virulensi beberapa isolat cendawan entomopatogen...
TRANSCRIPT

Prayogo: Virulensi isolat entomopatogen dan pengendalian penggerek ubijalar C. formicarius 738
VIRULENSI BEBERAPA ISOLAT CENDAWAN ENTOMOPATOGEN Beauveria bassiana (Balsamo) Vuillemen UNTUK MENGENDALIKAN
PENGGEREK UBIJALAR Cylas formicarius Yusmani Prayogo
Balai Penelitian Tanaman Kacang-kacangan dan Umbi-umbian
ABSTRAK Cylas formicarius merupakan salah satu hama yang sangat penting pada ubijalar karena
dapat menyebabkan kehilangan hasil hingga 100%. Penelitian ini bertujuan untuk mempe-lajari virulensi beberapa isolat cendawan entomopatogen Beauveria bassiana untuk mengen-dalikan C. formicarius. Penelitian dilakukan di laboratorium Entomologi, Balai Penelitian Tanaman Kacang-kacangan dan Umbi-umbian (BALITKABI). Perlakuan adalah 15 isolat B. bassiana yang diperoleh dari berbagai lokasi sentra produksi ubijalar di Indonesia. Hasil penelitian menunjukkan bahwa semua isolat B. bassiana yang diuji mampu menginfeksi dan membunuh C. formicarius dalam rentang waktu 7 hari setelah aplikasi (HSA). Infeksi cenda-wan ditandai dengan tumbuhnya miselium berwarna putih pada bagian organ tubuh serangga meliputi ruas tungkai, ruas antena, ruas toraks dan segmen-segmen abdomen. Kolo-nisasi miselium cendawan berkembang menyelimuti seluruh tubuh serangga sehingga serangga tertutup penuh oleh miselium yang berwarna putih membentuk seperti mumi. Diperoleh tiga isolat B. bassiana yang virulen dalam membunuh C. formicarius hingga 100%, yaitu isolat yang berasal dari Bogor (BGR 2), Probolinggo (PB 2) dan Tumpang (TMP 2). Isolat B. bassiana yang diisolasi dari serangga lebih virulen dibandingkan dengan isolat yang diisolasi dari permukaan tanah. Hasil uji kerapatan konidia menunjukkan bahwa aplikasi cendawan B. bassiana isolat BGR 2, PB 2 maupun TMP 2 menggunakan kerapatan konidia 108/ml lebih efektif dalam menekan kerusakan umbi. Kerusakan umbi pada perlakuan kontrol (tanpa aplikasi B. bassiana) hingga mencapai 100% sehingga umbi sudah tidak layak untuk dikonsumsi. Aplikasi B. bassiana dengan kerapatan konidia 108/ml mampu menekan keru-sakan umbi akibat C. formicarius mencapai 98%. Efikasi isolat B. bassiana PB 2 dan TMP 2 lebih tinggi dibandingkan isolat BGR 2 dalam menekan kerusakan umbi akibat C. formicarius. Ketiga isolat B. bassiana berpeluang besar dapat digunakan sebagai salah satu agens pengendalian C. formicarius.
Kata kunci: Virulensi, mortalitas, B. bassiana, Ubijalar, C. formicarius.
ABSTRACT Virulence of some isolates of entomopathogenic fungus Beauveria bassiana to control
Cylas formicarius. Cylas formicarius is one of the most important pests in sweet potato caused yield losses up to 100%. This research to study the virulence of some isolates of entomopathogenic fungus Beauveria bassiana to control C. formicarius. The research was conducted in the laboratory of Entomology at Indonesian Legumes and Tuber Crops Research Institute (ILETRI). The treatment were 15 isolates of B. bassiana obtained from a variety of central sweet potato production in Indonesia. The results showed that all isolates of B. bassiana were tested was able to infect and kill of C. formicarius within seven days after application (DAA). Fungus infection were characterized by white mycelial growth on the insect body organs including leg segment, antenna, thorac and abdominal. Colonization by the white mycelium covering the entire body formed like a mummy. Obtained three isolates of B.

Prosiding Seminar Hasil Penelitian Tanaman Aneka Kacang dan Umbi 2011 739
bassiana is virulent in killing C. formicarius up to 100%, i.e. isolates from Bogor (BGR 2), Probolinggo (PB 2) and Tumpang (TMP 2). B. bassiana were isolated from insects are more virulent than the isolate from the soil. The result density test indicate that the application of B. bassiana BGR 2, PB 2 and TMP 2 isolates using the 108/ml conidia more effective in suppressing tubers damage. Tuber damage up to 100% on untreated (without B. bassiana), so that the tuber not feasible for consumption. Application of B. bassiana by 108/ml conidia densities able reducing tuber damage were caused C. formicarius reached 98%. Efficacy of B. bassiana TMP 2 and PB 2 isolates more higher than BGR 2 in suppressing tuber damage caused by C. formicarius. Three isolates of B. bassiana most likely to be used as a agent to control of C. formicarius.
Key words: Virulence, mortality, B. bassiana, sweet potato, C. formicarius.
PENDAHULUAN Penggerek umbi (Cylas formicarius F.) (Coleoptera: Curculionidae) merupakan salah
satu hama yang sangat penting pada ubijalar karena dapat merusak tanaman mulai di lapangan hingga di tempat penyimpanan. Hama ini tersebar luas di seluruh dunia, terutama daerah yang memiliki iklim kering karena serangga tersebut akan berkembang optimal (Jansson et al. 1990; Miyaji & Tanaka 1998; Ohara et al. 2000; Masaki et al. 2002). Inang utama dari C. formicarius adalah Ipomoea batatas, sedangkan inang alternatifnya adalah jenis tanaman yang termasuk kelompok Convolvulaceae, Solanum tuberosum, sweet cassava, Manihot utilisima, Xanthosoma sagittifolium, Dioscorea sp., Calathes allouya dan Apium gravealens (Austin 1991). Kehilangan hasil ubijalar akibat serangan C. formicarius hingga mencapai 100% (Capinera 2002). C. formicarius selain menyebabkan kerusakan umbi, juga menghasilkan senyawa terpen dari bekas gerekan yang tertinggal sehingga dapat meracuni konsumen (Sato et al. 1982).
Pengendalian C. formicarius yang dilakukan oleh petani umumnya aplikasi insekti-sida kimia (Nonci & Sriwidodo 1994). Insektisida sistemik yang berbahan aktif karbo-furan dilaporkan dapat menekan populasi hama ini di lapangan, namun kualitas umbi yang dihasilkan sangat rendah karena masih ditemukan gerekan pada umbi sehingga kurang layak untuk dikonsumsi (Nonci 2005). Hal ini disebabkan hama yang mati sangat terbatas pada stadia imago yang umumnya menghuni sekitar batang dan daun di atas permukaan tanah. Sementara itu, stadia C. formicarius yang lain (telur dan larva) khususnya yang hidup di dalam umbi berada di dalam tanah, dengan demikian senyawa insektisida kimia tidak dapat menjangkau serangga (Yamaguchi 2000). Fakta lain membuktikan bahwa senyawa insektisida kimia belum ada yang mampu menggagalkan penetasan telur (ovicidal) C. formicarius.
Komi (2000) menganjurkan pengendalian C. formicarius dengan cara eradikasi lahan dari sumber inokulum, yaitu termasuk pembakaran bekas tanaman yang rusak termasuk umbi yang ada di dalam tanah. Kegiatan tersebut dianggap tindakan strategi pengen-dalian yang lebih menguntungkan dibandingkan cara pengendalian lain. Namun pengendalian C. formicarius secara terintegrasi dengan cara pengelolaan tanaman terpadu secara nyata membuktikan hasil yang lebih baik (Yasuda 2000; Komi 2000; Tarafdar & Sarkar 2005; Kumano et al. 2007). Pengelolaan tanaman terpadu (PHT) terhadap hama C. formicarius, yaitu memadukan berbagai teknologi pengendalian yang bertujuan untuk mempertahankan populasi hama yang ada hanya dibawah nilai ambang

