diterapkan di Indonesia dengan penyesuaian pada kondisi setempat.
Hal lain yang cukup menarik untuk diperhatikan dengan banyak
digunakannya bambu sebagai bahan bangunan adalah harganya yang relatif
murah, ramah lingkungan, dan ketersediaannya yang berlimpah (BMTPC 2007) .
Pengetahuan sifat-sifat mekanik bambu mendapatkan perhatian para periset
berkaitan dengan potensi bambu untuk digunakan sebagai bahan konstruksi yang
memberikan kekuatan tinggi namun ringan (Ghavami et al. 2003a). Bobot jenis
adalah sifat fisik yang paling penting yang mempengaruhi sifat kekakuan,
kekuatan dan penyusutan, dalam skala yang lebih luas lagi bobot jenis ini dapat
menentukan penggunaan akhir bahan berkayu (Wang et al. 2011). Hal yang
sama diungkapkan pula oleh Jansen (1987) yang menyatakan bahwa antara BJ
dengan sifat-sifat mekanis terkait erat. Dengan demikian kedua sifat tersebut
merupakan satu rangkaian yang saling terkait.
Terdapat berbagai jenis tanaman bambu yang dapat digunakan untuk
keperluan bangunan antara lain Dendrocalamus asper, D giganteus,
Gigantochloa atroviolacea (Surjokusumo 1997). Selama ini penggunaan bambu
berdasarkan kebiasaan turun temurun (Morisco 2011). Kondisi seperti ini
menyulitkan penggunaan bambu secara optimum dan tidak mudah untuk
menentukan manfaat setiap jenis bambu secara tepat. Hal ini didukung juga

dengan banyaknya jenis-jenis bambu yang belum dikenal dan belum diketahui
kesesuaian pemanfaatannya. Salah satu alternatif cara yang dapat dilakukan untuk
mengetahui sifat-sifat bambu adalah melalui pendekatan evaluasi pola ikatan
pembuluh yang ada di setiap jenis bambu.
Bambu memiliki berbagai pola ikatan pembuluh yang bersikap khas untuk
jenis bambu tertentu. Menurut Grosser dan Liese (1971) tanaman bambu
memiliki 4 pola ikatan pembuluh. Jenis bambu yang selama ini dipergunakan
sebagai bahan dasar konstruksi diduga umumnya memiliki pola ikatan pembuluh
pola 3 atau 4 (Nuriyatin 2000). Hal ini mendasari perlunya penelitian secara
mendalam tentang pola ikatan pembuluh sebagai variabel yang dapat digunakan
sebagai penduga sifat mekanis bambu dan penggunaannya sebagai bahan
konstruksi. Penelitian ini bertujuan untuk menganalisis korelasi antara pola ikatan
pembuluh dengan sifat fisik dan mekanik beberapa jenis bambu.
Bahan dan Metode
Bahan
Bahan penelitian adalah 3 jenis bambu yaitu Dendrocalamus giganteus
Wallich ex Munro, Gigantochloa apus (J.A. & J.H. Schultes) Kurs dan D. asper
(Schultes f.) Backer ex Heyne yang telah berumur 3-4 tahun. Sampel bambu
diambil sekitar Fakultas Kehutanan IPB serta di Kebun Raya Bogor. Sampel uji
diambil di bagian pangkal, tengah dan ujung bambu dengan ulangan 3 kali
kecuali untuk D giganteus pada bagian ujung tidak dapat diambil sampel untuk
sifat mekanis karena dinding batangnya sangat tipis.
Metode
Pembuatan contoh uji bobot jenis mengikuti standar ISO/TC165N314
(2001) sedangkan pembuatan contoh uji sifat mekanik terutama untuk MOE,
MOR berpedoman ke ASTM D 143-94 (2000) yang dimodifikasi. Dalam hal ini
tidak setiap jenis bambu memiliki ketebalan yang sama sehingga untuk
mendapatkan ketebalan tertentu dilakukan proses penyambungan secara
setangkup terutama dalam pembuatan sampel uji lentur. Bentuk dan ukuran
keteguhan tekan sejajar serat berpedoman ke ISO/TC165N314 (2001) dengan

panjang spesimen sama dengan diameter bambu terluar bahkan jika lebih kecil
atau sama dengan 20 mm maka panjangnya 2 kali diameter terluar. Bentuk dan
ukuran contoh uji keteguhan tarik sejajar serat berpedoman ke ASTM D 143-94
(2000) yang dimodifikasi dengan ukuran yang lebih panjang dan ketebalan
sampel sesuai dengan ketebalan bambu. Penelitian dilaksanakan di Laboratorium
Bagian Rekayasa dan Disain Bangunan Kayu Hasil Hutan Fakultas Kehutanan
serta di Laboratorium Fisik dan Mekanik, Pusat Penelitian dan Pengembangan
Hasil Hutan, Kementerian Kehutanan.
Bentuk contoh uji
Bentuk contoh uji bobot jenis, keteguhan tekan, keteguhan lentur dan
keteguhan tarik ditampilkan berturut-turut pada Gambar 49 sampai Gambar 52.
Gambar 49. Bentuk contoh uji bobot jenis (BJ)
Gambar 50. Bentuk contoh uji tekan sejajar serat
2.5 cm
2.5 cm
D
2D
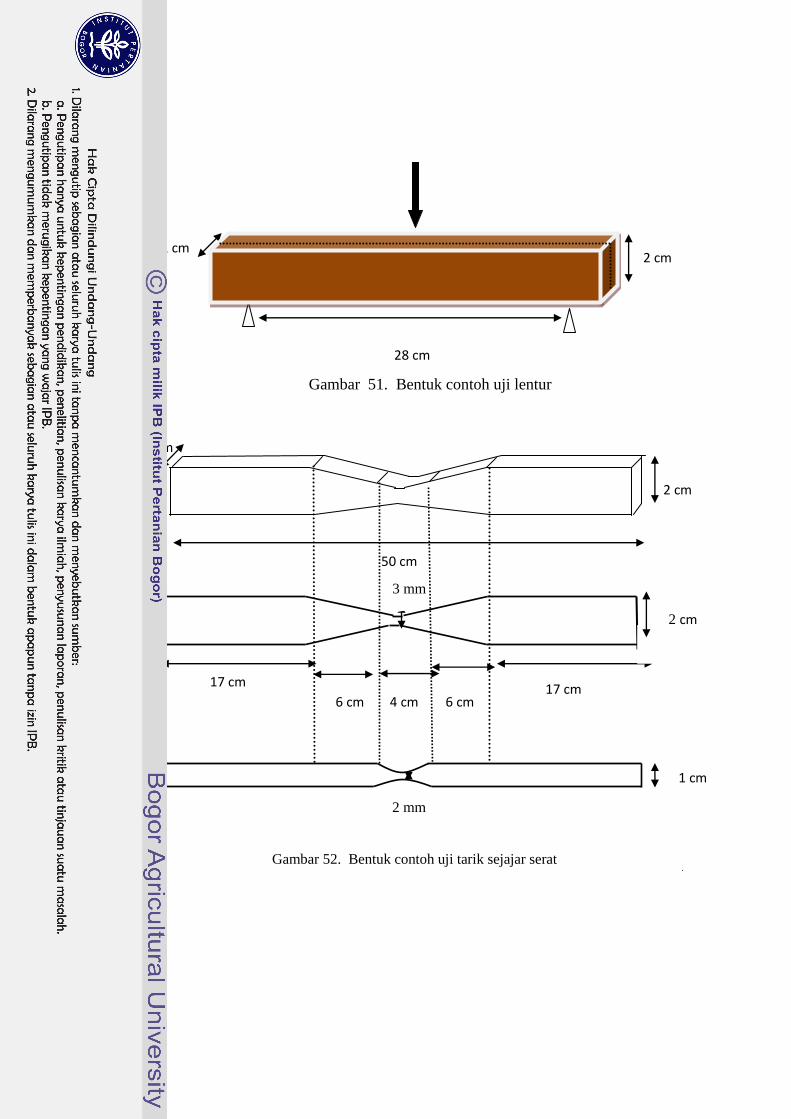
2 cm
2 cm
Gambar 52. Bentuk contoh uji tarik sejajar serat
Gambar 51. Bentuk contoh uji lentur
1 cm
28 cm
30 cm
50 cm
1cm
2 cm
17 cm 17 cm 6 cm 6 cm 4 cm
3 mm
2 mm
1 cm

Prosedur pengujian
Pengujian bobot jenis dilakukan dengan cara pengukuran volume sampel uji
dengan metoda pemindahan berat dan pengukuran bobot kering tanur (BKT)
sampel uji. Perhitungan bobot jenis (BJ) dengan menggunakan rumus(4):
Keterangan:
BJ : Bobot jenis
BKT : Bobot Kering Tanur
Vol : Volume saat basah
Pengujian MOE menggunakan mesin Instron dengan pembebanan
dilakukan di tengah-tengah contoh uji dengan panjang bentang 28 cm.
Perhitungan MOE menggunakan rumus(5):
Keterangan:
MOE : Modulus Elastisitas (kg/cm2)
P : Beban sebelum batas proporsi (kg)
L : Jarak sangga (cm)
y : Lenturan/defleksi pada beban P (cm)
b : Lebar contoh uji (cm)
h : Tinggi contoh uji (cm)
Keteguhan lentur patah (MOR)
Pengujian MOR menggunakan mesin Instron dengan pembebanan
dilakukan di tengah-tengah contoh uji dengan jarak sangga 28 cm. Perhitungan
MOR menggunakan rumus (6):
Keterangan :
MOR : Modulus of Rupture (kg/cm2)
P : Beban maksimal (kg)
MOE = PL3
4 ybh3
MOR = 3 P L
2 bh2
BJ = BKT
Vol. …………………………………………………(4)
………………………………………………….(5)
………………………………………………... (6)

