bab ii tinjauan pustaka 2.1 nira aren - sinta.unud.ac.id · 2.1.5 bakteri asam laktat dalam nira...
TRANSCRIPT

BAB II
TINJAUAN PUSTAKA
2.1 Nira Aren
Tanaman aren (Arenga pinnata MERR.) merupakan tanaman biji tertutup
(Angiospermae) yaitu biji buahnya terbungkus daging buah. Tanaman aren ini
termasuk suku pinang-pinangan (Aracaceae). Tanaman aren banyak terdapat
mulai dari pantai timur India sampai ke daerah Asia Tenggara. Indonesia
merupakan daerah terbesar produksi tanaman aren di dunia. Hampir 60% pohon
aren di dunia terdapat di Indonesia dan tersebar hampir di seluruh wilayah
nusantara yakni di Sulawesi, Maluku, Papua, dan Sumatera (Sunanto, 1992).
Sulawesi Utara merupakan salah satu daerah produksi tanaman aren yang cukup
besar.
Gambar 2.1Pohon Aren (Arenga pinnata. MERR)

Pohon aren (Gambar 2.1) umumnya ditemukan tumbuh secara liar (tidak
ditanam orang) dan hampir semua bagian dari pohon ini dapat dimanfaatkan, serta
memiliki nilai ekonomis tinggi mulai dari bagian-bagian fisik pohon maupun dari
hasil-hasil produksinya (Baharuddin et al., 2008). Pohon aren atau enau/seho
tumbuh mencapai ketinggian hingga 20 meter dengan garis tengah batang
mencapai 65 cm. Batang aren dibungkus oleh pelepah daun dan ijuk yang melekat
pada pangkal pelepah. Ijuk dapat dipanen setelah tanaman berumur 4 tahun dan
terus dipanen hingga 8-10 tahun, tergantung jenis dan pertumbuhan tanaman.
Batang berkulit keras yang membungkus jaringan gabus yang mengandung pati.
Kandungan pati mencapai maksimum sebelum tanaman berbunga dan menurun
drastis ketika tanaman disadap (Pribadi, 2009). Bagian bunga betina akan masak
dalam 1-3 tahun, bunga betina yang masih muda dapat diolah menjadi kolang-
kaling, dalam satu tandan buah aren masak tidak serempak. Bunga betina
menggandung 2-3 biji dengan kulit yang keras. Jumlah bunga betina berkisar 5-8
ribu biji pertandan. Nira diperoleh dari tangkai bunga jantan yang disadap ketika
tanaman aren berumur lima tahun, puncak produksi nira aren terjadi pada saat
tanaman aren berumur 15-20 tahun.
Proses penyadapan nira biasanya ditampung dalam bumbung (batang
bambu sepanjang satu meter), dan proses penampungan nira dapat berlangsung
hingga tiga bulan terus menerus tanpa henti. Setiap pohon dapat menghasilkan 10-
20 liter nira per hari dengan dua kali penyadapan yaitu waktu pagi dan sore hari
(Burhanuddin, 2005; Duryatmo dan Helmina, 2007). Nira aren dalam keadaan
segar berasa manis, berbau khas nira dan tidak berwarna. Nira yang baru menetes

dari tandan bunga mempunyai pH + 7, akan tetapi pengaruh keadaan sekitarnya
menyebabkan nira mudah terkontaminasi dan mengalami fermentasi secara alami
sehingga berubah menjadi asam (Lempang dan Mangopang, 2012). Kandungan
gula yang tinggi dalam nira aren memungkinkan nira aren dijadikan sebagai bahan
baku pembuatan gula aren. Selain itu juga nira aren bila didestilasi dapat
dikembangkan menjadi sumber biofuel (ethanol). Nira aren juga diolah secara
tradisional menjadi minuman beralkohol tinggi yang disebut “cap tikus”
(Tangkuman et al., 2010).
2.1.1 Komposisi kimia nira aren
Kandungan sukrosa nira aren lebih besar jika dibandingkan kandungan
sukrosa dari nira tebu dan nira siwalan. Hasil analisis komposisi kimia aren segar
asal Kabupaten Maros Propinsi Sulawesi Selatan dan Kota Tomohon Provinsi
Sulawesi Utara disajikan pada Tabel 2.1.
Nira aren mengandung beberapa zat gizi antara lain karbohidrat, protein,
lemak dan mineral. Rasa manis dari nira aren disebabkan kandungan karbohidrat
totalnya mencapai 11,28%. Komposisi nira aren tergantung pada letak daerah,
umur pohon dan umur tangkai bunga yang disadap (Sunanto, 1992). Tabel 2.2
menunjukkan komposisi kandungan nira pada beberapa jenis tanaman palma.
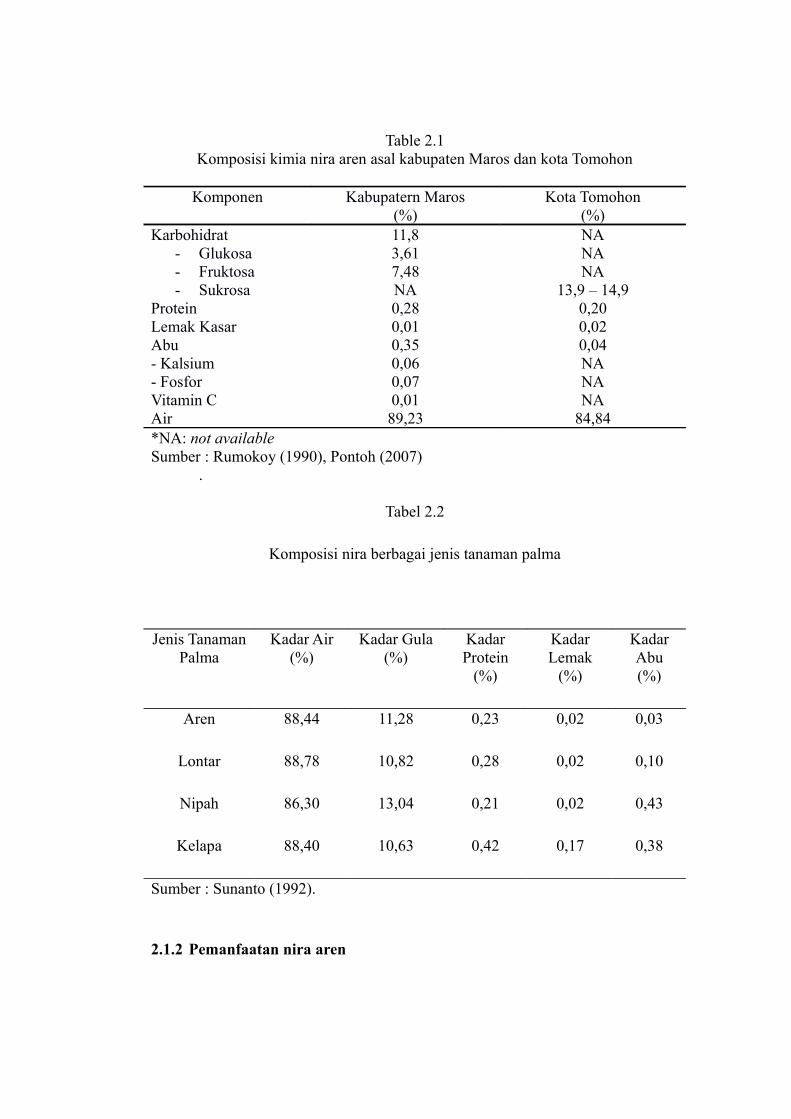
Table 2.1Komposisi kimia nira aren asal kabupaten Maros dan kota Tomohon
Komponen Kabupatern Maros(%)
Kota Tomohon(%)
Karbohidrat- Glukosa- Fruktosa- Sukrosa
11,83,617,48NA
NANANA
13,9 – 14,9Protein 0,28 0,20Lemak Kasar 0,01 0,02Abu- Kalsium - Fosfor
0,350,060,07
0,04NANA
Vitamin C 0,01 NAAir 89,23 84,84*NA: not availableSumber : Rumokoy (1990), Pontoh (2007)
.
Tabel 2.2
Komposisi nira berbagai jenis tanaman palma
Jenis TanamanPalma
Kadar Air(%)
Kadar Gula(%)
KadarProtein
(%)
KadarLemak
(%)
KadarAbu(%)
Aren 88,44 11,28 0,23 0,02 0,03
Lontar 88,78 10,82 0,28 0,02 0,10
Nipah 86,30 13,04 0,21 0,02 0,43
Kelapa 88,40 10,63 0,42 0,17 0,38
Sumber : Sunanto (1992).
2.1.2 Pemanfaatan nira aren