Prayogo: Virulensi isolat entomopatogen dan pengendalian penggerek ubijalar C. formicarius 740
kerusakan. Pengendalian dengan cara teknik budidaya dalam konsep PHT antara lain penggunaan stek harus terbebas dari infestasi telur maupun larva C. formicarius, membersihkan lahan dan membakar sisa bekas tanaman atau umbi yang rusak, rotasi tanaman bukan inang dan mempertahankan kondisi tanah tidak mengalami kekeringan (Jansson 1991; Yasuda 1999; Lagnaoui et al. 2000; Yasuda 2000). Tanah yang kering akan memicu terjadinya bongkahan/retakan permukaan tanah di sekitar batang ubijalar, kejadian tersebut menjadi salah satu pemicu penetrasi imago betina C. formicarius dalam proses oviposisi (peletakan telur) pada umbi di dalam tanah.
Pemanfaatan agens hayati cendawan entomopatogen Beauveria bassiana termasuk salah satu cara dalam konsep PHT dan mempunyai peluang yang besar untuk diterapkan (Yasuda 2000). Hal ini disebabkan, cendawan entomopatogen tersebut mampu hidup di dalam tanah sehingga sangat kondusif dalam menginfeksi serangga yang sebagian besar hidupnya berada di dalam tanah. Hasil penelitian Ondiaka et al. (2008) menunjukkan bahwa aplikasi cendawan B. bassiana selain efektif dalam membunuh C. formicarius, cendawan tersebut juga mampu menurunkan daya makan, kesuburan (fecundity) dan menggagalkan penetasan telur (ovicidal). Sedangkan Rosfi-ansyah (2009) memaparkan bahwa aplikasi cendawan B. bassiana dengan kerapatan konidia 108/ml mampu menekan kerusakan umbi sebesar 13%.
Menurut Ekesi (2001), isolat cendawan B. bassiana yang diisolasi dari hama Ootheca mutabilis Shalberg (Coleoptera: Chrysomelidae) mempunyai virulensi hingga 100% karena mampu membunuh serangga yang sama, yaitu imago O. mutabilis dalam rentang tujuh hari setelah aplikasi. Beberapa peneliti lain menyebutkan bahwa virulensi cendawan juga dipengaruhi oleh faktor daerah geografis asal isolat diperoleh. Seperti laporan Berreta et al. (1998) bahwa isolat B. bassiana yang berasal dari Argentina dan Brazil mempunyai virulensi yang berbeda terhadap hama Diatraea saccharalis (Lepidoptera: Pyralidae). Penelitian ini bertujuan untuk mendapatkan isolat B. bassiana yang virulen hasil isolasi dari berbagai lokasi sentra produksi ubijalar di Indonesia dan mendapatkan kerapatan konidia B. bassiana optimal untuk mengendalikan C. formicarius.
BAHAN DAN METODE Penelitian dilakukan di Laboratorium Entomologi, Balai Penelitian Tanaman Kacang-
kacangan dan Umbi-umbian (BALITKABI) yang dimulai dari bulan Januari sampai dengan Juli 2011. Rancangan percobaan yang digunakan adalah acak lengkap (RAL) dan diulang tiga kali. Penelitian ini terdiri dari dua tahap yaitu; (1) uji virulensi isolat cendawan entomopatogen B. bassiana terhadap C. formicarius dan (2) uji berbagai tingkat kerapatan konidia isolat B. bassiana yang virulen hasil penelitian tahap pertama untuk menekan kerusakan umbi ubijalar. Isolat cendawan B. bassiana yang diuji pada penelitian tahap pertama adalah 15 isolat yang diperoleh dari beberapa sentra produksi ubijalar di Indonesia, baik dari serangga mati maupun dari dalam tanah (Tabel 1). Sedangkan perlakuan pada penelitian kedua adalah tingkat kerapatan konidia isolat cendawan B. bassiana yang virulen hasil penelitian tahap pertama, yaitu kerapatan 106, 107, 108/ml dan kontrol (air steril).

Prosiding Seminar Hasil Penelitian Tanaman Aneka Kacang dan Umbi 2011 741
Tabel 1. Berbagai isolat cendawan B. bassiana yang diperoleh dari berbagai sumber dan lokasi produksi ubijalar di Indonesia.
Kode isolat Asal isolat Sumber isolat BGR 1 Kopo, Bogor Croccidolomia pavonana BGR 2 Cisarua, Bogor C. pavonana BGR 4 Cibeureum, Bogor Tanah MJ Pacet, Mojokerto Tanah LMB Lembang, Bandung C. pavonana BL Udanawu, Blitar Tanah MLG Kendalpayak, Malang Leptocoriza acuta BWG 1 Genteng, Banyuwangi Tanah BWG 2 Glenmore, Banyuwangi Tanah BGR 5 Dermaga, Bogor C. formicarius JB Jombang Tanah PB 1 Sumberasih, Probolinggo L. acuta PB 2 Muneng, Probolinggo L.acuta TMP 1 Sekarpuro,Tumpang Riptortus linearis TMP 2 Pakiskembar, Tumpang C. formicarius
Pelaksanaan penelitian sebagai berikut:
Uji virulensi berbagai isolat cendawan B. bassiana terhadap C. formicarius
Imago atau larva C. formicarius diperoleh dengan cara mengumpulkan umbi-umbi yang rusak karena tergerek oleh C. formicarius dari pasar Gadang (Malang). Selanjutnya, umbi-umbi yang rusak dimasukkan ke dalam kantong kain yang sudah berisi umbi segar kemudian kantong diikat erat menggunakan tali agar serangga tidak dapat keluar menembus kantong. Jumlah umbi segar yang disediakan di dalam kantong diusahakan dalam jumlah yang banyak dengan harapan imago maupun larva C. formicarius dapat berkembangbiak dengan optimal karena tersedia makanan yang cukup berlimpah. Setiap bulan dilakukan pemisahan serangga berdasarkan stadia untuk memperoleh populasi yang memiliki umur yang seragam. Serangga-serangga yang baru dipisahkan kemudian diberi pakan umbi yang masih segar dan dipelihara di dalam kotak plastik. Pemeliharaan serangga uji dilakukan terus menerus hingga memperoleh populasi minimal mencapai 3.000 ekor imago dengan umur yang sama. Imago C. formicarius yang baru terbentuk sebanyak 50 ekor setiap perlakuan dan setiap ulangan sebagai serangga uji dimasukkan ke dalam kotak plastik yang diberi pakan umbi segar dua buah setiap masing-masing kotak.
Masing-masing isolat cendawan B. bassiana sebagai perlakuan ditumbuhkan pada media potato dextrose agar (PDA) di dalam cawan Petri. Pada umur 21 hari setelah inokulasi (HSI), biakan cendawan di dalam setiap cawan ditambah air 10 ml kemudian koloni cendawan dikerok menggunakan kuas halus. Selanjutnya, suspensi konidia ditambah Tween 80 sebanyak 2 ml dan dikocok menggunakan vortex selama 30 detik dengan tujuan konidia B. bassiana yang bersifat hidropobik dapat bercampur secara homogen. Suspensi konidia dihitung menggunakan haemocytometer untuk memperoleh