L : Jarak sangga (cm)
b : Lebar contoh uji (cm)
h : Tinggi contoh uji (cm) Keteguhan Tekan Sejajar Serat (σ tk//)
Pengujian keteguhan tekan sejajar menggunakan mesin Baldwin.
Pembebanan dilakukan pada arah sejajar serat dengan kedudukan contoh uji
vertikal sehingga terjadi kerusakan pada contoh uji. Besarnya keteguhan tekan
sejajar serat dihitung dengan rumus (7):
Keterangan :
σtk// : Keteguhan tekan sejajar serat (kg/cm2)
P : Beban tekan maksimum (kg)
A : Luas penampang (cm2)
Keteguhan Tarik Sejajar Serat (σtr//)
Pengujian keteguhan tarik sejajar menggunakan mesin Instron dengan kedudukan
contoh uji vertikal yang dijepit pada kedua ujungnya kemudian ditarik sehingga terjadi
kerusakan pada dimensi terkecilnya. Besarnya keteguhan tarik sejajar serat dihitung
dengan rumus (8):
Keterangan :
σtr// : keteguhan tarik sejajar serat (kg/cm2)
P : Beban tekan maksimum (kg)
A : Luas penampang (cm2)
Analisis data
Data dianalisis melalui pendekatan regresi dengan peubah boneka. Peubah
boneka dalam analisis ini adalah tiga jenis bambu yang terwakili dalam peubah
X1-X2, posisi vertikal yaitu pangkal, tengah dan ujung yang terwakili dalam
peubah X3 dan X4, dua pola bambu yang terwakili dalam X5. Kontribusi
keseluruhan peubah akan dianalisis dalam persamaan regresi.
σ tk// : P
A
σ tr// = P
A
…………………………………………………… (7)
…………………………………………………..(8)
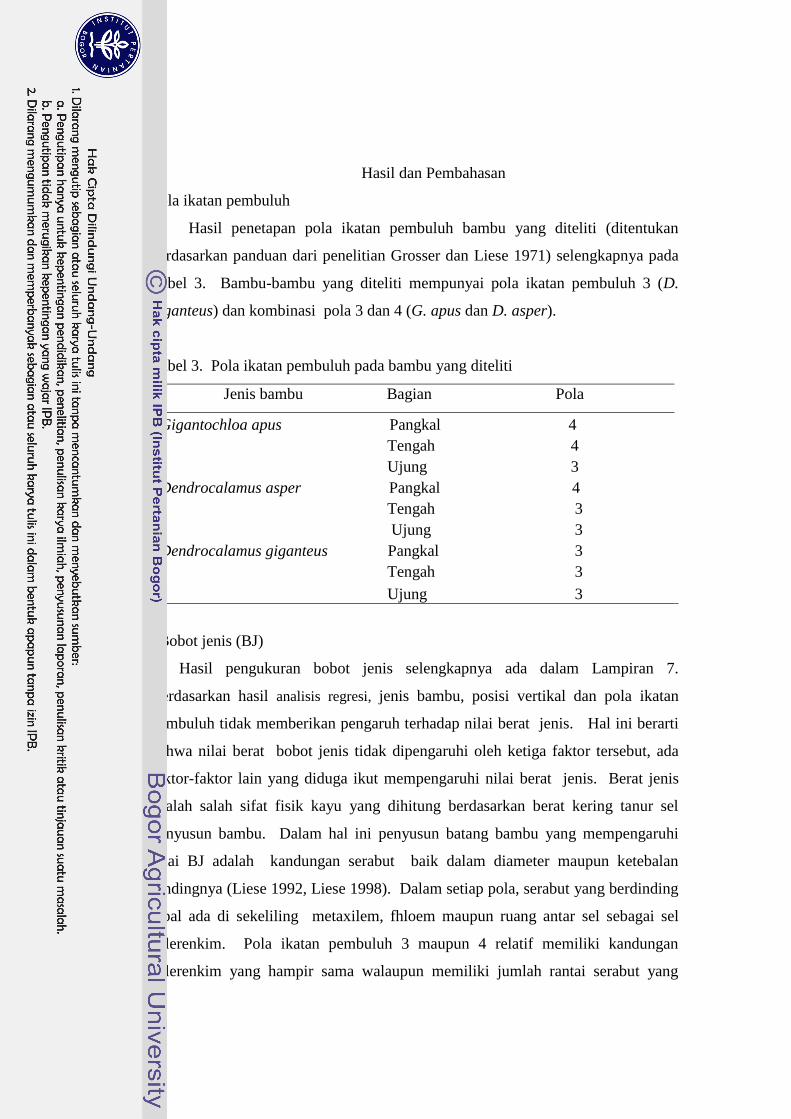
Hasil dan Pembahasan
Pola ikatan pembuluh
Hasil penetapan pola ikatan pembuluh bambu yang diteliti (ditentukan
berdasarkan panduan dari penelitian Grosser dan Liese 1971) selengkapnya pada
Tabel 3. Bambu-bambu yang diteliti mempunyai pola ikatan pembuluh 3 (D.
giganteus) dan kombinasi pola 3 dan 4 (G. apus dan D. asper).
Tabel 3. Pola ikatan pembuluh pada bambu yang diteliti
Jenis bambu Bagian Pola
Gigantochloa apus Pangkal 4
Tengah 4
Ujung 3
Dendrocalamus asper Pangkal 4
Tengah 3
Ujung 3
Dendrocalamus giganteus Pangkal 3
Tengah 3
Ujung 3
Bobot jenis (BJ)
Hasil pengukuran bobot jenis selengkapnya ada dalam Lampiran 7.
Berdasarkan hasil analisis regresi, jenis bambu, posisi vertikal dan pola ikatan
pembuluh tidak memberikan pengaruh terhadap nilai berat jenis. Hal ini berarti
bahwa nilai berat bobot jenis tidak dipengaruhi oleh ketiga faktor tersebut, ada
faktor-faktor lain yang diduga ikut mempengaruhi nilai berat jenis. Berat jenis
adalah salah sifat fisik kayu yang dihitung berdasarkan berat kering tanur sel
penyusun bambu. Dalam hal ini penyusun batang bambu yang mempengaruhi
nilai BJ adalah kandungan serabut baik dalam diameter maupun ketebalan
dindingnya (Liese 1992, Liese 1998). Dalam setiap pola, serabut yang berdinding
tebal ada di sekeliling metaxilem, fhloem maupun ruang antar sel sebagai sel
sklerenkim. Pola ikatan pembuluh 3 maupun 4 relatif memiliki kandungan
sklerenkim yang hampir sama walaupun memiliki jumlah rantai serabut yang
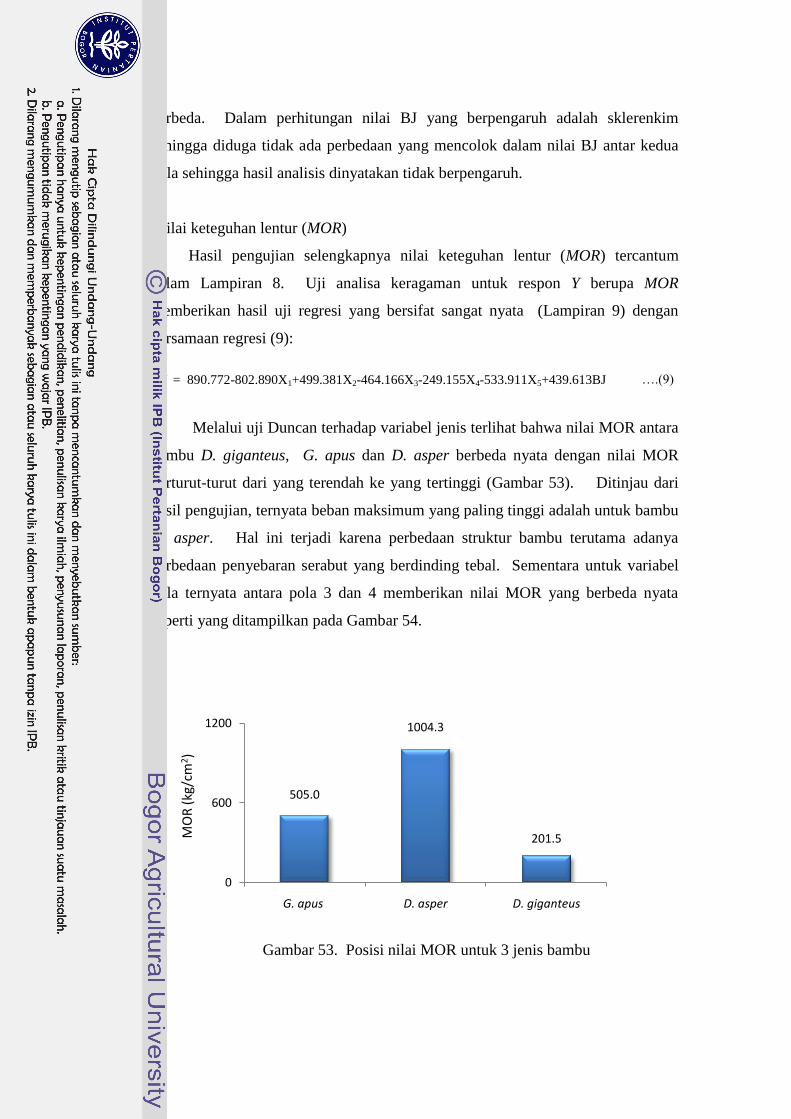
berbeda. Dalam perhitungan nilai BJ yang berpengaruh adalah sklerenkim
sehingga diduga tidak ada perbedaan yang mencolok dalam nilai BJ antar kedua
pola sehingga hasil analisis dinyatakan tidak berpengaruh.
Nilai keteguhan lentur (MOR)
Hasil pengujian selengkapnya nilai keteguhan lentur (MOR) tercantum
dalam Lampiran 8. Uji analisa keragaman untuk respon Y berupa MOR
memberikan hasil uji regresi yang bersifat sangat nyata (Lampiran 9) dengan
persamaan regresi (9):
Variabel-variabel yang mempengaruhi nilai MOR adalah jenisan p
Melalui uji Duncan terhadap variabel jenis terlihat bahwa nilai MOR antara
bambu D. giganteus, G. apus dan D. asper berbeda nyata dengan nilai MOR
berturut-turut dari yang terendah ke yang tertinggi (Gambar 53). Ditinjau dari
hasil pengujian, ternyata beban maksimum yang paling tinggi adalah untuk bambu
D. asper. Hal ini terjadi karena perbedaan struktur bambu terutama adanya
perbedaan penyebaran serabut yang berdinding tebal. Sementara untuk variabel
pola ternyata antara pola 3 dan 4 memberikan nilai MOR yang berbeda nyata
seperti yang ditampilkan pada Gambar 54.
505.0
1004.3
201.5
0
600
1200
G. apus D. asper D. giganteus
Gambar 53. Posisi nilai MOR untuk 3 jenis bambu
MO
R (
kg/c
m2)
Y = 890.772-802.890X1+499.381X2-464.166X3-249.155X4-533.911X5+439.613BJ ….(9)
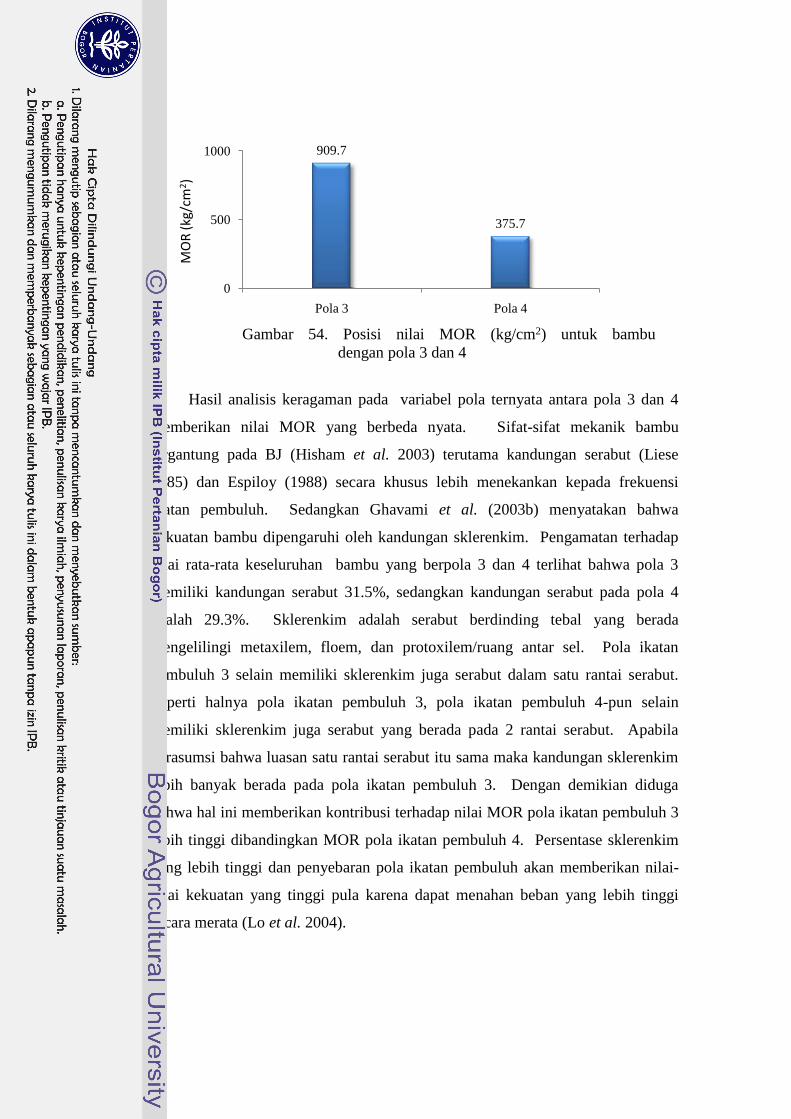
Hasil analisis keragaman pada variabel pola ternyata antara pola 3 dan 4
memberikan nilai MOR yang berbeda nyata. Sifat-sifat mekanik bambu
tergantung pada BJ (Hisham et al. 2003) terutama kandungan serabut (Liese
1985) dan Espiloy (1988) secara khusus lebih menekankan kepada frekuensi
ikatan pembuluh. Sedangkan Ghavami et al. (2003b) menyatakan bahwa
kekuatan bambu dipengaruhi oleh kandungan sklerenkim. Pengamatan terhadap
nilai rata-rata keseluruhan bambu yang berpola 3 dan 4 terlihat bahwa pola 3
memiliki kandungan serabut 31.5%, sedangkan kandungan serabut pada pola 4
adalah 29.3%. Sklerenkim adalah serabut berdinding tebal yang berada
mengelilingi metaxilem, floem, dan protoxilem/ruang antar sel. Pola ikatan
pembuluh 3 selain memiliki sklerenkim juga serabut dalam satu rantai serabut.
Seperti halnya pola ikatan pembuluh 3, pola ikatan pembuluh 4-pun selain
memiliki sklerenkim juga serabut yang berada pada 2 rantai serabut. Apabila
berasumsi bahwa luasan satu rantai serabut itu sama maka kandungan sklerenkim
lebih banyak berada pada pola ikatan pembuluh 3. Dengan demikian diduga
bahwa hal ini memberikan kontribusi terhadap nilai MOR pola ikatan pembuluh 3
lebih tinggi dibandingkan MOR pola ikatan pembuluh 4. Persentase sklerenkim
yang lebih tinggi dan penyebaran pola ikatan pembuluh akan memberikan nilai-
nilai kekuatan yang tinggi pula karena dapat menahan beban yang lebih tinggi
secara merata (Lo et al. 2004).
909.7
375.7
0
500
1000
Pola 3 Pola 4
Gambar 54. Posisi nilai MOR (kg/cm2) untuk bambu
dengan pola 3 dan 4
MO
R (
kg/c
m2)

Modulus elastisitas (MOE)
Hasil perhitungan nilai MOE selengkapnya terdapat pada Lampiran 8. Nilai
MOE bambu G. apus, D. asper, dan D. giganteus berturut-turut adalah 22005.3
kg/cm2, 40365.1 kg/cm
2, dan 56643.4 kg/cm
2 dan secara berturut-turut juga
merupakan urutan nilai MOE mulai yang terendah ke yang tertinggi. Perbedaan
nilai MOE terjadi karena perbedaan persentase sklerenkim yang dapat ditunjukan
oleh perbedaan nilai BJ (Janssen 1981).
Hasil pengolahan data menyatakan bahwa nilai modulus elastisitas tidak
dipengaruhi oleh jenis bambu, posisi vertikal, pola dan BJ. Persamaan regresi
yang terbentuk tidak bersifat nyata (Lampiran 9) bahkan koefisien determinasinya
pun bernilai kecil, yaitu sebesar 33.47% yang berarti bahwa hanya 33.47% variasi
Y yang dapat diterangkan oleh variabel X sedangkan sisanya yaitu 66.53%
diterangkan oleh variabel lain selain X. Dengan demikian ada variabel lain yang
berperan dan mempengaruhi nilai MOE.
Hamdan et al. (2009) menyatakan bahwa struktur anatomi mempengaruhi
sangat kuat pada sifat-sifat mekanik. Panjang serabut berkorelasi sangat kuat
terhadap nilai MOE (Liese 2003). Serabut tersusun atas sejumlah lapisan/lamella
dengan berbagai orientasi mikrofibril. Susunan sel serabut tersebut akan
memberikan kontribusi yang tinggi terhadap fleksibilitas bambu. Informasi
mengenai panjang serabut tidak muncul dalam bentuk pola ikatan pembuluh.
Dengan demikian diduga hal ini yang menyebabkan pola ikatan pembuluh tidak
memberikan kontribusi terhadap nilai MOE.
Keteguhan tekan sejajar serat
Hasil perhitungan nilai keteguhan sejajar serat tercantum pada Lampiran 10.
Persamaan regresi yang membentuk hubungan antara keteguhan tekan (Y) dan
peubah X (10) memberikan hasil yang bersifat sangat nyata dengan koefisien
determinasi (R2) sebesar 79% (Lampiran 11) membentuk persamaan:
Y = 3711.204-497.518X1+757.393X2-321.699X3-3631.46X4+894.204X5+524.266BJ …….(10)

Berdasar uji analisa keragaman ternyata faktor jenis bersifat nyata dan posisi
vertikal bersifat sangat nyata. Hasil uji beda Duncan terhadap jenis bambu
menyatakan tidak ada perbedaan artinya bahwa uji Duncan tidak cukup peka
terhadap perbedaan yang ada namun perbandingan diantara nilai-nilai keteguhan
tekan yang ada diantara jenis-jenis tersebut dari yang terendah ke yang tertinggi
berturut-turut adalah bambu G. apus, D. giganteus dan D. asper.
Posisi 3 jenis bambu berdasar nilai keteguhan tekan sejajar serat
tampilannya disajikan pada Gambar 55. Hasil pengamatan terhadap tampilan fisik
sampel uji keteguhan tekan sejajar serat ke-3 jenis bambu pada kondisi kering
udara ternyata Gigantochloa apus memiliki dinding batang yang paling tipis,
yaitu (0.2 cm-0.4 cm) dan luas permukaan paling kecil serta memerlukan tekanan
maksimum yang paling rendah dibandingkan bambu-bambu lain. Sementara pada
Dendrocalamus giganteus memiliki ketebalan dinding batang kurang lebih
berkisar dari 0.4 cm-0.6 cm dengan luas permukaan paling besar dibandingkan
bambu lain namun tekanan yang diperlukan sampai sampel uji rusak berada
diantara tekanan bambu D. asper dan G. apus. Pada D. asper meskipun
ketebalan dinding batangnya sekitar 0.5 cm dengan luas permukaan lebih kecil
dibandingkan D. giganteus namun memerlukan tekanan yang paling tinggi untuk
sampai pada posisi sampelnya rusak sehingga mempunyai nilai keteguhan tekan
yang paling tingggi diantara ke-3 jenis bambu. Menganalisis fakta-fakta yang ada
diduga bahwa yang menentukan dalam keteguhan tekan selain luas permukaan
penampang juga struktur dari bambunya sendiri. Perbedaan struktur terjadi
karena distribusi serabut pada penampang lintang bambu (Shao et al. 2010).

