elektrofisiologi jantung
DESCRIPTION
Tentang AritmiaTRANSCRIPT
ELEKTROFISIOLOGI JANTUNG DAN MEKANISME ARITMIA
PENDAHULUAN
Elektrofisologi adalah ilmu yang mempelajari mengenai mekanisme terbentuknya fenomena elektris
dan konsekuensinya dalam kehidupan suatu organisme (Dorland, 2007). Elektrofisiologi jantung
mempelajari tentang mekanisme, fungsi dan keberadaan aktivitas elektris di jantung termasuk
inisiasi impuls dan konduksi dari level seluler. Gangguan jantung dengan dasar gangguan proses
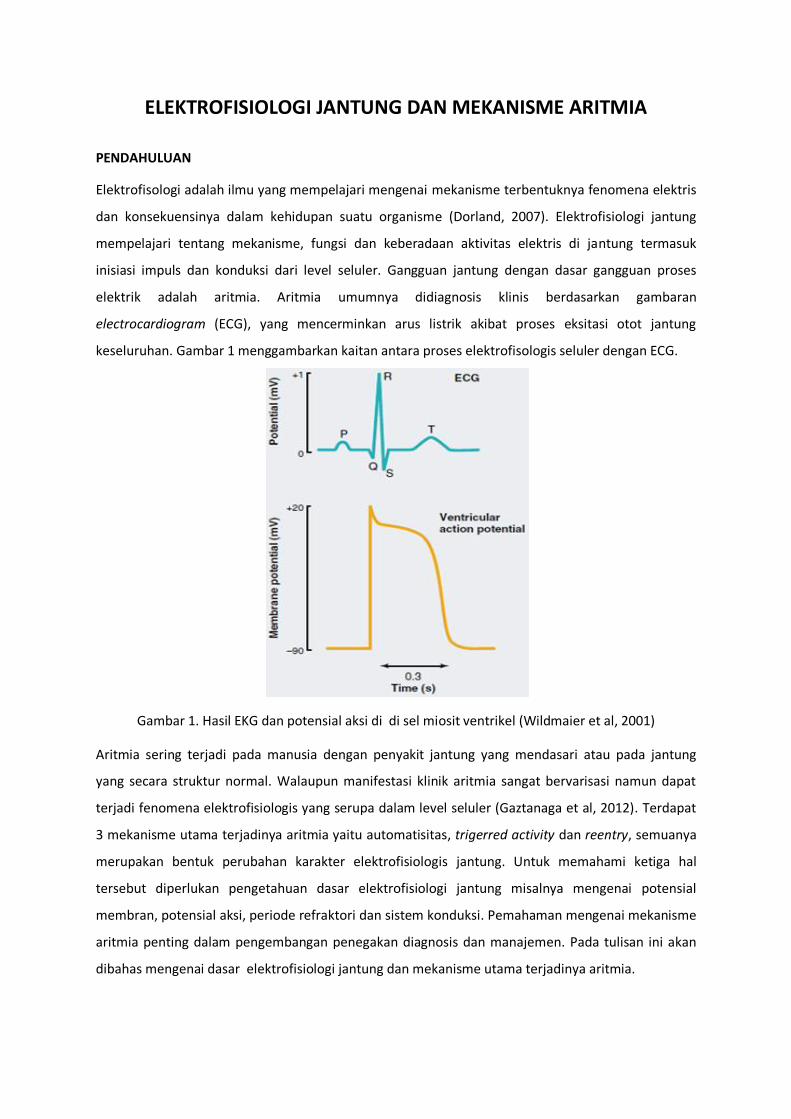
elektrik adalah aritmia. Aritmia umumnya didiagnosis klinis berdasarkan gambaran
electrocardiogram (ECG), yang mencerminkan arus listrik akibat proses eksitasi otot jantung
keseluruhan. Gambar 1 menggambarkan kaitan antara proses elektrofisologis seluler dengan ECG.
Gambar 1. Hasil EKG dan potensial aksi di di sel miosit ventrikel (Wildmaier et al, 2001)
Aritmia sering terjadi pada manusia dengan penyakit jantung yang mendasari atau pada jantung
yang secara struktur normal. Walaupun manifestasi klinik aritmia sangat bervarisasi namun dapat
terjadi fenomena elektrofisiologis yang serupa dalam level seluler (Gaztanaga et al, 2012). Terdapat
3 mekanisme utama terjadinya aritmia yaitu automatisitas, trigerred activity dan reentry, semuanya
merupakan bentuk perubahan karakter elektrofisiologis jantung. Untuk memahami ketiga hal
tersebut diperlukan pengetahuan dasar elektrofisiologi jantung misalnya mengenai potensial
membran, potensial aksi, periode refraktori dan sistem konduksi. Pemahaman mengenai mekanisme
aritmia penting dalam pengembangan penegakan diagnosis dan manajemen. Pada tulisan ini akan
dibahas mengenai dasar elektrofisiologi jantung dan mekanisme utama terjadinya aritmia.