Nira aren adalah cairan yang mudah mengalami kerusakan, penyebab
utama rusaknya nira adalah akibat kontaminasi oleh mikroorganisme. Jenis
mikroorganisme yang mengkontaminasi nira aren adalah khamir dan bakteri.
Bakteri yang mengkontaminasi nira aren adalah Saccharomyces sp. dan
Acetobacter sp., nira yang telah terkontaminasi oleh mikroorganisme, akan
mengalami proses fermentasi atau perombakan terhadap senyawa-senyawa
penyusunnya. Proses fermentasi ini melibatkan sukrosa yang terdapat dalam nira
yang akan berubah menjadi alkohol dan selanjutnya berubah menjadi asam
(Firdaus dan Sinda, 2003).
Produk pangan yang paling banyak dihasilkan dari bahan baku nira aren
adalah gula aren, cuka, dan alkohol. Adanya kemajuan teknologi saat ini
memungkinkan pengolahan nira aren menjadi bioethanol, pengolahan gula kristal
dan sebagai pengembang dalam bahan pangan. Efektifitas nira aren sebagai bahan
pengembang makanan lebih diperuntukkan dalam pembuatan kue, karena hanya
memerlukan pengembang dalam jumlah yang sedikit, untuk penggunaan nira aren
dalam pembuatan roti, belum dapat maksimal seperti penggunaan ragi yang dijual
di pasaran (Lempang dan Mangopang, 2012).
2.1.3 Fermentasi nira aren
Reaksi pertama yang terjadi pada nira adalah inversi sukrosa oleh enzim
yang terdapat dalam nira. Pada reaksi yang kedua glukosa dan fruktosa hasil
inversi difermentasi menjadi etanol, dan pada reaksi ketiga terjadi oksidasi etanol
oleh bakteri A. aceti menjadi asam asetat. Peristiwa inversi tersebut terjadi karena

sukrosa terhidrolisa menjadi D-glukosa dan D-fruktosa, hal ini disebabkan oleh
aktivitas enzim fruktoforanosidase (-h-fruktosidase, invertase) yang dihasilkan
mikroba. Jika terjadi fermentasi lanjut maka kadar gula akan menurun, kadar
alkohol meningkat kemudian terjadi peningkatan kadar asam sehingga pH
cenderung turun (IPBlink, 2013).
Asam piruvat yang dihasilkan dari proses fermentasi pertama dapat
dipecah menjadi alkohol, asam laktat, asam butirat, asam propionat, dan asam
asetat tergantung pada jenis bakteri yang berperan di dalamnya. Bakteri asam
laktat umumnya menghasilkan sejumlah besar asam laktat dari fermentasi substrat
berkarbohidrat melalui jalur glikolisis (Buckle et al., 2009).
Reaksi kimia yang terjadi dalam proses fermentasi nira adalah sebagai
berikut :
C12H22O11 + H2O C6H12O6 + C6H12O6 (1)
Sukrosa Air (invertasi) Glukosa Fruktosa
Apabila pH nira turun (asam) atau terdapat enzim invertase yang berasal dari
mikroorganisme, maka akan terjadi inversi sukrosa menjadi glukosa/fruktosa.
C6H12O6 2 C5H5OH + 2 CO2 (2)
Glukosa /Sukrosa etil alkohol karbondioksida