Prayogo: Virulensi isolat entomopatogen dan pengendalian penggerek ubijalar C. formicarius 742
kerapatan konidia rata-rata 107/ml. Suspensi dari masing-masing isolat cendawan disemprotkan pada 50 ekor imago C. formicarius, masing-masing perlakuan sebanyak 2 ml. Selanjutnya imago C. formicarius dimasukkan ke dalam kotak plastik yang diisi umbi segar sebagai pakan kemudian kotak plastik ditutup dengan kain kasa.
Peubah yang diamati adalah mortalitas C. formicarius yang terinfeksi B. bassiana setiap hari sampai dengan 21 hari setelah aplikasi (HSA) dan tingkat kolonisasi miselium isolat B. bassiana pada tubuh imago C. formicarius.
Uji kerapatan konidia isolat B. bassiana untuk mengendalikan C. formicarius
Imago C. formicarius yang digunakan dalam uji ini adalah yang berumur tujuh hari setelah terbentuk dari pupa dengan tujuan serangga betina sudah siap kopulasi dan segera bertelur. Jumlah imago C. formicarius yang digunakan adalah 25 pasang betina dan jantan (25 ekor) pada setiap perlakuan dan setiap ulangan.
Isolat cendawan yang virulen hasil penelitian tahap pertama ditumbuhkan pada media PDA di dalam cawan Petri. Pada umur 21 HSI, biakan cendawan di dalam cawan Petri ditambah dengan air 10 ml pada setiap cawan. Selanjutnya, koloni cendawan dikerok menggunakan kuas halus untuk mengambil kelompok konidia pada bagian permukaan koloni. Suspensi konidia kemudian dimasukkan ke dalam tabung reaksi dan ditambah Tween 80 sebanyak 2 ml sebagai perata selanjutnya dikocok menggunakan vortex selama 30 detik. Suspensi konidia dihitung menggunakan haemocytometer hingga memperoleh kerapatan 106, 107 dan 108/ml dan 0 (kontrol) sesuai perlakuan. Masing-masing suspensi konidia sesuai perlakuan diaplikasikan pada 50 ekor imago C. formicarius seperti kegiatan pertama. Selanjutnya, C. formicarius dimasukkan ke dalam kotak plastik yang diberi dua buah umbi segar sebagai pakan.
Peubah yang diamati adalah jumlah imago C. formicarius yang mati terinfeksi cendawan B. bassiana dan persentase kerusakan umbi akibat gerekan imago C. formicarius.
HASIL DAN PEMBAHASAN
Virulensi berbagai isolat cendawan B. bassiana terhadap imago C. formicarius
Virulensi isolat cendawan dinilai dari persentase imago C. fomicarius yang mati terinfeksi cendawan B. bassiana. Uji virulensi bertujuan untuk menyeleksi 15 isolat cendawan B. bassiana dengan tujuan memperoleh beberapa isolat yang efektif untuk mengendalikan C. formicarius. Hasil pengujian menunjukkan bahwa dari 15 isolat cendawan B. bassiana yang diaplikasikan ke imago C. formicarius, semuanya mampu menginfeksi dan mampu menyebabkan kematian imago C. formicarius. Pengamatan secara langsung pada serangga uji setelah disemprot dengan suspensi cendawan B. bassiana menunjukkan perilaku yang berbeda dengan serangga uji pada perlakuan kontrol. Serangga yang terinfeksi cendawan entomopatogen menampakkan pergerakan lamban bahkan cenderung diam dan kurang nafsu makan. Pengamatan lebih lanjut pada umbi yang disediakan di dalam kotak plastik, tampak hanya beberapa bagian yang

Prosiding Seminar Hasil Penelitian Tanaman Aneka Kacang dan Umbi 2011 743
dirusak oleh C. formicarius. Sementara itu, pada perlakuan kontrol dengan aplikasi air steril tampak umbi yang disediakan hampir habis digerek oleh serangga uji.
Kematian C. formicarius akibat infeksi cendawan B. bassiana baru tampak pada hari ketujuh setelah aplikasi (Tabel 2). Pada 7 HSA, mortalitas C. formicarius sangat bervariasi 10-63%. Kematian C. formicarius tertinggi terjadi pada aplikasi menggunakan isolat B. bassiana yang diperoleh dari BGR 2 (Cisarau, Bogor) dan TMP 2 (Pakiskembar, Tumpang) yaitu 63%. Sedangkan mortalitas C. formicarius terendah terjadi pada perla-kuan isolat B. bassiana yang diperoleh dari Jombang (JB), yaitu hanya 10%.
Pada 9 HSA, mortalitas C. formicarius terjadi peningkatan cukup nyata, kecuali perla-kuan isolat B. bassiana dari Jombang karena kematian serangga tampak tidak mengalami peningkatan. Isolat cendawan B. bassiana BGR 2, PB 1 (Sumberasih, Probolinggo), PB 2, TMP 1 (Sekarpuro, Tumpang) dan TMP 2 (Pakiskembar, Tumpang) tampak lebih unggul dibandingkan dengan isolat yang lain karena mampu membunuh C. formicarius masing-masing 87%; 66%; 76%; 60%, dan 90%. Sedangkan pada hari ke- 12 setelah aplikasi diperoleh dua isolat B. bassiana yang lebih virulen dalam membunuh imago C. formicarius, karena kematian serangga hingga mencapai 100%. Namun kedua perlakuan tersebut tidak berbeda nyata dengan isolat BGR 1, BGR 4, MJ (Mojokerto), LMB (Lembang, Bandung), BWG 1 (Genteng, Banyuwangi), BWG 2 (Glenmore, Banyuwangi) dan MLG (Kendalpayak, Malang). Pada 15 HSA, diperoleh tiga isolat B. bassiana yang memiliki virulensi tertinggi karena mampu membunuh serangga hingga 100% lebih cepat dibandingkan dengan 12 isolat B. bassiana lainnya, yaitu isolat BGR 2, PB 2 maupun TMP 2. Meskipun secara statistik ketiga isolat tersebut tidak berbeda nyata dengan isolat BGR 1, BGR 4, MJ, LMB, BL (Udanawu, Blitar), MLG, BWG 1, BWG 2, BGR 5 (Dermaga, Bogor) dan TMP 1.
Pengamatan hingga 21 HSA, menunjukkan bahwa ketiga isolat yaitu BGR 2, PB 2 dan TMP 2 tetap konsisten dengan mortalitas serangga uji tertinggi yaitu 100% dan tidak ada isolat lainnya yang mampu menandingi persentase kematian tersebut. Meskipun secara statistik persentase kematian C. formicarius tersebut tidak berbeda nyata dengan isolat BGR 1 (96,67%), MJ (86,67%), LMB (96,67%), MLG (96,67%), BWG 1 (96,67%), BWG 2 (90%), BGR 5 (93,33%) dan PB 1 (93,33%). Dilihat dari persentase mortalitas ketiga isolat (BGR 2, PB 2 dan TMP 2) mencapai 100% maka isolat-isolat tersebut dapat dikatakan memiliki virulensi yang tertinggi dibandingkan dengan 12 isolat lainnya.
Dari hasil penelitian ini mengindikasikan bahwa isolat B. bassiana yang diperoleh dari isolasi serangga mati memiliki virulensi lebih tinggi dibandingkan dengan isolat yang diperoleh dari tanah. Menurut Prayogo (2009) bahwa isolat cendawan entomopatogen yang diperoleh dari dalam tanah masih ada pengaruh berbagai cekaman, seperti akti-vitas pestisida kimia maupun senyawa metabolit bekas tanaman yang ada di permukaan tanah sehingga cendawan dalam keadaan fase saprob karena inang tidak ada. Oleh karena itu, pada waktu uji virulensi isolat cendawan tidak dapat mengekspresikan karakter fisiologi secara total termasuk karakter virulensi.
Menurut Ropek dan Para (2002), pertumbuhan cendawan entomopatogen Vertic-illium lecanii yang diperoleh dari tanah dipengaruhi oleh berbagai kontaminan dari logam berat seperti cadmium (Cd) dan plumbun (Pb). Selanjutnya, Popowska-Nowak et al. (2000) juga melaporkan bahwa pertumbuhan cendawan Paecilomyces fumosoroseus