POTENSIAL MEMBRAN
Setiap membran sel mempunyai potensial membran yang terjadi karena perbedaan konsentrasi ion
di sitoplasma dengan interstitial dan berubah bila ada aliran ion melintasi membran (Mohrman and
Haller, 2006). Potensial membran istirahat didefinisikan sebagai perbedaan potensial listrik (voltase)
pada membran sel eksitabel selama kondisi istirahat. Potensial membran istirahat sel tubuh berada
dalam rentang +5 sampai -100 mV. Pada sebagian besar sel, potensial membran istirahat berada
dalam kondisi terpolarisasi dengan nilai negatif, yang berarti bagian intrasel lebih negatif dari bagian
ekstrasel. Sel pacemaker jantung mempunyai potensial membran istirahat -60 mV sedangkan sel
otot jantung -90 mV (Tortora dan Dericsson, 2010).
PERANAN KANAL ION
Secara umum ion bersifat tidak larut lipid sehingga tidak mampu menembus membran lipid bilayer
dengan bebas. Ion keluar atau masuk sel melalui struktur protein transmembran yang dapat berupa
1) kanal ion, 2) ion exchanger, 3) pompa ion. Dari ketiganya (gambar 2), yang bertanggungjawab
terhadap potensial membran istirahat dan perubahan cepat pada saat timbulnya potensial aksi
adalah kanal ion.
Gambar 2. Protein transmembran untuk aliran ion (Jalife et al, 2009)
Otot jantung mempunyai 3 tipe kanal ion yaitu kanal ion gerbang voltase, kanal ion gerbang ligand
dan kanal ion gerbang mekanik. Pada kondisi normal kanal ion gerbang voltase yang memengang
peran utama dalam penjalaran potensial aksi. Kanal jenis ini mempunyai domain protein bermuatan
sebagai sensor voltase (gambar 3) yang sensitive terhadap perubahan potensial membran (Jalife et
al, 2009)
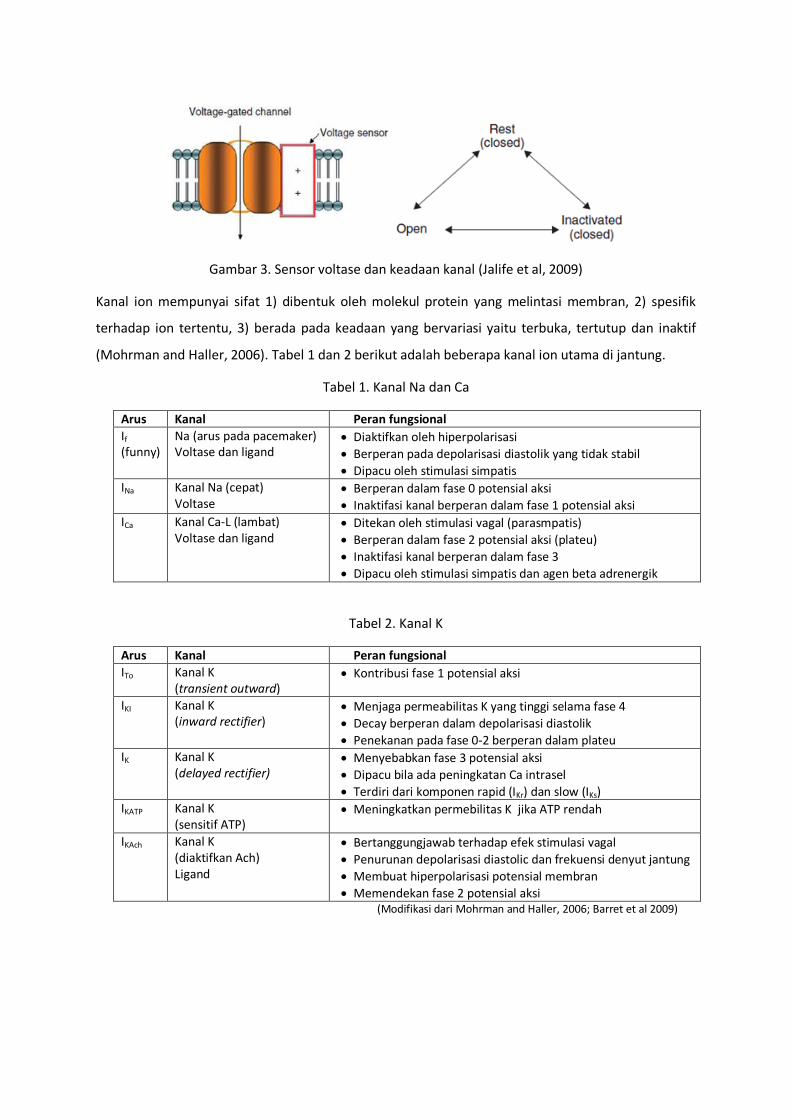
Gambar 3. Sensor voltase dan keadaan kanal (Jalife et al, 2009)
Kanal ion mempunyai sifat 1) dibentuk oleh molekul protein yang melintasi membran, 2) spesifik
terhadap ion tertentu, 3) berada pada keadaan yang bervariasi yaitu terbuka, tertutup dan inaktif
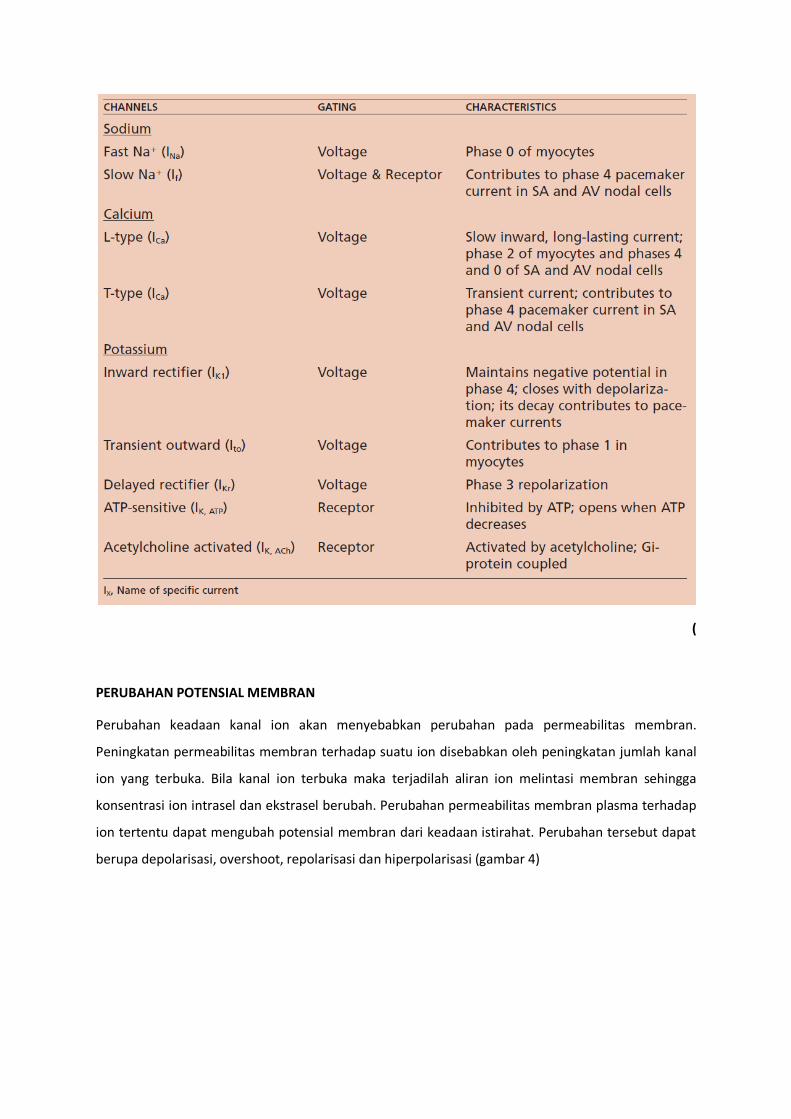
(Mohrman and Haller, 2006). Tabel 1 dan 2 berikut adalah beberapa kanal ion utama di jantung.
Tabel 1. Kanal Na dan Ca
Arus Kanal Peran fungsional
If
(funny) Na (arus pada pacemaker) Voltase dan ligand
Diaktifkan oleh hiperpolarisasi
Berperan pada depolarisasi diastolik yang tidak stabil
Dipacu oleh stimulasi simpatis INa Kanal Na (cepat)
Voltase Berperan dalam fase 0 potensial aksi
Inaktifasi kanal berperan dalam fase 1 potensial aksi ICa Kanal Ca-L (lambat)
Voltase dan ligand Ditekan oleh stimulasi vagal (parasmpatis)
Berperan dalam fase 2 potensial aksi (plateu)
Inaktifasi kanal berperan dalam fase 3
Dipacu oleh stimulasi simpatis dan agen beta adrenergik
Tabel 2. Kanal K
Arus Kanal Peran fungsional
ITo Kanal K (transient outward)
Kontribusi fase 1 potensial aksi
IKI Kanal K (inward rectifier)
Menjaga permeabilitas K yang tinggi selama fase 4
Decay berperan dalam depolarisasi diastolik
Penekanan pada fase 0-2 berperan dalam plateu IK Kanal K
(delayed rectifier) Menyebabkan fase 3 potensial aksi
Dipacu bila ada peningkatan Ca intrasel
Terdiri dari komponen rapid (IKr) dan slow (IKs) IKATP Kanal K
(sensitif ATP) Meningkatkan permebilitas K jika ATP rendah
IKAch Kanal K (diaktifkan Ach) Ligand
Bertanggungjawab terhadap efek stimulasi vagal
Penurunan depolarisasi diastolic dan frekuensi denyut jantung
Membuat hiperpolarisasi potensial membran
Memendekan fase 2 potensial aksi (Modifikasi dari Mohrman and Haller, 2006; Barret et al 2009)
(
PERUBAHAN POTENSIAL MEMBRAN
Perubahan keadaan kanal ion akan menyebabkan perubahan pada permeabilitas membran.
Peningkatan permeabilitas membran terhadap suatu ion disebabkan oleh peningkatan jumlah kanal
ion yang terbuka. Bila kanal ion terbuka maka terjadilah aliran ion melintasi membran sehingga
konsentrasi ion intrasel dan ekstrasel berubah. Perubahan permeabilitas membran plasma terhadap
ion tertentu dapat mengubah potensial membran dari keadaan istirahat. Perubahan tersebut dapat
berupa depolarisasi, overshoot, repolarisasi dan hiperpolarisasi (gambar 4)

Gambar 4. Perubahan Potensial Membran (Wildmaier et al, 2001)
Tiga ion utama yang berperan dalam menentukan potensial membran jantung adalah ion natrium
(Na), Kalium (K) dan Kalsium (Ca). Konsentrasi ion Na dan Ca lebih tinggi di ekstrasel sedangkan ion K
lebih tinggi di intrasel (Mohrman and Haller, 2006). Pembukaan kanal ion menyebabkan suatu
inward current atau outward current (gambar 5). Peningkatan permebilitas membran terhadap ion
Na dan Ca menyebabkan aliran ion bermuatan positif ke dalam sel (inward current) sehingga
polaritas membran berkurang (arus defleksi negatif, voltase depolarisasi). Sebaliknya peningkatan
permeabilitas membran terhadap ion K menyebabkan aliran ion positif keluar sel (outward current)
sehingga polaritas membran bertambah (arus defleksi positif, voltase repolarisasi). Penutupan atau
inaktivasi kanal ion menghentikan efek ini. Proses yang melibatkan aliran ketiga ion utama ini
mendasari terbentuknya potensial aksi di jantung.
Gambar 5. Inward dan outward current (Jalife et al, 2009)
Setelah terjadi proses potensial aksi maka komposisi ionik ekstra dan intrasel jantung menjadi
berubah. Kadar ion Na dan Ca intrasel menjadi bertambah sedangkan kadar K menjadi berkurang.
Untuk mencegah overload Ca intrasel dan mengembalikan komposisi ionik seperti semula maka
terdapat 3 mekanisme pompa ion (gambar 6). Tiga pompa ion itu adalah 1) pompa aktif Ca keluar sel
yang membutuhkan ATP, 2) mekanisme pompa NCX atau Na-Ca Exchanger dengan perbandingan Na
masuk dan Ca keluar 3:1, 3) transport aktif pompa ion Na/K ATPase mengeluarkan Na dan
memasukan K dengan perbandingan 3:2,