Pada reaksi ini terjadi proses fermentasi yang menghasilkan akohol dan
dilepaskannya CO2.
C5H5OH + O2 CH3COOH + H2O (3)
etil alkohol asam asetat
Pada reaksi ini terjadi penguraian alkohol menjadi asam asetat, apabila
terjadi oksidasi atau adanya aktivitas Acetobacter sp. Perubahan atau reaksi (1)
sampai (2) merupakan kegiatan mikroorganisme (ragi) Saccharomyces sp.
Sedangkan (C2H2O5) dari proses (2) sampai reaksi (3) merupakan kegiatan bakteri
Acetobacter sp. (Wibowo, 2006).
2.1.4 Mikroba dalam nira aren
Mikroba yang alami terdapat dalam nira aren adalah khamir dan bakteri,
jenis khamir dari genus Saccharomyses sp. yakni Saccharomyses cerivisae telah
dikenal sebagai mikroorganisme yang dapat memfermentasi gula (glukosa) dan
mengubahnya menjadi alkohol dan CO2 (Soeseno, 1992 ; Budiyanto, 2004 ).
Nira aren di Sulawesi Utara dijadikan minuman keras dengan sebutan Cap
Tikus, nira aren juga digunakan sebagai bahan baku pembuatan bioethanol.
Bioethanol yang dihasilkan dari proses produksi di daerah Minahasa Selatan
berkadar etanol 35% yang diperoleh dari 9 liter nira. Industri farmasi dan

kosmetik memerlukan etanol berkadar 99,6%. Menurut penelitian yang dilakukan
1 liter bioethanol 99,6% menghabiskan 15 liter nira aren (Pribadi, 2009).
Penelitian Muharani (2011), menunjukkan kemajuan pemanfaatan nira
aren ke arah bioteknologi dengan memanfaatkan mikroba yang terkandung dalam
nira aren sebagai minuman probiotik. Penelitian yang dilakukan mencakup
berbagai jenis nira antara lain nira aren, nira tebu dan nira kelapa. Total bakteri
probiotik yang diperoleh pada nira aren sebesar 45,35 x 1013 CFU/ml, sedangkan
pada nira tebu 34,50 x 1013CFU/ml dan nira kelapa sebesar 4,29 x 1013 CFU/ml.
James dan Chen (1985) menyatakan bahwa nira memiliki kandungan gula
yang tidak cukup untuk menghambat pertumbuhan mikroorganisme, bahkan
merupakan media yang baik untuk pertumbuhan mikroorganisme. Cahyaningsih
(2006) melakukan isolasi mikroorganisme pada beberapa waktu fermentasi nira
lontar (Tabel 2.3). Berdasarkan data ini maka dapat dinyatakan bahwa bakteri
asam laktat (BAL) merupakan mikroorganisme awal yang bertanggung jawab
dalam fermentasi awal nira.
Tabel 2.3
Mikroorganisme dari nira lontar pada berbagai waktu fermentasi
Waktufermentasi
(jam)
Nilai pH Jenis Mikroorganisme Total BAL(CFU/ml)
0 6,5 Khamir dan BAL 3,6 x 102
6 5,3 Bacillus, khamir dan BAL 6,8 x 107
12 4,8 Bacillus, khamir dan BAL 6,4 x 105

24 4,1 Bacillus, khamir dan BAL 7,1 x 103
36 3,6 Bacillus dan khamir NA48 3,6 Bacillus dan khamir NA
Sumber : Cahyaningsih (2006)
2.1.5 Bakteri asam laktat dalam nira aren
Kehadiran BAL di dalam fermentasi nira berlangsung selama 24 jam.
Setelah itu mikroorganisme yang tetap bertahan adalah khamir dan Bacillus.
Kehadiran BAL dalam proses fermentasi nira tidak diharapkan karena hasil
metabolisme BAL akan menyebabkan produk yang dihasilkan berasa asam
khususnya dalam pengolahan sari nira dan alkohol (Jay et al., 2005). Jenis bakteri
yang ditemukan pada nira antara lain genus Lactobacillus, Acetobacter, Sarcina,
Streptococcus, Leuconostoc, Bacillus, Eymomonsa, Brevibacterium, Micrococcus,
Serratia, Corynebacterium, Pediococcus dan Klebsiella (Frazier dan Westhoff,
1958).
Bakteri asam laktat yang teridentifikasi dalam penelitian Pinaria (2010)
(belum dipublikasikan), menghasilkan dua jenis bakteri asam laktat yakni
golongan Lactobacillus spp., Leuconostoc spp., dan Leu. mesenteriodes. Jenis
bakteri asam laktat yang teridentifikasi dalam penelitian ini mendukung penelitian
yang dilakukan Muharani (2011), yang berhasil mengisolasi potensi bakteri
probiotik dari nira aren yang memiliki total bakteri probiotik sebesar 45,35 x 1013
CFU/ml.

(1). Lactobacillus spp.
Lactobacillus spp. merupakan golongan bakteri asam laktat yang sering
dijumpai pada makanan fermentasi. Produk olahan, ikan, daging, susu dan buah-
buahan. Sejauh ini telah diketahui bahwa keberadaan bakteri ini tidak bersifat
patogen dan aman bagi kesehatan sehingga sering digunakan dalam industri
pengawetan makanan, minuman dan berpotensi sebagai probiotik. Sifat yang
mengguntungkan dari bakteri Lactobacillus sp. dalam bentuk probiotik adalah
dapat digunakan untuk mendukung peningkatan kesehatan khususnya kesehatan
pencernaan. Bakteri tersebut berperan sebagai flora normal dalam sistem
pencernaan. Fungsinya adalah untuk menjaga keseimbangan asam dan basa
sehingga pH dalam kolon konstan (Hadiningsih et al., 2006).
Lactobacillus heterofermentatif, menfermentasi gula dengan menghasilkan
asam asetat dan karbondioksida pada jalurnya. Golongan homofermentatif
menghasilkan alkohol dan asam laktat. Lactobacillus homofermentatif dapat
tumbuh pada suhu optimal 37oC atau di bawah temperatur tersebut antara lain L.
bulgaricus, L. helveticus, L. lactis, L. acidophilus, L. thermophilus dan L.
derburki. L. fermentum adalah salah satu contoh bakteri Lactobacillus
heterofermentatif yang dapat tumbuh pada temperatur yang tinggi. Golongan
bakteri homofermentatif yang dapat tumbuh pada suhu di bawah suhu optimal
adalah L. casei, L. plantarum, dan L. leichmannii, sedangkan heterofermentatif
yang dapat umbuh pada suhu rendah adalah L. brevis, L. buchneri, L. pastirianus.
Dari semua spesies yang adalah kecuali L. delbrueckii, L. leichmannii, L.

