ekologi, relung pakan, dan strategi adaptasi kelelawar...
TRANSCRIPT

EKOLOGI, RELUNG PAKAN, DAN STRATEGI ADAPTASI
KELELAWAR PENGHUNI GUA DI KARST GOMBONG
KEBUMEN JAWA TENGAH
FAHMA WIJAYANTI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2011

PERNYATAAN MENGENAI DISERTASI DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi : Ekologi, Relung Pakan, dan
Strategi Adaptasi Kelelawar Penghuni Gua di Karst Gombong Kebumen, Jawa
Tengah adalah karya saya dengan arahan dari komisi pembimbing dan belum
diajukan dalam bentuk apa pun kepada perguruan tinggi manapun. Sumber
informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak
diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam
Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Mei 2011
Fahma Wijayanti
NRP: G361060011

EKOLOGI, RELUNG PAKAN, DAN STRATEGI ADAPTASI
KELELAWAR PENGHUNI GUA DI KARST GOMBONG
KEBUMEN JAWA TENGAH
FAHMA WIJAYANTI
Disertasi
sebagai salah satu syarat untuk memperoleh gelar
Doktor pada
Program Studi Biologi Mayor Biosains Hewan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2011

Penguji pada Ujian Tertutp: Prof. Dr. Ani Mardistuti, MSc
(Departemen KSHE, Fakultas Kehutanan IPB)
: Prof. Wasmen Manalu, PhD
(Departemen Anatomi, Fisiologi, dan
Farmakologi FKH, IPB).
Penguji pada Ujian Terbuka: Dr. Henry Bastaman MES.
(Deputi Kemeterian Negara Lingkungan Hidup
Bidang Pembinaan Sarana Teknik Lingkungan
Dan Peningkatan Kapasitas)
: Dr. Ir. Novianto Bambang Wawandono, MSi.
Direktur Konservasi Dan Keanekaragaman Hayati
Kementerian Kehutanan RI)

ABSTRACT
The existence of cave dwelling bats of karst area need to be conserved, because bats have important roles for the ecosystem inside the cave as well as outside the cave. The objectives of this research were to know the biodiversity of cave dwelling bats, to identify physical factors which influenced the preference roosting place, to determine the prey preference of cave dwelling bats, to observe structural and physiological adaptation of the cave dwelling bats and to propose the karst management strategy based on the conservation status of bats. This study was conducted from September 2008 to July 2010 in twelve caves in Gombong karst area, Central Java. The sample of the bats were picked up at the roosting place during the day. The physical and microclimate parameters were measured under the bat roosts, three times in February, June and October 2009. Stomach gut content was collected and dissolved in aquadest. The material of insect were sorted and identified under microscope and compare to the insects that were collected by light trap in bat foraging area. The polen were collected from the intestine of fruit bats. Blood was taken from the interfemoral vein of bats. Lungs removed from body and were made preparations for histology. The diameter of alveoli was observed under the microscope. Then the amount of erythrocyte was counted by using hemocytometer and hemoglobin content was measured using Sahli’s method. The data was analyzed by ANOVA, Principle Component Analysis (PCA), Redundancy Analysis (RDA), Canonical Correspondence Analysis (CCA), Hybrid Canonical Correspondence Analysis (HCCA) and multiple regression. The result showed: 1) Fifteen spesies (eleven spesies of Microchiroptera and four spesies of Megachiroptera) indicated known in this research. 2) The microclimate factors which influenced the preference roosting place were the sound intensity, the distance from the cave mouth, the temperature, the humidity and the light intensity. The insects in gut content of insectivorous bats belong to 10 orders, distributed into 29 families. Otherwise pollen in gut content of frugivorous bats belong to 9 families, distributed into 33 spesies of plant. The niche overlap index between spesies of bats that occupy in one cave was less than 30%. The diameter of alveoli significantly correlated with humidity, temperature and ammonia levels. The amount of erythrocyte increased by the increase of humidity, decrease in the temperature and the increase of ammonia level. This tendency also revealed in hemoglobin change. Gombong karst area proposed as a karst region class I based on Kepmen ESDM No. 1456 K/20/MEM/2000
Key words: bats. Karst Gombong, roost preference, diet, adaptation, conservation.


RINGKASAN
FAHMA WIJAYANTI. Ekologi, Relung Pakan, dan Strategi Adaptasi Kelelawar Penghuni Gua di Karst Gombong Kebumen Jawa Tegah. Dibimbing oleh DEDY DURYADI SOLIHIN, HADI SUKADI ALIKODRA, dan IBNU MARYANTO
Persoalan yang timbul akibat pemanfaatan ekosistem karst dapat menyebabkan ekosistem karst tidak lagi memberikan manfaat ekonomi dan fungsi ekologi. Untuk menghindari hal tersebut, diperlukan upaya konservasi ekosistem karst. Hal ini hanya dapat dipenuhi jika pengetahuan yang mendasari kestabilan ekosistem karst ini dapat dipahami dengan baik. Penelitian ini menjawab beberapa aspek yang berkaitan dengan kelelawar penghuni gua di Karst Gombong. Aspek-aspek tersebut meliputi: biodiversitas dan struktur komunitas kelelawar gua; pola pemilihan sarang; relung pakan; serta adaptasi struktural dan fisiologi organ pernapasan kelelawar gua. Dari hasil penelitian tersebut dirancang strategi konservasi ekosistem gua, sehingga keberadaan kelelawar sebagai kunci penyedia energi ekosistem (key factor in cycle energy) dalam gua dan pemegang peran ekologis lainnya bagi ekosistem luar gua dapat dipertahankan.
Hasil penelusuran gua menunjukkan dari dua belas gua yang diteliti, sepuluh gua dihuni kelelawar, dua gua tidak dihuni kelelawar. Jenis-jenis kelelawar yang bersarang pada gua-gua tersebut terdiri atas empat jenis kelelawar Megachiroptera dan sebelas jenis kelelawar Microchiroptera. Hasil Redundancy Analysis (RDA) menunjukkan semakin panjang, tinggi, dan lebar lorong gua, semakin tinggi kelimpahan, indeks keanekaragaman jenis, dan kemerataan jenis kelelawar. Sebaliknya, jumlah pintu dan ventilasi gua tidak berkorelasi nyata. Gua dengan lorong sempit hanya dapat dihuni oleh jenis tertentu saja, yaitu jenis yang mampu malakukan manuver dengan baik. Sebaliknya, gua dengan lorong lebar, dapat dihuni kelelawar dengan kemampuan lebih beragam. Hal ini didukung oleh hasil penelitian Sevcik (2003) pada Plecotus auritus dan P. austriacus yang membuktian P. auritus lebih unggul mengeksploitasi habitat, karena lebih mampu melakukan manuver terbang. Semakin panjang, tinggi dan lebar lorong gua, semakin tinggi kelimpahan dan keanekaragaman jenis kelelawar yang dapat bersarang didalamnya. Hal ini sesuai dengan pendapat Maguran (1988) bahwa semakin luas habitat, semakin banyak jumlah dan jenis biota yang dapat hidup di dalamnya. Penelitian Schnitzler et al. (2003) membuktikan ketika terbang menuju lokasi sarang dan tempat pencarian makan, kelelawar cenderung menggunakan jalur yang sama. Hal ini menyebabkan jumlah pintu dan ventilasi gua tidak berpengaruh pada struktur komunitas kelelawar.
Hasil Canonical Component Analysis (CCA) menunjukkan lima parameter yang berpengaruh pada pemilihan sarang kelelawar berturut-turut dari yang paling kuat sampai yang paling lemah pengaruhnya adalah: intensitas suara, jarak dari pintu gua, suhu, kelembapan, dan intensitas cahaya. Berikut adalah beberapa alasan mengapa faktor faktor tersebut berpengaruh. 1) Hasil penelitian Schnitzler et al. (2003) membuktikan bahwa di ruang tertutup kelelawar lebih sulit menganalisis gelombang pantul (ekolokasi). Oleh kerena itu, di ruang tertutup seperti di dalam gua, gangguan suara sedikit saja akan menyebabkan kelelawar gagal menganalisis gelombang pantul. 2) Kelelawar yang mampu terbang dengan

manuver yang baik cenderung memilih lokasi sarang pada jarak yang jauh dari lorong gua karena lebih aman dari gangguan manusia. Sebaliknya kelelawar yang tidak mampu melewati lorong gua yang panjang memilih sarang di lokasi yang dekat dari pintu gua untuk memudahkan kelelawar tersebut keluar atau masuk ke dalam sarang. 3) Kelelawar merupakan homoikiloterm (suhu tubuh konstan) yang mempunyai batas toleransi sempit pada suhu lingkungan. Batas toleransi tersebut berbeda antara satu jenis kelelawar dan jenis lainnya, sehingga kelelawar memilih sarang yang sesuai dengan batas toleransi tubuhnya. 4) Membran petagium (sayap) kelelawar tersusun atas lapisan kulit tipis yang sangat peka pada kekeringan. Hal ini menyebabkan kelelawar yang mempunyai membran petagium tipis memilih lokasi sarang yang lembap, sedangkan yang memiliki membran petagium tebal mampu bersarang di lokasi gua yang cenderung kering. 5) Megachiroptera cenderung menggunakan penglihatannya untuk berorientasi pada ruang. Oleh karena itu jenis-jenis kelelawar Megachiroptera cenderung memilih lokasi di dalam gua yang mempunyai intensitas cahaya tinggi. Sebaliknya, jenis-jenis Microchiroptera lebih menggunakan kemampuan ekolokasinya untuk berorientasi pada ruang sehingga tidak memerlukan cahaya. Berdasarkan faktor mikroklimat yang berpengaruh tersebut, terdapat lima kelompok kelelawar. Setiap kelompok memiliki pola pemilihan sarang yang spesifik.
Kelelawar insektivora di Karst Gombong memangsa 29 famili serangga yang termasuk ke dalam 10 ordo, sedangkan kelelawar frugivora di Karst Gombong memakan polen 33 species tumbuhan yang termasuk dalam sembilan famili. Berdasarkan pemilihan pakannya tersebut, kelelawar Microchiroptera berkelompok menjadi lima kelompok, masing masing kelompok memangsa serangga dengan karakteristik berbeda. Kelelawar Megachiroptera mengelompok menjadi tiga kelompok, masing masing kelompok memilih ukuran polen berbeda dan berasal dari bunga dengan tipe tertentu. Hasil penghitungan indeks kesamaan relung pakan menunjukkan kelelawar yang berasosiasi dalam satu gua yang sama memiliki nilai indeks kesamaan relung pakan kecil (< 30%). Hal ini membuktikan bahwa jenis-jenis yang berasosiasi tersebut tidak berkompetisi memperebutkan makanan. Hal ini menjawab pertanyaan, mengapa satu gua dapat dihuni oleh beberapa jenis kelelawar dengan jumlah populasi yang sangat tinggi.
Hasil penelusuran sarang kelelawar membuktikan beberapa jenis-jenis kelelawar mampu bertahan hidup dalam sarang dengan kondisi dingin, lembap, rendah oksigen, dan tinggi kadar amonia. Hasil penelitian terdahulu membuktikan bahwa kondisi tersebut kurang menguntungkan bagi hewan mamalia. Penelitian ini menunjukkan adanya adaptasi struktural dan fisiologi organ pernapasan kelelawar yang bersarang pada kondisi suhu rendah, lembap, dan kadar amonia tinggi, yaitu dengan diameter alveolus sempit, rasio jumlah eritrosit/bobot tubuh tinggi dan rasio kadar hemoglobin/bobot tubuh tinggi. Pada kondisi udara dingin, lembap dan tinggi kadar amonia kelelawar membutuhkan lebih banyak energi untuk menghangatkan tubuh dan memproduksi zat antibodi. Oleh karenanya, sel tubuh membutuhkan lebih banyak oksigen. Diameter alveolus yang menyempit diperlukan agar absorbsi oksigen oleh pembuluh darah efektif. Jumlah eritrosit dan hemoglobin yang tinggi diperlukan agar kebutuhan sel akan oksigen segera terpenuhi akibat meningkatnya laju respirasi.
Hasil penelitian ini menunjukkan bahwa Karst Gombong perlu dilindungi karena memiliki keunikan ekosistem, dihuni oleh spesies yang terancam punah,

memiliki kanekaragaman spesies yang tinggi, memiliki fungsi perlindungan hidrologi dan potensial untuk wisata alam. Upaya perlindungan di Karst Gombong ini dapat dicapai dengan strategi konservasi. Konservasi sumber daya alam tersebut akan berhasil bila dilakukan atas dasar hasil penelitian yang akurat. Agar dapat dijadikan pedoman bagi semua unsur yang terlibat dalam pemanfaatan kawasan Karst Gombong, maka perlu ditetapkan status kawasan Karst Gombong sesuai dengan hasil penelitian di lapangan dan berpedoman pada peraturan perundang-undangan yang berlaku di Indonesia. Berdasarkan temuan pada penelitian ini kawasan Karst Gombong dapat diusulkan sebagai Kawasan karst kelas I sesuai dengan Kepmen ESDM Nomor 1456 K/20/MEM/2000 pasal 11. Hal ini karena kawasan Karst Gombong terbukti merupakan penyimpan air bawah tanah, merupakan ekosistem unik, habitat satwa khas dan satwa terancam punah, serta berpotensi wisata. Gua Macan, Gua Celeng, Gua Dempo, Gua Inten, Gua Jati jajar, Gua Kemit, Gua Liyah dan Gua Petruk berfungsi menyimpan air bawah tanah, memiliki dekorasi indah/speleotom aktif, habitat fauna khas/ unik, dan habitat fauna terancam punah. Oleh karenanya untuk mempertahankan fungsi ekologis gua dan komunitas biota yang dilindungi, ke-delapan gua tersebut perlu ditetapkan sebagai gua konservasi dan ditetapkan sebagai zona inti/zona perlindungan. Zona inti merupakan zona atau kawasan yang dilindungi dan kegiatan manusia dikendalikan secara ketat. Gua-gua yang ditetapkan sebagai zona inti tersebut dapat dimanfaatkan sebagai objek wisata dengan persyaratan khusus, yaitu dikelola sesuai dengan kebutuhan anatomi dan fisiologis kelelawar yang menghuni di dalamnya. Untuk menjamin kecukupan pakan kelelawar dan juga untuk mempertahankan mikroklimat di dalam gua kawasan hutan dan semak sekitar lima kilometer di luar zona inti perlu diusulkan sebagai zona penyangga/zona pemanfaatan tradisional. Pada zona penyangga kegiatan manusia diperkenankan tetapi, dengan pengendalian dan memenuhi ciri berkelanjutan. Kata kunci : Kelelawar, gua karst, relung pakan, adaptasi, strategi konservasi.


HALAMAN PENGESAHAN
Judul Disertasi : Ekologi, Relung Pakan, dan Strategi Adaptasi Kelelawar
Penghuni Gua di Karst Gombong Kebumen, Jawa Tengah Nama : Fahma Wijayanti NRP : G361060011 Program Studi : Biologi
Disetujui
Komisi Pembimbing
Dr. Ir. Dedy Duryadi Solihin, DEA Ketua
Prof. Dr. Ir. Hadi Sukadi Alikodra, MSc Prof. Ris. Dr. Ibnu Maryanto Anggota Anggota
Mengetahui
Ketua Program Studi Dekan Sekolah Pascasarjana Biologi
Dr. Ir. Dedy Duryadi Solihin, DEA Dr.Ir. Dahrul Syah, MSc Agr
Tanggal Ujian : 24 Mei 2011 Tanggal Lulus:


RIWAYAT HIDUP Penulis dilahirkan di Jakarta pada tanggal 17 Maret 1969 sebagai anak ke lima dari tujuh bersaudara, dari pasangan Prof. Dr. Hadjid Harna Widagdo dan Sudarsini. Menikah pada tahun 1993 dengan Nur Muhammad Busro, SE dan dikaruniai tiga orang putri : Iftina Amalia (16 tahun), Adelia Khairunnisa (10 tahun), dan Alysa Ilmi Aulia (4 tahun). Pendidikan Sarjana ditempuh di Fakultas Biologi, Universitas Jenderal Soedirman Purwokerto lulus tahun 1993. Pendidikan Pascasarjana ditempuh di Program Studi Biologi Program Pascasarjana Universitas Indonesia, lulus tahun 2001. Kesempatan melanjutkan program doktor pada Program Studi Biologi IPB diperoleh pada tahun 2006 dengan bantuan biaya pendidikan Program Doktor dari Departemen Agama RI. Penulis pertama kali bekerja sebagai dosen pada Universitas Muhammadiayah Hamka (UHAMKA) pada tahun 1994 dan pernah menjabat sebagai ketua Program Studi Pendidikan Biologi pada tahun 1999 s/d 2003. Pada tahun 2002 penulis diangkat sebagai dosen PNS di UIN Syarif Hidayatullah Jakarta, dan pada tahun 2004 s/d 2006 pernah menjabat sebagai Ketua Program Studi Biologi.
Selama mengikuti program S3 di IPB karya ilmiah berjudul Komunitas fauna Gua Petruk dan Gua Jatijajar telah disajikan pada Seminar Nasional Perhimpunan Biologi Indonesia di Malang pada tahun 2009, dan karya ilmiah berjudul Biodiversity and Pattern of Roosting Preference of Cave Dwelling Bats: Case Study at Several Caves in Gombong Karst Area, Kebumen, Central Java telah disajikan pada International 2nd South East Asian Bats Conference pada tanggal 6 Juni 2011 di Bogor. Tiga artikel telah diterbitkan yaitu: 1) Pengaruh Fisik Gua Pada Struktur Komunitas Kelelawar di Jurnal Biologi Lingkungan Volume 4 Nomor 1, April 2010; 2) Keanekaragaman Jenis Kelelawar serta Kondisi Mikroklimat Habitatnya pada Beberapa Gua di Kabupaten Kebumen di Jurnal Biologi Lingkungan Volume 4 Nomor 2, Oktober 2010; dan 3) Eritrosit dan Hemoglobin pada Kelelawar Gua di Kawasan Karst Gombong Kebumen Jawa Tengah di Jurnal Biologi Indonesia Volume 7 Nomor 11, Juni 2011. Sementara satu karya ilmiah lain sedang dalam proses penerbitan berjudul The Diet of Insectivorous Cave Dwelling Bats Based on Stomach Content di Journal of Tropical Biology yang akan terbit pada 2012.


KATA PENGANTAR
Berkat rahmat dan hidayah Allah yang maha pemurah, penyusunan disertasi berjudul Ekologi, Relung Pakan, dan Strategi Adaptasi Kelelawar Penghuni Gua di Karst Gombong Kebumen Jawa Tengah ini dapat diselesaikan. Penulis mengucapkan terima kasih kepada Dr. Ir. Dedy Duryadi Solihin, DEA selaku ketua komisi pembimbing, Prof.Dr. H.S.Alikodra MSc, dan Prof. Ris. Dr. Ibnu Maryanto, MSi selaku anggota komisi pembimbing yang telah memberikan arahan, petunjuk, dan motivasi sejak penyusunan proposal penelitian sampai pada penyempurnaan disertasi ini.
Banyak pihak telah ikut dalam pelaksanaan penelitian ini dan membantu penyelesaian disertasi baik di lapangan (Karst Gombong), di Jakarta, maupun di Bogor. Segala bantuan yang telah diberikan, baik moril maupun materil, dengan segala kerendahan hati penulis berkenan mengucapkan terima kasih. Kiranya segala bantuan tersebut tercatat sebagai ibadah, dan mendapat balasan kebaikan dari Allah SWT. Dalam melaksanakan penelitian maupun penulisan disertasi ini, apabila terdapat tingkah laku, tutur kata, sikap, maupun perbuatan penulis yang tidak berkenan, penulis mohon maaf yang sebesar-besarnya.
Penulis menyadari bahwa disertasi ini mungkin masih mengandung kekurangan atau kesalahan, meskipun penulis sudah berusaha sedimikian rupa untuk menyempurnakannya. Dengan berlapang dada kepada semua pihak yang mengetahui kekurangan atau kesalahan dalam disertasi ini, penulis sangat mengharapkan, menghormati, dan menghargai segala saran, kritikan, dan masukan-masukan untuk penyempurnaannya. Semoga disertasi ini dapat bermanfaat bagi kemaslahatan umat.
Bogor, Mei 2011 Penulis

ix
DAFTAR ISI
Halaman DAFTAR TABEL ........................................................................................... xi DAFTAR GAMBAR ...................................................................................... xiii DAFTAR LAMPIRAN ................................................................................... xv
PENDAHULUAN .......................................................................................... 1
Latar Belakang ....................................................................................... 1 Rumusan Masalah .................................................................................. 3 Tujuan Penelitian ................................................................................... 4 Hipotesis Penelitian ............................................................................... 4 Ruang Lingkup Penelitian ..................................................................... 5
TINJAUAN PUSTAKA ................................................................................. 7
BIODIVERSITAS DAN POLA PEMILIHAAN SARANG KELELAWAR: STUDI KASUS DI KAWASAN KARST GOMBONG KABUPATEN KEBUMEN JAWA TENGAH ....................................................................... 30
Abstract .................................................................................................. 30 Pendahuluan ........................................................................................... 31 Bahan dan Metode ................................................................................. 32 Hasil ....................................................................................................... 39 Pembahasan ........................................................................................... 48 Kesimpulan ............................................................................................ 55 Saran ...................................................................................................... 56
ANALISIS RELUNG PAKAN KELELAWAR INSEKTIVORA (Subordo: Microchiroptera) DAN KELELAWAR FRUGIVORA (Subordo: Megachiroptera) YANG BERSARANG DI GUA-GUA KARST GOMBONG KABUPATEN KEBUMEN JAWA TENGAH ......................... 57
Abstract .................................................................................................. 57 Pendahuluan ........................................................................................... 57 Bahan dan Metode ................................................................................. 59 Hasil ....................................................................................................... 62 Pembahasan ........................................................................................... 77 Kesimpulan ............................................................................................ 84 Saran ...................................................................................................... 84
ADAPTASI STRUKTURAL DAN FISIOLOGI PERNAPASAN KELELAWAR PENGHUNI GUA: STUDI KASUS DI KAWASAN KARST GOMBONG KABUPATEN KEBUMEN JAWA TENGAH .......... 85
Abstract .................................................................................................. 85 Pendahuluan ........................................................................................... 85 Bahan dan Metode ................................................................................. 88

x
Hasil ....................................................................................................... 91 Pembahasan ........................................................................................... 98 Kesimpulan ............................................................................................ 103 Saran ...................................................................................................... 103
PEMBAHASAN UMUM ............................................................................... 104 Pentingnya Upaya Konservasi bagi Ekosistem Karts Gombong ........... 104 Perhatian Pemerintah terhadap Ekosistem Karts Gombong .................. 107 Usulan Strategi Konservasi Ekosistem Karts Gombong ....................... 109
KESIMPULAN ............................................................................................... 116
SARAN …………………………………………………………………… ... 116
DAFTAR PUSTAKA ..................................................................................... 117
LAMPIRAN .................................................................................................... 124

xi
DAFTAR TABEL
Halaman
1. Kondisi fisik gua dan struktur komunitas kelelawar ............................. 40
2. Jenis dan kelimpahan kelelawar pada gua-gua yang dihuni kelelawar. . 41
3. Struktur komunitas kelelawar di setiap gua ........................................... 42
4. Hasil pengukuran mikroklimat sarang kelelawar .................................. 45
5. Urutan parameter fisik mikroklimat yang mempengaruhi pemilihan sarang kelelawar .................................................................................... 46
6. Pengelompokan kelelawar berdasarkan pola pemilihan sarang ............ 47
7. Hasil analisis isi perut kelelawar Microchiroptera ................................. 63
8. Karakteristik serangga pakan: bobot tubuh, panjang tubuh (berdasarkan serangga pembanding) dan kekerasan eksoskeleton (berdasarkan Aguirre et al. 2003) ........................................................... 64
9. Nilai kesamaan relung pakan (Morsita index) antarjenis kelelawar pada semua gua ..................................................................................... 67
10. Nilai kesamaan relung pakan (niche overlap) dan nilai uji chi square antarjenis kelelawar yang bersarang dalam satu gua yang sama .......... 68
11. Persentase polen pada saluran pencernaan tiap jenis kelelawar ............ 70
12. Jenis tanaman berdasarkan tipe bunga .................................................. 73
13. Jenis tanaman berdasarkan ukuran polen .............................................. 74
14. Nilai kesamaan relung pakan dan uji chi square antarjenis kelelawar Megachiroptera ...................................................................................... 77
15. Hasil uji korelasi Spearman hubungan antara diameter alveolus, jumlah eritrosit, dan kadar hemoglobin dengan kelembapan, suhu, kadar oksigen, dan kadar amonia ............................................................ 92
16. Hasil pengukuran mikroklimat sarang, jumlah eritrosit, kadar hemoglobin, dan diameter alveolus kelelawar ........................................ 93

xii
17. Hasil uji Tukey perbandingan diameter alveolus antarkoloni kelelawar 94
18. Hasil uji Tukey perbedaan rasio jumlah eritrosit per gram bobot badan antarkoloni kelelawar ............................................................................ 95
19. Hasil uji Tukey perbedaan rasio kadar hemoglobin per gram bobot badan antarkoloni kelelawar ................................................................... 97
20. Status konservasi kelelawar berdasarkan Red List IUCN versi 3.1 ( IUCN 2001) ........................................................................................ 104
21. Pedoman pengelolaan gua berdasarkan identifikasi fungsi gua ............ 108
22. Matriks kondisi fisik dan biota di gua-gua Karts Gombong .................. 111
23. Usulan pemanfaatan ruang gua berdasarkan jenis kelelawar yang bersarang dan prasyarat ekofisiologi yang dibutuhkan .......................... 112

xiii
DAFTAR GAMBAR
Halaman
1. Skema kerangka pemikiran ................................................................... 6
2. Formasi awal terbentuknya karst ........................................................... 8
3. Geomorfologi karst ................................................................................. 10
4. Denah pembagian mintakat pada Gua Jatijajar ....................................... 11
5. Suplai energi ke dalam gua ................................................................... 13
6. Rantai makanan di Gua Anak Takun Malaysia ....................................... 14
7. Struktur rangka kelelawar ....................................................................... 16
8. Organ pernapasan mamalia .................................................................... 25
9. Alveolus dan kapiler darah tempat difusi oksigen dan karbon dioksida 26
10. Struktur kimia hemoglobin ................................................................... 27
11. Peta citra satelit LANDSAT Karst Gombong Kabupaten Kebumen Jawa Tengah ............................................................................................ 32
12. Alat pemetaan roosting kelelawar dan pengukuran mikroklimat gua ... 33
13. Pemetaan gua dengan metode foreward ................................................ 35
14. Pengukuran kadar amonia udara ............................................................ 36
15. Metode penghitungan populasi kelelawar pada tiap sarang .................... 36
16. Pengambilan sampel kelelawar ............................................................... 37
17. Jenis-jenis kelelawar yang ditemukan pada setiap gua ......................... 41
18. Redundancy Analysis (RDA) hubungan kelimpahan (N), kekayaan jenis (S), keanekaragaman jenis (H), dan kemerataan jenis (E) kelelawar dengan panjang lorong gua (PG), lebar lorong gua (LG), tinggi lorong gua (TG), jumlah pintu gua (P), dan jumlah ventilasi gua (V) .......................................................................................................... 43
19. Grafik analisis Canonical Correspondence Analysis (CCA) jenis kelelawar berdasarkan kondisi fisik mikroklimat sarang ........................ 47

xiv
20. Hasil Principle Component Analysis (PCA) jenis kelelawar berdasarkan serangga pakannya ............................................................. 66
21. Hasil Principle Component Analysis (PCA) jenis kelelawar berdasarkan polen yang dimakan ......................................................... 71
22. Hybrid Canonical Correspondent Analysis (HCCA) pemilihan pakan jenis kelelawar Frugivora berdasarkan bentuk mahkota bunga ............. 75
23. Hybrid Canonical Correspondent Analysis (HCCA) pemilihan pakan jenis kelelawar Frugivora berdasarkan ukuran polen ............................. 75
24. Proses pembedahan dan perendaman paru-paru dalam laritan Bouin .... 90
25. Patung deorama cerita pewayangan Rama dan Shinta di lorong Gua Jatijajar ................................................................................................... 106
26. Usulan zonasi kawasan Karst Gombong Kabupaten Kebumen .............. 115

xv
DAFTAR LAMPIRAN
Halaman
1 Gambar jenis-jenis kelelawar penghuni gua Karst Gombong................. 124
2. Peta sebaran sarang kelelawar di gua-gua Karst Gombong ................... 128
3. Serangga pakan kelelawar insektivora/Microchiroptera ....................... 140
4. Polen tumbuhan sumber pakan kelelawar ............................................... 143
5. Gambar penampang melintang sayatan histologi alveolus kelelawar ..... 148
6. Nilai loading factor komponen Principle Component Analysis (PCA) jenis kelelawar berdasrkan jenis serangga pakannya dengan tiga komponen utama .................................................................................... 152
7 Nilai loading factor komponen Principle Component Analysis (PCA) jenis kelelawar berdasarkan polen tumbuhan pakannya ......................... 153

PENDAHULUAN
Latar Belakang
Indonesia merupakan negara yang kaya keanekaragaman hayati. Salah satu
bentuk keanekaragaman hayati Indonesia adalah ekosistem karst. Ekosistem
karst adalah kesatuan komunitas mahluk hidup dengan berbagai faktor
lingkungan yang terdapat pada suatu kawasan dengan batuan dasar berupa batu
gamping atau kapur. Ciri khas kawasan karst adalah adanya celah sinkholes
(sarang air), sungai bawah tanah, dan gua (Samodra 2006). Celah sinkholes dan
sungai bawah tanah pada ekosistem karst dapat menyimpan banyak air, sehingga
ekosistem karst berfungsi sebagai reservoar air (Vermeulen & Whitten 1999).
Selain itu, ekosistem karst juga berfungsi sebagai habitat biota khas gua karena
kondisi unik gua karst yang hanya dapat dihuni oleh fauna tertentu saja
(Epsinasa & Vuong 2008).
Menurut BAPPENAS (2003), terdapat sekitar 15.4 juta hektar kawasan
karst di wilayah Indonesia atau sekitar 10% dari seluruh luas daratan Indonesia.
Selama ini, pemerintah dan masyarakat memanfaatkan ekosistem karst tersebut
sebagai sumber pendapatan, di antaranya untuk kegiatan pertambangan dan obyek
wisata. Namun, pemanfaatan dua sektor ini belum didukung oleh dasar ilmu
pengetahuan (technopreunership) yang kuat, sehingga pemanfaatannya sering kali
menimbulkan persoalan. Menurut Ko (1999), persoalaan utama yang timbul
akibat pemanfaatan ekosistem karst adalah : 1) Adanya perubahan bentang alam
(landscape) dan struktur geologi karst akibat penambangan batu gamping; 2)
Menurunnya debit air bawah tanah akibat berkurangnya porositas batuan karst;
dan 3) Hilangnya keanekaragaman biota khas gua, akibat habitatnya rusak atau
terganggu.
Persoalan yang timbul akibat pemanfaatan ekosistem karst tersebut dapat
menyebabkan ekosistem karst tidak lagi memberikan manfaat ekonomi dan
fungsi ekologi. Untuk menghindari hal tersebut, diperlukan upaya konservasi
ekosistem karst. Hal ini hanya dapat dipenuhi jika pengetahuan yang mendasari
kestabilan ekosistem karst ini dapat dipahami dengan baik.

2
Penelitian untuk mendukung pemanfaatan ekosistem karst secara lestari
masih sangat sedikit. Selain itu, beberapa hasil penelitian yang telah dilakukan di
kawasan karst di Indonesia terutama hanya terfokus pada struktur geologi karst
saja. Penelitian mengenai komunitas biologi belum banyak dilakukan, padahal
komunitas biologi memegang peran penting dalam keseimbangan ekosistem karst.
Atas dasar alasan tersebut, penelitian mengenai komunitas biologi pada ekosistem
karst mutlak diperlukan.
Salah satu komunitas biologi yang berperan penting dalam ekosistem
karst adalah fauna troglozene, yaitu fauna yang bersarang di dalam gua, tetapi
mencari makan di luar gua (Epsinasa & Vuong 2008). Menurut Whitten et al.
(1999), hewan troglozene utama pada gua-gua karst di Indonesia adalah
kelelawar. Sebagai hewan troglozene, kelelawar mensuplai energi ke ekosistem
gua dengan guano (feses kelelawar) dan bangkainya. Tanpa kehadiran kelelawar,
aliran energi ke dalam ekosistem gua akan terhenti dan keanekaragaman biota
gua akan hilang.
Selain berperan penting dalam kestabilan ekosistem gua, kelelawar juga
memegang fungsi ekologi penting bagi ekosistem luar gua. Peran kelelawar bagi
ekosistem luar gua adalah sebagai pemencar biji, penyerbuk berbagai jenis
tumbuhan dan pengendali/predator serangga hama tanaman. Penelitian Tan et al.
(1998) membuktikan kelelawar Cynopterus brachyotis (Subordo:
Megachiroptera) di Bangi Malaysia memakan buah dan menyebarkan biji 17
famili tumbuhan hutan dan tanaman perkebunan. Penelitian Razakarivony et al.
(2005) di Malagasy membuktikan beberapa kelelawar subordo Microchiroptera
yang bersarang di gua (Hipposideros commersoni, Miniopterus manavi dan
Myotis goudoti) memakan serangga ordo Isoptera, Hymenoptera, Cooleoptera,
Lepidoptera, Orthoptera, Hemiptera, dan Homoptera. Anggota ordo serangga
tersebut tercatat sebagai serangga hama tanaman. Oleh sebab itu, hilangnya
komunitas kelelawar di dalam gua karst tidak hanya dapat menghancurkan
ekosistem dalam gua, tetapi juga dapat mempengaruhi ekosistem luar gua.
Potensi ekonomi ekosistem karst menyebabkan eksploitasi ekosistem
karst tidak dapat dihindari. Akibat eksploitasi untuk tujuan ekonomi ini,
ekosistem karst menanggung risiko yang cukup tinggi, salah satunya adalah

3
sarang kelelawar di gua yang telah dipertahankan dari generasi ke generasi tidak
lagi berfungsi sebagaimana mestinya. Hal ini karena kondisi fisik gua telah
berubah, menjadi tidak sesuai lagi dengan kebutuhan kelelawar. Oleh karena itu,
agar keberadaan kelelawar di gua dapat dipertahankan, diperlukan informasi yang
akurat mengenai segala aspek yang berkaitan dengan kehidupan kelelawar gua.
Penelitian ini akan menjawab beberapa aspek yang berkaitan dengan
kelelawar penghuni gua. Aspek-aspek tersebut meliputi biodiversitas dan struktur
komunitas kelelawar gua, pola pemilihan sarang, relung pakan, serta adaptasi
struktural dan fisiologi organ pernapasan kelelawar gua. Dari hasil penelitian
tersebut dirancang strategi konservasi ekosistem gua, sehingga keberadaan
kelelawar sebagai kunci penyedia energi ekosistem (key factor in cycle energy)
dalam gua dan pemegang peran ekologis lainnya bagi ekosistem luar gua dapat
dipertahankan.
Rumusan Masalah
Ekosistem karst berperan penting sebagai reservoar air dan habitat fauna
khas gua (Samodra 2006; Epsinasa & Vuong 2008). Akibat eksploitasi dan
pemanfaatan gua untuk kepentingan ekonomi yang kurang berwawasan
lingkungan, keberadaan dan peran ekosistem karst menjadi terancam. Salah satu
ancaman dari pemanfaatan ekosistem gua adalah rusaknya struktur gua (Ko
1999). Di samping permasalahan struktur gua, terdapat permasalahan lain yang
tidak kalah pentingnya akibat pemanfaatan gua. Permasalahan tersebut adalah
permasalahan komunitas biologi. Eksploitasi gua dengan kegiatan pertambangan
dan pengambilan guano sudah pasti merusak fisik gua dan mengancam komunitas
fauna gua. Demikian pula dengan pemanfaatan gua karst sebagai objek wisata.
Walaupun secara fisik tidak merusak struktur gua, kedatangan pengunjung dan
fasilitas wisata yang dibangun dapat mengganggu kenyamanan dan keamanan
fauna gua. Salah satu fauna gua yang terancam akibat pemanfaatan gua tersebut
adalah kelelawar. Untuk meminimalisir dampak pemanfaatan gua karst pada
komunitas kelelawar, diperlukan penelitian mengenai ekologi, relung pakan,
strategi adaptasi dan strategi konservasinya. Luaran penelitian ini adalah 1)
biodiversitas kelelawar gua, 2) pola pemilihan sarang kelelawar gua, 3) kesamaan
relung pakan kelelawar gua, 4) strategi adaptasi struktural dan fisilogi pernapasan

4
kelelawar gua, dan 5) strategi konservasi ekosistem Karst Gombong. Apabila
luaran di atas berhasil dicapai dengan baik, maka dapat dibuat pola pemanfaatan
gua yang tepat, yang dapat mempertahankan komunitas kelelawar dan
ekosistemnya.
Tujuan Penelitian
Tujuan penelitian ini adalah :
a. Mengkaji biodiversitas kelelawar penghuni gua berdasarkan faktor-faktor
fisik gua di beberapa gua Karst Gombong.
b. Mengidentifikasi faktor mikroklimat gua yang berpengaruh pada pemilihan
sarang kelelawar gua.
c. Mengidentifikasi pakan kelelawar gua dan menentukan kesamaan relung
(niche overlap) pakan kelelawar yang berasosiasi dalam satu gua yang
sama.
d. Mengkaji adaptasi struktural dan fisiologi pernapasan kelelawar yang
bersarang di gua-gua Karst Gombong.
e. Merancang strategi konservasi ekosistem Karst Gombong.
Hipotesis penelitian
Hipotesis penelitian ini adalah :
a. Keanekaragaman, kelimpahan, dan kemerataan jenis kelelawar dipengaruhi
secara nyata oleh panjang lorong, lebar lorong, tinggi lorong, serta jumlah
pintu, dan jumlah ventilasi gua.
b. Pola pemilihan sarang kelelawar gua dipengaruhi oleh jarak dari mulut
gua, tinggi atap gua, suhu, kelembapan udara, intensitas cahaya, kecepatan
angin, intensitas suara, kadar oksigen, dan kadar amonia di sekitar sarang.
c. Asosiasi bersarang dalam satu gua oleh beberapa jenis kelelawar dapat
dilakukan oleh jenis-jenis kelelawar yang memiliki indeks kesamaan relung
pakan yang kecil (< 50%).
d. Adaptasi kelelawar terhadap kondisi ruang gua yang dingin, lembap, rendah
oksigen, dan kadar amonia tinggi menentukan struktur dan fisiologi organ
pernapasan, yaitu diameter alveolus sempit (< 100µm), rasio jumlah

5
eritrosit/ bobot badan tinggi ( > 0.3 juta/ml /gr bobot badan) dan rasio kadar
hemoglobin/bobot badan tinggi ( > 0.9 g/ml /gr bobot badan).
Ruang Lingkup Penelitian
Secara sistematik, ruang lingkup penelitian dapat dilihat pada Gambar 1.
Namun, tahapan penelitian dilakukan secara bertahap mulai dari penelitian
tentang biodiversitas kelelawar, hingga strategi konservasi ekosistem karst
dilakukan secara rinci melalui beberapa topik penelitian sebagai berikut :
Penelitian 1 : Biodiversitas dan pola pemilihan sarang kelelawar penghuni gua:
Studi kasus di gua-gua Karst Gombong Kabupaten Kebumen
Jawa Tengah. Penelitian ini untuk mencapai tujuan a, b, dan e.
Penelitian 2 : Analisis relung pakan kelelawar insectivora ( Subordo
Microchiroptera) dan kelelawar frugivora (Subordo:
Megachiroptera) yang bersarang di Gua Karst Gombong.
Penelitian ini untuk mencapai tujuan c dan e.
Penelitian 3 : Adaptasi struktural dan fisiologi pernapasan kelelawar penghuni
gua di Karst Gombong Kabupaten Kebumen Jawa
Tengah. Penelitian ini untuk mencapai tujuan d dan e.
Penelitian 4 : Strategi konservasi ekosistem gua Karst Gombong. Penelitian ini
untuk mencapai tujuan e dan tertuang dalam pembahasan umum.
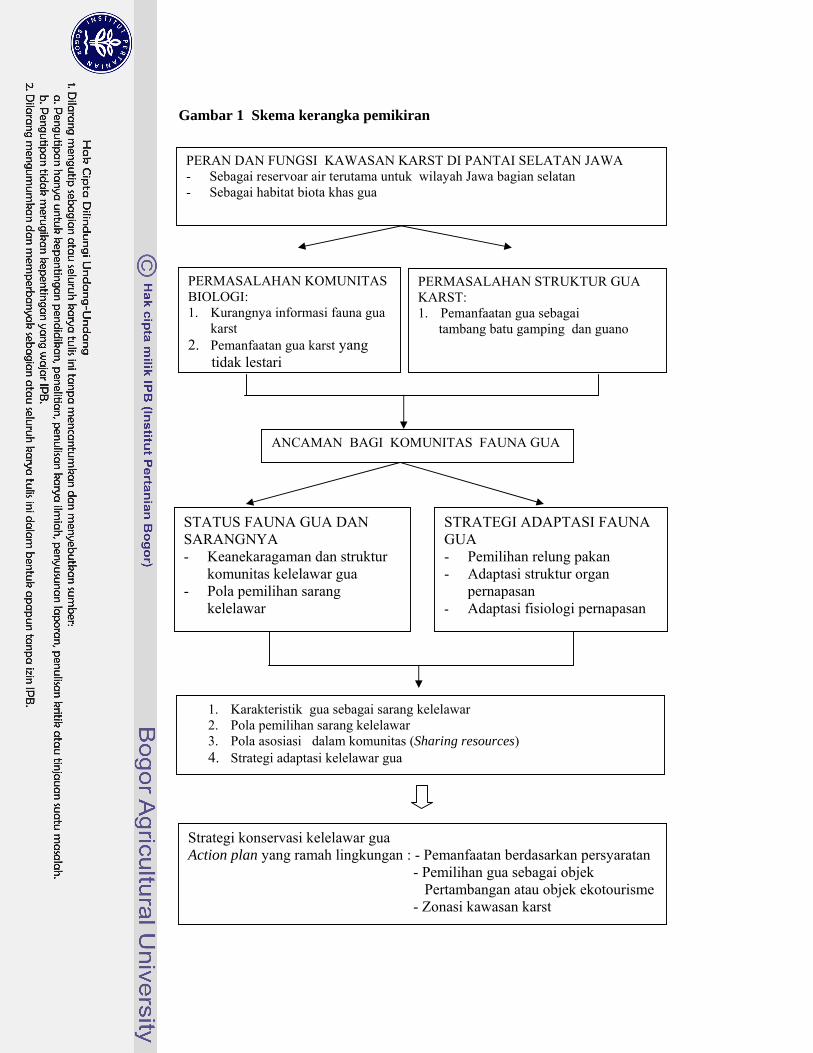
6
Gambar 1 Skema kerangka pemikiran
PERAN DAN FUNGSI KAWASAN KARST DI PANTAI SELATAN JAWA - Sebagai reservoar air terutama untuk wilayah Jawa bagian selatan - Sebagai habitat biota khas gua
PERMASALAHAN STRUKTUR GUA KARST: 1. Pemanfaatan gua sebagai tambang batu gamping dan guano
PERMASALAHAN KOMUNITAS BIOLOGI: 1. Kurangnya informasi fauna gua
karst 2. Pemanfaatan gua karst yang
tidak lestari
ANCAMAN BAGI KOMUNITAS FAUNA GUA
STATUS FAUNA GUA DAN SARANGNYA - Keanekaragaman dan struktur
komunitas kelelawar gua - Pola pemilihan sarang
kelelawar
1. Karakteristik gua sebagai sarang kelelawar 2. Pola pemilihan sarang kelelawar 3. Pola asosiasi dalam komunitas (Sharing resources) 4. Strategi adaptasi kelelawar gua
STRATEGI ADAPTASI FAUNA GUA - Pemilihan relung pakan - Adaptasi struktur organ
pernapasan - Adaptasi fisiologi pernapasan
Strategi konservasi kelelawar gua Action plan yang ramah lingkungan : - Pemanfaatan berdasarkan persyaratan
- Pemilihan gua sebagai objek Pertambangan atau objek ekotourisme - Zonasi kawasan karst

7
TINJAUAN PUSTAKA
Ekosistem Karst
Karstifikasi
Kata karst berasal dari bahasa Yugoslavia dan diperkenalkan oleh Cvijic
seorang geolog asal Jerman pada tahun 1850. Kata karst tersebut mengacu pada
kawasan batu gamping di Kota Trieste, Slovenia, Yugoslavia (Wirawan 2005).
Sampai saat ini, kata karst telah digunakan secara internasional dan telah diserap
secara utuh sebagai kata bahasa Indonesia. Salah satu definisi karst yang
dikemukakan oleh ahli geologi adalah bentang alam (landscape) pada lempeng
batuan gamping yang dibentuk oleh pelarutan batuan gamping. Pelarutan batu
gamping tersebut menghasilkan bentukan karst dengan ciri celah sinkhole
(lubang lari air), sungai bawah tanah, dan gua (Hamilton & Smith 2006;
Samodra 2006).
Proses terbentuknya karst (karstifikasi) berlangsung selama jutaan tahun
melalui peristiwa yang melibatkan faktor-faktor geologi, fisika, kimia, dan
biologi. Karstifikasi diawali dengan pergerakan lempeng bumi yang bersifat
dinamis. Pergerakan lempeng bumi tersebut menyebabkan lempeng saling
bertabrakan dan menghasilkan gaya tektonik yang mendorong sebagian lempeng
ke atas. Peristiwa ini menyebabkan sedimentasi sisa-sisa tumbuhan dan hewan
yang mengandung kapur (kalsium karbonat) terangkat dari dasar laut ke
permukaan (Gimes 2001). Menurut Yunqiu et al. (2006) biota laut tersebut
antara lain, koral (Pontes, Neandrina, Acropora, Siderastrea, Ginoid), Briozoa,
ganggang (Halimeda, Lithothamniam, Penicillus, Acialaria, Neomen),
Foraminifera, dan Moluska. Peristiwa yang disebabkan oleh gaya tektonik ini
menghasilkan deretan bukit kapur/gamping di permukaan laut. Gaya-gaya
tektonik tersebut dapat menyebabkan terjadinya patahan dan retakan yang saling
berasosiasi. Lempeng batuan yang terdeformasi oleh gaya-gaya tektonik ini
merupakan area yang sangat potensial untuk masuknya aliran air dan
terbentuknya perangkap-perangkap air (Eberhard 2006). Formasi awal
terbentuknya karst tersaji pada Gambar 2.

8
Gambar 2 Formasi awal terbentuknya karst (Sumber: Subterra 2004)
Setelah proses yang disebabkan oleh gaya tektonik, peristiwa selanjutnya
adalah pelarutan batuan karbonat oleh asam lemah. Reaksi karbon dioksida
(CO2) di udara dengan air hujan (H20) menghasilkan H2CO3 yang bersifat asam
lemah. Larutan tersebut mengalir melalui aliran air permukaan (run off) dan
akan melarutkan batu gamping sehingga terbentuk celah. Lebih rinci Samodra
(2006) menjelaskan reaksi kimia pelarutan batu gamping oleh asam lemah adalah
sebagai berikut :
H2O + CO2 H2CO3
H2CO3 HCO3 + H+
HCO3 + CaO CaCO3 + H2O
CaCO3 + H2O + CO2 CaH2C2O6
Celah yang dihasilkan oleh pelarutan tersebut semakin besar dari waktu ke waktu
sampai membentuk patahan dan rongga yang disebut karen (patahan), sinkhole
(lubang lari air), collapse sink/doline (rongga), dan gua (Gimes 2001). Gaya
tektonik yang terjadi pada masa berikutnya menyebabkan rongga dan gua saling
berasosiasi satu sama lain membentuk sistem perguaan dengan lorong yang
panjang (Samodra 2006). Persyaratan yang harus dipenuhi supaya lempeng
karen

9
batu gamping dapat membentuk morfologi karst menurut Hamilton & Smith
(2006) adalah : 1) lempeng batuan gamping mempunyai ketebalan yang cukup,
2) berada di wilayah dengan curah hujan tinggi, 3) batuan gamping banyak
mengandung celah atau rongga, 4) berada pada posisi lebih tinggi dibandingkan
lingkungan di sekitarnya.
Geomorfologi karst
Ahli geologi membagi geomorfologi karst menjadi karst luar atau exokarst
dan karst dalam atau endokarst (gua). Exokarst/epikarst dicirikan dengan: 1)
adanya bukit-bukit kapur berbentuk kerucut atau kubah, 2) permukaan kasar
berlubang-lubang membentuk dolina (cekungan), dan 3) adanya endapan
sedimen lempung berwarna merah hasil pelapukan batu gamping (Samodra
2006). Selain itu menurut Roemantyo & Noerdjito (2006), exokarst biasanya
tertutup oleh lapisan tanah yang tipis yang umumnya berasal dari batuan kapur
yang hancur dan terdekomposisi secara mekanik dan kimiawi. Lapisan tanah
tipis tersebut sebagian terkumpul pada cekungan. Proses pengayaan nutrisi pada
lapisan tanah exokarst dapat terjadi oleh adanya debu vulkanis, ataupun aliran air
hujan yang membawa humus dari tempat lain. Akibatnya exokarst dapat
ditumbuhi oleh jenis-jenis tumbuhan tertentu.
Endokarst (gua) menurut Hamilton & Smith (2006) merupakan ruang
dengan sirkulasi udara terbatas dan sangat sedikit atau bahkan sama sekali tidak
ada cahaya. Selain itu, menurut Wirawan (2005), ruang dalam gua biasanya
dilengkapi dengan ornamen-ornamen gua. Ornamen tersebut merupakan hasil
pengendapan kalsium karbonat (CaCO3) yang sebelumnya terlarutkan oleh
peristiwa karstifikasi. FINSPAC (1993) membagi ornamen-ornamen dalam gua
menjadi: 1) stalaktit, yaitu endapan kapur yang menggelantung pada langit-langit
gua, 2) stalakmit, yaitu endapan kapur yang terdapat pada lantai gua, 3) tiang
(column), yaitu pertemuan antara stalaktit dan stalakmit yang membentuk tiang,
4) tirai (drip curtain/drapery), yaitu endapan yang berbentuk lembaran tipis
vertikal, pada atap gua yang miring, dan 5) teras (travertin), yaitu endapan
kalsium karbonat pada lantai gua yang bertingkat sehingga membentuk
terrasering. Geomorfologi karst tersaji pada Gambar 3.