Gambar 6. Mekanisme mencegah overload Ca
Potensial aksi jantung di beberapa bagian jantung mempunyai karakter yang berbeda karena
perbedaan properti elektrofisiologisnya. Secara umum mekanisme timbulnya potensial aksi
dibedakan menjadi dua yaitu potensial aksi yang terjadi di pacemaker dan di otot kontraktil jantung.
Potensial aksi yang timbul di pacemaker merupakan “slow response action potential” sedangkan di
otot jantung merupakan “fast response action potential” (Tabel 3)
Tabel 3. Perbandingan Slow dan Fast Response Action Potential
Slow Response Action Potential Fast Response Action Potential
potensial aksi pacemaker
Inisiasi depolarisasi lebih lambat
amplitude potensial aksi yang lebih rendah
fase plateu yang pendek dan tidak stabil
repolarisasi yang lambat menuju potensial membran istirahat yang tidak stabil
potensial aksi sel otot jantung
depolarisasi cepat, overshoot tinggi (fase 0)
repolarisasi cepat setelah overshoot (fase 1)
adanya plateu yang lama (fase 2)
repolarisasi (fase 3) ke potensial membran istirahat yang stabil dan sangat negatif (fase 4).
(Modifikasi dari Mohrman and Haller, 2006)
MEKANISME POTENSIAL AKSI SEL PACEMAKER
Pacemaker jantung terdiri dari nodus sinoatrial (SA) dan nodus atrioventricular (AV) yang dapat
membentuk potensial aksi secara spontan (gambar 7). Pada kondisi normal nodus SA adalah
pacemaker utama jantung, memiliki potensial membran “istirahat” -60 mV yang tidak stabil yang
senantiasa terdepolarisasi dengan threshold potensial aksi -40mV. Kondisi ketidaksatabilan potensial
membran istirahat di pacemaker diistilahkan sebagai “depolarisasi fase 4”, “diastolik depolarisasi”
atau “potensial pacemaker” (Mohrman and Haller, 2006).
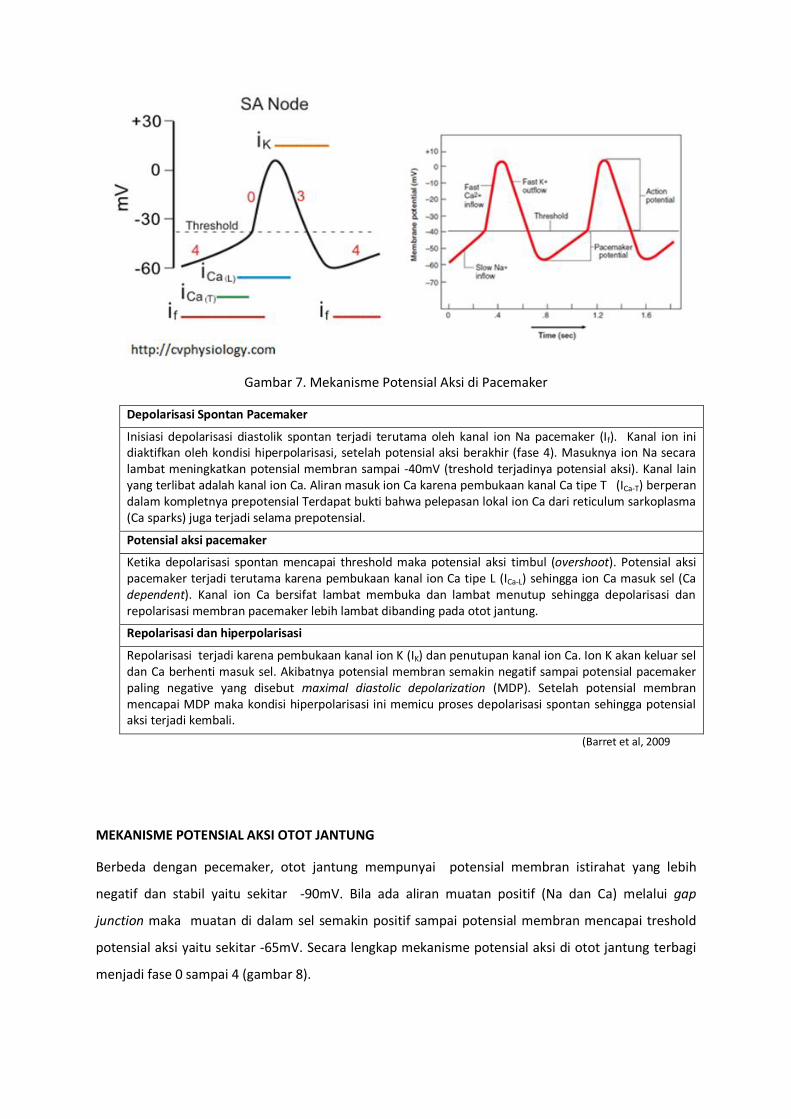
Gambar 7. Mekanisme Potensial Aksi di Pacemaker
Depolarisasi Spontan Pacemaker
Inisiasi depolarisasi diastolik spontan terjadi terutama oleh kanal ion Na pacemaker (If). Kanal ion ini diaktifkan oleh kondisi hiperpolarisasi, setelah potensial aksi berakhir (fase 4). Masuknya ion Na secara lambat meningkatkan potensial membran sampai -40mV (treshold terjadinya potensial aksi). Kanal lain yang terlibat adalah kanal ion Ca. Aliran masuk ion Ca karena pembukaan kanal Ca tipe T (ICa-T) berperan dalam kompletnya prepotensial Terdapat bukti bahwa pelepasan lokal ion Ca dari reticulum sarkoplasma (Ca sparks) juga terjadi selama prepotensial.
Potensial aksi pacemaker
Ketika depolarisasi spontan mencapai threshold maka potensial aksi timbul (overshoot). Potensial aksi pacemaker terjadi terutama karena pembukaan kanal ion Ca tipe L (ICa-L) sehingga ion Ca masuk sel (Ca dependent). Kanal ion Ca bersifat lambat membuka dan lambat menutup sehingga depolarisasi dan repolarisasi membran pacemaker lebih lambat dibanding pada otot jantung.
Repolarisasi dan hiperpolarisasi
Repolarisasi terjadi karena pembukaan kanal ion K (IK) dan penutupan kanal ion Ca. Ion K akan keluar sel dan Ca berhenti masuk sel. Akibatnya potensial membran semakin negatif sampai potensial pacemaker paling negative yang disebut maximal diastolic depolarization (MDP). Setelah potensial membran mencapai MDP maka kondisi hiperpolarisasi ini memicu proses depolarisasi spontan sehingga potensial aksi terjadi kembali.
(Barret et al, 2009
MEKANISME POTENSIAL AKSI OTOT JANTUNG
Berbeda dengan pecemaker, otot jantung mempunyai potensial membran istirahat yang lebih
negatif dan stabil yaitu sekitar -90mV. Bila ada aliran muatan positif (Na dan Ca) melalui gap
junction maka muatan di dalam sel semakin positif sampai potensial membran mencapai treshold
potensial aksi yaitu sekitar -65mV. Secara lengkap mekanisme potensial aksi di otot jantung terbagi
menjadi fase 0 sampai 4 (gambar 8).
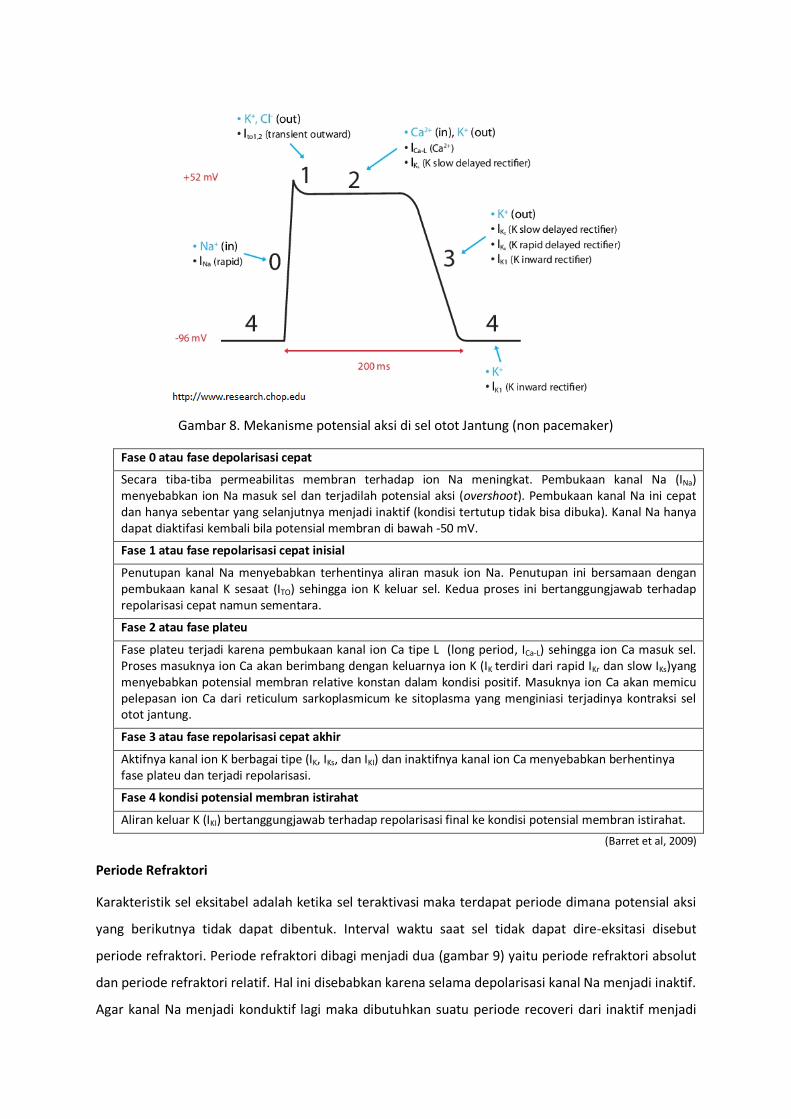
Gambar 8. Mekanisme potensial aksi di sel otot Jantung (non pacemaker)
Fase 0 atau fase depolarisasi cepat
Secara tiba-tiba permeabilitas membran terhadap ion Na meningkat. Pembukaan kanal Na (INa) menyebabkan ion Na masuk sel dan terjadilah potensial aksi (overshoot). Pembukaan kanal Na ini cepat dan hanya sebentar yang selanjutnya menjadi inaktif (kondisi tertutup tidak bisa dibuka). Kanal Na hanya dapat diaktifasi kembali bila potensial membran di bawah -50 mV.
Fase 1 atau fase repolarisasi cepat inisial
Penutupan kanal Na menyebabkan terhentinya aliran masuk ion Na. Penutupan ini bersamaan dengan pembukaan kanal K sesaat (ITO) sehingga ion K keluar sel. Kedua proses ini bertanggungjawab terhadap repolarisasi cepat namun sementara.
Fase 2 atau fase plateu
Fase plateu terjadi karena pembukaan kanal ion Ca tipe L (long period, ICa-L) sehingga ion Ca masuk sel. Proses masuknya ion Ca akan berimbang dengan keluarnya ion K (IK terdiri dari rapid IKr dan slow IKs)yang menyebabkan potensial membran relative konstan dalam kondisi positif. Masuknya ion Ca akan memicu pelepasan ion Ca dari reticulum sarkoplasmicum ke sitoplasma yang menginiasi terjadinya kontraksi sel otot jantung.
Fase 3 atau fase repolarisasi cepat akhir
Aktifnya kanal ion K berbagai tipe (IK, IKs, dan IKI) dan inaktifnya kanal ion Ca menyebabkan berhentinya fase plateu dan terjadi repolarisasi.
Fase 4 kondisi potensial membran istirahat
Aliran keluar K (IKI) bertanggungjawab terhadap repolarisasi final ke kondisi potensial membran istirahat.
(Barret et al, 2009)
Periode Refraktori
Karakteristik sel eksitabel adalah ketika sel teraktivasi maka terdapat periode dimana potensial aksi
yang berikutnya tidak dapat dibentuk. Interval waktu saat sel tidak dapat dire-eksitasi disebut
periode refraktori. Periode refraktori dibagi menjadi dua (gambar 9) yaitu periode refraktori absolut
dan periode refraktori relatif. Hal ini disebabkan karena selama depolarisasi kanal Na menjadi inaktif.
Agar kanal Na menjadi konduktif lagi maka dibutuhkan suatu periode recoveri dari inaktif menjadi
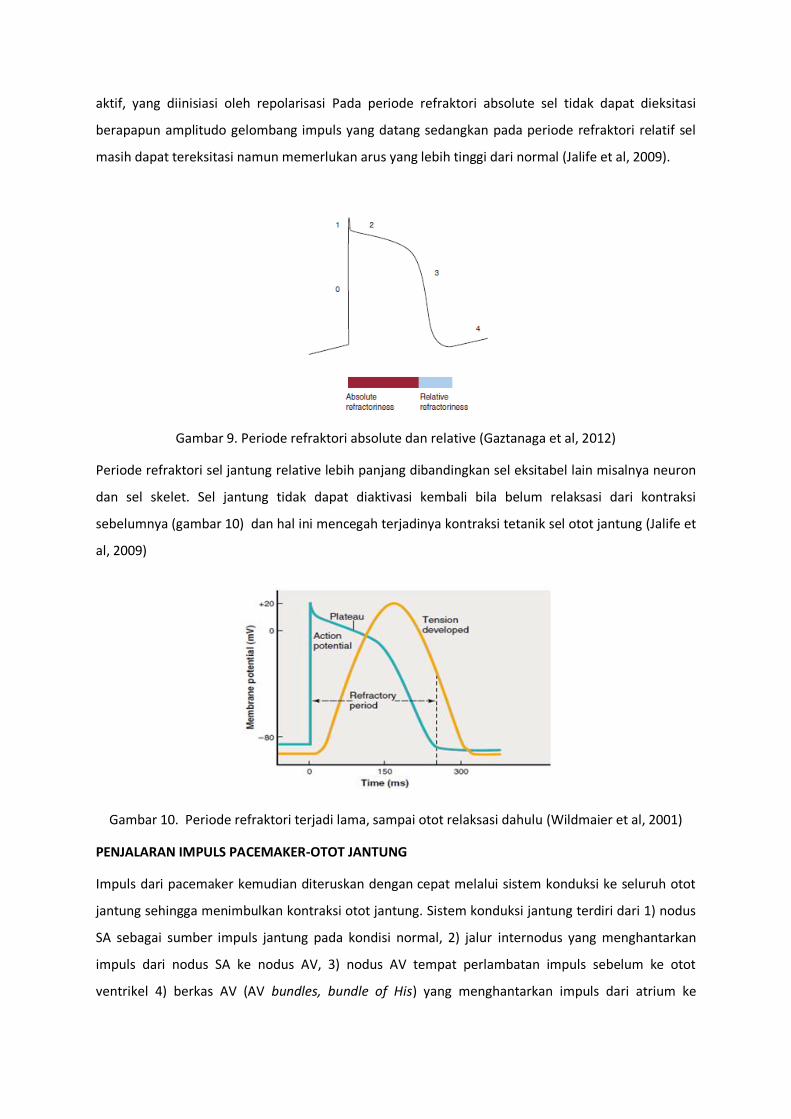
aktif, yang diinisiasi oleh repolarisasi Pada periode refraktori absolute sel tidak dapat dieksitasi
berapapun amplitudo gelombang impuls yang datang sedangkan pada periode refraktori relatif sel
masih dapat tereksitasi namun memerlukan arus yang lebih tinggi dari normal (Jalife et al, 2009).
Gambar 9. Periode refraktori absolute dan relative (Gaztanaga et al, 2012)
Periode refraktori sel jantung relative lebih panjang dibandingkan sel eksitabel lain misalnya neuron
dan sel skelet. Sel jantung tidak dapat diaktivasi kembali bila belum relaksasi dari kontraksi
sebelumnya (gambar 10) dan hal ini mencegah terjadinya kontraksi tetanik sel otot jantung (Jalife et
al, 2009)
Gambar 10. Periode refraktori terjadi lama, sampai otot relaksasi dahulu (Wildmaier et al, 2001)
PENJALARAN IMPULS PACEMAKER-OTOT JANTUNG
Impuls dari pacemaker kemudian diteruskan dengan cepat melalui sistem konduksi ke seluruh otot
jantung sehingga menimbulkan kontraksi otot jantung. Sistem konduksi jantung terdiri dari 1) nodus
SA sebagai sumber impuls jantung pada kondisi normal, 2) jalur internodus yang menghantarkan
impuls dari nodus SA ke nodus AV, 3) nodus AV tempat perlambatan impuls sebelum ke otot
ventrikel 4) berkas AV (AV bundles, bundle of His) yang menghantarkan impuls dari atrium ke
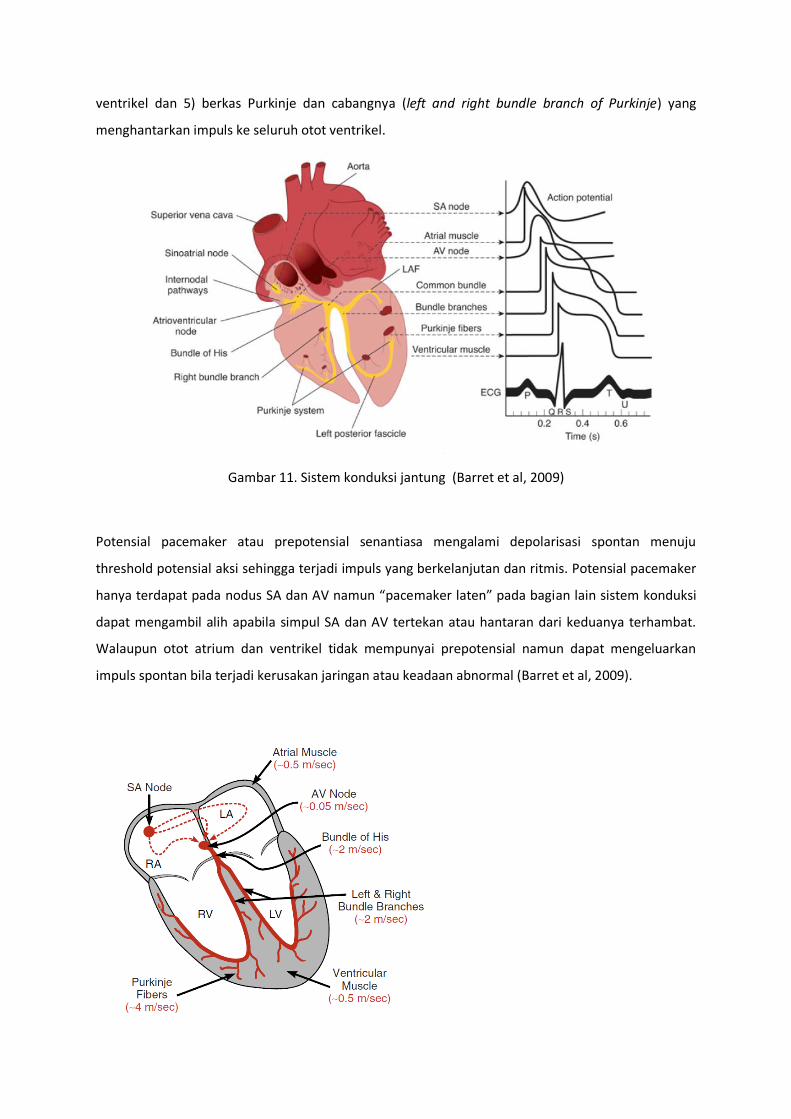
ventrikel dan 5) berkas Purkinje dan cabangnya (left and right bundle branch of Purkinje) yang
menghantarkan impuls ke seluruh otot ventrikel.
Gambar 11. Sistem konduksi jantung (Barret et al, 2009)
Potensial pacemaker atau prepotensial senantiasa mengalami depolarisasi spontan menuju
threshold potensial aksi sehingga terjadi impuls yang berkelanjutan dan ritmis. Potensial pacemaker
hanya terdapat pada nodus SA dan AV namun “pacemaker laten” pada bagian lain sistem konduksi
dapat mengambil alih apabila simpul SA dan AV tertekan atau hantaran dari keduanya terhambat.
Walaupun otot atrium dan ventrikel tidak mempunyai prepotensial namun dapat mengeluarkan
impuls spontan bila terjadi kerusakan jaringan atau keadaan abnormal (Barret et al, 2009).