hilgardii, L. trichodes dan sebagai dari L. brevis dapat menfermenstasi laktosa
dengan menghasilkan asam laktat dan sebagian dari bakteri ini sangat penting bagi
industri susu (Frazier dan Westhoff, 1985).
Asam laktat yang dihasilkan dapat menyebabkan terjadinya penurunan pH
lingkungan. pH yang rendah dapat menghambat kontaminasi mikroba pembusuk
dan mikroba patogen. Penurunan pH disebabkan karena adanya asam-asam
organik yang dihasilkan oleh BAL (Nur, 2005). Bakteri asam laktat mempunyai
efek pengawet karena menghasilkan senyawa-senyawa yang mampu menghambat
pertumbuhan berbagai mikroba. Sebagian besar efek antimikroba ini disebabkan
oleh pembentukan asam laktat dan asam asetat serta penurunan pH yang
dihasilkan. Selain itu bakteri asam laktat juga menghasilkan senyawa-senyawa
penghambat lain seperti hidrogen peroksida, diasetil, karbondioksida, reuterin dan
bakteriosin (de Vuyst and Vandamme, 1994).
(2). Leuconostoc spp.
Leuconostoc adalah golongan bakteri asam laktat Gram positif cenderung
anaerob fakultatif yang membutuhkan faktor tumbuh kompleks meliputi asam
amino, peptida, karbohidrat, vitamin dan ion logam. Bakteri L. mesenteroides
merupakan bakteri asam laktat yang melakukan fermentasi glukosa menggunakan
jalur pentosa fosfat untuk memproduksi asam laktat, etanol dan CO2. Leuconostoc
dapat diisolasi pada beberapa produk fermentasi termasuk kefir, saeurkraut, dan
produk susu (Salminen et al., 2004).

Leuconostoc merupakan salah satu jenis BAL yang sangat penting dalam
produksi makanan, dimana bakteri ini (1) mampu memproduksi diasetil sehingga
mampu memberi bau terhadap produk makanan, (2) toleran terhadap konsentrasi
garam yang tinggi, contohnya produk saeukraut dan pikel. L. mesenteroides
muncul pada awal fermentasi laktat, (3) terdapat pada setiap produk fermentasi
sayuran dan kebanyakan memproduksi banyak asam, (4) toleran terhadap
konsentrasi gula yang tinggi 50-60% untuk L. mesenteroides (Frazier dan
Westhoff, 1958).
Golongan bakteri Leuconostoc dibedakan menurut jalur fermentasinya,
pada jalur heterofermentatif dapat menghasilkan asam laktat, asam asetat, etanol
dan CO2 dari fermentasi glukosa. Pada jalur ini fruktosa mengalami degradasi
membentuk manitol. Strain dari Leuconostoc dapat membentuk dekstran atau
lendir. Bentuk sel dari Leuconostoc hampir sama dengan Streptococcus berbentuk
kokus tunggal dan berpasangan membentuk rantai pendek (Pederson, 1971).
Terdapat lima spesies dari genus Leuconostoc yaitu Leu. mesenteroides,
Leu. paramesenteroides, Leu. lactis, Leu. carnosum, dan Leu. gelidium, Leu.
mesenteroides mempunyai tiga subspecies yaitu Leu. mesenteroides subsp.
mesenteroides, Leu. mesenteroides subsp. dextranicum, dan Leu. mesenteroides
subsp. cremoris (Ray, 2004). Bakteri yang termasuk genus ini banyak dijumpai
pada permukaan tanaman, daging dan olahannya, produk susu seperti es krim,
mentega dan sirup. Genus Leuconostoc berperan dalam beberapa produk

fermentasi sayuran seperti acar dan sauerkraut. Leu. mesenteroides mempunyai
toleransi terhadap kadar gula yang tinggi (55-60%) (Frazier and Westhoff, 1985).
2.2 Peranan Bakteri Asam Laktat dalam Produksi Pangan
Peranan utama bakteri asam laktat dalam bahan pangan adalah sebagai
kultur starter prduk-produk yang melibatkan proses fermentasi untuk memperoleh
produk akhir dengan tingkat konsentrasi yang tinggi. Selain menghasilkan produk
akhir yang konsisten, bakteri asam laktat ternyata memiliki efek mengawetkan
pada produk fermentasi yang diinginkan. Peran bakteri asam laktat yang banyak
dikenal adalah sebagai pengawet alami atau biopreservatif, yaitu zat yang
digunakan pada pengawetan secara hayati untuk mencegah adanya mikroba
perusak dan pathogen. Pemakaiannya pada bahan pangan bertujuan untuk
mencegah dan mengurangi pertumbuhan mikroba pembusuk (Wirawati, 2002).
Bakteri asam laktat diklasifikasikan menjadi empat kelas yaitu genus
Lactobacillus, Leuconostoc, Streptococcus, dan Pediococcus. Klasifikasi tersebut
lebih didasarkan pada ciri morfologi, tipe fermentasi, kemampuan untuk tumbuh
pada suhu yang berbeda, sifat stereospesifik (D atau L laktat), serta toleransi
terhadap asam dan basa. Klasifikasi bakteri asam laktat berkembang sehingga
genus Lactobacillus menjadi Lactobacillus dan Carnobacterium. Sedangkan
genus Streptococcus menjadi empat yaitu Streptococcus, Lactococcus,
Vagococcus, dan Enterococcus. Genus Pediococcus menjadi Pedicoccus,

Tetragenococcus dan Aerococcus, genus Leuconostoc tetap sama. Klasifikasi yang
baru tersebut dihasilkan dengan mempertimbangkan komposisi asam lemak pada
membran sel, motilitas dan urutan rRNA, serta persen guanin dan sitosin pada
DNA (Kusumawati, 2000).
Bakteri asam laktat diisolasi dari produk olahan susu, seperti: produk susu
fermentasi Scandinavia, beberapa jenis yoghurt, susu fermentasi, keju, daging dan
sayur hasil fermentasi, yang berfungsi sebagai sumber bakteri asam laktat yang
memproduksi EPS. Eksopolisakarida dapat digunakan sebagai pengemulsi,
pembentuk gel dan pengikat air pada industri makanan, dan juga digunakan
sebagai prebiotik. Selain itu juga bermanfaat dibidang kesehatan untuk antitumor.
Penggunaan EPS untuk aplikasi medis juga telah dilaporkan. Inulin dapat
mengurangi Colitis (inflamasi di perut) tikus percobaan. Fruktan (FOS dan inulin)
dapat berfungsi sebagai penyedap rasa makanan (van Hijum et al., 2002).
Bakteri asam laktat mempunyai peranan esensial hampir dalam semua
proses fermentasi makanan dan minuman. Peran utama bakteri ini dalam industri
makanan adalah untuk pengasaman bahan mentah dengan memproduksi sebagian
besar asam laktat (homofermentatif) atau asam laktat, asam asetat, etanol
(heterofermentatif) dan digunakan dalam produk susu seperti yogurt, sour cream
(susu asam), keju, mentega dan produksi asam-asaman seperti asinan (Nur, 2005).
Golongan bakteri asam laktat homofermentatif dan heterofermentatif memiliki
peran dan fugsinya masing-masing (Fox et al., 2000) ;