10
Gambar 3 Geomorfologi karst (Sumber: FINSPAC 1993)
Hamilton & Smith (2006) membagi lingkungan dalam gua berdasarkan
pengaruh sinar matahari menjadi 3 mintakat, yaitu 1) mintakat I adalah mintakat
di dalam gua yang sinar matahari dapat masuk dan iklim dalam gua masih
dipengaruhi oleh iklim luar gua, 2) mintakat II adalah mintakat di dalam gua
yang tidak ada sinar matahari yang masuk, tetapi iklim di dalam mintakat tersebut
masih dipengaruhi oleh iklim di luar gua, dan 3) mintakat III adalah mintakat
yang tidak ada sinar matahari dan iklim di dalam mintakat ini stabil, tidak
dipengaruhi oleh fluktuasi iklim di luar gua. Contoh denah pembagian mintakat
pada Gua Jatijajar dapat dilihat pada Gambar 4.
Menurut Russo et al. (2003) dinding dan atap gua merupakan penyangga
efektif yang memisahkan lingkungan dalam gua dengan lingkungan luar gua. Oleh
karenanya, lingkungan dalam gua memiliki mikroklimat yang berbeda dari luar
gua. Menurut Samodra (2006) mikroklimat dalam gua cenderung lebih dingin
dan lebih lembap. Hal ini karena 1) adanya aliran sungai di lantai gua; 2) adanya
air rembesan di atap gua; 3) tidak ada sinar matahari, dan 4) sirkulasi udara
terbatas. Selain itu menurut Russo et al. (2003) mikroklimat tersebut dapat
berbeda antara satu zona (mintakat) dan zona lainnya. Hal ini dipengaruhi oleh
pengaruh sinar matahari, formasi gua, dan keberadaan mahluk hidup di
dalamnya. Penelitian Baudinette et al. (1994) di Gua Kelelawar dan Gua

11
Robertson Afrika Selatan membuktikan gua yang dihuni kelelawar dengan jumlah
besar dapat menaikkan suhu dalam gua hingga 3oC.
Gambar 4 Denah pembagian mintakat pada Gua Jatijajar (Sumber : DISPARBUD
Kabupaten Kebumen 2004 yang telah dimodifikasi)
Tipe-tipe gua
Aliran air merupakan faktor utama dalam pembentukan gua karst.
Karenanya menurut Hamilton & Smith (2006), berdasarkan penetrasi air pada
dinding dan atap gua, dapat dibedakan tiga-tipe gua karst, yaitu 1) gua fosil,
adalah gua karst yang pada dinding dan atapnya tidak ada lagi penetrasi air. Hal
ini menyebabkan pertumbuhan ornamen-ornamen gua terhenti dan mikroklimat
gua cenderung lebih kering dibandingkan tipe gua karst lainnya; 2) gua vedosa:
gua karst yang berada pada sarang air (water table) yang datar. Tipe gua ini
ditandai dengan sedikitnya penetrasi air pada atap gua sehingga tidak banyak
ditemukan ornamen gua. Oleh karena berada pada sarang air datar, maka banyak
terbentuk mata air di dinding gua. Banyaknya mata air tersebut menyebabkan
mikroklimat di dalam gua cenderung dingin dan lembap; 3) gua pheartic adalah
gua karst yang berada pada bidang miring, yang penetrasi air pada atap gua
berlangsung sangat efektif. Tipe gua ini ditandai dengan banyaknya tetesan air

12
pada atap gua dan biasanya lantai gua dilalui sungai bawah tanah. Ornamen gua
membentuk formasi yang kompleks dan masih aktif berkembang. Keberadaan
sungai bawah tanah dan banyaknya tetesan air pada atap gua menyebabkan gua
pheartic dingin dan lembap (Samodra 2006).
Pada gua fosil, vedosa maupun pheartic terbentuk zonasi atau mintakat-
mintakat. Mintakat tersebut sangat dipengaruhi oleh besarnya mulut gua,
banyaknya ventilasi gua maupun formasi gua. Gua tipe pheartic memiliki
formasi gua yang lebih kompleks dibandingkan tipe gua lainnya (Samodra 2006).
Hal ini disebabkan lorong gua yang berliku-liku, adanya aliran sungai, dan
banyaknya ornamen-ornamen gua. Kondisi ini menghasilkan mintakat III yang
lebih luas, yaitu mintakat gelap dengan suhu dan kelembapan stabil tidak
dipengaruhi suhu luar gua.
Gua dapat diartikan sebagai ruang /lorong yang berada di bawah permukaan
tanah. Selain tipe-tipe gua karst yang telah dijelaskan di atas, menurut Ko (2004)
terdapat tiga tipe gua lain, yaitu 1) gua lava: gua yang terbentuk karena aktivitas
vulkanik atau gunung berapi, 2) gua litoral /gua laut: gua yang terbentuk kerena
gelombang laut yang memecah karang di pantai, dan 3) gua sandstone: gua yang
terbentuk karena erosi air dan angin pada batuan pasir. Ketiga tipe gua tersebut
memiliki lorong pendek, formasi gua sederhana, dan cahaya matahari dapat
masuk hampir keseluruh bagian lorong gua. Karena itu tidak terdapat mintakat-
mintakat seperti halnya pada gua karst dan tidak membentuk ekosistem yang
kompleks.
Komunitas fauna gua karst
Dinding dan atap gua merupakan pembatas yang memisahkan lingkungan
dalam gua dengan luar gua. Dinding dan atap tersebut tidak tembus sinar
matahari. Akibatnya, kondisi dalam gua menjadi gelap dan tumbuhan hijau
(autotrof) tidak ditemukan. Meskipun demikian, menurut Ko (2004), ruang
dalam gua dapat ditempati oleh mahluk hidup. Hal ini karena sumber energi
didatangkan dari luar gua melalui unsur hara yang terlarut dalam aliran air, debu
zat-zat organik yang terbawa oleh udara serta bahan nutrisi yang berasal dari

13
hewan yang bersarang di dalam gua tetapi mencari makan di luar gua (hewan
Troglozene).
Menurut Ko (2004), di kawasan karst penghubung utama antara ekosistem
luar gua dan ekosistem dalam gua adalah burung dan Mamalia. Jenis-jenis
burung di antaranya adalah walet (Aerodramus fuciphagus) dan sriti (Hirundo
tahitica), sedangkan kelompok Mamalia adalah ordo Chiroptera (kelelawar).
Menurut Whitten et al. (1999) dan Sinaga et al. (2006) fauna troglozene utama di
gua-gua karst di Pulau Jawa adalah kelelawar. Bahkan jumlah populasi kelelawar
tersebut dapat mencapai jutaan individu dalam satu gua. Secara sederhana suplai
energi ke dalam gua disajikan dalam Gambar 5.
Gambar 5 Suplai energi ke dalam gua (Sumber: Subterra 2004)
Berdasarkan sumber energinya, jenis-jenis fauna yang hidup di gua menurut
Ko (2004) dibedakan menjadi: 1) necrophagus, yaitu fauna pemakan bangkai 2)
cocroaphagus, yaitu fauna pemakan kotoran/feses 3) parasit, yaitu fauna yang
hidup pada fauna lain dan 4) predator, yaitu fauna pemakan fauna lain. Penelitian
McCure (1985) di Gua Batu Malaysia mendapatkan necrophagus terdiri atas :
lalat (Muscoidae:Insekta) dan semut (Formicidae: Insekta); cocroaphagus terdiri
atas ekor pegas (Collembola: Insekta/Hexapoda), kumbang (Stratiomyiidae:
Insekta), kecoa (Blattidae: Insekta), kumbang (Tineidae: Insekta), jangkerik
(Gryllothalpidae: Insekta) dan jangkerik (Gryllidae: Insekta); parasit terdiri atas :

14
kutu (Ichneumonidae: Insekta); dan predator terdiri atas : kala jengking
(Scorpionidae: Arachnida) , semut (Formicidae: Inseta) dan ular (Elaphe taeniura:
Reptilia). Penelitian Wirawan (2004) di Gua Pawon Jawa Tengah mendapatkan
ekor pegas (Collembola: Insekta), lalat (Diptera:Insekta), kecoa
(Blatodea:Insekta), dan kumbang (Colleoptera:Insekta) sebagai pemakan guano.
Fauna-fauna tersebut kemudian dimakan oleh kodok (Bufo: Amphibia) dan laba-
laba (Arachnidae: Decapoda). Dunn (1978) mendapatkan rantai makanan di
dalam Gua Anak Takun Malaysia seperti pada Gambar 6.
GUANO
Gambar 6 Rantai makanan di Gua Anak Takun Malaysia (Dunn, 1978)
Ruang dalam gua yang gelap dan lembap menyebabkan fauna gua harus
beradaptasi pada keadaan tersebut. Adaptasi oleh fauna gua ini memerlukan
waktu yang panjang. Hasil adaptasi tersebut menurut Espinasa & Vuong (2008)
menghasilkan ciri-ciri sebagai berikut: 1) tubuh tidak berpigmen, 2) mempunyai
alat gerak yang ramping dan panjang, 3) indera peraba atau pendengar
berkembang; 4) mata tereduksi atau hilang sama sekali, 5) metabolisme lambat.
Menurut Suyanto (2001) dan Espinasa & Vuong (2008) berdasarkan
tingkat adaptasinya, fauna gua dibedakan menjadi :
1) Troglobit, yaitu hewan yang telah mengalami modifikasi khusus sesuai
dengan kondisi gua yang gelap, seperti tidak berpigmen dan lebih
KELELAWAR (CHIROPTERA)
lalat (Muscoidae)
kumbang (Lathridiidae)
jangkerik (Gryllidae)
kecoa (Blattidae
kala jengking (Scorpionodae)
laba-laba (Arachnidae)
semut (Formicidae)
ular (Boidae)
kodok (Anura)
ekor Pegas/ Collembola

15
berfungsinya indera peraba, penciuman, dan pendengaran. Troglobit
merupakan penghuni tetap gua yang tidak dapat hidup di habitat lain. Oleh
karena itu, hewan troglobit merupakan kelompok yang paling fragil di
antara kelompok lainnya. Espinasa & Vuong (2008) mendapatkan serangga
troglobit: Nicoletiid (Zygentoma: Insecta) di Gua Oaxaca, Mexico.
Menurut Whitten et al. (1999) fauna troglobit yang sering ditemukan di
gua-gua karst di Pulau Jawa adalah kepiting (Sesarmoides jacobsoni:
Crustacea ), Udang putih (Macrobrachium poeti: Crustacea) dan ikan buta
(Puntius binotatus: Osteicthyes). Penelitian Wijayanti (2001) di Gua Petruk
dan Gua Jatijajar Jawa Tengah mendapatkan fauna troglobit: ikan buta
(Amblyopsis spelaeus: Osteicthyes), udang gua (Macrobrachium pilimanus:
Crustacea), laba-laba gua (Stigophrynus darmamani: Arachnidea), dan
kumbang gua (Eustra saripaensis: Insekta). Hasil penelitian Rachmadi
(2003) di gua Karst Ngerong, Tuban, Jawa Timur, mendapatkan fauna
troglobit: kalajengking gua (Chaerilus sabinae: Scorpionidae), kepiting gua
(Cancrocaeca xenomorpha: Cruatacea), kepiting mata kecil (Sesarmoides
emdi: Crustacea), isopoda gua (Cirolana marosina: Isopoda), kumbang gua
(Eustra saripaensis:Insecta), dan ekor pegas gua (Pseudosinella maros:
Insecta).
2) Troglozene, yaitu fauna yang secara teratur masuk ke dalam gua untuk
berlindung, beristirahat, dan berkembang biak, tetapi mencari makan di luar
gua. Meskipun hanya sebagian hidupnya berada di dalam gua, hewan
troglozene telah beradaptasi dengan kondisi gua yang gelap. Menurut
Vermeulen & Whitten (1999), fauna troglozene mempunyai kemampuan
echolokasi, yaitu kemampuan menangkap gelombang pantul (gema)
berfrekuensi ultrasonik (>20 KHz). Echolokasi ini berguna untuk
mendeteksi mangsa dan orientasi ruang tanpa mengunakan mata.
Kelompok fauna troglozene merupakan spesies kunci dalam ekosistem gua,
karena fauna troglozene memindahkan energi dari luar gua ke dalam gua.
Fauna troglozene yang sering ditemukan di gua karst di Indonesia adalah
burung walet (Collocalia fuciphaga/Aerodramus fuciphagus), burung sriti
(Hirundo tahitica), dan kelelawar (ordo Chiroptera) (Whitten et al. 1999).

16
3) Troglophil, yaitu fauna yang hidup di dalam gua, tetapi belum mengalami
modifikasi khusus. Fauna ini selama hidupnya berada dalam gua, tetapi
jenis yang sama juga ditemukan di luar gua. Bila terjadi gangguan di dalam
gua, fauna troglophil dapat pindah ke habitat luar gua. Penelitian Castillo et
al. (2009) di Los Ricos Cave, Queretaro, Mexico mendapatkan kodok
(Eleutherodactylus longipes: Anura) sebagai fauna troglophil yang secara
musiman memasuki gua. Menurut Whitten et al. (1999) jangkerik
(Rhapidophora dammarmani: Insekta), kumbang (Collasoma scrutater:
Insekta), laba-laba (Liphistius sp: Arachnidae), dan keong (Thiara scabra:
Gastropoda) merupakan troglophil yang sering dijumpai di gua-gua karst di
Pulau Jawa.
Kelelawar sebagai Kelompok Fauna Troglozene
Kelelawar merupakan fauna troglozene utama di gua-gua karst di Indonesia
(Whitten et al.1999; dan Suyanto 2001). Kelelawar adalah Mamalia yang
termasuk dalam ordo Chiroptera. Ciri khas ordo ini adalah tulang telapak tangan
(metacarpal) dan tulang jari (digiti) mengalami pemanjangan sehingga berfungsi
sebagai kerangka sayap. Sayap tersebut terbentuk dari selaput tipis (petagium)
yang membentang antara tulang-tulang telapak dan jari tangan sampai sepanjang
sisi tubuh (Nowak 1994; Altringham 1996). Nowak (1994) menggambarkan
struktur rangka kelelawar seperti terlihat pada Gambar 7.
Gambar 7 Struktur rangka kelelawar (Sumber : Nowak 1994)
digiti 1
digiti 2
digiti 3
digiti 4
digiti 5
petagium

17
Ordo Chiroptera terdiri atas 2 subordo, yaitu Megachiroptera dan
Microchiroptera. Kedua subordo ini diduga tidak mempunyai hubungan
kekerabatan dan merupakan hasil evolusi konvergen, yaitu evolusi yang terjadi
pada dua spesies yang berbeda tetapi beradaptasi dengan cara yang sama
sehingga menghasilkan morfologi yang mirip (Altringham 1996). Salah satu
alasan yang mendukung adalah : saraf superior colliculus (s.c) kanan pada otak
tengah Microchiroptera mengatur retina mata kiri dan sebaliknya s.c kiri mengatur
retina mata kanan. Hal ini ditemukan pada semua Mamalia, kecuali primata. Pada
Megachiroptera, saraf superior colliculus kanan otak tengah mengatur retina mata
kiri dan mata kanan sekaligus. Keadaan ini hanya ditemukan pada Primata,
Dermoptera, dan Megachiroptera (Corbet & Hill 1992; Altringham 1996). Karena
alasan tersebut maka diduga Megachiroptera berasal dari nenek moyang Primata,
sedangkan Microchiroptera diduga berasal dari nenek moyang bukan Primata.
Penelitian HanGuan et al. (2006) tentang philogenetika kelelawar juga
mendapatkan bahwa kelelawar Megachiroptera memiliki kekerabatan lebih dekat
dengan primata dibandingkan dengan Microchiroptera. Saat ini diketahui
terdapat 18 famili, 192 genus dan sekitar 1111 jenis kelelawar yang ada di dunia
(Safi & Kerth 2004). Menurut Suyanto et al. (1998) terdapat 10 famili, 49 genus,
dan sekitar 151 jenis terdapat di Indonesia.
Anggota subordo Megachiroptera makanan utamanya adalah buah
(frugivora), selain itu juga memakan serbuk sari (polen) dan nektar. Subordo ini
terdiri atas 1 famili, yaitu Pteropodidae dengan 42 genus dan 166 spesies (Nowak
1994). Menurut Altringham (1996) anggota subordo Megachiroptera memiliki
ukuran yang relatif besar (bobot minimum 10 gram maksimum 1500 gram dengan
bentangan sayap maksimum 1700 mm); memiliki mata besar; telinga tidak
memiliki tragus; moncong sederhana dan ekor tidak berkembang; jari kedua dan
jari ketiga terpisah relatif jauh dan memiliki cakar pada jari kedua, kecuali pada
Eonycteris, Dobsonia, dan Neopterix.
Anggota subordo Microchiroptera kebanyakan pemakan serangga
(insectivora). Selain itu, ada juga yang penghisap darah (sanguivora), misalnya
Desmodus vampirus; dan penghisap madu misalnya (Leptonycteris curasoae).
Subordo ini terdiri atas 17 famili, 150 genus, dan 945 spesies. Ciri

18
Microchiroptera adalah berukuran kecil (bobot minimum 2 gram, maksimum 196
gram dengan bentangan sayap maksimum 70 mm); memiliki mata kecil; telinga
memiliki tragus (tonjolan dari dalam daun telinga) atau anti tragus (tonjolan dari
luar daun telinga); jari sayap tidak bercakar dan moncong sangat bervariasi,
terutama famili Rhinolophidae dan Hipposideridae memiliki daun hidung
(noselea) yang kompleks.
Klasifikasi kelelawar menurut Corbet & Hill (1992) adalah sebagai berikut :
Kingdom : Animalia
Filum : Chordata
Sub Filum : Vertebrata
Kelas : Mamalia
Ordo : Chiroptera
Subordo : Megachiroptera
Famili : Pteropodidae
Subordo : Microchiroptera
Famili : Rhinolophidae, Hipposideridae,
Megadermatidae, Craseonycteridae,
Rhinopomatidae, Nycteridae, Emballonuridae,
Phyllostomidae, Mormoopidae, Noctilionidae,
Furipteridae, Thyropteridae, Mystacinidae,
Myzopodidae, Vespertilionidae, Molosidae dan
Natalidae
Menurut Nowak ( 1994), kelelawar ditemukan di seluruh permukaan bumi,
kecuali di daerah kutub dan pulau-pulau terpencil. Kemampuan terbang kelelawar
merupakan faktor penting dalam persebaran hewan ini. Selain itu, jenis pakannya
sangat bervariasi sehingga memungkinkan hidup di berbagai tipe habitat. Menurut
Altringham (1996), sekitar 200 spesies kelelawar ditemukan di Madagaskar dan
Afrika; 300 spesies ditemukan di Amerika Selatan dan Amerika Tengah; 240
jenis ditemukan di Asia dan Australia; dan sekitar 40 spesies ditemukan di
Amerika Utara dan Eropa. Menurut Suyanto et al. (1998), di Indonesia terdapat
151 jenis kelelawar. Jenis-jenis tersebut menyebar di seluruh kepulauan

19
Indonesia. Lebih lanjut Kunz & Pierson (1994) menjelaskan bahwa kelelawar
merupakan Mamalia paling berhasil, karena dapat ditemukan di berbagai tipe
habitat dengan ketinggian mulai 10 m dpl sampai 3000 m dpl. Winkelmann et al.
(2000) meneliti penggunaan habitat oleh kelelawar Synconycteris australis di
Papua New Guinea. Menurut Winkelmann et al. (2000) faktor-faktor yang dapat
mempengaruhi keberadaan dan kelimpahan kelelawar pada suatu habitat ialah 1)
struktur fisik habitat, 2) iklim mikro habitat, 3) ketersediaan pakan dan sumber
air, 4) keamanan dari predator, 5) kompetisi, dan 6) ketersediaan sarang.
Perilaku bersarang
Sarang merupakan salah satu komponen penting dalam hidup kelelawar.
Kebanyakan jenis kelelawar hidup berkoloni dalam bersarang dan pencarian
makan. Menurut Zukal et al. (2005) beberapa keuntungan hidup dalam koloni
adalah adanya transfer informasi, keamanan pada predator, keberhasilan
reproduksi, dan thermoregulasi. Altringham (1996) menjelaskan tiga perilaku
produk transfer informasi yang dilakukan dalam koloni kelelawar, yaitu 1)
mengikuti (following behaviour), yaitu perilaku yang menyebabkan anggota
dalam koloni bersama-sama menuju suatu lokasi tempat pencarian makan atau
tempat bersarang; 2) penanda hubungan sosial (sosial signal), yaitu pemahaman
signal-signal intensional, termasuk signal tanda bahaya; 3) belajar (learning
behaviour), yaitu proses pembelajaran dari induk ke anak yang menyebabkan
kelelawar muda mampu mengembangkan teknik pencarian makan, menghindar
dari predator, serta hal-hal yang menguntungkan bagi kehidupannya. Willis &
Brigham (2004) meneliti pembagian sarang (roost sharing) dan kebersamaan
sosial (sosial cohesion) kelelawar Eptesicus fuscus (Microchiroptera) di Cypres
Hill Canada. Hasil penelitian membuktikan bahwa interaksi sosial dan kerja sama
intraspesifik dalam koloni dapat menghasilkan ketahanan terhadap gangguan
predator dan cuaca buruk. Penelitian Baudinette et al. (1994) di Gua Kelelawar
dan Gua Robertson Australia membuktikan gua yang dihuni kelelawar dengan
jumlah besar dapat menaikkan suhu dalam gua hingga 3oC. Pada musim dingin,
keadaan ini menguntungkan kelelawar karena mengurangi energi yang diperlukan
untuk menghangatkan tubuh.

20
Setiap jenis kelelawar mempunyai beberapa alternatif dalam memilih
lokasi sarang, di antaranya adalah pohon yang tinggi, di balik batu, di atap rumah,
dan di dalam gua. Menurut Altringham (1996), pemilihan sarang mempengaruhi
distribusi lokal dan global, kepadatan, strategi pencarian makan, strategi kawin,
struktur sosial, dan pergerakan musiman. Menurut Zahn & Hager (2005) proses
yang terlibat dalam memilih tempat bersarang cukup kompleks. Ketersediaan
tempat bersarang yang cocok misalnya, akan mempengaruhi perilaku pencarian
makan, tetapi perilaku bersarang sendiri juga dipengaruhi oleh kelimpahan dan
penyebaran makanan.
Menurut Baudinette et al. (1994), Russso et al. (2003), dan Willis &
Brigham (2004), dengan memilih sarang, kelelawar dapat memperoleh beberapa
keuntungan, yaitu perlindungan dari cuaca buruk, perlindungan dari predator,
memperkecil energi termoregulasi, keberhasilan reproduksi, serta transfer
informasi tempat mencari makan dan tempat bersarang. Penelitian Willis &
Brigham (2004) dan juga penelitian Seckerdieck et al. (2005) membuktikan
bahwa kelelawar mempunyai home instink yang kuat, sarang yang dipilih
kelelawar dipertahankan sampai beberapa generasi. Namun demikian apabila
sarang mendapat ganggguan dan kelelawar tidak nyaman dan aman, sarang ini
akan ditinggalkan (Willis & Brigham 2004).
Menurut Altringham (1996), kebanyakan kelelawar pemakan buah
(Megachiroptera) bersarang di pohon dengan jumlah koloni besar. Pohon sarang
Megachiroptera biasanya tinggi dan besar, tetapi tidak berdaun rimbun. Menurut
Campbell et al. (1996), pohon tempat bersarang kelelawar biasanya menyediakan
akses yang mudah menuju tempat pencarian makan (central place foraging) dan
mempunyai pencahayaan yang cukup bagi perkembangan anakan. Penelitian
Storz et al. (2000) pada sarang kelelawar Cyanopterus sphinx (Megachiroptera) di
India Barat mendapatkan tanaman palem (Caryota urens : Palmaea) ditempati
oleh 1 individu jantan dewasa, 37 individu betina dewasa, dan 33 individu anakan.
Penelitian Soegiharto & Kartono (2009) mendapatkan kelelawar Megachiroptera:
Pteropus vampirus menempati tanaman kelapa (Cocos nucifera: Palmaea), kepuh
(Sterculia foetida: Malvaceae), dan kapuk (Ceiba pentandra: Malvaceae) di

21
Kebun Raya Bogor. Tanaman yang dipilih memiliki ketinggian yang cukup untuk
menghindar dari gangguan predator serta bertajuk relatif lebar dan mendatar.
Jenis Megachiroptera yang bersarang di gua biasanya dalam koloni kecil
atau bahkan hanya satu individu saja. Jenis-jenis tersebut adalah Rousettus
amplexicaudatus, Megaderma lyra dan Eonysteris spelaea (Suyanto 2001).
Penelitian Doyle (1979) di Gua Pondok Malaysia mendapatkan Eonycteris
spelaea dengan jumlah dua puluh individu dan Megaderma lyra hanya lima
individu bersarang dalam gua.
Sebaliknya, ordo Microchiroptera bersarang di pohon dalam jumlah sedikit.
Microchiroptera lebih menyukai bersarang di bangunan buatan manusia, di celah
batuan atau di gua dibandingkan pada dahan pohon. Penelitian Campbell et al.
(1996) di hutan Pasific Nortwest Amerika Serikat mendapatkan kelelawar
Lasionycteri noctivagans (Vespertilionidae: Microchiroptera) bersarang pada
pohon pinus (Pinus ponderosa: Pinaceae) dan pinus putih (Pinus monticola:
Pinaceae). Law & Chidel (2002) meneliti sarang dan ekologi pencarian makan
kelelawar Kerivoula papuensis (Vespertilionidae : Microchiroptera) di hutan
hujan New South Wales Australia. Sebanyak 11 individu kelelawar ditangkap di
sekitar hutan dan dipasangi radiotracking. Lima puluh empat persen (54%) di
antaranya bersarang di pohon yang jaraknya 5.2 km dari sungai, dan dua puluh
tiga persen (23%) bersarang di pohon yang jaraknya 2.7 km dari sungai, dua puluh
tiga persen (23%) bersarang dipohon yang jaraknya 2 km. Jumlah individu dalam
koloni sarang ternyata tidak lebih dari 10 individu. Tanaman yang digunakan
sebagai sarang adalah pohon jeruk (Flindersia australis: Rutaceae). Russo et al.
(2003) meneliti seleksi sarang oleh kelelawar jenis Barbastella barbastellus
(Vespertilionidae: Microchiroptera) di hutan Italia. Tanaman pada hutan yang
tidak ditebang lebih banyak dihuni kelelawar Barbastella barbastellus daripada
di hutan yang telah mengalami penebangan. Hal ini karena di hutan yang belum
ditebang lebih banyak terdapat tanaman tua (hampir mati) dengan kulit kayu
mengelupas, tinggi, dan sedikit daun.
Kebanyakan jenis Microchiroptera bersarang di gua dalam jumlah besar.
Menurut Altringham (1996) dan Zahn & Hager (2005) beberapa jenis kelelawar
memilih gua sebagai tempat bersarang karena kondisi gua yang lembap, suhu

22
stabil, dan jauh dari kebisingan. Dengan kondisi demikian, kelelawar kelompok
Microchiroptera dapat meminimalkan kekurangan air akibat evaporasi, dapat
memilih suhu yang tepat untuk tubuhnya, dan dapat menghindari kebisingan yang
dapat mengganggu bahkan dapat menyebabkan kematian. Menurut Ruczynsi et al.
(2007) kelelawar Microchiroptera memiliki alat pendengaran yang sangat
sensitif pada gelombang suara, terutama gelombang pantul (echolokasi)
berfrekuensi ultrasonik ( > 20 KHerzt).
Hasil penelitian Seckerdieck et al. (2005) membuktikan bahwa koloni
kelelawar betina Rhinolopus hipposideros (Rhinolophidae : Microchiroptera)
pada masa produktif cenderung memilih ruang bawah tanah dengan suhu rata
rata 2oC lebih dingin dan lebih stabil dibandingkan lokasi lainnya. Pada masa
hamil dan menyusui, sebenarnya kelelawar lebih membutuhkan suhu hangat.
Meskipun lebih dingin, ruang bawah tanah lebih dipilih sebagai tempat bersarang
karena mempunyai suhu stabil. Penelitian Zahn & Hager (2005) juga
menunjukkan bahwa kelelawar jenis Myotis daubentonii (Vespertilionidae:
Microchiroptera) ditemukan bereproduksi di gua-gua di Eropa Tengah yang juga
dijadikan tempat bersarang bagi kelelawar muda dan jantan dewasa. M.
daubentonii jantan biasanya menempati lokasi yang lebih dingin dibandingkan
M. daubentonii betina.
Apriandi et al. (2008) melakukan penelitian pada kelelawar penghuni gua di
Karst Cibinong. Hasil penelitan menunjukkan dalam satu gua ditemukan dua
koloni kelelawar Hipposideros larvatus (Hipposideridae: Microchiroptera)
bersarang di satu gua yang sama. Tiap jenis kelelawar memilih sarang dalam gua
dengan jarak dari mulut gua berbeda. Hasil penelitian Maryanto &
Maharadatunkamsi (1991) pada gua-gua di Pulau Sumbawa mendapatkan jenis
Rhinolophus luctus (Rhinolopodidae: Microchiroptera) menyukai tempat
bersarang di ujung gua. Dunn (1978) mendapatkan Hipposideros diadema
(Hipposideridae: Microchiroptera) dan H.armiger di atap gua pada jarak 200 kaki
dari mulut Gua Anak Takun Malaysia. Dengan memilih sarang jauh dari mulut
gua, kelelawar dapat terhindar dari gangguan manusia dan predator serta dapat
memilih mikroklimat yang stabil dan sesuai bagi tubuhnya. Tetapi, pemilihan
sarang dengan jarak jauh dari mulut gua harus didukung oleh kemampuan

23
orientasi ruang dalam keadaan gelap dan kemampuan terbang dalam ruang dengan
banyak rintangan. Penelitian Safi & Kerth (2004) pada 35 jenis kelelawar
Microchiroptera di zona temperate Eropa dan Amerika Utara menunjukkan bahwa
kelelawar yang mempunyai tulang-tulang jari (phalanges) sayap panjang hanya
mampu mengeksploitasi habitat dengan kanopi terbuka. Sebaliknya kelelawar
yang memiliki tulang jari sayap pendek, lebih mampu mengeksploitasi habitat
berkelok-kelok dan banyak rintangan. Oleh karena itu, kelelawar yang mampu
bersarang pada lokasi jauh dari mulut gua kemungkinan adalah kelelawar dengan
tulang jari sayap pendek.
Adaptasi struktural dan fisiologi pernapasan
Kondisi ruang gua yang sempit, sirkulasi udara terbatas, dan banyak dihuni
kelelawar menyebabkan udara dalam gua menjadi rendah oksigen (hypoxic),
tinggi karbon dioksida (hypercapnic), dan tinggi gas amonia (Baudinette et al.
1994). Keadaan ini kurang menguntungkan bagi hewan gua karena: 1)
kurangnya oksigen dapat menyebabkan respirasi terhambat (Guyton 1995); 2)
tingginya karbon dioksida dapat menyebabkan afinitas hemoglobin pada oksigen
menurun (Guyton 1995); dan 3) tingginya konsentrasi gas amonia (NH3) dapat
menyebabkan gangguan metabolisme, iritasi epitel organ pernapasan serta
gangguan fisiologi saraf (Hutabarat et al. 2000). Oleh karena itu, agar dapat
bertahan hidup dalam gua, kelelawar harus beradaptasi pada keadaan tersebut.
Hal ini sesuai dengan pendapat Alikodra (2010) bahwa untuk bertahan hidup dan
berkembang dalam suatu habitat, hewan harus mengembangkan strategi, di
antaranya strategi adaptasi pada habitat.
Penelitian Baudinette et al. (1994) di Afrika Selatan membuktikan bahwa
kelelawar Miniopterus schreibersii (Vespertilionidae: Microchiroptera) dapat
hidup pada gua dengan kondisi udara rendah oksigen (hypoxic) dan tinggi karbon
dioksida (hypercapnic). Jumlah populasi kelelawar yang besar pada gua tersebut
menyebabkan oksigen yang digunakan untuk respirasi lebih besar dibandingkan
oksigen yang masuk ke dalam gua. Sebaliknya karbon diokasida yang dihasilkan
dari proses respirasi menambah jumlah karbon dioksida yang terperangkap di
dalam gua (Baudinette et al. 1994).

24
Menurut Suyanto (2001), kelelawar juga dapat bertahan hidup pada gua
dengan kandungan amonia tinggi. Penelitian Sridhar et al. (2006) mendapatkan
urin dan feses (guano) kelelawar Hipposideros speoris (Hipposideridae:
Microchiroptera) tersusun atas 5.7 ± 1.5% nitrogen (N) berbentuk amonia (NH3).
Amonia tersebut merupakan hasil katabolisme protein. Amonia dalam guano
dapat menguap menjadi gas bercampur dengan komponen udara lainnya. Hal ini
menyebabkan kandungan amonia udara meningkat tajam (Shidar et al. 2006).
Hutabarat (2000) melakukan penelitian pada karyawan pabrik lateks yang terkena
paparan amonia sebesar 500 ppm sampai 600 ppm selama 60 hari. Hasil
penelitian menunjukkan karyawan yang terkena paparan amonia mengalami
gejala sebagai berikut : tenggorokan kering (80%); jalan pernapasan kering
(73.3%); mata perih (66.67%); batuk (53.3%); dan pingsan (6.67%). Menurut
Suyanto (2001) kelelawar gua dapat bertahan pada kandungan amonia udara
mencapai 5000 ppm, sedangkan manusia hanya mampu bertahan pada kandungan
amonia udara maksimum sebesar 100 ppm.
Penelitian tentang strategi adaptasi kelelawar yang bersarang di gua dengan
kondisi dingin dan lembap pernah dilakukan oleh Baudinette et al. (2000) di
Australia. Hasil penelitian menunjukkan laju respirasi kelelawar Macroderma
gigas (Megadermatidae: Microchiroptera) dan Rhinonycteris aurantias
(Hipposideridae: Microchiroptera) menyesuaikan dengan suhu dan kelembapan
udara dalam gua. Pada saat kondisi udara kering dan dingin (kelembapan <60%
dan suhu < 5.6oC) laju respirasi sama dengan pada saat kondisi udara lembap dan
hangat ( kelembapan > 80% ;suhu > 9.8oC). Tetapi bila kondisi udara lembap
dan dingin (kelembapan < 60% dan suhu < 9o) laju respirasinya meningkat
tajam. Meningkatnya laju respirasi tersebut merupakan strategi agar tubuh tetap
hangat. Namun demikian, sejauh ini belum ada penelitian mengenai strategi
adaptasi fisiologi dan anatomi pernapasan kelelawar untuk bertahan hidup pada
kondisi hypoxic, hypercapnic, dan tinggi amonia.
Menurut Frandson (1992) organ respirasi Mamalia terdiri atas: 1) lubang
hidung (nasale), berfungsi menyaring, menghangatkan, dan melembapkan udara
yang masuk ke dalam organ pernapasan; 2) laring, berfungsi melindungi jalan
napas bawah dari obstruksi (masuknya) benda asing; 3) trakhea, berfungsi
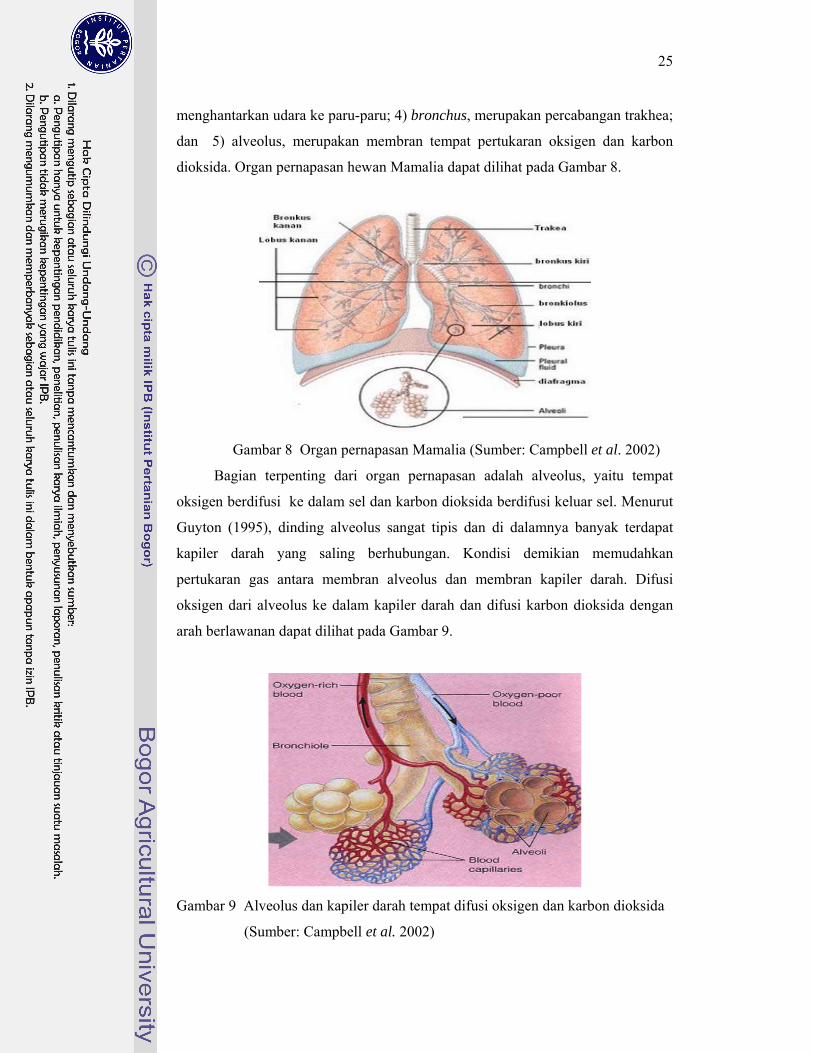
25
menghantarkan udara ke paru-paru; 4) bronchus, merupakan percabangan trakhea;
dan 5) alveolus, merupakan membran tempat pertukaran oksigen dan karbon
dioksida. Organ pernapasan hewan Mamalia dapat dilihat pada Gambar 8.
Gambar 8 Organ pernapasan Mamalia (Sumber: Campbell et al. 2002)
Bagian terpenting dari organ pernapasan adalah alveolus, yaitu tempat
oksigen berdifusi ke dalam sel dan karbon dioksida berdifusi keluar sel. Menurut
Guyton (1995), dinding alveolus sangat tipis dan di dalamnya banyak terdapat
kapiler darah yang saling berhubungan. Kondisi demikian memudahkan
pertukaran gas antara membran alveolus dan membran kapiler darah. Difusi
oksigen dari alveolus ke dalam kapiler darah dan difusi karbon dioksida dengan
arah berlawanan dapat dilihat pada Gambar 9.
Gambar 9 Alveolus dan kapiler darah tempat difusi oksigen dan karbon dioksida
(Sumber: Campbell et al. 2002)

26
Menurut Guyton (1995) pertukaran gas respirasi melewati membran
alveolus berlangsung sangat efektif. Hal ini karena permukaan membran alveolus
sangat tipis. Selain itu, dengan bentuk seperti gelembung-gelembung udara, luas
permukaan membran menjadi sangat besar. Hasil penelitian Setiadi (2000)
menunjukkan kelelawar Scotophilus kuhli (Vespertilionidae: Microchiroptera)
memiliki alveolus lebih kecil dan rapat dibandingkan alveolus tikus (Mus
musculus). Menurut Plopper & Adams (1993), alveolus beberapa Mamalia
berukuran kecil dan rapat sehingga permukaan respirasinya lebih luas. Permukaan
respirasi yang luas menyebabkan paru-paru mampu mengikat oksigen lebih
banyak (Plopper & Adams 1993). Menurut Guyton (1995), luas permukaan
membran alveolus ini dapat berkurang karena rusaknya dinding alveolus.
Kerusakan tersebut di antaranya adalah karena emfisema, yaitu keadaan alveolus
bersatu disebabkan terpapar gas racun atau karena infeksi kuman.
Setelah oksigen berdifusi dari alveolus ke kapiler darah dan karbon
dioksida berdifusi dari kapiler darah ke alveolus, selanjutnya oksigen akan
diangkut oleh darah ke seluruh jaringan tubuh oleh sel darah merah (eritrosit).
Karena adanya unsur hemoglobin dalam eritrosit, maka eritrosit dapat mengikat
oksigen dan karbon dioksida. Menurut Ganong (2001), eritrosit Mamalia
berbentuk cakram bikonkaf. Cakram bikonkaf tersebut memiliki permukaan yang
relatif luas untuk pertukaran oksigen.
Menurut Ganong (2001) jumlah eritrosit juga berbeda antara satu individu
dengan individu lainnya. Faktor yang mempengaruhi jumlah eritrosit hewan
adalah kondisi nutrisi, aktivitas fisik, umur, dan lingkungan. Selain itu, menurut
Ganong (2001), penurunan kadar oksigen (hipoksia) akan merangsang ginjal
untuk melepaskan enzim eritrogenin (erythrogenin). Selanjutnya, enzim tersebut
akan mengaktifkan eritropoietinogen yang merupakan prekusor dalam
pembentukan eritopoietin. Di dalam sumsum tulang, eritropoietin akan
meningkatkan jumlah sel bakal (stem cell). Sel bakal ini akan menjadi prekusor
darah merah dan selanjutnya menjadi sel darah merah (Ganong 2001). Menurut
Guyton (1995), binatang yang terpapar oksigen rendah selama bertahun tahun
menunjukkan peningkatan jaringan hipoksik dan peningkatan jumlah sel darah
merah sampai 20%.

27
Kemampuan sel darah merah mengikat oksigen disebabkan adanya
hemoglobin. Hemoglobin merupakan molekul protein yang berikatan dengan
porphyrin. Di bagian tengah molekul porphyrin tersebut terdapat satu atom besi
(Fe). Menurut Ganong (2001) hemoglobin Mamalia tersusun atas empat subunit
protein berbentuk globul (bola). Satu subunit dapat membawa satu molekul
oksigen, dengan demikian setiap molekul hemoglobin dapat membawa empat
molekul oksigen. Struktur hemoglobin dapat dilihat pada Gambar 10.
Gambar 10 Struktur kimia hemoglobin (Sumber: Campbell et al. 2002)
Menurut Ganong (2001) hemoglobin yang mengikat oksigen disebut
oksihemoglobin. Afinitas (daya ikat) hemoglobin pada oksigen dipengaruhi oleh
suhu tubuh, keasaman (pH) darah, dan konsentrasi bahan-bahan kimia dalam
darah. Pada keadaan suhu udara rendah, tekanan oksigen tinggi dan keasaman
darah tinggi, afinitas hemoglobin pada oksigen meningkat. Sebaliknya pada
kondisi tekanan oksigen rendah, suhu tinggi, dan keasaman rendah afinitas
hemoglobin pada oksigen menurun (Ganong 2001). Menurut Guyton (1995),
dalam keadaan normal, 100 ml darah manusia mengandung 15 gram hemoglobin
yang mampu mengangkut 0.03 gram oksigen. Penelitian Fatmawati (2007),
mendapatkan kandungan hemoglobin anjing (Canis familiaris) 11.8 ± 1.13 g/mm3.
Menurut Frandson (1992) jumlah hemoglobin hewan Mamalia berada pada
kisaran 12 – 18 g/mm3.

28
Kesamaan relung (niche overlap) pakan Kelelawar dari jenis (species) berbeda dapat memanfaatkan gua yang sama
sebagai sarang. Hal ini dibuktikan oleh: penelitian Dunn (1978) yang
mendapatkan jenis Hipposideros armiger, H. cineraceus, Rhinolophus affinis dan
R. malayanus di Gua Anak Takun Malaysia; penelitian Zukal et al. (2005) yang
mendapatkan jenis Myotis myotis dan Rhinolophus hipposideros di Gua
Katerinska Cekoslvakia; dan penelitian Apriandi et al. (2008) yang mendapatkan
jenis Miniopterus australis, Myotis adversus dan Rhinolophus affinis di Gua
Gudawang Bogor. Menurut Kunz (1982) dan Willis & Brigmann (2004) sarang
yang dipilih kelelawar memiliki akses yang mudah pada sumber pakan. Oleh
karena itu, apabila jenis-jenis kelelawar yang bersarang dalam satu gua tersebut
bergantung pada sumber pakan yang sama, akan terjadi kompetisi, terutama bila
ketersediaan sumber pakan terbatas. Sebaliknya, bila sumber pakan berbeda,
kompetisi tidak terjadi.
Menurut Cox (2002), penggunaan relung yang sama (niche overlap)
menyebabkan interaksi kompetitif, yaitu tiap populasi yang berkompetisi
memberikan pengaruh yang merugikan bagi pesaingnya (competitor). Menurut
Reynold & Ludwig (1984) nilai niche overlap berkisar antara nol (0) sampai
dengan satu (1). Apabila nilai niche overlap pakan mendekati satu berarti kedua
jenis hewan tersebut memiliki pakan yang sama dan berpotensi untuk
berkompetisi.
Kelelawar anggota subordo Megachiroptera adalah pemakan buah
(frugivora) ataupun serbuk sari (polinator), sedangkan anggota subordo
Microchiroptera kebanyakan pemakan serangga (insektivora). Menurut
Altringham (1996), berdasarkan strategi pencarian makannya, kelelawar
dibedakan menjadi tipe spesialis (selektif) dan opportunis (generalis). Kelelawar
tipe spesialis hanya memakan jenis tertentu. Tipe ini bisa menghabiskan banyak
waktu dan energi dalam pencarian makan, tetapi makanan yang didapatkan
memiliki profit (nilai gizi) tinggi. Tipe opportunis menghabiskan lebih sedikit
waktu dan energi dalam pencarian makannya, tetapi makanan yang didapatkan
mungkin lebih sedikit nilai gizinya dibandingkan kelelawar tipe spesialis.
Beberapa Megachiroptera, misalnya Rousettus amplexicaudatus (Pteropodidae:

29
Makrochiroptera) adalah pemakan buah tipe oportunis, sedangkan Macroglosus
sabrinus (Pteropodidae: Makrochiroptera) adalah tipe spesialis. Menurut Nowak
(1994) R. amplexicaudatus memakan jambu biji (Psidium guajava), pisang (Musa
paradisiaca); sawo (Manilkara kauki), dan buah masak lainnya. Sedangkan
Macroglosus sabrinus terspesialissasi untuk memakan nectar bunga durian
(Durio zibethinus) dan bunga patai (Parkis speciosa) (Nowak 1994).
Microchiroptera adalah pemakan serangga dengan tipe spesialis ataupun
oportunis. Hal ini dibuktikan dengan hasil penelitian Razakarivony et al. (2005)
yang meneliti makanan kelelawar jenis Myzopoda aurita (Myzopodidae:
Microchiroptera) di Madagaskar. Pada saat ngengat (Lepidoptera) melimpah,
prosentase ngengat dalam feses juga meningkat. Hasil penelitian ini
membuktikan bahwa kelelawar ini memanfaatkan kesempatan (oportunis) dalam
memilih jenis makanannya.

30
BIODIVERSITAS DAN POLA PEMILIHAN SARANG
KELELAWAR: STUDI KASUS DI KAWASAN
KARST GOMBONG KABUPATEN KEBUMEN
JAWA TENGAH
ABSTRACT
The existence of cave dwelling bats of karst area need to be conserved, because bats have important roles for the ecosystem inside as well as outside the cave. The objectives of this research were to know the biodiversity of cave dwelling bats; the physical factors influencing the community structure of the bats and the physical factors influencing the preference roosting place. This study was conducted from September 2008 to March 2009 in twelve caves in Gombong Karst Area, Central Java. The mapping of the roosting place was carried out using forward method. The sample of the bats were picked up at the roosting place during the day. The physical and microclimate parameters were measured under the bat roosts, three times in February, June and October 2009. The data were analyzed by ANOVA, Redundancy analysis (RDA) and canonical correspondence analysis (CCA). The result showed: 1) Fifteen species (eleven species of Microchiroptera and four spesies of Megachiroptera) indicated known in this research. 2) The length, height, and width of the cave corridor influenced the community structure of the bats; 3) The microclimate factors which influenced the roosting place preference were the sound intensity, the distance from the cave entrance, the temperature, the humidity, and the light intensity. Based on the factors mentioned, there were five goup of bats, each of which has specific patterns of roosting preference. Key words: cave, bat, roosting preference, Gombong karst.
Pendahuluan
Kawasan Karst Gombong terletak di Jawa Tengah bagian selatan, tepatnya
pada 7°27'-7°50' LS dan 109°22' - 109°50' BT. Menurut Whitten et al. (1999),
Paparan Sunda awalnya merupakan lautan tropik dangkal yang dasarnya banyak
mengendap kalsium karbonat yang dihasilkan oleh binatang berkerangka kapur
dan foraminifera. Dasar laut tersebut terdorong ke atas oleh gaya tektonik,
akibatnya terbentuk barisan bukit karst. Hal ini diperkuat oleh pernyataan Pusat
Survei Geologi Badan Geologi Nasional (2006), bahwa Karst Gombong berada di
atas batuan gamping yang mengandung fosil biota laut: Radiolaria, Hedbergella,
Ratalipora dan Bolivisoides cf. exculpta. Pelarutan batu gamping ini berlanjut

31
dengan proses abrasi sehingga menghasilkan gua yang bercabang-cabang, gelap,
lembap, temperatur stabil, dan sirkulasi udara terbatas.
Di kawasan Karst Gombong terdapat sekitar 112 gua karst, dan lebih dari 60
gua di antaranya dihuni kelelawar (DISPARHUB Kebumen 2004). Menurut
Boudinette et al. (1994), Duran & Centano (2002), dan Twente (2004), dinding
dan atap gua membentuk fisik dan mikroklimat yang berbeda antara satu gua
dengan gua lainnya. Perbedaan fisik dan mikroklimat tersebut menyebabkan
setiap gua membentuk ekosistem yang unik dan dihuni oleh keanekaragaman jenis
fauna yang khas. Berdasarkan hasil-hasil penelitian tersebut diduga setiap gua di
kawasan Karst Gombong akan memiliki keanekaragaman jenis kelelawar berbeda.
Namun, belum ada hasil penelitan yang menunjukkan ke arah itu.
Hasil-hasil penelitian mengenai kelelawar di luar Indonesia membuktikan
bahwa tiap jenis kelelawar cenderung memilih sarang di dalam gua dengan
kondisi yang sesuai bagi kebutuhan tubuhnya. Penelitian Zahn & Hager (2005)
mendapatkan Myotis daubentonii jantan menempati lokasi yang lebih dingin
dibandingkan M. daubentonii betina di gua-gua di Eropa Tengah. Penelitian
Duran & Centano (2002) membuktikan kelelawar Pteronotus quadridens
bersarang di gua dengan suhu 28oC s/d 35oC, dan Erophylla sezekorni bersarang
di gua dengan suhu 25 oC s/d 28 oC. Di samping itu, diduga terdapat faktor-faktor
fisik dan mikroklimat lain selain suhu yang mempengaruhi pemilihan sarang
kelelawar di gua-gua Karst Gombong.
Penelitian ini bertujuan untuk: 1) Mengkaji biodiversitas kelelawar
penghuni gua berdasarkan faktor-faktor fisik gua di Karst Gombong; 2)
Mengidentifikasi faktor mikroklimat gua yang berpengaruh terhadap pola
pemilihan sarang kelelawar gua. Hipotesis penelitian ini adalah : 1) )
Keanekaragaman, kelimpahan, dan kemerataan jenis kelelawar dipengaruhi secara
nyata oleh panjang lorong, tinggi lorong, serta jumlah pintu, dan jumlah ventilasi
gua; 2) Pola pemilihan sarang kelelawar gua dipengaruhi oleh jarak dari pintu
gua, tinggi atap gua, suhu, kelembapan, kecepatan angin, intensitas cahaya,
intensitas suara, kadar oksigen, dan kadar amonia di sekitar sarang.
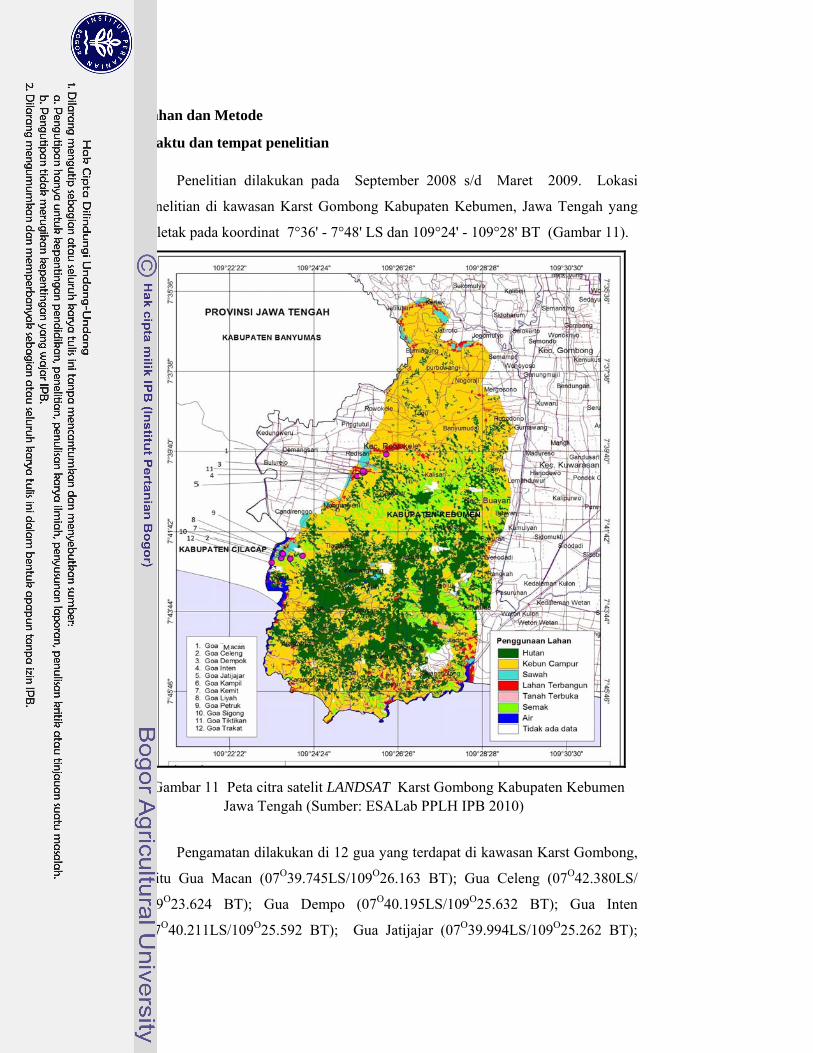
32
Bahan dan Metode
Waktu dan tempat penelitian Penelitian dilakukan pada September 2008 s/d Maret 2009. Lokasi
penelitian di kawasan Karst Gombong Kabupaten Kebumen, Jawa Tengah yang
terletak pada koordinat 7°36' - 7°48' LS dan 109°24' - 109°28' BT (Gambar 11).
Gambar 11 Peta citra satelit LANDSAT Karst Gombong Kabupaten Kebumen Jawa Tengah (Sumber: ESALab PPLH IPB 2010)
Pengamatan dilakukan di 12 gua yang terdapat di kawasan Karst Gombong,
yaitu Gua Macan (07O39.745LS/109O26.163 BT); Gua Celeng (07O42.380LS/
109O23.624 BT); Gua Dempo (07O40.195LS/109O25.632 BT); Gua Inten
(07O40.211LS/109O25.592 BT); Gua Jatijajar (07O39.994LS/109O25.262 BT);

33
Gua Kampil (07O42.389LS/109O23.836 BT); Gua Kemit (07O42.247LS/
109O23.638 BT); Gua Liyah (07O42.392LS/109O23.838 BT); Gua Petruk
(07O42.315LS/109O24.130BT); Gua Sigong (07O42.487LS/109O23.389 BT); Gua
Tiktikan (07O40.166LS/109O25.595BT); dan Gua Tratag (07O42.267LS/
109O23.66BT). Identifikasi sampel dilakukan di Laboratorium Mamalia PUSLIT
Biologi LIPI Cibinong.
Bahan dan alat
Bahan yang digunakan adalah chloroform dan alkohol 70%. Alat yang
digunakan adalah: Global Positioning System (GPS) merk Garmin; altimeter merk
Krisbow, kompas merk Sunto; pita ukur (50 m); mist net; hand net; harp trap;
kantong spesimen; bambu; timbangan digital; jangka sorong; lux meter;
anemometer; sound level meter; digital thermometer; digital hygrometer; oksigen
meter; generator; pompa vacum; dan air quality checker (Gambar 12).
Gambar 12 Alat pemetaan roosting kelelawar dan pengukuran mikroklimat gua
Cara kerja
Sebelum menentukan gua pengamatan, dilakukan survei pendahuluan
dengan tujuan mencari informasi mengenai kondisi umum, sebaran gua, jumlah
gua, dan tipe gua di kawasan Karst Gombong. Survei pendahuluan dilakukan
dengan cara menjelajah kawasan Karst Gombong dan mencari data sekunder ke
PEMDA Kabupaten Kebumen; Dinas Pariwisata dan Perhubungan Kabupaten

34
Kebumen; serta ke Badan Geologi Nasional di Bandung. Penentuan gua
pengamatan dilakukan dengan metode purposive random sampling, yaitu gua
pengamatan ditentukan sebanyak 10% dari seluruh gua yang ada. Berdasarkan
survei pendahuluan diketahui bahwa di kawasan Karst Gombong terdapat 112
gua. Maka ditentukan sekitar 10% atau dua belas gua sebagai objek penelitian.
Dua belas gua tersebut dibagi menjadi empat kelompok berdasarkan panjang
lorong gua, yaitu lorong gua pendek (kurang dari 100 m) sebanyak tiga gua (Gua
Tiktikan, Gua Tratag, Gua Sigong); lorong gua sedang (antara 100 m s/d 200 m)
sebanyak tiga gua (Gua Macan, Gua Dempo, Gua Kampil); lorong gua panjang
(antara 200 m s/d 350 m) sebanyak tiga gua (Gua Inten, Gua Kemit, Gua
Jatijajar), dan lorong gua sangat panjang (lebih dari 350 m) sebanyak tiga gua
(Gua Petruk, Gua Celeng, Gua Liyah).
Titik koordinat setiap gua ditentukan dengan menggunakan GPS.
Permeating lokasi sarang kelelawar dilakukan dengan menelusuri semua lorong
gua, mulai dari mulut/pintu gua (entrance) sampai ujung gua (duck). Metode
pemetaan yang digunakan adalah metode foreward (HIKESPI 2004) yang
dilakukan dengan cara sebagai berikut: orang pertama berdiri di titik pertama
(mulut gua) dan orang kedua di titik kedua (belokan/simpangan); setelah
pembacaan alat selesai, orang pertama berdiri di titik kedua, orang kedua di titik
ketiga (belokan/simpangan berikutnya). Begitu seterusnya sampai titik terakhir di
ujung gua (Gambar 13). Berdasarkan kriteria BCRA (British Cave Researche
Association) grade pemetaan yang dihasilkan dengan metode ini adalah grade III
(Subterra 2004). Hal ini karena alat yang digunakan terbatas pada kompas dan pita
ukur, dimana derajat kesalahan pengukuran sudut adalah ± 2.5o, dan derajat
kesalahan pengukuran jarak ± 50 cm (Subterra 2004).