Melihat penyebaran serabut pada 2 jenis bambu khususnya D. asper dan D.
giganteus pada batang bagian tengah terlihat bahwa permukaan penampang
lintang D. asper memiliki kandungan serabut yang lebih tinggi dibandingkan D.
giganteus pada berbagai bagian permukaan batang. Demikian pula pada bagian
ujung nampak bahwa persen serabut pada bambu D. asper lebih tinggi. Data
selengkapnya ada pada Tabel 4.
Tabel 4. Persentase serabut pada 2 jenis bambu
Jenis bambu Posisi vertikal Posisi horizontal % serabut
D. asper Tengah tepi 49.1
tengah 31.5
pusat 30.2
dalam 24.5
D. giganteus Tengah tepi 41.7
tengah/pusat 22.0
dalam 19.0
D. asper Ujung tepi …
tengah 37
pusat 33
dalam ….
D. giganteus Ujung tepi 49.6
tengah/pusat 27.9
dalam 22.3
2126.1
2883.5
2386
0
2000
4000
G. apus D. asper D. giganteus
Gambar 55. Posisi nilai keteguhan tekan pada 3 jenis bambu
Ket
eguhan
tek
an /
/ se
rat
(kg/c
m2)
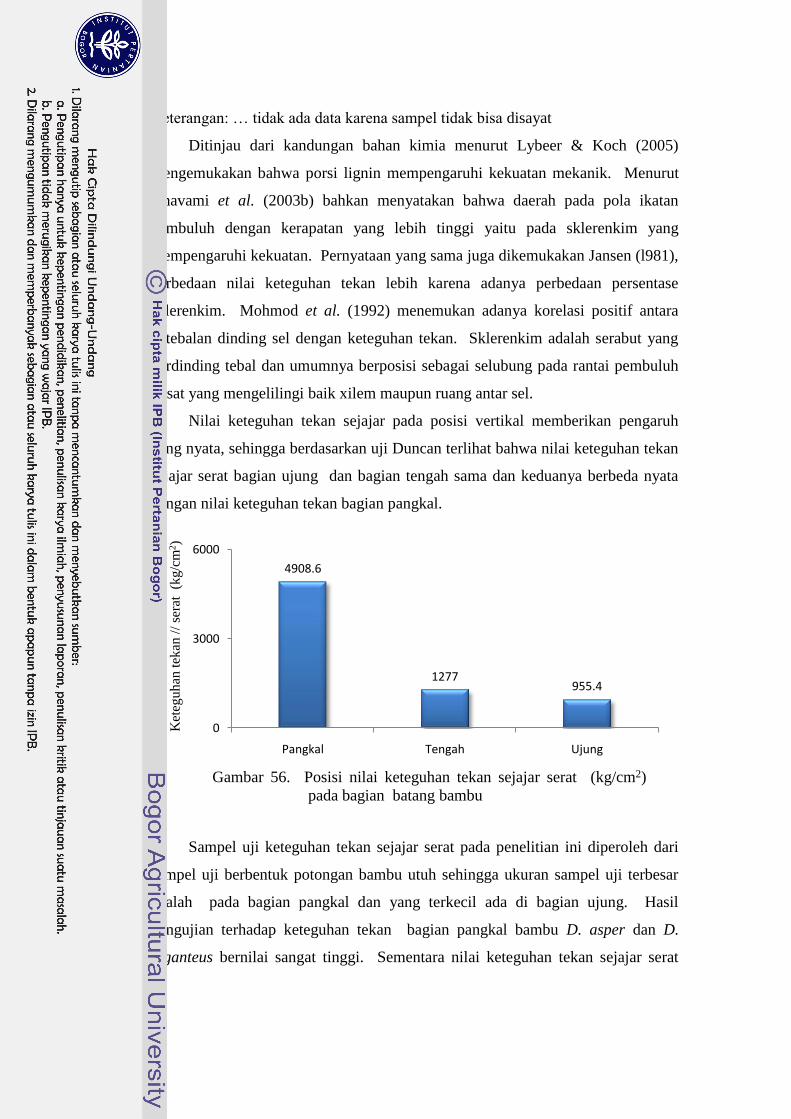
Keterangan: … tidak ada data karena sampel tidak bisa disayat
Ditinjau dari kandungan bahan kimia menurut Lybeer & Koch (2005)
mengemukakan bahwa porsi lignin mempengaruhi kekuatan mekanik. Menurut
Ghavami et al. (2003b) bahkan menyatakan bahwa daerah pada pola ikatan
pembuluh dengan kerapatan yang lebih tinggi yaitu pada sklerenkim yang
mempengaruhi kekuatan. Pernyataan yang sama juga dikemukakan Jansen (l981),
perbedaan nilai keteguhan tekan lebih karena adanya perbedaan persentase
sklerenkim. Mohmod et al. (1992) menemukan adanya korelasi positif antara
ketebalan dinding sel dengan keteguhan tekan. Sklerenkim adalah serabut yang
berdinding tebal dan umumnya berposisi sebagai selubung pada rantai pembuluh
pusat yang mengelilingi baik xilem maupun ruang antar sel.
Nilai keteguhan tekan sejajar pada posisi vertikal memberikan pengaruh
yang nyata, sehingga berdasarkan uji Duncan terlihat bahwa nilai keteguhan tekan
sejajar serat bagian ujung dan bagian tengah sama dan keduanya berbeda nyata
dengan nilai keteguhan tekan bagian pangkal.
Sampel uji keteguhan tekan sejajar serat pada penelitian ini diperoleh dari
sampel uji berbentuk potongan bambu utuh sehingga ukuran sampel uji terbesar
adalah pada bagian pangkal dan yang terkecil ada di bagian ujung. Hasil
pengujian terhadap keteguhan tekan bagian pangkal bambu D. asper dan D.
giganteus bernilai sangat tinggi. Sementara nilai keteguhan tekan sejajar serat
4908.6
1277955.4
0
3000
6000
Pangkal Tengah Ujung
Ket
eguhan
tek
an /
/ se
rat
(kg/c
m2)
Gambar 56. Posisi nilai keteguhan tekan sejajar serat (kg/cm2)
pada bagian batang bambu
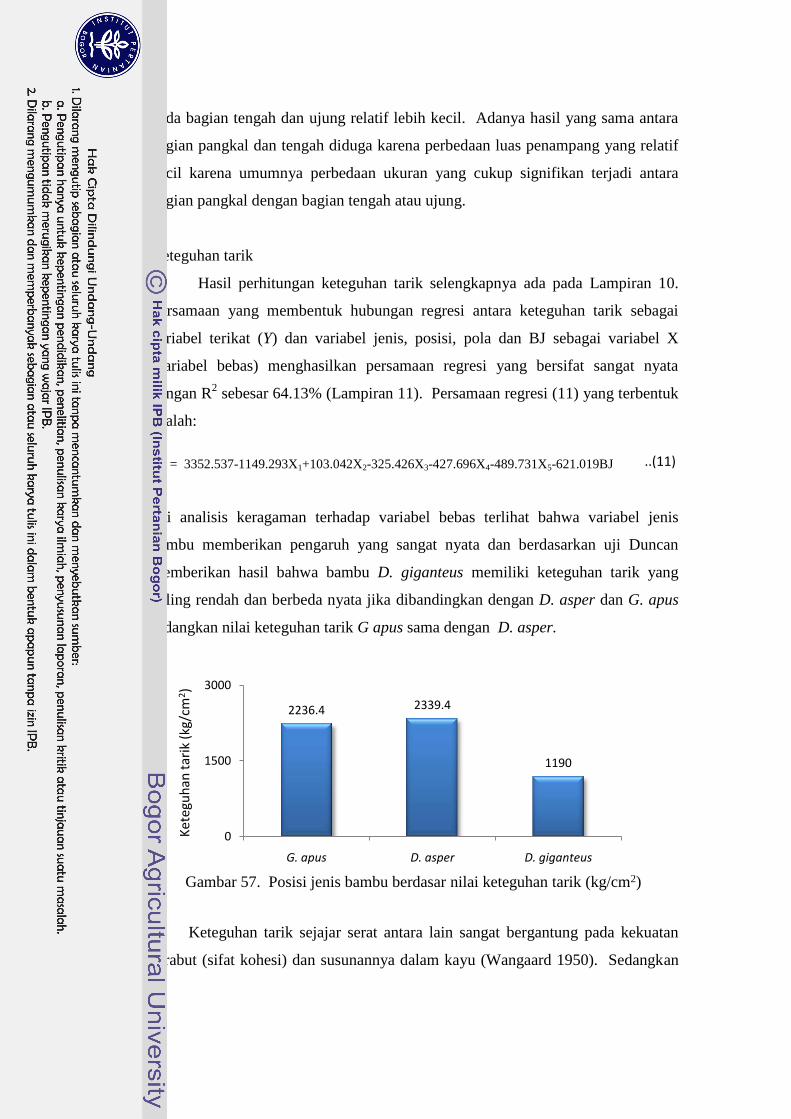
pada bagian tengah dan ujung relatif lebih kecil. Adanya hasil yang sama antara
bagian pangkal dan tengah diduga karena perbedaan luas penampang yang relatif
kecil karena umumnya perbedaan ukuran yang cukup signifikan terjadi antara
bagian pangkal dengan bagian tengah atau ujung.
Keteguhan tarik
Hasil perhitungan keteguhan tarik selengkapnya ada pada Lampiran 10.
Persamaan yang membentuk hubungan regresi antara keteguhan tarik sebagai
variabel terikat (Y) dan variabel jenis, posisi, pola dan BJ sebagai variabel X
(variabel bebas) menghasilkan persamaan regresi yang bersifat sangat nyata
dengan R2 sebesar 64.13% (Lampiran 11). Persamaan regresi (11) yang terbentuk
adalah:
Uji analisis keragaman terhadap variabel bebas terlihat bahwa variabel jenis
bambu memberikan pengaruh yang sangat nyata dan berdasarkan uji Duncan
memberikan hasil bahwa bambu D. giganteus memiliki keteguhan tarik yang
paling rendah dan berbeda nyata jika dibandingkan dengan D. asper dan G. apus
sedangkan nilai keteguhan tarik G apus sama dengan D. asper.
Keteguhan tarik sejajar serat antara lain sangat bergantung pada kekuatan
serabut (sifat kohesi) dan susunannya dalam kayu (Wangaard 1950). Sedangkan
2236.4 2339.4
1190
0
1500
3000
G. apus D. asper D. giganteus
Gambar 57. Posisi jenis bambu berdasar nilai keteguhan tarik (kg/cm2)
Ke
tegu
han
tar
ik (
kg/c
m2)
Y = 3352.537-1149.293X1+103.042X2-325.426X3-427.696X4-489.731X5-621.019BJ ..(11)

Janssen (1981) menyatakan bahwa kekuatan tarik tergantung kepada persentase
sklerenkim yang dimiliki oleh bambu. Hal ini diperkuat pula oleh Wang et al.
(2011) yang mengemukakan bahwa sklerenkim memberikan kontribusi dalam
stabilitas kekuatan, sementara Lo et al. (2004) menyatakan bahwa kerapatan
serabut dalam jaringan sklerenkim adalah indikator yang baik dalam pendugaan
kekuatan bambu.
Simpulan dan Saran
Nilai bobot jenis (BJ) tidak dipengaruhi oleh jenis bambu, posisi vertikal
batang dan pola pada bambu. Sementara nilai MOR secara bersama-sama
dipengaruhi oleh faktor–faktor yang diujikan yaitu jenis bambu, posisi vertikal
batang dan pola ikatan.pembuluh. Jenis bambu dan pola ikatan pembuluh
memberikan pengaruh yang berbeda nyata terhadap nilai MOR. Dendrocalamus
asper memberikan kontribusi yang terbesar terhadap nilai MOR (1004 kg/cm2)
sedangkan kontribusi yang terkecil diberikan oleh Dendrocalamus giganteus (505
kg/cm2). Demikian pula pola ikatan pembuluh 3 memberikan kontribusi yang
lebih tinggi terhadap nilai MOR (376 kg/cm2) dibandingkan pola ikatan pembuluh
4 (910 kg/cm2).
Nilai keteguhan lentur (MOE) tidak dipengaruhi oleh jenis bambu, posisi
vertikal, pola dan BJ pada bambu. Nilai keteguhan tekan dipengaruhi oleh jenis
bambu dan posisi vertikal dan hasil uji yang berbeda ditunjukan oleh keteguhan
tarik yang hanya dipengaruhi oleh jenis bambu saja. Dendrocalamus asper
cenderung memberikan kontribusi yang tertinggi terhadap nilai keteguhan tekan
sejajar serat (2884 kg/cm2) dibandingkan Dendrocalamus giganteus (2386
kg/cm2) dan Gigantochloa apus (2126 kg/cm
2). Hasil uji yang berbeda ditunjukan
oleh kekuatan keteguhan tarik yang hanya dipengaruhi oleh jenis bambu saja.
Dendrocalamus asper memberikan kontribusi yang tertinggi pada nilai keteguhan
tarik (2340 kg/cm2) sedangkan Dendrocalamus giganteus memberikan kontribusi
yang terendah (1190 kg/cm2).
Di antara berbagai jenis bambu yang diujikan, D. asper memiliki sifat
mekanis yang tertinggi yaitu dalam nilai MOR, keteguhan tekan sejajar serat, dan
keteguhan tarik. D. giganteus memiliki sifat mekanis yang terendah terutama

dalam nilai MOR dan keteguhan tarik. Sementara G. apus mempunyai nilai
keteguhan tekan yang terendah dibandingkan bambu yang lain.
Hasil pengujian pada berbagai variabel yang mempengaruhi nilai sifat
mekanis, pola ikatan pembuluh muncul sebagai salah satu variabel yang
berpengaruh terhadap nilai MOR. Hal ini berarti bahwa pola ikatan pembuluh
dapat dipertimbangkan sebagai variabel yang berperan dalam sifat mekanis.
Mengingat pemanfaatan bambu untuk konstruksi sangat dipengaruhi oleh
kualitas kekuatan mekanik, maka perlu dibuat suatu standar kekuatan untuk
bambu. Keberadaan pola ikatan pembuluh diharapkan dapat menjadi bagian
pengelompokan bambu berdasarkan kekuatannya seperti halnya standar kekuatan
kayu.
DAFTAR PUSTAKA
[ASTM] American Society for Testing and Materials. D 143-94 (Reapproved
2000). Standard test methods for small clear specimens of timber. Astm
Intl.
[BMTPC] Building Materials & Technology Promotion Council. 2007. The
Technological base of the building materials industry bamboo in housing &
building construction. Ministry of Housing and Urban Proverty Alleviation,
Government of India.
Draper N, Smith H. 1992. Analisis Regresi Terapan. Ed ke-2. Jakarta: PT
Gramedia Pustaka Utama.
Erakhrumen AA, Ogunsawo OY. 2009. Water absorption, anti-swell efficiency,
and dimensional stability properties of neem seed-oil treated wild grown
Bambusa vulgaris Schrad.Ex J.C. Wendl. In Southwest Nigeria.
BioResources 4(4): 1417-1429.
Espiloy ZB, Tesoro FO. 1988. Bamboo Research in the Philippines. Di dalam:
Rao R, Gnanaharan R, Sastry CB. Bamboos Current Research Proceedings
of the International Bamboo Workshop; India.
Ghavami K, Rodrigues CS, Paciornik S. 2003a. Bamboo: functionally graded
composite material. J civ eng (building and housing) 4 (1): 1-10

Ghavami K, Allameh SM, Sanchez ML, Soboyejowo. 2003b. Multiscale study
of bamboo Phyllostachys edulis. Departement of civil engineering, Rio de
Janeiro.
Hamdan H, Anwar U M K, Zaidon A, Tamizi M M. 2009. Mechanical properties
and failure behavior of Gigantochloa scortechinii. J Tropic For Sci 21(4):
336-334.
Hisham N, Mohmod AL, Sulaiman O. 2003. Variation of moisture content and specific
gravity of Gigantochloa scortechinii Gamble along the internodes sixth Height.
World Forestry Congress XII. Canada.
ISO/TC165N314. 2001, Determination of physical and mechanical properties of
bamboo. INBAR.
Jansen JJA. 1981. The relationship between the mechanical properties and the
biological and chemical composition of bamboo. Pp. 27-32 in Higuchi, T.
(Ed.) Bamboo Production and Utilization. Proceedings of the Congress
Group 5.3A. Production and Utilization of Bamboo and Related Species.
XVII IUFRO World Congress. September 6 -17, 1981. Kyoto, Japan.
Janssen JJA. 1987. Bamboo research at the Eindhoven University of Technology.
Lybeer1 B, Koch G. 2005. Lignin distribution in the tropical bamboo spesies
Gigantochloa levisia IAWA J, Vol. 26 (4): 443–456
Liese W. 1985. Anatomy and properties of bamboo. Di dalam : Rao, A.N.,
Dhanarajan, G. dan Sastry, C.B, editor. Recent Research on Bamboo.
Proceedings of the International Bamboo Workshop, Hangzholu, People's
Republic of China, 6-14 Oktober, 1985. Academy of Forestry, People's
Republic of China & International ,Development Research Centre, Canada,
hal: 196-208.
Liese W. 1998. The anatomy of bamboo culms. INBAR Technology Report No
18.
Liese W. 2003. Structures of bamboo culm affecting its utilization. Di dalam
Xuhe C, Yiping L, Ying H, editor. Proceedings of International Workshop
on Bamboo Industrial Utilization. Hubei dan Xianning, Oktober 2003. hlm
6–10.
Lo T, Cui H, Leung H. 2004. The effect of fiber density on strength capacity of
bamboo. Matterials Letters 58 (21): 2595-2598.
Morisco. 2011. Pemberdayaan bambu untuk kesejahteraan rakyat dan kelestarian
lingkungan. Http://www.moriscobamboo.com/artikel_02.html.[17 Desember
2011).

Mohmod, AL. Amin, A. Kasim, J. Jusuh MZ. 1992. Effects of anatomical
characteristics on the physical and mechanical properties of Bambusa
blumeana. J Tropil For Sci 6(2): 159-170 159
Nuriyatin N. 2000. Studi analisa sifat-sifat dasar bambu pada beberapa tujuan
penggunaan [tesis]. Bogor: Jurusan Ilmu Pengetahuan Kehutanan, Institut
Pertanian Bogor.
Purwito. 2008. Standarisasi bambu sebagai bahan bangunan alternatif pengganti
kayu. Prosiding PPI Standardisasi 2008, 25 November 2008
Shao Z P, Zhou L, Liu Y M, Wu Z M, Arnaud C. 2010. Differences in
structure and strength between internode and node section of moso bamboo.
J Tropic For Sci 22 (2): 133-138.
Surjokusumo, HMS. 1997. Pemanfaatan bambu untuk bangunan. Dalam panel
diskusi bambu, 4 Desember 1997, Jakarta.
Wang X, Ren H, Zhang B, Fei B, Burgert I. 2011. Cell wall structure and
formation of maturing fibres of moso bamboo (Phyllostachys pubescens)
increase buckling resistance . J. R. Soc. Interface rsif20110462.
Wangaard FF. 1950. The Mechanical Properties of Wood. John Willey & Sons,
Inc. New York, Chapman & Hill Limited London.