Prayogo: Virulensi isolat entomopatogen dan pengendalian penggerek ubijalar C. formicarius 744
(Wize) Brown & Smith (Deuteromycotina: Hyphomycetes) dipengaruhi oleh aktivitas logam berat. Sementara itu, Klingen et al. (2002) memaparkan bahwa senyawa meta-bolit sekunder tanaman juga dapat menghambat pertumbuhan cendawan entomopa-togen Metarhizium anisopliae (Metsch.) (Deuteromycotina: Hyphomycetes) meskipun dalam konsentrasi yang rendah, sendangkan pada konsentrasi yang tinggi akan menyebabkan toksik. Senyawa metabolit sekunder dari berbagai serasah tanaman khususnya dari kelompok Brassicaceae sangat berpengaruh terhadap pertumbuhan dan perkembangan cendawan B. bassiana meskipun cendawan tersebut sudah dikulturkan beberapa kali (Sudirman et al. 2008).
Tabel 2 Mortalitas imago C. formicarius yang terinfeksi berbagai isolat cendawan B. bassiana Persentase mortalitas C. formicarius pada ke ..... n HSA* Asal isolat
7 9 12 15 18 21 BGR 1 23,33 bc 46,67 bcd 73,33 ab 90,00 abc 90,00 abc 96,67 ab BGR 2 63,33 a 86,67 a 100,00 a 100,00 a 100,00 a 100,00 a BGR 4 40,00 ab 70,00 abc 76,67 ab 73,33 abc 73,33 abc 73,33 cd MJ 20,00 bc 46,67 bcd 76,67 ab 80,00 abc 80,00 abc 86,67 abc LMB 20,00 bc 43,33 bcde 76,67 ab 93,33 abc 96,67 ab 96,67 ab BL 23,33 bc 40,00 cde 50,00 b 60,00 cd 66,67 bc 66,67 d MLG 26,67 bc 50,00 bcd 73,33 ab 86,67 abc 90,00 abc 96,67 ab BWG 1 16,67 bc 33,33 de 63,33 ab 70,00 abcd 83,33 abc 90,00 ab BWG 2 20,00 bc 43,33 bcde 70,00 ab 80,00 abc 83,33 abc 90,00 ab BGR 5 33,33 bc 36,67 cde 50,00 b 76,67 abc 90,00 abc 93,33 ab JB 10,00 c 10,00 e 10,00 c 36,67 d 33,33 d 33,33 e PB 1 36,67 abc 66,67 abcd 50,00 b 63,33 bcd 63,33 cd 93,33 ab PB 2 26,67 bc 76,67 ab 90,00 a 100,00 a 100,00 a 100,00 a TMP 1 43,33 ab 60,00 abcd 76,67 ab 76,67 abc 83,33 abc 83,33 bc TMP 2 63,33 a 90,00 a 100,00 a 100,00 a 100,00 a 100,00 a Kontrol (air) 0,00 c 0,00 e 0,00 c 0,00 e 0,00 e 0,00 f LSD 0,05 26,79 34,74 39,13 35,15 31,52 14,85 KK (%) 24,55 29,77 22,17 19,28 18,83 22,35
*Angka-angka selajur yang diikuti oleh huruf yang sama, tidak berbeda nyata pada uji DMRT (Duncan’s Multiple Range Test) pada taraf 0,05. Keterangan: BGR1 (Kopo, Bogor), BGR2 (Cisarua, Bogor), BGR4 (Cibeureum, Bogor), MJ (Mojokerto), LMB (Lembang, Bandung), BL (Blitar), MLG (Kendalpayak, Malang), BWG1 (Genteng, Banyuwangi), BWG2 (Glenmore, Banyuwangi), BGR5 (Dermaga, Bogor), JB (Jombang), PB1 (Sumberasih, Probolinggo), PB2 (Muneng, Probolinggo), TMP1 (Sekarpuro, Tumpang), dan TMP2 (Pakiskembar, Tumpang).
Hasil penelitian ini menunjukkan bahwa virulensi isolat cendawan tidak berkaitan dengan sumber isolat cendawan maupun serangga uji yang digunakan. Kedua isolat yaitu BGR 2 dan PB 2, meskipun diperoleh dari sumber yang berbeda dengan serangga uji ternyata memiliki virulensi yang sama tingginya dengan isolat TMP 2 yang diperoleh dari sumber yang sama dengan serangga uji (C. formicarius). Isolat yang diperoleh dalam satu lokasi dan dari sumber yang sama juga mempunyai keragaman virulensi yang berbeda. Hal ini disebabkan perbedaan virulensi cendawan dipengaruhi oleh kera-gaman intraspesies (Fatiha et al. 2007). Sementara itu, keragaman intraspesies cen-dawan dipengaruhi oleh keragaman genetik isolat (Sugimoto et al. 2003a & 2003b). Namun untuk memudahkan dalam mendapatkan isolat cendawan entomopatogen yang memiliki virulensi tinggi oleh Prayogo (2009) disarankan mengamati karakter fisiologi