35
Gambar 13 Pemetaan gua dengan metode foreward (Hikespi 2004)
Parameter fisik gua yang diukur adalah: panjang lorong gua, lebar lorong
gua, tinggi lorong gua, jumlah mulut gua dan jumlah ventilasi gua. Panjang
lorong gua diukur mulai dari mulut gua sampai ujung gua dengan menggunakan
pita meter. Bila terdapat percabangan lorong gua, semua percabangan tersebut
juga diukur dan hasil pengukurannya dijumlahkan. Lebar lorong gua diukur
dengan cara sebagai berikut: ditentukan lima lokasi di dalam lorong gua secara
acak, kelima lokasi tersebut diukur lebarnya (tegak lurus dari satu dinding gua ke
dinding lain yang berseberangan) menggunakan pita meter, kemudian dihitung
rata-ratanya. Tinggi lorong gua juga diukur pada lima lokasi yang dipilih secara
acak dan dihitung rata-ratanya. Cara yang dilakukan bergantung pada kondisi
gua. Cara tersebut adalah: 1) menggunakan tali rafia yang diikat dengan batu,
batu dilempar tegak lurus hingga menyentuh atap gua, panjang tali rafia yang
terbawa lemparan batu diukur (cara ini dilakukan pada gua yang mempunyai atap
tinggi dan sedikit stalaktit); 2) menggunakan 3 potongan bambu yang disambung
fleksibel (cara ini dilakukan pada gua yang mempunyai atap tidak terlalu tinggi).
Mengingat tingkat kesulitan yang cukup tinggi, pengukuran fisik gua hanya
dilakukan satu kali, yaitu pada bulan Februari sampai Maret 2009.
Pengukuran parameter fisik mikroklimat sarang dilakukan di bawah
sarang kelelawar pada jarak terdekat dari sarang yang mungkin terjangkau.
Pengukuran kadar amonia udara dilakukan dengan cara: udara di bawah sarang
kelelawar dihisap menggunakan pipa vacum dan dialirkan ke pingel yang telah
Step III
Step II
Step I

36
berisi absorban NH3 selama 120 menit (Gambar 14). Larutan absorban yang telah
bercampur dengan NH3 dari udara dipindahkan ke dalam tabung yang tertutup
rapat. Pembacaan konsentrasi NH3 dilakukan di laboratorium dengan
menggunakan spektrofotometer. Pengukuran parameter lingkungan sarang
kelelawar diulang sebanyak 3 kali pada bulan yang berbeda (Februari; Juni;
Oktober 2009). Kecuali pengukuran amonia udara hanya dilakukan satu kali
(Maret 2010).
Gambar 14 Pengukuran kadar amonia udara
Estimasi jumlah kelelawar di setiap sarang dilakukan dengan menghitung
langsung (direct count) kelelawar di sarangnya. Penghitungan dilakukan pada
siang hari (10.00 WIB s/d 15.00 WIB) saat kelelawar bersarang di dalam gua
dengan cara (Saroni 2005) : 1) Diukur luas sarang dengan membuat proyeksi
sarang ke lantai gua; 2) Tiap satu sarang dibuat tiga kuadrat secara acak masing-
masing berukuran 1 meter persegi; 3) Pada setiap kuadrat dihitung jumlah
kelelawar; 4) Jumlah kelelawar tiap sarang adalah luas sarang dikalikan jumlah
kelelawar rata- rata pada setiap kuadrat (Gambar 15).
.
Gambar 15 Metode penghitungan populasi kelelawar pada tiap sarang
kuadrat I
kuadrat III
1m
kuadrat II

37
Pengambilan sampel kelelawar dilakukan pada setiap sarang. Cara yang
dilakukan bergantung pada kondisi gua (formasi gua, keberadaan stalaktit dan
tinggi atap gua). Cara tersebut adalah : 1) menggunakan hand net, apabila formasi
gua sederhana dan sarang terjangkau hand net; 2) menggunakan mist net yang
dipasang di sekitar mulut gua, apabila sarang pada posisi tinggi, dan lorong gua
lebar; 3) menggunakan harpa trap yang dipasang di mulut gua, apabila sarang
berada pada posisi tinggi dan lorong gua sempit (Gambar 16).
a) hand net b) mist net c) harpa trap
Gambar 16 Pengambilan sampel kelelawar
Jumlah minimal sampel kelelawar yang diambil pada setiap sarang adalah
lima ekor. Sampel kelelawar yang tertangkap dimasukkan ke dalam kantong
blacu, kemudian dilakukan pengukuran morfometri untuk identifikasi awal.
Setelah itu sampel dibius dengan menggunakan chloroform dan direndam dalam
alkohol 70%. Identifikasi sampel dilakukan di laboratoriun Mamalia LIPI
Cibinong dengan menggunakan buku kunci identifikasi Mamalia: The Mammals
of the Indomalayan Region: a Systematic Review (Corbet & Hill 1992).
Analisis data
Kelimpahan populasi kelelawar pada setiap sarang dihitung dengan cara
pendugaan berdasarkan luas sarang, dengan rumus (Saroni 2005):
P= D x L
Keterangan : P= kelimpahan populasi kelelawar (individu) D= kepadatan ( individu/meter2) L= luas hunian (meter2 ).

38
Struktur komunitas yang diamati meliputi: kelimpahan, indeks
keanekaragaman jenis (H’), kekayaan jenis (S), dan indeks kemerataan jenis (E).
Untuk menentukan indeks keanekaragaman jenis kelelawar pada setiap gua
digunakan rumus indeks keanekaragaman (H’) Shannon & Wiener (Magurran
2004) sebagai berikut:
H’= - ∑ ( ni/N) ln (ni/N)
Keterangan :
H’ = Indeks keanekaragaman Shannon-Wiener ni = Jumlah individu jenis ke i N = Jumlah total individu Indeks keanekaragaman jenis kelelawar ini dihitung menggunakan software
ecological methodology versi 5.1.
Indeks kemerataan jenis kelelawar dianalisis dengan digunakan indeks
kemerataan Simpson (E), dengan rumus sebagai berikut (Magurran 2004):
E = H’/ ln S
Keterangan : E = indeks kemerataan H’ = indeks keanekaragaman S = jumlah spesies
Indeks kemerataan jenis kelelawar dihitung menggunakan software ecological
methodology versi 5.1.
Kecenderungan hubungan antara parameter fisik gua dengan struktur
komunitas kelelawar dianalisis dengan analisis multivariat RDA (Redundancy
analysis). RDA merupakan penjabaran dari regesi linear berganda memakai
model linear dengan variabel X dan Y. Adapun parameter fisik gua yang
dianalisis adalah panjang lorong gua, tinggi lorong gua, lebar lorong gua, jumlah
ventilasi, dan jumlah pintu gua. RDA dihitung menggunakan software Canoco for
windows 4.5 (Leps & Smilauer 1999; Koneri 2007). Untuk mengetahui nilai
korelasi faktor fisik gua dengan struktur komunitas kelelawar dilakukan uji
korelasi Spearmen dengan taraf kepercayaan 95% menggunakan software SPSS
versi 15.
Pengaruh mikroklimat gua terhadap pemilihan sarang kelelawar, dianalisis
dengan analisis multivariat CCA (canonical correspondence analysis)
menggunakan software canoco for windows 4.5. Penggunaan metode CCA ini

39
bertujuan untuk menentukan hubungan dalam bentuk grafik serta mengungkap
informasi maksimum dari suatu matriks data dengan faktor lingkungan secara
bersamaan. Matriks data tersebut terdiri atas jenis kelelawar dan faktor
lingkungan yang terdiri atas 9 parameter yaitu; jarak dari mulut gua, tinggi
sarang, suhu, kelembapan, intensitas cahaya, intensitas suara, kecepatan angin,
persentasi oksigen udara, dan kadar amonia udara. Untuk mengurutkan
mikroklimat yang paling berpengaruh terhadap pemilihan sarang kelelawar
digunakan RDA (Redundancy analysis) dengan metode forward selection dan
diuji menggunakan monte carlo permutation dengan 199 permutasi acak (Leps &
Smilauer 1999; Koneri 2007).
Hasil
Biodiversitas dan sebaran sarang kelelawar
Hasil penelusuran gua menunjukkan bahwa dari dua belas gua yang diteliti,
sepuluh gua dihuni kelelawar, dan dua gua tidak dihuni kelelawar. Jenis-jenis
kelelawar yang bersarang pada gua-gua tersebut terdiri atas empat jenis kelelawar
Megachiroptera dan sebelas jenis kelelawar Microchiroptera. Kelelawar
Megachiroptera terdiri atas: Cynopterus horsfieldii Gay, 1843 Cynopterus
brachyotis (Muller, 1838); Rousettus amplexicaudatus (Geoffroy, 1810); dan
Eonycteris spelaea (Dobson,1871). Kelelawar Microchiroptera terdiri atas
(Chaerophon plicata (Buchannan,1800), Hipposideros ater Templeton, 1848;
Hipposideros cf.ater; Hipposideros sp; Hipposideros bicolor (Temminck, 1834);
Hipposideros sorenseni Kitchener & Maryanto, 1993; Hipposideros diadema
(Geoffroy,1813); Rhinilophus borneensis Peters, 1861; Rhinolophus affinis
Horsfield, 1823; Miniopterus australis Tomes, 1858; dan Miniopterus
schreibersii (Kuhl, 1819). Gambar jenis-jenis kelelawar yang ditemukan dapat
dilihat pada Lampiran 1.
Gua Petruk paling banyak dihuni oleh kelelawar yaitu sembilan jenis,
diikuti oleh Gua Celeng (4 jenis), Gua Dempo, Gua Liyah, Gua Inten, Gua Kemit,
Gua Jatijajar (masing-masing 3 jenis), Gua Macan (2 jenis), Gua Sigong dan Gua
Tratag (1 jenis). Gua Tiktikan dan Gua Kampil tidak dihuni kelelewar. Kondisi
fisik gua dan struktur komunitas kelelawar yang ditemukan di gua-gua Karst
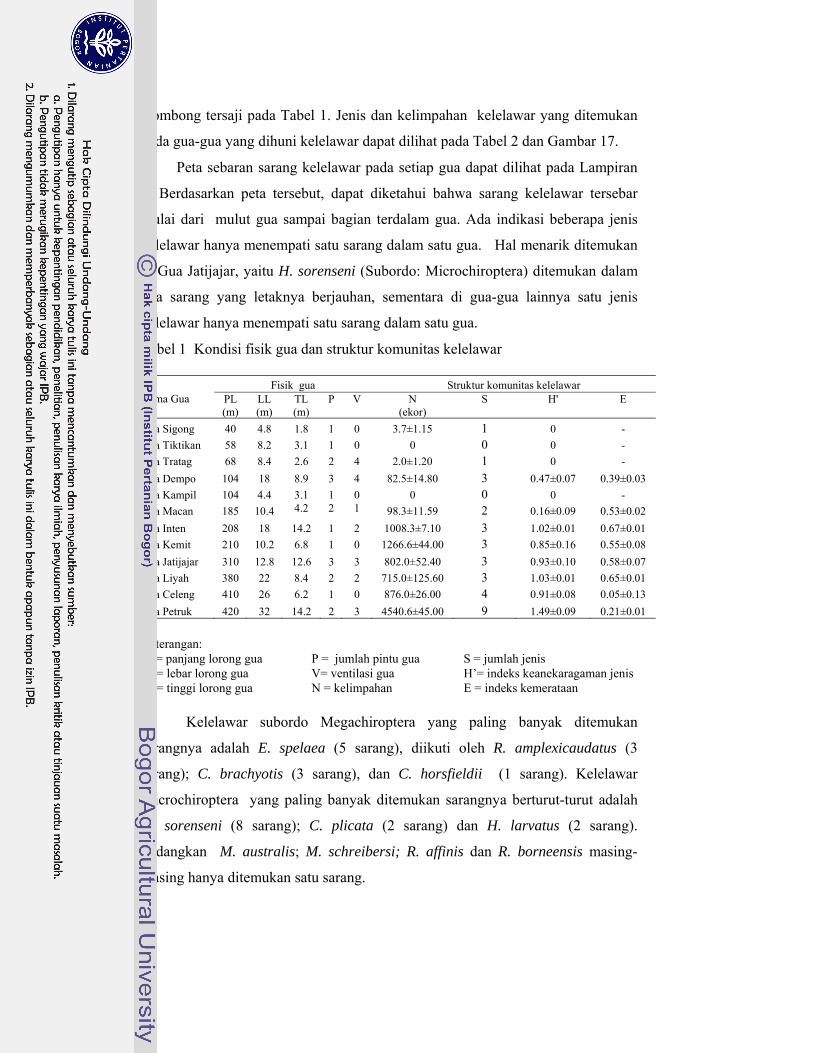
40
Gombong tersaji pada Tabel 1. Jenis dan kelimpahan kelelawar yang ditemukan
pada gua-gua yang dihuni kelelawar dapat dilihat pada Tabel 2 dan Gambar 17.
Peta sebaran sarang kelelawar pada setiap gua dapat dilihat pada Lampiran
2. Berdasarkan peta tersebut, dapat diketahui bahwa sarang kelelawar tersebar
mulai dari mulut gua sampai bagian terdalam gua. Ada indikasi beberapa jenis
kelelawar hanya menempati satu sarang dalam satu gua. Hal menarik ditemukan
di Gua Jatijajar, yaitu H. sorenseni (Subordo: Microchiroptera) ditemukan dalam
dua sarang yang letaknya berjauhan, sementara di gua-gua lainnya satu jenis
kelelawar hanya menempati satu sarang dalam satu gua.
Tabel 1 Kondisi fisik gua dan struktur komunitas kelelawar
Nama Gua
Fisik gua Struktur komunitas kelelawar PL (m)
LL (m)
TL (m)
P
V
N (ekor)
S
H'
E
Gua Sigong 40 4.8 1.8 1 0 3.7±1.15 1 0 - Gua Tiktikan 58 8.2 3.1 1 0 0 0 0 - Gua Tratag 68 8.4 2.6 2 4 2.0±1.20 1 0 - Gua Dempo 104 18 8.9 3 4 82.5±14.80 3 0.47±0.07 0.39±0.03 Gua Kampil 104 4.4 3.1 1 0 0 0 0 - Gua Macan 185 10.4 4.2 2 1 98.3±11.59 2 0.16±0.09 0.53±0.02 Gua Inten 208 18 14.2 1 2 1008.3±7.10 3 1.02±0.01 0.67±0.01 Gua Kemit 210 10.2 6.8 1 0 1266.6±44.00 3 0.85±0.16 0.55±0.08 Gua Jatijajar 310 12.8 12.6 3 3 802.0±52.40 3 0.93±0.10 0.58±0.07 Gua Liyah 380 22 8.4 2 2 715.0±125.60 3 1.03±0.01 0.65±0.01 Gua Celeng 410 26 6.2 1 0 876.0±26.00 4 0.91±0.08 0.05±0.13 Gua Petruk 420 32 14.2 2 3 4540.6±45.00 9 1.49±0.09 0.21±0.01 Keterangan: PL= panjang lorong gua P = jumlah pintu gua S = jumlah jenis LL= lebar lorong gua V= ventilasi gua H’= indeks keanekaragaman jenis TL= tinggi lorong gua N = kelimpahan E = indeks kemerataan
Kelelawar subordo Megachiroptera yang paling banyak ditemukan
sarangnya adalah E. spelaea (5 sarang), diikuti oleh R. amplexicaudatus (3
sarang); C. brachyotis (3 sarang), dan C. horsfieldii (1 sarang). Kelelawar
Microchiroptera yang paling banyak ditemukan sarangnya berturut-turut adalah
H. sorenseni (8 sarang); C. plicata (2 sarang) dan H. larvatus (2 sarang).
Sedangkan M. australis; M. schreibersi; R. affinis dan R. borneensis masing-
masing hanya ditemukan satu sarang.
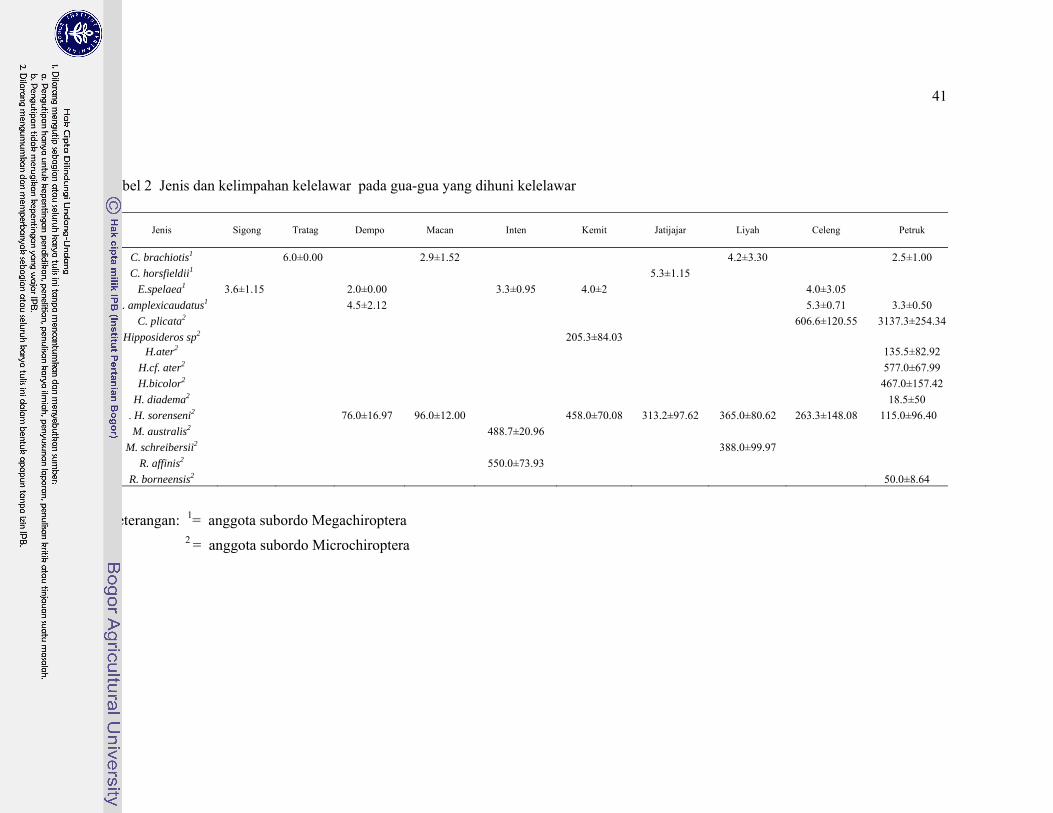
41
Tabel 2 Jenis dan kelimpahan kelelawar pada gua-gua yang dihuni kelelawar
Keterangan: 1= anggota subordo Megachiroptera
2 = anggota subordo Microchiroptera
Jenis
Sigong
Tratag
Dempo
Macan
Inten
Kemit
Jatijajar
Liyah
Celeng
Petruk
C. brachiotis1 6.0±0.00 2.9±1.52 4.2±3.30 2.5±1.00 C. horsfieldii1 5.3±1.15
E.spelaea1 3.6±1.15 2.0±0.00 3.3±0.95 4.0±2 4.0±3.05 R. amplexicaudatus1 4.5±2.12 5.3±0.71 3.3±0.50
C. plicata2 606.6±120.55 3137.3±254.34Hipposideros sp2 205.3±84.03
H.ater2 135.5±82.92 H.cf. ater2 577.0±67.99 H.bicolor2 467.0±157.42
H. diadema2 18.5±50. H. sorenseni2 76.0±16.97 96.0±12.00 458.0±70.08 313.2±97.62 365.0±80.62 263.3±148.08 115.0±96.40 M. australis2 488.7±20.96
M. schreibersii2 388.0±99.97 R. affinis2 550.0±73.93
R. borneensis2 50.0±8.64
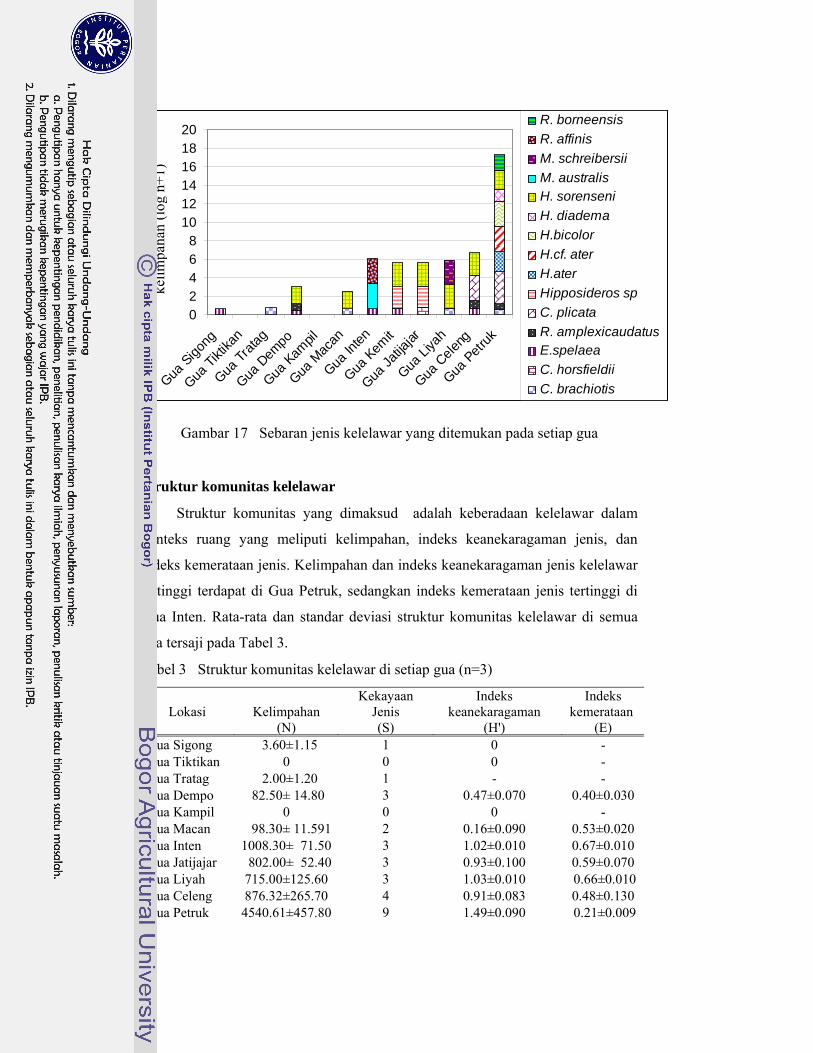
02468
101214161820
Gua Sigo
ng
Gua Ti
ktika
n
Gua Tr
atag
Gua D
empo
Gua Kam
pil
Gua M
acan
Gua In
ten
Gua Kem
it
Gua Ja
tijajar
Gua Li
yah
Gua C
eleng
Gua Petr
uk
R. borneensisR. affinisM. schreibersiiM. australisH. sorenseniH. diademaH.bicolorH.cf. ater H.aterHipposideros spC. plicataR. amplexicaudatusE.spelaeaC. horsfieldiiC. brachiotis
Gambar 17 Sebaran jenis kelelawar yang ditemukan pada setiap gua
Struktur komunitas kelelawar
Struktur komunitas yang dimaksud adalah keberadaan kelelawar dalam
konteks ruang yang meliputi kelimpahan, indeks keanekaragaman jenis, dan
indeks kemerataan jenis. Kelimpahan dan indeks keanekaragaman jenis kelelawar
tertinggi terdapat di Gua Petruk, sedangkan indeks kemerataan jenis tertinggi di
Gua Inten. Rata-rata dan standar deviasi struktur komunitas kelelawar di semua
gua tersaji pada Tabel 3.
Tabel 3 Struktur komunitas kelelawar di setiap gua (n=3)
Lokasi
Kelimpahan (N)
Kekayaan Jenis (S)
Indeks keanekaragaman
(H')
Indeks kemerataan
(E) Gua Sigong 3.60±1.15 1 0 - Gua Tiktikan 0 0 0 - Gua Tratag 2.00±1.20 1 - - Gua Dempo 82.50± 14.80 3 0.47±0.070 0.40±0.030 Gua Kampil 0 0 0 - Gua Macan 98.30± 11.591 2 0.16±0.090 0.53±0.020 Gua Inten 1008.30± 71.50 3 1.02±0.010 0.67±0.010 Gua Jatijajar 802.00± 52.40 3 0.93±0.100 0.59±0.070 Gua Liyah 715.00±125.60 3 1.03±0.010 0.66±0.010 Gua Celeng 876.32±265.70 4 0.91±0.083 0.48±0.130 Gua Petruk 4540.61±457.80 9 1.49±0.090 0.21±0.009
kelim
paha
n (lo
g n+
1)
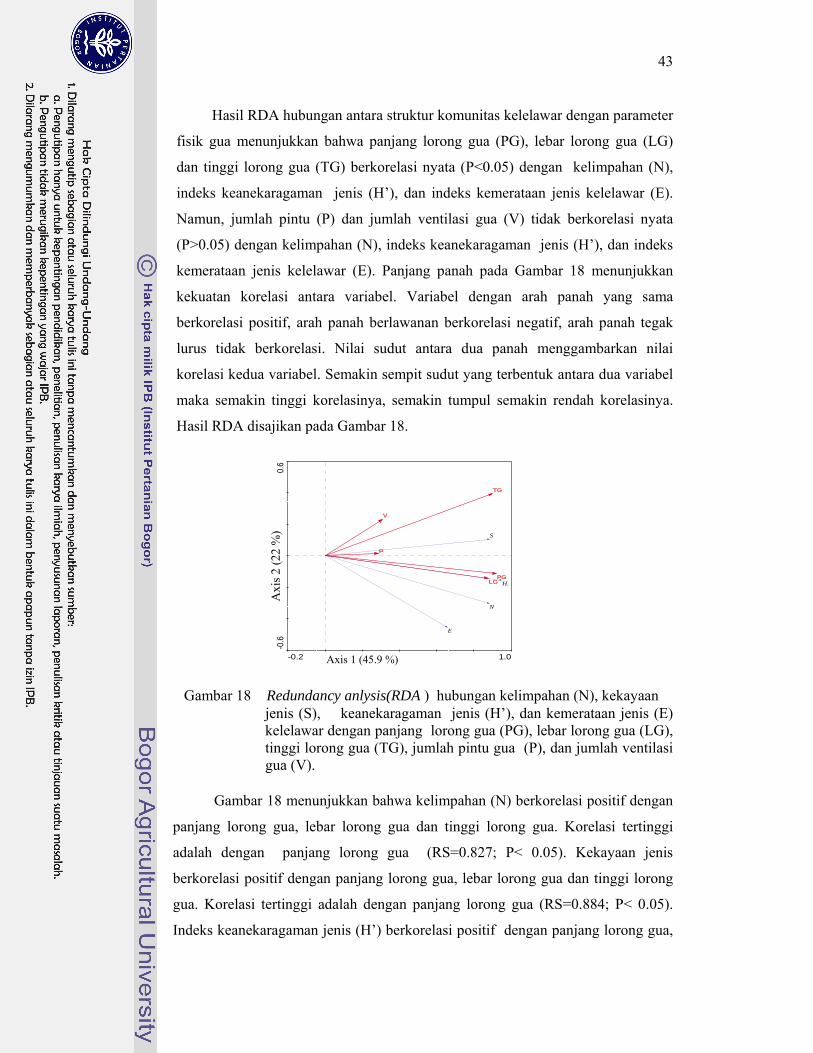
43
Hasil RDA hubungan antara struktur komunitas kelelawar dengan parameter
fisik gua menunjukkan bahwa panjang lorong gua (PG), lebar lorong gua (LG)
dan tinggi lorong gua (TG) berkorelasi nyata (P<0.05) dengan kelimpahan (N),
indeks keanekaragaman jenis (H’), dan indeks kemerataan jenis kelelawar (E).
Namun, jumlah pintu (P) dan jumlah ventilasi gua (V) tidak berkorelasi nyata
(P>0.05) dengan kelimpahan (N), indeks keanekaragaman jenis (H’), dan indeks
kemerataan jenis kelelawar (E). Panjang panah pada Gambar 18 menunjukkan
kekuatan korelasi antara variabel. Variabel dengan arah panah yang sama
berkorelasi positif, arah panah berlawanan berkorelasi negatif, arah panah tegak
lurus tidak berkorelasi. Nilai sudut antara dua panah menggambarkan nilai
korelasi kedua variabel. Semakin sempit sudut yang terbentuk antara dua variabel
maka semakin tinggi korelasinya, semakin tumpul semakin rendah korelasinya.
Hasil RDA disajikan pada Gambar 18.
-0.2 1.0
-0.6
0.6
E
S
H.
N
PGLG
TG
P
V
Gambar 18 Redundancy anlysis(RDA ) hubungan kelimpahan (N), kekayaan
jenis (S), keanekaragaman jenis (H’), dan kemerataan jenis (E) kelelawar dengan panjang lorong gua (PG), lebar lorong gua (LG), tinggi lorong gua (TG), jumlah pintu gua (P), dan jumlah ventilasi gua (V).
Gambar 18 menunjukkan bahwa kelimpahan (N) berkorelasi positif dengan
panjang lorong gua, lebar lorong gua dan tinggi lorong gua. Korelasi tertinggi
adalah dengan panjang lorong gua (RS=0.827; P< 0.05). Kekayaan jenis
berkorelasi positif dengan panjang lorong gua, lebar lorong gua dan tinggi lorong
gua. Korelasi tertinggi adalah dengan panjang lorong gua (RS=0.884; P< 0.05).
Indeks keanekaragaman jenis (H’) berkorelasi positif dengan panjang lorong gua,
Axi
s 2 (2
2 %
)
Axis 1 (45.9 %)

44
lebar lorong gua, dan tinggi lorong gua. Korelasi tertinggi adalah dengan lebar
lorong gua (RS=0.898; P < 0.05). Indeks kemerataan jenis (E) berkorelasi positif
dengan panjang lorong gua, lebar lorong gua, dan tinggi lorong gua. Korelasi
tertinggi adalah dengan lebar lorong gua (RS=0.757; P < 0.05). Dengan demikian
dapat disimpulkan bahwa semakin panjang, lebar dan tinggi lorong gua
menyebabkan semakin banyak jumlah populasi kekelawar, semakin tinggi
keanekaragaman jenis, dan semakin merata sebaran kelelawar yang bersarang di
dalamnya. Sebaliknya, hasil uji RDA menunjukkan tidak adanya korelasi
signifikan (P>0.05) antara kelimpahan, indeks keanekaragaman jenis, dan indeks
kemerataan jenis kelelawar dengan jumlah ventilasi dan jumlah pintu gua. Artinya
kelimpahan, indeks keanekaragaman jenis, dan indeks kemerataan jenis kelelawar
di gua-gua Karst Gombong tidak dipengaruhi oleh banyaknya jumlah pintu dan
jumlah ventilasi gua.
Hal menarik terlihat pada dua gua yang mempunyai lorong pendek (<
100m) yaitu Gua Sigong dan Gua Tratag. Kedua gua tersebut dihuni oleh
kelelawar, tetapi hanya satu jenis Megachiroptera dan jumlahnya sangat sedikit ( <
4 ekor). Sementara gua lain, yaitu Gua Kampil yang mempunyai lorong sedang
(100m s/d 200m) sama sekali tidak dihuni kelelawar.
Pola pemilihan sarang kelelawar
Hasil pengukuran mikroklimat sarang kelelawar pada setiap gua dapat
diterangkan sebagai berikut : di Gua Macan, C.brachyotis bersarang di lokasi
dengan jarak 2 ± 0.7 m, tinggi 8.8 ± 0.6 m, suhu 2 ± 0.7oC, kelembapan 54.6 ±
1.15%, intensitas cahaya 81.4 ± 50 lux, intensitas suara 289.5 ± 115 db, kecepatan
angin 0.001 ± 0 m/s, kadar oksigen 21.1 ± 0.57 %, dan kadar amonia udara 944 ±
0 ppm. Secara lengkap, hasil pengukuran mikroklimat sarang semua jenis
kelelawar pada setiap gua dapat dilihat pada Tabel 4.
Hasil analisis RDA dengan metode seleksi langkah maju (forward
selection) menunjukkan terdapat lima parameter fisik mikroklimat yang
mempengaruhi pemilihan sarang kelelawar, yaitu jarak dari pintu gua, suhu,
kelembapan, intensitas cahaya, dan intensitas suara. Dari ke-lima parameter
tersebut yang paling dominan mempengaruhi pemilihan sarang oleh kelelawar
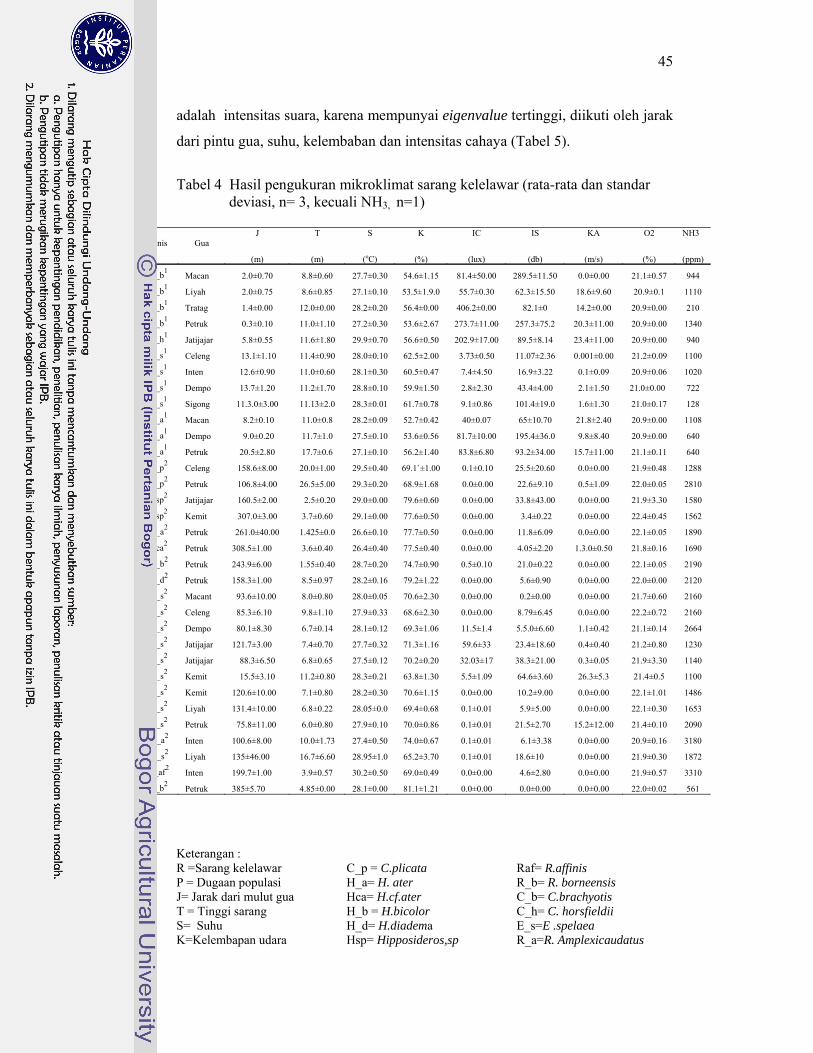
45
adalah intensitas suara, karena mempunyai eigenvalue tertinggi, diikuti oleh jarak
dari pintu gua, suhu, kelembaban dan intensitas cahaya (Tabel 5).
Tabel 4 Hasil pengukuran mikroklimat sarang kelelawar (rata-rata dan standar
deviasi, n= 3, kecuali NH3, n=1)
Jenis Gua J
T
S
K
IC
IS
KA
O2
NH3
(m) (m) (oC) (%) (lux) (db) (m/s) (%) (ppm)
C_b1 Macan 2.0±0.70 8.8±0.60 27.7±0.30 54.6±1.15 81.4±50.00 289.5±11.50 0.0±0.00 21.1±0.57 944
C_b1 Liyah 2.0±0.75 8.6±0.85 27.1±0.10 53.5±1.9.0 55.7±0.30 62.3±15.50 18.6±9.60 20.9±0.1 1110
C_b1 Tratag 1.4±0.00 12.0±0.00 28.2±0.20 56.4±0.00 406.2±0.00 82.1±0 14.2±0.00 20.9±0.00 210
C_b1 Petruk 0.3±0.10 11.0±1.10 27.2±0.30 53.6±2.67 273.7±11.00 257.3±75.2 20.3±11.00 20.9±0.00 1340
C_h1 Jatijajar 5.8±0.55 11.6±1.80 29.9±0.70 56.6±0.50 202.9±17.00 89.5±8.14 23.4±11.00 20.9±0.00 940
E_s1 Celeng 13.1±1.10 11.4±0.90 28.0±0.10 62.5±2.00 3.73±0.50 11.07±2.36 0.001±0.00 21.2±0.09 1100
E_s1 Inten 12.6±0.90 11.0±0.60 28.1±0.30 60.5±0.47 7.4±4.50 16.9±3.22 0.1±0.09 20.9±0.06 1020
E_s1 Dempo 13.7±1.20 11.2±1.70 28.8±0.10 59.9±1.50 2.8±2.30 43.4±4.00 2.1±1.50 21.0±0.00 722
E_s1 Sigong 11.3.0±3.00 11.13±2.0 28.3±0.01 61.7±0.78 9.1±0.86 101.4±19.0 1.6±1.30 21.0±0.17 128
R_a1 Macan 8.2±0.10 11.0±0.8 28.2±0.09 52.7±0.42 40±0.07 65±10.70 21.8±2.40 20.9±0.00 1108
R_a1 Dempo 9.0±0.20 11.7±1.0 27.5±0.10 53.6±0.56 81.7±10.00 195.4±36.0 9.8±8.40 20.9±0.00 640
R_a1 Petruk 20.5±2.80 17.7±0.6 27.1±0.10 56.2±1.40 83.8±6.80 93.2±34.00 15.7±11.00 21.1±0.11 640
C_p2 Celeng 158.6±8.00 20.0±1.00 29.5±0.40 69.1`±1.00 0.1±0.10 25.5±20.60 0.0±0.00 21.9±0.48 1288
C_p2 Petruk 106.8±4.00 26.5±5.00 29.3±0.20 68.9±1.68 0.0±0.00 22.6±9.10 0.5±1.09 22.0±0.05 2810
Hsp2 Jatijajar 160.5±2.00 2.5±0.20 29.0±0.00 79.6±0.60 0.0±0.00 33.8±43.00 0.0±0.00 21.9±3.30 1580
Hsp2 Kemit 307.0±3.00 3.7±0.60 29.1±0.00 77.6±0.50 0.0±0.00 3.4±0.22 0.0±0.00 22.4±0.45 1562
H_a2 Petruk 261.0±40.00 1.425±0.0 26.6±0.10 77.7±0.50 0.0±0.00 11.8±6.09 0.0±0.00 22.1±0.05 1890
Hca2 Petruk 308.5±1.00 3.6±0.40 26.4±0.40 77.5±0.40 0.0±0.00 4.05±2.20 1.3.0±0.50 21.8±0.16 1690
H_b2 Petruk 243.9±6.00 1.55±0.40 28.7±0.20 74.7±0.90 0.5±0.10 21.0±0.22 0.0±0.00 22.1±0.05 2190
H_d2 Petruk 158.3±1.00 8.5±0.97 28.2±0.16 79.2±1.22 0.0±0.00 5.6±0.90 0.0±0.00 22.0±0.00 2120
H_s2 Macant 93.6±10.00 8.0±0.80 28.0±0.05 70.6±2.30 0.0±0.00 0.2±0.00 0.0±0.00 21.7±0.60 2160
H_s2 Celeng 85.3±6.10 9.8±1.10 27.9±0.33 68.6±2.30 0.0±0.00 8.79±6.45 0.0±0.00 22.2±0.72 2160
H_s2 Dempo 80.1±8.30 6.7±0.14 28.1±0.12 69.3±1.06 11.5±1.4 5.5.0±6.60 1.1±0.42 21.1±0.14 2664
H_s2 Jatijajar 121.7±3.00 7.4±0.70 27.7±0.32 71.3±1.16 59.6±33 23.4±18.60 0.4±0.40 21.2±0.80 1230
H_s2 Jatijajar 88.3±6.50 6.8±0.65 27.5±0.12 70.2±0.20 32.03±17 38.3±21.00 0.3±0.05 21.9±3.30 1140
H_s2 Kemit 15.5±3.10 11.2±0.80 28.3±0.21 63.8±1.30 5.5±1.09 64.6±3.60 26.3±5.3 21.4±0.5 1100
H_s2 Kemit 120.6±10.00 7.1±0.80 28.2±0.30 70.6±1.15 0.0±0.00 10.2±9.00 0.0±0.00 22.1±1.01 1486
H_s2 Liyah 131.4±10.00 6.8±0.22 28.05±0.0 69.4±0.68 0.1±0.01 5.9±5.00 0.0±0.00 22.1±0.30 1653
H_s2 Petruk 75.8±11.00 6.0±0.80 27.9±0.10 70.0±0.86 0.1±0.01 21.5±2.70 15.2±12.00 21.4±0.10 2090
M_a2 Inten 100.6±8.00 10.0±1.73 27.4±0.50 74.0±0.67 0.1±0.01 6.1±3.38 0.0±0.00 20.9±0.16 3180
M_s2 Liyah 135±46.00 16.7±6.60 28.95±1.0 65.2±3.70 0.1±0.01 18.6±10 0.0±0.00 21.9±0.30 1872
R_af2 Inten 199.7±1.00 3.9±0.57 30.2±0.50 69.0±0.49 0.0±0.00 4.6±2.80 0.0±0.00 21.9±0.57 3310
R_b2 Petruk 385±5.70 4.85±0.00 28.1±0.00 81.1±1.21 0.0±0.00 0.0±0.00 0.0±0.00 22.0±0.02 561
Keterangan : R =Sarang kelelawar P = Dugaan populasi J= Jarak dari mulut gua T = Tinggi sarang S= Suhu K=Kelembapan udara
C_p = C.plicata H_a= H. ater Hca= H.cf.ater H_b = H.bicolor H_d= H.diadema Hsp= Hipposideros,sp
Raf= R.affinis R_b= R. borneensis C_b= C.brachyotis C_h= C. horsfieldii E_s=E .spelaea R_a=R. Amplexicaudatus
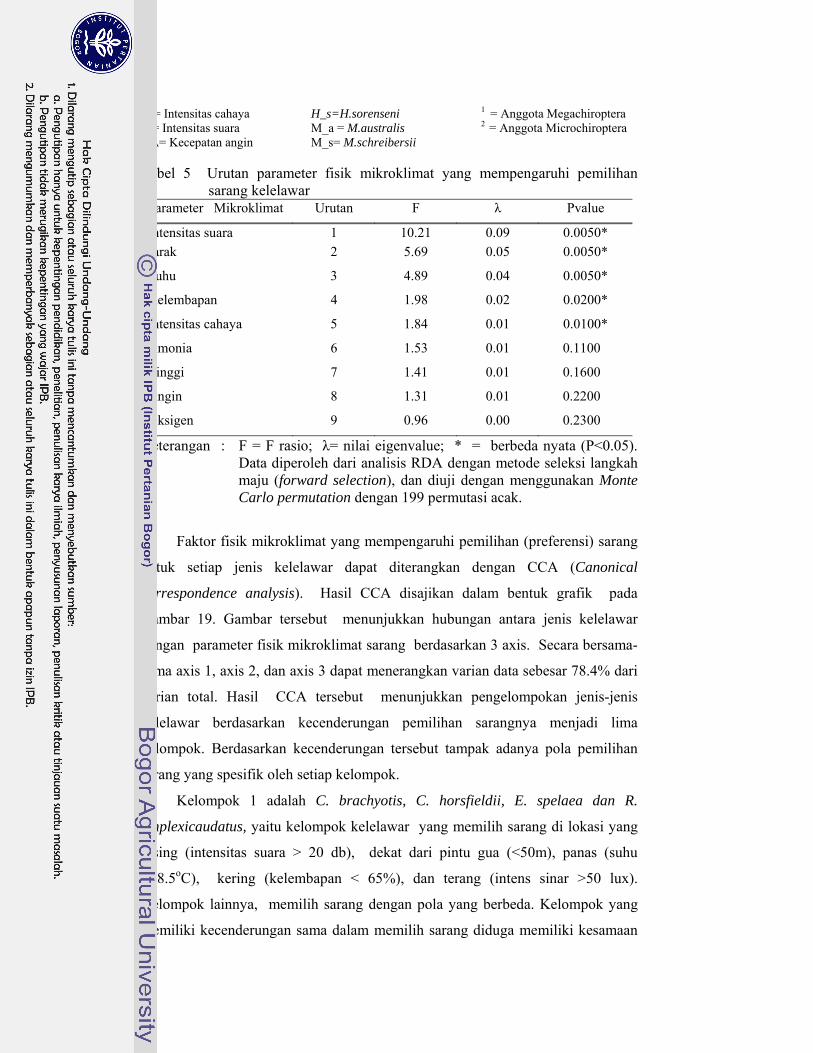
46
IC= Intensitas cahaya IS= Intensitas suara KA= Kecepatan angin
H_s=H.sorenseni M_a = M.australis M_s= M.schreibersii
1 = Anggota Megachiroptera 2 = Anggota Microchiroptera
Tabel 5 Urutan parameter fisik mikroklimat yang mempengaruhi pemilihan sarang kelelawar
Parameter Mikroklimat Urutan F λ Pvalue
Intensitas suara 1 10.21 0.09 0.0050* Jarak 2 5.69 0.05 0.0050*
Suhu 3 4.89 0.04 0.0050*
Kelembapan 4 1.98 0.02 0.0200*
Intensitas cahaya 5 1.84 0.01 0.0100*
Amonia 6 1.53 0.01 0.1100
Tinggi 7 1.41 0.01 0.1600
Angin 8 1.31 0.01 0.2200
Oksigen 9 0.96 0.00 0.2300
Keterangan : F = F rasio; λ= nilai eigenvalue; * = berbeda nyata (P<0.05). Data diperoleh dari analisis RDA dengan metode seleksi langkah maju (forward selection), dan diuji dengan menggunakan Monte Carlo permutation dengan 199 permutasi acak.
Faktor fisik mikroklimat yang mempengaruhi pemilihan (preferensi) sarang
untuk setiap jenis kelelawar dapat diterangkan dengan CCA (Canonical
correspondence analysis). Hasil CCA disajikan dalam bentuk grafik pada
Gambar 19. Gambar tersebut menunjukkan hubungan antara jenis kelelawar
dengan parameter fisik mikroklimat sarang berdasarkan 3 axis. Secara bersama-
sama axis 1, axis 2, dan axis 3 dapat menerangkan varian data sebesar 78.4% dari
varian total. Hasil CCA tersebut menunjukkan pengelompokan jenis-jenis
kelelawar berdasarkan kecenderungan pemilihan sarangnya menjadi lima
kelompok. Berdasarkan kecenderungan tersebut tampak adanya pola pemilihan
sarang yang spesifik oleh setiap kelompok.
Kelompok 1 adalah C. brachyotis, C. horsfieldii, E. spelaea dan R.
amplexicaudatus, yaitu kelompok kelelawar yang memilih sarang di lokasi yang
bising (intensitas suara > 20 db), dekat dari pintu gua (<50m), panas (suhu
>28.5oC), kering (kelembapan < 65%), dan terang (intens sinar >50 lux).
Kelompok lainnya, memilih sarang dengan pola yang berbeda. Kelompok yang
memiliki kecenderungan sama dalam memilih sarang diduga memiliki kesamaan
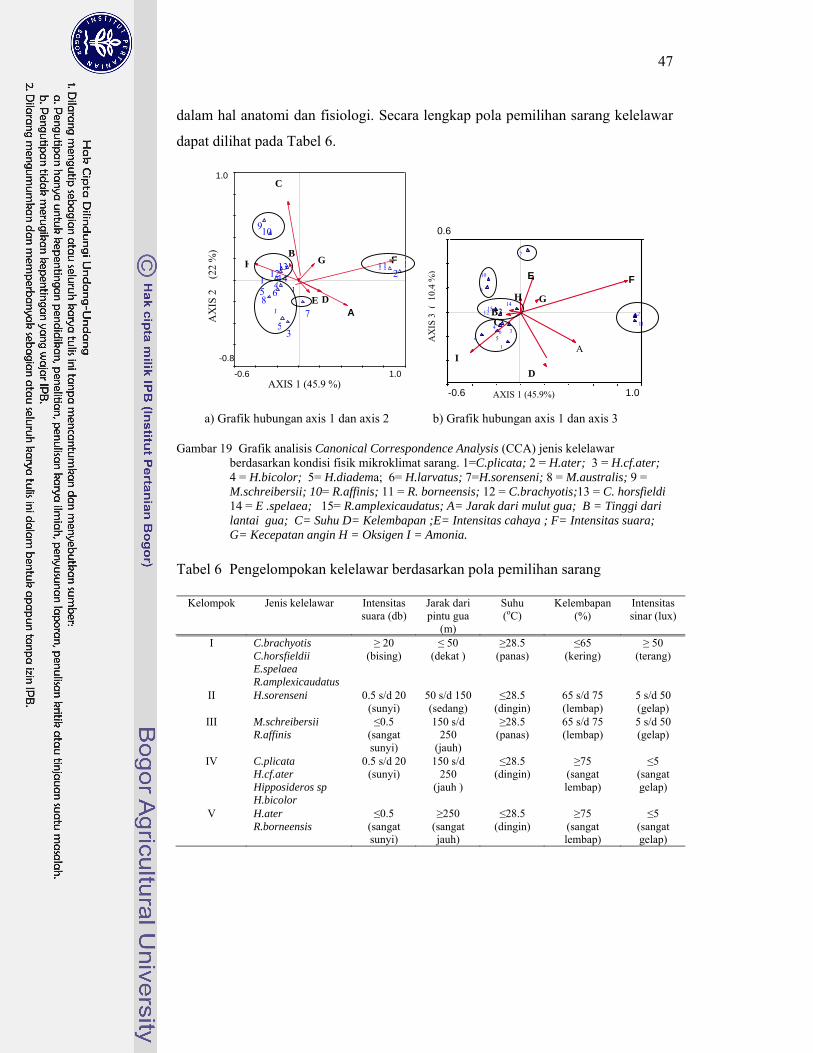
47
dalam hal anatomi dan fisiologi. Secara lengkap pola pemilihan sarang kelelawar
dapat dilihat pada Tabel 6.
a) Grafik hubungan axis 1 dan axis 2 b) Grafik hubungan axis 1 dan axis 3 Gambar 19 Grafik analisis Canonical Correspondence Analysis (CCA) jenis kelelawar
berdasarkan kondisi fisik mikroklimat sarang. 1=C.plicata; 2 = H.ater; 3 = H.cf.ater; 4 = H.bicolor; 5= H.diadema; 6= H.larvatus; 7=H.sorenseni; 8 = M.australis; 9 = M.schreibersii; 10= R.affinis; 11 = R. borneensis; 12 = C.brachyotis;13 = C. horsfieldi 14 = E .spelaea; 15= R.amplexicaudatus; A= Jarak dari mulut gua; B = Tinggi dari lantai gua; C= Suhu D= Kelembapan ;E= Intensitas cahaya ; F= Intensitas suara; G= Kecepatan angin H = Oksigen I = Amonia.
Tabel 6 Pengelompokan kelelawar berdasarkan pola pemilihan sarang
Kelompok Jenis kelelawar Intensitas suara (db)
Jarak dari pintu gua
(m)
Suhu (oC)
Kelembapan (%)
Intensitas sinar (lux)
I C.brachyotis C.horsfieldii E.spelaea R.amplexicaudatus
≥ 20 (bising)
≤ 50 (dekat )
≥28.5 (panas)
≤65 (kering)
≥ 50 (terang)
II H.sorenseni 0.5 s/d 20 (sunyi)
50 s/d 150 (sedang)
≤28.5 (dingin)
65 s/d 75 (lembap)
5 s/d 50 (gelap)
III M.schreibersii R.affinis
≤0.5 (sangat sunyi)
150 s/d 250
(jauh)
≥28.5 (panas)
65 s/d 75 (lembap)
5 s/d 50 (gelap)
IV C.plicata H.cf.ater Hipposideros sp H.bicolor
0.5 s/d 20 (sunyi)
150 s/d 250
(jauh )
≤28.5 (dingin)
≥75 (sangat lembap)
≤5 (sangat gelap)
V H.ater R.borneensis
≤0.5 (sangat sunyi)
≥250 (sangat jauh)
≤28.5 (dingin)
≥75 (sangat lembap)
≤5 (sangat gelap)
-0.6 1.0
0.6
12
7
1514
18
10
6
9
4
53
2
11
A
B3C
D
E F
GH
I
AX
IS 3
( 1
0.4
%)
AXIS 1 (45.9%)
-0.6 1.0
-0.8
1.0
12
7
14
1
910
6 8
4
53
2
A
B
C
D E
FG
I
11
15
AXIS 1 (45.9 %)
AX
IS 2
(2
2 %
)
H 13

48
Pembahasan
Jumlah semua jenis kelelawar yang ditemukan di gua-gua Karst Gombong
mencapai 15 jenis atau 10% dari 151 jenis yang pernah dilaporkan terdapat di
Indonesia (Suyanto et al. 1998). Bila dibandingkan dengan penelitian yang
dilakukan di kawasan karst lain di Indonesia maupun di luar Indonesia, jumlah
jenis yang ditemukan dalam penelitian ini tergolong tinggi. Di Indonesia,
penelitian Maryanto & Maharadatunkamsi (1991) di gua-gua Karst Sumbawa,
mendapatkan delapan jenis kelelawar, penelitian Saroni (2005) di gua-gua
kawasan Karst Sangkulirang-Mangkaliat Kalimantan Timur mendapatkan
sembilan jenis kelelawar, penelitian Pujirianti (2006) di gua-gua kawasan Karst
Alas Purwo mendapatkan 13 jenis kelelawar, dan penelitian Apriandi et al. (2008)
di gua-gua kawasan Karst Gudawang Bogor mendapatkan 10 jenis kelelawar. Di
luar Indonesia, penelitian Furman & Ozgul (2002) mendapatkan delapan jenis
kelelawar di Karst Istambul Turki, dan penelitian Parsons et al. (2002)
mendapatkan 11 jenis kelelawar di Britain Inggris. Hasil penelitian ini
menunjukkan bahwa Karst Gombong menyimpan kekayaan jenis kelelawar yang
cukup tinggi.
Banyaknya jumlah jenis yang ditemukan dalam penelitian ini disebabkan
gua-gua di kawasan Karst Gombong memiliki ukuran dan geomorfologi yang
berbeda-beda, sehingga terbentuk lingkungan fisik yang sangat bervariasi. Setiap
kondisi lingkungan gua yang berbeda menghasilkan mikro habitat unik yang
dihuni jenis kelelawar berbeda-beda. Selain itu ketersediaan pakan dan sumber
air di kawasan Karst Gombong juga mendukung banyak jenis kelelawar untuk
hidup dan bertahan. Gua-gua yang diteliti seluruhnya berada di lingkungan hutan
karst yang dikelilingi oleh lahan pertanian. Menurut Riswan et al. (2006) vegetasi
hutan karst di kawasan Karst Gombong terdiri atas 187 jenis tumbuhan yang
termasuk dalam 125 marga dan 60 suku tumbuhan. Jenis-jenis tanaman tersebut
menyediakan pakan bagi kelelawar.
Jumlah jenis kelelawar dalam satu gua yang ditemukan di gua-gua karst di
Indonesia pada penelitian-penelitian sebelumnya, antara satu sampai enam jenis
kelelawar (Maryanto & Maharadatunkamsi 1991; Saroni 2005; Pujirianti 2006;
Apriandi 2006). Beberapa hasil penelitian pada kawasan karst lain di luar

49
Indonesia mendapatkan jumlah yang bervariasi antara satu sampai tiga jenis
kelelawar dalam satu gua. Misalnya penelitian Seckerdieck et al. (2005) di Gua
Alterberga Jerman hanya mendapatkan satu jenis kelelawar: Rhinolophus
hipposideros (Microchiroptera) bersarang dalam satu gua. Penelitian Dunn
(1978) di Gua Anak Takun Malaysia; penelitian Duran & Centano (2002) di Gua
Bonita India Barat ; dan penelitian Zukal et al. (2005) di Gua Katerinska
Chekoslovakia masing-masing menemukan dua jenis kelelawar bersarang dalam
satu gua. Penelitian Zahn dan Hager (2005) mendapatkan tiga jenis kelelawar
bersarang dalam satu gua yang berlokasi di Bavaria Jerman. Dalam penelitian ini
terdapat satu gua, yaitu Gua Petruk yang dihuni oleh sembilan jenis kelelawar.
Jumlah ini jauh lebih tinggi dibandingkan penemuan-penemuan sebelumnya.
Banyaknya jenis kelelawar yang ditemukan di Gua Petruk disebabkan
variasi lingkungan yang terbentuk di Gua Petruk. Hal ini sesuai dengan pendapat
Castillo et al. (2009) bahwa kondisi lingkungan di dalam satu gua dapat berbeda
antara satu zona (mintakat) dengan zona lainnya, dan dapat menyebabkan
pemisahan mikroklimat dalam ruang gua. Pemisahan mikroklimat tersebut dapat
mengundang keanekaragaman jenis mahluk hidup.
Megachiroptera yang ditemukan dalam penelitian, dua jenis di antaranya
yaitu Cynopterus horsfieldi dan Cynopterus brachiotis, tidak pernah didapatkan
bersarang di gua karst pada penelitian sebelumnya baik yang dilakukan di
Indonesia maupun di luar Indonesia. Kedua jenis kelelawar tersebut lebih sering
ditemukan bersarang di pepohonan, sesuai dengan hasil penelitian Ruczynski et
al. (2007) dan Soegiharto & Kartono (2009). Menurut Altringham (1996),
kelelawar genus Cynopterus memilih bersarang di pepohonan karena
mengandalkan penglihatannya untuk mengenali lingkungan. Ditemukannya jenis
C. horsfieldii dan C. brachyotis hanya dalam penelitian ini, menunjukkan gua-
gua yang dihuni jenis tersebut intensitas cahayanya tinggi, sehingga masih
memungkinkan C. horsfieldii dan C. brachyotis menggunakan penglihatannya.
Hal ini didukung oleh hasil pengukuran mikroklimat sarang, terbukti C.
horsfielsdii memilih sarang di tempat yang terang (intensitas cahaya > 50 lux).
Sementara, dua jenis Megachiroptera lainnya yaitu R.amplexicaudatus dan E.
spelaea pernah ditemukan bersarang di dalam gua karst pada beberapa penelitian