a) Homofermentatif memiliki aldolase, sehingga bakteri ini tidak dapat
menfermentasi pentosa dan hanya dapat memfermentasi heksosa melalui
jalur glikolisis, yang hanya menghasilkan asam laktat dari metabolisme
gula. Golongan yang termasuk adalah Pediococcus, Streptococcus, dan
Leuconostoc, Lactobacillus (Frazier and Westhoff, 1985).
b) Heterofermentatif memiliki fosfoketolase sehingga mampu
memfermentasi gula secara heterofermentatif menghasilkan asam laktat,
etanol dan CO2. Bakteri yang termasuk golongan ini adalah Leuconostoc,
Lactobacillus dan Carnobacterium (Frazier and Westhoff, 1985).
2.3 Eksopolisakarida (EPS)
Eksopolisakarida adalah suatu polisakarida yang diproduksi dan
dieksresikan oleh mikroba. EPS dihasilkan oleh beberapa strain dari spesies
bakteri asam laktat, diantaranya jenis homopolisakarida dihasilkan oleh
Leuconostoc mesenteroides, sedangkan jenis heteropolisakarida dihasilkan oleh
Streptococcus thermophilus OR 901, Lactobacillus bulgaricus CNRZ 1187 (de
Vuyst et al., 1998). Strain-strain tersebut belum digunakan secara ekstensif dalam
produksi komersial karena produksi EPSnya dalam jumlah sedikit (kurang dari
500 mg/ liter) dan biosintesisnya sangat tidak stabil (de Vuyst and Degeest, 1999).
Malik et al., (2008) menunjukkan bahwa beberapa EPS yang diproduksi
oleh bakteri asam laktat telah digunakan dalam bidang kesehatan dan produksi

makanan seperti pada Tabel 2.4. Karakteristik EPS adalah berbentuk seperti kapur
berongga yang melekat dan sulit dikeluarkan. Bakteri asam laktat yang
menghasilkan lendir telah banyak digunakan dalam industri susu. Bakteri ini telah
digunakan di Finland sejak abad kesembilan belas, khususnya dalam produksi
produk susu fermentasi kental. Jenis bakteri yang menghasilkan EPS secara luas
digunakan untuk meningkatkan kualitas reologi yoghurt untuk, menghambat
sineresis, serta mampu mengganti starter yang lain yang biasanya digunakan
dalam pengolahan yoghurt (Salminen et al., 2004).
Tabel 2.4
Kegunaan beberapa jenis EPS dalam industi pangan dan farmasi
Jenis EPS Struktur EPS Kegunaan SumberDekstran α 1,6 dan
α 1,3 glikosidikAnti inflamasi, anti
trombotik, anti koagulan,Drug delivery system
Veronese danCeliceti, 2006 ;
Dinoto et al.,2011Kefiran D-glukosa/
D-galaktosaAnti bakteri, anti jamur,
anti kankerVu et al.,2009
Gellan D-glukoronat/ L-ramnosa
Drug delivery system Vu et al.,2009
Curdlan β – 1,3 glikosidik Drug delivery system Rehm, 2009 ;Gunadi et al.,2005
Xanthan α – D- manosa Pengemulsi dan bidangkosmetik, makanan dan
farmasi
Sutherland, 1998
Sumber : Malik et al., (2008)
2.3.1 Bakteri asam laktat penghasil eksopolisakarida

Sebagian besar BAL yang menghasilkan EPS termasuk genus
Streptococcus, Lactobacillus, Lactococcus, Leuconostoc, dan Pediococcus (Tabel
2.5) (Ruas-Madiedo and Reyes-Gavilan, 2006). Berdasarkan komposisinya,
eksopolisakarida dapat dibedakan menjadi dua jenis berdasarkan komposisi
kimianya, yaitu ;
(1). Homopolisakarida (HoPSs)
Eksopolisakarida jenis homopolisakarida ini tersusun atas satu jenis gula
monosakarida saja (utamanya glukosa dan fruktosa) (Tallon et al., 2004).
Homopolisakarida juga dapat menghasilkann dekstran, mutan, alternan, reuteran,
pullulan,levan, inulin, curdlan dan lain-lain (Patel et al., 2011).
Tabel 2.5
Total EPS dari beberapa jenis BAL pada beberapa media
dan tipe pertumbuhan.
Mikroorganisme Media Temp(0C)
Time(jam)
pH Yield(mg/l)
L. rahmnosus 9595 M BMM 32-37 72 6 1000L. delb. bulgaricus RR Whey 38 24-28 5 95-110L. rhamnosus R BMM 37 72 6 500L. delb. bulgaricus Milk 42 24 6 110L. delb. bulgaricus MRS 40 18 7 263L. rhamnosus GG Milk 37 20 7 83L. delb. bulgaricus 291 Skimmed Milk 37 22 7 80L. casei CG11 BMM 25 48 7 130L. helventicus Skimmed milk 37 60 5 730L. delb.bulgaricus Whey
(proteinfree)37 18 6 800
L.rhamnosus 9595 Whey permeate 37 24 6 2775

supplementedL. paracasei BMM 32-37 72 6 80Sumber : Badel et. al. (2011)
Homopolisakarida yang terdiri dari glukosa adalah α-glukan (dekstran,
mutan dan alteran) dan β-glukan, sedangkan homopolisakarida yang terdiri dari
fruktosa adalah fruktan (levan dan inulin). Contoh bakteri yang menghasilkan
homopolisakarida dekstran dari L. mesenteroides, selulosa dari Acetobacter,
Achromo, Achromobacter, Agobacterium, Alcaligenes, Pseudomonas, Rhizobium,
Sarcibna dan Zooglea seperti pada Tabel 2.6. HoPSs disekresikan oleh
Lactobacillus sp. mengandung glukosa atau fruktosa sebagai satu-satunya
monosakarida dan diklasifikasikan sebagai glukan dan fruktan seperti pada Tabel
2.7 .
Tabel 2.6 Homopolisakarida yang dihasilkan oleh BAL berdasarkan tipe polisakarida
Tipe Spesis Ikatanα-D-glukan
- Dextran
- Mutan
- Alteran
L. mesenteroides subsp.mesenteroidesL. mesenteroides subsp.dextranicumStreptococcus mutansStreptococcus sobrinusLeuconostocmesenteroides
α-D-Glcpb (16)
α-D-Glcpb (13)
α-D-Glcpb (13)/ (16)
β-D-glukan Pediococcus spp.Streptococcus spp.
β-D-Glcpb (13)
Fruktan- Levan- Innulin –like
Streptococcus salivariusStreptococcus mutans
β-D-Frup (2 6)
Sumber : Ruas-Madiedo and Reyes-Gavilan, (2005)