Faktor ekstrinsik yang mempengaruhi kecepatan konduksi
PRINSIP MEKANISME ARITMIA
Mekanisme utama terjadinya aritmia adalah automatisitas, triggered activity dan reentry.
Mekanisme aritmia ini selanjutnya dikelompokan menjadi dua yaitu gangguan pembentukan impuls
dan gangguan konduksi impuls (tabel 4). Automatisitas dan trigerred activity termasuk dalam
kelompok gangguan pembentukan impuls sedangkan reentry adalah bentuk gangguan konduksi
impuls. Aritmia yang terjadi dapat disebabkan oleh terjadinya salah satu gangguan atau kombinasi
keduanya.
Tabel 4. Pembagian mekanisme aritmia jantung
(Gaztanaga et al, 2012)
GANGGUAN PEMBENTUKAN IMPULS
AUTOMATISITAS
Pada kondisi normal nodus SA paling cepat membentuk potensial aksi spontan (fastest rate of firing)
sehingga mencegah pembentukan impuls spontan pacemaker lain yang lebih lambat (gambar 12).
Setiap kali nodus SA terjadi potensial aksi maka impuls akan dijalarkan ke nodus AV, Bundle His dan
Purkinje. Sebelum pacemaker laten tersebut mencapai potensial threshold intrinsik, sudah
mendapat impuls kembali dari nodus SA sehingga mereka tidak sempat mencetuskan impuls sendiri.