IV. KORELASI POLA IKATAN PEMBULUH PADA
KANDUNGAN KIMIA 4 JENIS BAMBU
Abstrak
Informasi tentang sifat kimia bambu selama ini terbatas baik dari jenis
bambu maupun posisi pengambilan sampelnya, pengamatan variabel, dan
hubungan di antara variabel respons. Penelitian ini dilakukan untuk mendapatkan
informasi lengkap dengan mempertimbangkan pola ikatan pembuluh dan
melihat pengaruhnya pada sifat kimia bambu. Selain itu, diharapkan juga dapat
mengeksplorasi keberadaan pola ikatan pembuluh serta kontribusinya pada sifat
kimia bambu. Bahan penelitian adalah 4 jenis bambu yaitu, Arundinaria hundsii,
Cephalostachyum pergracile, Dendrocalamus giganteus, dan Dendrocalamus
asper yang diambil dari Kebun Raya Bogor dan di sekitar Fakultas Kehutanan
Insitut Pertanian Bogor. Variabel yang diamati adalah kandungan ekstraktif,
abu, lignin, alfa selulosa, dan pati yang dianalisis dengan berbagai pengujian
standar. Data diolah dengan analisi deskripsi dan analisis keragaman.
Kandungan ekstraktif pada berbagai jenis dan pola ikatan pembuluh bambu
berkisar dari 4% hingga 9.9%. Berdasarkan pengujian, interaksi antara jenis
bambu dan pola ikatan pembuluh berpengaruh pada kadar ekstraktif. Interaksi
antara jenis dan pola ikatan pembuluh bambu juga berpengaruh pada nilai kadar
abu. Kandungan abu bambu berkisar dari 1.6% hingga 4.3%. Adanya interaksi

antara jenis dan pola bambu berpengaruh pada nilai kadar lignin. Nilai kadar
lignin pada bambu yang diteliti berkisar dari 28.9% sampai 32%. Pengaruh
interaksi juga muncul dalam penentuan analisis kadar pati yang nilainya berkisar
dari 0.1% sampai 1.4%. Tidak ada interaksi antara jenis dan pola ikatan
pembuluh bambu yang mempengaruhi nilai kadar alfa selulosa, demikian pula
setiap jenis dan pola ikatan pembuluh bambu tidak berpengaruh pada nilai
respons. Kesimpulannya ialah interaksi antara jenis dan pola ikatan pembuluh
bambu berpengaruh pada beberapa sifat kimia kayu.
Kata kunci: sifat kimia bambu, pola ikatan pembuluh bambu, jenis bambu
Abstract
Information about chemical properties of bamboo is limited both in species
of the bamboo, position of sampling, observation variables, and relationship
among variables with response. This study was done to obtain complete
information, by considering vessel bundle pattern of bamboo and evaluate its
effect on the chemical properties of bamboo. In addition, it is expected to
explore the existence of the vessel bundle pattern of bamboo as well as its
contribution to the chemical properties of the bamboo. The material research is 4
species of bamboo which are Arundinaria hundsiii, Cephalostachyum pergracile,
Dendrocalamus giganteus and D. asper taken from the Bogor Botanical Garden
and the area of Faculty of Forestry, Bogor Agricultural University. The observed
variables were content of extractives, ash, lignin, alpha cellulose, and starch
using various testing standards. The data was processed by description analysis
and varian analysis. The extractive content of the species various and the vessel
bundle patterns of bamboo range from 4% to 9.9%. The results showed that
interaction between the species and the vessel bundle patterns of bamboo
influences the extractive content. Ash content of the bamboo range from 1.6% to
4.3%. The interaction between the species and vessel bundle pattern of bamboo
effects the lignin level. Lignin in bamboo in this study ranges from 28.9% to
31.9%. The interaction effect also appears in the starch content, which ranges
from 0.1% to 1%. There was no interaction between the species and the vessel
bundle pattern of bamboo that affect the alpha cellulose content. This study
concludes that interaction between the species and the vessel bundle pattern of
bamboo affect some chemical properties of the bamboo.
Keyword: bamboo chemical properties, bamboo bonding pattern, bamboo species
Pendahuluan
Sifat-sifat kimia beragam berdasar spesies, kondisi pertumbuhan, umur,
bagian batang bambu dan faktor-faktor eksternal topografi dan efek musim ( Lwin
et al. 2007). Bambu terdiri atas sekitar 50-70% holoselulosa, 30% pentosan, dan
20-25% lignin. Kandungan silika 0.5-5% dan mempengaruhi pemotongan bambu
dan mutu pulping dan umumnya ada di daerah epidermis (Liese l992) sedangkan

menurut Qisheng et al. (2001) komponen organik bambu sama seperti kayu
terutama terdiri atas selulosa (±55%), lignin (±25%) dan hemiselulosa (pentosan
±20%) (Lwin et al. 2007). Menurut Liese (2006), bambu memiliki beberapa
ekstraktif yang disimpan dalam dinding sel, dalam sel lumina sebagai tambahan
korteks atau dalam lakuna. Bahan organik pada bambu antara lain adalah lilin dan
pati. Bahan anorganik seperti silika adalah penyusun utama epidermis dengan
nilai berkisar diantara 1.5% dan 6.4%.
Kebanyakan studi menyediakan informasi secara umum dari beberapa spesies
bambu dan terpusat pada satu spesies atau hanya satu asfek saja (Li 2004).
Selama ini informasi sifat kimia yang diperoleh berasal dari jenis-jenis bambu
tertentu dengan perolehan beragam dari berbagai posisi vertikal dan posisi
horizontal. Sebagai upaya pemanfaatan bambu secara optimum memang
diperlukan berbagai informasi yang akurat yang sifatnya lebih mendalam.
Penelitian ini bertujuan menggali sifat kimia bambu khususnya yang terkait
dengan pola ikatan pembuluh karena setiap jenis bambu tampil khas dalam
penampang melintang batang. Melalui hasil penelitian ini diharapkan dapat
diperoleh informasi yang lebih lengkap termasuk mengenai posisi pola ikatan
pembuluh dalam kontribusinya terhadap sifat kimia.
Bahan dan Metode
Bahan
Bahan penelitian adalah 4 jenis bambu yang telah berumur 3-4 tahun serta
memiliki pola ikatan pembuluh 1 sampai 4 yang ditentukan berdasarkan panduan
penetapan pola ikatan pembuluh bambu oleh Grosser dan Liese (1971) dengan
setiap ulangan 2 kali. Spesies bambu tersebut ialah Arundinaria hundsii (Ah),
Cephalostachyum pergracile (Cp) dan Dendrocalamus asper (Da), D. giganteus
(Dg) yang diambil dari Kebun Raya Bogor dan dari daerah Fakultas Kehutanan
Institut Pertanian Bogor.
Penentuan Komponen Kimia Bambu
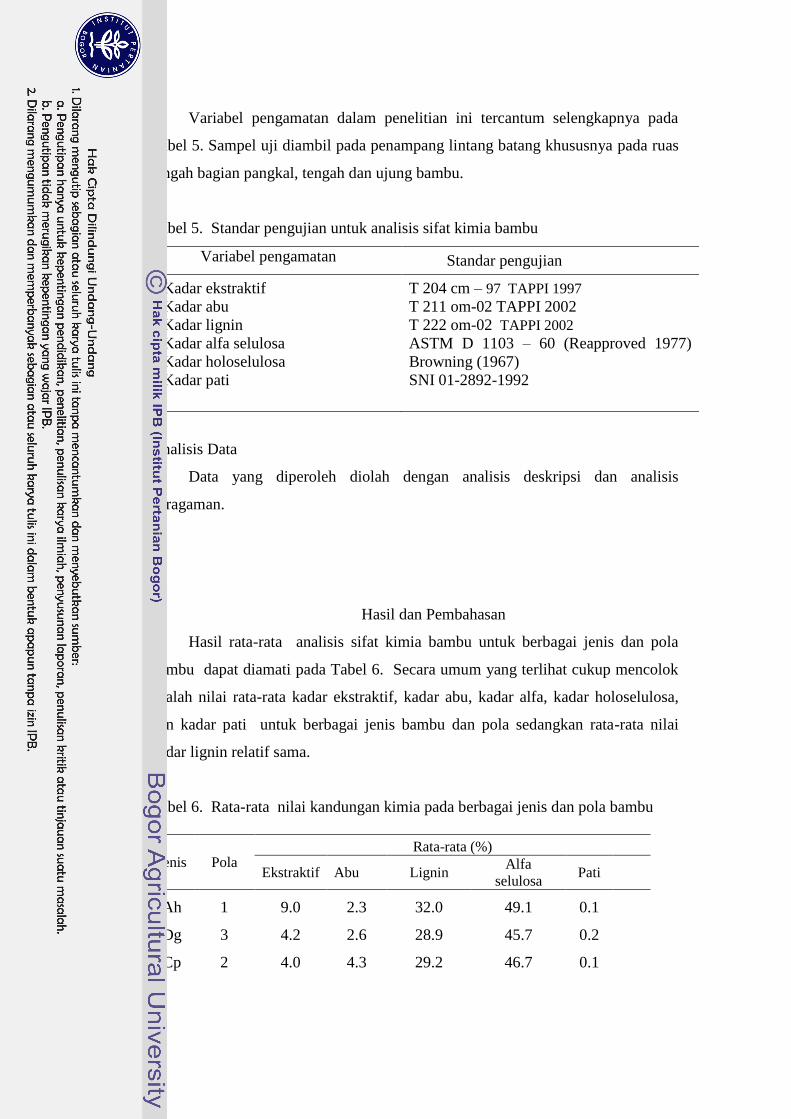
Variabel pengamatan dalam penelitian ini tercantum selengkapnya pada
Tabel 5. Sampel uji diambil pada penampang lintang batang khususnya pada ruas
tengah bagian pangkal, tengah dan ujung bambu.
Tabel 5. Standar pengujian untuk analisis sifat kimia bambu
Variabel pengamatan Standar pengujian
Kadar ekstraktif
Kadar abu
Kadar lignin
Kadar alfa selulosa
Kadar holoselulosa
Kadar pati
T 204 cm – 97 TAPPI 1997
T 211 om-02 TAPPI 2002
T 222 om-02 TAPPI 2002
ASTM D 1103 – 60 (Reapproved 1977)
Browning (1967)
SNI 01-2892-1992
Analisis Data
Data yang diperoleh diolah dengan analisis deskripsi dan analisis
keragaman.
Hasil dan Pembahasan
Hasil rata-rata analisis sifat kimia bambu untuk berbagai jenis dan pola
bambu dapat diamati pada Tabel 6. Secara umum yang terlihat cukup mencolok
adalah nilai rata-rata kadar ekstraktif, kadar abu, kadar alfa, kadar holoselulosa,
dan kadar pati untuk berbagai jenis bambu dan pola sedangkan rata-rata nilai
kadar lignin relatif sama.
Tabel 6. Rata-rata nilai kandungan kimia pada berbagai jenis dan pola bambu
Jenis Pola Rata-rata (%)
Ekstraktif Abu Lignin Alfa
selulosa Pati
Ah 1 9.0 2.3 32.0 49.1 0.1
Dg 3 4.2 2.6 28.9 45.7 0.2
Cp 2 4.0 4.3 29.2 46.7 0.1

Da 4 7.0 1.6 29.2 44.4 1.0
Da 3 9.9 3.0 30.7 40.2 1.4
Kadar ekstraktif hasil penelitian pada berbagai jenis bambu dan pola
berkisar dari 4.o% (Cephalostachyum pergracile) hingga 9.9% (Dendrocalamus
asper). Hasil penelitian Li (1983) pada bambu Phyllostachys pubescens ternyata
nilai kandungan ekstraktifnya adalah sekitar 7%. Demikian pula dengan
kandungan abu hasil penelitian untuk berbagai jenis bambu berkisar dari 1.6%
hingga 4.3% sedangkan kandungan abu untuk bambu Phyllostachys pubescens
rata-rata 1.3% (Li 2007). Kandungan lignin hasil penelitian berkisar dari 28.9%
hingga 32.0%. Hasil penelitian memberikan hasil yang relatif lebih tinggi
dibandingkan dengan lignin rata-rata bambu yaitu 20-25% (Liese 1992). Kisaran
kandungan alfa selulosa hasil penelitian ini adalah 40.2% hingga 49.1%
sedangkan kandungan alfa selulosa hasil penelitian Li et al (2007) adalah 47%.
Kadar pati yang diperoleh melalui hasil penelitian adalah berkisar dari 0.1%-0.4%
sedangkan kadar pati pada bambu D. asper berkisar dari 0.27%-2.8% (Sulthoni
1985). Secara umum hasil penelitian tidak jauh berbeda dengan hasil penelitian
lain. Perbedaan diduga disebabkan karena perbedaan lokasi/tempat tumbuh,
spesies dan musim (Lwin et al. 2007).
Untuk mengetahui interaksi antara jenis dan pola yang berpengaruh pada
nilai kandungan kimia maka semua variabel pengamatan sifat kimia bambu harus
diamati khususnya dengan membandingkan jenis bambu yang berbeda tetapi
memiliki pola yang sama (D. giganteus) serta jenis bambu yang sama dengan pola
yang berbeda (D. asper) untuk melihat interaksi jenis dan pola yang
mempengaruhi kandungan kimia bambu. Tahap selanjutnya untuk memudahkan
dalam pemahaman olahan data maka penulisan jenis bambu dan pola dapat
digantikan oleh notasi Ah/1, Dg/3, dan seterusnya.
Kadar ekstraktif
Nilai rata-rata kandungan ekstraktif pada berbagai jenis dan pola bambu
nilainya berkisar dari 4.0% (C.pergracile) sampai 9.9% (D. asper) (Gambar 58).
Pengolahan lebih lanjut terhadap data diawali dengan memeriksa kemungkinan
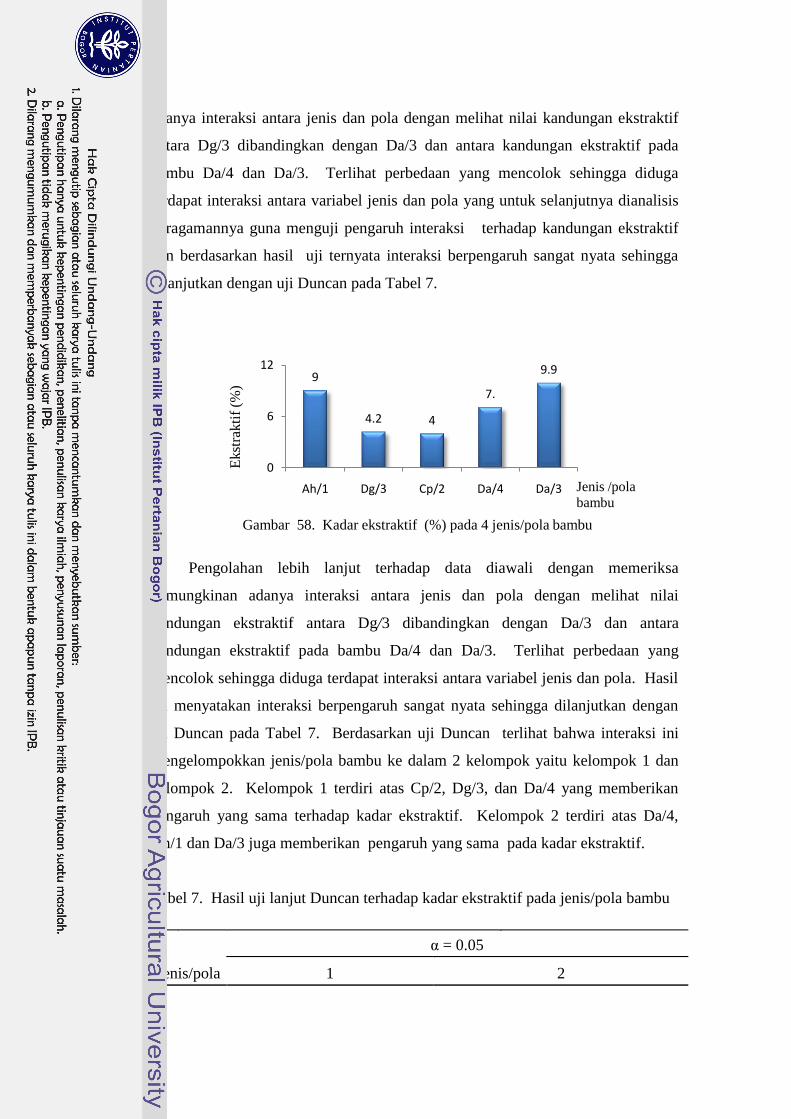
adanya interaksi antara jenis dan pola dengan melihat nilai kandungan ekstraktif
antara Dg/3 dibandingkan dengan Da/3 dan antara kandungan ekstraktif pada
bambu Da/4 dan Da/3. Terlihat perbedaan yang mencolok sehingga diduga
terdapat interaksi antara variabel jenis dan pola yang untuk selanjutnya dianalisis
keragamannya guna menguji pengaruh interaksi terhadap kandungan ekstraktif
dan berdasarkan hasil uji ternyata interaksi berpengaruh sangat nyata sehingga
dilanjutkan dengan uji Duncan pada Tabel 7.
Pengolahan lebih lanjut terhadap data diawali dengan memeriksa
kemungkinan adanya interaksi antara jenis dan pola dengan melihat nilai
kandungan ekstraktif antara Dg/3 dibandingkan dengan Da/3 dan antara
kandungan ekstraktif pada bambu Da/4 dan Da/3. Terlihat perbedaan yang
mencolok sehingga diduga terdapat interaksi antara variabel jenis dan pola. Hasil
uji menyatakan interaksi berpengaruh sangat nyata sehingga dilanjutkan dengan
uji Duncan pada Tabel 7. Berdasarkan uji Duncan terlihat bahwa interaksi ini
mengelompokkan jenis/pola bambu ke dalam 2 kelompok yaitu kelompok 1 dan
kelompok 2. Kelompok 1 terdiri atas Cp/2, Dg/3, dan Da/4 yang memberikan
pengaruh yang sama terhadap kadar ekstraktif. Kelompok 2 terdiri atas Da/4,
Ah/1 dan Da/3 juga memberikan pengaruh yang sama pada kadar ekstraktif.
Tabel 7. Hasil uji lanjut Duncan terhadap kadar ekstraktif pada jenis/pola bambu
Jenis/pola
α = 0.05
1 2
9
4.2 4
7.
9.9
0
6
12
Ah/1 Dg/3 Cp/2 Da/4 Da/3
Gambar 58. Kadar ekstraktif (%) pada 4 jenis/pola bambu
Jenis /pola
bambu
Ekst
rakti
f (%
)