Prosiding Seminar Hasil Penelitian Tanaman Aneka Kacang dan Umbi 2011 745
cendawan. Isolat cendawan entomopatogen yang memiliki karakter fisiologi sebagai berikut; hifa atau kelompok hifa agak panjang, tebal, berbentuk seperti wol (wolly) maka isolat cendawan tersebut memiliki virulensi lebih tinggi. Koloni yang tebal akan mengha-silkan jumlah konidia yang lebih banyak dibandingkan dengan koloni cendawan yang tipis (pellicular) maupun koloni agak panjang dan menyebar kesegala arah (cottony) (Prayogo 2009).
Kematian C. formicaroius pada 7 HSA ditandai dengan tumbuhnya miselium pada berbagai organ embelan-embelan serangga, yaitu meliputi segmen-segmen antena maupun segmen-segmen tungkai. Namun demikian, miselium juga ditemukan pada bagian toraks maupun segmen-segmen abdomen meskipun tidak setebal miselium yang tumbuh pada antena maupun tungkai (Gambar 1). Semakin bertambah hari, kolonisasi miselium cendawan pada masing-masing isolat semakin menutupi seluruh tubuh serangga. Hasil pengamatan terhadap mortalitas pada beberapa imago C. formicarius yang terinfeksi berbagai isolat B. bassiana tampak pada perlakuan isolat BGR 2, PB 2 dan TMP 2 tumbuh lebih cepat dibandingkan dengan 12 isolat B. bassiana yang lain (Gambar 2).
Gambar 1. Miselium isolat cendawan B. bassiana BGR 2 yang tumbuh pada berbagai organ embelan-embelan C. formicarius pada umur 7 HSA. (a) tungkai,
(b) antena, (c) torak dan abdomen (d). Pertumbuhan miselium pada tiga isolat cendawan B. bassiana, yaitu BGR 2, PB 2
dan TMP 2 yang virulen tampak bertambah tebal dan tumbuh lebih cepat dalam mengkolonisasi seluruh tubuh imago C. formicarius. Oleh karena itu, seluruh tubuh terbungkus oleh miselium yang berwarna putih dibandingkan dengan isolat B. bassiana
a
b c d

Prayogo: Virulensi isolat entomopatogen dan pengendalian penggerek ubijalar C. formicarius 746
yang avirulen (JB) (Gambar 3). Kolonisasi tubuh C. formicarius tidak hanya dipenuhi oleh kumpulan miselium cendawan, namun juga dipenuhi oleh kumpulan konidia. Karakter isolat cendawan yang demikian akan sangat menguntungkan apabila digunakan sebagai agens hayati di lapangan karena isolat tersebut akan lebih cepat dalam mengkolonisasi populasi hama yang ada di lokasi yang dikendalikan. Hal ini terjadi karena kumpulan konidia yang terbentuk lebih cepat dalam penyebaran (transmisi) dari serangga yang mati ke serangga inang yang sehat sehingga proses diseminasi patogen di lokasi lebih cepat dalam menyebabkan epizooti.
Gambar 2. Perkembangan pertumbuhan miselium isolat cendawan B. bassiana BGR 2 pada beberapa organ tungkai, antena dan abdomen C. formicarius pada umur 9 HSA.
Hasil penelitian ini seperti yang dilaporkan oleh Kassa (2003), dikatakan bahwa
berbagai jenis isolat yang diisolasi dari berbagai lokasi di Ethiopia dan Sudan memiliki tingkat kolonisasi yang berbeda-beda bergantung pada tingkat virulensi isolat. Sementara itu, hasil penelitian Varela dan Morales (1996) sangat berbeda karena pertumbuhan koloni antar isolat yang diperoleh tidak berbeda nyata. Hal ini diduga karena jumlah isolat B. bassiana yang diuji terlalu sedikit sehingga keragaman isolat rendah. Pertum-buhan koloni yang lebih cepat mengindikasikan isolat cendawan tersebut lebih meng-untungkan seperti yang pernah dilaporkan oleh Trizelia (2005) bahwa isolat cendawan B. bassiana yang tumbuh lebih cepat akan lebih menguntungkan digunakan sebagai agens hayati. Hal ini disebabkan hanya sedikit waktu yang dibutuhkan untuk mempro-duksi konidia cendawan, karena konidia berfungsi sebagai organ infektif yang digunakan dalam proses pemencaran dan proses infeksi (Wraight et al. 2001; Lerche et al. 2004). Semakin banyak jumlah konidia yang diproduksi, semakin efektif cendawan tersebut sebagai agens hayati dalam menyebabkan epizooti di lapangan (Purlong & Pell 2001; Ganga-Visalakshy et al. 2004). Di lapangan, umumnya sistem transmisi cendawan ento-mopatogen terjadi secara horizontal artinya konidia pindah dari serangga yang mati ke serangga inang yang sehat dengan bantuan angin, air, serangga maupun aktivitas manu-sia dan bukan faktor keturunan (Wagner & Lewis 2000; Chun & Mingguang 2004).

Prosiding Seminar Hasil Penelitian Tanaman Aneka Kacang dan Umbi 2011 747
Efikasi kerapatan konidia tiga isolat B. bassiana terhadap mortalitas C. formicarius
Isolat cendawan B. bassiana yang digunakan adalah BGR 2, PB 2 dan TMP 2 yaitu yang menunjukkan virulen tinggi hasil penelitian tahap pertama. Sedangkan serangga C. formicarius yang digunakan untuk uji efikasi kerapatan konidia tiga isolat cendawan B. bassiana adalah imago yang sudah tujuh hari terbentuk dari stadia pupa. Keadaan tersebut ditujukan agar imago sudah berkopulasi dan sudah menghasilkan telur. Semen-tara itu, telur yang menetas membentuk larva 1 sudah mempunyai kemampuan untuk merusak umbi. Hasil penelitian ini menunjukkan bahwa semakin tinggi kerapatan konidia B. bassiana yang diaplikasikan, semakin efektif dalam membunuh imago C. formicarius. Hal ini terbukti dari ketiga isolat B. bassiana yang digunakan, yaitu baik isolat yang berasal dari Cisarua, Bogor (BGR 2), Muneng, Probolinggo (PB 2) maupun Pakiskembar, Tumpang (TMP 2). Waktu kematian C. formicarius pada uji kerapatan konidia ini lebih pendek, yaitu hanya dalam rentang waktu tiga hari apabila dibanding-kan dengan uji virulensi (penelitian pertama).
Gambar 3. Mumifikasi (kolonisasi) miselium B. bassiana isolat BGR 2 (a), B. bassiana isolat
PB 2 (b), B. bassiana isolat TMP 2 (c) dan B. bassiana isolat JB (c) pada tubuh C. formicarius setelah umur 15 HSA.
Aplikasi ke imago C. formicarius menggunakan kerapatan konidia 108/ml, setelah tiga
hari sudah membunuh serangga uji hingga di atas 60% (Tabel 3). Mortalitas C. formicarius semakin meningkat dengan bertambahnya jumlah hari pengamatan. Dilihat dari persentase mortalitas serangga uji mengindikasikan bahwa isolat yang berasal dari
a b
c
d