Gambar 12. Frekuensi intrinsik pacemaker (Despopoulos, 2003)
Kecepatan pembentukan potensial aksi spontan ditentukan oleh interaksi 3 faktor yaitu a)
kemiringan depolarisasi fase 4 (prepotential, potensial pacemaker), b) potensial threshold untuk
inisiasi potensial aksi, dan c) maximum diastolic potential (MDP). Perubahan dari ketiga hal tersebut
dapat mengubah kecepatan inisiasi impuls (gambar 13).
Gambar 13. Faktor yang mempengaruhi potensial pacemaker (Despopoulos, 2003)
Tanda khas dari automatisitas normal adalah adanya “overdrive suppression”. Pade pacemaker laten
yang diambil alih (overdriving), frekuensi terbentuknya impuls terjadi lebih cepat dari frekuensi
intrinsiknya dan terjadi penurunan kemiringan fase 4 sebagian besar karena peningkatan aktivitas
pompa Na/K (terjadi hiperpolarisasi).
Pada pacemaker laten, arus depolarisasi dari pacemaker primer memicu masuknya Na ke dalam sel
(gambar 14). Peningkatan kadar natrium intrasel ini memicu pompa Na/K ATPase untuk
mengeluarkan natrium dan bertukar dengan kalium. Semakin sering terjadi arus depolarisasi maka
semakin aktif kerja pompa. Karena perbandingan ion Na masuk dan K keluar adalah 3:2 maka
potensial membrane semakin negatif (hiperpolarisasi). Hal ini mengimbangi proses depolarisasi

otomatis oleh kanal “funny” (If) sehingga sel pacemaker laten tidak pernah terdepolarisasi otomatis
mencapai potensial tresholdnya dan mencegah terjadinya potensial aksi spontan sesuai irama
intrinsiknya.
Gambar 14. Mekanisme ionik overdrive supression
Ketika stimulasi “overdrive” dihentikan (gambar 15), frekuensi pembentukan impuls kembali sesuai
dengan frekuensi intrinsik secara bertahap (disebut periode “warm-up”). Karena overdrive, terjadi
overload Na intrasel sehingga ketika “overdrive” berhenti, pompa Na/K memerlukan waktu untuk
kembali ke pola aktivitas dasarnya. Hal ini yang menjelaskan adanya jeda beberapa saat sebelum
pacemaker laten tersebut kembali ke frekuensi intrinsik. Tingkat supresi dan waktu recover
proporsional dengan frekuensi dan durasi stimulasi yang diberikan.
Gambar 15. Overdrive suppression dengan periode “warm up” (Castanaganza et al, 2012)
Mekanisme ini berperan penting dalam menjaga irama sinus, yang secara berkelanjutan
menghambat pacemaker laten dibawahnya. Pada pasien dengan pacemaker eksternal, irama
intrinsik juga disupresi melalui mekanisme ini. Tidak adanya “overdrive suppression”
mengindikasikan bahwa aritmia yang terjadi ditimbulkan oleh mekanisme selain peningkatan
automatisitas normal.
Aktivitas pacemaker dikontrol oleh sistem saraf otonom (gambar 16) dan dapat dipengaruhi oleh
berbagai factor sistemik termasuk gangguan metabolik, substansi endogen atau obat-obatan.

Gambar. 16. Frekuensi Instrinsik rate nodus SA dan interaksi dengan tonus otonom
Keadaan yang mempengaruhi firing rate nodus SA
AUTOMATISASI NORMAL YANG BERUBAH
Aktivitas parasimpatik mengurangi kecepatan pembentukan impuls pacemaker dengan melepaskan
acetylcholine (Ach) sehingga terjadi hiperpolarisasi sel melalui peningkatan konduktansi kanal K (IK-
Ach). Selain itu juga mengurangi aktivitas kanal Ca (ICa-L) dan aktivitas kanal Na funny (If) sehingga
makin memperlambat pembentukan impuls (gambar 17).
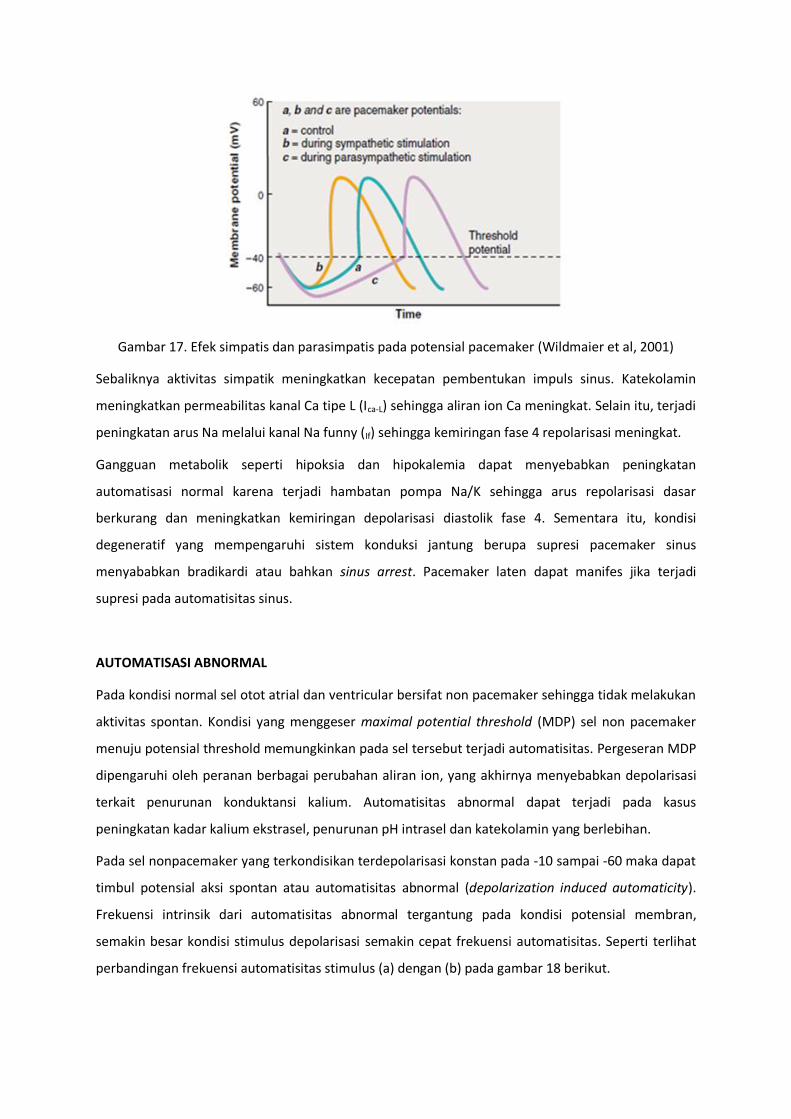
Gambar 17. Efek simpatis dan parasimpatis pada potensial pacemaker (Wildmaier et al, 2001)
Sebaliknya aktivitas simpatik meningkatkan kecepatan pembentukan impuls sinus. Katekolamin
meningkatkan permeabilitas kanal Ca tipe L (Ica-L) sehingga aliran ion Ca meningkat. Selain itu, terjadi
peningkatan arus Na melalui kanal Na funny (If) sehingga kemiringan fase 4 repolarisasi meningkat.
Gangguan metabolik seperti hipoksia dan hipokalemia dapat menyebabkan peningkatan
automatisasi normal karena terjadi hambatan pompa Na/K sehingga arus repolarisasi dasar
berkurang dan meningkatkan kemiringan depolarisasi diastolik fase 4. Sementara itu, kondisi
degeneratif yang mempengaruhi sistem konduksi jantung berupa supresi pacemaker sinus
menyababkan bradikardi atau bahkan sinus arrest. Pacemaker laten dapat manifes jika terjadi
supresi pada automatisitas sinus.
AUTOMATISASI ABNORMAL
Pada kondisi normal sel otot atrial dan ventricular bersifat non pacemaker sehingga tidak melakukan
aktivitas spontan. Kondisi yang menggeser maximal potential threshold (MDP) sel non pacemaker
menuju potensial threshold memungkinkan pada sel tersebut terjadi automatisitas. Pergeseran MDP
dipengaruhi oleh peranan berbagai perubahan aliran ion, yang akhirnya menyebabkan depolarisasi
terkait penurunan konduktansi kalium. Automatisitas abnormal dapat terjadi pada kasus
peningkatan kadar kalium ekstrasel, penurunan pH intrasel dan katekolamin yang berlebihan.
Pada sel nonpacemaker yang terkondisikan terdepolarisasi konstan pada -10 sampai -60 maka dapat
timbul potensial aksi spontan atau automatisitas abnormal (depolarization induced automaticity).
Frekuensi intrinsik dari automatisitas abnormal tergantung pada kondisi potensial membran,
semakin besar kondisi stimulus depolarisasi semakin cepat frekuensi automatisitas. Seperti terlihat
perbandingan frekuensi automatisitas stimulus (a) dengan (b) pada gambar 18 berikut.