Cp/2 4.0
Dg/3 4.2
Da/4 7.0 7.0
Ah/1 9.0
Da/3 9.9
Apabila ditelusuri lebih lanjut dari pengelompokan interaksi itu sendiri
terlihat bahwa pengaruh interaksi pada kelompok 2 (kecuali pada Da/4)
memberikan pengaruh yang lebih besar terhadap nilai kadar ekstraktif
dibandingkan dengan kelompok 1. Kelompok 1 terdiri atas bambu-bambu dengan
pola 2 dan 3 sedangkan kelompok 2 terdiri atas bambu-bambu yang memiliki
pola 1 dan 3. Analisa lebih lanjut terhadap nilai kadar ekstraktif tidak akan
terlepas dari letak ekstraktif itu sendiri dalam batang bambu yang menurut Liese
(2006) bambu memiliki ekstraktif yang disimpan dalam dinding sel, dalam rongga
sel atau dalam lakuna (rongga batang bambu). Dengan demikian kandungan
ekstraktif suatu bambu tidak akan terlepas dari struktur selnya. Selanjutnya analis
akan diutamakan pada pola 3 yang terpisah pengelompokannnya. Kadar ekstraktif
yang berasal dari D. asper mempunyai nilai yang lebih besar dibandingkan
ekstraktif dari bambu D. giganteus. Pengamatan terhadap struktur sel khususnya
ketebalan dinding sel dikombinasikan dengan persen serabut selengkapnya dapat
dilihat pada Tabel 8. Tebal dinding sel serabut D. asper bagian tengah dan ujung
(6.3 µm) lebih kecil daripada tebal dinding sel serabut D. giganteus (7.2 µm),
namun persentase serabut yang lebih tinggi (34.4%) dimiliki oleh bambu D. asper
sementara D. giganteus mempunyai persentase serabut 28.8%. Dengan demikian
karena jumlah sel D. asper lebih banyak dengan dinding sel yang relatif tebal
maka bambu D. asper berpeluang mendapatkan kadar ekstraktif yang lebih
tinggi. Hal ini sesuai dengan hasil penelitian dengan nilai kadar ekstraktif D.
asper lebih besar yaitu 9.9% sedangkan kadar ekstratif D. giganteus hanya 4.2%.
Bambu C. pergracile yang memiliki pola 2 ternyata mempunyai tebal
dinding serabut sebesar 5.8 µm (Tabel 8) relatif lebih kecil dibandingkan
ketebalan dinding serabut A. hundsii (5.9 µm) tapi C. pergracile memiliki

persentase serabut lebih tinggi (40.3% dibandingkan 38.4%). Kondisi seperti ini
sulit menjelaskan adanya kontribusi yang berbeda terhadap kadar ekstraktif kedua
jenis bambu tersebut. Seperti yang telah dikemukakan oleh Liese (2006) bahwa
ekstraktif juga ada pada rongga sel. Pengamatan terhadap lebar rongga sel serabut
A. hundsii (6.9 µm ) yang lebih lebar dibandingkan diameter rongga sel serabut
pada C. pergracile (3.4 µm) maka kemungkinan akan ada perbedaan kandungan
ekstraktif. Hasil analisa lebih lanjut terhadap kandungan ekstraktif (Tabel 7)
memperlihatkan hasil bahwa kandungan ekstraktif pada bambu A. hundsii lebih
tinggi dibandingkan dengan kandungan ekstraktif C. pergracile.
Tabel 8. Ketebalan dinding sel dan persen serabut D. asper, D. giganteus,
C. pergracile dan A. hundsii
Jenis bambu Bagian Pola Tebal dinding
(mikron)
Persen serabut
(%)

Ekstraktif memainkan peranan penting dalam menentukan penggunaan
sejumlah spesies selain itu juga dapat mengontrol keawetan, warna, bau dan rasa.
Dalam beberapa spesies, ekstraktif yang bersifat fenolik menyediakan ketahanan
terhadap busuk dan serangan serangga (Liese 2006). Di antara bambu-bambu
yang diujikan terlihat bahwa bambu A. hundsii (pola 1) dan D. asper (pola 3)
memiliki kandungan ekstraktif yang relatif lebih besar sehingga diduga memiliki
keawetan yang lebih tinggi.
Kadar abu
Nilai kandungan abu rata-rata selengkapnya ditampilkan pada Gambar 59.
Perbandingan antara D/3 dengan Da/3 dan Da/4 dengan Da/3 kelihatan berbeda
secara mencolok sehingga kemungkinan ada interaksi antara jenis dan pola.
D.asper
D.giganteus
C.pergracile
A.hundsii
Tengah
Ujung
Pangkal
Tengah
Ujung
Pangkal
Tengah
Ujung
Tengah
3
3
3
3
3
2
2
2
1
5.4
7.1
Rata2 6.3
6.3
8.4
6.8
Rata2 7.2
6.0
5.8
5.5
Rata2 5.8
5.9
33.8
35.0
Rata2 34.4
25.9
27.6
33.3
Rata2 28.8
39.5
45.3
36.1
40.3
38.4
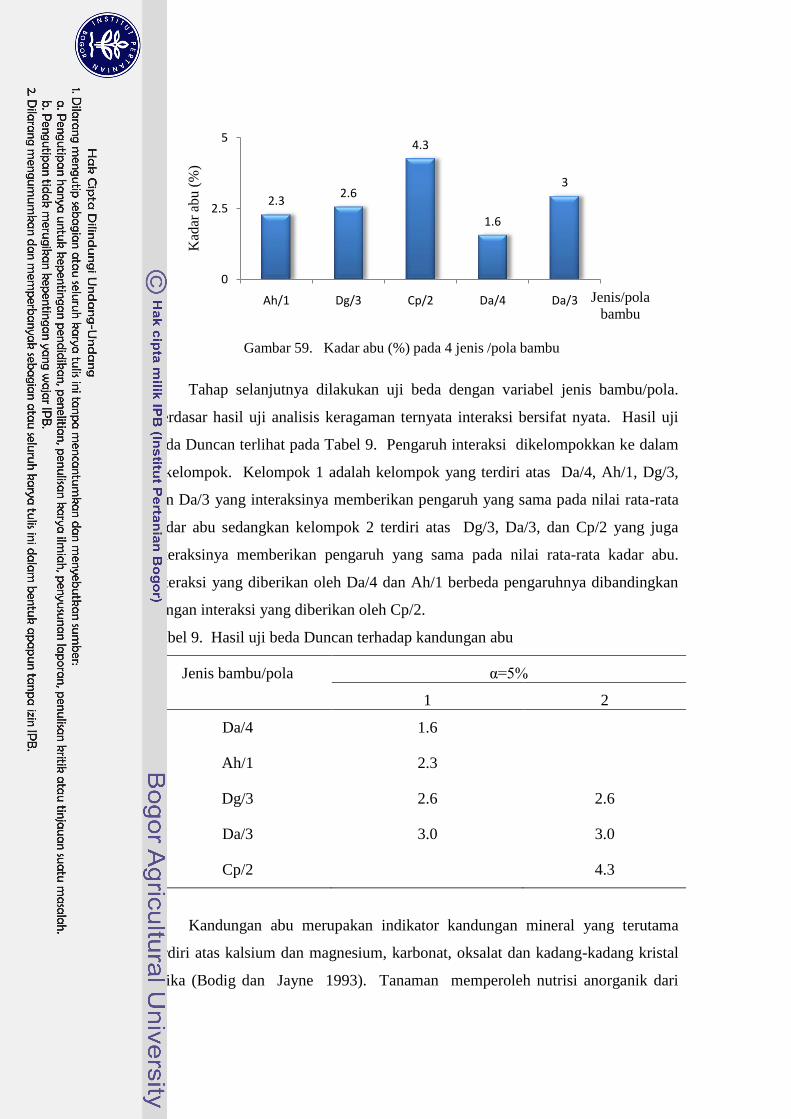
Tahap selanjutnya dilakukan uji beda dengan variabel jenis bambu/pola.
Berdasar hasil uji analisis keragaman ternyata interaksi bersifat nyata. Hasil uji
beda Duncan terlihat pada Tabel 9. Pengaruh interaksi dikelompokkan ke dalam
2 kelompok. Kelompok 1 adalah kelompok yang terdiri atas Da/4, Ah/1, Dg/3,
dan Da/3 yang interaksinya memberikan pengaruh yang sama pada nilai rata-rata
kadar abu sedangkan kelompok 2 terdiri atas Dg/3, Da/3, dan Cp/2 yang juga
interaksinya memberikan pengaruh yang sama pada nilai rata-rata kadar abu.
Interaksi yang diberikan oleh Da/4 dan Ah/1 berbeda pengaruhnya dibandingkan
dengan interaksi yang diberikan oleh Cp/2.
Tabel 9. Hasil uji beda Duncan terhadap kandungan abu
Jenis bambu/pola α=5%
1 2
Da/4
Ah/1
Dg/3
Da/3
Cp/2
1.6
2.3
2.6
3.0
2.6
3.0
4.3
Kandungan abu merupakan indikator kandungan mineral yang terutama
terdiri atas kalsium dan magnesium, karbonat, oksalat dan kadang-kadang kristal
silika (Bodig dan Jayne 1993). Tanaman memperoleh nutrisi anorganik dari
2.32.6
4.3
1.6
3
0
2.5
5
Ah/1 Dg/3 Cp/2 Da/4 Da/3 Jenis/pola
bambu
Gambar 59. Kadar abu (%) pada 4 jenis /pola bambu
Kad
ar a
bu (
%)

dalam tanah atau dari air hujan (Austin et al. 1974) sehingga nutrisi yang diambil
tergantung dari spesies, tanah, iklim dan faktor eksternal lain (Chen et al. 1987,
Kozlowskin & Pallardy 1997).
Pada tanaman bambu Phylostahys pubescens, akumulasi nutrisi mineral
bervariasi dari satu bagian ke bagian yang lain. Sebagai contoh nutrisi mineral
pada rizoma lebih tinggi dibandingkan di bagian batang namun lebih rendah
dibanding daun. Tapi untuk akumulasi K, Ca dan Mn lebih besar di bagian batang
dibandingkan rizoma (Pai-hui 1985). Sebagai pendukung transport air dan nutrisi
adalah ikatan pembuluh (Wang et al. 2011) khususnya metaxilem yang terdiri atas
2 pembuluh besar (Liese dan Kumar 2003). Unsur hara diserap oleh akar dalam
bentuk cairan bersama-sama dengan air dan aliran ini mengalir melalui unsur
metaxilem (Salisbury dan Ross 1992). Dengan demikian metaxilem adalah
bagian yang sangat penting dalam transportasi hara sehingga keberadaannya
termasuk ukuran diameter metaxilem akan mempengaruhi proses penyaluran
unsur hara. Metaxilem pada penampang lintang bambu terlihat secara jelas
berada dalam setiap pola ikatan pembuluh sehingga untuk mengetahui jumlah
metaxilem harus diketahui pula jumlah pola ikatan dalam luasan tertentu
(kerapatan ikatan pembuluh). Tabel 10 menjelaskan tentang kerapatan ikatan
pembuluh dan ukuran metaxilem. Arundinaria hundsii dengan pola 1 memiliki
kerapatan ikatan pembuluh tertinggi namun dengan diameter metaxilem yang
terkecil, bambu D. asper dengan kerapatan ikatan pembuluh terkecil mempunyai
diameter metaxilem terbesar sedangkan C. pergracil dengan pola 2 memiliki
kerapatan dan diameter metaxilem yang berada diantara kedua bambu D. asper
dan A. hundsii. Hasil uji lanjut Duncan menyatakan bahwa yang berpengaruh
pada kontribusi kadar abu yang lebih tinggi adalah hasil dari interaksi antara jenis
bambu C. pergracil dengan pola 2. Berpengaruhnya perbedaan ukuran metaxilem
ini terkait dengan akumulasi nutrisi yang berbeda terkait dengan laju siklus nutrisi
yang berbeda pula (Kozlowskin & Pallardy 1997). Bambu A.hundsii dengan pola
1 memiliki kerapatan ikatan pembuluh yang tertinggi dengan diameter metaxilem
yang paling kecil akan mengalami hambatan yang paling besar karena adanya
gaya tarik antarlarutan dengan dinding sel metaxilem/adhesi akan lebih besar
dibanding kohesinya juga ditambah oleh gaya tahanan dari gravitasi. Bambu