50
sebelumnya (Maryanto & Maharadatunkmsi 1991; Suyanto 2001; Pujirianti
2006). Menurut Altringham (1996), R. amplexicaudatus dan E. spelaea mampu
menggunakan ekholokasi untuk memahami ruang, meskipun kemampuan
ekholokasinya tidak sebaik kelelawar Microchiroptera. Hal ini menyebabkan R.
amplexicaudatus dan E. spelaea sering ditemukan bersarang di dalam gua dengan
kondisi gelap ataupun terang.
Ke-lima genus Microchiroptera yang ditemukan dalam penelitian ini
seluruhnya pernah dilaporkan bersarang di gua-gua karst di luar Indonesia
maupun di Indonesia (Dunn 1978; Maryanto & Maharadatunkamsi 1991; Furman
& Ozgul 2002; Parsons et al. 2002; Seckerdieck et al. 2004; Zahn dan Hager
2005; Saroni 2005; Pujirianti 2006 dan Apriandi et al. 2008). Hal ini
menunjukkan genus tersebut merupakan penghuni gua yang sudah teradaptasi
dengan kondisi lingkungan gua. Menurut Altringham (1996) dan Zahn & Hager
(2005), beberapa jenis Microchiroptera memilih gua sebagai sarang karena
kondisi gua lembap, suhu stabil dan jauh dari kebisingan. Dengan kondisi
demikian kelelawar dapat meminimalkan kekurangan air akibat evaporasi, dapat
memilih suhu yang tepat untuk tubuhnya, dan dapat menghindari kebisingan yang
dapat mengganggu bahkan dapat menyebabkan kematian. Kondisi gua yang gelap
tidak menjadi masalah bagi kelelawar Microchiroptera. Hal ini karena
Microchiroptera memiliki ekholokasi, yaitu kemampuan mendeteksi benda-
benda di sekitarnya, dengan menggunakan gelombang pantul (echo/ gema)
berfrekuensi ultrasonik. Ekholokasi ini menyebabkan dalam kondisi gelap,
kelelawar mampu berorientasi terhadap ruang gua (Verboom et al. 1999; Ulrich
et al. 2003).
Berdasarkan jumlah temuan sarang (Gambar 16), kelelawar
Megachiroptera yang paling banyak ditemukan sarangnya adalah E. spelaea.
Microchiroptera yang paling banyak ditemukan sarangnya adalah H. sorenseni.
Hal ini menunjukkan, meskipun membutuhkan kondisi lingkungan tertentu,
kondisi lingkungan yang diinginkan oleh kedua kelelawar tersebut tersedia di
banyak gua di Karst Gombong. Sebaliknya, C. horsfieldii (Megachiroptera), M.
australis; M. schreibersi; R. affinis dan R. borneensis hanya ditemukan di satu

51
gua. Hal ini menunjukkan bahwa jenis-jenis kelelawar tersebut menghendaki
kondisi lingkungan yang spesifik yang hanya tersedia di satu gua saja.
Hal menarik ditemukan di Gua Jatijajar, dimana kelelawar H. sorenseni
(Subordo: Microchiroptera) ditemukan dalam dua sarang yang letaknya berjauhan,
sementara di gua-gua lainnya satu jenis kelelawar hanya menempati satu sarang.
Hal ini dapat dijelaskan dengan hasil penelitian Rossiter et al. (2002) yang
menguji genetik 15 individu betina Rhinolophus ferrumequinum (Rhinolophidae :
Microchiroptera) di Barat Daya Britain dengan menggunakan microsatellite.
Hasilnya menunjukkan, individu jenis sama tetapi berasal dari koloni berbeda
memiliki jarak genetik antara 0.17 – 0.64. Sementara, individu yang berasal dari
satu koloni yang sama memiliki jarak genetik mendekati nol (0.03). Hal ini
menunjukkan anggota koloni yang sama memiliki hubungan kekerabatan yang
sangat dekat atau berasal dari induk yang sama sedangkan koloni yang berbeda,
meskipun dari jenis yang sama memperlihatkan hubungan kekerabatan yang
cukup jauh atau berasal dari induk yang berbeda. Berdasarkan pernyataan
tersebut, dapat diduga bahwa dua koloni H. sorenseni yang ditemukan di Gua
Jatijajar merupakan koloni yang berasal dari induk yang berbeda.
Hasil RDA menunujukkan semakin panjang, tinggi, dan lebar lorong gua,
semakin tinggi kelimpahan, indeks keanekaragaman jenis, dan kemerataan jenis
kelelawar. Hal ini sesuai dengan pendapat Maguran (2004) bahwa semakin luas
habitat, semakin banyak mahluk hidup yang dapat hidup di dalamnya. Lebih
lanjut Baudinette et al. (1994) menjelaskan bahwa lorong gua yang panjang dapat
menyebabkan pemisahan mikroklimat ruang gua. Semakin banyak mikroklimat
yang terbentuk, maka semakin banyak jenis kelelawar yang dapat bersarang di
ruang-ruang tersebut. Hasil penelitian Sevcik (2003) pada kelelawar Plecotus
auritus dan P. austriacus telah mampu menerangkan mengapa semakin lebar
lorong gua, semakin tinggi indeks keanekaragaman jenis kelelawar. Menurut
Sevcik (2003), P. auritus lebih leluasa melakukan manuver daripada P.
austriacus. Akibatnya P. auritus memiliki keunggulan tersendiri dalam eksploitasi
habitat. Berdasarkan hasil penelitian tersebut, dapat dikatakan bahwa gua dengan
lorong sempit hanya dapat dihuni oleh jenis tertentu saja, yaitu jenis yang mampu
malakukan manuver dengan baik. Sebaliknya pada gua dengan lorong lebar, dapat

52
dihuni kelelawar dengan kemampuan lebih beragam. Akibatnya, semakin lebar
lorong gua, maka semakin banyak jenis yang dapat bersarang di dalamnya.
Gua-gua dengan lorong pendek (< 100m) dapat dihuni oleh Megachiroptera.
Sementara Microchiroptera hanya menghuni gua dengan panjang lorong lebih
dari 100 meter. Hal ini karena gua dengan lorong pendek hanya memiliki
mikroklimat dengan kondisi panas, terang, kering dan bising. Berdasarkan hasil
identifikasi mikroklimat sarang, diketahui bahwa kondisi ini tidak disukai oleh
Microchiroptera, sementara Megachiroptera dapat bertahan dengan kondisi
tersebut. Hal menarik terlihat di Gua Kampil yang memiliki panjang lorong lebih
dari 100 meter, lebar lorong 4.2 meter dan tinggi lorong 3.1 meter. Di gua tersebut
tidak ditemukan kelelawar. Diduga ada faktor lain, selain faktor fisik dan
mikroklimat gua yang menyebabkan gua tersebut tidak dihuni kelelawar. Faktor
tersebut adalah gangguan oleh manusia. Berdasarkan hasil pengamatan, di lorong
Gua Kampil ditemukan bekas galian yang ditinggalkan oleh penambang ilegal.
Kegiatan penambangan tersebut diduga mengganggu kelelawar, dan
menyebabkan gua tersebut tidak dihuni kelelawar.
Tidak berpengaruhnya jumlah pintu gua dan jumlah ventilasi gua pada
struktur komunitas kelelawar (kelimpahan, keanekaragaman jenis, dan
kemerataan jenis) disebabkan meskipun mempunyai beberapa pintu dan ventilasi,
semua jenis kelelawar yang bersarang dalam satu gua cenderung menggunakan
satu pintu atau ventilasi yang sama untuk keluar masuk gua. Hal ini sesuai
dengan hasil penelitian Schnitzler et al. (2003) yang membuktikan ketika terbang
menuju lokasi sarang dan tempat pencarian makan, kelelawar cenderung
menggunakan jalur yang sama. Transfer informasi penggunaan jalur terbang ini
dilakukan dari orang tua (induk) kepada anak melalui perilaku mengikuti
(following behavior).
Hasil CCA menunjukkan lima parameter yang berpengaruh pada pemilihan
sarang kelelawar berturut-turut dari yang paling kuat sampai yang paling lemah
pengaruhnya adalah: intensitas suara, jarak dari pintu gua, suhu, kelembapan dan
intensitas cahaya. Penelitian Schnitzler et al. (2003) membuktikan durasi sinyal
ekholokasi yang digunakan oleh kelelawar Nyctalus noctula dan Rhinolophus
ferrumequinum di habitat tertutup lebih panjang daripada di habitat terbuka. Hal

53
ini karena, di ruang tertutup gema yang dihasilkan lebih kompleks sehingga
kelelawar lebih sulit menganalisis gelombang pantul (ekholokasi). Oleh kerena
itu, di ruang tertutup seperti di dalam gua, gangguan suara sedikit saja akan
menyebabkan kelelawar gagal menganalisis gelombang pantul. Akibatnya,
beberapa jenis kelelawar yang memiliki pendengaran sangat sensitif memilih
sarang dengan intensitas suara mendekati nol, sedangkan jenis-jenis kelelawar
yang pendengarannya kurang sensitif dapat bersarang di lokasi dengan intensitas
suara tinggi.
Jarak dari pintu gua merupakan parameter kedua setelah intensitas suara
yang berpengaruh nyata pada pemilihan sarang kelelawar. Hal ini karena untuk
dapat terbang dalam lorong gua yang panjang dan sempit, dibutuhkan gerakan
manuver yang baik. Sebagai akibatnya, dalam memilih lokasi sarang, kelelawar
yang mampu terbang dengan manuver yang baik cenderung memilih lokasi sarang
pada jarak yang jauh dari lorong gua karena lebih aman dari gangguan manusia.
Sebaliknya, kelelawar yang tidak mampu melewati lorong gua yang panjang
memilih sarang di lokasi yang dekat dari pintu gua untuk memudahkan kelelawar
tersebut keluar atau masuk ke dalam sarang.
Suhu merupakan parameter fisik mikroklimat urutan ke-tiga yang
berkorelasi nyata pada pemilihan sarang kelelawar. Penelitian Zukal et al. (2005)
membuktikan bahwa kelelawar Myotis myotis and Rhinolophus hipposideros
memilih ruang yang hangat (20oC s/d 26oC) ketika melakukan hibernasi di musim
dingin. Hal ini karena kelelawar merupakan homoikioterm (suhu tubuh konstan)
yang mempunyai batas toleransi sempit pada suhu lingkungan. Batas toleransi
tersebut berbeda antara satu jenis kelelawar dengan jenis lainnya, sehingga setiap
jenis kelelawar memilih sarang yang sesuai dengan batas toleransi tubuhnya.
Kelembapan merupakan parameter urutan ke-empat yang berkorelasi nyata
pada pemilihan sarang kelelawar. Menurut Baudinete et al. (1994) membran
petagium (sayap) kelelawar tersusun atas lapisan kulit tipis yang sangat peka pada
kekeringan. Hal ini menyebabkan kelelawar yang mempunyai membran petagium
tipis memilih lokasi sarang yang lembap, sedangkan yang memiliki membran
petagium tebal mampu bersarang di lokasi gua yang cenderung kering.
Berdasarkan kekuatan korelasinya, intensitas cahaya merupakan parameter

54
terakhir yang berkorelasi pada pemilihan sarang kelelawar. Megachiroptera
cenderung menggunakan penglihatannya untuk berorientasi pada ruang. Oleh
karena itu, jenis-jenis kelelawar Megachiroptera cenderung memilih lokasi di
dalam gua yang mempunyai intensitas cahaya tinggi. Sebaliknya, jenis-jenis
Microchiroptera lebih menggunakan kemampuan ekholokasinya untuk
berorientasi pada ruang, sehingga tidak memerlukan cahaya dan memilih sarang
di ruang gua yang intensitas cahayanya rendah.
Hasil analisis CCA menunjukkan 5 kelompok kelelawar berdasarkan
kecenderungan pola pemilihan sarangnya. Kelompok I seluruhnya adalah anggota
Megachiroptera. Menurut Altringham (1996) kebanyakan kelelawar pemakan
buah (Magachiroptera) lebih sering bersarang di pohon daripada di gua. Hal ini
karena kelelawar Megachiroptera masih mengandalkan penglihatan daripada
ekholokasi, sehingga untuk berorientasi pada lingkungan masih dibutuhkan
cahaya. Pola pemilihan sarang oleh Megachiroptera ini menyebabkan
Megachiroptera dapat menghuni gua dengan panjang lorong kurang dari 100
meter, misalnya di Gua Sigong dan Gua Tratag. Karena gua-gua yang lorongnya
pendek biasanya hanya memiliki ruang dengan mikroklimat panas, terang, kering
dan bising.
Kelompok II, hanya terdiri atas kelelawar H. sorenseni. Berdasarkan hasil
pemetaan sarang, kelompok ini paling banyak ditemukan di gua-gua Karst
Gombong (8 sarang). Hal ini menunjukkan, kondisi lingkungan sarang yang
diinginkan oleh kelelawar tersebut (dingin, lembap, gelap, dan sunyi) paling
banyak tersedia di gua-gua Karst Gombong. Sejauh ini belum ada penelitian
yang menjelaskan kecenderungan pemilihan sarang jenis tersebut, kecuali
penelitian di Karst Gombong ini.
Kelompok III terdiri atas M. schreibersii dan R. affinis. Hasil penelitian ini
didukung oleh hasil penelitian sebelumnya, yaitu penelitian Apriandi et al. (2008)
yang mendapatkan jenis R. affinis di beberapa gua Karst Gudawang Bogor berada
pada jarak rata rata 180 m dari pintu gua, suhu 27.5oC dan kelembapan 96%.
Kelompok IV terdiri atas C. plicata, H.cf. ater, Hipposideros sp, dan H. bicolor.
Hasil penelitian ini sejalan dengan hasil penelitian Twente (2004) yang
mendapatkan C. plicata dengan jumlah ribuan ekor di kubah gua di Istambul

55
Turki berada pada jarak 200 m dari pintu gua , suhu 27.5oC. Kelompok V terdiri
atas H. ater dan R. borneensis. Pemilihan sarang R. borneensis yang ditemukan
dalam penelitian ini (berjarak > 250 m dari pintu gua) sedikit berbeda dari hasil
penelitian Saroni (2005) yang mendapatkan jenis tersebut bersarang pada jarak
152 m dari pintu gua, suhu 25oC dan kelembapan 85%.
Kesimpulan
Dari hasil penelitian ini dapat disimpulkan :
1. Jenis-jenis kelelawar yang bersarang di gua-gua Karst Gombong ada 15 jenis,
yaitu 4 jenis Megachiroptera terdiri atas Cynopterus horsfieldii Gay, 1843;
Cynopterus brachyotis (Muller, 1838); Rousettus amplexicaudatus (Geoffroy,
1810); Eonycteris spelaea (Dobson,1871), dan 11 jenis Microchiroptera
terdiri atas Chaerophon plicata (Buchannan,1800), Hipposideros ater
Templeton, 1848; Hipposideros cf.ater; Hipposideros sp; Hipposideros
bicolor (Temminck, 1834); Hipposideros sorenseni Kitchener & Maryanto,
1993; Hipposideros diadema (Geoffroy,1813); Rhinilophus borneensis
Peters, 1861; Rhinolophus affinis Horsfield, 1823; Miniopterus australis
Tomes, 1858; dan Miniopterus schreibersii (Kuhl, 1819). Kelimpahan,
indeks keanekaragaman jenis dan kemerataan jenis kelelawa di gua-gua Karst
Gombong berkorelasi nyata dengan panjang lorong gua, lebar gua, dan tinggi
lorong gua. Semakin panjang, lebar, dan tinggi lorong gua, maka semakin
tinggi kelimpahan, keanekaragaman jenis, dan kemerataan jenis
kelelawarnya. Sementara jumlah pintu gua dan jumlah ventilasi gua tidak
berkorelasi nyata dengan struktur komunitas kelelawar.
2. Parameter fisik yang paling berpengaruh pada pemilihan sarang kelelawar
berturut-turut adalah : intensitas suara, jarak dari mulut gua, suhu,
kelembapan, dan intensitas cahaya. Berdasarkan perameter tersebut terdapat
5 kelompok kelelawar yang memililih tempat bersarang dengan pola yang
spesifik.

56
Saran
1. Karst Gombong perlu diusulkan sebagai kawasan konservasi, mengingat
kawasan karst tersebut memiliki keanekaragaman jenis kelelawar yang tinggi.
2. Perlu dibuat zonasi pemanfaatan ruang gua berdasarkan pola pemilihan
sarang sesuai dengan informasi ilmiah yang didapat dari penelitian ini.

57
ANALISIS RELUNG PAKAN
KELELAWAR INSEKTIVORA (Subordo: Microchiroptera)
DAN KELELAWAR FRUGIVORA (Subordo: Megachiroptera)
YANG BERSARANG DI GUA-GUA KARST GOMBONG
KABUPATEN KEBUMEN JAWA TENGAH
ABSTRACT
The research aims to determine the diet of cave dwelling bats in Gombong Karst Area, Kebumen, Central Java. The research was done in June 2009 to March 2010. Stomach contents was collected and dissolved in aquadest. The material of insect were sorted and identified under microscope and compare to the insects that were collected by light trap in bat foraging area. The polen were collected from the intestine of fruit bats. The data were analyzed by Principle Component Analysis (PCA), chi square and niche overlap index. There were found eleven species of insectivorous bat and five species of frugivorous bat (four of those frugivorous bats roost in cave). The insects in gut content of insectivorous bats belong to 10 orders, distributed into 29 families. Otherwise, pollen in gut content of frugivorous bats belong to 9 families, distributed into 33 species of plant. Based on prey preference, the insectivorous bats can be classified into four goups. Based on flower and polen preference, the frugivorous bats can be classified into three goups. The niche overlap index between species of bats that occupy in one cave was less than 30%.
Key words : bat, insectivorus, frugivorous, cave,niche overlap index.
Pendahuluan
Ketersediaan pakan menentukan kelimpahan dan keberadaan kelelawar di
suatu habitat (Feeler & Pierson, 2002; Russso et al. 2003). Oleh karena itu,
kelelawar cenderung memilih sarang yang dekat dengan sumber pakan atau
memiliki akses pada sumber pakan. Hal ini dibuktikan oleh beberapa hasil
penelitian terdahulu. Menurut Law & Chidel (2002), Kerivoula papuensis
(Microchiroptera) bersarang pada jarak maksimum 2.1 km dari tempat pencarian
makannya di hutan hujan di New South Wales. Hodgkison et al. (2004)
membuktikan Balionycteris maculata mencari makan pada jarak satu kilometer di
sekitar sarangnya. Penelitian Agosta (2002) membuktikan Eptesicus fuscus
memilih sarang di dekat permukiman manusia, dan hasil analisis isi perut
menunjukkan bahwa makanannya adalah serangga yang banyak terdapat di

58
sekitar lampu penerangan permukiman tersebut. Demikian pula halnya dengan
kelelawar yang bersarang di gua-gua karst. Menurut Furman & Ozgul (2002),
selain memenuhi persyaratan fisik yang sesuai dengan tubuh kelelawar, gua yang
dipilih oleh kelelawar harus memiliki akses pada sumber pakan.
Hasil penelitian lain mengenai sarang kelelawar di gua-gua karst
menunjukkan bahwa satu gua dapat dihuni oleh beberapa jenis kelelawar. Duran
& Centano (2002) mendapatkan kelelawar insektivora Pteronotus quadridiens
dan Erophylla quadridiens bersarang dalam satu gua di Gua Bonita, Los Perez.
Furman & Ozgul (2002) mendapatkan tiga sampai lima jenis kelelawar bersarang
dalam satu gua di Istambul Turki. Apriandi et al. (2008) mendapatkan tiga
sampai delapan jenis kelelawar bersarang dalam satu gua di Karst Gudawang
Bogor. Mengingat gua tersebut harus memiliki akses pada sumber makanannya,
diduga jenis-jenis kelelawar yang bersarang dalam gua yang sama mencari makan
di tempat yang sama. Sejauh ini belum ada penelitian yang menjelaskan
pembagian relung pakan kelelawar yang bersarang di gua karst dan seberapa
besar kompetisi pakan di antara jenis-jenis kelelawar yang bersarang dalam satu
gua tersebut. Informasi dasar ini sangat diperlukan dalam upaya melestarikan
kelelawar. Terutama dalam kaitannya dengan pengelolaan gua sebagai habitat
sarang dan pengelolaan habitat pencarian makannya.
Menurut Chairunnisa (1997) diduga Microchiroptera berasal dari nenek
moyang Rodensia yang cenderung insektivora, sedangkan Megachiroptera berasal
dari nenek moyang Primata yang cenderung frugivora. Hal ini didukung oleh
pendapat Altringham (1996) bahwa anggota subordo Microchiroptera sebagian
besar adalah pemakan serangga, sedangkan anggota subordo Megachiroptera
makanan utamanya adalah buah (frugivora). Selain itu menurut Nowak (1994)
selain memakan buah sebagai pakan utamanya, kelelawar Megachiroptera juga
memakan serbuk sari (polen) dan madu sebagi pakan tambahan (sumber protein).
Oleh karena itu, relung pakan kelelawar Microchiroptera dapat dianalisis
berdasarkan serangga yang dimakan oleh kelelawar Microchiroptera, dan relung
pakan Megachiroptera dapat dianalisis berdasarkan polen yang dimakan oleh
Megachiroptera.

59
Tujuan penelitian ini adalah : mengidentifikasi pakan kelelawar insektifora
dan kelelawar frugivora penghuni gua dan menentukan kesamaan relung pakan
kelelawar yang berasosisasi dalam satu gua yang sama. Hipotesis penelitian ini
adalah : asosiasi bersarang dalam satu gua oleh beberapa jenis kelelawar dapat
dilakukan oleh jenis-jenis yang memiliki indeks kesamaan relung pakan kecil (<
50%).
Bahan dan Metode
Penelitian dilakukan pada bulan Juni 2009 s/d Maret 2010 di sepuluh
gua yang dihuni kelelawar di Kawasan Karst Gombong Selatan, Kabupaten
Kebumen, Jawa Tengah, yaitu: Gua Celeng, Gua Dempo, Gua Inten, Gua
Jatijajar, Gua Kemit, Gua Liyah, Gua Macan, Gua Petruk, Gua Sigong, dan Gua
Tratag. Identifikasi sampel dilakukan di Laboratorium Hewan Pusat Penelitian
Sumberdaya Hayati dan Biodiversitas (PPSHB) IPB Bogor.
Bahan yang digunakan adalah chloroform, alkohol 70%, gliserin dan cat
kuku. Alat yang digunakan adalah: mist net, harpa trap, hand net, insect net, light
trap, alat bedah, mikroskop cahaya merk olympus, mikrometer merk olympus,
sentrifuge, kaca objek, kaca penutup, kaca arloji, pipet, pinset, gelas ukur,
cawan petri, tabung reaksi, lampu spiritus, kertas hisap, dan kertas label.
Pengambilan sampel kelelawar Microchiroptera dilakukan di setiap sarang,
masing-masing sebanyak 5 s/d 10 individu. Sampel diambil dengan
menggunakan hand net, mist net ataupun harpa trap sesaat setelah kelelawar
kembali dari pencarian makan (pukul 05.00 s/d 06.00 WIB). Kelelawar yang
tertangkap segera dibius dan dimasukkan ke dalam alkohol 70%. Pengambilan
sampel Megachiroptera dilakukan di lokasi pencarian makan, hal ini untuk
mencegah tindakan destruktif, mengingat jumlah individu Megachiroptera yang
bersarang di gua hanya berkisar 2 s/d 6 individu saja. Apabila sampel diambil di
lokasi sarang, dikhawatirkan jumlahnya semakin berkurang. Sampel diambil di
tiga lokasi pencarian makan (di persawahan Desa Ayah; di hutan lindung
Candirenggo; dan di sekitar permukiman Desa Jatijajar) sebanyak 5 s/d 10
individu setiap jenis dengan menggunakan mist net dan harpa trap.
Penangkapan serangga sebagai bahan pembanding dalam proses identifikasi
isi perut Microchiroptera, dilakukan dengan menggunakan light trap yang

60
dipasang di tiga lokasi pencarian makan kelelawar, yaitu di persawahan Desa
Jatijajar, di hutan lindung Desa Candi Renggo, dan di sekitar permukiman Desa
Jatijajar.
Pengamatan isi perut dilakukan dengan cara: 1) Sampel yang tertangkap
segera dibius dengan chloroform dan direndam dalam alkohol 70%; 2) Sampel
dibedah viseral, dan saluran pencernaan mulai dari kerongkongan (osefagus)
hingga anus dikeluarkan dari tubuh; 3) Isi saluran pencernaan dikumpulkan pada
cawan petri dan dilarutkan dengan aquadest sampai terpisah antara lemak,
kotoran, dan sisa-sisa tubuh serangga; 4) Sisa-sisa tubuh serangga diamati di
bawah mikroskop perbesaran 10 X 10 kemudian diidentifikasi dengan kunci
identifikasi serangga oleh Whitaker (1988) dan kunci identifikasi serangga
(Borror et al. 1996), serta dibandingkan dengan serangga hasil koleksi di area
pencarian makan kelelawar.
Identifkasi isi perut Megachiroptera dilakukan dengan cara (Soegiharto et
al. 2010): 1) Sampel kelelawar dibedah dan saluran pencernaan dikeluarkan dari
tubuh; 2) Isi saluran pencernaan ditampung di cawan petri dan direndam dalam
alkohol 70%, kemudian disentrifugasi dengan putaran 2000 rpm selama 30
menit; 3) Cairan alkohol yang terpisah dengan endapan isi perut dibuang dan
diganti dengan alkohol yang baru. Sentrifugasi dan pencucian dengan alkohol ini
dilakukan sebanyak tiga kali; 4) Endapan yang dihasilkan diletakan di gelas objek
sebanyak satu tetes, ditetesi dengan gliserol, kemudian ditutup dengan kaca
penutup. Pada bagian tepinya direkatkan menggunakan cat kuku; 5) Dilakukan
identifikasi polen di bawah mikroskop dengan perbesaran 100x dan 400x dengan
menggunakan buku kunci identifikasi polen (Erdtman 1952).
Analisis data
Transformasi data dilakukan untuk mengubah skala pengukuran data asli
menjadi bentuk lain sehingga data dapat memenuhi asumsi-asumsi yang
mendasari analisis ragam. Pada penelitian ini digunakan transformasi arcsin (n +
0.5) dikarenakan data tersaji dalam proporsi (persentase) dan banyak data yang
bernilai nol (Syahid 2009)

61
Variasi pakan kelelawar dianalisis menggunakan analisis statistik
multivariat yang didasarkan pada analisis komponen utama (Principal Component
Analysis) (Ludwig & Reynolds 1988). Analisis Komponen Utama merupakan
metode statistik deskriptif yang bertujuan untuk menampilkan dalam bentuk
grafik, maksimum informasi yang terdapat dalam suatu matriks data. Matriks data
yang dimaksud terdiri atas jenis kelelawar (pada baris) dan jenis pakan sebagai
variabel kuantitatif (kolom). Analisis Komponen Utama ini menggunakan
software SPSS versi 15.
HCCA (Hybrid Canonical Correspondence Analysis) digunakan untuk
menganalisis kecenderungan kelelawar Megachiroptera memilih pakannya
berdasarkan jenis tanaman, tipe mahkota bunga dan ukuran polen. Untuk analisis
HCCA digunakan software canoco for windows 4.5 ( Leps & Smilauer 1999).
Uji chi kuadrat (chi square) digunakan untuk mengetahui apakah terdapat
perbedaan yang nyata antara jenis pakan kelelawar satu dengan lainnya. Hipotesis
yang digunakan adalah:
H0 = pakan kelelawar satu dengan yang lain tidak berbeda nyata.
H1= pakan kelelawar satu dengan yang lain berbeda nyata.
Nilai chi square dihitung dengan rumus (Ludwig & Reynold 1988):
Keterangan:
X2 = chi square
Xi = banyaknya jenis serangga pakan pada kelelawar jenis ke i
µi = banyaknya jenis serangga yang diharapkan pada kelelawar jenis-i
Jika X2 hitung < X2 tabel maka H0 diterima, jika X2 hitung > X2 tabel maka H1
diterima. Uji chi square dilakukan dengan bantuan software SPSS versi 15.
Metode analisis yang digunakan untuk menganalisis kesamaan relung pakan
kelelawar adalah metode niche overlap index (Ludwig & Reynolds 1988), yaitu
metode yang didasarkan pada kesamaan proporsi jenis pakan yang ditemukan di
saluran pencernaan hewan yang dibandingkan. Persamaan yang digunakan adalah
persamaan simplified Morsita index dengan rumus sebagai berikut (Ludwig &
Reynolds 1988):
X2 = ∑ (Xi - µi)2
µi

62
2∑ PijPik
CH = ∑ Pij
2 + Pik
2
Keterangan: CH = indeks simplified morisita antara kelelawar jenis ke-j dan kelelawar jenis
ke-k Pij = proporsi famili serangga yang digunakan oleh kelelawar jenis ke-j (n/N) Pik = proporsi famili serangga yang digunakan oleh kelelawar jenis ke-k (n/N) n = jumlah famili serangga seluruhnya Penghitungan nilai indeks kesamaan relung pakan ini menggunakan software
ecological methodology versi 5.1.
Hasil
Relung pakan kelelawar insektivora
Hasil analisis isi perut, ditemukan 29 famili serangga yang termasuk dalam
sepuluh ordo. Kelelawar jenis H. sorenseni memakan paling banyak serangga dari
famili berbeda ( 17 famili); diikuti oleh Hipposideros sp (8 famili); M.
schreibersii (6 famili); C. plicata (8 famili); H. bicolor (5 famili); M.australis (6
famili); R.affinis (5 famili) H.diadema (5 famili); H.ater (4 famili); H.cf. ater (4
famili); dan R. borneensis (1 famili). Serangga yang ditemukan dan proporsi
temuan pada setiap jenis kelelawar di semua sarang tersaji pada Tabel 7.
Berdasarkan hasil pengamatan terhadap serangga yang dikoleksi di lokasi
pencarian makan kelelawar (Lampiran 3), serta hasil studi pustaka berdasarkan
Aguirre et al. (2002), dapat diketahui karakteristik serangga pakan yang meliputi
bobot tubuh, panjang tubuh, dan kekerasan eksoskeleton serangga pakan.
Karakteristik serangga pakan dapat dilihat pada Tabel 8.
Hasil Principle Component Analysis (PCA) dengan 3 komponen utama
dapat menggambarkan 85% dari varian total. Komponen 1,2,dan 3 masing masing
menerangkan 47.5%, 23.5%, 14.0%. Nilai loading faktor dari masing masing
komponen dapat dilihat pada Lampiran 6. Penggambaran hasil PCA dengan 3
komponen utama dapat dilihat pada Gambar 20.
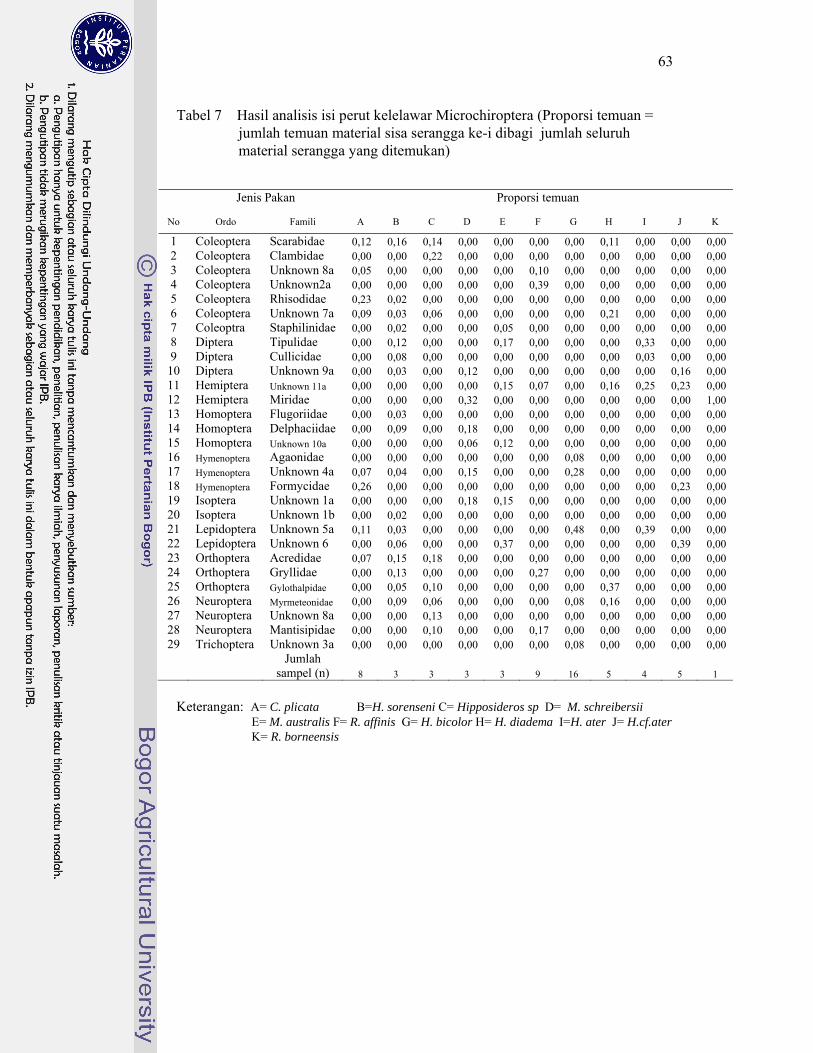
63
Tabel 7 Hasil analisis isi perut kelelawar Microchiroptera (Proporsi temuan = jumlah temuan material sisa serangga ke-i dibagi jumlah seluruh material serangga yang ditemukan)
Keterangan: A= C. plicata B=H. sorenseni C= Hipposideros sp D= M. schreibersii E= M. australis F= R. affinis G= H. bicolor H= H. diadema I=H. ater J= H.cf.ater K= R. borneensis
Jenis Pakan Proporsi temuan
No Ordo Famili A B C D E F G H I J K
1 Coleoptera Scarabidae 0,12 0,16 0,14 0,00 0,00 0,00 0,00 0,11 0,00 0,00 0,00 2 Coleoptera Clambidae 0,00 0,00 0,22 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 3 Coleoptera Unknown 8a 0,05 0,00 0,00 0,00 0,00 0,10 0,00 0,00 0,00 0,00 0,004 Coleoptera Unknown2a 0,00 0,00 0,00 0,00 0,00 0,39 0,00 0,00 0,00 0,00 0,00 5 Coleoptera Rhisodidae 0,23 0,02 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 6 Coleoptera Unknown 7a 0,09 0,03 0,06 0,00 0,00 0,00 0,00 0,21 0,00 0,00 0,007 Coleoptra Staphilinidae 0,00 0,02 0,00 0,00 0,05 0,00 0,00 0,00 0,00 0,00 0,00 8 Diptera Tipulidae 0,00 0,12 0,00 0,00 0,17 0,00 0,00 0,00 0,33 0,00 0,00 9 Diptera Cullicidae 0,00 0,08 0,00 0,00 0,00 0,00 0,00 0,00 0,03 0,00 0,00
10 Diptera Unknown 9a 0,00 0,03 0,00 0,12 0,00 0,00 0,00 0,00 0,00 0,16 0,00 11 Hemiptera Unknown 11a 0,00 0,00 0,00 0,00 0,15 0,07 0,00 0,16 0,25 0,23 0,00 12 Hemiptera Miridae 0,00 0,00 0,00 0,32 0,00 0,00 0,00 0,00 0,00 0,00 1,0013 Homoptera Flugoriidae 0,00 0,03 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 14 Homoptera Delphaciidae 0,00 0,09 0,00 0,18 0,00 0,00 0,00 0,00 0,00 0,00 0,00 15 Homoptera Unknown 10a 0,00 0,00 0,00 0,06 0,12 0,00 0,00 0,00 0,00 0,00 0,0016 Hymenoptera Agaonidae 0,00 0,00 0,00 0,00 0,00 0,00 0,08 0,00 0,00 0,00 0,00 17 Hymenoptera Unknown 4a 0,07 0,04 0,00 0,15 0,00 0,00 0,28 0,00 0,00 0,00 0,00 18 Hymenoptera Formycidae 0,26 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,23 0,0019 Isoptera Unknown 1a 0,00 0,00 0,00 0,18 0,15 0,00 0,00 0,00 0,00 0,00 0,00 20 Isoptera Unknown 1b 0,00 0,02 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 21 Lepidoptera Unknown 5a 0,11 0,03 0,00 0,00 0,00 0,00 0,48 0,00 0,39 0,00 0,0022 Lepidoptera Unknown 6 0,00 0,06 0,00 0,00 0,37 0,00 0,00 0,00 0,00 0,39 0,00 23 Orthoptera Acredidae 0,07 0,15 0,18 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 24 Orthoptera Gryllidae 0,00 0,13 0,00 0,00 0,00 0,27 0,00 0,00 0,00 0,00 0,00 25 Orthoptera Gylothalpidae 0,00 0,05 0,10 0,00 0,00 0,00 0,00 0,37 0,00 0,00 0,00 26 Neuroptera Myrmeteonidae 0,00 0,09 0,06 0,00 0,00 0,00 0,08 0,16 0,00 0,00 0,00 27 Neuroptera Unknown 8a 0,00 0,00 0,13 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 28 Neuroptera Mantisipidae 0,00 0,00 0,10 0,00 0,00 0,17 0,00 0,00 0,00 0,00 0,00 29 Trichoptera Unknown 3a 0,00 0,00 0,00 0,00 0,00 0,00 0,08 0,00 0,00 0,00 0,00
Jumlah sampel (n) 8 3 3 3 3 9 16 5 4 5 1
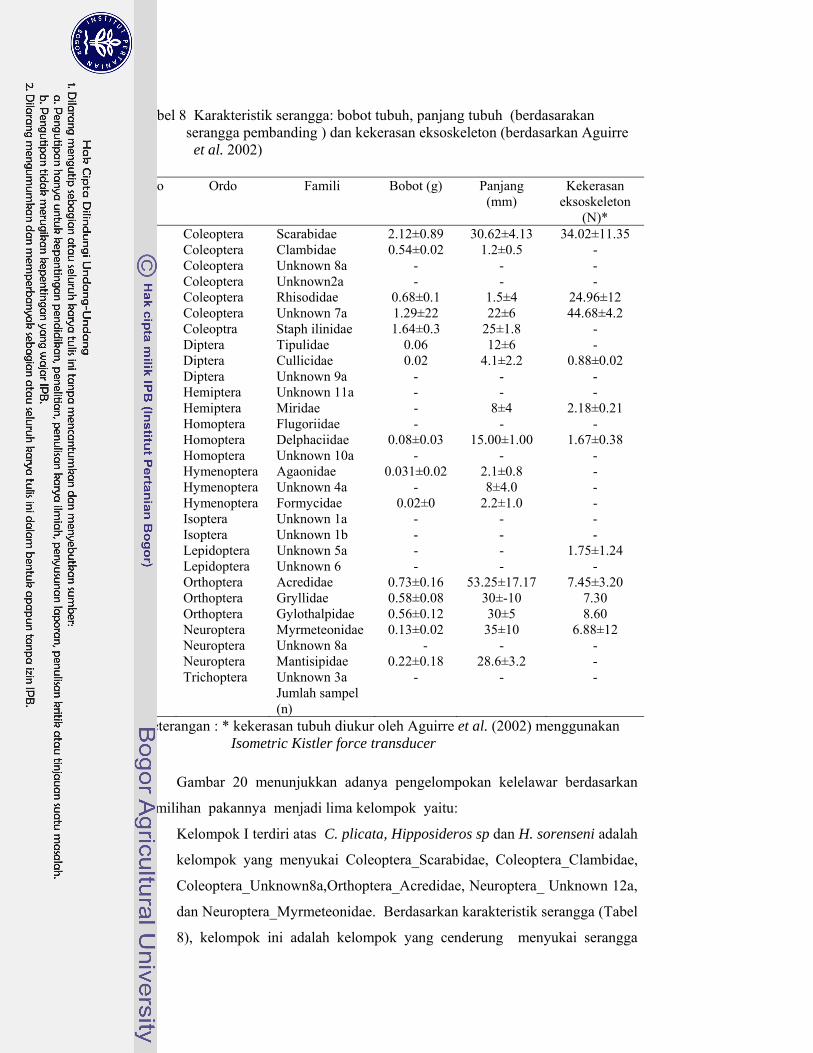
64
Tabel 8 Karakteristik serangga: bobot tubuh, panjang tubuh (berdasarakan serangga pembanding ) dan kekerasan eksoskeleton (berdasarkan Aguirre et al. 2002)
Keterangan : * kekerasan tubuh diukur oleh Aguirre et al. (2002) menggunakan Isometric Kistler force transducer
Gambar 20 menunjukkan adanya pengelompokan kelelawar berdasarkan
pemilihan pakannya menjadi lima kelompok yaitu:
1. Kelompok I terdiri atas C. plicata, Hipposideros sp dan H. sorenseni adalah
kelompok yang menyukai Coleoptera_Scarabidae, Coleoptera_Clambidae,
Coleoptera_Unknown8a,Orthoptera_Acredidae, Neuroptera_ Unknown 12a,
dan Neuroptera_Myrmeteonidae. Berdasarkan karakteristik serangga (Tabel
8), kelompok ini adalah kelompok yang cenderung menyukai serangga
No Ordo Famili Bobot (g) Panjang (mm)
Kekerasan eksoskeleton
(N)* 1 Coleoptera Scarabidae 2.12±0.89 30.62±4.13 34.02±11.35 2 Coleoptera Clambidae 0.54±0.02 1.2±0.5 - 3 Coleoptera Unknown 8a - - - 4 Coleoptera Unknown2a - - - 5 Coleoptera Rhisodidae 0.68±0.1 1.5±4 24.96±12 6 Coleoptera Unknown 7a 1.29±22 22±6 44.68±4.2 7 Coleoptra Staph ilinidae 1.64±0.3 25±1.8 - 8 Diptera Tipulidae 0.06 12±6 - 9 Diptera Cullicidae 0.02 4.1±2.2 0.88±0.02 10 Diptera Unknown 9a - - - 11 Hemiptera Unknown 11a - - - 12 Hemiptera Miridae - 8±4 2.18±0.21 13 Homoptera Flugoriidae - - - 14 Homoptera Delphaciidae 0.08±0.03 15.00±1.00 1.67±0.38 15 Homoptera Unknown 10a - - - 16 Hymenoptera Agaonidae 0.031±0.02 2.1±0.8 - 17 Hymenoptera Unknown 4a - 8±4.0 - 18 Hymenoptera Formycidae 0.02±0 2.2±1.0 - 19 Isoptera Unknown 1a - - - 20 Isoptera Unknown 1b - - - 21 Lepidoptera Unknown 5a - - 1.75±1.24 22 Lepidoptera Unknown 6 - - - 23 Orthoptera Acredidae 0.73±0.16 53.25±17.17 7.45±3.20 24 Orthoptera Gryllidae 0.58±0.08 30±-10 7.30 25 Orthoptera Gylothalpidae 0.56±0.12 30±5 8.60 26 Neuroptera Myrmeteonidae 0.13±0.02 35±10 6.88±12 27 Neuroptera Unknown 8a - - - 28 Neuroptera Mantisipidae 0.22±0.18 28.6±3.2 - 29 Trichoptera Unknown 3a - - -
Jumlah sampel (n)

65
berukuran besar (bobot > 0.5 g, panjang > 20 mm) dan keras (kekerasan
eksoskeleton > 7.5 N)
2. Kelompok II terdiri atas H.diadema adalah kelelawar yang menyukai
Coleoptera_Unknown 7a dan Orthoptera_Gryllothalpidae Berdasarkan
karakteristik serangga (Tabel 8), kelompok ini adalah kelompok yang sama
dengan kelompok I, cenderung menyukai serangga berukuran besar (bobot
> 0.5 g, panjang > 20 mm) dan keras (kekerasan eksoskeleton > 7.5 N).
3. Kelompok III terdiri atas M. schreibersii, M. australis, H.cf.ater, R.
borneensis adalah kelompok yang menyukai Diptera_unknown 9a,
Hemiptera_Unknown 11a, dan Lepidoptera_Unknown 6a. dan Isoptera_
Unknown 1a . Berdasarkan karakteristik serangga (Tabel 8), kelompok ini
adalah kelompok yang cenderung menyukai serangga berukuran sedang
(panjang 5mm s/d 20 mm) dan lunak (kekerasan eksoskeleton < 7.5 N)
4. Kelompok IV terdiri atas R. affinis adalah kelompok yang menyukai
Coleoptera_Unknown 8a, Coleoptera_Unknown 2a, Orthoptera_Gryllidae
dan Neuroptera_Mantisipidae. Berdasarkan karakteristik serangga (Tabel
8), kelompok ini cenderung menyukai serangga berukuran kecil (bobot <
0.5 g, panjang < 20 mm) dan keras (kekerasan eksoskeleton > 7.5 N).
5. Kelompok V terdiri atas H. bicolor dan H. ater adalah kelompok kelelawar
yang menyukai Lepidoptera Unknown 5a, Trichoptera_unknown 3a,
Hymenoptera_Unknown4a, Hymenoptera _agaonidae. Berdasarkan
karakteristik serangga (Tabel 8), kelompok cenderung menyukai serangga
berukuran kecil ( panjang < 5 mm) dan lunak (kekerasan eksoskeleton < 7.5
N)
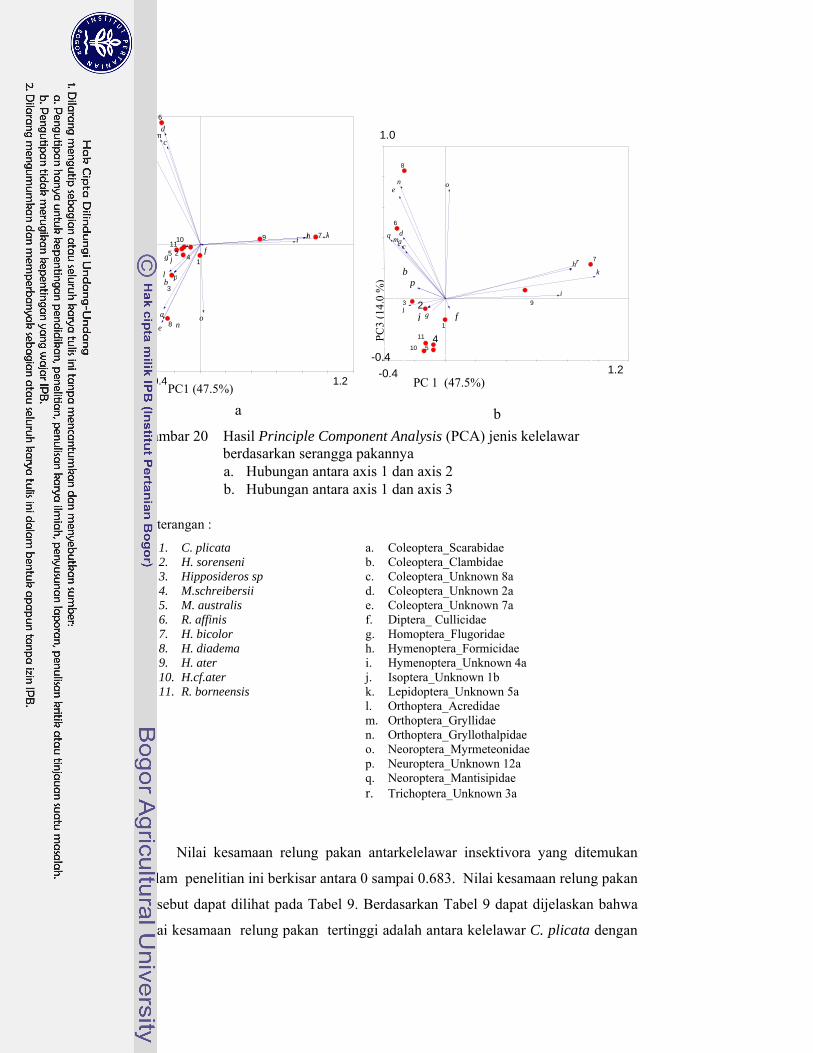
66
Gambar 20 Hasil Principle Component Analysis (PCA) jenis kelelawar berdasarkan serangga pakannya a. Hubungan antara axis 1 dan axis 2 b. Hubungan antara axis 1 dan axis 3
Keterangan :
1. C. plicata 2. H. sorenseni 3. Hipposideros sp 4. M.schreibersii 5. M. australis 6. R. affinis 7. H. bicolor 8. H. diadema 9. H. ater 10. H.cf.ater 11. R. borneensis
a. Coleoptera_Scarabidae b. Coleoptera_Clambidae c. Coleoptera_Unknown 8a d. Coleoptera_Unknown 2a e. Coleoptera_Unknown 7a f. Diptera_ Cullicidae g. Homoptera_Flugoridae h. Hymenoptera_Formicidae i. Hymenoptera_Unknown 4a j. Isoptera_Unknown 1b k. Lepidoptera_Unknown 5a l. Orthoptera_Acredidae m. Orthoptera_Gryllidae n. Orthoptera_Gryllothalpidae o. Neoroptera_Myrmeteonidae p. Neuroptera_Unknown 12a q. Neoroptera_Mantisipidae r. Trichoptera_Unknown 3a
Nilai kesamaan relung pakan antarkelelawar insektivora yang ditemukan
dalam penelitian ini berkisar antara 0 sampai 0.683. Nilai kesamaan relung pakan
tersebut dapat dilihat pada Tabel 9. Berdasarkan Tabel 9 dapat dijelaskan bahwa
nilai kesamaan relung pakan tertinggi adalah antara kelelawar C. plicata dengan
-0.4 1.2-0.4
1.0
a
b
c
d
e
fg
h
i
j
k
l
m
n o
p
q
r
1
23
45
6
7
8
9
10
11
PC 1 (47.5%) -0.4 1.2
-1.0
1.0
a
b
c
d
e
f g
h i
j
k
l
m
no
p
q
r
12
3
4 5
6
7
8
910 11
PC 2
(23.
5 %
)
PC1 (47.5%) PC
3 (1
4.0
%)
ba
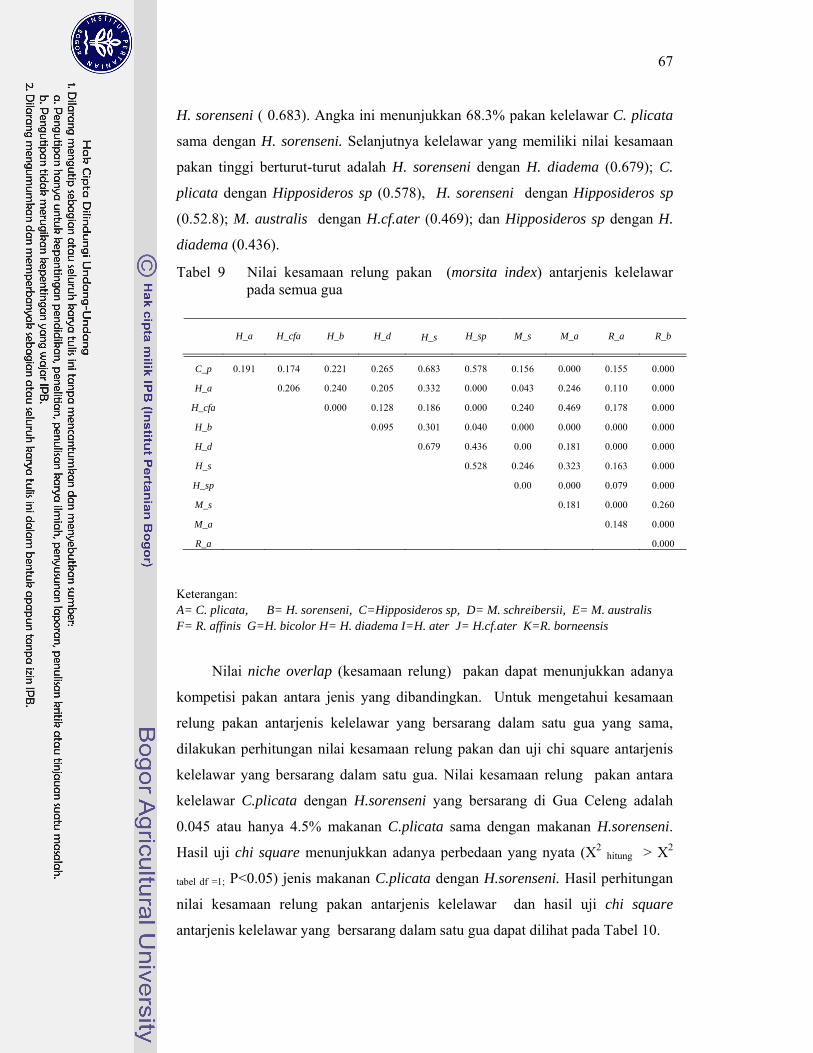
67
H. sorenseni ( 0.683). Angka ini menunjukkan 68.3% pakan kelelawar C. plicata
sama dengan H. sorenseni. Selanjutnya kelelawar yang memiliki nilai kesamaan
pakan tinggi berturut-turut adalah H. sorenseni dengan H. diadema (0.679); C.
plicata dengan Hipposideros sp (0.578), H. sorenseni dengan Hipposideros sp
(0.52.8); M. australis dengan H.cf.ater (0.469); dan Hipposideros sp dengan H.
diadema (0.436).
Tabel 9 Nilai kesamaan relung pakan (morsita index) antarjenis kelelawar pada semua gua
H_a
H_cfa
H_b
H_d
H_s
H_sp
M_s
M_a
R_a
R_b
C_p 0.191 0.174 0.221 0.265 0.683 0.578 0.156 0.000 0.155 0.000
H_a 0.206 0.240 0.205 0.332 0.000 0.043 0.246 0.110 0.000
H_cfa 0.000 0.128 0.186 0.000 0.240 0.469 0.178 0.000
H_b 0.095 0.301 0.040 0.000 0.000 0.000 0.000
H_d 0.679 0.436 0.00 0.181 0.000 0.000
H_s 0.528 0.246 0.323 0.163 0.000
H_sp 0.00 0.000 0.079 0.000
M_s 0.181 0.000 0.260
M_a 0.148 0.000
R_a 0.000
Keterangan: A= C. plicata, B= H. sorenseni, C=Hipposideros sp, D= M. schreibersii, E= M. australis F= R. affinis G=H. bicolor H= H. diadema I=H. ater J= H.cf.ater K=R. borneensis
Nilai niche overlap (kesamaan relung) pakan dapat menunjukkan adanya
kompetisi pakan antara jenis yang dibandingkan. Untuk mengetahui kesamaan
relung pakan antarjenis kelelawar yang bersarang dalam satu gua yang sama,
dilakukan perhitungan nilai kesamaan relung pakan dan uji chi square antarjenis
kelelawar yang bersarang dalam satu gua. Nilai kesamaan relung pakan antara
kelelawar C.plicata dengan H.sorenseni yang bersarang di Gua Celeng adalah
0.045 atau hanya 4.5% makanan C.plicata sama dengan makanan H.sorenseni.
Hasil uji chi square menunjukkan adanya perbedaan yang nyata (X2 hitung > X2
tabel df =1; P<0.05) jenis makanan C.plicata dengan H.sorenseni. Hasil perhitungan
nilai kesamaan relung pakan antarjenis kelelawar dan hasil uji chi square
antarjenis kelelawar yang bersarang dalam satu gua dapat dilihat pada Tabel 10.
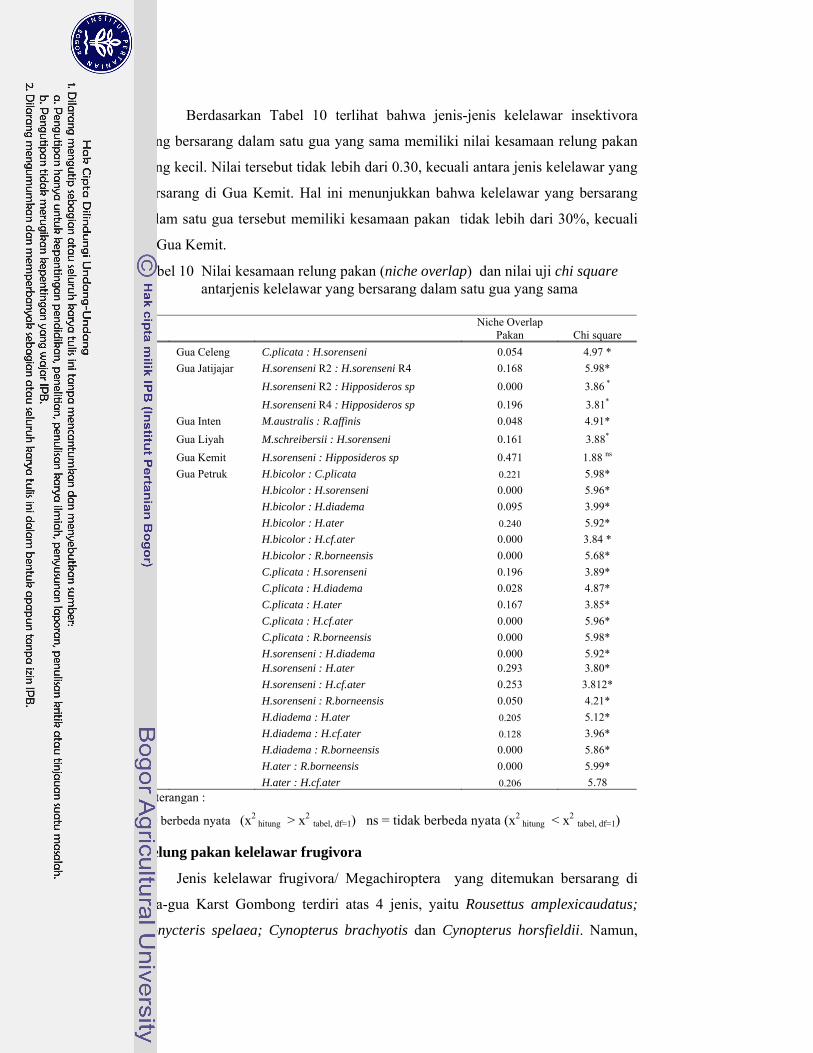
68
Berdasarkan Tabel 10 terlihat bahwa jenis-jenis kelelawar insektivora
yang bersarang dalam satu gua yang sama memiliki nilai kesamaan relung pakan
yang kecil. Nilai tersebut tidak lebih dari 0.30, kecuali antara jenis kelelawar yang
bersarang di Gua Kemit. Hal ini menunjukkan bahwa kelelawar yang bersarang
dalam satu gua tersebut memiliki kesamaan pakan tidak lebih dari 30%, kecuali
di Gua Kemit.
Tabel 10 Nilai kesamaan relung pakan (niche overlap) dan nilai uji chi square antarjenis kelelawar yang bersarang dalam satu gua yang sama
Niche Overlap Pakan Chi square
1 Gua Celeng C.plicata : H.sorenseni 0.054 4.97 * 2 Gua Jatijajar H.sorenseni R2 : H.sorenseni R4 0.168 5.98* H.sorenseni R2 : Hipposideros sp 0.000 3.86 * H.sorenseni R4 : Hipposideros sp 0.196 3.81* 3 Gua Inten M.australis : R.affinis 0.048 4.91* 4 Gua Liyah M.schreibersii : H.sorenseni 0.161 3.88* 5 Gua Kemit H.sorenseni : Hipposideros sp 0.471 1.88 ns 6 Gua Petruk H.bicolor : C.plicata 0.221 5.98* H.bicolor : H.sorenseni 0.000 5.96* H.bicolor : H.diadema 0.095 3.99* H.bicolor : H.ater 0.240 5.92* H.bicolor : H.cf.ater 0.000 3.84 * H.bicolor : R.borneensis 0.000 5.68* C.plicata : H.sorenseni 0.196 3.89* C.plicata : H.diadema 0.028 4.87* C.plicata : H.ater 0.167 3.85* C.plicata : H.cf.ater 0.000 5.96* C.plicata : R.borneensis 0.000 5.98* H.sorenseni : H.diadema 0.000 5.92* H.sorenseni : H.ater 0.293 3.80* H.sorenseni : H.cf.ater 0.253 3.812* H.sorenseni : R.borneensis 0.050 4.21* H.diadema : H.ater 0.205 5.12* H.diadema : H.cf.ater 0.128 3.96* H.diadema : R.borneensis 0.000 5.86* H.ater : R.borneensis 0.000 5.99* H.ater : H.cf.ater 0.206 5.78
Keterangan :
* = berbeda nyata (x2 hitung > x2 tabel, df=1) ns = tidak berbeda nyata (x2
hitung < x2 tabel, df=1) Relung pakan kelelawar frugivora
Jenis kelelawar frugivora/ Megachiroptera yang ditemukan bersarang di
gua-gua Karst Gombong terdiri atas 4 jenis, yaitu Rousettus amplexicaudatus;
Eonycteris spelaea; Cynopterus brachyotis dan Cynopterus horsfieldii. Namun,