Prayogo: Virulensi isolat entomopatogen dan pengendalian penggerek ubijalar C. formicarius 748
Tumpang sangat konsisten dan memiliki efikasi lebih tinggi dibandingkan dengan isolat dari Probolinggo maupun Bogor. Kenyataan tersebut tampak pada 4 HSA, mortalitas serangga sudah mencapai 90% dengan aplikasi kerapatan konidia 108/ml, sementara itu mortalitas dari aplikasi isolat PB 2 maupun BGR 2 masing-masing hanya 71% dan 75%. Namun pengamatan lebih lanjut pada 7 HSA, aplikasi menggunakan isolat B. bassiana PB 2 mampu menandingi efikasi isolat TMP 2 karena secara statistik tidak berbeda nyata.
Pengendalian C. formicarius menggunakan kerapatan konidia B. bassiana 106 dan 107/ml masih dinilai kurang optimal dalam membunuh serangga karena mortalitas serangga uji yang diperoleh masih lebih tinggi apabila menggunakan kerapatan konidia 108/ml. Kenyataan ini terjadi pada ketiga isolat B. bassiana yang dinyatakan virulen hasil penelitian dari tahap pertama yang digunakan. Hasil penelitian ini mengindikasikan bahwa kedua isolat B. bassiana PB 2 dan TMP 2 mempunyai peluang yang sama besarnya dapat digunakan sebagai salah satu agens hayati untuk mengendalikan C. formicarius terutama menggunakan kerapatan konidia 108/ml.
Tabel 3. Mortalitas imago C. formicarius yang dikendalikan menggunakan tiga isolat cendawan entomopatogen B. bassiana dengan kerapatan konidia berbeda.
Mortalitas imago C. formicarius pada ke..n HSA (%)* Kerapatan konidia B. bassiana per ml 3 4 5 6 7 BGR 2 106 11,67 f 23,33 h 23,33 gh 30,00 g 36,67 g 107 30,00 d 41,67 e 41,67 e 53,33 d 60,00 d 108 60,00 b 71,67 c 71,67 c 86,67 b 95,00 b PB 2 106 10,00 f 21,67 i 21,67 h 31,67 g 36,67 g 107 18,33 e 30,00 f 30,00 f 43,33 e 51,67 e 108 61,67 b 75,00 b 75,00 b 85,00 b 98,33 a TMP 2 106 16,67 e 25,00 g 25,00 g 35,00 f 40,00 f 107 38,33 c 51,67 d 51,67 d 68,33 c 76,67 c 108 76,67 a 90,00 a 90,00 a 93,33 a 98,33 a Kontrol (air) 0,00 i 0,00 i 0,00 i 2,00 h 2,00 h DMRT (0,05) 1,64 1,65 1,69 1,88 1,81 KK (%) 24,55 29,77 22,17 19,28 18,83
* Angka-angka selajur yang diikuti oleh huruf yang sama, tidak berbeda nyata pada uji DMRT taraf 5%. Keterangan: BGR 2 (Cisarua, Bogor), PB (Muneng, Probolinggo) dan TMP 2 (Pakiskembar, Tumpang).
Persentase kerusakan ubijalar akibat C. formicarius yang dikendalikan menggunakan cendawan B. bassiana dengan
perbedaan kerapatan konidia Persentase kerusakan umbi dinilai dari besarnya umbi yang tergerek oleh larva C.
formicarius yang sudah terbentuk dari telur yang dihasilkan oleh imago yang sudah terinfeksi konidia B. bassiana sebagai perlakuan. Persentase kerusakan umbi diamati pada hari ke- 21 setelah aplikasi cendawan B. bassiana. Hasil pengamatan menunjukkan bahwa semakin tinggi kerapatan konidia yang diaplikasikan pada tubuh C. formicarius, semakin rendah tingkat kerusakan umbi yang terjadi. Aplikasi suspensi konidia B.
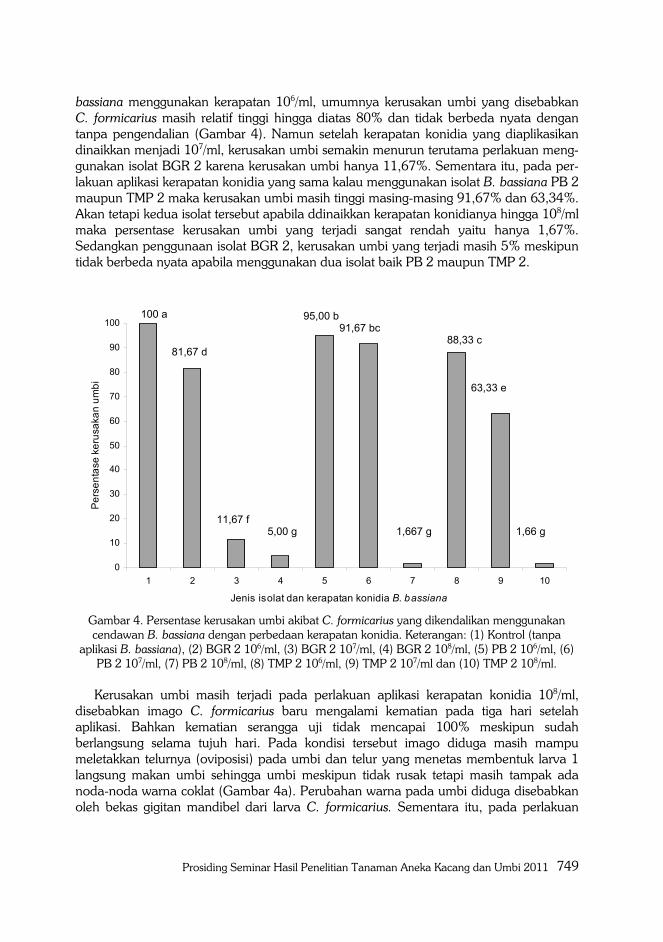
Prosiding Seminar Hasil Penelitian Tanaman Aneka Kacang dan Umbi 2011 749
bassiana menggunakan kerapatan 106/ml, umumnya kerusakan umbi yang disebabkan C. formicarius masih relatif tinggi hingga diatas 80% dan tidak berbeda nyata dengan tanpa pengendalian (Gambar 4). Namun setelah kerapatan konidia yang diaplikasikan dinaikkan menjadi 107/ml, kerusakan umbi semakin menurun terutama perlakuan meng-gunakan isolat BGR 2 karena kerusakan umbi hanya 11,67%. Sementara itu, pada per-lakuan aplikasi kerapatan konidia yang sama kalau menggunakan isolat B. bassiana PB 2 maupun TMP 2 maka kerusakan umbi masih tinggi masing-masing 91,67% dan 63,34%. Akan tetapi kedua isolat tersebut apabila ddinaikkan kerapatan konidianya hingga 108/ml maka persentase kerusakan umbi yang terjadi sangat rendah yaitu hanya 1,67%. Sedangkan penggunaan isolat BGR 2, kerusakan umbi yang terjadi masih 5% meskipun tidak berbeda nyata apabila menggunakan dua isolat baik PB 2 maupun TMP 2.
0
10
20
30
40
50
60
70
80
90
100
1 2 3 4 5 6 7 8 9 10
Jenis isolat dan kerapatan konidia B. bassiana
Per
sent
ase
keru
saka
n um
bi
Gambar 4. Persentase kerusakan umbi akibat C. formicarius yang dikendalikan menggunakan cendawan B. bassiana dengan perbedaan kerapatan konidia. Keterangan: (1) Kontrol (tanpa
aplikasi B. bassiana), (2) BGR 2 106/ml, (3) BGR 2 107/ml, (4) BGR 2 108/ml, (5) PB 2 106/ml, (6) PB 2 107/ml, (7) PB 2 108/ml, (8) TMP 2 106/ml, (9) TMP 2 107/ml dan (10) TMP 2 108/ml. Kerusakan umbi masih terjadi pada perlakuan aplikasi kerapatan konidia 108/ml,
disebabkan imago C. formicarius baru mengalami kematian pada tiga hari setelah aplikasi. Bahkan kematian serangga uji tidak mencapai 100% meskipun sudah berlangsung selama tujuh hari. Pada kondisi tersebut imago diduga masih mampu meletakkan telurnya (oviposisi) pada umbi dan telur yang menetas membentuk larva 1 langsung makan umbi sehingga umbi meskipun tidak rusak tetapi masih tampak ada noda-noda warna coklat (Gambar 4a). Perubahan warna pada umbi diduga disebabkan oleh bekas gigitan mandibel dari larva C. formicarius. Sementara itu, pada perlakuan
100 a
81,67 d
11,67 f 5,00 g
95,00 b 91,67 bc
1,667 g
88,33 c
63,33 e
1,66 g