D.asper/4 memiliki kerapatan ikatan pembuluh yang rendah dengan diameter
yang besar juga akan mengalami hambatan dalam mengalirkan nutrisinya karena
selain oleh gaya adhesi dan kohesi yang besar juga tertahan oleh gaya gravitasi.
Dengan demikian kondisi optimum dalam proses penyerapan akan dimiliki oleh
bambu C.pergracil/2 dengan kerapatan dan diameter metaxilem berada di antara
bambu D.asper/4 dan A.hundsii/1 sehingga siklus nutrisi berjalan dengan lancar
dan otomatis akan memberikan kontribusi terhadap kandungan abu yang tinggi
pula.
Tabel 10. Kerapatan ikatan pembuluh dan diameter metaxilem bambu
Jenis bambu/pola Kerapatan ikatan pembuluh (/mm2) Diameter metaxilem(µm)
A. hundsii/1 3.2 80.2
D. asper/4 0.6 203.6
C. pergracil/2 2.8 118.4
Lignin
Nilai rata-rata kandungan lignin pada beberapa jenis bambu/pola tercantum
selengkapnya pada Gambar 60. Untuk melihat interaksi antara jenis dan pola
maka dibandingkan antara Dg/3 dengan Da/3 dan Da/4 dengan Da/3, terlihat
terdapat perbedaan yang mencolok sehingga ada dugaan terdapat interaksi.
Berdasar perbedaan tersebut maka dilakukan uji beda dengan variabel jenis
bambu/pola dan teruji bahwa interaksi antar jenis dan pola memberikan pengaruh
yang sama terhadap kadar lignin. Hal ini terjadi karena perbedaan kandungan
lignin antar jenis bambu/pola relatif kecil.
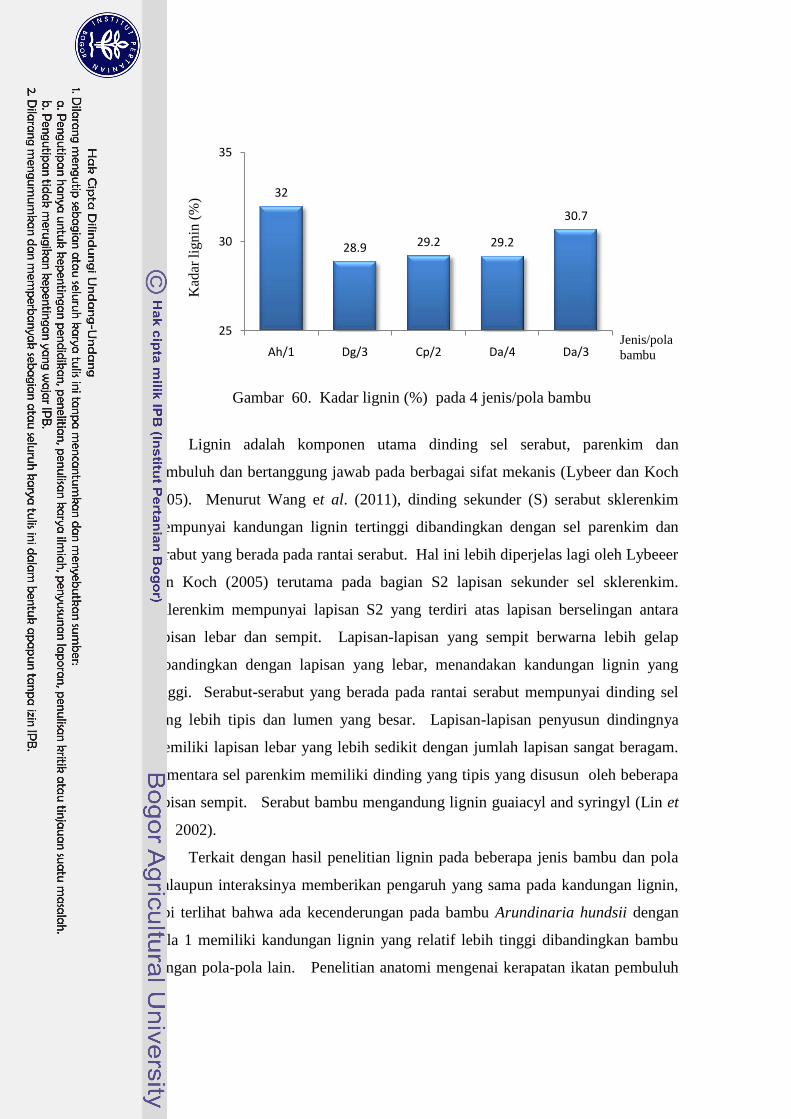
Lignin adalah komponen utama dinding sel serabut, parenkim dan
pembuluh dan bertanggung jawab pada berbagai sifat mekanis (Lybeer dan Koch
2005). Menurut Wang et al. (2011), dinding sekunder (S) serabut sklerenkim
mempunyai kandungan lignin tertinggi dibandingkan dengan sel parenkim dan
serabut yang berada pada rantai serabut. Hal ini lebih diperjelas lagi oleh Lybeeer
dan Koch (2005) terutama pada bagian S2 lapisan sekunder sel sklerenkim.
Sklerenkim mempunyai lapisan S2 yang terdiri atas lapisan berselingan antara
lapisan lebar dan sempit. Lapisan-lapisan yang sempit berwarna lebih gelap
dibandingkan dengan lapisan yang lebar, menandakan kandungan lignin yang
tinggi. Serabut-serabut yang berada pada rantai serabut mempunyai dinding sel
yang lebih tipis dan lumen yang besar. Lapisan-lapisan penyusun dindingnya
memiliki lapisan lebar yang lebih sedikit dengan jumlah lapisan sangat beragam.
Sementara sel parenkim memiliki dinding yang tipis yang disusun oleh beberapa
lapisan sempit. Serabut bambu mengandung lignin guaiacyl and syringyl (Lin et
al. 2002).
Terkait dengan hasil penelitian lignin pada beberapa jenis bambu dan pola
walaupun interaksinya memberikan pengaruh yang sama pada kandungan lignin,
tapi terlihat bahwa ada kecenderungan pada bambu Arundinaria hundsii dengan
pola 1 memiliki kandungan lignin yang relatif lebih tinggi dibandingkan bambu
dengan pola-pola lain. Penelitian anatomi mengenai kerapatan ikatan pembuluh
32
28.9 29.2 29.2
30.7
25
30
35
Ah/1 Dg/3 Cp/2 Da/4 Da/3Jenis/pola
bambu
Gambar 60. Kadar lignin (%) pada 4 jenis/pola bambu
Kad
ar lig
nin
(%)

pada pola 1 memperlihatkan nilai yang paling tinggi dibandingkan pola-pola lain.
Serabut pada pola 1 ada sebagai sklerenkim (tidak ada serabut yang berposisi
dalam rantai serabut) sehingga dapat dipastikan semua serabut berdinding tebal
dengan kandungan lignin yang tinggi. Dalam pengamatan di lapangan diduga
kontribusi lignin dan kerapatan memberikan pengaruh pada kondisi pertumbuhan
batang yang berdiri tegak. Sama halnya dengan kandungan serabut pada bambu
Cephalostachym pergracile (Cp) dengan pola 2 terdiri atas serabut sklerenkim
sehingga memiliki kandungan lignin yang relatif tinggi pula.
Kandungan lignin pada bambu Dendrocalamus asper (Da) memiliki nilai
yang berbeda antara pola 3 dengan pola 4. Kedua pola mempunyai serabut dalam
posisi sebagai rantai serabut, tapi kandungan serabut pada posisi sebagai rantai
serabut dalam pola 3 lebih sedikit dibandingkan dengan pola 4. Hasil
perhitungan nilai rata-rata kandungan serabut pada pola 3 adalah 34% sedangkan
pada pola 4 adalah 23%, hal ini berarti bahwa porsi sklerenkim lebih tinggi pada
bambu dengan pola 3 sehingga kandungan ligninnya pun relatif lebih tinggi.
Kandungan lignin pada bambu Dendrocalamus giganteus (Dg/3) lebih
rendah dibandingkan dengan kandungan lignin pada Dendrocalamus asper (Da/3).
Hasil perhitungan persentase serabut pada kedua bambu pun mendukung kondisi
tersebut. Bambu D. giganteus mempunyai nilai rata-rata 29% serabut, sementara
D. asper memiliki 34% serabut. Perbedaan kandungan serabut akan berkontribusi
pada kandungan lignin.
Kandungan pati
Hasil penelitian nilai rata-rata kandungan kadar pati dapat dilihat pada
Gambar 61. Terdapat interaksi antara pola dan jenis dengan perbandingan kadar
pati antara Dg/3 dengan Da/3 dan antara Da/4 dengan Da/3 yang cukup
mencolok. Di antara berbagai jenis/pola yang diujikan terlihat bahwa bambu D.
asper memiliki kandungan pati yang tertinggi sedangkan bambu A. hundsii/pola 1
dan C. pergracile/pola 2 memiliki kandungan pati yang terendah.
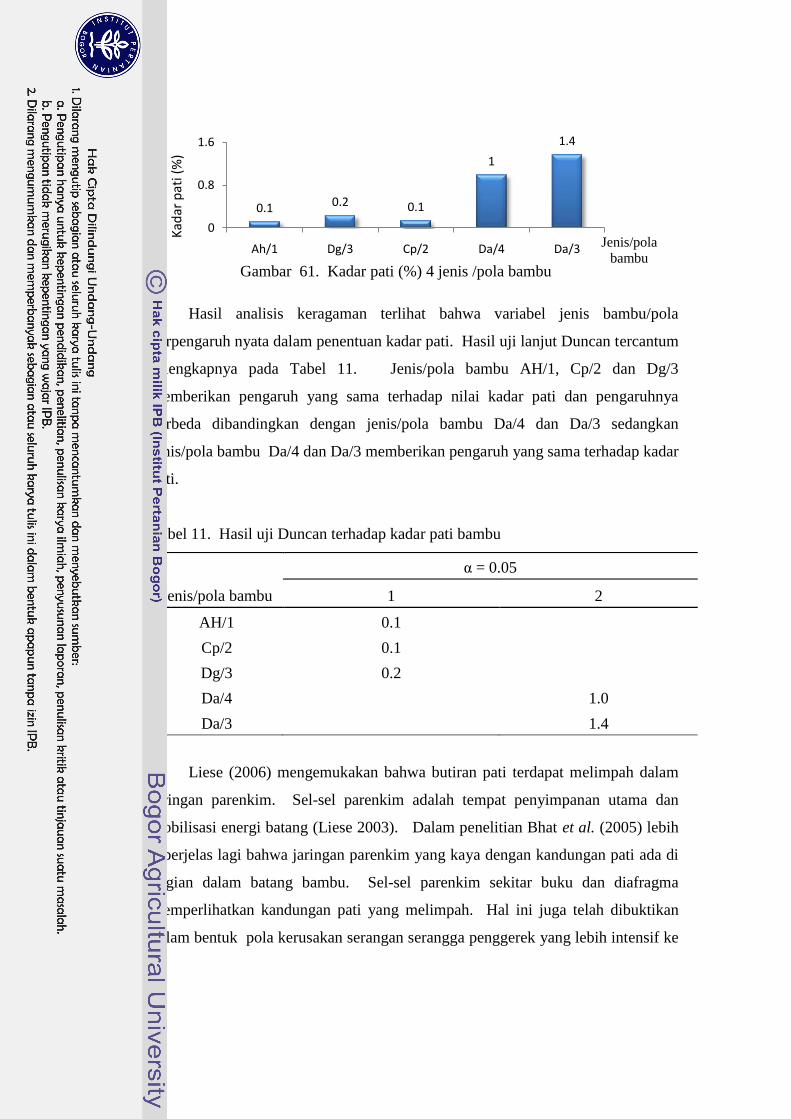
Hasil analisis keragaman terlihat bahwa variabel jenis bambu/pola
berpengaruh nyata dalam penentuan kadar pati. Hasil uji lanjut Duncan tercantum
selengkapnya pada Tabel 11. Jenis/pola bambu AH/1, Cp/2 dan Dg/3
memberikan pengaruh yang sama terhadap nilai kadar pati dan pengaruhnya
berbeda dibandingkan dengan jenis/pola bambu Da/4 dan Da/3 sedangkan
jenis/pola bambu Da/4 dan Da/3 memberikan pengaruh yang sama terhadap kadar
pati.
Tabel 11. Hasil uji Duncan terhadap kadar pati bambu
Jenis/pola bambu
α = 0.05
1 2
AH/1 0.1
Cp/2 0.1
Dg/3 0.2
Da/4 1.0
Da/3 1.4
Liese (2006) mengemukakan bahwa butiran pati terdapat melimpah dalam
jaringan parenkim. Sel-sel parenkim adalah tempat penyimpanan utama dan
mobilisasi energi batang (Liese 2003). Dalam penelitian Bhat et al. (2005) lebih
diperjelas lagi bahwa jaringan parenkim yang kaya dengan kandungan pati ada di
bagian dalam batang bambu. Sel-sel parenkim sekitar buku dan diafragma
memperlihatkan kandungan pati yang melimpah. Hal ini juga telah dibuktikan
dalam bentuk pola kerusakan serangan serangga penggerek yang lebih intensif ke
0.1 0.2 0.1
1
1.4
0
0.8
1.6
Ah/1 Dg/3 Cp/2 Da/4 Da/3 Jenis/pola
bambuGambar 61. Kadar pati (%) 4 jenis /pola bambu
Kad
ar p
ati (
%)