Gambar 18. Frekuensi automatisitas meningkat dengan peningkatan depolarisasi (Jalife et al, 2009)
Berbeda dengan peningkatan automatisitas normal, pada automatisitas abnormal berciri kurang
sensitive terhadap “overdrive suppression” walaupun pada kondisi tertentu dapat teramati.
TRIGERRED ACTIVITY
Trigerred activity (TA) terjadi setelah inisiasi impuls karena “afterdepolarisasi” (oskilasi potensial
membran selama atau segera setelah potensial aksi). Afterdepolarisasi hanya terjadi jika ada
potensial aksi yang mendahului (sebagai trigger) dan bila sudah mencapai potensial threshold akan
membentuk potensial aksi yang baru. Hal ini dapat sebagai sumber terpicunya respon baru, menjadi
potensial aksi tersendiri.
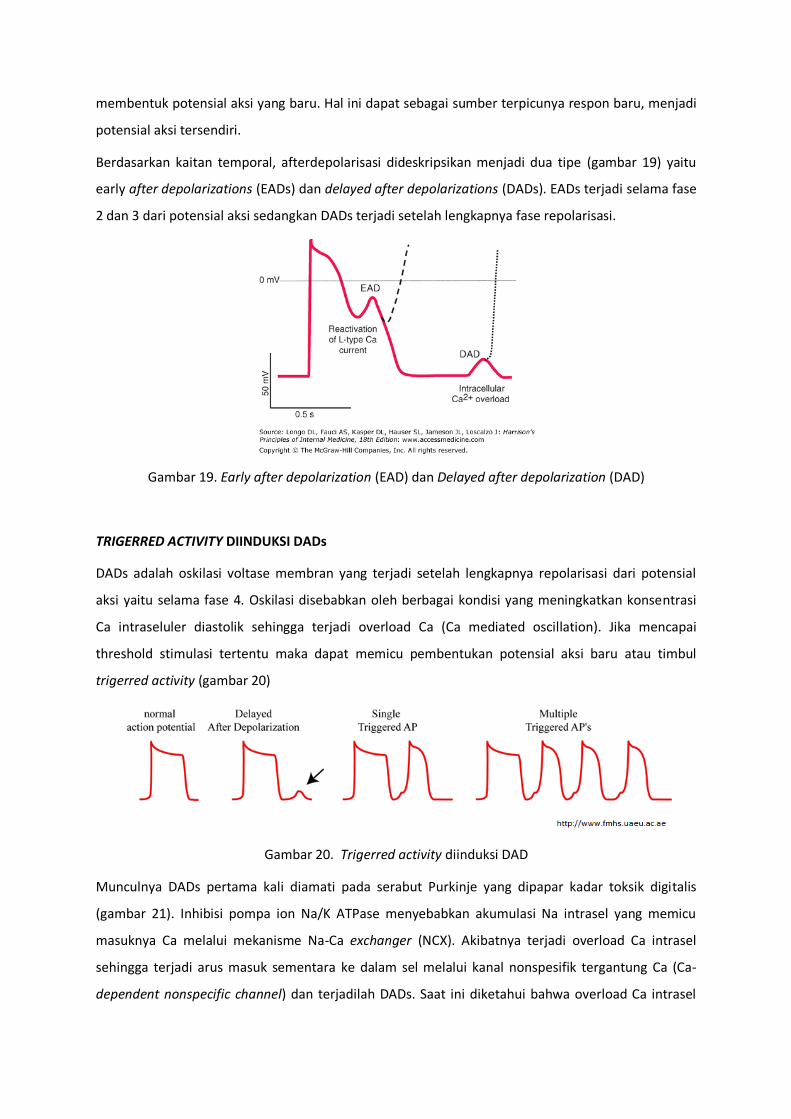
Berdasarkan kaitan temporal, afterdepolarisasi dideskripsikan menjadi dua tipe (gambar 19) yaitu
early after depolarizations (EADs) dan delayed after depolarizations (DADs). EADs terjadi selama fase
2 dan 3 dari potensial aksi sedangkan DADs terjadi setelah lengkapnya fase repolarisasi.
Gambar 19. Early after depolarization (EAD) dan Delayed after depolarization (DAD)
TRIGERRED ACTIVITY DIINDUKSI DADs
DADs adalah oskilasi voltase membran yang terjadi setelah lengkapnya repolarisasi dari potensial
aksi yaitu selama fase 4. Oskilasi disebabkan oleh berbagai kondisi yang meningkatkan konsentrasi
Ca intraseluler diastolik sehingga terjadi overload Ca (Ca mediated oscillation). Jika mencapai
threshold stimulasi tertentu maka dapat memicu pembentukan potensial aksi baru atau timbul
trigerred activity (gambar 20)
Gambar 20. Trigerred activity diinduksi DAD
Munculnya DADs pertama kali diamati pada serabut Purkinje yang dipapar kadar toksik digitalis
(gambar 21). Inhibisi pompa ion Na/K ATPase menyebabkan akumulasi Na intrasel yang memicu
masuknya Ca melalui mekanisme Na-Ca exchanger (NCX). Akibatnya terjadi overload Ca intrasel
sehingga terjadi arus masuk sementara ke dalam sel melalui kanal nonspesifik tergantung Ca (Ca-
dependent nonspecific channel) dan terjadilah DADs. Saat ini diketahui bahwa overload Ca intrasel