69
untuk keperluan analisis relung pakan kelelawar Megachiroptera, sampel
ditangkap di lokasi pencarian makan (tiga lokasi). Hal ini karena jumlah individu
tiap sarang hanya 1 s/d 6 individu saja. Dalam penangkapan sampel tersebut, juga
berhasil ditangkap satu jenis kelelawar Megachiroptera yang tidak bersarang di
gua, yaitu Macroglossus sobrinus. Dengan demikian dalam analisis relung pakan
kelelawar Megachiroptera ini, terdapat 5 jenis kelelawar yang dianalisis relung
pakannya.
Setelah dilakukan identifikasi polen yang ditemukan di dalam saluran
pencernaan, diperoleh 33 species tumbuhan yang termasuk dalam sembilan famili.
Berdasarkan polen yang ditemukan, polen tumbuhan yang paling banyak
dikonsumsi adalah polen tumbuhan anggota famili Fabaceae (Acasia sp.),
Myrtaceae (Syzygium malaccense, Syzygium aqueum, Psidium guajava), dan
Bombacaceae (Ceiba sp1, Ceiba sp2). Persentase keberadaan polen pada saluran
pencernaan setiap jenis kelelawar frugivora dapat dilihat pada Tabel 11.
Hasil PCA berdasarkan polen yang ditemukan pada isi perut
Megachiroptera menggunakan 3 komponen utama dapat menjelaskan 88.536%
dari total varian. Komponen utama I menjelaskan 41.485%, kompenen utama II
menjelaskan 27.370%, komponen utama III menjelaskan 19.68%. Nilai loading
faktor dari masing masing komponen setelah direduksi menjadi 3 komponen
utama dapat dilihat pada pada Lampiran 7. Penggambaran hasil analisis PCA
dengan 3 axis tertera pada Gambar 21.
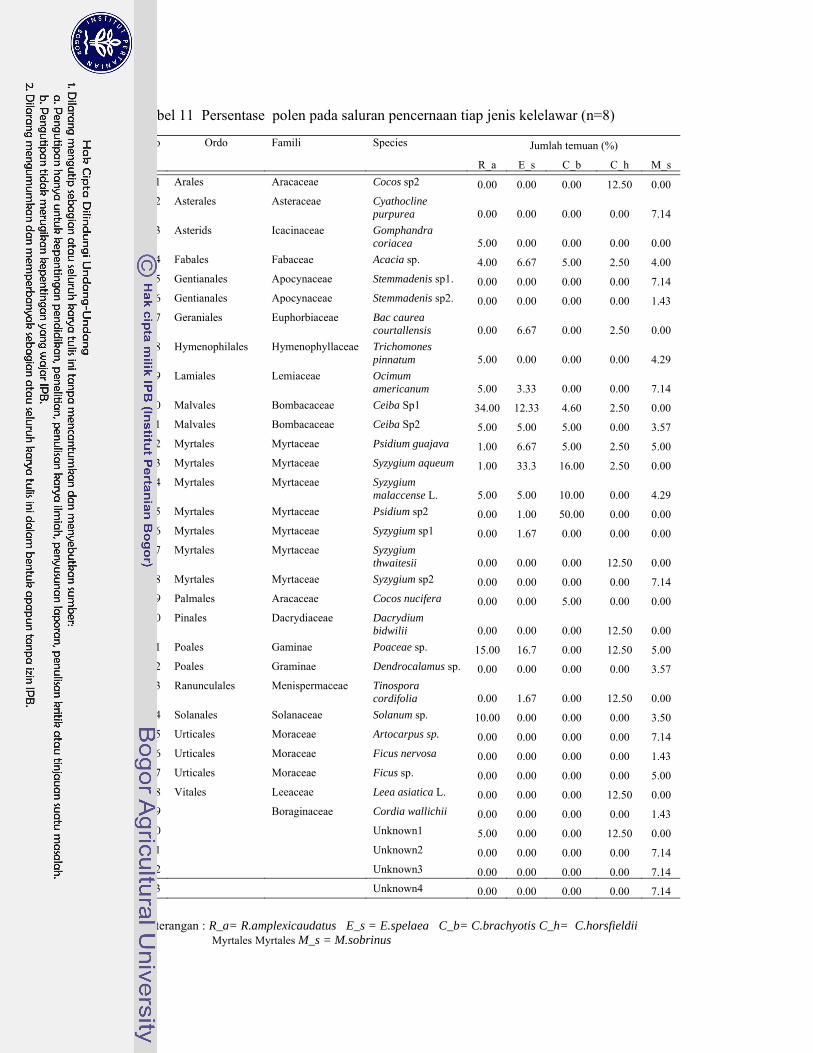
70
Tabel 11 Persentase polen pada saluran pencernaan tiap jenis kelelawar (n=8)
No Ordo Famili Species Jumlah temuan (%) R_a E_s C_b C_h M_s
1 Arales Aracaceae Cocos sp2 0.00 0.00 0.00 12.50 0.00 2 Asterales Asteraceae Cyathocline
purpurea 0.00 0.00 0.00 0.00 7.14 3 Asterids Icacinaceae Gomphandra
coriacea 5.00 0.00 0.00 0.00 0.00 4 Fabales Fabaceae Acacia sp. 4.00 6.67 5.00 2.50 4.00 5 Gentianales Apocynaceae Stemmadenis sp1. 0.00 0.00 0.00 0.00 7.14 6 Gentianales Apocynaceae Stemmadenis sp2. 0.00 0.00 0.00 0.00 1.437 Geraniales Euphorbiaceae Bac caurea
courtallensis 0.00 6.67 0.00 2.50 0.00 8 Hymenophilales Hymenophyllaceae Trichomones
pinnatum 5.00 0.00 0.00 0.00 4.299 Lamiales Lemiaceae Ocimum
americanum 5.00 3.33 0.00 0.00 7.1410 Malvales Bombacaceae Ceiba Sp1 34.00 12.33 4.60 2.50 0.00 11 Malvales Bombacaceae Ceiba Sp2 5.00 5.00 5.00 0.00 3.57 12 Myrtales Myrtaceae Psidium guajava 1.00 6.67 5.00 2.50 5.00 13 Myrtales Myrtaceae Syzygium aqueum 1.00 33.3 16.00 2.50 0.00 14 Myrtales Myrtaceae Syzygium
malaccense L. 5.00 5.00 10.00 0.00 4.29 15 Myrtales Myrtaceae Psidium sp2 0.00 1.00 50.00 0.00 0.00 16 Myrtales Myrtaceae Syzygium sp1 0.00 1.67 0.00 0.00 0.00 17 Myrtales Myrtaceae Syzygium
thwaitesii 0.00 0.00 0.00 12.50 0.00 18 Myrtales Myrtaceae Syzygium sp2 0.00 0.00 0.00 0.00 7.14 19 Palmales Aracaceae Cocos nucifera 0.00 0.00 5.00 0.00 0.00 20 Pinales Dacrydiaceae Dacrydium
bidwilii 0.00 0.00 0.00 12.50 0.00 21 Poales Gaminae Poaceae sp. 15.00 16.7 0.00 12.50 5.00 22 Poales Graminae Dendrocalamus sp. 0.00 0.00 0.00 0.00 3.57 23 Ranunculales Menispermaceae Tinospora
cordifolia 0.00 1.67 0.00 12.50 0.00 24 Solanales Solanaceae Solanum sp. 10.00 0.00 0.00 0.00 3.50 25 Urticales Moraceae Artocarpus sp. 0.00 0.00 0.00 0.00 7.14 26 Urticales Moraceae Ficus nervosa 0.00 0.00 0.00 0.00 1.43 27 Urticales Moraceae Ficus sp. 0.00 0.00 0.00 0.00 5.00 28 Vitales Leeaceae Leea asiatica L. 0.00 0.00 0.00 12.50 0.00 29 Boraginaceae Cordia wallichii 0.00 0.00 0.00 0.00 1.43 30 Unknown1 5.00 0.00 0.00 12.50 0.00 31 Unknown2 0.00 0.00 0.00 0.00 7.14 32 Unknown3 0.00 0.00 0.00 0.00 7.1433 Unknown4 0.00 0.00 0.00 0.00 7.14
Keterangan : R_a= R.amplexicaudatus E_s = E.spelaea C_b= C.brachyotis C_h= C.horsfieldii
Myrtales Myrtales M_s = M.sobrinus
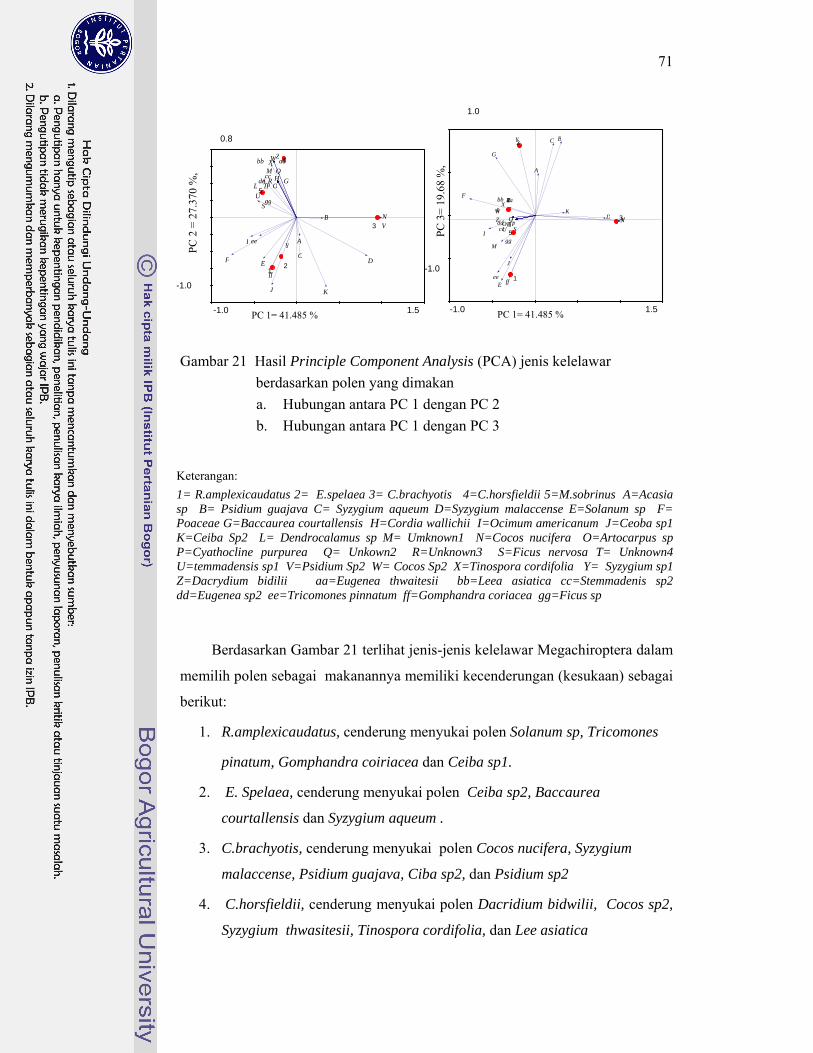
71
Gambar 21 Hasil Principle Component Analysis (PCA) jenis kelelawar
berdasarkan polen yang dimakan a. Hubungan antara PC 1 dengan PC 2 b. Hubungan antara PC 1 dengan PC 3
Keterangan: 1= R.amplexicaudatus 2= E.spelaea 3= C.brachyotis 4=C.horsfieldii 5=M.sobrinus A=Acasia sp B= Psidium guajava C= Syzygium aqueum D=Syzygium malaccense E=Solanum sp F= Poaceae G=Baccaurea courtallensis H=Cordia wallichii I=Ocimum americanum J=Ceoba sp1 K=Ceiba Sp2 L= Dendrocalamus sp M= Umknown1 N=Cocos nucifera O=Artocarpus sp P=Cyathocline purpurea Q= Unkown2 R=Unknown3 S=Ficus nervosa T= Unknown4 U=temmadensis sp1 V=Psidium Sp2 W= Cocos Sp2 X=Tinospora cordifolia Y= Syzygium sp1 Z=Dacrydium bidilii aa=Eugenea thwaitesii bb=Leea asiatica cc=Stemmadenis sp2 dd=Eugenea sp2 ee=Tricomones pinnatum ff=Gomphandra coriacea gg=Ficus sp
Berdasarkan Gambar 21 terlihat jenis-jenis kelelawar Megachiroptera dalam
memilih polen sebagai makanannya memiliki kecenderungan (kesukaan) sebagai
berikut:
1. R.amplexicaudatus, cenderung menyukai polen Solanum sp, Tricomones
pinatum, Gomphandra coiriacea dan Ceiba sp1.
2. E. Spelaea, cenderung menyukai polen Ceiba sp2, Baccaurea
courtallensis dan Syzygium aqueum .
3. C.brachyotis, cenderung menyukai polen Cocos nucifera, Syzygium
malaccense, Psidium guajava, Ciba sp2, dan Psidium sp2
4. C.horsfieldii, cenderung menyukai polen Dacridium bidwilii, Cocos sp2,
Syzygium thwasitesii, Tinospora cordifolia, dan Lee asiatica
-1.0 1.5
-1.0
1.0
A
BC
D
E
F
G
HI
J
K
L
M
N O PQ
R
S
T
UV
WX
Y
Z
aabb
ccdd
eeff
gg
1
2
34
5
PC 1= 41.485 %
PC 3
= 19
.68
%,
-1.0 1.5
-1.0
0.8
A
B
C DE F
G H
I
J K
L
M
N
OP
Q R
S
T U
V
W X
Y
Z aabb
cc dd
ee
ff
gg
1 2
3
4
5
PC 1= 41.485 %
PC 2
= 2
7.37
0 %
,

72
5. M. sobrinus, cenderung menyukai polen: Ficus nervosa, Stemmodenis
sp1, ficus sp, Dendrocalamus sp,Unknown4, Cyathocline purpurea,
Stemodenis sp2, dan Syzygium sp2.
Berdasarkan Van Steenis (2006), tipe bunga (bentuk kelopak bunga) dari 33
jenis tanaman yang dikunjungi oleh kelelawar Megachiroptera di atas, dapat
dikelompokkan menjadi delapan bentuk, yaitu tabung, lonceng, cawan, corong,
bulir majemuk, mangkuk, membulat, dan simetri labiatus (kupu-kupu). Tipe
bunga dari jenis-jenis tanaman yang dikunjungi kelelawar Megachiroptera di
Karst Gombong dapat dilihat pada Tabel 12.
Berdasarkan ukurannya, polen yang dikonsumsi oleh kelelawar
Megachiroptera yang ditemukan dalam penelitian ini, dapat dikelompokkan
menjadi polen berukuran kecil (diameter < 40 µm); polen berukuran sedang
(diameter antara 40 s/d 60 µm ) dan polen berukuran besar (diameter > 60 µm)
(Lampiran 4). Pengelompokan tanaman berdasarkan ukuran polennya dapat
dilihat pada Tabel 13.
Pengelompokan jenis kelelawar berdasarkan jenis tanaman dan bentuk
mahkota bunga berdasarkan HCCA dengan 3 axis dapat dilihat pada Gambar 22a
dan 22b. Variasi data yang dapat diterangkan adalah 90.5%. Axis 1 menerangkan
66.0%; axis = 11.6%, dan axis 3= 12.9%. HCCA jenis kelelawar berdasarkan
jenis tanaman dan ukuran polen dengan 3 axis dapat dilihat pada Gambar 23a dan
23b. Variasi data yang dapat diterangkan adalah 96.522%, axis 1 menerangkan
43.6%, axis 2 = 39.28%, dan axis 3 = 13.59%.
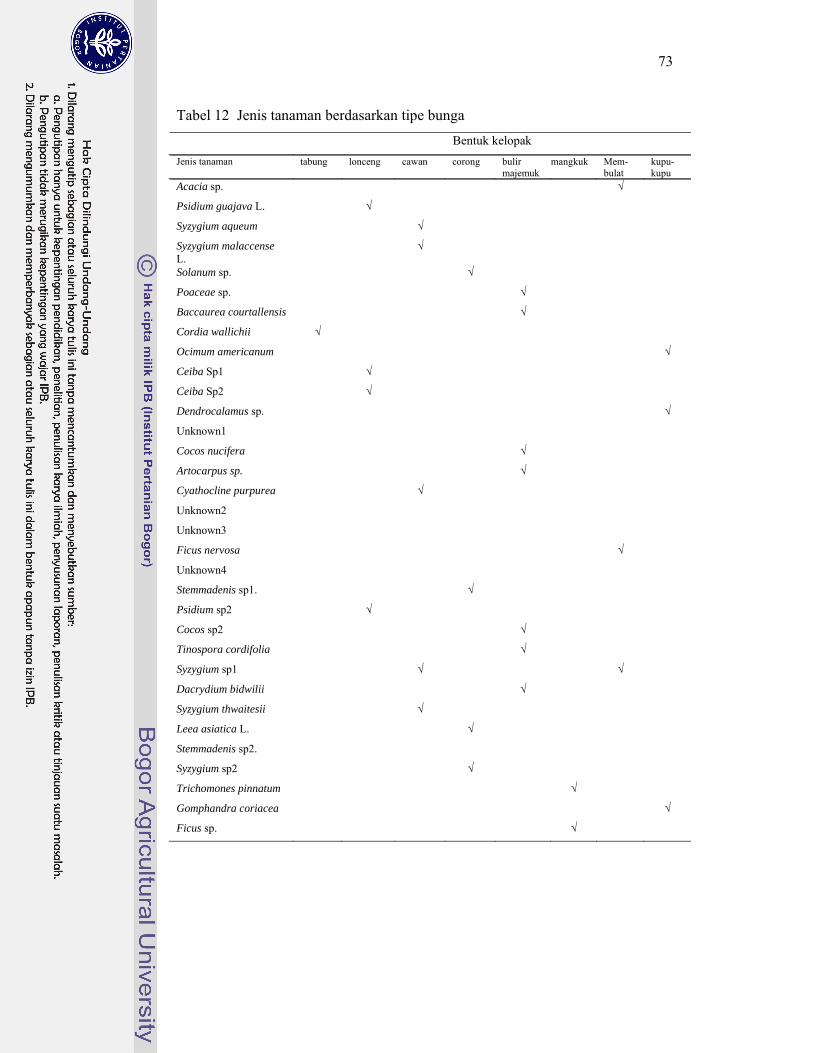
73
Tabel 12 Jenis tanaman berdasarkan tipe bunga Bentuk kelopak Jenis tanaman tabung lonceng cawan corong bulir
majemuk mangkuk Mem-
bulat kupu- kupu
Acacia sp. √
Psidium guajava L. √
Syzygium aqueum √
Syzygium malaccense L.
√
Solanum sp. √
Poaceae sp. √
Baccaurea courtallensis √
Cordia wallichii √
Ocimum americanum √
Ceiba Sp1 √
Ceiba Sp2 √
Dendrocalamus sp. √
Unknown1
Cocos nucifera √
Artocarpus sp. √
Cyathocline purpurea √
Unknown2
Unknown3
Ficus nervosa √
Unknown4
Stemmadenis sp1. √
Psidium sp2 √
Cocos sp2 √
Tinospora cordifolia √
Syzygium sp1 √ √
Dacrydium bidwilii √
Syzygium thwaitesii √
Leea asiatica L. √
Stemmadenis sp2.
Syzygium sp2 √
Trichomones pinnatum √
Gomphandra coriacea √
Ficus sp. √
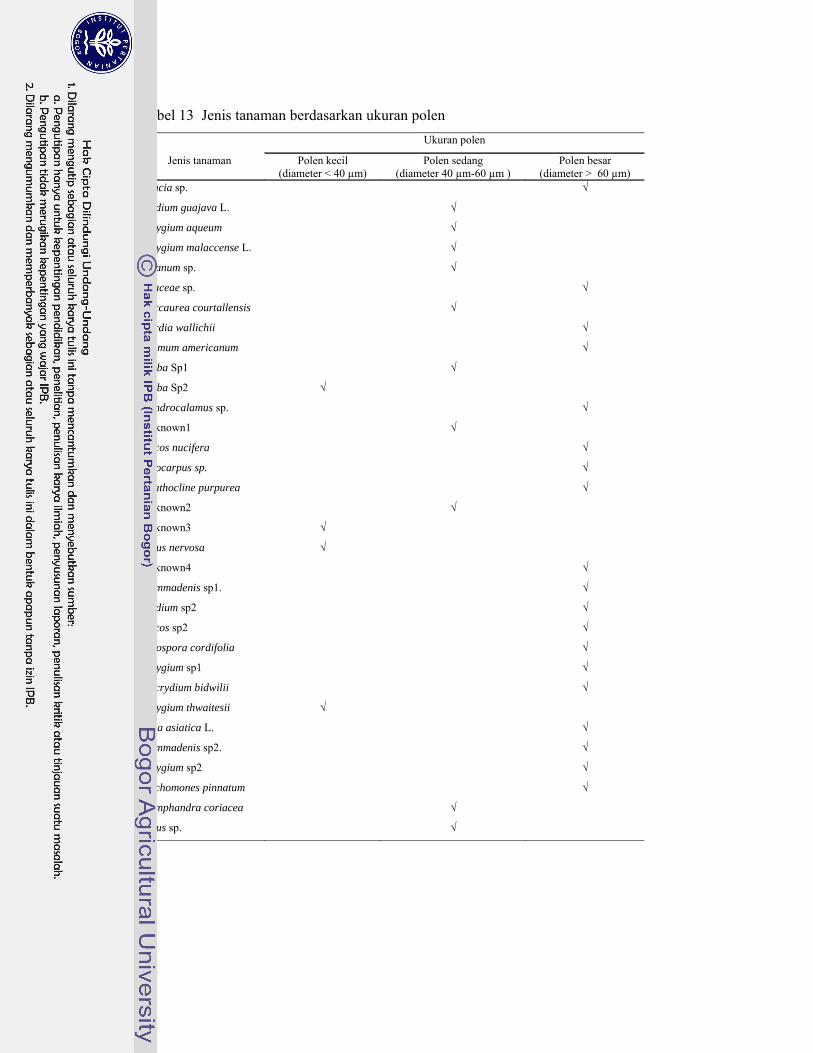
74
Tabel 13 Jenis tanaman berdasarkan ukuran polen Ukuran polen
Jenis tanaman Polen kecil (diameter < 40 µm)
Polen sedang (diameter 40 µm-60 µm )
Polen besar (diameter > 60 µm)
Acacia sp. √
Psidium guajava L. √
Syzygium aqueum √
Syzygium malaccense L. √
Solanum sp. √
Poaceae sp. √
Baccaurea courtallensis √
Cordia wallichii √
Ocimum americanum √
Ceiba Sp1 √
Ceiba Sp2 √
Dendrocalamus sp. √
Unknown1 √
Cocos nucifera √
Artocarpus sp. √
Cyathocline purpurea √
Unknown2 √
Unknown3 √
Ficus nervosa √
Unknown4 √
Stemmadenis sp1. √
Psidium sp2 √
Cocos sp2 √
Tinospora cordifolia √
Syzygium sp1 √
Dacrydium bidwilii √
Syzygium thwaitesii √
Leea asiatica L. √
Stemmadenis sp2. √
Syzygium sp2 √
Trichomones pinnatum √
Gomphandra coriacea √
Ficus sp. √
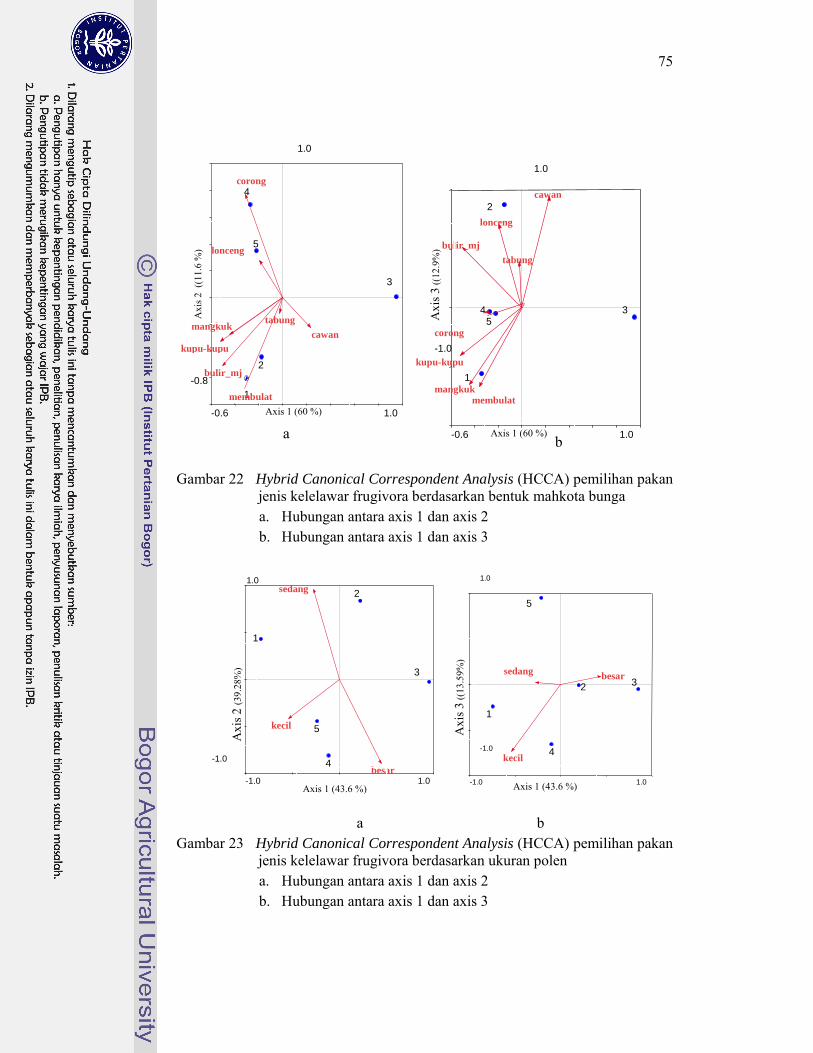
75
Gambar 22 Hybrid Canonical Correspondent Analysis (HCCA) pemilihan pakan jenis kelelawar frugivora berdasarkan bentuk mahkota bunga a. Hubungan antara axis 1 dan axis 2 b. Hubungan antara axis 1 dan axis 3
a b Gambar 23 Hybrid Canonical Correspondent Analysis (HCCA) pemilihan pakan
jenis kelelawar frugivora berdasarkan ukuran polen a. Hubungan antara axis 1 dan axis 2 b. Hubungan antara axis 1 dan axis 3
-0.6 1.0
-0.81
2
3
4
5
tabung
bulir_mj
kupu-kupu
corong
mangkuk
membulat
lonceng
cawan
Axis 1 (60 %)
Axi
s 2 (
(11.
6 %
)
a -0.6 1.0
-1.0
1.0
1
2
3 45
tabung bulir_mj
kupu-kupu
corong
mangkuk membulat
lonceng
cawan
Axis 1 (60 %) A
xis 3
((12
.9%
) b
-1.0 1.0
-1.0
1.0sedang
besar
kecil
1
2
3
4
5
Axis 1 (43.6 %)
Axi
s 2 (3
9.28
%)
-1.0 1.0
-1.0
1.0
sedang besar
kecil
1
2 3
4
5
Axi
s 3 ((
13.5
9%)
1.0
Axis 1 (43.6 %)

76
Gambar 22a, 22b, 23a dan 23b menunjukkan adanya kecenderungan
memilih bentuk mahkota bunga dan ukuran polen, menjadi 3 kelompok kelelawar
yaitu :
1. Kelompok 1, R. amplexicaudatus dan E. spelaea cenderung memilih
polen berukuran sedang (diameter 40 µm - 60 µm ) dari bunga
berbentuk mangkuk, bulir majemuk, dan membulat.
2. Kelompok 2, C. brachyotis cenderung memilih polen berukuran besar
(diameter > 60 µm ) dari bunga berbentuk cawan.
3. Kelompok 3, C. horsfieldii dan M. sobrinus cenderung memilih
polen berukuran kecil (diameter < 40 µm) dari bunga bentuk lonceng
dan corong.
Hasil penghitungan nilai kesamaan relung pakan menunjukkan nilai
kesamaan relung pakan (niche overlap) antara R. amplexicaudatus dan E. spelaea
adalah 0.48. Hal ini berarti bahwa 48% pakan R. amplexicaudatus sama dengan
pakan E. spealea. Hasil uji chi square (X2) antarjenis Megachiroptera berdasarkan
jenis polen yang dimakannya menunjukkan bahwa pakan kelelawar R.
amplexicaudatus tidak berbeda nyata (xhitung > x tabel, df=1) dari pakan kelelawar E.
spelaea, pakan kelelawar C. brachyotis, pakan C. horsfieldii, dan pakan kelelawar
M. sobrinus. Pakan antarjenis kelelawar lainnya juga menunjukkan tidak adanya
perbedaan yang nyata, kecuali antara kelelawar C.horsfieldii dan M. sobrinus.
Nilai kesamaan relung pakan C. horsfieldii dan M. sobrinus 0.397 dan hasil uji
chi square menunjukkan adanya perbedaan yang nyata (xhitung > x tabel, df=1). Nilai
kesamaan relung pakan dan hasil uji chi square antarjenis kelelawar
Megachiroptera dapat dilihat pada Tabel 14.
Berdasarkan Tabel 14 dapat dijelaskan bahwa nilai kesamaan relung pakan
tertinggi adalah antara R. amplexicaudatus dengan E. spelaea yaitu 0,837. Hal ini
berarti 83,7% polen yang dimakan kedua jenis kelelawar tersebut sama. Nilai
kesamaan relung pakan berikutnya yang tinggi adalah antara R. amplexicaudatus
dengan C. horsfieldii (0.786) dan antara C. brachyotis dangan C. horsfieldii
(0.628). Sementara nilai kesamaan relung pakan antara M. sobrinus dengan semua
jenis kelelawar lainnya kurang dari 50%, bahkan kesamaan relung pakan M.
sobrinus dengan C. horsfieldii hanya 0.397. Hasil uji chi square menunjukkan

77
bahwa berdasarkan polen yang dimakan, antarjenis kelelawar Megachiroptera
tidak berbeda nyata (x hitung > x tabel, df=1), kecuali antara M. sobrinus dengan C.
horsfieldii polen yang dimakan berbeda nyata.
Tabel 14 Nilai kesamaan relung pakan dan uji chi antarjenis kelelawar
Megachiroptera
E_s C_b C_h M_s
N_o X2 N_o X2 N_o X2 N_o X2
R._a 0.837 4.037 ns 0.624 0.003ns 0.786 0,063ns 0,499 0.310ns
E_s - - 0.435 1.700ns 0.534 0.224ns 0.452 0.056ns
C_b - - 0.628 0.76 ns 0.458 1.360 ns
C_h - - 0.397 4.9*
M_s - -
Keterangan: R_a= Rousettus amplexicaudatus, E_s= Eonycteris spelaea, C_b=
Cynopterus brachyotis, C_h=Cynopterus horsfieldi, M_s= Macroglosuss sobrinus N_o = indeks niche overlap X2 = nilai chi square * = berbeda nyata (x hitung > x tabel, df=1) ns = tidak berbeda nyata (x hitung > x tabel, df=1)
Pembahasan
Serangga pakan kelelawar insektivora yang ditemukan pada penelitian ini
terdiri atas 29 famili yang termasuk dalam delapan ordo. Dari delapan ordo
serangga pakan tersebut keseluruhannya pernah ditemukan dalam isi perut
kelelawar Microchiroptera pada penelitian-penelitian sebelumnya. Razakarivony
et al. (2005) mendapatkan ordo Orthoptera, Hemiptera, Araneae dan Homoptera
pada pencernaan kelelawar Emballonuridae sp, Hipposideros commersoni,
Triaenops rufus, Miniopterus manavi dan Myotis goudoti di Madagascar. Agosta
(2002) mendapatkan Coleoptera, Lepidoptera, Hymenoptera, Hemiptera, Diptera,
dan Lepidoptera pada pencernaan kelelawar Eptesicus fuscus di Amerika Utara.
Aguirre et al.(2003) mendapatkan Coleoptera, Odonata,Orthoptera, Lepidoptera,
Homoptera, dan Arachnida pada penceraan sepuluh jenis Microchiroptera di
Taman Nasional Espiritu Bolivia. Hal ini menunjukkan serangga yang ditemukan
dalam penelitian ini merupakan serangga yang biasa dimakan kelelawar
Microchiroptera/insektivora.

78
Hasil penelitian ini menunjukkan terdapat kecenderungan kelelawar
Microchiroptera dalam memilih pakannya. Menurut Razakarivony et al. (2005),
kecenderungan (kesukaan) dalam pemilihan pakan ini ditentukan oleh strategi
pencarian makan (foraging strategy), fisiologi dan anatomi tubuh, serta
kelimpahan makanan. Menurut Altringham (1996), dalam strategi pencarian
makan, kelelawar dikelompokkan menjadi tipe spesialis (selektif) dan
opportunis. Kelelawar tipe spesialis hanya memangsa jenis tertentu dengan profit
tinggi. Tipe ini bisa menghabiskan banyak waktu dan energi dalam mencari
makan, dan hanya memilih makanan dengan kriteria tertentu saja. Kelelawar tipe
opportunis menghabiskan lebih sedikit waktu dan energi dalam pencarian
makannya, karena memakan banyak jenis makanan, tetapi keuntungan (profit)
yang didapatkan mungkin lebih sedikit dibandingkan kelelawar tipe spesialis.
Dalam penelitian ini, tampaknya kelelawar H. sorenseni dan Hipposideros sp
merupakan tipe oportunis, karena memakan banyak famili serangga. Sementara R.
affinis, H. diadema, H. ater, H.cf. ater, dan R. borneensis adalah tipe selektif,
karena hanya memilih satu sampai lima famili serangga sebagai makanannya.
Hasil analisis PCA menunjukkan lima kelompok kelelawar
Microchiroptera berdasarkan makanan kesukaannya. Adanya
kecenderungan/kesukaan pada serangga pakan dengan karakteristik khas ini
sejalan dengan penelitian Aguirre et al.(2003) dan Zhang et al. (2005). Menurut
Aguirre (2003), kekerasan serangga dan ukuran serangga yang dipilih ditentukan
oleh kekuatan rahang dan morfologi gigi kelelawar. Menurut Zhang et al. (2005)
terdapat korelasi antara besar tubuh kelelawar dengan besar mangsa, dimana
kelelawar T. pachypus yang bertubuh lebih besar dari pada kelelawar T. robustula
memakan serangga berukuran lebih besar dibandingkan serangga yang dimakan T.
robustula.
Selain tingkat kekerasan dan besar tubuh, diduga ketinggian terbang
serangga juga berpengaruh pada pemilihan pakan kelelawar. Menurut
Altringham (1996), terdapat dua kategori cara pencarian makan kelelawar, yaitu
aerial hawking dan flycatching. Aerial hawking dilakukan dengan cara
mendeteksi keberadaan mangsa, mengejar dan memakannya sambil terbang.
Flycatching dilakukan dengan cara mendeteksi keberadaan mangsa dari

79
ketinggian, mengejar, menangkap, membawanya kembali ke tempat yang tinggi
dan kemudian memakannya. Kelelawar yang berperilaku makan aerial hawking
cenderung memangsa serangga yang terbang tinggi, sebaliknya flycatching
dilakukan oleh kelelawar yang memangsa serangga tanah atau yang terbang
rendah (Altringham 1996). Dalam penelitian ini, ketinggian terbang serangga
pakan tidak teramati. Namun, berdasarkan komunikasi pribadi dengan ahli
serangga di Museum Zoologi LIPI (Sutrisno H. 12 November 2010, komunikasi
pribadi) diketahui bahwa kebanyakan anggota ordo Lepidoptera, Orthoptera,
Coleoptera, dan Diptera cenderung berada di dekat permukaan tanah, sedangkan
anggota ordo Hemiptera, Homoptera, Trichoptera, dan Hymenoptera dapat
terbang tinggi dari permukaan tanah. Dengan demikian dapat diduga kelelawar
C.plicata, H.sorenseni, Hipposideros sp dan H.diadema, termasuk jenis yang
berperilaku flycatching, sementara R. borneensis, H. bicolor, H. ater, H.cf. ater
dan M. schreibersii termasuk jenis yang berperilaku aerial hawking. Jenis
lainnya, yaitu R. affinis dan M. australis termasuk jenis yang berperilaku
kombinasi keduanya flycatching, dan aerial hawking.
Menurut Ludwig & Reynolds (1988) nilai kesamaan relung pakan berkisar
antara nol sampai satu. Semakin mendekati nilai satu, berarti semakin tinggi
kesamaan pakan kedua jenis hewan yang dibandingkan, atau kedua jenis hewan
tersebut memakan makanan yang sama. Sebaliknya, nilai kesamaan relung pakan
mendekati nol berarti kedua jenis hewan memakan makanan yang berbeda. Nilai
kesamaan relung pakan kelelawar Microchiroptera yang ditemukan tergolong
kecil. Hasil penelitian ini menunjukkan tidak terjadi overlap pakan pada
kelelawar Microchiroptera yang bersarang di gua-gua Karst Gombong. Hal ini
karena masing-masing jenis menyukai makanan yang berbeda.
Hasil penelitian ini menunjukkan bahwa asosiasi bersarang dalam satu gua
terjadi antara jenis-jenis kelelawar yang memiliki nilai kesamaan relung pakan
(nilai niche overlap) kecil. Nilai kesamaan relung pakan tersebut tidak melebihi
30 %, kecuali di Gua Kemit 47.1%. Menurut Cox (2002) penggunaan relung
yang sama (niche overlap) menyebabkan interaksi kompetitif, dimana tiap
populasi yang berkompetisi memberikan pengaruh yang merugikan bagi
pesaingnya (competitor). Hasil penelitian ini membuktikan bahwa kompetisi

80
makanan tidak terjadi antara jenis-jenis Microchiroptera yang bersarang dalam
satu gua yang sama. Hasil penelitian ini dapat menjawab pertanyaan mengapa
kelelawar insektivora (Microchiroptera) yang bersarang dalam satu gua dapat
bertahan dari generasi ke generasi dan mencapai jumlah populasi yang tinggi
tanpa berkompetisi memperebutkan makanan. Meskipun mencari makanan di
lokasi yang sama, kelelawar tersebut membagi relung pakannya dan memilih
makanan dengan karakteristik berbeda. Hal ini diperkuat dengan hasil temuan
bahwa nilai niche overlap pakan antarjenis kelelawar insektivora yang bersarang
dalam satu gua, lebih kecil bila dibandingkan dengan nilai kesamaan relung
pakan antarjenis kelelawar yang bersarang pada gua berbeda. Sebagai contoh
adalah antara H. sorenseni dengan Hipposideros sp. Nilai niche overlap antara
H. sorenseni dengan Hipposideros sp. yang bersarang pada gua berbeda adalah
0.528. Sementara nilai niche overlap pakan H. sorenseni dengan Hipposideros sp
yang bersarang bersama di Gua Jatijajar adalah 0.169 . Demikian pula halnya
dengan C. plicata dengan H. sorenseni. Nilai niche overlap antara C. plicata
dengan H. sorenseni yang bersarang pada gua berbeda adalah 0.683. Namun
nilai niche overlap antara C. plicata dengan H. sorenseni yang bersarang bersama
di Gua Celeng adalah 0.054. Hal ini mengindikasikan, dua jenis kelelawar
insektivora yang bersarang dalam satu gua, membagi relung pakannya untuk
menghindari kompetisi.
Kesamaan jenis pakan lebih dari 30% (yaitu: 47.9%) ditemukan antara
H.sorenseni dengan Hipposideros sp yang bersarang bersama di Gua Kemit.
Sebagaimana telah dijelaskan sebelumnya, berdasarkan jenis-jenis serangga yang
dimakan, kelelawar H. sorenseni dengan Hipposideros sp dapat digolongkan
sebagai jenis kelelawar oportunis karena memakan banyak jenis serangga.
Berdasarkan hasil pengamatan, Gua Kemit berada di Hutan Lindung Kawasan
Karst Gombong Selatan sebelah Timur. Vegetasi di wilayah tersebut lebih
didominasi oleh semak belukar daripada pepohonan. Berbagai jenis serangga
terutama ordo Coleoptera dan Orthoptera terlihat melimpah di semak belukar
tersebut pada saat penelitian ini dilakukan. Karena pakan tersedia dalam jumlah
banyak, kesamaan relung pakan yang tinggi antara kedua jenis kelelawar tersebut
tidak menyebabkan kompetisi negatif.

81
Menurut Altringham (1996) kelelawar subordo Megachiroptera tidak
mempunyai hubungan kekerabatan dengan kelelawar subordo Microchiroptera.
Salah satu alasan yang mendukung adalah : Saraf superior colliculus (s.c) kanan
pada otak tengah Microchiroptera mengatur retina mata kiri dan sebaliknya s.c
kiri mengatur retina mata kanan. Hal ini ditemukan pada semua Mamalia kecuali
Primata. Pada Megachiroptera, saraf superior colliculus kanan otak tengah
mengatur retina mata kiri dan mata kanan sekaligus. Keadaan ini hanya
ditemukan pada Primata, Dermoptera dan Megachiroptera (Altringham 1996).
Terpisahnya kekerabatan Megachiroptera dengan Microchiroptera ini dipertegas
oleh Han Guan et al. (2006) yang meneliti filogenetik kelelawar, bahwa
Megachiroptera memiliki kekerabatan lebih dekat dengan primata daripada
dengan Microchiroptera. Oleh karena berbeda kekerabatan, perilaku makan
kelelawar Microchiroptera jauh berbeda dengan kelelawar Megachiroptera.
Menurut Chairunnisa (1997) diduga Megachiroptera berasal dari nenek moyang
bangsa Primata yang cenderung frugivora, sedangkan Microchiroptera berasal dari
nenek moyang Rodensia yang cenderung insektivora.
Famili tumbuhan yang ditemukan dalam pencernaan Megachiroptera pada
penelitian ini, beberapa di antaranya pernah ditemukan di saluran pencernaan
Megachiroptera pada penelitian-penelitian sebelumnya. Di antaranya adalah
penelitian Tan et al. (1998) yang mendapatkan famili Myrtaceae dan
Euphorbiaceae pada kelelawar C. Brachyotis dan penelitian Soegiharto et al.
(2010) yang mendapatkan polen tumbuhan Bombacaceae, Fabaceae, Myrtaceae
dan Euphorbiaceae pada pencernaan kelelawar Cynopterus minutus, C.
brachyotis, C.titthaheilus, M. sobrinus dan R. amplexicaudatus.
Polen tumbuhan yang paling banyak dijumpai dalam saluran pencernaan
kelelawar Megachiroptera adalah dari famili Myrtaceae (83,3%), Bombacaceae
(83,3%) dan Fabaceae (83,3%). Myrtaceae (jambu-jambuan), Bombacaceae
(kapuk), dan Fabaceae (akasia) merupakan tumbuhan yang biasa ditemukan pada
daerah-daerah hutan karst. Hasil foto satelit citra LANDSAT (Lampiran 2),
terlihat dua per tiga dari Karst Gombong adalah hutan karst dan kebun campur.
Di antara hutan tersebut terdapat persawahan dan permukiman penduduk.
Berdasarkan hasil penelitian Riswan et al. (2006) jenis tumbuhan yang dijumpai

82
di hutan kasrt Gombong terdiri atas 187 jenis tumbuhan yang termasuk dalam 125
famili dan 60 ordo. Jenis-jenis tersebut merupakan jenis asli hutan karst, seperti
Alstonia scholaris (pulai), Piper aduncum (sirih-sirihan), dan Saccharum
spontaneum (glagah) serta tumbuh-tumbuhan yang ditanam untuk maksud
reboisasi yang disarankan oleh PERHUTANI yaitu pohon serba guna yang
bernilai ekonomi. Famili tumbuhan yang ditemukan di saluran pencernaan
kelelawar Megachiroptera yang ditemukan pada penelitian ini, sebagain besar
termasuk dalam daftar famili tumbuhan hutan Karst Gombong yang telah
diidentifikasi oleh Riswan et al. (2006) tersebut. Di antaranya adalah Fabaceae,
Myrtaceae, Euphorbiacea, Aracaceae, Moraceae, Asteraceae, Bombacaceae, dan
Apocynaceae. Dengan demikian, dapat dikatakan bahwa kelelawar
Megachiroptera berperan penting dalam kelangsungan hidup tanaman hutan Karst
Gombong karena membantu penyerbukannya.
Berdasarkan polen tumbuhan yang dimakan, hasil analisis PCA
menunjukkan R. amplexicaudatus, E. spelaea, C. brachyotis, C. horsfieldii dan M.
sobrinus memiliki kecenderungan memilih polen dari tumbuhan yang berbeda.
Namun demikian, hasil perhitungan nilai kesamaan relung pakan dan uji chi
square menunjukkan bahwa perbedaan tersebut tidak nyata, kecuali antara
C.horsfieldii dan M.sobrinus.
Berdasarkan karakteristik bunga dan ukuran polen, hasil HCCA
menunjukkan adanya pengelompokan jenis kelelawar menjadi 3 kelompok.
Menurut Barth (1991) secara alami koevousi bunga Angiosfermae beradaptasi
pada hewan penyerbuk, begitu pula sebaliknya. Oleh karena itu, hasil penelitian
ini menunjukkan adanya kesesuaian morfologi moncong kelelawar dengan
morfologi bunga. Bila dilihat morfologi moncongnya, kelompok 1, yaitu R.
amplexicaudatus dan E. spelaea bermoncong pendek, sehingga cenderung
memilih polen berukuran sedang (diameter 40 µm - 60 µm ) dari bunga
berbentuk mangkuk, bulir majemuk, dan membulat. Kelompok 2 C. brachyotis
bermoncong besar dan lebar, sehingga cenderung memilih polen berukuran besar
dari bunga berbentuk cawan. Kelompok 3 yaitu C. horsfieldii, dan M. sobrinus
bermoncong ramping dan panjang, cenderung memilih polen berukuran kecil dari
bunga berbentuk tabung dan corong. Namun, hasil penelitian ini sedikit berbeda

83
dari hasil penelitian Soegiharto et al. (2010) yang membuktikan bahwa
karakteristik tumbuhan yang diserbuki kelelawar memiliki bunga berwarna putih,
ukuran polen besar, dan banyak, bentuk mahkota bunga mangkuk, menghasilkan
nektar banyak, dan mengeluarkan bau menyengat.
Nilai kesamaan relung pakan antarjenis kelelawar Megachiroptera yang
ditemukan tinggi ( > 43%), kecuali antara C.horsfieldii dengan M. sobrinus ( < 43
%). Demikian pula halnya dengan hasil uji chi kuadrat, menunjukkan bahwa
berdasarkan polen yang dimakannya, antara jenis-jenis kelelawar Megachiroptera
tidak berbeda nyata, kecuali antara C.horsfieldii dengan M. sobrinus. Hal ini
menunjukkan adanya kesamaan pakan antara jenis-jenis tersebut. Meskipun
memiliki kesamaan relung pakan yang tinggi, diduga kompetisi negatif antarjenis-
jenis Megachiroptera di Karst Gombong tidak terjadi. Hal ini karena kelelawar
Megachiroptera memiliki wilayah jelajah yang luas dalam pencarian pakannya.
Sebagaimana dinyatakan oleh Kunz & Pierson (1994) bahwa kelelawar
Megachiroptera mampu menjelajah dengan jarak 28 km dalam satu kali
pencarian makannya. Selain itu kawasan Karst Gombong seluruhnya berada di
areal hutan lindung yang menyediakan pakan cukup banyak bagi kelelawar
Megachiroptera.
Kesimpulan
1. Berdasarkan kesukaannya pada serangga pakan, kelelawar insektivora di
Karst Gombong dibagi menjadi lima kelompok yaitu: kelompok 1 adalah
C. plicata, Hipposideros sp, dan H. sorenseni cenderung memilih serangga
berukuran besar dan keras; kelompok 2 adalah H.diadema yang juga
cenderung memilih serangga berukuran besar dan keras tetapi dari jenis
yang berbeda dengan kelompok 1; kelompok 3 adalah M. Schreibersii, M.
australis, H.cf. ater, R.borneensis, cenderung memilih serangga berukuran
sedang dan lunak; kelompok 4 adalah R. affinis , cenderung memilih
serangga berukuran kecil dan keras; kelompok 5 adalah H. bicolor dan H.
ater cenderung memilih serangga berukuran kecil dan lunak. Berdasarkan
kesukaan pada polen tumbuhan, kelelawar frugivora di Karst Gombong
dibagi menjadi tiga kelompok: kelompok 1 adalah R. amplexicaudatus

84
dan E. spelaea yang cenderung memilih polen berukuran sedang
(diameter 40 µm - 60 µm ) dari bunga berbentuk mangkuk, bulir majemuk
dan membulat; kelompok 2 adalah C. brachyotis yang cenderung memilih
polen berukuran besar (diameter > 60 µm ) dari bunga berbentuk cawan,
dan kelompok 3 adalah C. horsfieldii dan M. sobrinus yang cenderung
memilih polen berukuran kecil (diameter < 40 µm) dari bunga bentuk
lonceng dan corong.
2. Nilai kesamaan relung pakan antarjenis kelelawar insektivora yang
bersarang dalam satu gua kurang dari 30%. Berarti kompetisi pakan
antarjenis kelelawar yang bersarang dalam satu gua relatif kecil.
Saran
1. Diperlukan penelitian lebih lanjut untuk mengetahui kelimpahan serangga
pakan dan waktu kemunculan serangga tersebut, sehingga dapat diketahui
pengaruh kelimpahan serangga pada pemilihan relung pakan dan pembagian
relung pakan berdasarkan waktu.
2. Keanekaragaman jenis tumbuhan di hutan lindung dan kebun campur yang
berada di sekitar gua, perlu dijaga kelestariannya. Mengingat hutan tersebut
mnyediakan pakan bagi kelelawar yang bersarang di gua-gua karst.