bagian dalam dinding batang yang menandakan adanya kandungan pati yang lebih
banyak (Bhat et al. 2005, Liese 2006).
Analisis hasil penelitian ini terkait penelitian Liese (2006) dan Bhat et al.
(2005) memerlukan informasi tentang kondisi bagian dalam batang yang
dicerminkan dalam bentuk kerapatan ikatan pembuluh jenis-jenis bambu yang
diteliti (Tabel 12). Pola penyebaran kerapatan ikatan pembuluh penampang
lintang batang bambu secara umum adalah kerapatan pada bagian dalam paling
rendah dibandingkan bagian lainnya. Kerapatan ikatan pembuluh yang rendah
berarti mengandung parenkim/jaringan dasar yang tinggi sehingga memungkinkan
mengandung pati yang paling banyak.
Hasil pengujian (Tabel 11) menyatakan bahwa baik AH/1, Dg/3, maupun
Cp/2 memberikan pengaruh yang sama pada kadar pati . Demikian pula Da/4 dan
Da/3 memberikan kontribusi sama pada kadar pati. Bambu AH/1, Dg/3, maupun
Cp/2 memberikan pengaruh pada kandungan pati yang relatif lebih rendah
dibandingkan dengan jenis/pola bambu Da/4 dan Da/3. Apabila hal ini dikaitkan
dengan kondisi kerapatan ikatan pembuluh terutama pada bagian dalam terlihat
bahwa bambu AH/1, Dg/3, maupun Cp/2 memiliki persentase parenkim yang
lebih kecil (kerapatan ikatan pembuluh tinggi) dibandingkan dengan bambu Da/4
dan Da/3 yang berarti bambu AH/1, Dg/3, maupun Cp/2 memiliki kandungan pati
yang lebih sedikit dibandingkan dengan jenis/pola bambu Da/4 dan Da/3.
Pati yang terkandung dalam bambu memainkan peranan penting dalam
keawetan dan masa pakai bambu. Ketahanan bambu terhadap jamur dan serangga
penggerek terkait erat dengan komposisi kimia (Li 2004). Pati yang terdapat pada
bambu merupakan nutrisi untuk jamur dan serangga penggerek. Bambu dengan
nilai kadar pati yang tinggi memiliki peluang kurang resisten terhadap serangan
organisme (Hidalgo 2011, Liese 2003, Sulistyowati 1997). Dalam penelitian
ternyata bambu dengan kadar pati yang tinggi dimiliki oleh bambu D.asper. Hasil
penelitian Sulthoni (1985) memperlihatkan hasill diantaranya bahwa kandungan
pati pada bambu D. asper berfluktuasi diantara 0.3%-3.0%. Dengan demikian
bambu D. asper rentan terhadap serangan organisma penggerek dan posisi
ketahanan terhadap serangan serangga lebih tinggi dibandingkan Bambusa
vulgaris namun lebih rendah dibandingkan Gigantochloa apus dan G. atter.
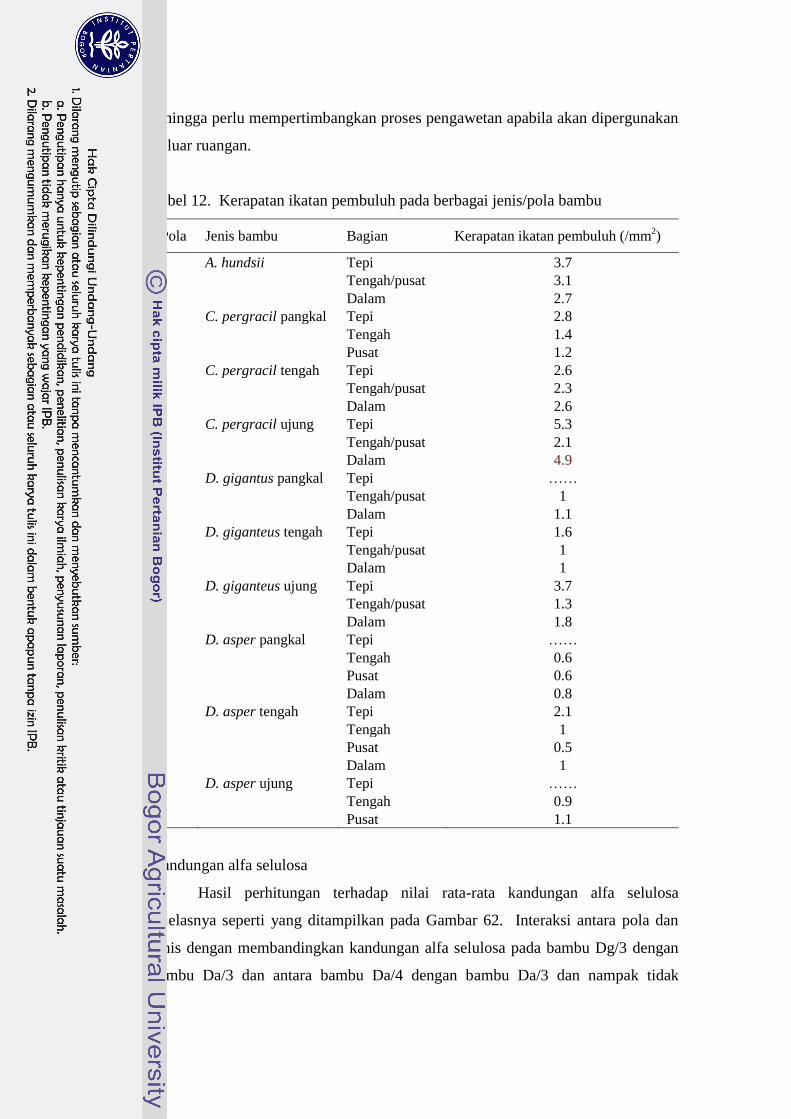
sehingga perlu mempertimbangkan proses pengawetan apabila akan dipergunakan
di luar ruangan.
Tabel 12. Kerapatan ikatan pembuluh pada berbagai jenis/pola bambu
Pola Jenis bambu Bagian Kerapatan ikatan pembuluh (/mm2)
1 A. hundsii Tepi 3.7
Tengah/pusat 3.1
Dalam 2.7
2 C. pergracil pangkal Tepi 2.8
Tengah 1.4
Pusat 1.2
2 C. pergracil tengah Tepi 2.6
Tengah/pusat 2.3
Dalam 2.6
2 C. pergracil ujung Tepi 5.3
Tengah/pusat 2.1
Dalam 4.9
3 D. gigantus pangkal Tepi ……
Tengah/pusat 1
Dalam 1.1
3 D. giganteus tengah Tepi 1.6
Tengah/pusat 1
Dalam 1
3 D. giganteus ujung Tepi 3.7
Tengah/pusat 1.3
Dalam 1.8
4 D. asper pangkal Tepi ……
Tengah 0.6
Pusat 0.6
Dalam 0.8
3 D. asper tengah Tepi 2.1
Tengah 1
Pusat 0.5
Dalam 1
3 D. asper ujung Tepi ……
Tengah 0.9
Pusat 1.1
Kandungan alfa selulosa
Hasil perhitungan terhadap nilai rata-rata kandungan alfa selulosa
sejelasnya seperti yang ditampilkan pada Gambar 62. Interaksi antara pola dan
jenis dengan membandingkan kandungan alfa selulosa pada bambu Dg/3 dengan
bambu Da/3 dan antara bambu Da/4 dengan bambu Da/3 dan nampak tidak
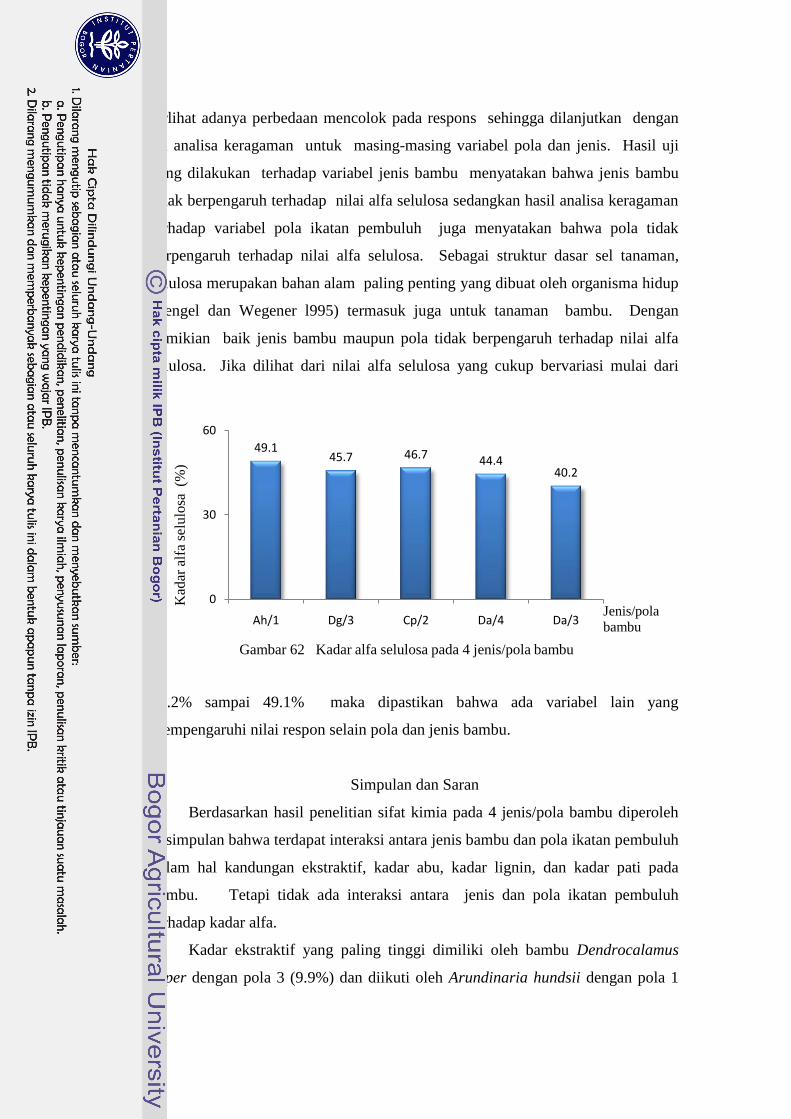
terlihat adanya perbedaan mencolok pada respons sehingga dilanjutkan dengan
uji analisa keragaman untuk masing-masing variabel pola dan jenis. Hasil uji
yang dilakukan terhadap variabel jenis bambu menyatakan bahwa jenis bambu
tidak berpengaruh terhadap nilai alfa selulosa sedangkan hasil analisa keragaman
terhadap variabel pola ikatan pembuluh juga menyatakan bahwa pola tidak
berpengaruh terhadap nilai alfa selulosa. Sebagai struktur dasar sel tanaman,
selulosa merupakan bahan alam paling penting yang dibuat oleh organisma hidup
(Fengel dan Wegener l995) termasuk juga untuk tanaman bambu. Dengan
demikian baik jenis bambu maupun pola tidak berpengaruh terhadap nilai alfa
selulosa. Jika dilihat dari nilai alfa selulosa yang cukup bervariasi mulai dari
40.2% sampai 49.1% maka dipastikan bahwa ada variabel lain yang
mempengaruhi nilai respon selain pola dan jenis bambu.
Simpulan dan Saran
Berdasarkan hasil penelitian sifat kimia pada 4 jenis/pola bambu diperoleh
kesimpulan bahwa terdapat interaksi antara jenis bambu dan pola ikatan pembuluh
dalam hal kandungan ekstraktif, kadar abu, kadar lignin, dan kadar pati pada
bambu. Tetapi tidak ada interaksi antara jenis dan pola ikatan pembuluh
terhadap kadar alfa.
Kadar ekstraktif yang paling tinggi dimiliki oleh bambu Dendrocalamus
asper dengan pola 3 (9.9%) dan diikuti oleh Arundinaria hundsii dengan pola 1
49.145.7 46.7 44.4
40.2
0
30
60
Ah/1 Dg/3 Cp/2 Da/4 Da/3Jenis/pola
bambu
Gambar 62 Kadar alfa selulosa pada 4 jenis/pola bambu
Kad
aral
fa s
elulo
sa (%
)

(9%). Bambu Cephalostachyum pergracile dengan pola 2 memiliki kadar abu
tertinggi (4.3%), sementara bambu Arundinaria hundsii dengan pola 1 memiliki
kandungan lignin dan alfa selulosa tertinggi berturut-turut 32% dan 49.1%.
Bambu Dendrocalamus asper mempunyai kandungan pati tertinggi yang berkisar
dari 1%-1.4%.
Adanya peranan interaksi antara jenis dan pola bambu yang memberikan
kontribusi terhadap beberapa sifat kimia menandakan bahwa jenis dan pola bambu
penting untuk diperhatikan karena terkait dengan beberapa sifat kimia.
Pola ikatan pembuluh memiliki hubungan erat dengan sifat kimia bambu
dan akan berpengaruh terhadap pemanfaatan bambu. Dengan demikian hubungan
sifat kimia dengan pola ikatan pembuluh bambu perlu dikembangkan lebih
mendalam agar bermanfaat dalam membantu menentukan arah penggunaan
bambu.

DAFTAR PUSTAKA
Austin R, Euda K, Levy D. 1974. Bamboo: its Growth and Cultivation. New
York: Weatherhill.
Bhat KV, Varma RV, Paduvil R, Pandalai RC. Santhoshkumar R. 2005.
Distribution of starch in the culms of Bambusa bambos (L.)Voss and its
influence on borer damage. J Americ Bamb Socie 19(1): 1-4
Bodig J, Jayne BA. 1993. Mechanics of Wood and Wood Composites. Florida:
Krieger Publishing Company.
Chen Y, Qin W, LI X, Gong J, Nimanna. 1987. The chemical composition of ten
bamboo species. Di dalam: Rao AN et al, editor. Proceedings of the
International Workshop; Hangzhou. 6–14 October 1985. hlm 110–113.
Espiloy, Z.B. 1983. Variability of specific gravity, silica content and fiber
measurements in kauayan-tinik (B. blumeana). NSTA Technology Journal
8(2): 42-74.
Grosser D, Liese W. 1971. On the anatomy of Asian bamboos, with spesial
reference to their vaskular bundles. Wood Sci and Tech 5: 290-312.
Hidalgo H. 2011. When and how to harvest bamboo?.
http://www.guaduabamboo.com/starch-bamboo.htm[23 Oktober 2011]
Kozlowski TT, Pallardy SG. 1997. Physiology of Woody Plants. United States
of America: Academic Pr.
Li, 1983. Report. Institute of Wood Industry,Chinese Academy of Forestry,
Beijing
Li XB, Shupe TF, Peter GF, Hse CY, Eberhardt TL. 2007. Chemical changes
with maturation of the bamboos spesies Phyllostachys pubescens.
Li XB. 2004. Physical, chemical and mechanical properties of bamboo and its
utilization potential for fibreboard manufacturing [Thesis]. Chinese
Academy of Forestry.
Li XB, Peter GF, Hse CY, Eberhardt TL. 2007. Chemical changes with
maturation of the bamboo spesies Phyllostachys puberscens. J Trop Fort
Sci 19(1): 612 (2007).

Lybeer B, Koch G. 2005. Lignin distribution in the tropical bamboo spesies
Gigantochloa levis. J IAWA 26 (4): 443–456.
Liese W. 1992. The Structure of bamboo in relation to its properties and utilization .
Di dalam: Bamboo and its use. International Symposium On Industrial Use
Of Bamboo. Beijing, China, 7-11 Desember 1992. hlm 1 – 6.
Liese W. 2003. Structures of bamboo culm affecting its utilization. Di dalam
Xuhe C, Yiping L, Ying H, editor. Proceedings of International Workshop
on Bamboo Industrial Utilization. Hubei dan Xianning, Oktober 2003. hlm
6 – 10.
Liese W. 2006. The Anatomy of Bamboo Culms. Http://www.inbar.int/
/txt/tr18/default2.htm. [24 Desember 2006].
Liese W, Kumar S. 2003. Bamboo Preservation Compendium. INBAR Tech.
Rep. No. 22.
Lin J, He X, Hu Y, Kuang T, Ceulemans R. 2002. Lignification and lignin
heterogeneity for various age classes of bamboo (Phyllostachys pubescens)
stems. Physiologia plantarum 112: 296-302.
Lwin KM, Han YY, Maung W, Moe AKZ, Than SBM. 2007. An investigation on
morphology, anatomy and chemical properties of some Myanmar bamboos.
http://www.myanmar.gov.mm/Ag/Jur/ProcFo01.10.[3 Nov. 2007]
Pai-hui H. 1985. A Study on the Mineral Nutrition of Phyllostachys pubescens.
Di dalam : Rao, A.N., Dhanarajan, G. dan Sastry, C.B, editor. Recent
Research on Bamboo. Proceedings of the International Bamboo Workshop,
Hangzholu, People's Republic of China, 6-14 Oktober, 1985.
Qisheng Z, Shenxue J, Yongyu T. 2001. Physical properties of bamboo
material. Industrial Utilization On Bamboo Technical Report No.26
Salisbury FB, Ross CW. 1995. Fisiologi Tumbuhan. Bandung: ITB
Sulistyowati A. 1997. Pengawetan bambu. Wacana No 6.
Sulthoni A. 1985. Traditional preservation of bamboo in Java, Indonesia. Di
dalam : Rao, A.N., Dhanarajan, G. dan Sastry, C.B, editor. Recent
Research on Bamboo. Proceedings of the International Bamboo Workshop,
Hangzholu, People's Republic of China, 6-14 Oktober, 1985.

Wang X, Ren H, Zhang B, Fei B, Burgert I. 2011. Cell wall structure and
formation of maturing fibres of moso bamboo (Phyllostachys pubescens)
increase buckling resistance. J R Soc Interface.
V. PEMBAHASAN DAN KESIMPULAN UMUM
Pembahasan Umum
Pemanfaatan bambu berdasarkan pola
Lebih dari 1000 spesies bambu yang tersebar di dunia memiliki sifat fisik
dan mekanik yang berbeda (Liese 1997). Perbedaan di antara spesies tanaman
bambu tersebut terkait dengan perbedaan dalam sifat anatomi dan kimia (Lwin et
al 2007 ). Dengan demikian studi pada sifat anatomi, fisik dan mekanik harus
dibawa ke arah penjaminan mutu bambu. Karena itu diperlukan informasi yang
menyeluruh dari sifat-sifat yang dimiliki bambu tersebut (Liese 1997). Pola
ikatan pembuluh sebagai variabel sifat anatomi mempunyai karakter yang
mewakili sifat-sifat suatu jenis bambu.
Pola ikatan pembuluh (disingkat pola) berperan dalam sifat mekanik
sebagai faktor yang mempengaruhi kekuatan. Pola ikatan pembuluh juga
mempunyai peran penting karena bersama-sama dengan jenis bambu berpengaruh
pada beberapa sifat kimia bambu yang menjadi dasar dalam pemanfaatan bambu
secara luas.
Bambu pada pola ikatan pembuluh 1
Dalam penelitian sifat anatomi bambu khususnya dalam penentuan
kerapatan ikatan pembuluh ternyata salah satunya dipengaruhi oleh jenis bambu
dan pola. Pola 1 memiliki kerapatan ikatan pembuluh tertinggi dibandingkan
dengan pola-pola yang ada. Kerapatan ikatan pembuluh umumnya terkait dengan
nilai BJ. Hal ini berarti bahwa pola 1 berpeluang mempunyai nilai BJ yang
tinggi. Hasil pengukuran nilai BJ bambu Arundinaria hundsii adalah 0.8
sedangkan nilai BJ bambu Arundinaria javonica adalah 0.6. Bambu dengan nilai
BJ yang relatif tinggi dapat digunakan untuk pemanfaatan struktural dan
termasuk ke dalam kelas kuat II jika dilihat dari sisi BJ saja (Lampiran 12).