Tabel 2.7 Lactobacillus sp. golongan homopolisakarida berdasarkan tipe polisakarida
Spesis Monosakarida Ikatan TipePolisakarida
L. hilgardii D-Glcp α-(1,6) DekstranL. fermentum D-Glcp α-(1,6) DekstranL. reuteri 180 D-Gcp α-(1,6) DekstranL.reuteri ML1 D-Glcp α-(1,3) MutanL. reuteri 121 A D-Glcp α-(1,4) Reuteran L.reuteri 121 D-Fruf β-(2,6) LevanL.reuteri LB 121 D-Fruf β-(2,1) InulinL. sakei Kg15 D-Glcp α-(1,6) DekstranL.parabuchneri 33 D-Glcp α-(1,6) DekstranSumber : Badel et al. (2010).
(2). Heteropolisakarida (HePSs)
Heteropolisakarida terdiri dari 3-7 jenis gula monosakarida, paling sedikit
2 monosakarida. Selain itu dapat juga ditemukan gugus asetil, fosfat, n-gliserol-
phospat dan N-asetil-aminosugars. Sedangkan menurut Harrah et al., (2006), unit
penyusun heteropolisakarida bervariasi antara 3-8 sakarida dan seringkali
mengandung kombinasi α-glukosa, α-galaktosa, L-ramnosa, dan kadang terdapat
N-asetilglukosamin, N-asetilgalaktosamin, asam glukoronik dan subsituten non-
karbohidrat seperti fosfat, asetil dan gliserol. Contoh heteropolisakarida yaitu
Xanthan dari Xanthomonas campestris sebagai pengental, penguat tekstur, dan
pengembang. HePSs dari Lactobacillus spp. (L. delbrueckii bulgaricus, L.
rhamnosus dan L. helveticus) memiliki unit berulang terdiri dari tujuh
monosakarida, dimana glukosa, galaktosa dan rhamnose adalah gula utama seperti
pada Tabel 2.8.
Tabel 2.8

Lactobacillus sp. golongan Heteropolisakarida
berdasarkan tipe polisakarida penyusunnya
Strain PrincipalMonosaccharides
Subtituent and/ or secondarymonosaccharides
L. acidophilus LMG 9433 Glc, Gal GlcNAc + GluAL.sakei 0-1 Glc, RhaL.delb. subsp, bulgaricus 291 Glc, GalLY 03, 24, 25 Glc, Gal RhaNCFB2074 Glc, GalL.belveticus ATCC 15807 Glc, Gal, (Rha) Phosphoric acidL. paracasei 34-1 Gal GalINAc, glycerol 3-PL.pentosus LPS 26 Glc, Rha
Glc ManL. plantarum EP56 Glc, Gal N-GlcNAc
Glc, Gal, Rha Glycerol, phospheglycerolL. rhamnosus C83 Glc, GalL. rhamnosus GG Gal, Rha GlcNAcL. rhamnosus RW-9595 M orR
Glc, Gal, Rha
L. casei CG11 Glc, RhaL. kefiranofactens subsp.kefiranofaciens
Glc, Gal
Sumber : Badel et al. (2010).
Kefir adalah contoh heteropolisakarida yang disintesis dari Lactobacillus
kefiri dan Lactobacillus kefiranofaciens yang ditemukan dalam susu fermentasi,
kefir bersifat larut dalam air (Maeda et al., 2004). Struktur polimer kefiran
dibentuk dari monomer D-glukosa atau heteropolisakarida, D-galaktosa yang
mengalami percabangan pada dua unit rantai serta delapan unit rantai. Polimer ini
menunjukkan aktivitas anti bakteri, anti jamur, dan anti kanker (Vu et al., 2009).
2.3.2 Metabolisme eksopolisakarida

Polisakarida yang terdapat pada permukaan sel bakteri secara umum
terdapat dalam dua bentuk yaitu lipopolisakarida dan eksopolisakarida (Weiner et
al. 1995). Lipopolisakarida mengikat permukaan sel melalui penempelan dengan
lipid, sementara eksopolisakarida (EPS) berasosiasi dengan permukaan sel
sebagai sebuah kapsul. EPS memiliki heterogenisitas dalam strukturnya, mulai
dari struktur sederhana, ikatan α 1-4 berupa rantai glukosa tidak bercabang yang
disebut dekstran sampai dengan struktur yang kompleks dengan ikatan bercabang
atau substitusi heteropolisakarida yang tersusun dari sub unit berulang
oligosakarida yang disebut dengan xantan dan asam kolanik. EPS dapat juga
disubstitusi, umumnya ikatan ester dan N dengan piruvat, asetat, sulfat, fosfat dan
gugus lainnya (Weiner et al. 1995).
Eksopolisakarida disintesa dalam fase-fase pertumbuhan yang berbeda
dengan kondisi yang bervariasi tergantung dari jenis mikroorganismenya. Proses
sintesa dapat dibagi menjadi dua prinsip dasar yaitu tempat sintesa dan prekursor
alami misalnya sintesa di luar dinding sel atau pada membran sel. Sintesa
heteropolisakarida berbeda dengan sintesa homopolisakarida yang disintesa pada
membran sitoplasma dengan memanfaatkan prekursor yang terbentuk intraselular.
Gula nukleotida berperan penting dalam sintesa heteropolisakarida sehingga
peranannya dalam interkonvensi monosakarida atau disakrida (gula) sebaik
aktivasi gula yang dibutuhkan untuk polimerisasi monosakarida menjadi
polisakrida (Cerning, 1990).