85
ADAPTASI STRUKTURAL DAN FISIOLOGI PERNAPASAN
KELELAWAR PENGHUNI GUA:
STUDI KASUS DI KAWASAN KARST GOMBONG
KABUPATEN KEBUMEN JAWA TENGAH
ABSTRACT
The purpose of this study was to observe the structural and physiological adaptation of the cave bats conducted from September 2009 to March 2010 in Karst Gombong, Kebumen Regency, Central Java. In each cave where the bats roosting, the temperature, also the humidity, oxygen level, and amonia level were measured. Three individual samples were caught from every bat roost during the day when the bats were staying in their roosts. Blood was taken from the vein interfemoral of bats. Then the erythrocyte count was measured by using hemocytometer and hemoglobin content was measured using Sahli’s method. Lungs removed from body and were made preparations for histology. The diameter of alveoli was observed under the microscope. The data were analyzed using ANOVA and multiple regessions. It was concluded from this study that humidity, temperature, and amonia correlated significantly with erythrocyte and hemoglobin content. The diameter of alveoli significantly correlated with humidity, temperature, and amonia levels. The amount of erythrocyte increased by the increase of humidity, decrease in the temperature, and the increase of amonia level. This tendency also revealed in hemoglobin change. Key words : cave bat, adaptation, alveolus, erythrocyte, hemoglobin. Pendahuluan
Hasil penelitian membuktikan bahwa mikroklimat lingkungan di sekitar
sarang berpengaruh pada pemilihan sarang oleh kelelawar. Hal ini karena setiap
jenis kelelawar memilih sarang sesuai dengan kebutuhan fisiologis tubuhnya.
Berkaitan dengan persyaratan mikroklimat tersebut, beberapa jenis kelelawar
terbukti dapat bersarang di dalam gua dengan kondisi rendah oksigen, lembap,
tinggi karbon dioksida, dan tinggi kadar amonia. Keadaan ini sebenarnya kurang
menguntungkan bagi kelelawar, karena beberapa hasil penelitian pada hewan
Mamalia membuktikan bahwa: 1) Kekurangan oksigen dapat menyebabkan
kematian sel dan gangguan metabolisme (Kunz & Nagy 1988); 2) Paparan gas
amonia (NH3) dapat menyebabkan iritasi epitel organ pernapasan serta gangguan
fisiologi saraf (Hutabarat, 2002); dan 3) Berada pada lingkungan bersuhu
rendah dan kelembapan udara tinggi dapat menyebabkan gangguan psikomotorik

86
dan terhambatnya aktivitas kerja enzim (Baudinette et al. 2000). Namun, karena
mampu beradaptasi, kelelawar dapat bertahan hidup dan berkembang dalam
sarang dengan kondisi tidak menguntungkan. Bahkan menurut Furman & Ozgul
(2002), sarang kelelawar tersebut dipertahankan dari generasi ke generasi.
Kelelawar merupakan hewan troglozene (bersarang di dalam gua, mencari
makan di luar gua), sehingga tidak semua aktivitas harian kelelawar dilakukan di
dalam gua. Namun, menurut Zahn & Hager (2005) kelelawar berada di sarangnya
minimal delapan jam dalam satu hari. Beberapa hasil penelitian pada Mamalia
lain, mengindikasikan paparan gas racun lebih dari dua jam dapat menyebabkan
gangguan pernapasan. Heinrich et al. (1986) meneliti pengaruh gas emisi
pembakaran bensin pada sistem pernapasan hamster (Cricetulus barabensis) dan
tikus (Mus musculus). Hasil penelitian menunjukkan paparan gas emisi selama
120 menit menyebabkan edema paru, yaitu penimbunan cairan serosa atau
serosanguinosa secara berlebihan di dalam ruang interstisial dan alveolus
(Heinrich et al. 1986). Hutabarat (2002) melakukan penelitian pada karyawan
pabrik latexs yang terkena paparan amonia sebesar 500 ppm sampai 600 ppm
selama empat jam per hari dalam 60 hari. Hasil penelitian menunjukkan,
karyawan tersebut mengalami gejala tenggorokan kering (80%), mata perih
(66.67%), batuk (53.3%), dan pingsan (6.67%). Oleh karena itu, apabila tidak
beradaptasi, kelelawar yang berada dalam gua delapan jam per hari selama
hidupnya, sangat beresiko mengalami gangguan pernapasan.
Strategi adaptasi kelelawar yang bersarang di gua dengan suhu udara dingin
dan lembap pernah diteliti oleh Baudinette et al.(2000) di Australia. Hasil
penelitian menunjukkan laju respirasi kelelawar Macroderma gigas
(Microchiroptera) dan Rhinonycteris aurantias (Microchiroptera) menyesuaikan
suhu dan kelembapan udara dalam gua. Pada saat kondisi udara kering dan
dingin (kelembapan <60% dan suhu < 5.6oC) laju respirasi sama dengan pada
saat kondisi udara lembap dan hangat (kelembapan > 80% ;suhu > 9.8oC) yaitu
1.96 ± 0.08 BMR (1 BMR/ Basal Metabolic Rate = 1 ml O2 g-1 h-1). Tetapi bila
kondisi udara lembap dan dingin (kelembapan <60% dan suhu < 9o) laju
respirasinya meningkat tajam, yaitu 13.47± 0.44 BMR. Meningkatnya laju
respirasi tersebut merupakan strategi agar tubuh tetap hangat. Namun, sejauh ini

87
belum ada penelitian yang menjelaskan adaptasi struktural dan fisiologi
pernapasan kelelawar sebagai strategi untuk bertahan hidup di gua dengan
kondisi udara dingin, lembap, kadar amonia tinggi, dan kadar oksigen rendah.
Informasi ilmiah ini sangat diperlukan terutama untuk merancang strategi
pelestarian kelelawar yang tepat, sesuai dengan kebutuhan fisio-ekologinya.
Perbedaan diameter alveolus, jumlah eritosit dan kadar hemoglobin
antarkoloni kelelawar yang bersarang di lingkungan dengan kondisi udara
berbeda, dapat dijadikan indikator dalam memahami strategi adaptasi struktural
dan fisiologi pernapasan kelelawar. Hal ini didukung oleh beberapa penelitian
terdahulu. Menurut Ganong (2001), semakin banyak dan kecil diameter alveolus,
semakin luas permukaan membran alveolus dan semakin banyak oksigen dan
karbon dioksida yang dapat dipertukarkan. Hasil penelitian Setiadi (2000) pada
kelelawar Scotophilus kuhli (Vespertilionidae: Mikrochiroptera) membuktikan
alveolus Scotophilus kuhli lebih kecil dan rapat dibandingkan alveolus tikus (Mus
musculus). Menurut Setiadi (2000), sebagai Mamalia terbang, kebutuhan oksigen
Scotophilus kuhli lebih banyak dibandingkan kebutuhan oksigen Mus musculus,
akibatnya pertukaran gas di alveous Scotophilus kuhli berlangsung lebih efektif
daripada pertukaran gas di alveolus Mus musculus. Menurut Ganong (2001)
penurunan kadar oksigen (hipoksia) akan merangsang ginjal untuk melepaskan
enzim erythrogenin yang memacu pembentukan sel darah merah. Binatang yang
terpapar oksigen rendah selama bertahun-tahun menunjukkan peningkatan
jaringan hipoksik dan peningkatan jumlah sel darah merah sampai 20% (Ganong
1995). Karena alasan tersebut di atas, diduga kelelawar yang bersarang di gua
yang dingin, lembap, kadar oksigen rendah dan kadar amonia tinggi beradaptasi
dengan diameter alveolus kecil, jumlah eritrosit tinggi dan kadar hemoglobin
tinggi.
Tujuan penelitian ini adalah : mengkaji adaptasi struktural dan fisiologi
pernapasan kelelawar yang bersarang di gua-gua Karst Gombong. Hipotesis
penelitian ini adalah: adaptasi kelelawar terhadap kondisi gua yang dingin,
lembap, rendah kadar oksigen, dan tinggi kadar amonia menentukan struktur dan
fisiologi organ pernapasan, yaitu diameter alveolus sempit (< 100 µm) rasio

88
jumlah eritrosit/bobot badan tinggi (> 0.3 juta ml/g bobot badan) dan rasio kadar
hemoglobin/bobot badan tinggi (> 0,9 g/ml/g bobot badan).
Bahan dan Metode
Penelitian dilakukan pada September 2009 s/d Maret 2010. Lokasi
penelitian di tujuh goa yang terdapat di kawasan Karst Gombong Kabupaten
Kebumen Jawa Tengah yaitu Gua Petruk, Gua Celeng, Gua Kemit, Gua Jatijajar,
Gua Inten, Gua Liyah dan Gua Dempo. Pengamatan laboratorium dilakukan di
laboratorium hewan PPSHB PAU IPB, dan Laboratorium Fisiologi dan Anatomi
FKH IPB.
Bahan yang digunakan adalah chloroform, larutan fisiologis NaCl 0.9%,
HCl 0.1 N, aquadest, sampel darah, antikoagulan Ethylene Diamine Tetraacetic
Acid (EDTA), alkohol 70%, alkohol 80%, alkohol 90%, larutan bouin, parafin,
xylol dan pewarna Hematoksilin Eosin (HE). Alat yang digunakan adalah hand
net, mist net dan harpa net, peralatan bedah; timbangan digital, mikroskop
(Olympus), jangka sorong digital (Krisbow), spuit, kapas, venoject (Therumo,
Japan) berisi EDTA, tabung reaksi, gelas objek, kaca penutup, pipet eritrosit,
aspirator, kamar hitung hemocytometer, tabung Sahli, mikrotom putar, pisau
mikrotom ukuran 5 µm dan blok parafin, dan mikrometer.
Cara kerja
Pengukuran mikroklimat udara dilakukan di sekitar sarang kelelawar pada
jarak terdekat yang mungkin terjangkau. Parameter mikroklimat yang diukur
adalah: suhu, kelembapan udara relatif, kadar oksigen, dan kadar amonia udara.
Sampel kelelawar Microchiroptera ditangkap di sarangnya di dalam gua,
sedangkan sampel Megachiroptera ditangkap di sekitar gua (Megachiroptera tidak
dapat ditangkap di dalam gua karena berjumlah sedikit/ maksimal 6 ekor). Sampel
yang dipilih adalah kelelawar dewasa berjenis kelamin jantan, sebanyak 3 ekor
setiap sarang. Pengukuran mikroklimat sarang dan penangkapan sampel dilakukan
pada siang hari pada saat kelelawar berada di sarangnya.
Penghitungan jumlah eritrosit dan hemoglobin dilakukan dengan cara: darah
diambil dari vena interfemoral sebanyak 1 ml menggunakan spuit ukuran 1 ml

89
(Kunz & Nagy 1988). Darah yang terambil segera dimasukkan ke dalam tabung
venoject yang telah berisi EDTA. Penghitungan jumlah eritrosit dilakukan dengan
cara (Sastradipradja 1989): 1) Sampel darah dihisap menggunakan pipet eritrosit
hingga tanda tera 0.5 pada aspirator; 2) Ujung pipet eritrosit dibersihkan
menggunakan kertas tissu, selanjutnya pengencer hayem dihisap hingga tanda tera
101 pada aspirator; 3) Pipet digerakkan memutar dengan membentuk arah angka
delapan selama tiga menit; 4) Larutan yang tidak tercampur dibuang dengan
menempelkan kertas tissu pada ujung pipet; 5) Satu tetes larutan diteteskan ke
dalam hemositometer; 6) Dilakukan penghitungan jumlah sel eritrosit yang
terdapat pada lima kotak eritrosit hemositometer (satu kotak kanan atas, satu
kotak kiri atas, satu kotak kiri bawah, satu kotak kiri bawah dan satu kotak
tengah) di bawah mikroskop perbesaran 40 X 10. Jumlah eritrosit kelelawar
adalah jumlah total eritrosit dalam lima kotak hemositometer dikali 10 4 /mm3
(Sastradipradja 1989) .
Pengukuran kadar hemoglobin darah dilakukan dengan metode Sahli yaitu
(Sastradipradja 1989) : 1) Larutan HCl 0.1 N diteteskan pada tabung Sahli sampai
tanda tera 1.0; 2) Sampel darah dihisap dengan mengunakan pipet Sahli sampai
tanda tera 2.0; 3) Sampel darah dimasukkan ke dalam tabung Sahli dan
dibiarkan selama 3 menit; 4) Larutan ditambah dengan aquades sedikit demi
sedikit sambil diaduk dengan pengaduk Sahli hingga warna larutan sama dengan
warna standar hemoglobinometer pada kit Sahli; 5) Nilai kadar hemoglobin (g/ml)
adalah angka yang tertera pada tabung hemoglobin.
Kelelawar yang sudah diambil sampel darahnya segera dibius dengan
chloroform dan dilakukan pembedahan. Paru-paru dikeluarkan dari tubuh dan
direndam dalam larutan Bouin selama 24 jam, setelah itu dipindahkan ke dalam
larutan alkohol 70% untuk kemudian dilakukan proses pembuatan preparat
histologi (Gambar 24).
Pembuatan preparat histologi dilakukan dengan cara (Sobota & Hemmersen
1993): 1) Paru-paru dipotong sebesar 0.5 cm2, kemudian didehidrasi untuk
menarik air dari jaringan dengan cara direndam dalam larutan alkohol bertingkat
70% (selama 24 jam); 80% (selama 24 jam) dan 90% (selama 12 jam); 2)
Potongan paru-paru yang telah didehidrasi dijernihkan dengan cara direndam

90
dalam xylol selama 30 menit; 3) Potongan paru-paru yang sudah dijernihkan,
dilakukan infiltrasi dengan parafin cair dalam inkubator parafin; 4) Dilakukan
embedding (penanaman) pada blok parafin; 5) Dilakukan pemotongan dengan
mikrotom putar dengan ketebalan 5 µm; 6) Sayatan diletakkan pada gelas objek
dan dilakukan pewarnaan dengan Hematoxcillin Eosin (HE).
a b c
Gambar 24 Proses pembedahan dan perendaman paru-paru dalam larutan Bouin a. situs viscerum setelah dilakukan pembedahan longitidinal
ventral. b. Paru paru setelah dikeluarkan dari tubuh. c. Perendaman dalam larutan Bouin
Pengukuran diameter alveolus dilakukan dengan cara: 1) Mikrometer
dipasang pada lensa okuler dan dilakukan kalibrasi skala dengan perbesaran 40 x
10; 2) Alveolus dipilih secara acak pada lima lapang pandang, setiap lapang
pandang diukur 10 alveolus.
Analisis data
Transformasi data dilakukan untuk mengubah skala pengukuran data asli
menjadi bentuk lain sehingga data dapat memenuhi asumsi-asumsi yang
mendasari analisis ragam. Pada penelitian ini digunakan transformasi log (n+1),
dikarenakan data tersaji dalam sebaran tidak normal (Gaspersz 1995). Selain itu
jumlah eritrosit dan kadar amonia dianalisis berdasarkan proporsi per gam bobot
badan, hal ini karena sampel kelelawar yang diamati memiliki bobot tubuh yang
sangat beragam.
Perbedaan diameter alveolus, jumlah eritrosit dan kadar hemoglobin
antarkoloni kelelawar yang bersarang pada ruang gua yang berbeda, dianalisis
dengan sidik ragam (ANOVA). Untuk melihat perbedaan antarkoloni kelelawar
dilakukan uji lanjut Tukey pada tingkat selang kepercayaan 95%. Hubungan

91
antara kondisi udara di sekitar sarang (suhu, kelembapan, kadar oksigen dan kadar
amonia) dengan diameter alveolus, jumlah eritrosit, dan kadar hemoglobin
dianalisis dengan uji korelasi Spearman, uji ANOVA, uji Tukey dan uji regesi
linear berganda (multiple regession) pada tingkat selang kepercayaan 95 %.
Pengolahan data dilakukan dengan menggunakan software SPSS versi 17
(Gaspersz 1995).
Hasil
Sebanyak 40 sampel kelelawar dewasa berjenis kelamin jantan berhasil
ditangkap dan dilakukan pengamatan pada diameter alveolus, jumlah eritrosit, dan
kadar hemoglobinnya. Kelelawar tersebut berasal dari 14 sarang (14 koloni) yang
tersebar di tujuh gua. Setelah dilakukan identifikasi, diketahui kelelawar tersebut
terdiri atas tiga jenis Megachiroptera (Cynopterus brachyotis (Muller, 1838);
Rousettus amplexicaudatus (E.Geoffroy, 1810); dan Eonycteris spelaea
(Dobson,1871) dan delapan jenis kelelawar Microchiroptera (Chaerophon plicata
(Buchannan,1800), Hipposideros ater Templeton, 1848; Hipposideros cf. ater;
Hipposideros sp; Hipposideros sorenseni Kitchener & Maryanto, 1993,
Rhinolophus affinis Horsfield, 1823; Miniopterus australis Tomes, 1858; dan
Miniopterus schreibersii (Kuhl, 1819)).
Hasil pengukuran mikroklimat udara di sekitar sarang Chaerophon plicata
yang berada di Gua Petruk menunjukkan kelembapan udara = 70.4± 0.1% , suhu
udara= 29.5±0.1%, kadar oksigen= 22.0± 0.8%, dan kadar amonia= 2810 ppm.
Setelah dilakukan pengamatan pada sampel kelelawar, diketahui bobot tubuh
Chaerophon plicata tersebut adalah 12.8± 0.75 g (n=3), jumlah eritrosit=
4.080.000± 200.000 /ml (n=3), dan kadar hemoglobin =12.4 ± 0.5 g/ml (n=3).
Hasil pengukuran pada sayatan histologi paru-paru, menunjukkan diameter
alveolus kelelawar ini adalah = 114.2 ± 24 µm (n=50). Hasil pengukuran
mikroklimat sarang, jumlah eritrosit, kadar hemoglobin dan diameter alveolus
koloni kelelawar lainnya dapat dilihat pada Tabel 16.
Hasil uji korelasi Spearman menunjukkan: 1) Diameter alveolus berkorelasi
nyata dengan kadar amonia (R=-0.342,P<0.05), tetapi tidak berkorelasi nyata
dengan suhu, kelembapan dan kadar oksigen; 2) Rasio jumlah eritrosit per gram
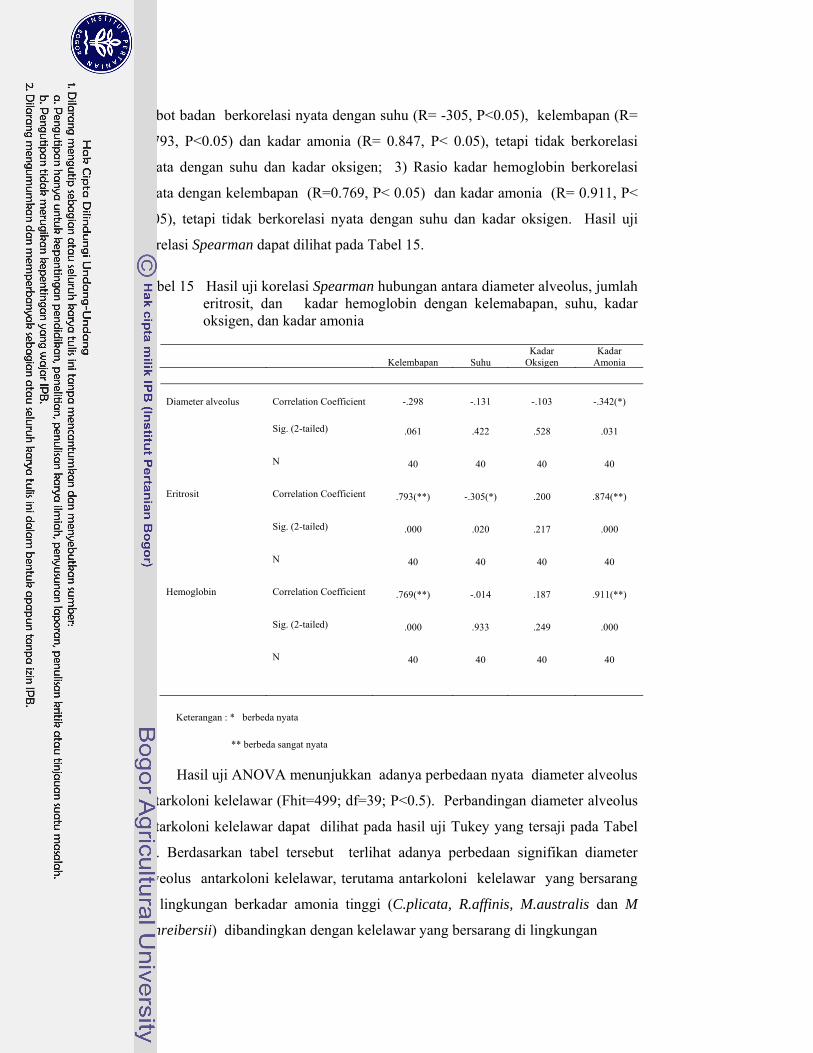
92
bobot badan berkorelasi nyata dengan suhu (R= -305, P<0.05), kelembapan (R=
0.793, P<0.05) dan kadar amonia (R= 0.847, P< 0.05), tetapi tidak berkorelasi
nyata dengan suhu dan kadar oksigen; 3) Rasio kadar hemoglobin berkorelasi
nyata dengan kelembapan (R=0.769, P< 0.05) dan kadar amonia (R= 0.911, P<
0.05), tetapi tidak berkorelasi nyata dengan suhu dan kadar oksigen. Hasil uji
korelasi Spearman dapat dilihat pada Tabel 15.
Tabel 15 Hasil uji korelasi Spearman hubungan antara diameter alveolus, jumlah eritrosit, dan kadar hemoglobin dengan kelemabapan, suhu, kadar oksigen, dan kadar amonia
Kelembapan Suhu Kadar
Oksigen Kadar
Amonia
Diameter alveolus
Correlation Coefficient
-.298
-.131
-.103
-.342(*)
Sig. (2-tailed) .061 .422 .528 .031
N 40 40 40 40
Eritrosit Correlation Coefficient .793(**) -.305(*) .200 .874(**)
Sig. (2-tailed) .000 .020 .217 .000
N 40 40 40 40
Hemoglobin Correlation Coefficient .769(**) -.014 .187 .911(**)
Sig. (2-tailed) .000 .933 .249 .000
N 40 40 40 40
Keterangan : * berbeda nyata
** berbeda sangat nyata
Hasil uji ANOVA menunjukkan adanya perbedaan nyata diameter alveolus
antarkoloni kelelawar (Fhit=499; df=39; P<0.5). Perbandingan diameter alveolus
antarkoloni kelelawar dapat dilihat pada hasil uji Tukey yang tersaji pada Tabel
17. Berdasarkan tabel tersebut terlihat adanya perbedaan signifikan diameter
alveolus antarkoloni kelelawar, terutama antarkoloni kelelawar yang bersarang
di lingkungan berkadar amonia tinggi (C.plicata, R.affinis, M.australis dan M
schreibersii) dibandingkan dengan kelelawar yang bersarang di lingkungan
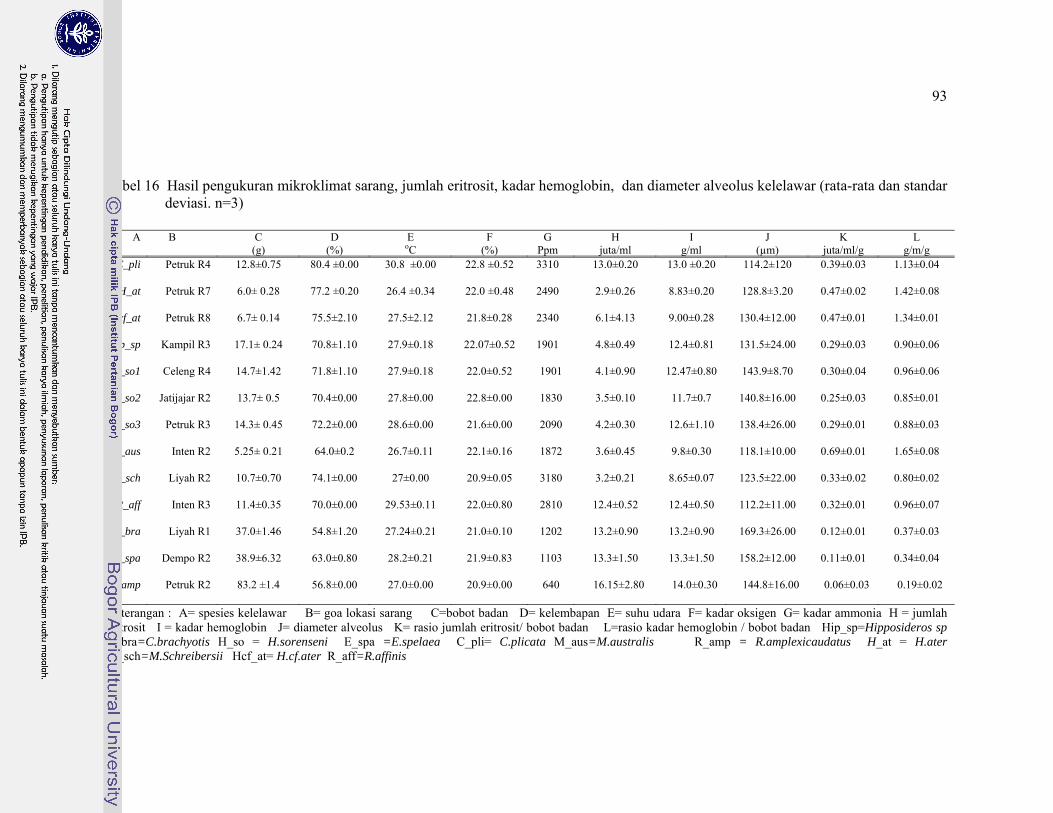
93
Tabel 16 Hasil pengukuran mikroklimat sarang, jumlah eritrosit, kadar hemoglobin, dan diameter alveolus kelelawar (rata-rata dan standar deviasi. n=3)
A
B
C (g)
D (%)
E oC
F (%)
G Ppm
H juta/ml
I g/ml
J (µm)
K juta/ml/g
L g/m/g
C_pli
Petruk R4
12.8±0.75
80.4 ±0.00
30.8 ±0.00
22.8 ±0.52
3310
13.0±0.20
13.0 ±0.20
114.2±120
0.39±0.03
1.13±0.04
H_at
Petruk R7
6.0± 0.28
77.2 ±0.20
26.4 ±0.34
22.0 ±0.48
2490
2.9±0.26
8.83±0.20
128.8±3.20
0.47±0.02
1.42±0.08
H.cf_at
Petruk R8
6.7± 0.14
75.5±2.10
27.5±2.12
21.8±0.28
2340
6.1±4.13
9.00±0.28
130.4±12.00
0.47±0.01
1.34±0.01
Hip_sp
Kampil R3
17.1± 0.24
70.8±1.10
27.9±0.18
22.07±0.52
1901
4.8±0.49
12.4±0.81
131.5±24.00
0.29±0.03
0.90±0.06
H_so1
Celeng R4
14.7±1.42
71.8±1.10
27.9±0.18
22.0±0.52
1901
4.1±0.90
12.47±0.80
143.9±8.70
0.30±0.04
0.96±0.06
H_so2
Jatijajar R2
13.7± 0.5
70.4±0.00
27.8±0.00
22.8±0.00
1830
3.5±0.10
11.7±0.7
140.8±16.00
0.25±0.03 0.85±0.01
H_so3
Petruk R3
14.3± 0.45
72.2±0.00 28.6±0.00 21.6±0.00 2090 4.2±0.30
12.6±1.10 138.4±26.00 0.29±0.01 0.88±0.03
M_aus
Inten R2
5.25± 0.21
64.0±0.2
26.7±0.11
22.1±0.16
1872
3.6±0.45
9.8±0.30
118.1±10.00
0.69±0.01
1.65±0.08
M_sch
Liyah R2
10.7±0.70
74.1±0.00 27±0.00 20.9±0.05 3180 3.2±0.21
8.65±0.07 123.5±22.00 0.33±0.02 0.80±0.02
R_aff
Inten R3
11.4±0.35
70.0±0.00
29.53±0.11
22.0±0.80
2810
12.4±0.52
12.4±0.50
112.2±11.00
0.32±0.01
0.96±0.07
C_bra
Liyah R1
37.0±1.46
54.8±1.20 27.24±0.21 21.0±0.10 1202 13.2±0.90
13.2±0.90 169.3±26.00 0.12±0.01 0.37±0.03
E_spa
Dempo R2
38.9±6.32
63.0±0.80
28.2±0.21
21.9±0.83
1103
13.3±1.50
13.3±1.50
158.2±12.00
0.11±0.01
0.34±0.04
R_amp
Petruk R2
83.2 ±1.4
56.8±0.00
27.0±0.00
20.9±0.00
640
16.15±2.80
14.0±0.30 144.8±16.00
0.06±0.03
0.19±0.02
Keterangan : A= spesies kelelawar B= goa lokasi sarang C=bobot badan D= kelembapan E= suhu udara F= kadar oksigen G= kadar ammonia H = jumlah eritrosit I = kadar hemoglobin J= diameter alveolus K= rasio jumlah eritrosit/ bobot badan L=rasio kadar hemoglobin / bobot badan Hip_sp=Hipposideros sp C_bra=C.brachyotis H_so = H.sorenseni E_spa =E.spelaea C_pli= C.plicata M_aus=M.australis R_amp = R.amplexicaudatus H_at = H.ater M_sch=M.Schreibersii Hcf_at= H.cf.ater R_aff=R.affinis

94
berkadar amonia rendah (Hipposideros sp, H.sorenseni1, C.brachiotis, E.speleae
dan R.amplexicaudatus).
Tabel 17 Hasil uji Tukey perbandingan diameter alveolus antarkoloni kelelawar
C_pl1
H_at
H.cf_a
Hip_sp
H_s_1
H_s-2
H_s-3
M_au
M_sc
R_af
C_br
E_sp
R_a
C_pl 1
* * * * * * ns ns * * * *
H_at 1
ns * * * * * * * * * *
H.cf_a1
* * * * * * * * * *
Hp_sp11
Ns * * * * * * * *
H_s-11
ns
ns
ns
*
*
*
*
*
H_s_21
ns
*
*
*
*
*
*
H_s_31
* * * * * *
M_au1
*
ns
*
*
*
M_sc2 * * * *
R_af 2 * * *
C_br 2 ns ns
E_sp2 ns
Keterangan : ns= tidak berbeda nyata pada taraf 0.05 *= berbeda nyata pada taraf 0.05
1 = Microcheroptera 2 = Megachiroptera
Berdasarkan hasil uji korelasi Spearman (Tabel 16), terbukti bahwa
diameter alveolus berkorelasi nyata dengan kadar amonia, dan tidak berkorelasi
nyata dengan suhu, kelembapan, dan kadar oksigen. Korelasi antara diameter
alveolus dengan kadar amonia bersifat negatif (R= -0,342, P< 0,05), artinya
semakin tinggi kadar amonia, diameter alveolus kelelawar semakin kecil. Bentuk
hubungan antara kelembapan, suhu, kadar oksigen, dan kadar amonia dengan
diameter alveolus kelelawar dapat dijelaskan dengan persamaan regesi linear
berganda sebagai berikut:
Y= 245.8 + 0.121 X1 + 0.006 X2 – 0.209X3 – 0.828 X4
(R= 0.838; F= 4.72; P=0.03)
Y = diameter alveolus X1 = suhu
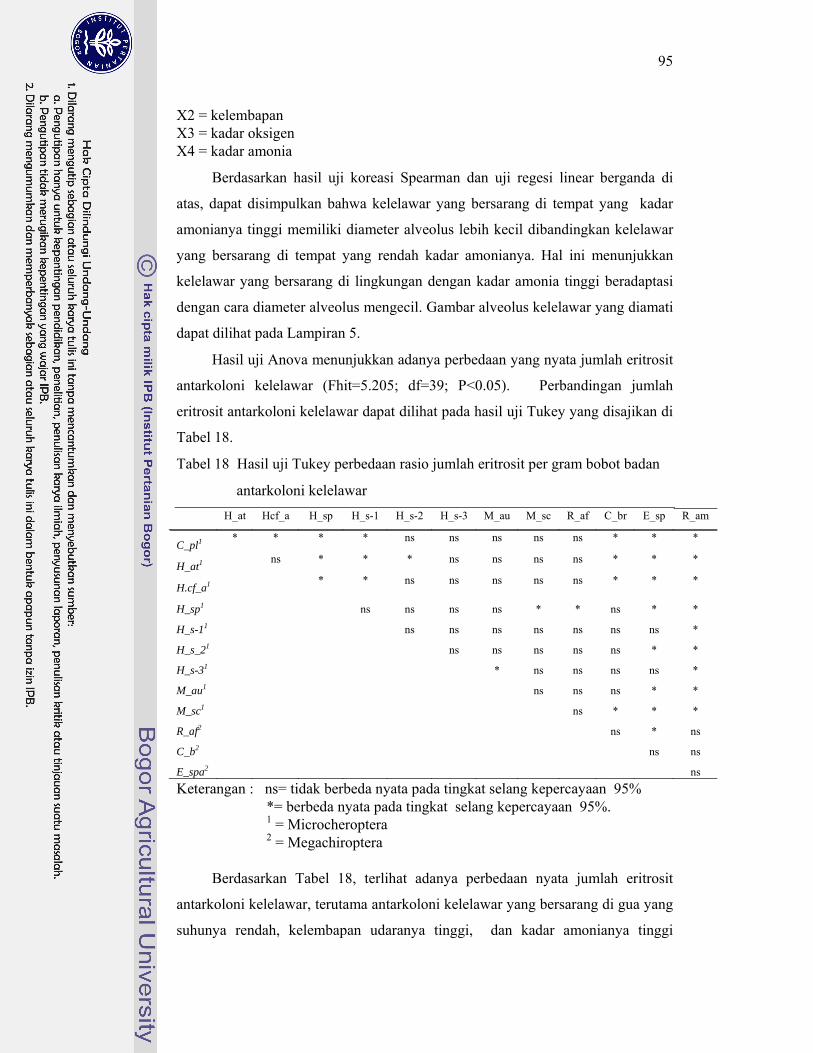
95
X2 = kelembapan X3 = kadar oksigen X4 = kadar amonia
Berdasarkan hasil uji koreasi Spearman dan uji regesi linear berganda di
atas, dapat disimpulkan bahwa kelelawar yang bersarang di tempat yang kadar
amonianya tinggi memiliki diameter alveolus lebih kecil dibandingkan kelelawar
yang bersarang di tempat yang rendah kadar amonianya. Hal ini menunjukkan
kelelawar yang bersarang di lingkungan dengan kadar amonia tinggi beradaptasi
dengan cara diameter alveolus mengecil. Gambar alveolus kelelawar yang diamati
dapat dilihat pada Lampiran 5.
Hasil uji Anova menunjukkan adanya perbedaan yang nyata jumlah eritrosit
antarkoloni kelelawar (Fhit=5.205; df=39; P<0.05). Perbandingan jumlah
eritrosit antarkoloni kelelawar dapat dilihat pada hasil uji Tukey yang disajikan di
Tabel 18.
Tabel 18 Hasil uji Tukey perbedaan rasio jumlah eritrosit per gram bobot badan
antarkoloni kelelawar
H_at Hcf_a H_sp H_s-1 H_s-2 H_s-3 M_au M_sc R_af C_br E_sp R_am
C_pl1 * * * * ns ns ns ns ns * * *
H_at1 ns * * * ns ns ns ns * * *
H.cf_a1 * * ns ns ns ns ns * * *
H_sp1
ns ns ns ns * * ns * *
H_s-11 ns ns ns ns ns ns ns *
H_s_21 ns ns ns ns ns * *
H_s-31 * ns ns ns ns *
M_au1 ns ns ns * *
M_sc1 ns * * *
R_af2 ns * ns
C_b2 ns ns
E_spa2 ns Keterangan : ns= tidak berbeda nyata pada tingkat selang kepercayaan 95% *= berbeda nyata pada tingkat selang kepercayaan 95%.
1 = Microcheroptera 2 = Megachiroptera Berdasarkan Tabel 18, terlihat adanya perbedaan nyata jumlah eritrosit
antarkoloni kelelawar, terutama antarkoloni kelelawar yang bersarang di gua yang
suhunya rendah, kelembapan udaranya tinggi, dan kadar amonianya tinggi

96
(H. ater, H. cf. ater , dan M. schreibersii) dibandingkan jumlah eritrosit kelelawar
yang bersarang di gua yang suhunya tinggi, kelembapan udaranya rendah, dan
kadar amonianya rendah (H. sorenseni 1, E. spelaea, C. brachyotis dan R.
amplexicaudatus). Hal ini sesuai dengan hasil uji korelasi Spearman pada Tabel
16, yang menunjukkan adanya korelasi nyata antara jumlah eritrosit kelelawar
dengan suhu, kelembapan udara, dan kadar amonia. Bentuk hubungan antara
kadar eritrosit kelelawar dengan suhu, kelembapan, kadar oksigen dan kadar
amonia dapat dijelaskan dengan persamaan regesi linear berganda sebagai
berikut:
Y= 1601471.037 – 0.487 X1 + 0.384 X2 – 0.043X3 + 0.732X4
(R= 0.962; F= 24.6; P= 0.00).
Y= diameter alveolus X1 = suhu X2 = kelembapan X3 = kadar oksigen X4 = kadar amonia
Berdasarkan hasil uji korelasi Spearman pada Tabel 16 tampak bahwa
korelasi antara jumlah eritrosit dengan suhu udara bersifat negatif (R= -305,
P<0.05), artinya kelelawar yang bersarang di tempat bersuhu rendah mempunyai
jumlah eritrosit lebih tinggi dibandingkan kelelawar yang bersarang di tempat
bersuhu tinggi. Korelasi antara jumlah eritrosit dengan kelembapan udara bersifat
positif (R= R= 0.793, P<0.05), artinya kelelawar yang bersarang di tempat
berkelembapan udara tinggi memiliki jumlah eritrosit lebih tinggi dibandingkan
kelelawar yang bersarang di tempat berkelembapan udara rendah. Demikian pula
halnya dengan kadar amonia. Korelasi antara jumlah eritrosit dengan kadar
amonia berkorelasi positif, artinya kelelawar yang bersarang di tempat berkadar
amonia tinggi memiliki jumlah eritrosit lebih tinggi dibandingkan kelelawar yang
bersarang di tempat berkadar amonia rendah. Persamaan regesi linear berganda
menunjukkan jumlah eritrosit bertambah seiring dengan menurunnya suhu udara,
bertambahnya kelembapan udara dan meningkatnya kadar amonia. Hal ini
membuktikan adanya adaptasi fisiologi kelelawar terhadap mikroklimat dingin,
lembap dan kadar amonia tinggi yaitu dengan jemlah eritrosit yang meningkat.
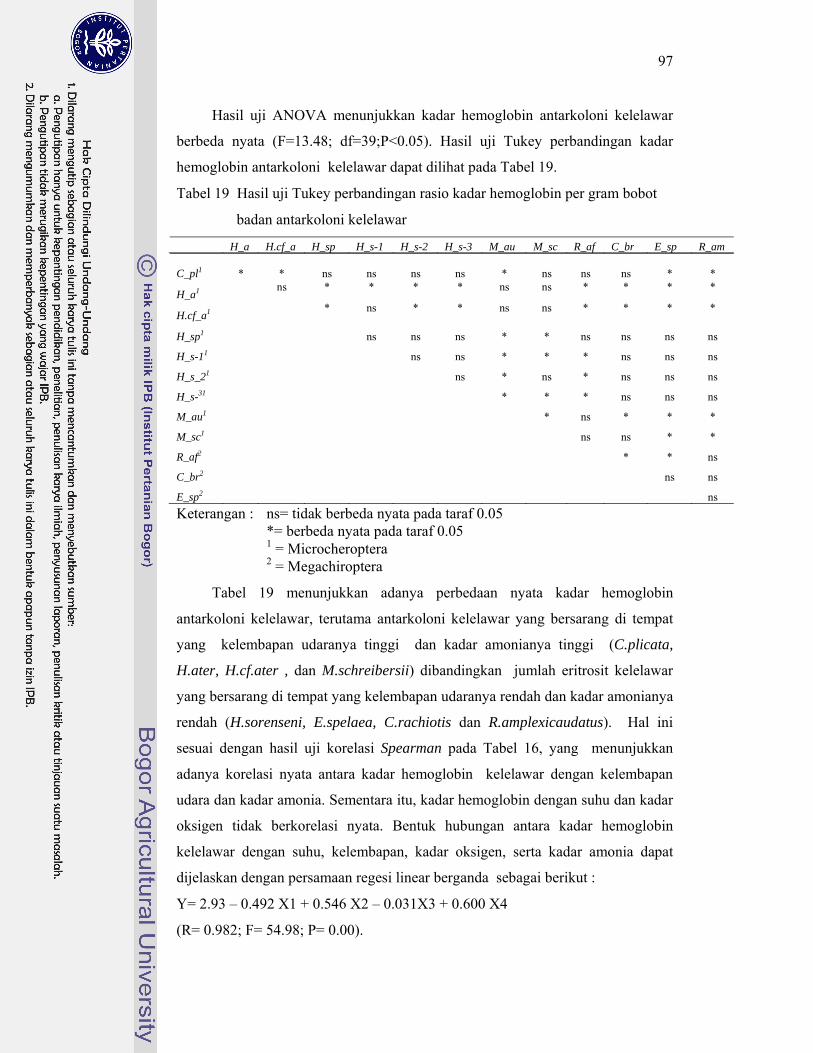
97
Hasil uji ANOVA menunjukkan kadar hemoglobin antarkoloni kelelawar
berbeda nyata (F=13.48; df=39;P<0.05). Hasil uji Tukey perbandingan kadar
hemoglobin antarkoloni kelelawar dapat dilihat pada Tabel 19.
Tabel 19 Hasil uji Tukey perbandingan rasio kadar hemoglobin per gram bobot
badan antarkoloni kelelawar H_a H.cf_a H_sp H_s-1 H_s-2 H_s-3 M_au M_sc R_af C_br E_sp R_am
C_pl1 *
*
ns
ns
ns
ns
*
ns
ns
ns
* *
H_a1 ns * * * * ns ns * * * *
H.cf_a1 * ns * * ns ns * * * *
H_sp1
ns ns ns * * ns ns ns ns
H_s-11 ns ns * * * ns ns ns
H_s_21 ns * ns * ns ns ns
H_s-31 * * * ns ns ns
M_au1 * ns * * *
M_sc1 ns ns * *
R_af2 * * ns
C_br2 ns ns
E_sp2 ns Keterangan : ns= tidak berbeda nyata pada taraf 0.05 *= berbeda nyata pada taraf 0.05
1 = Microcheroptera 2 = Megachiroptera
Tabel 19 menunjukkan adanya perbedaan nyata kadar hemoglobin
antarkoloni kelelawar, terutama antarkoloni kelelawar yang bersarang di tempat
yang kelembapan udaranya tinggi dan kadar amonianya tinggi (C.plicata,
H.ater, H.cf.ater , dan M.schreibersii) dibandingkan jumlah eritrosit kelelawar
yang bersarang di tempat yang kelembapan udaranya rendah dan kadar amonianya
rendah (H.sorenseni, E.spelaea, C.rachiotis dan R.amplexicaudatus). Hal ini
sesuai dengan hasil uji korelasi Spearman pada Tabel 16, yang menunjukkan
adanya korelasi nyata antara kadar hemoglobin kelelawar dengan kelembapan
udara dan kadar amonia. Sementara itu, kadar hemoglobin dengan suhu dan kadar
oksigen tidak berkorelasi nyata. Bentuk hubungan antara kadar hemoglobin
kelelawar dengan suhu, kelembapan, kadar oksigen, serta kadar amonia dapat
dijelaskan dengan persamaan regesi linear berganda sebagai berikut :
Y= 2.93 – 0.492 X1 + 0.546 X2 – 0.031X3 + 0.600 X4
(R= 0.982; F= 54.98; P= 0.00).

98
Y= kadar hemoglobin X1 = suhu X2 = kelembapan X3 = kadar oksigen X4 = kadar amonia
Berdasarkan hasil uji korelasi Spearman pada Tabel 16, tampak bahwa
korelasi antara kadar hemoglobin dan kelembapan udara bersifat positif, artinya
kelelawar yang bersarang di tempat berkelembapan udara tinggi memiliki kadar
hemoglobin lebih tinggi dibandingkan kelelawar yang bersarang di tempat
berkelembapan udara rendah. Demikian pula halnya dengan kadar amonia.
Korelasi antara kadar hemoglobin dengan kadar amonia bersifat positif, artinya
kelelawar yang bersarang di tempat berkadar amonia tinggi memiliki kadar
hemoglobin lebih tinggi dibandingkan dengan kelelawar yang bersarang di gua
dengan kadar amonia rendah. Persamaan regesi linear berganda menunjukkan
kadar amonia bertambah seiring dengan menurunnya suhu, bertambahnya
kelembapan udara dan bertambahnya kadar amonia. Hal ini membuktikan adanya
adaptasi fisiologi kelelawar yang bersarang di tempat dengan kelembapan tinggi
dan kadar amonia tinggi yaitu dengan kadar hemoglobin meningkat.
Pembahasan
Hasil pengukuran suhu di sekitar sarang kelelawar, menunjukkan semua
sarang berada dalam kisaran suhu optimal untuk hewan Mamalia yaitu 25oC s/d
30 oC . Hal ini sesuai dengan pendapat Frandson (1992) bahwa suhu lingkungan
optimal untuk Mamalia di daerah tropis berkisar antara 25oC s/d 32oC. Hasil
pengukuran kadar oksigen menunjukkan tidak terjadi penurunan kadar oksigen
pada sarang kelelawar yang berada di dalam gua. Hal ini berbeda dengan hasil
penelitian Baudinette et al. (1994) di gua-gua di Australia yang mendapatkan
kondisi udara di sekitar sarang kelelawar yang berada di dalam gua memiliki
kadar oksigen 10% lebih rendah dibandingkan kadar oksigen di luar gua. Pada gua
karst yang diamati dalam penelitian ini, kadar oksigen tidak menurun karena pori-
pori karst pada dinding dan atap gua dapat dilewati udara sehingga oksigen dapat
masuk melalui pori-pori tersebut. Masuknya udara melalui pori-pori karst ini
menyebabkan oksigen yang berkurang karena digunakan oleh kelelawar, dapat

99
bertambah lagi. Oleh sebab itu, dalam penelitian ini kadar oksigen tidak
berpengaruh pada diameter alveolus, jumlah eritrosit dan kadar hemoglobin,
karena kadar oksigen udara di semua sarang berada dalam kisaran normal ( >
20.9%).
Hasil pengukuran kelembapan udara menunjukkan beberapa sarang
memiliki kelembapan udara tinggi ( mencapai 80%). Tingginya kelembapan
udara di beberapa sarang kelelawar tersebut disebabkan banyaknya rembesan air
di dinding dan atap gua, serta adanya aliran sungai bawah tanah. Sebagai
pembanding, menurut peraturan Menteri Kesehatan RI No. 24 tahun 2001, batas
optimal kelembapan udara relatif di lingkungan kerja manusia adalah 45% s/d
56% (DEPKES RI 2001). Hasil pengukuran kadar amonia di beberapa lokasi
sarang kelelawar juga menunjukkan kadar amonia yang tinggi (mencapai 3310
ppm). Tingginya kadar amonia tersebut berasal dari urine dan feses kelelawar.
Penelitian Sridhar et al. (2006) mendapatkan urin dan feses (guano) kelelawar
Hipposideros speoris (Hipposideridae: Mikrochiroptera) tersusun atas 5.7 ± 1.5%
nitrogen (N) berbentuk amonia (NH3). Amonia dalam guano ini dapat menguap
menjadi gas bercampur dengan komponen udara lainnya. Hal ini menyebabkan
kandungan amonia udara meningkat tajam (Shidar et al. 2006). Sebagai
pembanding, menurut Peraturan Pemerintah RI No 46 Tahun 1999, kadar amonia
udara yang boleh ada pada udara ambien normal tidak lebih dari 60 µg/m3 atau
60 ppm (KLH 2003). Hasil penelitian ini menunjukkan, bila dibandingkan
dengan lingkungan manusia, kelembapan dan kadar amonia di beberapa sarang
kelelawar jauh di atas batas normal. Oleh karena itu, agar dapat bertahan hidup
dalam sarang tersebut, kelelawar perlu beradaptasi. Hal ini sesuai dengan
pendapat Alikodra (2010) bahwa untuk bertahan di suatu habitat, hewan harus
mengembangkan strategi adaptasi.
Hasil penelitian ini membuktikan adanya adaptasi struktural organ
pernapasan kelelawar sebagai strategi untuk dapat bertahan hidup di sarang
dengan kadar amonia tinggi. Hal ini dibuktikan dengan adanya korelasi yang
nyata antara diameter alveolus kelelawar dengan kadar amonia. Kelelawar yang
bersarang di gua berkadar amonia tinggi, memiliki diameter alveolus lebih kecil
dibandingkan kelelawar yang bersarang di gua berkadar amonia rendah. Menurut

100
Guyton (1995), dinding alveolus sangat tipis dan di dalamnya banyak terdapat
kapiler darah. Kondisi demikian memudahkan pertukaran gas antara alveolus
dengan kapiler darah. Menurut Plopper & Adams (1993), alveolus yang
berukuran kecil dan rapat, menyebabkan permukaan membran respirasi menjadi
lebih luas. Permukaan membran respirasi yang luas menyebabkan paru-paru
mampu mengikat oksigen lebih banyak (Plopper dan Adams 1993). Selain itu
menurut Guyton (1995), semakin sempit diameter alveolus, semakin besar
tekanan parsialnya, hal ini menyebabkan semakin cepat absorbsi oksigen oleh
kapiler darah. Kemampuan mengikat oksigen lebih banyak dan absorbsi oksigen
lebih cepat diperlukan oleh kelelawar yang bersarang di lingkungan berkadar
amonia tinggi, karena kelelawar memerlukan lebih banyak energi untuk memacu
terbentuknya limfosit dan zat antibodi. Hasil penelitian Setiadi (2000)
menunjukkan kelelawar Scotophilus kuhli (Vespertilionidae: Mikrochiroptera)
memiliki diameter alveolus lebih kecil dan rapat dibandingkan alveolus tikus
(Mus musculus), karena sebagai Mamalia terbang, kelelawar membutuhkan energi
lebih banyak dibandingkan tikus.
Dalam fisiologi pernapasan Mamalia, setelah oksigen berdifusi ke kepiler
darah, oksigen akan diangkut oleh darah ke seluruh jaringan tubuh oleh sel darah
merah (eritrosit). Kemampuan eritrosit mengangkut oksigen disebabkan adanya
pigmen respirasi atau hemoglobin (Frandson 1992). Menurut Campbell et al.
(2002) oksigen sangat tidak larut dalam air sehingga untuk mengangkut oksigen,
diperlukan hemoglobin. Hemoglobin tersebut terdiri atas empat subunit protein
berbentuk globul (bola), setiap globul memiliki satu gugus hem dengan satu atom
besi (Fe). Atom besi inilah yang dapat mengikat oksigen. Semakin banyak jumlah
eritrosit, semakin banyak kadar hemoglobin dalam darah Mamalia, semakin
banyak oksigen yang dapat ditransportasikan.
Jumlah eritrosit kelelawar Microchiroptera yang ditemukan dalam
penelitian ini berkisar antara 2.9 juta/ml s/d 13.03 juta/ ml, dan jumlah eritrosit
kelelawar Megachiroptera yang ditemukan dalam penelitian ini berkisar antara
13.2 juta/ml s/d 16.15 juta/ml. Riedesel (1977) mencatat hasil-hasil penelitian
mengenai eritrosit beberapa jenis kelelawar adalah sebagai berikut : Myotis
sodalis 9.5 juta/ml; Nyctalus noctula 10.61 juta/ml s/d 14.38 juta/ml; Tadarida

101
brasiliensis mexicana 10.11 juta/ml s/d 16.53 juta/ml; Desmodus rotundus
murinus 8.20 juta/ml s/d 12.39 juta/ml dan Eptesicus fuscus 11.96 juta/ml. Bila
dibandingkan dengan penelitian terdahulu, jumlah eritrosit kelelawar yang
ditemukan dalam penelitian ini berada dalam kisaran yang lebih luas ( 2.9 juta/ml
s/d 16.15 juta/ml). Jumlah eritrosit yang sangat bervariasi ini disebabkan sampel
kelelawar yang diteliti memiliki ukuran tubuh dan habitat yang sangat bervariasi
pula. Menurut Ganong (2001), faktor yang mempengaruhi jumlah eritrosit hewan
adalah kondisi nutrisi, aktivitas fisik, umur dan lingkungan (Ganong 2001).
Cunningham (2002) menjelaskan jumlah eritrosit jenis-jenis hewan Mamalia
sebagai berikut : kucing (Felis felis domesticus) 6 juta/ ml-8 juta/ml; sapi (Bos
javanicus) 6 juta/ ml -8 juta/ml; anjing (Canis familiaris) 6 juta/ml s/d 8 juta/ml ;
kambing (Capra aegagus) 13 juta/ml s/d 14 juta/ml; babi (Sus barbatus) 6 juta/ml
s/d 8juta/ml ; dan kelinci (Lepus negicollis) 5.5 juta/ml s/d 6.5 juta/ml. Bila
dibandingkan dengan Mamalia lain, jumlah eritrosit kelelawar jauh lebih tinggi
dibandingkan jumlah eritrosit Mamalia lain. Menurut Altringham (1996), pada
waktu terbang kelelawar membutuhkan banyak energi sehingga lebih banyak
oksigen yang dikonsumsi. Oleh karena itu, untuk memenuhi kebutuhan oksigen
yang besar, kelelawar memiliki jumlah eritrosit lebih banyak dibandingkan
Mamalia lain.
Hasil penelitian ini membuktikan adanya adaptasi fisiologi pernapasan oleh
kelelawar yang bersarang di tempat yang dingin, lembap dan berkadar amonia
tinggi yaitu dengan peningkatan rasio jumlah eritrosit per bobot badan dan rasio
kadar hemoglobin per bobot badan. Rasio jumlah eritrosit per bobot badan
kelelawar yang bersarang di gua yang dingin dan lembap, lebih tinggi
dibandingkan rasio jumlah eritrosit per bobot badan kelelawar yang bersarang di
gua yang hangat dan kering. Hal ini karena kelelawar yang bersarang pada tempat
dingin dan lembap harus memacu metabolisme tubuhnya agar tubuh tetap hangat.
Hasil penelitian ini sejalan dengan hasil penelitian Baudinette et al. (2000) yang
membuktikan, kelelawar yang bersarang pada kondisi udara dingin dan lembap
mengalami peningkatan laju respirasi hingga 13.47± 0.44 BMR (Basal Metabolic
Rate = 1 ml O2 g-1 h1). Menurut Atringham (1996), kelelawar merupakan
hewan homoikiloterm. Thermoregulasi kelelawar menyebabkan suhu tubuh tetap

102
hangat, walaupun berada di lingkungan yang dingin. Meningkatnya laju respirasi
yang didukung oleh meningkatnya jumlah eritrosit merupakan salah satu bentuk
thermoregulasi kelelawar di lingkungan yang dingin agar suhu tubuh tetap hangat.
Rasio jumlah eritrosit per gram bobot badan kelelawar yang bersarang di
gua berkadar amonia tinggi, juga lebih banyak dibandingkan rasio jumlah eritrosit
per gram bobot badan kelelawar yang bersarang di gua yang kadar amonianya
rendah. Menurut Guyton (1995), masuknya zat racun ke dalam darah dapat
menurunkan afinitas (daya ikat) oksigen oleh hemoglobin. Hal ini dapat
menyebabkan sel kekurangan oksigen (hipoksia). Sebagai bentuk strategi adaptasi
pada keadaan ini, kelelawar memacu produksi sel eritrositnya. Mekanisme
peningkatan jumlah eritrosit dalam tubuh Mamalia adalah sebagai berikut
(Ganong 2001): penurunan kadar oksigen (hipoksia) akan merangsang ginjal
untuk melepaskan enzim eritrogenin (erythrogenin). Selanjutnya enzim tersebut
akan mengaktifkan eritropoietinogen yang merupakan prekusor dalam
pembentukan eritopoietin. Di dalam sumsum tulang, eritropoietin akan
meningkatkan jumlah sel bakal (stem cell). Sel bakal ini akan menjadi prekusor
darah merah dan selanjutnya menjadi sel darah merah (Ganong 2001). Selain itu
menurut Guyton (1995), apabila zat asing masuk ke dalam tubuh, tubuh akan
membentuk antibodi dan meningkatkan jumlah sel limfosit untuk menghancurkan
zat asing. Oleh karena itu, apabila amonia masuk ke dalam tubuh, kelelawar
membutuhkan lebih banyak energi untuk memacu peningkatan antibodi dan sel
limfosit tersebut. Kebutuhan energi yang banyak ini diimbangi dengan
peningkatan jumlah eritrosit.
Peningkatan kadar hemoglobin kelelawar sejalan dengan peningkatan
jumlah eritrosit. Oleh karena itu, kelelawar yang bersarang di gua dengan kondisi
udara dingin, lembap, dan amonia tinggi memiliki kadar hemoglobin lebih tinggi
dibandingkan kelelawar yang bersarang di gua dengan kondisi hangat, kering, dan
kadar amonia rendah. Peningkatan kadar hemoglobin tersebut merupakan
strategi agar laju respirasi meningkat. Dengan demikian, suhu tubuh dapat
dipertahankan dan masuknya racun ke dalam tubuh dapat segera dinetralisir.
Selain itu menurut Campbell et al. (2002) kadar hemoglobin yang tinggi
merupakan adaptasi hewan untuk menyimpan oksigen. Hal ini dibuktikan pada

103
Mamalia air : anjing laut Weddell, yang mampu menyimpan 70% kebutuhan
oksigennya dalam darah. Dengan kadar hemoglobin yang tinggi, kelelawar dapat
menyimpan oksigen dalam jumlah besar dan kekurangan oksigen dapat dihindari.
Riedesel (1977) mencatat hasil-hasil penelitian mengenai hemoglobin
beberapa jenis kelelawar adalah sebagai berikut: Nyctalus noctula 11.1 g/ml;
Myotis myotis 10.9 g/ml; Plecotus auritus 9.8 g/ml; Myotis nattereri 10.9; dan
Artibeus lituratus 17.0 g/ml. Menurut Guyton (1995) dalam keadaan normal 1 ml
darah manusia mengandung 14 gram (14 g/mm3) hemoglobin yang mampu
mengangkut 0.03 gram oksigen. Penelitian Nugraha (2007) mendapatkan
kandungan hemoglobin anjing (Canis familiaris) usia 3 sampai 7 bulan berkisar
5.77 s/d 10.94 g/mm3. Fatmawati (2007) mendapatkan hemoglobin anjing ras
Dobermean dewasa berkisar 11.8 ± 1.13 g/mm3. Bila dibandingkan dengan hewan
Mamalia lain, kadar hemoglobin dalam darah kelelawar yang ditemukan dalam
penelitian ini cenderung lebih tinggi. Hal ini karena, sebagai Mamalia terbang
kelelawar membutuhkan energi lebih banyak dibandingkan Mamalia lain.
Kesimpulan
Strategi adaptasi struktural dan fisiologi kelelawar yang bersarang di gua
dengan kondisi udara dingin, lembap dan kadar amonia tinggi adalah dengan
diameter alveolus yang kecil, jumlah eritrosit meningkat dan kadar hemoglobin
meningkat.
Saran
Penelitian ini merupakan penelitian pendahuluan dalam rangka memahami
strategi adaptasi kelelawar penghuni gua. Oleh karena itu masih diperlukan
penelitian lebih lanjut yang meliputi : struktur trakhea, struktur jantung,
hematologi darah, serta laju metabolisme kelelawar penghuni gua.