Prayogo: Virulensi isolat entomopatogen dan pengendalian penggerek ubijalar C. formicarius 750
kerapatan konidia B. bassiana 107/ml (BGR 2, PB 2 dan TMP 2), kerusakan umbi tampak lebih parah (Gambar 4b) dibandingkan dengan perlakuan kerapatan konidia 108/ml. Namun, pada perlakuan kerapatan konidia 106/ml, diduga jumlah telur yang dihasilkan C. formicarius juga masih banyak sehingga larva yang hidup dan merusak umbi juga lebih banyak dibandingkan perlakuan kerapatan konidia yang lebih tinggi (Gambar 4c & 4d).
Gambar 4. Persentase kerusakan umbi akibat tergerek C. formicarius yang diaplikasi menggunakan B. bassiana isolat TMP 2. Aplikasi konidia 108/ml (a), aplikasi
konidia 107/ml (b), aplikasi konidia 106/ml (c) dan kontrol (d).
Capinera (2009) menyatakan bahwa telur-telur yang dihasilkan dalam rentang lima sampai enam hari akan menetas dan terbentuk larva instar 1 selanjutnya larva langsung makan dan merusak umbi. Dilaporkan Capinera lebih lanjut bahwa C. formicarius yang merusak umbi adalah hanya stadia larva saja, sedangkan stadia stadia imago menyerang organ batang maupun daun. Hasil pembelahan umbi yang rusak tergerek C. formicarius pada perlakuan kerapatan konidia 106/ml, hanya ditemukan larva yang aktif makan umbi dan tidak ditemukan imago (Gambar 5). Hasil pengamatan pada kerusakan umbi yang tergerek oleh C. formicarius tampak bahwa penggunaan ketiga isolat B. bassiana, baik isolat dari BGR 2, PB 2 maupun TMP 2 dapat melindungi umbi dengan baik apabila menggunakan kerapatan konidia 108/ml. Namun, untuk mempertahankan umbi
c d
a b

Prosiding Seminar Hasil Penelitian Tanaman Aneka Kacang dan Umbi 2011 751
dalam keadaan baik terbebas dari gerekan C. formicarius maka dianjurkan untuk aplikasi menggunakan isolat PB 2 atau TMP 2 dengan kerapatan konidia 108/ml. Hal ini disebabkan kerusakan umbi yang terjadi sangat rendah dibandingkan dengan isolat BGR 2. Umbi yang rusak akibat gerekan C. formicarius meskipun hanya dalam jumlah kecil dapat mengakibatkan rendahnya kualitas umbi sehingga tidak layak untuk dikonsumsi. Umbi yang tergerek C. formicarius dapat menghasilkan senyawa terpen yang dapat menyebabkan toksik bagi yang mengkonsumsi. Oleh karena itu, keselamatan umbi perlu mendapat perhatian lebih agar tidak mengalami kerusakan akibat C. formicarius. Ditinjau dari efikasi cendawan B. bassiana dalam mengendalikan C. formicarius dan perlindungan kerusakan umbi, maka ketiga jenis isolat cendawan tersebut berpeluang besar dapat dikembangkan dalam PHT ubijalar sebagai alternatif pengganti insektisida kimia.
Gambar 5. Larva instar II C. formicarius yang ditemukan pada umbi yang rusak pada perlakuan aplikasi kerapatan konidia B. bassiana 106/ml.
KESIMPULAN Hasil penelitian ini dapat disimpulkan sebagai berikut:
1. Dari lima belas isolat cendawan B. bassiana yang diuji diperoleh tiga isolat yang memiliki virulensi cukup tinggi karena mampu membunuh C. formicarius hingga 100%, yaitu B. bassiana yang diperoleh dari Bogor (BGR 2), Probolinggo (PB 2) dan Tumpang (TMP 2).
2. Isolat B. bassiana yang memiliki virulensi tinggi ditandai dengan pertumbuhan miselium lebih cepat dan lebih tebal dalam mengolonisasi tubuh C. formicarius dengan rentang waktu yang sangat singkat dibandingkan isolat yang avirulen.
3. B. bassiana isolat PB 2 dan TMP 2 dengan kerapatan konidia 108/ml lebih efektif untuk mengendalikan C. formicarius dan menekan kerusakan umbi dibandingkan dengan isolat BGR 2.
SARAN

Prayogo: Virulensi isolat entomopatogen dan pengendalian penggerek ubijalar C. formicarius 752
Masih diperlukan kajian penelitian metode aplikasi dan waktu aplikasi cendawan B. bassiana yang tepat di lapangan untuk pengendalian C. formicarius.
DAFTAR PUSTAKA Austin DF. 1991. Association between the plant family Convolvulaceae and Cylas formicarius
weevil. In: Jansen RK and KV. Raman (eds.) Sweet Potato Pest Management: A Global Perspective. Westview Press, Boulder. p.45-47.
Berretta MF, Lecuona LE, Zandomeni RO Grau O. 1998. Genotyping isolates of the entomo-pathogenic fungus Beauveria bassiana by RAPD with fluorescenst labels. J Invertebr Pathol 71: 145-150.
Capinera JL. 2009. Sweet potato weevil. Departement of Entomology and Nematology, Devision of Plant Industry. http://entnemdept.ufl.edu/creatures/ve/potato/ sweetpotato_ weevil.htm [12 Feb 2012].
Chun C, Mingguang F. 2004. Observation on the initial inoculums source and dissemination of entomophthorales caused epizootics in populations of cereal aphids. Sci China C Life Sci 47(1): 38-43.
Ekesi S. 2001. Pathogenicity and antifeedant activity of entomopathogenic Hyphomycetes to the cowpea leaf beetle Ootheca mutabilis Shalberg. Insect Sci Appl 21(1): 55-60.
Fatiha L, Ali S, Ren S, Rizal M. 2007. Biological characteristics and pathogenicity of Vertici-llium lecanii against Bemisia tabaci (Homoptera: Aleyrodidae) on eggplant. J Pak Entomol 29(2): 63-73.
Ganga-Visalakshy PN, Manoj-Kumar A, Krisnamoorthy A. 2004. Epizootics of fungal patho-gen Verticillium lecanii (Zimmermann) on Thrips palmi Karny. Insect Environ 10(3): 134-135.
Jansson RK, Hunsbeerger ABG, Lecrone SH, O’Hair SK. 1990. Seasonal abundance, population growth and within-plant distribution of sweet potato weevil (Coleoptera: Curculionidae) on sweet potato in Southern Florida. Environ Entomol 19(2): 313-321.
Jansson RK. 1991. Biological control of Cylas spp. Pages 169-201. In: Sweet potato pest management: A global pespective Ed. by Jasson, R.K.& Raman, K.V. Sweet Potato Pest Management: A Global Perspective. Westview Press, Boulder.
Kassa A. 2003. Development and testing of mycoinsecticides based on submerged spores and aerial conidia of the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae (Deuteromycotina: Hyphomycetes) for control of locust grasshoppers and storage pests. [dissertation]. Gottingen.
Klingen I, Hajek A, Meadow R, Renwick JAA. 2002. Effect of brassicaceous plants on the survival and infectivity of insect pathogenic fungi. Biol Contr 47: 411-425.
Komi K. 2000. Eradication of sweet potato weevil Cylas formicarius (Fabricius) from Muroto city, Kochi, Japan, Food & Fertilizer Technology Center. http://www.ffc.agnet.org/ library/abstract/eb493c.html [13 Agu 2011].
Kumano N, Kohama T, Ohno S. 2007. Effects of irradiation on dispersal ability of male sweet potato weevils (Coleoptera: Curculionidae) in the field. J of Econ Entomol 100(3): 730-736.
Lagnaouli A, Cisneros F, Alcazar J, Molares F. 2000. A sustainable pest management strategy fpr sweet potato weevil in Cuba: A Succes Story. Food & Fertilizer Technology Center (FFTC). http://www.fftc.agnet.org/library/ abstract/ eb493a. html. [12 Agu 2011]