dapat terjadi selain mekanisme inhibisi pompa ion Na/K ATPase. Stimulus lain yang menyebabkan
overload Ca intrasel adalah katekolamin, iskemia, hypertofi, hipokalemia dan hiperkalemia.
Gambar 21. Mekanisme Ionik DADs (Jalife, 2009)
Katekolamin menyebabkan overload Ca intraseluler via peningkatan aliran masuk Ca melalui kanal
Ca tipe L (ICa-L) dan peningkatan aliran Na-Ca exchanger. DADs yang diinduksi iskemia diduga
dimediasi oleh akumulasi lipofosfogliserida pada jaringan iskemik dengan diikuti peningkatan Na dan
Ca intrasel. Overload Ca intrasel juga dapat disebabkan oleh fungsi reticulum sarkoplasmikum yang
abnormal (misal mutasi reseptor ryanodin), memfasilitasi aritmia klinis seperti catecholaminergic
polymorphic VT.
Adenosin telah dipakai sebagai uji diagnosis DADs. Adenosin mengurangi aliran masuk ion Ca secara
indirek dengan menghambat adenilat siklase dan cyclic adenosine monophosphate (cAMP).
Adenosin dapat menghilangkan DADs yang diinduksi katekolamin namun tidak mengubah DADs yang
diiduksi inhibisi pompa Na/K. Adanya interupsi VT dengan pemberian adenosine menunjukan bahwa
mekanisme aritmia adalah karena DADs yang diinduksi katekolamin.
Karakteristik utama DADs adalah dapat kemunculanya semakin dipicu oleh peningkatan frekuensi
stimulasi. Frekuensi dan durasi stimulasi selanjutnya terkait dengan amplitudo dan jumlah triggered
activity. Misalnya pada skema munculnya DADs pada serabut Purkinje yang dipapar digitalis kadar
toksik (gambar 22). Stimulasi potensial aksi dengan frekuensi yang relative lambat akan diikuti oleh
fase 4 yang normal (a). Jika stimulasi semakin cepat maka terdapat dua gelombang oskilasi selama
fase 4 (b). Apabila stimulasi lebih dipercepat maka DADs mencapai threshold sehingga munculah
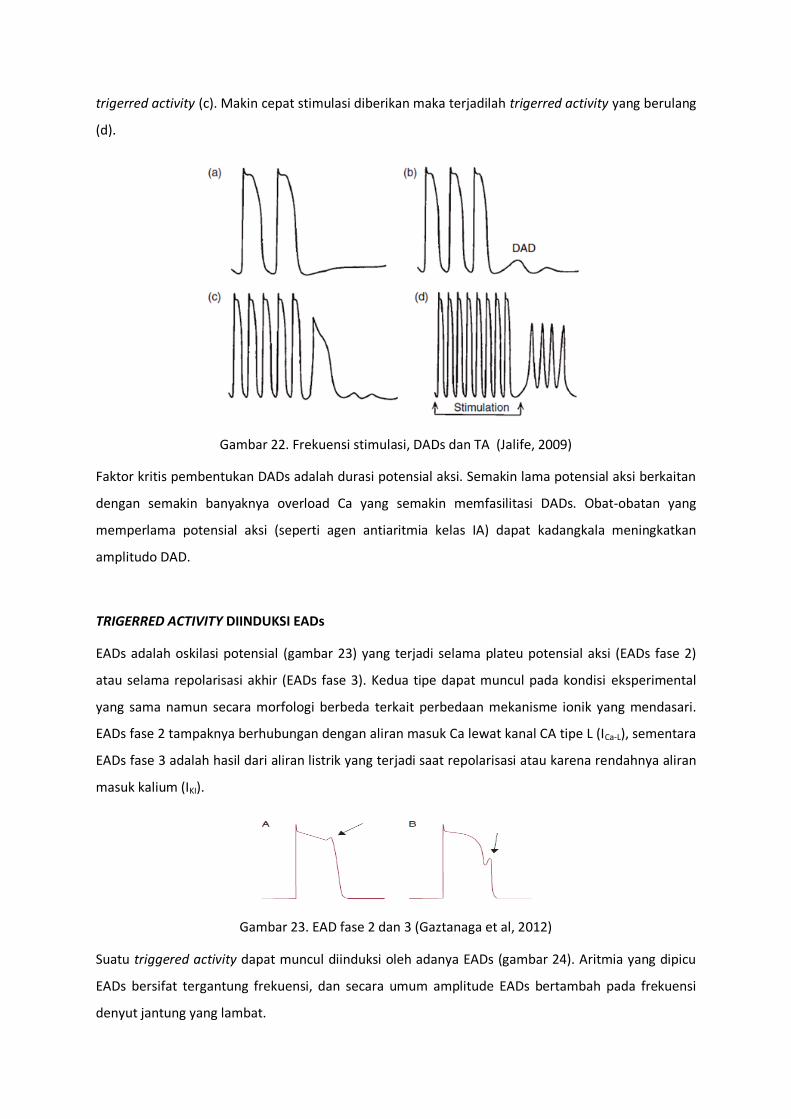
trigerred activity (c). Makin cepat stimulasi diberikan maka terjadilah trigerred activity yang berulang
(d).
Gambar 22. Frekuensi stimulasi, DADs dan TA (Jalife, 2009)
Faktor kritis pembentukan DADs adalah durasi potensial aksi. Semakin lama potensial aksi berkaitan
dengan semakin banyaknya overload Ca yang semakin memfasilitasi DADs. Obat-obatan yang
memperlama potensial aksi (seperti agen antiaritmia kelas IA) dapat kadangkala meningkatkan
amplitudo DAD.
TRIGERRED ACTIVITY DIINDUKSI EADs
EADs adalah oskilasi potensial (gambar 23) yang terjadi selama plateu potensial aksi (EADs fase 2)
atau selama repolarisasi akhir (EADs fase 3). Kedua tipe dapat muncul pada kondisi eksperimental
yang sama namun secara morfologi berbeda terkait perbedaan mekanisme ionik yang mendasari.
EADs fase 2 tampaknya berhubungan dengan aliran masuk Ca lewat kanal CA tipe L (ICa-L), sementara
EADs fase 3 adalah hasil dari aliran listrik yang terjadi saat repolarisasi atau karena rendahnya aliran
masuk kalium (IKI).
Gambar 23. EAD fase 2 dan 3 (Gaztanaga et al, 2012)
Suatu triggered activity dapat muncul diinduksi oleh adanya EADs (gambar 24). Aritmia yang dipicu
EADs bersifat tergantung frekuensi, dan secara umum amplitude EADs bertambah pada frekuensi
denyut jantung yang lambat.

Gambar 24. Trigerred activity diinduksi EAD
Plateu dari potensial aksi adalah periode ketika kondisi resistensi membran tinggi dan aliran arus
sedikit. Akibatnya, perubahan kecil pada arus yang menyebabkan repolarisasi atau depolarisasi
dapat berpengaruh terhadap durasi dan profil potensial aksi. Berbagai jenis zat dan kondisi yang
menurunkan arus keluar atau meningkatkan arus masuk (sehingga menggeser current outward yang
normal) dapat menyebabkan kondisi yang diperlukan untuk terjadinya EADs.
Tabel 5. Obat yang dapat menginduksi EAD
(Gaztanaga et al, 2012).
Kondisi utama yang mendasari pembentukan EADs adalah pemanjangan potensial aksi yang tampak
pada electrocardiogram (ECG) sebagai pemanjangan interval QT. Bebrapa obat antiaritmia, terutama
kelas IA dan III dapat menjadi proaritmia karena efek terapinya adalah pemanjangan potensial aksi.
Banyak obat lainya dapat sebagai predisposisi pembentukan EADs terutama ketika dihubungkan
dengan kondisi hipokalemia dan atau bradikardia sebagai faktor tambahan yang menyebabkan
pemanjangan potensial aksi. Katekolamin mungkin dapat meningkatkan EADs dengan meningkatkan
arus Ca, walaupun resultan peningkatan denyut jantung bersamaan dengan peningkatan arus K
secara efektif mengurangi durasi potensial aksi sehingga meniadakan EADs.

GANGGUAN KONDUKSI IMPULS
Perlambatan atau blok konduksi terjadi ketika impuls yang menjalar gagal untuk dihantarkan.
Berbagai faktor aktif dan pasif pada membran menentukan kecepatan dan suksesnya konduksi.
Misalnya adalah faktor efikasi impuls yang menstimulasi dan eksitabilitas jaringan dimana impuls
dihantarkan. Gap junction (gambar 25) berperan penting dalam cepat dan suksesnya konduksi
impuls.
Gambar 25. Gap Junction (Mohrman and Haller, 2006)
Umumnya blok konduksi impuls terjadi pada frekuensi impuls yang tinggi sebagai akibat dari
recoveri yang tidak lengkap dari kondisi refraktori. Ketika suatu impuls berikutnya sampai pada
jaringan yang masih dalam periode refraktori maka impuls tidak dapat dihantarkan atau dihantarkan
dengan aberasi. Hal ini merupakan mekanisme khas yang menjelaskan beberapa fenomena seperti
blok konduksi berkas cabang dari denyut premature, fenomena Ashman selama atrial fibrillation
(AF) dan acceleration dependent aberration.
Bradikardia atau deceleration dependent block diduga disebabkan berkurangnya eksitabilitas pada
interval diastolik yang panjang dengan berkurangnya amplitude potensial aksi.
Banyak faktor yang dapat mempengaruhi konduksi, termasuk frekuensi, tonus otonom, obat (misal
Ca channel blocker, digitalis, adenosine/adenosine trifosfat) atau proses degeneratif (dengan
mengubah fisiologi jaringan dan kapasitas untuk konduksi impuls).
REENTRY
Ketika impuls normal dari nodus sinoatrial telah menjalar ke seluruh ventrikel maka ventrikel akan
masuk ke periode refraktori. Selanjutnya impuls akan berhenti dan tidak akan menjalar lebih jauh
lagi. Apabila terdapat serabut otot tertentu yang tidak teraktivasi saat inisial impuls menjalar (misal
damaged area), maka bagian tersebut dapat lebih dullu mengalami recoveri eksitabilitasnya
sebelum impuls menghilang. Akibatnya bagian tersebut dapat berperan sebagai penghubung untuk
terjadinya reeksitasi di area lain yang sebelumnya terdepolarisasi saat penjalaran impuls inisial tetapi
telah recoveri. Reentry terjadi ketika impuls yang menjalar gagal menghilang dan kemudian me-

reeksitasi jantung yang telah selesai periode refraktorinya (Guyton & Hall, 2006). Karena terjadi
penjalaran berulang impuls yang kembali lagi ke tempat asal untuk mereaktivasi, maka fenomena ini
disebut reentry, reentrant excitation, circus movement, reciprocal/echo beats, atau reciprocating
tachycardia (RT).
Gambar 26. Anatomic reentry dan Functional reentry (Veenhuyzen et al, 2004)
Reentry dibagi menjadi dua kelompok yaitu anatomical/classic reentry dan functional reentry
(gambar 26). Pada anatomical reentry terdapat struktur anatomi yang terlibat sedangkan pada
functional reentry terdapat hambatan fungsional tanpa kelainan struktur. Reentry adalah mekanisme
aritmia yang paling sering dijumpai di klinis (Gaztanaga et al, 2012).
ANATOMICAL REENTRY
Nama lain dari anatomical reentry adalah classic reentry dan digambarkan dengan ring model
(gambar 26). Mekanisme classic reentry didasarkan pada hambatan anatomis berupa area tidak
dapat tereksitasi dikelilingi jalur sirkuler sehingga gelombang awal dapat masuk kembali (reenter),
membentuk sirkuit reentrant yang stabil (Gaztanaga et al, 2012).