104
PEMBAHASAN UMUM
Pentingnya upaya konservasi bagi ekosistem Karst Gombong
Hasil penelitian ini menunjukkan bahwa di gua-gua Karst Gombong hidup
lima belas jenis kelelawar, yang terdiri atas empat jenis Megachiroptera, dan
sebelas jenis Microchiroptera. Hal ini menunjukkan bahwa Karst Gombong
menyimpan kekayaan jenis kelelawar yang jauh lebih tinggi dibandingkan karst
lain yang berada di Indonesia maupun di luar Indonesia, yaitu di Karst Sumbawa
delapan jenis (Maryanto & Maharadatunkamsi 1991), di Karst Sangkulirang-
Mangkaliat Kalimantan Timur sembilan jenis (Saroni 2005), di Karst Istambul
Turki: delapan jenis (Furman & Ozgull 2002) dan di Karst Britain Inggis: sebelas
jenis (Parsons et al. 2002).
Lima belas jenis kelelawar yang bersarang di gua-gua Karst Gombong
tersebut, delapan jenis di antaranya termasuk jenis yang dilindungi berdasarkan
Red List IUCN versi 3.1 (IUCN 2001). Satu jenis termasuk dalam kategori
vulnerable, satu jenis termasuk dalam kategori near threatened dan enam jenis
termasuk dalam kategori lower risk /least concern. Status kelelawar berdasarkan
Red List IUCN versi 3.1 dapat dilihat pada Tabel 20.
Tabel 20 Status konservasi kelelawar berdasarkan Red List IUCN versi 3.1
(IUCN 2001)
Jenis Kelelawar
Status Konservasi
(Red List IUCN versi 3.1)
Keterangan status
H. sorenseni Vulnerable Memiliki risiko tinggi untuk punah di alam karena penurunan jumlah populasi yang tinggi dan penyebaran yang sangat terbatas.
M. schreibersii Near threatened Diperkirakan akan terancam punah dalam waktu dekat
H. ater M. australis H. bicolor R. borneensis R. affinis R. amplexicaudatus
Lower risk / least concern
Tidak memenuhi syarat untuk dimasukkan ke dalam critically endangered, endangered, vulnerable ataupun near threatened karena memiliki risiko rendah untuk punah.

105
Hasil analisis relung pakan membuktikan bahwa kelelawar
Microchiroptera yang bersarang di Karst Gombong merupakan predator 29 famili
serangga yang berpotensi sebagai hama pertanian sedangkan kelelawar
Megachiroptera yang ditemukan di Karst Gombong merupakan polinator 33
genus tumbuhan hutan dan tanaman perkebunan yang bernilai ekonomi. Menurut
John et al. (1990) suatu kawasan perlu dilindungi atas dasar beberapa kriteria
yaitu 1) Karakteristik atau keunikan ekosistem; 2) Spesies khusus yang diminati,
bernilai penting, kelangkaan atau terancam 3) Memiliki keanekaragaman spesies
tinggi 4) Landskap atau ciri geofisik yang bernilai eksotik; 5) Fungsi
perlindungan hidrologi; 6) Potensial untuk wisata alam; 7) Tempat peninggalan
budaya. Berdasarkan kriteria tersebut, hasil penelitian ini menunjukkan bahwa
Karst Gombong perlu dilindungi karena memiliki keunikan ekosistem, dihuni oleh
spesies yang terancam, memiliki kanekaragaman spesies yang tinggi, memiliki
fungsi perlindungan hidrologi, dan potensial untuk wisata alam.
Selama ini pengelolaan gua yang dilakukan PEMDA Kabupaten Kebumen
lebih terfokus pada pemanfaatan gua sebagai sumber Pendapatan Asli Daerah
(PAD), yaitu dengan memanfaatkan gua karst sebagai objek wisata. Sejak tahun
1976, PEMDA Kabupaten Kebumen telah mengembangkan Gua Jatijajar sebagai
objek wisata minat umum. Selanjutnya pada tahun 1984 Gua Petruk juga
ditetapkan sebagai objek wisata. Namun, berbeda dari Gua Jatijajar, Gua Petruk
ditetapkan sebagai objek wisata minat khusus. Menurut Dinas Pariwisata
Kabupaten Kebumen (2004), objek wisata alam minat umum adalah kawasan
yang memiliki keindahan alam dan dikelola untuk menarik wisatawan sebanyak
mungkin. Objek wisata minat khusus adalah kawasan wisata yang dikelola bagi
wisatawan dengan persyaratan khusus, misalnya kegiatan penelusuran gua yang
memerlukan peralatan standar dan pemandu yang terampil. Sangat disayangkan,
pemanfaatan Gua Jatijajar dan Gua Petruk untuk tujuan wisata tersebut, selama ini
masih belum dilandasi oleh dasar ilmu pengetahuan yang kuat sehingga sangat
berisiko menimbulkan kerusakan ekosistem. Hal ini terlihat pada pengelolaannya
yang lebih berorientasi bisnis tanpa mempertimbangkan resiko kerusakan
ekosistem. Sebagai contoh, untuk menarik kedatangan wisatawan di Gua Jatijajar,
pada dinding dan atap gua dipasang lampu penerangan dan dibuat berbagai

106
deorama yang menggambarkan cerita pewayangan Rama dan Shinta (Gambar 25).
Selain itu, di luar gua juga dibangun berbagai fasilitas wisata seperti pasar
souvenir, taman bermain, panggung hiburan dan jalan. Hasil penelitian ini
membuktikan bahwa di Gua Jatijajar bersarang (dihuni) lima jenis kalelewar, dan
dua jenis di antaranya (R.amplexicaudatus dan H.sorenseni) termasuk dalam
jenis-jenis yang perlu dilindungi berdasarkan Red List IUCN versi 3.1. Dalam
memilih sarangnya, jenis-jenis kelelawar tersebut memerlukan persyaratan fisik
mikroklimat yang spesifik, sesuai dengan fisiologis tubuhnya. Pembangunan
fasilitas wisata di Gua Jatijajar dapat menyebabkan perubahan mikroklimat gua
menjadi tidak sesuai lagi dengan kondisi yang dibutuhkan kelelawar. Kondisi ini
akan mengakibatkan kepunahan jenis-jenis kelelawar tersebut. Berapa lama hal ini
akan terjadi sangat bergantung pada besarnya tekanan pada gua ini. Oleh karena
itu PEMDA Kabupaten Kebumen harus mempertimbangkan pola pengelolaan
yang diterapkan selama ini.
Gambar 25 Patung deorama cerita pewayangan Rama dan Shinta di
lorong Gua Jatijajar (Foto: Wijayanti 2010)
Di Gua Petruk, meskipun ditetapkan sebagai objek wisata minat khusus,
jumlah wisatawan yang menelusuri gua tidak dibatasi. Apalagi pada
pelaksanaannya tidak ada tata tertib yang mengatur aktivitas penelusuran gua
tersebut. Hasil penelitian ini membuktikan bahwa Gua Petruk dihuni oleh
sembilan jenis kelelawar dengan kelimpahan yang sangat tinggi. Di antara

107
sembilan jenis tersebut, lima jenis di antaranya termasuk jenis yang perlu
dilindungi berdasarkan Red List IUCN versi 3.1. Kedatangan wisatawan dengan
perilaku dan jumlah yang tidak terkontrol sangat berisiko mengganggu
kenyamanan kelelawar, dan pada akhirnya dapat menyebabkan kepunahan jenis
kelelawar tersebut. Sebetulnya risiko ini dapat di minimalisir apabila objek wisata
gua dikelola sesuai dengan persyaratan mikroklimat yang dibutuhkan kelelawar.
Persyaratan mikroklimat tersebut telah diketahui melalui hasil penelitian ini.
Untuk itu diperlukan kebijakan PEMDA yang mendukung ke arah pengelolaan
gua yang sesuai dengan persyaratan mikroklimat yang dibutuhkan sebagaimana
telah diketahui dari hasil penelitian ini.
Perhatian pemerintah terhadap ekosistem Karst Gombong
Menyadari sifat ekosistem karst yang unik dan rentan, sudah seharusnya
pemerintah pusat dan daerah mengeluarkan kebijakan yang mengarah pada
kelestariannya. Perhatian pemerintah pusat pada Karst Gombong mulai terlihat
sejak tahun 1987. Pada tahun tersebut, Dirjen PHKA (saat itu bernama Dirjen
PHPA) melakukan identifikasi fungsi gua di Provinsi Jawa Tengah, termasuk di
Karst Gombong, dan mengeluarkan pedoman pengelolaan gua berdasarkan hasil
identifikasi fungsi gua. Menurut pedoman pengelolaan tersebut, suatu gua bisa
ditetapkan sebagai gua konservasi, gua pendidikan, gua wisata, gua sumber air,
gua budaya, dan gua tambang berdasarkan kriteria pokok seperti tercantum pada
Tabel 21. Namun, identifikasi fungsi gua yang dilaksanakan pada tahun 1987
tersebut tidak pernah tuntas hingga saat ini sehingga rekomendasi pengelolaan
gua sesuai dengan pedoman yang ada tidak pernah terwujud. Dari 112 gua yang
ada di Karst Gombong hanya 3 gua yang teridentifikasi, yaitu Gua Jatijajar, Gua
Petruk, dan Gua Liyah, dan itupun baru sampai pada pemetaan lorong gua dan
identifikasi geohidrologi gua. Oleh karenanya, hingga saat ini, pedoman
pengelolaan gua tersebut hanya menjadi konsep saja bagi pengelola kawasan
Karst Gombong.
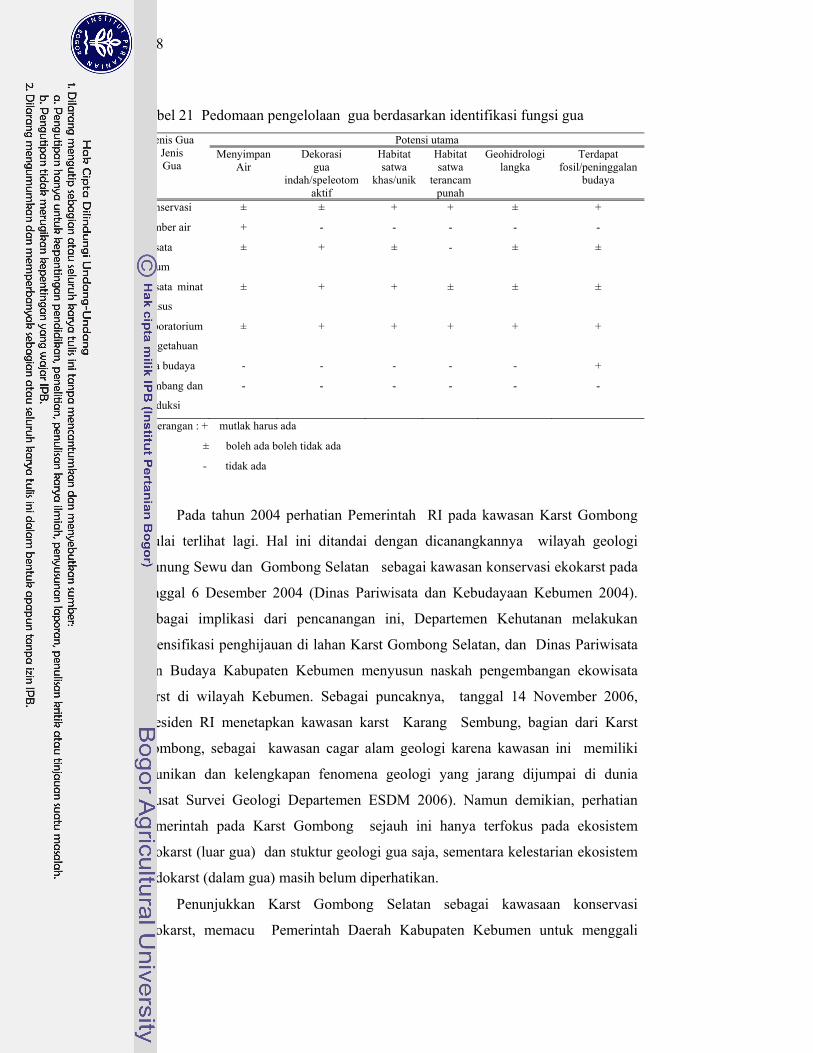
108
Tabel 21 Pedomaan pengelolaan gua berdasarkan identifikasi fungsi gua Jenis Gua
Jenis Gua
Potensi utama Menyimpan
Air Dekorasi
gua indah/speleotom
aktif
Habitat satwa
khas/unik
Habitat satwa
terancam punah
Geohidrologi langka
Terdapat fosil/peninggalan
budaya
Konservasi ± ± + + ± +
Sumber air + - - - - -
Wisata
umum
± + ± - ± ±
Wisata minat
khusus
± + + ± ± ±
Laboratorium
pengetahuan
± + + + + +
Gua budaya - - - - - +
Tambang dan
produksi
- - - - - -
Keterangan : + mutlak harus ada
± boleh ada boleh tidak ada
- tidak ada
Pada tahun 2004 perhatian Pemerintah RI pada kawasan Karst Gombong
mulai terlihat lagi. Hal ini ditandai dengan dicanangkannya wilayah geologi
Gunung Sewu dan Gombong Selatan sebagai kawasan konservasi ekokarst pada
tanggal 6 Desember 2004 (Dinas Pariwisata dan Kebudayaan Kebumen 2004).
Sebagai implikasi dari pencanangan ini, Departemen Kehutanan melakukan
intensifikasi penghijauan di lahan Karst Gombong Selatan, dan Dinas Pariwisata
dan Budaya Kabupaten Kebumen menyusun naskah pengembangan ekowisata
karst di wilayah Kebumen. Sebagai puncaknya, tanggal 14 November 2006,
Presiden RI menetapkan kawasan karst Karang Sembung, bagian dari Karst
Gombong, sebagai kawasan cagar alam geologi karena kawasan ini memiliki
keunikan dan kelengkapan fenomena geologi yang jarang dijumpai di dunia
(Pusat Survei Geologi Departemen ESDM 2006). Namun demikian, perhatian
pemerintah pada Karst Gombong sejauh ini hanya terfokus pada ekosistem
exokarst (luar gua) dan stuktur geologi gua saja, sementara kelestarian ekosistem
endokarst (dalam gua) masih belum diperhatikan.
Penunjukkan Karst Gombong Selatan sebagai kawasaan konservasi
exokarst, memacu Pemerintah Daerah Kabupaten Kebumen untuk menggali

109
potensi ekosistem Karst Gombong. Pada tahun 2004, Dinas Pariwisata dan
Budaya Kabupaten melakukan survei gua di seluruh kawasan Karst Gombong
(DISPARHUB Kabupaten Kebumen 2004). Meskipun survei yang dilakukan
hanya sebatas identifikasi titik koordinat gua dan letak administratif gua, hasil
survei ini telah berhasil memberikan informasi penting mengenai jumlah dan
sebaran gua-gua karst di kawasan Karst Gombong. Melalui informasi ini,
sebetulnya dapat digali lebih dalam kondisi ekosistem gua-gua di kawasan Karst
Gombong. Dengan demikian, gua-gua yang telah teridentifikasi tersebut dapat
dikelola sesuai dengan potensi yang ada. Namun, sangat disayangkan, antusias
PEMDA Kebumen untuk menggali lebih jauh ekosistem gua-gua di Karst
Gombong sangat rendah. Akibatnya, hasil inventarisasi gua yang telah dilakukan
tujuh tahun lalu hanya berfungsi sebagai data base tanpa diikuti dengan
kebijakan pengelolaannya.
Usulan strategi konservasi ekosistem Karst Gombong
Upaya perlindungan di Karst Gombong dapat dicapai dengan strategi
konservasi. Menurut Alikodra (1988) yang dimaksud dengan konservasi adalah
upaya pengelolaan sumber daya alam yang menjamin: a) perlindungan pada
berlangsungnya proses-proses ekologis dan sistem penyangga kehidupan; b)
pengawetan sumber daya alam dan keanekaragaman sumber plasma nutfah; dan
c) pemanfaatan secara lestari sumber daya alam dan lingkungannya. Konservasi
sumberdaya alam tersebut akan berhasil bila dilakukan atas dasar hasil penelitian
yang akurat. Hasil penelitian ini perlu dijadikan pedoman dalam upaya
konservasi ekosistem gua di kawasan Karst Gombong.
Pelestarian kawasan karst harus bersifat lintas sektoral dan melibatkan
berbagai unsur masyarakat yang terlibat dalam kawasan karst tersebut. Agar dapat
dijadikan pedoman bagi semua unsur yang terlibat dalam pemanfaatan kawasan
Karst Gombong, maka perlu ditetapkan status kawasan Karst Gombong sesuai
dengan hasil penelitian di lapangan dan berpedoman pada peraturan perundang-
undangan yang berlaku di Indonesia.
Hingga saat ini peraturan normatif yang berkaitan langsung dengan
pengelolaan kawasaan karst di indonesia hanya Kepmen ESDM Nomor 1456

110
K/20/MEM/2000 tentang Pedoman Pengelolaan Kawasan Karst. Menurut
Samodra (2006) meskipun masih bersifat sektoral, Keputusan Menteri ini
membuka kesempatan sektor lain untuk menyempurnakannya.
Berdasarkan Kepmen ESDM Nomor 1456 K/20/MEM/2000 pasal 11,
kawasan karst dibagi menjadi : kawasan karst kelas I, kawasan karst kelas II, dan
kawasan karst kelas III. Kawasan kelas I merupakan kawasan yang memiliki
salah satu atau lebih kriteria berikut :
a. Berfungsi sebagai penyimpan air bawah tanah secara tetap (permanen)
dalam bentuk akuifer, sungai bawah tanah, telaga ataupun danau
bawah tanah yang keberadaannya mencukupi fungsi umum hidrologi.
b. Mempunyai gua-gua dan sungai bawah tanah aktif yang kumpulannya
mepunyai jaringan baik tegak ataupun mendatar.
c. Gua-gua mempunyai speleotem aktif dan atau peninggalan sejarah
sehingga berpotensi dikembangkan sebagai objek wisata dan budaya.
d. Mempunyai kandungan flora dan fauna khas yang memenuhi arti
fungsi sosial, ekonomi, serta pengembangan ilmu pengetahuan.
Kawasan karst kelas II merupakan kawasan yang memiliki salah satu atau semua
kriteria berikut :
a. Berfungsi sebagai penimbun air bawah tanah, berupa daerah
tangkapan air hujan yang mempengaruhi naik turunnya muka air
bawah tanah di kawasan karst, sehingga masih memegang fungsi
umum hidrologi
b. Mempunyai jaringan lorong-lorong bawah tanah hasil bentukan
sungai dan gua yang sudah kering, mempunyai speleotem yang sudah
tidak aktif dan menjadi tempat tinggal tetap fauna yang semuanya
memberi nilai dan manfaat ekonomi.
Kawasan karst kelas III merupakan kawasan yang tidak memiliki kriteria
sebagaimana dimaksud dalam kriteria kelas I dan kriteria kelas II.
Sebelum penelitian ini dilakukan, status kawasan Karst Gombong belum
dapat ditetapkan karena belum cukup data yang mendukung penetapan status
kawasan tersebut. Namun, berdasarkan temuan pada penelitian ini, kawasan Karst
Gombong dapat diusulkan sebagai kawasan karst kelas I sesuai dengan Kepmen

111
ESDM Nomor 1456 K/20/MEM/2000 pasal 11. Hal ini karena kawasan Karst
Gombong terbukti merupakan penyimpan air bawah tanah, merupakan ekosistem
unik, habitat satwa khas dan satwa terancam punah, serta berpotensi wisata.
Setelah kawasan karst ditetapkan Kawasan Karst kelas I, gua-gua di
kawasan Karst Gombong harus dimanfaatkan sesuai dengan kondisi fisik dan
status komunitas biota yang terkandung di dalamnya. Matriks pada Tabel 22 dapat
menggambarkan kondisi fisik dan biota dua belas gua yang dikaji dalam
penelitian ini.
Tabel 22 Matriks kondisi fisik dan biota di gua-gua Karst Gombong Komponen Pertimbangan
1
2
3
4
5
6
7
8
9
10 11
12
1. berfungsi penyimpan air bawah tanah
+ + - + + - + + + - - -
2. Dekorasi gua indah /speleotom aktif
+ + - + + - + + + - - -
3. habitat fauna khas/unik
+ + + + + - + + + - - -
4. habitat fauna terancam punah
+ + + + + - + + + - - -
5. terdapat peninggalan budaya
- - - - - - - - - - - -
Keterangan: 1= Gua Macan 2= Gua Celeng 3 = Gua Dempo 4 = Gua Inten 5= Gua Jatijajar 6= Gua Kampil 7 = Gua Kemit 8 = Gua Liyah 9= Gua Petruk 10= Gua Sigong 11= Gua Tratag 12 = Gua Tiktikan
Berdasarkan matriks tersebut terlihat bahwa Gua Macan, Gua Celeng, Gua
Dempo, Gua Inten, Gua Jati jajar, Gua Kemit, Gua Liyah, dan Gua Petruk
berfungsi menyimpan air bawah tanah, memiliki dekorasi indah/speleotom aktif,
habitat fauna khas/ unik, dan habitat fauna terancam punah. Oleh karenanya,
untuk mempertahankan fungsi ekologis gua dan komunitas biota yang
dilindungi, ke-delapan gua tersebut perlu ditetapkan sebagai gua konservasi.
Sebagai gua konservasi, ke-delapan gua tersebut dapat dimanfaatkan sebagai
objek wisata dengan persyaratan khusus tetapi tidak boleh dimanfaatkan untuk
kegiatan pertambangan. Sebaliknya, Gua Kampil, Gua Sigong, Gua Tratag dan
Gua Tiktikan tidak berfungsi menyimpan air tanah, tidak memiliki speleotom

112
aktif, bukan habitat fauna khas/ unik, dan bukan habitat fauna terancam punah.
Oleh karenanya, keempat gua tersebut dapat dimanfaatkan untuk kegiatan
pertambangan dengan syarat tidak merusak struktur gua. Apabila Gua Macan,
Gua Celeng, Gua Dempo, Gua Inten, Gua Jati jajar, Gua Kemit, Gua Liyah, dan
Gua Petruk akan dimanfaatkan sebagai gua wisata alam (ekotourism), bentuk
wisata yang dapat dikembangakan adalah wisata minat khusus yang
pengelolaannya berdasarkan pada prasyarat ekofisiologi yang dibutuhkan
kelelawar sebagaimana telah diketahui dari hasil penelitian ini. Dengan cara
demikian diharapkan jenis-jenis kelelawar yang bersarang di dalamnya dapat
dipertahankan.
Berdasarkan hasil penelitian ini, diketahui bahwa dalam memilih sarang,
terdapat lima faktor fisik yang berpengaruh nyata, yaitu intensitas suara,
kelembaban udara, suhu udara, intensitas cahaya, dan jarak dari pintu gua. Untuk
mempertahankan keberadaan kelelawar, ruang gua yang dihuni kelelawar
tersebut, harus dikelola sesuai dengan persyaratan mikroklimat yang dibutuhkan
kelelawar. Dari hasil penelitian ini dapat diusulkan pemanfaatan ruang gua sesuai
dengan kebutuhan kelelawar tersebut. Usulan pemanfaatan ruang gua yang
digunakan sebagai sarang kelelawar tersebut dapat dilihat pada Tabel 24.
Ekosistem dalam gua tidak dapat dipisahkan degan ekosistem luar gua. Oleh
karena itu, upaya pelestarian kawasan karst perlu dilakukan secara holistik dan
terpadu antara ekosistem dalam gua dan ekosistem luar gua. Untuk maksud
tersebut perlu ditetapkan zonasi kawasan Karst Gombong. Zonasi kawasan karst
ini harus menjadi acuan dalam pemanfaatan dan pengeloaan kawasan Karst
Gombong oleh semua unsur yang terkait.
Tabel 24 Usulan pemanfaatan ruang gua berdasarkan jenis kelelawar yang bersarang dan prasyarat ekofisiologi yang dibutuhkan
Ruang sarang kelelawar
Prasyarat mikroklimat Usulan pengelolaan ruang gua
C.brachyotis C.horsfieldii E.spelaea R.amplexicaudatus
Intensitas suara ≥ 2 db Suhu ≥28.5oC Kelembaban ≤65% Intensitas cahaya ≥ 50 lux
- Boleh dikunjungi manusia. - Pada dinding dan atap gua boleh dilakukan pembangunan untuk menarik wisatawan, dengan syarat pembangunan tersebut

113
Menurut Cughley & Gunn 1995, zonasi dalam kawasan perlindungan
sangat diperlukan agar dalam kawasan tersebut dapat dilakukan kegiatan
pelestarian sekaligus pemanfaatan oleh manusia. Zonasi dalam kawasan Karst
Gombong diusulkan meliputi zona inti/zona perlindungan dan zona
penyangga/zona pemanfaatan tradisional. Zona inti merupakan zona yang
kawasan tersebut dilindungi dan kegiatan manusia dikendalikan secara ketat dan
tidak diperbolehkan adanya kegiatan penambangan. Zona penyangga merupakan
kawasan yang menyangga kelestarian zona inti. Penetapan zona penyangga di
kawasan Karst Gombong ini, selain untuk menjamin kecukupan pakan kelelawar
juga untuk mempertahankan mikroklimat di dalam gua. Karena menurut Samodra
(2006), keadaan di dalam gua sangat ditentukan oleh vegetasi, tanah, dan air di
luar gua. Masyarakat sekitar dapat memenfaatkan zona penyangga dengan
tidak merusak struktur gua. - Pada lorong gua boleh dipasang lampu penerangan.
H.sorenseni
Intensitas suara:0.5 s/d 20 db Suhu ≤ 28.5oC Kelembapan : 65 s/d 75% Intensitas cahaya : 5 s/d 50 lux
- Boleh dikunjungi manusia tetapi jumlah dan kegiatannya diawasi dengan ketat. - Dinding dan atap gua tidak boleh Dilakukan pembangunan - Pada lorong gua tidak boleh dipasang lampu penerangan.
C.plicata H.cf.ater Hipposideros sp
Intensitas suara 0.5 s/d 20 db Suhu≤28.5oC Kelembapan ≥75% Intensitas cahaya ≤5 lux
- Boleh dikunjungi manusia tetapi jumlah dan kegiatannya diawasi dengan ketat
- Tidak boleh ada pembangunan di dalam maupun di luar gua
- Tidak boleh dipasang lampu penerangan
M.schreibersii R.affinis
Intensitas suara ≤0.5 db Suhu ≥28.5oC Kelembapan :65 s/d 75% Intensitas cahaya : 5 s/d 50 lux
- Tidak boleh dikunjungi manusia - Tidak boleh dilakukan pembangunan pada dinding dan atap gua - Tidak boleh dipasang lampu penerangan
H.ater R.borneensis
Intensitas suara ≤0.5 db Suhu ≤28oC Kelembapan ≥75% Intensitas cahaya ≤5 lux
- Tidak boleh dikunjungi manusia - Tidak boleh ada pembangunan baik
di dalam maupun di luar gua - Tidak boleh dipasang lampu
penerangan

114
kegiatan ekonomi tradisional seperti penggembalaan hewan ternak atau
mengambil hasil hutan tanpa menebang.
Berdasarkan hasil penelitian ini, diketahui bahwa delapan gua di kawasan
Karst Gombong merupakan habitat sarang kelelawar terancam punah. Gua-gua
tersebut adalah Gua Dempo, Gua Inten, Gua Jatijajar, Gua Petruk, Gua Kemit,
Gua Liyah, Gua Macan, dan Gua Celeng. Selain itu, ke-delapan gua tersebut juga
memiliki speleotem aktif dan berfungsi sebagai penyimpan air. Karena itu, ke-
delapan gua tersebut perlu diusulkan sebagai zona inti.
Hasil penelitian ini membuktikan bahwa makanan kelelawar
Microchiroptera yang bersarang di gua-gua Karst Gombong adalah 29 famili
serangga yang termasuk dalam 10 ordo. Serangga-serangga tersebut hidup di
persawahan, semak belukar, dan hutan karst yang berada di sekitar gua.
Kelelawar Megachiroptera yang ditemukan dalam penelitian ini terbukti
mengunjungi 33 genus tumbuhan yang termasuk dalam sembilan famili untuk
memakan polennya. Tumbuhan tersebut adalah tumbuhan yang hidup di hutan
karst dan perkebunan yang berada di sekitar gua. Oleh karena itu, untuk
mendukung kehidupan kelelawar, kawasan sekitar gua yang merupakan tempat
pencarian makan kelelawar perlu ditetapkan sebagai zona penyangga/ zona
pemanfaatan tradisonal.
Penelitian Law & Chidel (2004) membuktikan bahwa Kerivoula papuensis
(Microchiroptera) bersarang pada jarak maksimum 2.1 km dari tempat pencarian
makannya di hutan hujan di New South Wales. Feelers & Pierson (2002)
membuktikan kelelawar Corynorhinus townsendii (Microchiroptera) mencari
makan dengan jarak ± 10.2 km dari sarangnya di karst California. Zahn et al. 2005
membuktikan Myotis myiotis (Microchiroptera) mencari makan dengan jarak 2.5
sampai dengan 8.9 km dari sarangnya. Dalam penelitian ini, tidak diteliti jarak
pencarian makan kelelawar yang bersarang di gua-gua Karst Gombong. Namun,
berdasarkan hasil penelitian terdahulu, hutan dan semak dengan jarak 5 km s/d 8
km di sekitar zona inti perlu diusulkan sebagai zona penyangga. Usulan zonasi
kawasan Karst Gombong dapat dilihat pada peta satelit Lansat yang tersaji pada
Gambar 26. Apabila semua pihak yang terkait dalam pemanfaatan kawasan Karst
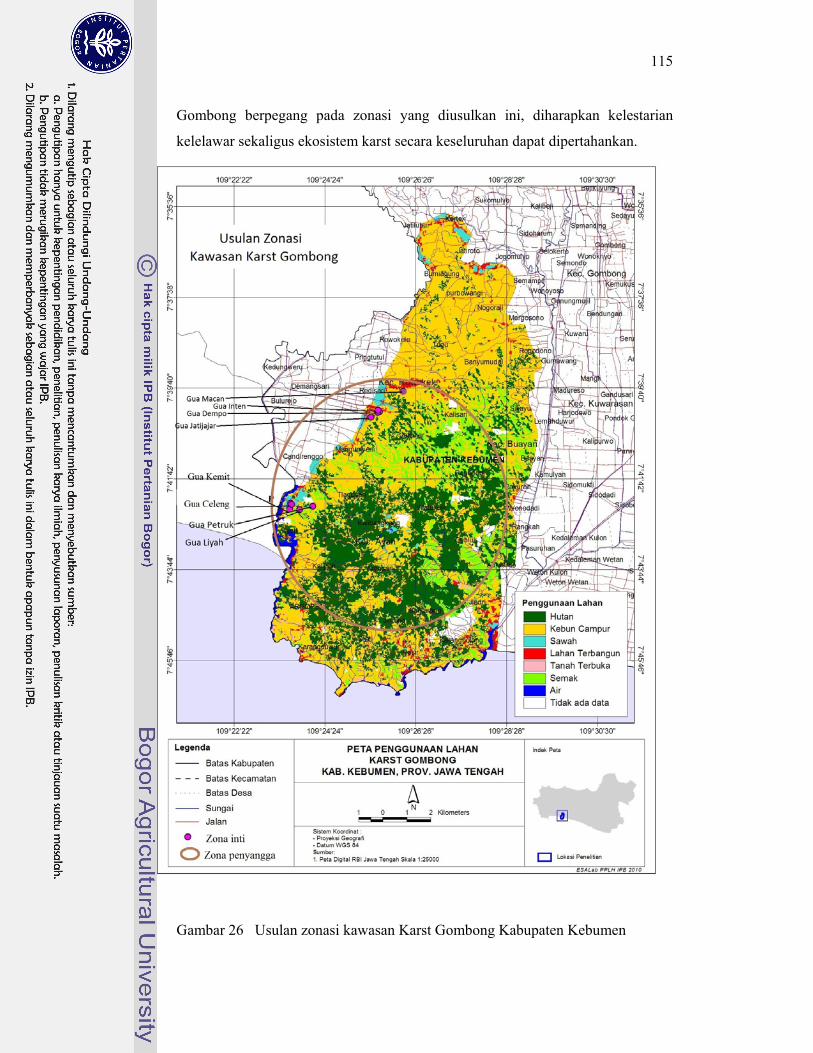
115
Gombong berpegang pada zonasi yang diusulkan ini, diharapkan kelestarian
kelelawar sekaligus ekosistem karst secara keseluruhan dapat dipertahankan.
Gambar 26 Usulan zonasi kawasan Karst Gombong Kabupaten Kebumen

116
KESIMPULAN
Hasil penelitian ini dapat disimpulkan :
1. Biodiversitas kelelawar di kawasan Karst Gombong tinggi, dan terdiri dari
kelelawar yang termasuk dalam Red List IUCN Versi 3.1 sehingga perlu
dilakukan upaya perlindungan
2. Gua-gua yang ada di kawasan Karst Gombong memiliki kondisi
mikroklimat berbeda, sehingga pengelolaannya harus disesuaikan dengan
kebutuhan kelelawar.
3. Asosiasi bersarang (roosting) menunjukkan gambaran asosiasi yang tidak
menyebabkan kompetisi negatif
4. Strategi adaptasi kelelawar untuk bertahan hidup di gua dengan kondisi
dingin, lembab dan tinggi kadar amonia adalah dengan alveolus
menyempit, jumlah eritrosit tinggi dan kadar hemoglobin tinggi
5. Karst Gombong perlu diusulkan sebagai Kawasan karst kelas I sesuai
dengan Kepmen ESDM Nomor 1456 K/20/MEM/2000. Gua Macan, Gua
Celeng, Gua Dempo, Gua Inten, Gua Jati jajar, Gua Kemit, Gua Liyah,
dan Gua Petruk perlu ditetapkan sebagai zona inti, mengingat gua-gua
tersebut dihuni oleh jenis-jenis kelelawar yang perlu dilidungi menurut
Red List IUCN versi 3.1, dan kawasan hutan di sekitar gua-gua tersebut
perlu ditetapkan sebagai zona penyangga mengingat kawasan tersebut
merupakan tempat pencarian makan kelelawar.
SARAN
1. Hasil penelitian ini perlu dijadikan pedoman dalam pengelolaan kawasan
Karst Gombong oleh semua fihak yang terkait.
2. Diperlukan penelitian lebih lanjut mengenai habitat pencarian pakan
kelelawar gua, guna menentukan zona penyangga yang lebih tepat di
kawasan Karst Gombong
3. Hasil penelitian ini dapat dipergunakan sebagai percontohan dalam
pengelolaan kawasan karst lain yang ada di Indonesia.

117
DAFTAR PUSTAKA
Agosta SJ. 2002. Habitat use, diet and roost selection by the big brown bat
(Eptesicus fuscus) in North America: a case for conserving an abundant spesies. Mam Review. 32:179-198.
Aguirre LF, Herrel A, Van Damme R & Matthysen E. 2002. Ecomorphological analysis of tropic niche partioning in a tropical savana bat community. Proceeding of the Royal Society of London. 269:1271-1278.
Aguirre LF, Herrel A, Van Damme R & Matthysen E. 2003. The implications of food hardness for diet in bats. Functional Ecology. 17: 201-212.
Alikodra HS. 1988. Konservasi Sumber Daya Alam. Diktat Kuliah. Fakultas Kehutanan IPB. Bogor.
Alikodra HS. 2010. Teknik Pengelolaan Satwa Liar Dalam Rangka Mempertahankan Keanekaragaman Hayati Indonesia. IPB Press. Bogor.
Altringham JD. 1996. BATS. Biologi and Behaviour. Oxford University Press. New York.
Apriandi J, Kartono AP & I Maryanto. 2008. Keanekaragaman dan kekerabatan jenis kelelawar berdasarkan kondisi mikroklimat tempat bertenger pada beberapa goa di kawasan Gua Gudawang. J. Biol. Indo. 5(2): 121-134
Arita HT. 1996. The conservation of cave-roosting bats in Yacatan, Mexico. Biological Conservation. 12: 188-194.
[BAPPENAS] Badan Perencana Pembangunan Nasional. 2003. Strategi dan Rencana Aksi Keanekaragaman Hayati Indonesia 2003-2010. LIPI Press. Bogor.
Barth FG. 1991. Insect and Flowers. The Biology of a Partnership. Princeton University Press. New Jersey.
Bartonicka T & Zukal J. 2003. Flight activity and habitat use of four bat spesies in a small town revealed bats revisited: feedback controls both a decrease and an increase in call frequency . Mamm.Biol. 5: 282-290.
Baudinette RV, Wlls RT, Sanderson KJ & Clark B. 1994. Microclimat conditions in maternity caves of the Bent-wing bat Miniopterus schreibersii: an attempt restoration of a former maternity site. Wildl. Res. 21: 607-619
Baudinette RV, Churchill SK, Christian KA, Nelson JE & Hudson PJ. 2000. Energy, water balance and the roost microenvironment in three Australian cave-dwelling bats (Microchiroptera). J. Comp. Phy. Biol. 170: 439-446
Borror DJ, Triplehorn & Johnson NF. 1996. An Introduction to The Study of Insects. Sixth edition. Saunders College Publishing.
Campbell LA, Hallet JG & O’Connel MA. 1996. Conservation of bats in managed forest: use of roost by brown bats. Eptesicus fuscus, conform to the fission- fusion model. Animal Behaviour. 68: 495-505.
Campbell NA, Reece JB & Mitchell LG. 2002. Biologi. Edisi kelima. Jilid 1. Penerjemah: Rahayu Lestari et al. Terjemahan dari Biology. Erlangga. Jakarta.
Castillo AE, Meneses GC, Davilla-Montes MJ, Anaya MM & Leon PR. 2009. Seasonal distribution and circadian activity in the troglophile long-footed robber frog Eleutherodactylus longipes (Anura: Brachycephalidae) at Los

118
Riscos Cave, Queretaro, Mexico: Field and laboratory studies. J. Cave and Karst Studies 71(1):121-128
Caughley G & Gunn A. 1995. Conservation Biology in Theory and Practice. Blackwell Science. Canada.
Chairunnisa. 1997. Studi komparatif morfologi saluran pencernaan kelelawar pemakan serangga (Scotophilus kuhli) dan kelelawar pemakan buah (Cynopterus brachyotis). Tesis. Progam Studi Biologi. Sekolah Pasca Sarjana IPB. Bogor.
Corbet GB & JE Hill. 1992. The mammal of the Indomalayan region. Asystematic review. Natural history museum publications. Oxford University Press.
Cox GW. 2002. General Ecology. Laboratory Manual. Mc Gaw-Hill Higher Education. New York.
Cunningham JG. 2002. Textbook of Veterinary Physiologi. Edisi ke-3. WB Saunders company. Philadelphia.
Departemen Kehutanan Republik Indonesia.1987. Pola pengelolaan fungsi gua berdasarkan hasil identifikasi fungsi gua di Provinsi Jawa Tengah. Laporan. Proyek Pembinaan Kelestarisn Sumber Daya Alam Hayati Pusat. DIRJEN PHPA. Bogor.
Departemen Kesehatan Republik Indonesia. 2001. Pedoman Tata Laksana Lingkungan Kerja . DEPKES RI. Jakarta.
[DISPARHUB] Dinas Pariwisata dan Perhubungan Kabupaten Kebumen (2004). Laporan hasil survei goa di di wilayah Kabupaten Kebumen. Dinas
Pariwisata dan Perhubungan Kebumen. Doyle ME. 1979. Factor affecting distribution of fauna in Gua Pondok. The
Malayan Nature J. 25: 21-26. Dunn FL. 1978. Gua Anak Takun ecological observation. The Malayan Nature J.
19(1): 75-87 Duran AR & Centano JAS. (2002) Temperature selection by tropical bats
roosting in caves. J. Thermal Biol. 28: 465-468. Eberhard SM. 2006. Ecology and hidrology of a threatened gound water
dependent ecosystem: the jewel cave karst system in Western Australia. Helictite. J. Austrlia Speleological Research. 39(1):21-22.
Epsinasa L & Vuong NH. 2008. A new spesies of cave adapted Nicoletiid (Zygentoma: Insecta) from Sistema Huautla, Oaxaca, Mexico: The tenth deepest cave in the world. J. cave and karst studies 70 (2): 289-298.
Erdtman G. 1952. Polen Morphology and Taxonomy : An Introduction to the Study Polen and Spores. Copenhagen. Munksgard.
ESALab PPLH IPB 2010. Kawasan Karst Gombong Kabupaten Kebumen (Peta citra satelit LANDSAT).PPLH IPB. Bogor. 3 lembar.
Fatmawati F. 2007. Gambaran hematology anjing pelacak operasional ras dobermen di Subdit Satwa POLRI Depok. Skripsi. Fakultas Kedokteran Hewan IPB. Bogor.
Feeler GM & Pierson ED. 2002. Habitat use and foraging behavior of Townsend’s big-eared bat (Corynorhinus townsendii) in coastal California. J.Mamm. 83 (1) : 167-177.
[FINSPAC] Federation of Indonesian Speleological Activities. 1993. Java caves. Report of a visit to Indonesian by Australian and British cavers. Finspac. Bogor.

119
Frandson RD. 1992. Anatomi dan Fisiologi Ternak. Edisi IV. Penerjemah: B.Srigandono dan K Praseno. Gadjah Mada University Press. Yogyakarta.
Furman A & Ozgul A. 2002. Distribution of cave dwelling bats and conservation status of undergound habitats in the Istanbul area. Ecological Research. 17: 69-77.
Ganong WF. 2001. Fisiologi Kedokteran. Edisi ke-3. Penerbit buku kedokteran EGC. Jakarta.
Gasperszs V. 1995. Teknik Analisis dalam Penelitian Percobaan. Jilid 2. Tarsito. Bandung.
Gimes KG. 2001. Syngenetic karst in Australlia: a review. Helictite J. Austral Spel. Res 39 (2): 27-38.
Guyton AC. 1995. Fisiologi Manusia dan Mekanisme Penyakit. Edisi ke- III. Alih bahasa: Petrus Andrianto. Penerbit Buku Kedokteran ECG. Jakarta.
Hamilton E & Smith. 2006. Thinking about karst and world heritage. Helictite J. Australia speleological research. 39 (2): 51-54.
HanGuan AK, Esa Y, Sallehin AA, Ryan JR, Julaihi AM, Kumaran JV & Abdullah MT. 2006. Roosting ecology and social organization of geater horsehoe bat (Rhinolophus ferrumequinum). Behav. Ecol Sociobiol. 51: 510-518.
Heinrich U, Muhle H & Takenaka S. 1986. Chronic effects on the respiratory tract of hamsters, mice and rats after long-term inhalation of high-concentrations of filtered and unfiltered diesel-engine emissions. Anatomy and Physiologi. 6: 383-395.
[HIKESPI] Himpunan Kegiatan Speleologi Indonesia. 2004. Kursus tingkat dasar, lanjutan dan diklat speleologi caving, kumpulan makalah dan peraga kursus. HIKESPI. Bogor.
Hodgkison R, Balding ST, Zubaid A & Kunz TH. 2004. Habitat structure, wing morphology, and the horseshoe bats revisited: feedback controls both a decrease and an increase in call insectivorous bats. J.Applied Ecology. 39: 605-617.
Hutabarat IO.2002. Analisis dampak gas amonia dan klorin pada faal paru pekerja pabrik sarung tangan “X” di Medan. Laporan penelitian. Lembaga Penelitian dan Pengabdian Masyarakat USU. Medan.
[IUCN] The International Union for Concervation of Nature Spesies Survival Commission. 2001. IUCN Red List Catagories and Criteria. Version 3.1. IUCN. Switzerland.
John, Mackinnon K, Child G & Thorsell JM. 1990. Pengelolaan Kawasan yang Dilindungi di Daerah Tropika. Penerjemah: Harry harsono Amir. Gadjah Mada University Press. Yogyakarta.
Keller V & Bollmann K. 2004. From red list to spesies of conservation concern. Conservation biology. 18(6): 1636-1644.
Keputusan Menteri Energi dan Sumberdaya Mineral no.1456 Tahun 2000 tentang Pengelolaan Kawasan Karst.
[KLH] Kementrian Negara Lingkungan Hidup. 2003. Himpunan Peraturan Perundang Undangan di Bidang Pengelolaan Lingkungan Hidup dan Pengendalian Dampak Lingkungan Era Otonomi Daerah Buku II. Jakarta.

120
Koneri, R. 2007. Bioekologi dan konservasi kumbang lucanid (Coleoptera: Lucanidae) di hutan gunung salak, Jawa Barat. Disertasi. Sekolah Pasca Sarjana IPB. Bogor.
Ko RKT. 1999. Dampak Penambangan pada Ekosistem Karst. Himpunan Kegiatan Speleologi Indonesia. Bogor.
Ko RKT. 2004. Pengertian kawasan karst sebagai suatu sistem energi. Makalah kunci pada lokakarya nasional karst di banda aceh Juli 2004. Himpunan Kegiatan Speleologi Indonesa. Bogor.
Kunz TH & Nagy KA. 1988. Methods of energy budget analysis. Dalam : Ecological and Behavioral Methods for The Study of Bats. Kunz TH, editor. Smithonsonian Institution Press. Washington D.C. London.
Kunz TH & Pierson ED. 1994. Bats of the world. An Introduction. Dalam : Walker’s Bats of the World.The John Hopkins University press. Baltimore and London
Law B & Chidel M. 2002. Tracks and riparian zones facilitate the use of Australian regrowth forest by acquisition in echolocating bats. Trends in Ecol. and Ev. 18 (8): 386-394.
Leps J & Smilauer. 1999. Multivariate Analysis of Ecolocical Data. Faculty of Biological Science University of South Bohemia.
Ludwig JA & Reynolds JE. 1988. Statistical Ecology. A Primer on Methodts and Computing. John wiley London.
Maguran AE. 1988. Ecological diversity and its measurement. Croom Helm Limited. London.
Maguran AE. 2004. Measuring biological diversity. Malden: Blackwell Publishing.
Mancina CA, Balseiro F & Herrera LG. 2005. Polen digestion by nectarivorous and frugivorous Antillean bats. Mamm.Bol. 70 (5): 282-290
Maryanto I & Maharadatunkamsi. 1991. Kecenderungan jenis jenis kelelawar dalam memilih tempat bertengger pada beberapa gua di Kabupaten Sumbawa. Media Konservasi. 3:29-34
McCure HE.1985. Microcosm of batu caves. Malay Nat. J. 19(1): 65-74. Metzner W, S Zhang & Smotherman M. 2002. Doppler-shift compensation
behavior in horseshoe bats revisited: feedback controls both a decrease and an increase in call frequency . J. Exp Biol. 205: 1607–1616.
Meyer CF, Schwarz CJ & Fahr J. 2004. Activity pattern and habitat preference of insectivorous bats in a Microchiroptera based on stomach contents. Mamm. Biol. 70(5): 312-316
Nugraha KN. 2007. Gambaran darah anjing kampung jantan (Canis familiaris) umur 3 sampai 7 bulan. Skripsi. Fakultas Kedokteran Hewan IPB.Bogor.
Nowak RM.1994. Walker’s bats of the word. The Johns Hopkins University Press. Baltimore and London.
Parsons KN, Jones G, Watts ID & Geenaway G. 2002. Swarming of bats at Under- gound sites in Britain-implications for conservation. Biol.Cons. 111:63-70.
Plopper CG & Adams DR.1993. Respiratory System Dalam: Textbook of veterinary histology. Edisi IV. Dellmann HD. Editor. A Wavarley Company. Tokyo.

121
Pujirianti I. 2006. Keanekaragaman jenis dan pola penggunaan ruang bertengger kelelawar di beberapa goa di Taman nasional alas Purwo Jawa Timur. Skripsi. Depertemen Konservasi Sumberdaya Hutan dan Ekowisata Fakultas Kehutanan IPB Bogor.
Pusat Survei Geologi Badan Geologi Departemen Energi dan Sumber Daya Mineral. 2006. Panduan Geowisata Daerah Kebumen dan Sekitarnya. Departemen ESDM. Jakarta
Rachmadi. 2003. Keanekaragaman Arthropoda di gua Ngerong, Tuban, Jawa Timur, Zoo Indo. 29: 19-26.
Razakarivony V, Rajemison B & Goodman SM. 2005. The diet of Malagsy Microchiroptera based on stomach contents. Mamm. Biol. 70(5): 312-316.
Riedesel ML. 1977. Blood physiology. Dalam: Biology of Bats. Volume III. William A. Wimsatt, editor. Academic press. New York.
Riswan R, Noerdjito M & Rachman I. 2006 Vegetasi hutan karst: Kasus kawasan Gombong Selatan-Ayah Kebumen. Dalam : Manajemen Bioregional : Karst, masalah dan pemecahannya. Dilengkapi kasus Jabodetabek. Maryanto, Noerdjito M dan Ubaidillah R, editor. Bogor: PUSLIT Biologi. LIPI..
Roemantyo & Noerdjito M. 2006. Vegatasi hutan karst: Kasus kawasan Gombong Selatan, Ayah, Kebumen, Jawa Tengah. Dalam : Manajemen Bioregional : Karst, masalah dan pemecahannya. Dilengkapi kasus Jabodetabek. Maryanto, Noerdjito M dan Ubaidillah R, editor. Bogor: PUSLIT Biologi. LIPI.
Rossiter SJ, Jones G, Ransome RD & Barratt EM. 2002. Related structure and kin biased foraging in Nyctalus noctula. J. Experimental Biology. 210: 3607-3615.
Ruczynski I, Kalko EKV &Siemers BM. 2007. The sensory basis of roost finding in a forest bat. Mam Biol. 26: 162-163
Russo D, Cistrone L, Jones G & Mazzoleni S. 2003. Roost Selection by Barbastelle Bats in beech Woodland of Central Italy. J. Biologycal Conservation. 117: 73-81.
Safi K & Kerth G. 2004. A comparative analysis of specialization and extinction risk in temperate-zone bats. Conservation Biology. 18(5): 262-269.
Samodra H. 2006. Hubungan antara struktur geologi dengan pembentukan sistem pergoaan : Studi kasus di segmen karst Cigudeg. Dalam : Manajemen Bioregional : Karst, masalah dan pemecahannya. Dilengkapi kasus Jabodetabek. Maryanto I, Noerdjito M dan Ubaidillah R, editor. Bogor: PUSLIT Biologi. LIPI.
Saroni. 2005. Penyusunan metode pendugaan populasi kelelawar penghuni goa: Studi kasus di Kawasan Karst Sangkulirang Mangkaliat Kabupaten Kutai Timur. Skripsi. Depertemen Konservasi Sumberdaya Hutan dan Ekowisata Fakultas Kehutanan IPB Bogor.
Sastradipradja 1989. Penuntun Praktikum Fisiologi Veteriner. Depdikbud. Dirjen. Dikti. PAU Ilmu hayat.IPB.
Schnitzler HU, Moss CF & Denzinger A. 2003. From spatial orientation to food acquisition in echolocating bats. Trends in ecology and evolution. 18 (8): 386-394.