Biosintesis EPS menjelaskan tentang konversi gula (glikolisis) seperti
polisakarida yang umumnya digunakan untuk dinding sel. EPS yang dihasilkan
oleh bakteri menggunakan dua jalur sintesis sesuai dengan lokasinya yakni
ektraseluler dan intraseluler. Pada homopolisakarida (Gambar 2.2) EPS disintesis
di dalam sel oleh glycansucrase ektraseluler atau glycosiltransferase (Gtf)
(Coutinho and Henrissat, 1999). Glycosiltransferase (GTF) adalah enzim yang
tergabung dalam kelompok keluarga glikosida hidrolase, dan merupakan
kelompok enzim sukrase yang disebut sebagai glukansukrase, dan fruktansukrase.
Enzim-enzim tersebut diketahui terlibat dalam sintesis eksopolisakarida dari
mikroba (Malik et al., 2008).
Biosintesis heteropolisakarida (Gambar 2.3) mekanisme pembentukannya
lebih kompleks dari pada biosintesis homopolisakarida. EPS disintesis oleh
bakteri secara intraseluler di dalam sitoplasma. Gula dari medium dimasukkan
kedalam sitoplasma dan diaktifkan melalui fosforilasi selama penyimpanan di
dalam sel oleh Phosphoenolpyruvate - Phosphotranferase (PEP-PTS). Biosintesis
EPS membutuhkan energi di dalam prosesnya. Pengulangan polimerasi sangat
kompleks dan melibatkan beberapa enzim dan protein secara berurutan. Langkah
terakhir dari biosintesis EPS adalah pemindahan polisakrida ke luar sel dan
disekresikan ke lingkungan (EPS slime) atau penempelan pada sel (EPS capsular).
Mekanisme polimerisasi dari unit pengulangan dalam BAL dan pengeluaran
subsekuen masih belum jelas (de Vuyst and Degeest, 1999).

Ket : katalis dominan, N-batas signal, Glukosa-residu, Fruktosa-residu
Gambar 2.2
Jalur Biosintesis EPS Homopolisakarida
Pertumbuhan sel bakteri berbanding terbalik dengan produksi EPS.
Mekanisme pembentukan EPS yang tinggi diperoleh pada saat laju pertumbuhan
sel yang terjadi dengan kecepatan rendah, sel-sel bakteri akan lebih banyak
menghasilkan Isoprenoid glycosyl lipid carriers yang berperan sebagai prekursor
untuk pembentukan dinding sel dan produksi EPS. Isomer isopreonid glycosyl
lipid carriers berada di dalam membran sel yang berfungsi sebagai penerima

residu molekul gula. Semakin banyak prekursor yang dihasilkan maka semakin
tinggi prduksi EPS. Sebaliknya pada laju pertumbuhan yang cepat prekursor ini
akan lebih banyak dimanfaatkan dalam pembentukan dinding sel (Sutherland,
1990).
Phosphoenolpyruvate - Phosphotranferase antibodi
galaktosa
Laktosa 6P laktosa
Galaktosa 6P Glukosa
Tagatosa 6-P Glukosa 6-P
Fruktosa 6P
Glukosa 1 P
Tagatosa 1,6 dIP

Fruktosa 1,6 dIP dTDP-glukosa
Glikolisis UDP-Glukosa
dTDP-4 keto-6-deoksimanosa
ATP UDP-galaktosa
dTDP-ramnosa
Biomasa
Unit Berulang
EPS
Gambar 2.3 Jalur Biosintesis EPS Heteropolisakarida
Keterangan : jalur yang terlibat dalam katabolisme laktosa (kiri dan kanan atas) dan Biosintesiseksopolisakrida (kanan bawah) dalam fermentasi glukosa dan fruktosa Lactococus lactis (glukosamengirim melalui glukosa fosfotransferase pada sistem tranportasi primer). Angka menunjukkanenzim yang terlibat 1). Glukokinase, 2). Fosfoglukametase 3). UDP-glukosa fosforilase, 4). UDP-galaktosa-4-epimerase, 5). dTDP-glukosa firofosforelase, 6). Dehidratase, 7). Reduktaseepimerase, 8). Glikosil tranferase, 9). 1-fosfofruktokinase, 10). Isomerase fosfoglukosa, 11). 6-fosfofruktokinase, 12). Fruktosa-1,6 bisfosfatase, 13). Aldolase fruktosa 1.6-difos, de Vuyst danDegeest, (1999).

2.3.3 Faktor-faktor yang mempengaruhi produksi eksopolisakarida
Beberapa faktor yang berpengaruh terhadap produksi EPS bakteri asam
laktat adalah :
(1). Suhu dan waktu inkubasi.
Beberapa peneliti menunjukkan bahwa setiap bakteri starter kultur yang
berbeda memiliki kemampuan memproduksi EPS yang berbeda pada suhu dan
waktu inkubasi yang tertentu dan pada umumnya bukan pada suhu pertumbuhan
optimumnya. Sebagai contoh Lactobacillus casei yang merupakan starter kultur
yang digunakan dalam pembuatan yakult mampu memproduksi EPS sebesar 121
mg/l pada suhu inkubasi 30oC dan waktu inkubasi 24 jam, bila waktu inkubasi
diperpanjang sampai 72 jam produksi akan menurun pada suhu 30oC dan
37oC. Hasil ini menunjukkan bahwa efisiensi sel adalah terbaik dalam
mengkonversi karbohidrat menjadi polimer pada kondisi kultur tersebut (Mozzi et
al., 1996).
Sebelumnya Cerning (1990) dengan menggunakan kultur yogurt
Streptococcus thermophilus, L. delbrueckii sub sp. bulgaricus, starter keju
Lactococcus lactis ssp. cremoris menyatakan bahwa produksi EPS terbaik pada
suhu di bawah suhu pertumbuhan optimumnya. Sementara Malaka et al. (2004)
berdasarkan hasil penelitiannya menemukan bahwa Lb.delbrueckii ssp.

bulgaricus yaitu starter yogurt menghasilkan EPS optimal sebesar 359,2 mg/l
pada medium SSR 10% dengan suhu inkubasi 30oC dan waktu inkubasi selama 16
jam (Malaka, 2012).
(2). pH medium pertumbuhan
pH optimum pertumbuhan bagi kebanyakan bakteri terletak antara 6,5 dan
7,5. Namun beberapa spesies dapat tumbuh dalam keadaan sangat masam atau
sangat alkalin. Bagi kebanyakan spesies, dengan nilai pH minimum untuk tetap
hidup yaitu 4 dan nilai pH maksimum ialah 9 (Pelczar and Chan, 2005). Mozzi et
al. (1996) menemukan bahwa Lactobacillus casei memproduksi EPS lebih baik
pada pH awal 4,0 dari pada pH 5,0; 5,5 dan 6,5. Sementara Lb. delbrueckii sub
sp. bulgaricus memproduksi EPS optimal pada pH 6,5 dengan menghasilkan EPS
sebesar 326,2 mg/l.
(3). Sumber Karbon dalam médium pertumbuhan
BAL membutuhkan nutrisi kompleks seperti asam amino, peptida, derivat
asam nukleat, vitamin, garam, asam lemak, serta unsur pertumbuhandasar bakteri
seperti karbon, nitrogen, oksigen, sulfur, fosfor, magnesium, zat besi, dan
sejumlah kecil logam lainnya. Sumber karbon terbaik menurut Mozzi et al. (1995)
adalah galaktosa (56 mg/l) dibanding sumber karbon lainnya fruktosa, sukrosa
dan laktosa. Chu et al. (2001) melaporkan bahwa Bifidobacterium longum BB-
79 mampu memanfaatkan laktosa untuk memproduksi EPS. Suatu fermentasi
dilakukan secara anaerobik dengan konsentrasi laktosa mula-mula antara 2-
5%. Temperatur dan pH pada bioreaktor dijaga yaitu berturut-turut 37oC dan