Diameter dan tebal batang bambu A. hundsii pada bagian pangkal berturut-turut
adalah 1.6 cm dan 0.6 cm, sedangkan diameter dan tebal batang bambu A.
javonica pada bagian pangkal berturut-turut adalah 2 cm dan 0.6 cm. Ditinjau dari
segi fisik, baik bambu A. hundsii maupun A. javonica memiliki kekurangan jika
digunakan sebagai bahan konstruksi secara langsung. Dengan demikian untuk
memanfaatkan kedua jenis bambu ini memerlukan dukungan teknologi khususnya
dengan memanfaatkan teknologi laminasi bambu (Puslitbang Pemukiman 2009)
sehingga dapat menghasilkan balok dengan kekuatan tinggi. Alternatif lain
pemanfaatan bambu dengan diameter kecil adalah dengan memproses lebih lanjut
sebagai bahan papan partikel bambu (Renjie et al. 2003). Penggunaan bambu
pola 1 secara langsung adalah sebagai asesori elemen bangunan (Surjokusumo
1997) ataupun sebagai tanaman hias. Penggunaan bambu pola 1 sebagai asesori
elemen bangunan didukung juga oleh potensi keawetannya karena termasuk
kelompok bambu dengan kandungan ekstraktif yang tinggi dibandingkan pola
lain. Kandungan pati yang rendah pada pola 1 akan memberikan ketahanan
terhadap serangan serangga penggerek. Ditinjau dari sisi panjang serat, walaupun
panjang serat yang dimiliki pola 1 paling rendah dibandingkan dengan pola-pola
lain namun masih termasuk ke dalam kelompok kelas I apabila digunakan sebagai
bahan baku serat/pulp dan kertas (acuan persyaratan kayu sebagai bahan baku
pulp dan kertas pada Lampiran 13 dan hasil penetapan berdasarkan acuan pada
Lampiran 14). Demikian pula potensi kandungan alfa selulosa yang relatif paling
tinggi (49.1%) dibandingkan pola lain mengindikasikan bahwa pola 1 mempunyai
kandungan serat yang paling tinggi. Namun kandungan ligninnya yang tinggi
sehingga meningkatkan kebutuhan bahan kimia pemasak tinggi pula.
Bambu pada pola ikatan pembuluh 2
Pola 2 yang dimiliki oleh bambu Cephalostachyum pergracile dan
Melocanna baccifera memiliki nilai kerapatan ikatan pembuluh yang berada
setingkat lebih rendah dibandingkan kerapatan ikatan pembuluh pola 1. Sama
halnya seperti pola 2, bambu dengan pola 1 pun berpotensi memiliki nilai BJ yang
tinggi. Bambu yang tergolong pola 2 yaitu bambu Melocanna baccifera dan
Cephalostachyum pergracile masing-masing memiliki nilai BJ 0.7 dan 0.6

(termasuk kelas kuat II apabila dilihat dari sisi BJ-nya). Hasil pengukuran
terhadap diameter batang M. baccifera bagian pangkal adalah 3.4 cm dan tebal
dinding batangnya adalah 0.7 cm. Diameter batang bambu C. pergracile adalah
4.7 cm sedangkan tebal dinding batangnya adalah 1.4 cm. Seperti halnya bambu
A. hundsii dan A. javonica, bambu M. baccifera dan bambu C. pergracile pun
tidak dapat dipergunakan secara langsung sebagai bahan konstruksi, tapi dapat
diaplikasikan dalam bentuk bambu lamina dan bambu lapis. Hal ini sesuai
dengan hasil penelitian Nuriyatin (2000) yang mengumpulkan informasi tentang
jenis-jenis bambu serta kegunaannya, di antaranya menyatakan bahwa bambu C.
pergracile umumnya dimanfaatkan sebagai bahan konstruksi, sedangkan bambu
M. baccifera umumnya digunakan sebagai bahan bangunan, anyaman, kerajinan,
dan bahan baku pulp dan kertas (Dransfield dan Widjaja 1995). Hal yang harus
diperhatikan untuk bambu berpola 2 adalah kandungan ekstraktifnya lebih rendah
dibandingkan pola lain. Kandungan abu yang tinggi juga dimiliki oleh bambu ini.
Diduga kadar silika yang tinggi ada pada bambu C. pergracile sehingga harus
diwaspadai pada saat proses pemotongan karena akan menumpulkan
pisau/gergaji. Namun bambu C. pergracile memiliki kadar pati yang rendah
yang berarti relatif lebih tahan terhadap serangan serangga penggerek.
Penggunaan bambu M. baccifera sebagai bahan kerajinan (Nuriyatin 2000)
menghasilkan produk ramah lingkungan dan sangat diminati masyarakat dunia
(Anonim 2011).
Bambu C.pergracile dan M.baccifera juga dapat digunakan sebagai bahan
baku serat termasuk bahan baku pulp dan kertas. Panjang serabut bambu pada
pola 2 termasuk ke dalam kelas I karena mempunyai panjang serat lebih dari 2000
mikron (Lampiran 14). Nilai alfa selulosanya pun mendukung karena mempunyai
nilai yang relatif tinggi (46.7%) sehingga berpotensi untuk dijadikan bahan baku
pulp. Kandungan lignin yang dimiliki pola 2 berada diantara pola-pola lain
sehingga bahan kimia yang dibutuhkan diduga berada di antara pola-pola
tersebut.
Bambu pada pola ikatan pembuluh 3

Nilai kerapatan ikatan pembuluh untuk pola 3 berada pada posisi paling
rendah dibandingkan pola-pola lain. Bambu-bambu yang memiliki pola 3 adalah
Dendrocalamus strictus, D. giganteus, D. asper bagian tengah dan ujung, serta
Gigantochloa apus bagian ujung. Ditinjau dari BJ, nilai BJ keseluruhan bambu
yang diteliti adalah 0.6 namun khusus bambu-bambu untuk keperluan konstruksi
struktural, nilai BJ-nya adalah 0.8. D. asper, dan Gigantochloa apus secara
tampilan fisik sesuai untuk penggunaan struktural. Berdasarkan hasil pengujian,
salah satu variabel yang dilibatkan dalam standar kekuatan kayu adalah MOR
yang dipengaruhi pola ikatan pembuluh. Posisi pola 3 dalam perannya sebagai
bahan dasar konstruksi termasuk ke dalam kelas kuat II (Lampiran 12).
Bambu D. giganteus juga dapat dimanfaatkan sebagai elemen non
struktural, bahan baku pulp dan kertas, kerajinan dan lain-lain. Bambu G. apus
bagian ujung juga dapat digunakan sebagai bahan angklung. Dalam penelitian
Nuriyatin (2000) dikemukakan bahwa diantara 5 jenis bambu yang diujikan
ternyata bambu G. atroviolacea memiliki kualitas suara terbaik. Diduga hal ini
terjadi selain karena nilai persentase serabut yang berdinding tebal cukup tinggi
(BJ-nya adalah 0.7) juga kondisi serabut tersebar secara merata pada penampang
lintang batang bambu. Bambu G. apus berada pada posisi ke-2 sebagai bahan
angklung setelah bambu G. atroviolacea.
Seperti halnya pola-pola lain, bambu-bambu yang tergolong ke dalam pola 3-
pun dapat dimanfaatkan sebagai bahan baku serat/pulp dan kertas (Lampiran 14).
Hal ini didukung ukuran serabut yang panjang sehingga termasuk ke dalam
kualitas I jika digunakan sebagai bahan baku pulp dan kertas. Pola 3 yang berasal
dari bambu D. asper memiliki sifat kimia yang khas karena memiliki kandungan
ekstraktif serta kadar pati yang paling tinggi dibanding pola lain. Keadaan yang
berbeda dimiliki oleh bambu D. giganteus yang mempunyai kandungan ekstraktif
dan pati yang rendah.
Bambu pada pola ikatan pembuluh 4
Bambu-bambu yang memiliki pola 4 adalah adalah D. asper bagian pangkal
dan G. apus bagian pangkal dan tengah. Bambu-bambu tersebut memiliki
diameter batang yang besar serta dinding batang yang tebal sehingga sesuai jika

digunakan sebagai bahan baku struktural. Ditinjau dari sisi BJ-pun memiliki nilai
yang cukup tinggi pula yaitu 0.6. Dengan nilai BJ yang tinggi diharapkan dimensi
lebih stabil berdasarkan sifat mekanis serta mengacu kepada standar kayu sebagai
bahan baku konstruksi maka bambu-bambu yang memiliki pola 4 dapat
dikelompokan ke dalam kelas kuat III (Lampiran 14).
Hasil penelitian Nuriyatin (2000) menyatakan bahwa bambu D. asper
mempunyai kegunaan sebagai bahan konstruksi, kerajinan, mebel, dan lain-lain.
Sebagai bahan mebel, bambu D. asper memenuhi beberapa persyaratan termasuk
karena kekokohannya (Ridwanti 2000). Hal ini sesuai apabila dikaitkan dengan
persyaratan fisik mekanik kayu untuk mebel yang menuntut kekuatan dan
keawetan bahan baku minimum kelas III (SNI 1989). Bambu G. apus selain
dimanfaatkan sebagai bahan konstruksi juga digunakan sebagai bahan baku pulp
dan kertas. Hasil penelitian Bachtiar (2008) pada bambu G. apus menyatakan
bahwa bambu ini dapat digunakan sebagai komponen konstruksi rangka batang
ruang.
Bambu-bambu yang tergolong ke dalam pola 4 juga memiliki peluang yang
baik sebagai bahah pulp dan kertas (Lampiran 14). Hal ini didukung dengan nilai
serabut yang terpanjang dibandingkan pola-pola lain sehingga nilai daya tenun
pun sangat baik (kelas I). Tapi di antara pola yang ada, kadar alfa selulosa pada
pola 4 paling rendah sehingga dapat berpengaruh pada kekuatan serat. Bambu
yang termasuk ke dalam pola 4 juga mempunyai kandungan ekstraktif yang tidak
terlalu tinggi, kadar abu paling rendah, dan kadar pati yang tinggi sehingga harus
diperhitungkan apabila persyaratan penggunannya memerlukan sifat keawetan
tinggi.
Pola dalam Pemanfaatan Bambu
Pengamatan terhadap keseluruhan hasil penelitian memberikan gambaran
pemanfaatan bambu dalam penggunaan tertentu dengan memanfaatkan pola yang
dimiliki oleh setiap jenis bambu. Dalam penggunaan bambu sebagai bahan
konstruksi, pola menunjukkan hubungan yang cukup erat terutama terhadap nilai
MOR . Penelitian ini merupakan langkah awal dalam memanfaatkan bambu
secara optimum. Langkah ke depan diharapkan adanya pengembangan standar

penggunaan bambu sebagai bahan dasar konstruksi dengan melibatkan pola ikatan
pembuluh.
Penggunaan bambu sebagai bahan berserat (pulp dan kertas) telah lama
dilakukan industri di berbagai negara. Penelitian terhadap bambu sebagai bahan
serat dilakukan banyak peneliti yang menunjukkan prospek bambu. Hasil
penelitian yang diperoleh dengan menguji melalui morfologi serat dan turunannya
adalah bahwa bambu dengan pola ikatan pembuluh 1 sampai 4 memberikan hasil
yang sama, yaitu setiap pola menghasilkan daya tenun kelas I sementara untuk
nilai lainnya termasuk kelas III. Pengamatan lebih mendalam terhadap karakter
pola ikatan pembuluh terkait dengan perannya sebagai bahan baku pulp dan kertas
terlihat bahwa bambu pada pola 1 mempunyai ciri tertentu yaitu memiliki
fleksibilitas yang paling tinggi dibandingkan dengan pola-pola lain sementara ciri
pada pola ikatan pembuluh 4 adalah mempunyai panjang serabut dan daya tenun
yang paling tinggi.
Hasil penelaahan terhadap penggunaan bambu sebagai bahan kerajinan
terlihat bahwa seluruh bambu yang tergabung dalam pola 4 dapat digunakan
sebagai bahan kerajinan, demikian pula sebagian bambu yang tergabung dalam
pola 2 (Cephalostachyum pergracile). Secara fisik, selama ini bambu-bambu
yang dipergunakan sebagai bahan kerajinan berukuran besar baik diameter
maupun ketebalan dinding batangnya.
Dalam aplikasi penggunaan, ciri-ciri yang dimiliki setiap pola akan
memberikan kontribusi ketahanan terhadap serangan penggerek, tingkat
kandungan serat dan kelancaran dalam proses pemesinan. Khusus bambu yang
tergolong pola 1 selain memiliki kandungan serat tertinggi, ketahanan terhadap
serangan penggerek juga tinggi. Sementara pada pola 2, diindikasikan bambu
memiliki ketahanan terhadap serangan penggerek tinggi, juga diduga
mengandung silika tinggi yang berpengaruh pada proses pemesinan. Bambu pada
pola 3 mempunyai kandungan serat sedang dengan ketahanan terhadap serangan
rayap beragam. Salah satu ciri yang dimiliki oleh bambu yang tergolong pola 4
adalah selain mempunyai kandungan serat paling rendah juga ketahanan terhadap
penggereknya pun paling rendah.

Gambaran umum yang dapat dibaca dari keberadaan pola ikatan pembuluh
bambu terkait dengan penggunaannya dapat dilihat pada Gambar 63.
Gambar 63. Pemanfaatan bambu berdasarkan pola
Simpulan umum
Pola ikatan pembuluh bambu (pola) adalah variabel sifat anatomi selain
dapat digunakan sebagai kunci identifikasi juga menunjukkan karakter yang
mewakili sifat-sifat suatu jenis bambu. Pola tersebut memiliki fungsi dan
keterkaitan dengan sifat-sifat dasar yang berguna dalam arah pemanfaatan bambu.
Pola ikatan pembuluh muncul pada berbagai jenis bambu baik dalam bentuk
pola tunggal atau bentuk pola kombinasi antara dua pola. Pola pada bagian
pangkal batang terdiri dari pola tunggal sedangkan pola kombinasi terbentuk pada
bagian tengah dan ujung batang bambu. Perbedaan pola berkaitan dengan
kerapatan ikatan pembuluh, panjang serabut dan persentase kandungan serabut.
Pengujian sifat fisik dan mekanik bambu menunjukkan bahwa pola
berpengaruh pada nilai MOR kecuali pada BJ, MOE, keteguhan tekan sejajar
Pola 1
•Konstruksi
•Laminasi
•Papan partikel
•Serat/pulp dan kertas
Pola 2
•Konstruksi
•Kerajinan
•Laminasi
•Papan partikel
•Serat/pulp dan kertas
Pola 3
•Konstruksi
•Meubel
•Kerajianan
•Alat musik
•Laminasi
•Papan partikel
•Serat/pulp dan kertas
Pola 4
•Konstruksi
•Meubel
•Kerajinan
•Laminasi
•Papan partikel
•Serat/pulp dan kertas

serat, dan keteguhan tarik sejajar serat. Meskipun demikian pola tersebut cukup
kuat dalam menduga kekuatan bambu sehingga dapat dipakai dalam arah
pemanfaatan untuk bahan konstruksi, bahan bangunan, mebel dan sebagainya.
Demikian pula pola dapat mengarahkan pemanfaatan untuk keperluan non
struktural seperti kerajinan, asesori bahan bangunan, anyaman dan lain-lain.
Pengujian terhadap sifat kimia terbukti adanya hubungan erat antara jenis
bambu dan pola dengan kadar ekstraktif, kadar abu, kadar pati, dan lignin kecuali
dengan selulosa. Hal ini dapat menjadi dasar hubungan erat antara jenis dan pola
dengan sifat kimia yang menjadi dasar pemanfaatan bambu seperti untuk bahan
baku pulp dan kertas, dan papan serat.
Berdasarkan kajian hubungan antara pola dengan sifat-sifat dasar dan
kaitannya dengan parameter pemanfaatan bambu, dapat disimpulkan bahwa pola
tersebut dapat dipakai sebagai penduga kemungkinan pemanfaatan bambu dalam
praktek. Selain itu penerapan di lapangan lebih mudah dan praktis. Untuk itu
perlu dilengkapi dengan pembuatan matriks pemanfaatan dan parameter yang
dipersyaratkan untuk tujuan pemakaian tertentu dengan memasukan
penggolongan jenis bambu berdasar pola ikatan pembuluh.
Mengingat banyaknya jenis bambu, maka perlu dilakukan inventarisasi
bambu-bambu yang tumbuh di Indonesia sekaligus menetapkan pola ikatan
pembuluhnya. Selanjutnya untuk mempertegas peranan ikatan pembuluh dalam
pendugaan penggunaannya, sebaiknya penelitian dikembangkan pula untuk
bambu-bambu lainnya. Tahap selahjutnya diharapkan aplikasi pemanfaatan pola
dapat dijadikan acuan dalam penentuan pemanfaatan bambu dengan dilibatkan
secara langsung dalam klasifikasi kelas penggunaan bambu.

DAFTAR PUSTAKA
Anonim. 2011. Usaha Kerajinan Bambu Indonesia Diminati Pasar Dunia. Http://
bisnisukm.com/usaha-kerajinan-bambu-indonesia-diminati-pasar-
dunia.html.[17 Desember 2011]
Bactiar G. 2008. Pemanfaatan buluh bambu tali sebagai komponen pada
konstruksi rangka batang ruang [disertasi]. Bogor. Program Pascasarjana,
Institut Pertanian Bogor.
Dransfield S, Widjaja EA, editor. 1995. Bamboos. No 7. Indonesia: Prosea
Foundation Bogor.
Lwin KM, Han YY, Maung W, Moe AKZ, Than SBM. 2007. An investigation on
morphology, anatomy and chemical properties of some Myanmar bamboos. Nuriyatin N. 2000. Studi analisa sifat-sifat dasar bambu pada beberapa tujuan
penggunaan [tesis]. Bogor: Program Pascasarjana, Institut Pertanian Bogor.
Puslitbang Pemukiman. 2009, Pengembangan Bambu Komposit sebagai Bahan
Bangunan Alternatif Pengganti Kayu
Renjie, Z, Zhe C, Jianhui Z. 2003. Technological innovative course and prospect
of bamboo-based panel of China. Proceedings of International Workshop
on Bamboo Industrial Utilization.
Ridwanti. 2002. Pemanfaatan bambu di Indonesia. Fakultas Pertanian,
Universitas Sumatra Utara.
[SNI] Standar Nasional Indonesia. 1989. Persyaratan kayu sebagai bahan mebel.
Surjokusumo, HMS. 1997. Pemanfaatan bambu untuk bangunan. Dalam panel
diskusi bambu, 4 Desember 1997. Jakarta.