122
Seckerdieck A, Walther B & Halle S. 2005. Alternative use of two different roost types by a maternity colony of the lesser horseshoe bat (Rhinolophus hipposideros). Mam. Biol. 8: 216-224.
Setiadi H. 2000. Morfologi saluran pernapasan kelelawar pemakan serangga (Scotophilus kuhlii, Leach 1821) dengan tinjauan khusus pada trakhea dan paru-paru. Skripsi. Fakultas Kedokteran Hewan IPB. Bogor.
Sevcik M. 2003. Does wing morphology reflect different foraging strategies in sibling bat spesies sibling bat spesies zone bats: Plecotus auritus and P austriacus ?. Fol. Zool. 52: 672-679
Sinaga MH, Achmadi AS & Maryanto I. 2006. Peran kelelawar gua dalam keseimbangan ekosistem. Dalam : Manajemen Bioregional : Karst, maslah dan pemecahannya. Dilengkapi kasus Jabodetabek. Maryanto, Noerdjito M dan Ubaidillah R, editor. Bogor: PUSLIT Biologi. LIPI.
Sobotta & Hammersen F.1993. Histologi. Atlas Berwarna Anatomi Mikroskopik. Edisi III. Penerjemah: Petrus Andrianto. Penerbit buku kedokteran EGK. Jakarta.
Soegiharto S & AP Kartono. 2009. Karakteristik tipe pakan kelelawar pemakan buah dan nektar di daerah perkotaan: studi kasus di Kebun Raya Bogor. J. Biol Indo. 6 (1): 119-130.
Soegiharto S, Kartono AP & Maryanto I. 2010 Pengelompokkan kelelawar pemakan buah dan nektar berdasarkan karakteristik jenis pakan polen di Kebun Raya Bogor. J. Biol. Indon. 6 (2): 225-235.
Sridhar KR, Ashwini KW, Seena S & Sreepada KS. 2006. Manure qualilities of guano of incectivorous cave bat Hippsideros speoris. Tropical and subtropical agoecosystems. 6: 103-110.
Storzt JF, Bhat HB & Kunz TH. 2000. Social structure of polygynous tent-making bat, Cyanopterus sphinx (Megachiroptera) J. Zool. 251:151-165.
Subterra. 2004. Grade Pemetaan Goa. http://www.subterra.or.id Diunduh : 14 Maret 2009.
Surono. 2006. Petrologi batuan karbonat pembentuk karst. Dalam : Manajemen Bioregional : Karst, masalah dan pemecahannya. Dilengkapi kasus Jabodetabek. Maryanto, Noerdjito M dan Ubaidillah R. Editor. Bogor: PUSLIT Biologi. LIPI.
Suyanto, A. 2001. Kelelawar di Indonesia. Bogor: Pusat Penelitian dan Pengembangan Biologi. LIPI
Suyanto A, Yoneda M, Maryanto I, Maharadatunkamsi & Sugardjito J. 1998. Cheklist of the mammals of Indonesia. LIPI-JICA. Bogor.
Syahid A. 2009. Transformasi Data. http://abdulsyahid forum.blogspot.com /2009/04/ transformasi-data.html. Diunduh 20 Juni 2009.
Tan KH, Zuabid A & Kunz TH. 1998. Food habits of Cynopterus brachyotis (Muller) (Chiroptera: Pteropodidae) in Peninsular Malaysia. J. Trop Ecol. 14: 299-307.
Twente Jr JW. 2004. Some aspects of habitat selection and other behavior of cavern dwelling bats. Ecology. 36(4): 706-732.
Ulrich HS, Cynthia FM & Annette D. 2003. From spatial orientation to food acquisition in echolocating bats. Trend in Ecol and Ev. 18 (8): 386-394

123
Van Steenis CGGJ. 2006. Flora. Untuk sekolah di Indonesia. Terjemahan : Moeso S, Soenarto H, Soeryo SA, Wibisono dan Margono P. PT Pranya Paramita. Jakarta.
Verboom B, Boonman AM & Limpens HJGA. 1999. Acoustic perception of landscape elements by the pond bat ( Myotis dasycneme). J. Zoology. 248 : 59-66
Vermeulen J & Whitten T. 1999. Biodiversity and Cultural Property in The Management of Limestone Resources, Lesson from East Asia. The World Bank, Washington DC.
Watt A. 1982. Illustrated Dictionary of Geology. Libraire du lban, Longman York.
Whitaker Jr JO. 1988. Food Habits Analysis of Insectivorous Bats. Thomas H. Kunz, editor. Ecologycal and Behavioral Methods for The Study of Bats. Smithsonian Institution. Uniteted States of Amerika.
Whitten T, Soeriaatmadja RE & Suraya AA. 1999. Ekologi Jawa dan Bali. Seri Ekologi Indonesia. Jilid II. Kartikasari SN, editor. Alih bahasa : SN Kartikasari, TB Utami & A Widiantoro. Prenhallindo. Jakarta.
Wijayanti. 2001. Komunitas fauna Gua Petruk dan Gua Jatijajar Kabupaten kebumen Jawa Tengah. Tesis. Progam Pasca Sarjana Universitas Indonesia. Jakarta.
Willis CKR & Brigham. M. 2004. Roost switching, roost sharing and social cohesion : Forest-dwelling big brown bats, Eptesicus fuscus, conform to the fission-fusion model. Animal Behavior 68: 495-505.
Winkelmann JR, Bonaccorso FJ & Strickler TL. 2000. Home range of southern blossom bat, Syconycteris australis in Papua New Guinea. Tropical Biology. 66: 126-132.
Wirawan R. 2005. Pengelolaan kawasan karst (Studi kasus kawasan karst Gua Pawon Desa Gunungmasigit Kecamatan Cipatat Kabupaten Bandung Jawa Barat). Tesis. Progam Studi Ilmu Lingkungan Progam Pascasarjana Universitas Indonesia. Jakarta.
Yoder JA, Joshua B, Benoit, Crhistensen BS & Hobbs HH. 2009. Entomopathogenic fungi carried by the cave orb weaver spider, meta ovalis (Araneae, Tetragnathidae) with implications for mycoflora transfer to cave crickets. J. cave and karst studies 70 (2): 786-792.
Yunqiu X, Cheng , Yong L & Zhenping D. 2006. Environmental reconstruction of karst using a honey suckle spesies widely used in traditional Chinese medicine. Helictite J. Australia speleological research. 39 (2): 47-50.
Zahn A & Hager I. 2005. A cave dwelling colony of Myotis daubentonii in Bavaria, Germani. J. Mam. Biol. 70 : 242-165.
Zhang L, Jones G, Rossiter S, Ades G, Liang B & Zhang S. 2005. Diet of flat headed bats, Tylonycters zone bats. Cons. Biol.18 (5) 482-489.
Zukal J, Berkova H & Rehak Z. 2005. Activity shelter selection by Myotis myotis and Rhinolophus hipposideros hibernating in the katerinska cave. J. Mam Biol 70: 271-281.
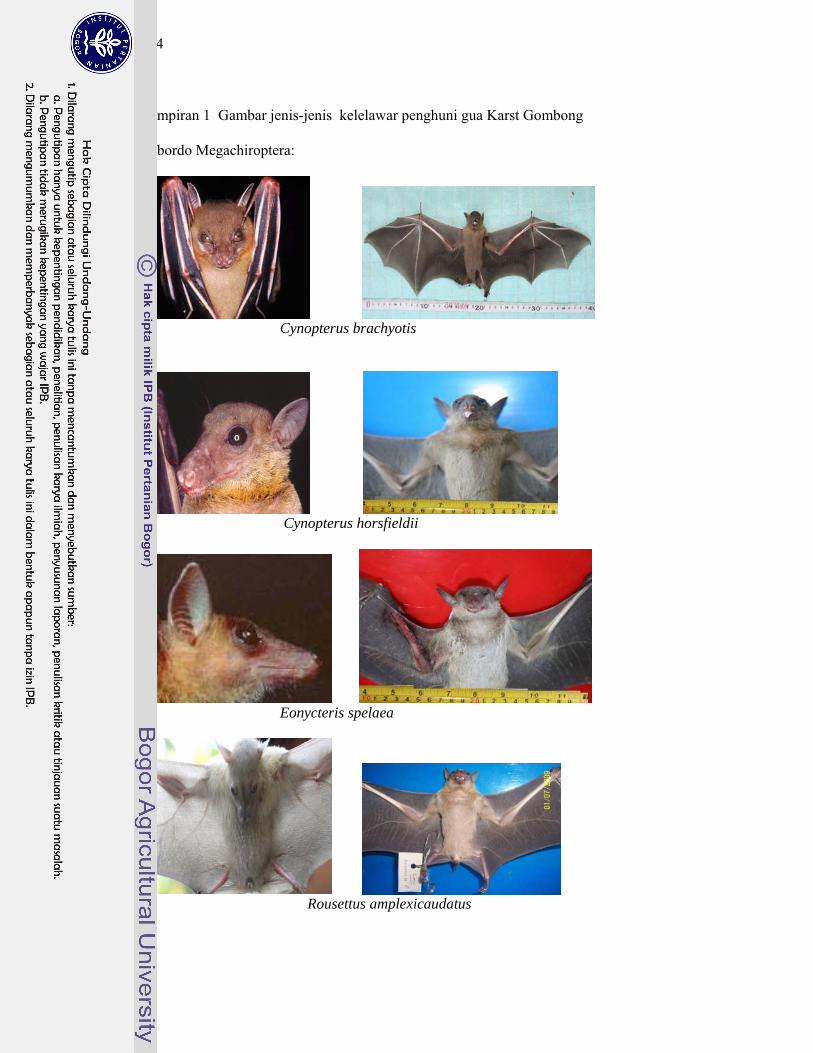
124
Lampiran 1 Gambar jenis-jenis kelelawar penghuni gua Karst Gombong Subordo Megachiroptera:
Cynopterus brachyotis
Cynopterus horsfieldii
Eonycteris spelaea
Rousettus amplexicaudatus

125
Subordo Microchiroptera:
Hipposideros ater
Hipposideros cf. ater
Hipposideros bicolor
Hipposideros sorenseni
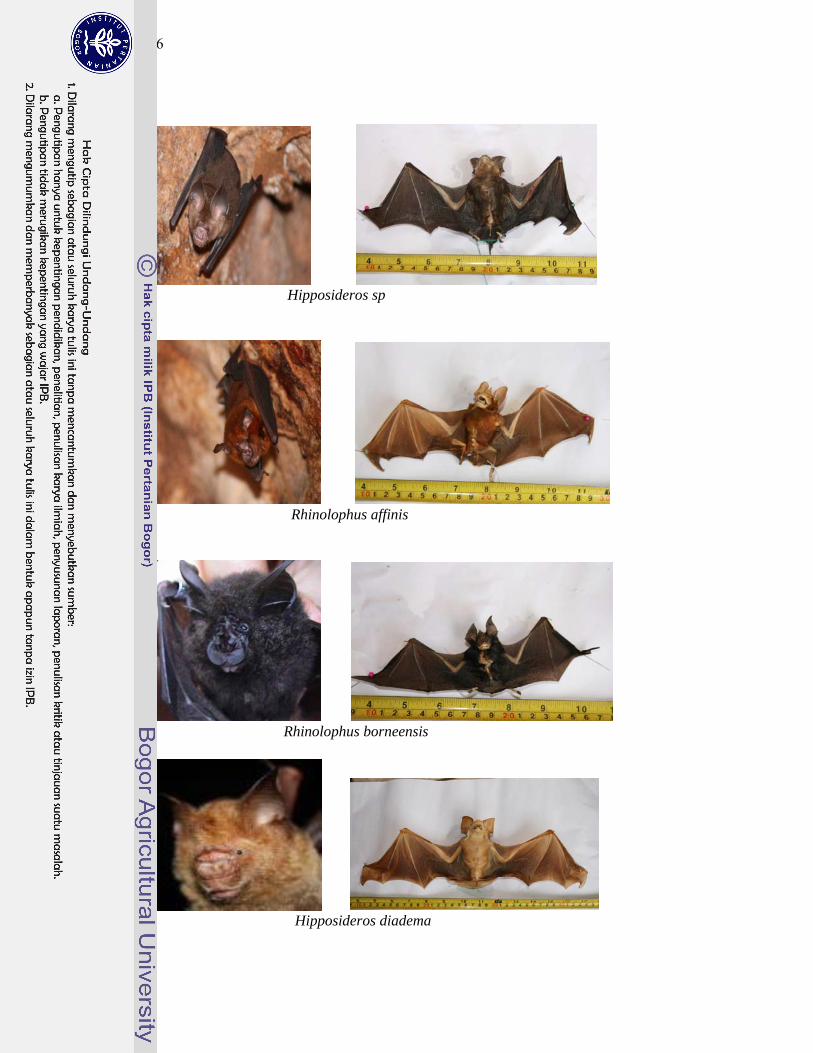
126
Hipposideros sp
Rhinolophus affinis
Rhinolophus borneensis
Hipposideros diadema

127
Miniopterus schreibersii
Chaerophon plicata
Minioptrus australis

128
Lampiran 2 Peta sebaran sarang kelelawar di gua-gua Karst Gombong

129

130

131
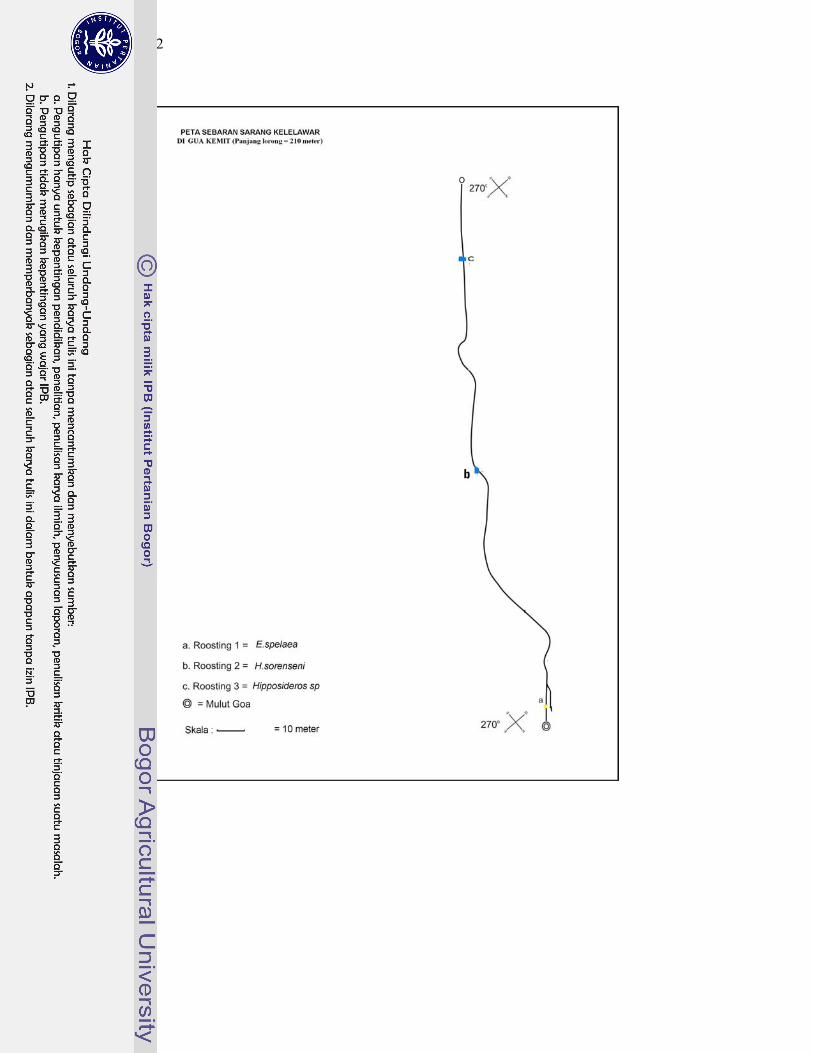
132

133

134

135

136

137

138

139
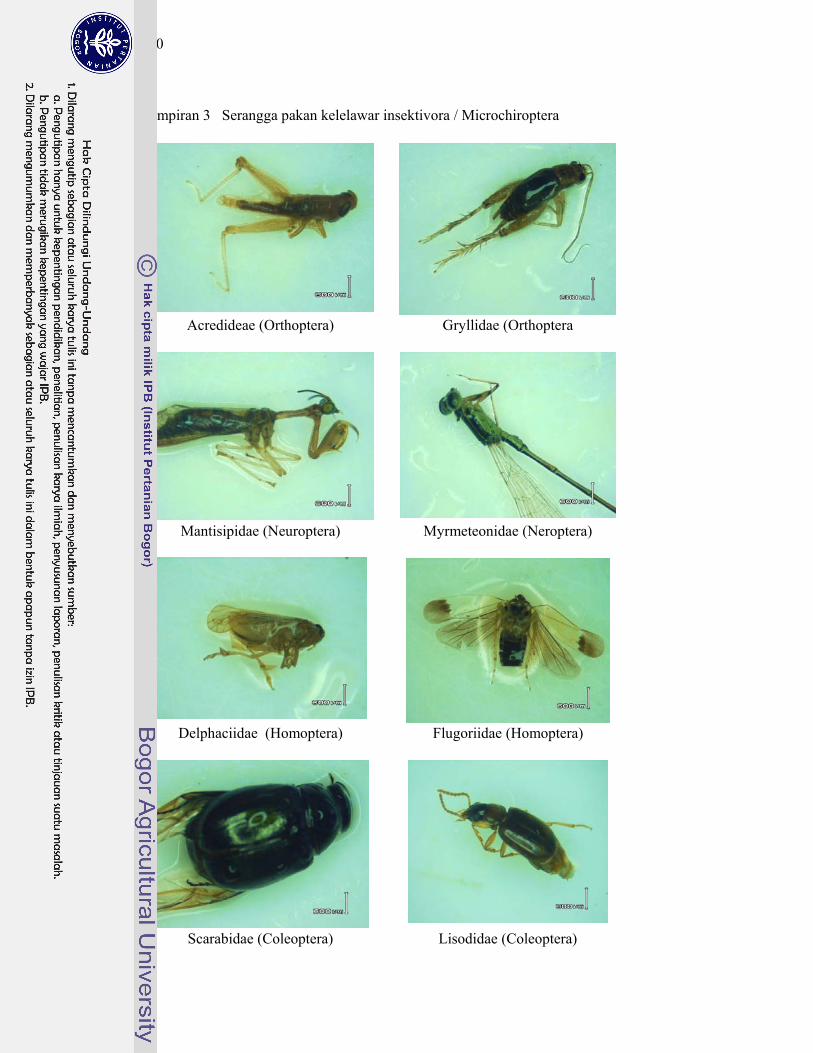
140
Lampiran 3 Serangga pakan kelelawar insektivora / Microchiroptera
Acredideae (Orthoptera) Gryllidae (Orthoptera
Mantisipidae (Neuroptera)
Myrmeteonidae (Neroptera)
Delphaciidae (Homoptera) Flugoriidae (Homoptera)
Scarabidae (Coleoptera) Lisodidae (Coleoptera)

141
Gryllothalpidaea (Orthoptera) Staphilinidae (Coleoptera)
Tipulidae (Diptera) Hymenoptera
Miridae (Hemiptera) Formicidae ( Hymenoptera)
Coleoptera (Lygaeidae) Eurymiideae (Hemiptera)
142
Coleoptera Homoptera
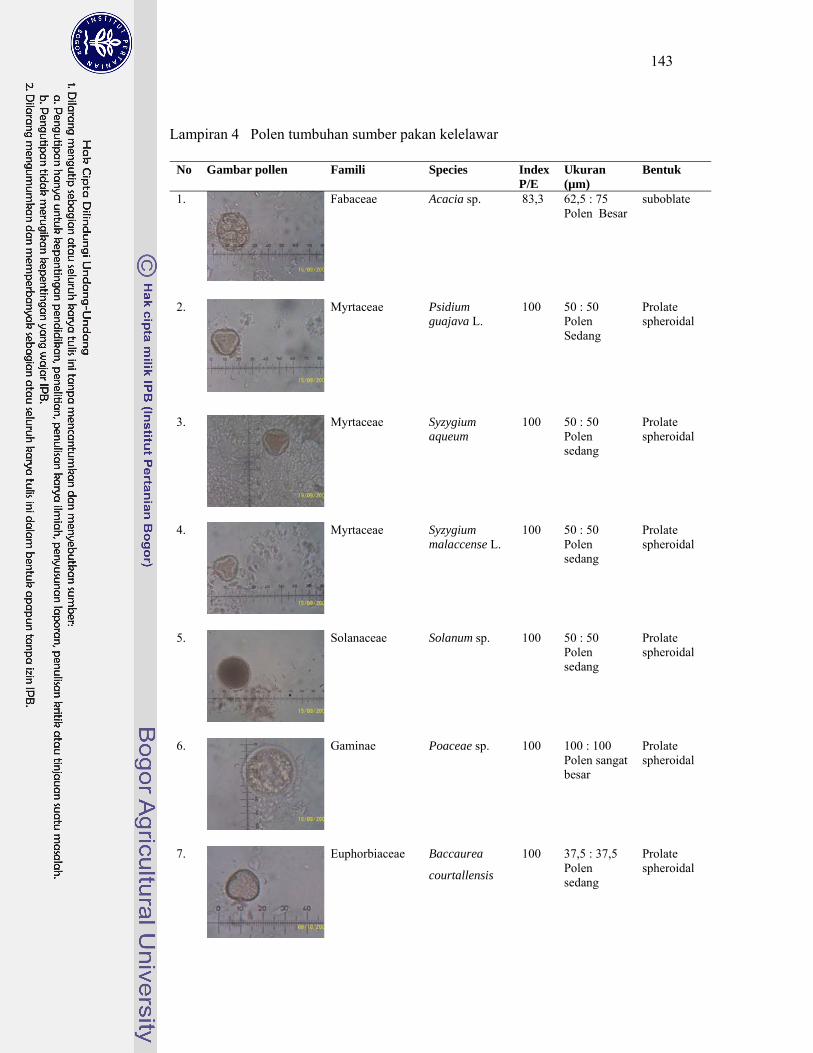
143
Lampiran 4 Polen tumbuhan sumber pakan kelelawar
No Gambar pollen Famili Species Index P/E
Ukuran (µm)
Bentuk
1. Fabaceae
Acacia sp. 83,3 62,5 : 75 Polen Besar
suboblate
2. Myrtaceae
Psidium guajava L.
100 50 : 50 Polen Sedang
Prolate spheroidal
3. Myrtaceae
Syzygium aqueum
100 50 : 50 Polen sedang
Prolate spheroidal
4. Myrtaceae
Syzygium malaccense L.
100 50 : 50 Polen sedang
Prolate spheroidal
5. Solanaceae
Solanum sp.
100 50 : 50 Polen sedang
Prolate spheroidal
6. Gaminae
Poaceae sp. 100 100 : 100 Polen sangat besar
Prolate spheroidal
7. Euphorbiaceae
Baccaurea
courtallensis
100 37,5 : 37,5 Polen sedang
Prolate spheroidal
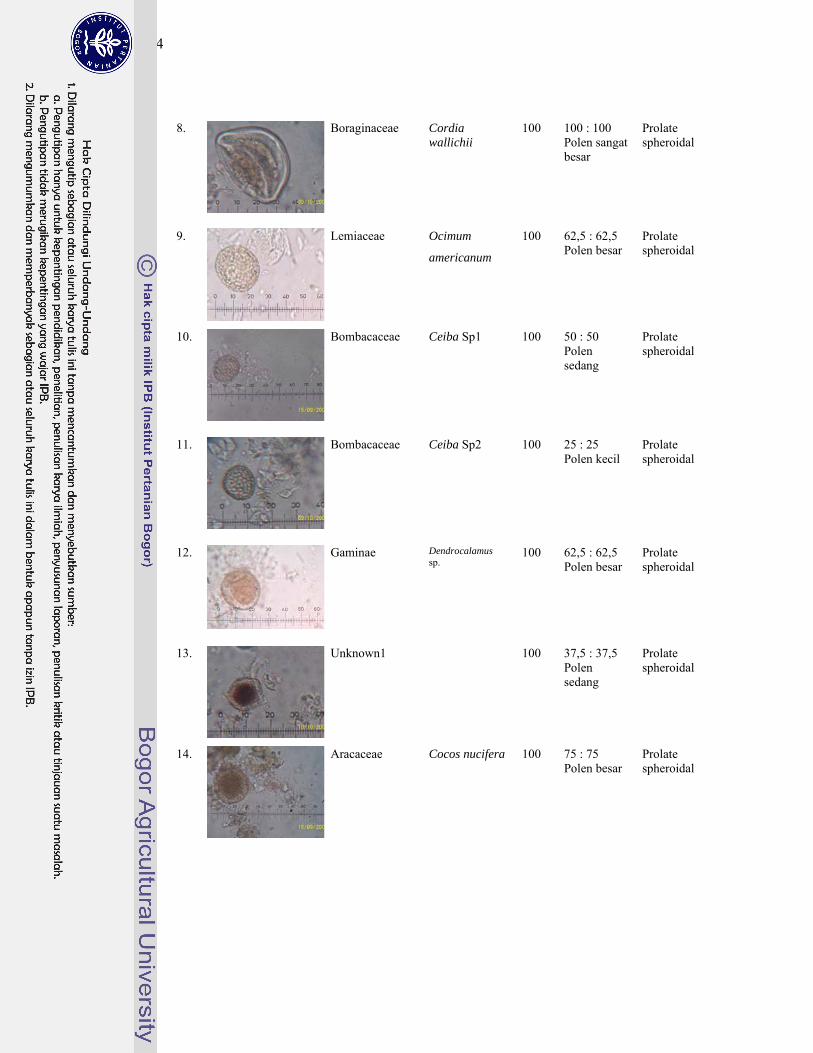
144
8. Boraginaceae
Cordia wallichii
100 100 : 100 Polen sangat besar
Prolate spheroidal
9. Lemiaceae
Ocimum
americanum
100 62,5 : 62,5 Polen besar
Prolate spheroidal
10. Bombacaceae
Ceiba Sp1
100 50 : 50 Polen sedang
Prolate spheroidal
11. Bombacaceae
Ceiba Sp2 100 25 : 25 Polen kecil
Prolate spheroidal
12. Gaminae
Dendrocalamus sp.
100 62,5 : 62,5 Polen besar
Prolate spheroidal
13. Unknown1
100 37,5 : 37,5 Polen sedang
Prolate spheroidal
14. Aracaceae
Cocos nucifera
100 75 : 75 Polen besar
Prolate spheroidal
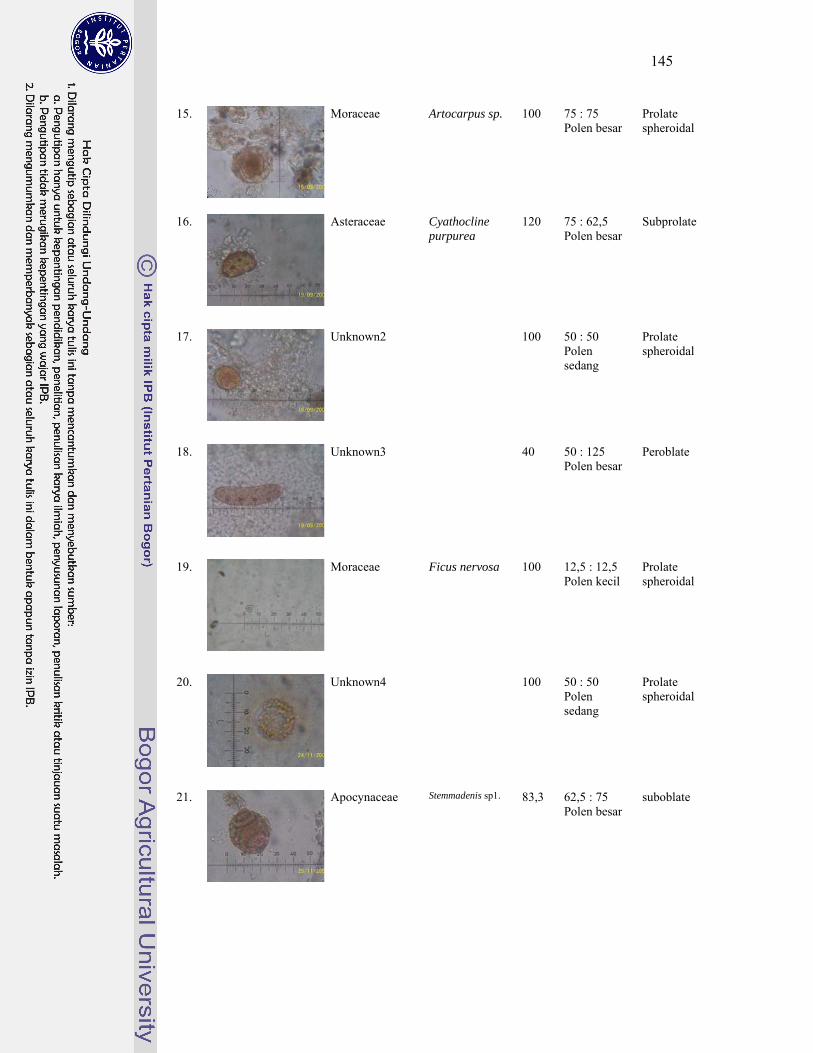
145
15. Moraceae
Artocarpus sp. 100 75 : 75 Polen besar
Prolate spheroidal
16. Asteraceae
Cyathocline purpurea
120 75 : 62,5 Polen besar
Subprolate
17. Unknown2
100 50 : 50 Polen sedang
Prolate spheroidal
18. Unknown3
40 50 : 125 Polen besar
Peroblate
19. Moraceae
Ficus nervosa 100 12,5 : 12,5 Polen kecil
Prolate spheroidal
20. Unknown4
100 50 : 50 Polen sedang
Prolate spheroidal
21. Apocynaceae Stemmadenis sp1. 83,3 62,5 : 75 Polen besar
suboblate
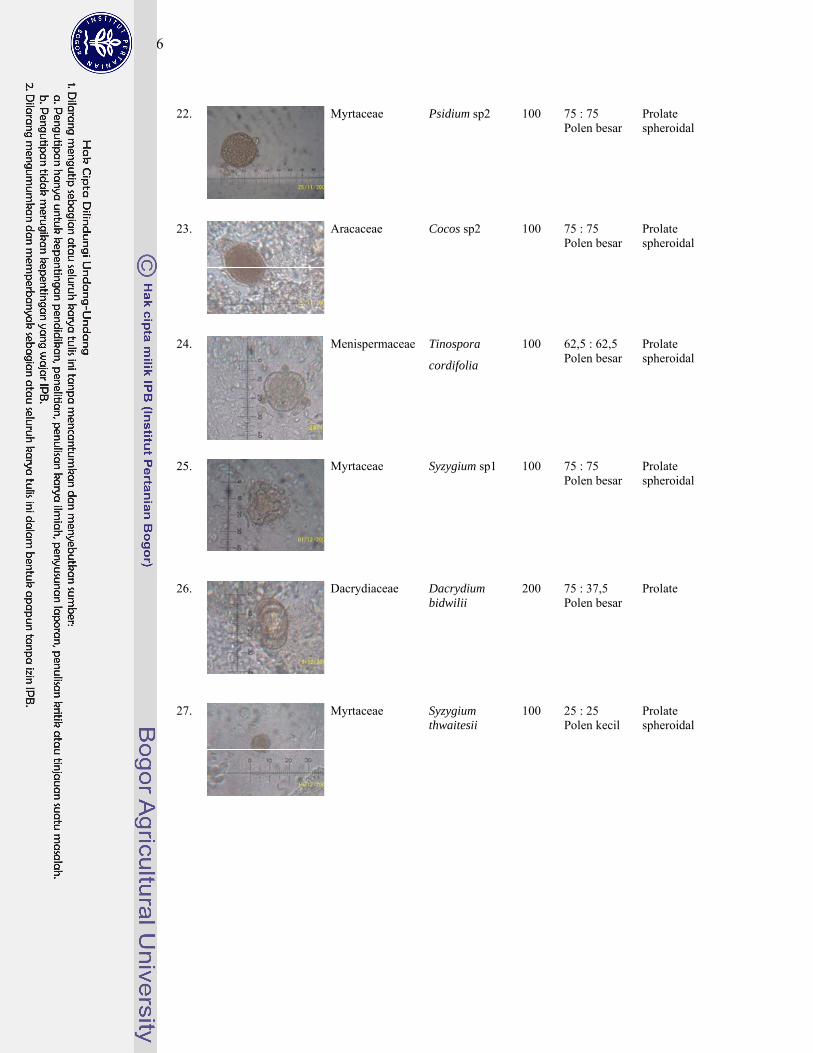
146
22. Myrtaceae
Psidium sp2 100 75 : 75 Polen besar
Prolate spheroidal
23. Aracaceae
Cocos sp2 100 75 : 75 Polen besar
Prolate spheroidal
24. Menispermaceae
Tinospora
cordifolia
100 62,5 : 62,5 Polen besar
Prolate spheroidal
25. Myrtaceae
Syzygium sp1
100 75 : 75 Polen besar
Prolate spheroidal
26. Dacrydiaceae
Dacrydium bidwilii
200 75 : 37,5 Polen besar
Prolate
27. Myrtaceae
Syzygium thwaitesii
100 25 : 25 Polen kecil
Prolate spheroidal
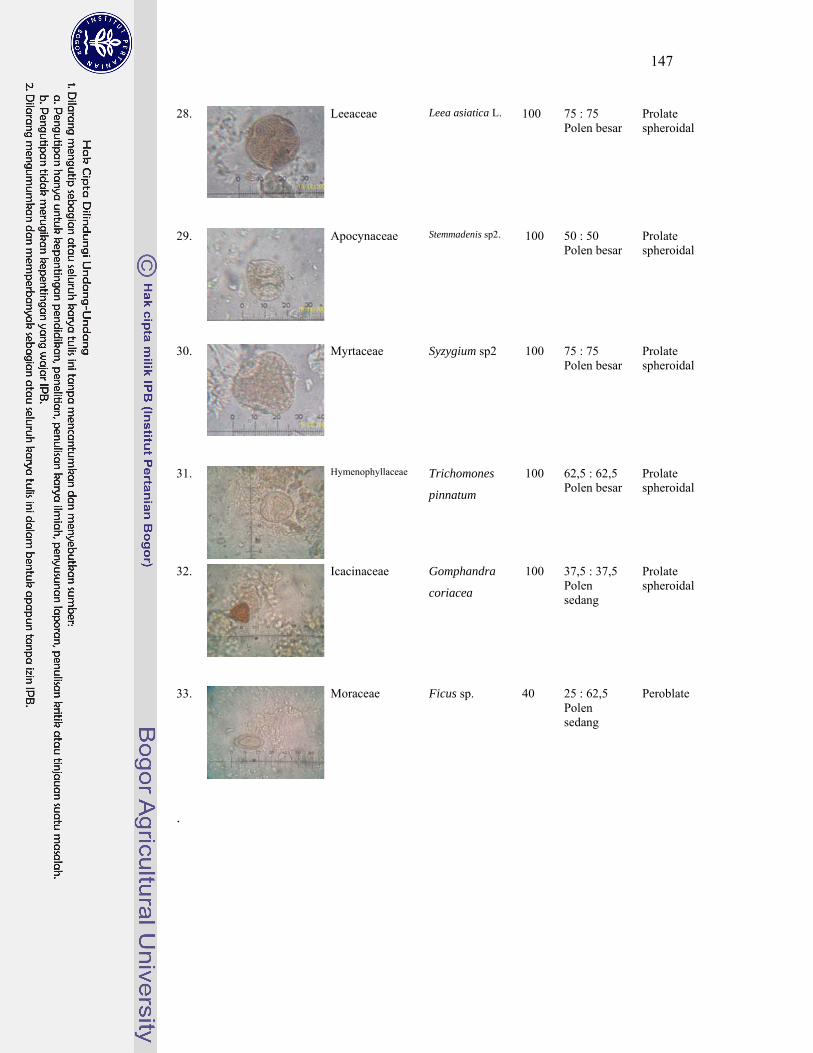
147
.
28. Leeaceae
Leea asiatica L.
100 75 : 75 Polen besar
Prolate spheroidal
29. Apocynaceae
Stemmadenis sp2. 100 50 : 50 Polen besar
Prolate spheroidal
30. Myrtaceae
Syzygium sp2
100 75 : 75 Polen besar
Prolate spheroidal
31. Hymenophyllaceae
Trichomones
pinnatum
100 62,5 : 62,5 Polen besar
Prolate spheroidal
32. Icacinaceae
Gomphandra
coriacea
100 37,5 : 37,5 Polen sedang
Prolate spheroidal
33. Moraceae
Ficus sp.
40 25 : 62,5 Polen sedang
Peroblate
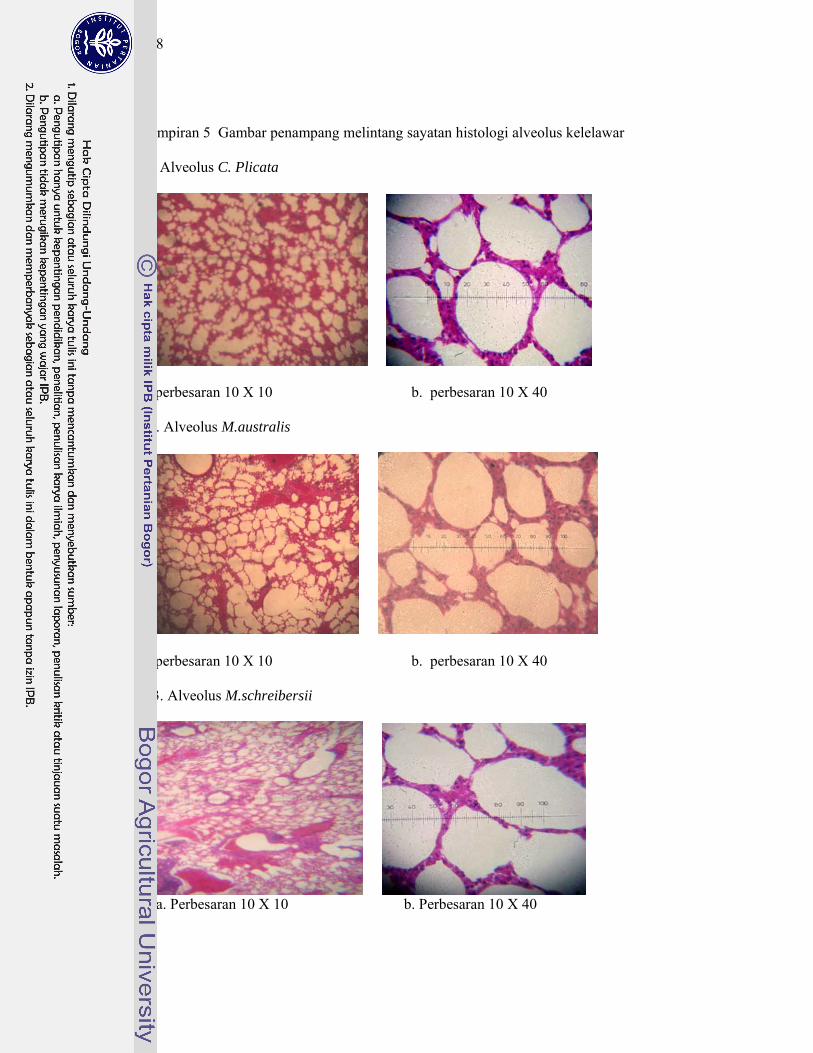
148
Lampiran 5 Gambar penampang melintang sayatan histologi alveolus kelelawar 1. Alveolus C. Plicata
a. perbesaran 10 X 10 b. perbesaran 10 X 40
2. Alveolus M.australis
a. perbesaran 10 X 10 b. perbesaran 10 X 40 3. Alveolus M.schreibersii
a. Perbesaran 10 X 10 b. Perbesaran 10 X 40
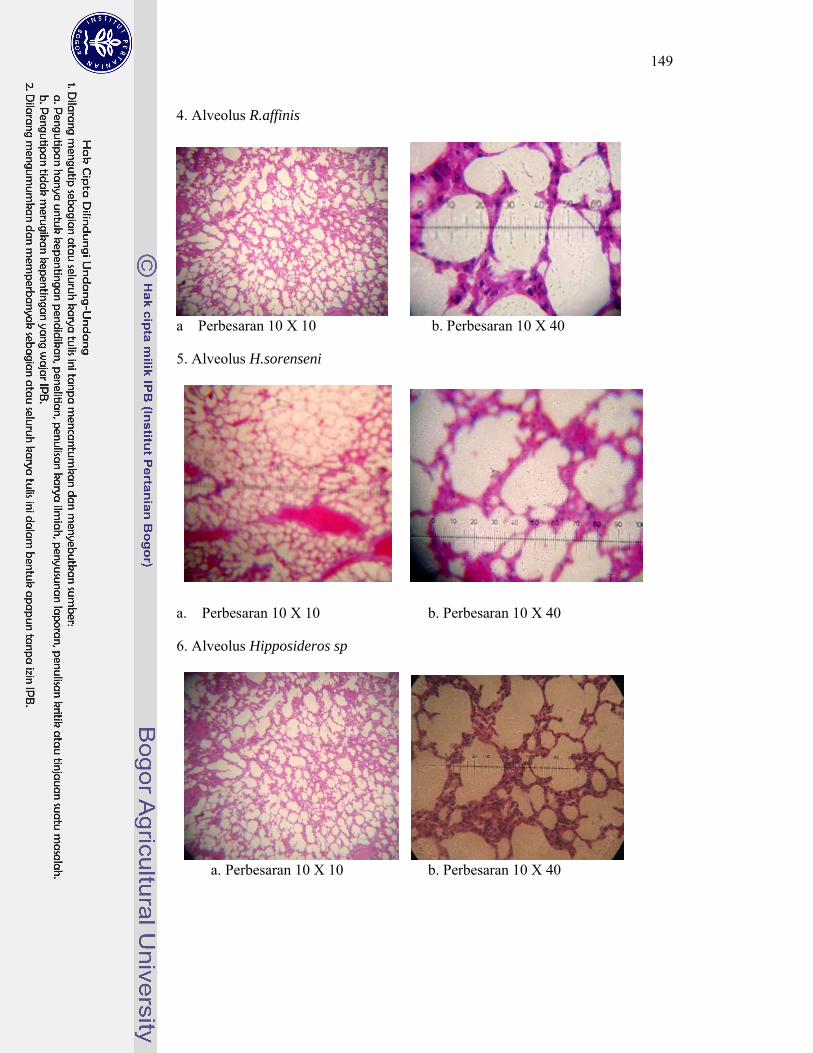
149
4. Alveolus R.affinis
a Perbesaran 10 X 10 b. Perbesaran 10 X 40
5. Alveolus H.sorenseni
a. Perbesaran 10 X 10 b. Perbesaran 10 X 40
6. Alveolus Hipposideros sp
a. Perbesaran 10 X 10 b. Perbesaran 10 X 40
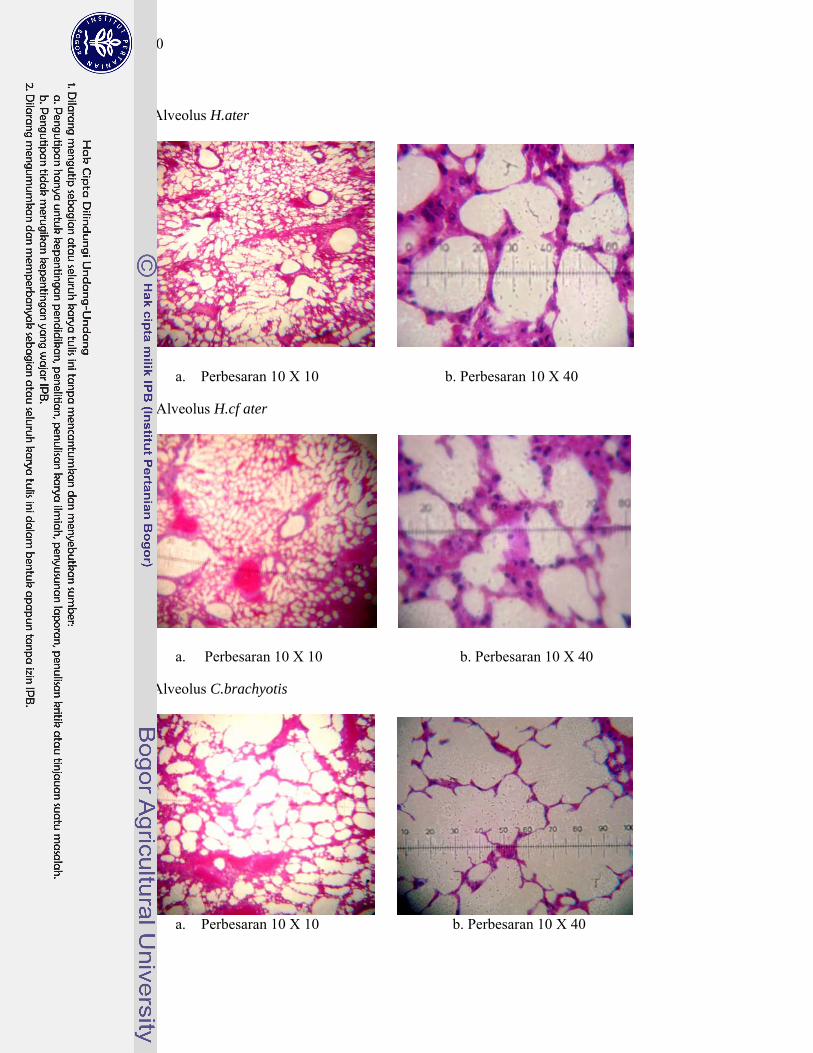
150
7.Alveolus H.ater
a. Perbesaran 10 X 10 b. Perbesaran 10 X 40
8. Alveolus H.cf ater
a. Perbesaran 10 X 10 b. Perbesaran 10 X 40
9.Alveolus C.brachyotis
a. Perbesaran 10 X 10 b. Perbesaran 10 X 40
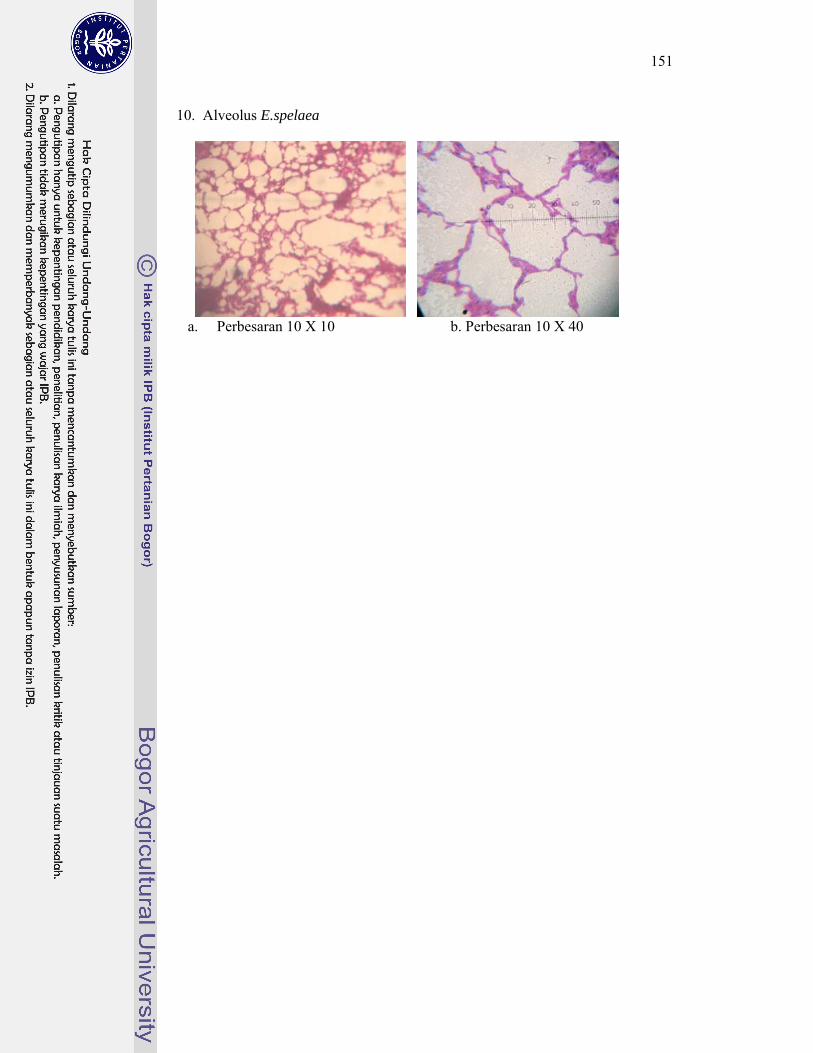
151
10. Alveolus E.spelaea
a. Perbesaran 10 X 10 b. Perbesaran 10 X 40
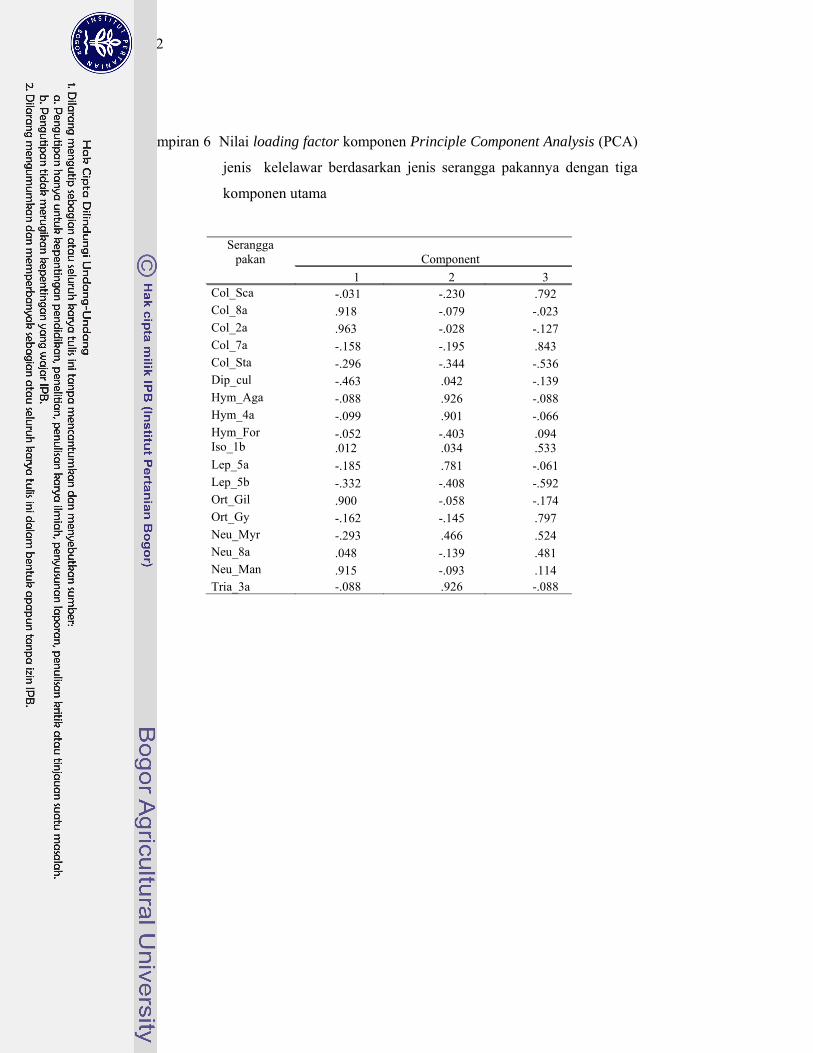
152
Lampiran 6 Nilai loading factor komponen Principle Component Analysis (PCA)
jenis kelelawar berdasarkan jenis serangga pakannya dengan tiga
komponen utama
Serangga
pakan Component 1 2 3 Col_Sca -.031 -.230 .792 Col_8a .918 -.079 -.023 Col_2a .963 -.028 -.127 Col_7a -.158 -.195 .843 Col_Sta -.296 -.344 -.536 Dip_cul -.463 .042 -.139 Hym_Aga -.088 .926 -.088 Hym_4a -.099 .901 -.066 Hym_For Iso_1b
-.052 .012
-.403 .034
.094
.533 Lep_5a -.185 .781 -.061 Lep_5b -.332 -.408 -.592 Ort_Gil .900 -.058 -.174 Ort_Gy -.162 -.145 .797 Neu_Myr -.293 .466 .524 Neu_8a .048 -.139 .481 Neu_Man .915 -.093 .114 Tria_3a -.088 .926 -.088
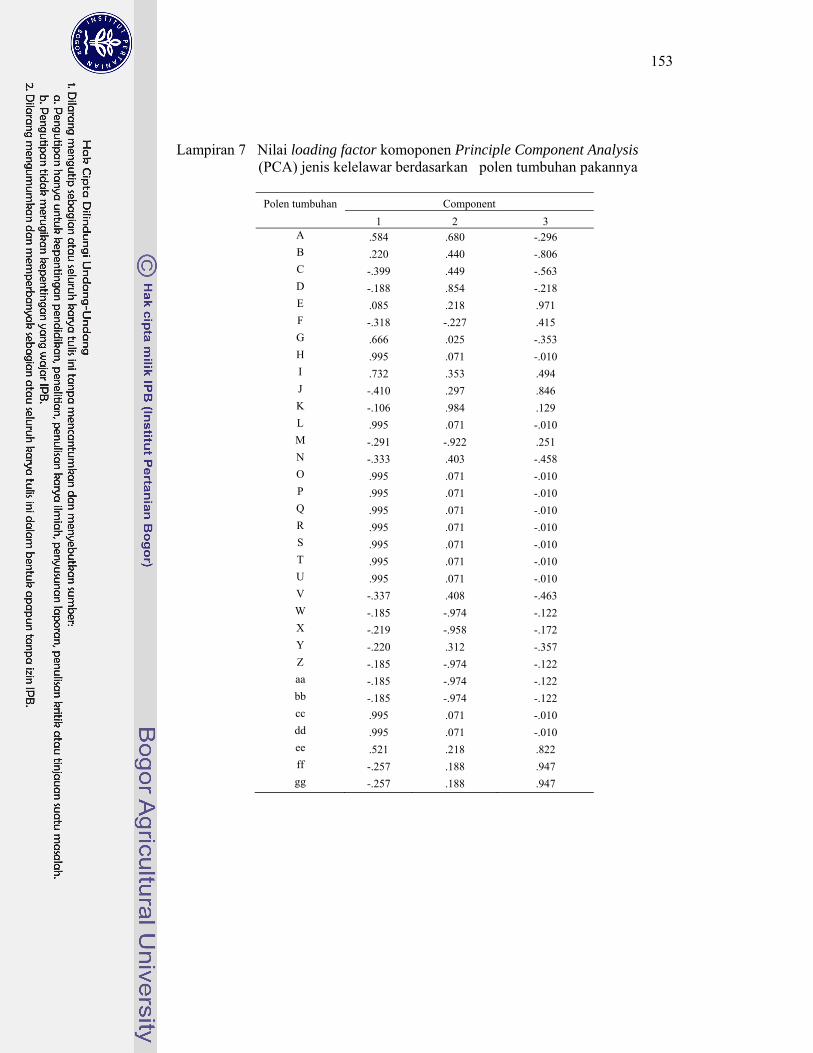
153
Lampiran 7 Nilai loading factor komoponen Principle Component Analysis
(PCA) jenis kelelawar berdasarkan polen tumbuhan pakannya
Polen tumbuhan Component 1 2 3
A .584 .680 -.296 B .220 .440 -.806 C -.399 .449 -.563 D -.188 .854 -.218 E .085 .218 .971 F -.318 -.227 .415 G .666 .025 -.353 H .995 .071 -.010 I .732 .353 .494 J -.410 .297 .846 K -.106 .984 .129 L .995 .071 -.010 M -.291 -.922 .251 N -.333 .403 -.458 O .995 .071 -.010 P .995 .071 -.010 Q .995 .071 -.010 R .995 .071 -.010 S .995 .071 -.010 T .995 .071 -.010 U .995 .071 -.010 V -.337 .408 -.463 W -.185 -.974 -.122 X -.219 -.958 -.172 Y -.220 .312 -.357 Z -.185 -.974 -.122 aa -.185 -.974 -.122 bb -.185 -.974 -.122 cc .995 .071 -.010 dd .995 .071 -.010 ee .521 .218 .822 ff -.257 .188 .947 gg -.257 .188 .947