(Gaztanaga et al, 2012)
Inisiasi dan keberlangsungan reentry tergantung pada kecepatan konduksi dan periode refraktori di
setiap jalur yang menentukan panjangnya gelombang depolarisasi (wavelength = kecepatan
konduksi x periode refraktori). Agar reentry terjadi panjang gelombang depolarisasi (wavelength)
harus lebih pendek dari panjang jalur (pathway) agar pada tempat semula telah recoveri atau selesai
dari periode refraktori w. Hal ini akan memungkinkan terbentuknya excitable gap (gambar 27).
Gambar 27. Excitable gap (Gaztanaga et al, 2012)
Excitable gap adalah konsep kunci yang penting untuk memahami mekanisme reentry. Excitable gap
adalah excitable myocardium yang berada diantara “head” gelombang reentrant dan “tail” dari
gelombang yang mendahuluinya. Karena area tersebut telah recoveri dari periode refraktori maka
gelombang reentrant dapat kembali menjalar di sirkuit (Gaztanaga et al, 2012).
Gambar 26. Anatomical reentry
Hambatan secara structural anatomi di bagian
sentral akan membentuk dua jalur. Pada
penjalaran impuls, bila terjadi blok searah pada
satu jalur (unidirectional block) dan konduksi
yang lambat melalui jalur lainya maka akan
memungkinkan terjadinya reentry.

Gambar 28. Skema terjadinya reentry
Gambar 28 diatas merupakam skema terjadinya sirkuit reentry (A) Sirkuit memiliki dua jalur, salah
satunya jalur konduksi lambat (kanan). (B) Terdapat blok konduksi anterograde di jalur cepat dan
konduksi tetap terjadi di jalur lambat. Selanjutnya, karena jalur cepat telah recoveri maka
gelombang aktivasi dapat masuk kembali (reenter) ke jalur cepat (secara retrograde). (C) Selama
reentry yang berkelanjutan akan membentuk sebuah sirkuit dengan gap (excitable gap) berada
diantara “head” gelombang reentrant dan “tail” dari gelombang yang mendahuluinya
Inisiasi reentry yang diawali terbentuknya excitable gap ditentukan oleh 3 faktor yaitu 1) Jalur yang
akan dilalui impuls terlalu panjang (too long pathway), 2) kecepatan konduksi melambat, 3) periode
refraktori memendek. Ketiga hal tersebut terjadi pada kondisi patologis yang berbeda. Jalur yang
memanjang khas terjadi pada dilatasi jantung sedangkan penurunan kecepatan konduksi terjadi
pada blok pada sistem Purkinje, iskemi otot, hiperkalemia dan lainya. Pemendekan periode
refraktori terjadi karena obat seperti epinefrin atau stimulasi elektrik yang berulang (Guyton & Hall,
2006).
FUNCTIONAL REENTRY
Pada reentry fungsional, sirkuit yang terbentuk tidak ditentukan oleh hambatan anatomis namun
oleh heterogenitas dinamik properti elektrofisiologik jaringan yang terlibat. Lokasi dan ukuran dari
sirkuit reentrant fungsional dapat bervariasi namun biasanya kecil dan tidak stabil. Reentry
fungsional terjadi karena mekanisme yang berbeda-beda, terdiri dari leading cycle reentry,
anisotropic reentry, figure of eight reentry, reflection, spiral wave activity (Gaztanaga et al, 2012).
MEMBEDAKAN MEKANISME ARITMIA
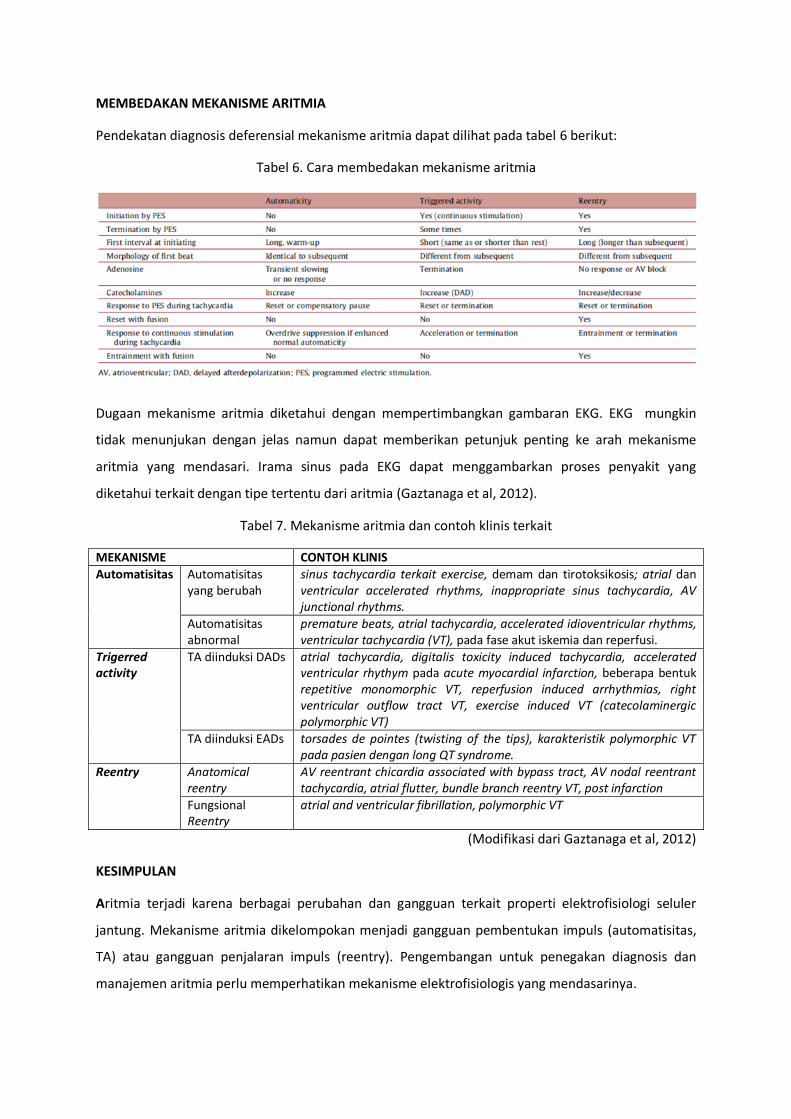
Pendekatan diagnosis deferensial mekanisme aritmia dapat dilihat pada tabel 6 berikut:
Tabel 6. Cara membedakan mekanisme aritmia
Dugaan mekanisme aritmia diketahui dengan mempertimbangkan gambaran EKG. EKG mungkin
tidak menunjukan dengan jelas namun dapat memberikan petunjuk penting ke arah mekanisme
aritmia yang mendasari. Irama sinus pada EKG dapat menggambarkan proses penyakit yang
diketahui terkait dengan tipe tertentu dari aritmia (Gaztanaga et al, 2012).
Tabel 7. Mekanisme aritmia dan contoh klinis terkait
MEKANISME CONTOH KLINIS
Automatisitas Automatisitas yang berubah
sinus tachycardia terkait exercise, demam dan tirotoksikosis; atrial dan ventricular accelerated rhythms, inappropriate sinus tachycardia, AV junctional rhythms.
Automatisitas abnormal
premature beats, atrial tachycardia, accelerated idioventricular rhythms, ventricular tachycardia (VT), pada fase akut iskemia dan reperfusi.
Trigerred activity
TA diinduksi DADs atrial tachycardia, digitalis toxicity induced tachycardia, accelerated ventricular rhythym pada acute myocardial infarction, beberapa bentuk repetitive monomorphic VT, reperfusion induced arrhythmias, right ventricular outflow tract VT, exercise induced VT (catecolaminergic polymorphic VT)
TA diinduksi EADs torsades de pointes (twisting of the tips), karakteristik polymorphic VT pada pasien dengan long QT syndrome.
Reentry Anatomical reentry
AV reentrant chicardia associated with bypass tract, AV nodal reentrant tachycardia, atrial flutter, bundle branch reentry VT, post infarction
Fungsional Reentry
atrial and ventricular fibrillation, polymorphic VT
(Modifikasi dari Gaztanaga et al, 2012)
KESIMPULAN
Aritmia terjadi karena berbagai perubahan dan gangguan terkait properti elektrofisiologi seluler
jantung. Mekanisme aritmia dikelompokan menjadi gangguan pembentukan impuls (automatisitas,
TA) atau gangguan penjalaran impuls (reentry). Pengembangan untuk penegakan diagnosis dan
manajemen aritmia perlu memperhatikan mekanisme elektrofisiologis yang mendasarinya.

DAFTAR PUSTAKA
Barrett K E, Barman S M, Boitano S, Brooks H. Ganong's Review of Medical Physiology, 23th ed.
McGraw-Hill. 2009.
Despopoulos, A, Silbernagl S. Color Atlas Of Physiology. Thieme. 2003.
Dorland, W. A. Newman. Dorland's illustrated medical dictionary. Philadelphia, PA: Saunders. 2007.
Gaztañaga L, Marchlinski FE, Betensky BP. Mechanisms of cardiac arrhythmias. Rev Esp Cardiol. 2012 Feb;65(2):174-85.
Guyton AC, Hall JE. Textbook of Medical Physiology, 11th ed. Philadelphia, PA: Elsevier, 2006.
Jalife J, Delmar M, Davidenko, Anumonwo J, Berenfeld O, Anumonwo KJ. Basic cardiac electrophysiology for the clinician. 2nd ed. New Jersey: Wiley-Blackwell; 2009.
Mohrman D, Heller L. Cardiovascular Physiology 6th ed McGraw Hill 2006.
Widmaier EP, Raff H, Strang KT. Vander, Sherman and Luciano's Human Physiology: the Mechanisms
of Body Function, 8th ed. Boston, MA: McGraw-Hill Higher Education, 2001.
Veenhuyzen G D, Simpson C S, Abdollah H. Atrial fibrillation. CMAJ September 28, 2004 vol. 171