6,9. Produksi EPS tertinggi dengan kecepatan 0,24 g/jam dicapai dengan
konsentrasi laktosa di bawah 5% dan konsentrasi EPS akhir adalah 1,45 g/l.
Marshall et al. (1995) melaporkan bahwa Lactococcus lactis ssp.cremoris sebagai
starter kultur untuk pembuatan keju memproduksi 2 tipe EPS dalam medium yang
yang mengandung glukosa, laktosa dan galaktosa. Total EPS yang diproduksi
adalah 25 μg/ml.
(4). Sumber mikromineral
Mineral dibutuhkan bakteri sebagai akseptor elektron dalam metabolisme
gula. Dalam reaksi polimerisasi EPS pembentukan rantai karbon membutuhkan
mineral sebagai akseptor elektron yang mengikat antara monomer satu dengan
monomer lainnya. Menurut Mozzi et al. (1995) garam mineral sangat
mempengaruhi produksi EPS dari L. Casei pada kondisi inkubasi 37oC selama 48
jam. Percobaan itu memberi hasil bahwa semua jenis garam
meningkatkan produksi EPS dengan hasil terbanyak pada suplementasi dengan
MgSO4 yakni menghasilkan EPS sebesar 107 mg/l. Hal ini disebabkan karena
Mg2+ mempunyai efek stimulator untuk sintesa polimer.
2.4 Optimalisasi Produksi EPS menggunakan Respons Surface Methode (RSM)
Metode permukaan respon Response Surface Method (RSM) merupakan
sekumpulan teknik matematika dan statistika yang berguna untuk menganalisis
permasalahan dimana beberapa variabel independen mempengaruhi variabel
respon dan tujuan akhirnya adalah untuk mengoptimalkan respon. Metode ini

pertama kali diajukan sejak tahun 1951 dan sampai saat ini telah banyak
dimanfaatkan baik dalam dunia penelitian maupun aplikasi industri (Sudjana,
1994). Metode permukaan respon memiliki peran penting dalam merancang,
merumuskan dan menganalisa suatu kajian ilmiah dan sering diaplikasikan dalam
bidang industri dan ilmu klinis, ilmu sosial dan ilmu pangan.
Menurut Montgomery (2009) RSM bertujuan untuk mengoptimalkan
respon. Ide dasar metode ini adalah memanfaatkan desain eksperimen statistika
untuk mencari nilai optimal dari suatu respon. Proses optimasi seringkali
dilakukan di dunia industri sebagai bentuk upaya meningkatkan mutu dan kualitas
produk yang dihasilkan. Selain digunakan dalam proses optimasi produk, RSM
juga digunakan dalam bidang yang lain seperti bidang ilmu pangan, biologi, ilmu
kedokteran dan kesehatan (Myers et al., 1971).
Menurut Gaspersz (1995) RSM dapat digunakan oleh peneliti untuk: (1)
mencari suatu fungsi pendekatan yang cocok untuk meramalkan respon yang akan
datang, serta (2) menentukan nilai-nilai dari variabel bebas yang
mengoptimumkan respons yang dipelajari. Dalam metode permukaan respon,
variabel-variabel bebas akan didefinisikan sebagai X1, X2, …, Xk, dimana variabel
bebas ini diasumsikan merupakan variabel kontinyu dan dapat dikendalikan oleh
peneliti tanpa kesalahan, sedangkan respon yang didefinisikan sebagai variabel
terikat (Y) diasumsikan merupakan variabel acak (random variable).
Pada dasarnya analisis RSM serupa dengan analisis regresi, yaitu
menggunakan prosedur pendugaan parameter fungsi respon berdasarkan metode
kuadrat terkecil, hanya saja dalam RSM diperluas dengan teknik-teknik

matematik untuk menentukan titik-titik optimum agar dapat ditemukan respon
yang optimum (maksimum atau minimum) (Gaspersz, 1995). RSM adalah desain
dan model yang bekerja dengan berbagai treatment secara terus menerus ketika
menemukan nilai optimum atau menggambarkan respon sesuai tujuan
(Hinkelmann and Kempthorne, 2005; Box and Draper, 2007). Tujuan utama dari
RSM adalah untuk menemukan respon optimal. Bila ada lebih dari satu respon
maka penting untuk menemukan optimum kompromi yang tidak mengoptimalkan
hanya saja satu respon (Box dan Draper, 2007). Bila ada kendala pada data desain,
maka desain eksperimental harus memenuhi persyaratan kendala. Tujuan kedua
adalah memahami bagaimana resepon perubahan dalam arah tertentu dengan
menyesuaikan variabel desain. Secara umum, permukaan respon dapat
divisualisasikan dalam bentuk grafis. Grafik sangat membantu untuk melihat
bentuk permukaan respon; bukit, lembah, dan garis bridge (Myers et al., 1989).
Keuntungan utama dari penggunaan metode RSM adalah mengurangi jumlah
percobaan yang harus dilakukan untuk mengevaluasi kondisi terbaik.
Tahapan yang perlu dilakukan untuk memulai pelaksanaan response
surface methode adalah sebagi berikut (Hanafiah, 2005) :
1. Menentukan model persamaan orde pertama dimana suatu desain eksperimen
dilakukan untuk pengumpulan dalam dan arah penelitian selanjutnya
ditentukan dengan menggunakan steepest descent.
2. Setelah arah penelitian diketahui, selanjutnya penentuan level faktor untuk
pengumpulan data selanjutnya.

3. Menentukan model persamaan orde kedua. Penentuan model dilakukan
dengan menentukan desain eksperimen dengan level yang telah ditetapkan
setelah metode steepest descent dilakukan.
4. Menentukan titik optimum dari faktor-faktor yang diteliti.