Prosiding Seminar Hasil Penelitian Tanaman Aneka Kacang dan Umbi 2011 753
Lerche S, Meyer U, Sermann H, Buether S. 2004. Dissemination of the entomopathogenic fungus Verticillium lecanii (Zimmermann) Viegas (Deuteromycotina: Hyphomecetes) in population of Frankliniella occidentalis Pergande, 1895 (Thysanoptera: Thripidae). Commun Agric Appl Biol Sci 69(3): 195-200.
Masaki S, Mizuno T, Toubaru K, Sasaki M, Kobayashi K, Kaneda M. 2002. Occurrence of Cylas formicarius and Euscepes postfasciatus in mountainous areas of Okinawa and islands (Japanese). Res Bull of The Plant Protect Service Japan 38: 39-43.
Miyaji K, Tanaka T. 1998. Geographic distribution of the sweet potato weevil Cylas formicarius, Euscepes postfasciatus Fairmaire (Coleoptera: Curculionidae) in the Amami islands (Japanese). Proccedings of The Association for Plant Protect of Kyushu 44: 88-92.
Nonci N, Sriwidodo. 1994. Pengaruh pengendalian Cylas formicarius pada ubijalar terhadap kerusakan ubi pada penyimpanan. Laporan Hasil Penelitian Jagung dan Ubi. (3): 89-97. Balai Penelitian Tanaman Pangan Maros.
Nonci N. 2005. Bioekologi dan pengendalian kumbang Cylas formicarius Fabricius (Coleoptera: Curculionidae). Jurnal Litbang Pertanian 24(2): 65-69.
Ondiaka S, Maniania NK, Nyamasyo GHN, Nderitu JH. 2008. Virulence of the entomopathogenic fungi Beauveria bassiana and Metarhizium anisopliae to sweet potato weevil Cylas formicarius effects on fecundity and egg viability. Ann of Appl Biol 153(1): 41-48.
Ohara K, Mishima S, Sakamoto T, Kaku K. 2000. Distribution of the sweet potato weevil Cylas formicarius (Fabricius) in Amami-Oshima island (Japanese). Res Bull od The Plant Protect Service Japan 36: 67-70.
Popowska-Nowak E, Bienkowski P, Bajan C, Tyrawaska D. 2000. Influence of some heavy metal ions on biological activity of two strains of entomopathogenic fungus Paecilomyces farinosus. Chem Ins Ekol 7(11): 1121-1128.
Prayogo, Y. 2009. Kajian cendawan entomopatogen entomopatogen Lecanicillium lecanii (Zimm.) (Viegas) Zare & Gams untuk menekan perkembangan telur hama pengisap polong kedelai Riptortus linearis (F.) (Hemiptera: Alydidae). [disertasi]. Sekolah Pascasarjana, Departemen Proteksi, Institut Pertanian Bogor.
Purlong MJ, Pell JK. 2001. Horizontal transmission of entomopathogenic fungi by the diamondback moth. Biol Contr 22: 288-299.
Ropek D, Para A. 2002. The effect of heavy metal ions and their complexons upon the growth, sporulation, and pathogenicity of the entomopathogenic fungus Verticillium lecanii. J Invertebr Pathol 79: 123-125.
Rosfiansyah. 2009. Pengaruh aplikasi Beauveria bassiana (Bals.) Vuillemin dan Heteror-habditis sp. terhadap serangan hama ubijalar Cylas formicarius (Fabricius) (Coleoptera: Brentidae). (tesis) Sekolah Pascasarjana, Departemen Proteksi Tanaman, Institut Pertanian Bogor.
Sato K, Uritani I, Saito T. 1982. Properties of terpene-inducing factor extracted from adults of the sweet potato weevil Cylas formicarius Fabricius (Coleoptera: Brethidae). Appl Entomol Zool 17(3): 368-374.
Sudirman LI, Prayogo Y, Yunimar, Ginting S. 2008. Effect of leaf litters and soils on viability of entomopathogenic fungi Beauveria bassiana (Bals.) Vuill. Hayati J Biosci 15(3): 93-98.

Prayogo: Virulensi isolat entomopatogen dan pengendalian penggerek ubijalar C. formicarius 754
Sugimoto M, Koike M, Nagao H, Okumura K, Tani M. 2003a. Genetic diversity of the entomopathogen Verticillium lecanii on the basis of vegetative compatibility. Phytopar 31: 450-457.
Sugimoto M, Koike M, Hiyana N, Nagao H. 2003b. Genetic, morphological and virulence characterization on the entomopathogenic fungus Verticillium lecanii. J Invertebr Pathol 82: 176-187.
Tarafdar J, Sarkar MA. 2005. Managing sweet potato weevil Cylas formicarius in West Bengal, India by some chemicals, bioproducts and sex pheromone traps. Acta Hort. Proc. of the 2nd Internat. Symp. on sweet potato and cassava: (Inovative technologies for commercialization) Kuala Lumpur, Malaysia. June 14-17, 2005. pp.189-196.
Trizelia. 2005. Cendawan entomopatohen Beauveria bassiana (Bals.) Vuill. (Deuteromy-cotina: Hyphomycetes): keragaman genetik, karakterisasi fisiologi, dan virulensinya ter-hadap Crocidolomia pavonana (F.) (Lepidoptera: Pyralidae). [disertasi]. Departemen Proteksi Tanaman, Institut Pertanian Bogor.
Varela A, Morales E. 1996. Characterization of some Beauveria bassiana isolates and their virulence toward the coffe berry borer Hypothenemus hampei. J Invertebr Pathol 67: 147-152.
Wagner BL, Lewis LC. 2000. Colonization of corn Zea maize by the entomopathogenic fungus Beauveria bassiana. Appl Environ Microbiol 66(8): 3468-3473.
Wraight SP, Jackson MA, de-Kock SL. 2001. Production, stabilization, and formulation of fungal biocontrol agents. In: Butt TM, Jackson C, Magan N, editor. Fungi as Biocontrol Agents. United Kingdom: CABI Publ.
Yamaguchi T, Setokuchi O, Miyaji K. 2000. Development and adult survival of the sweet potato weevil, Cylas formicarius (Fabricius) (Coleoptera: Curculionidae), during winter on Amami-Oshima island, Japan. Appl Entomol & Zool 35(4): 451-458.
Yasuda K. 1999. Auto-infection system for the sweet potato weevil Cylas formicarius (Fabri-cius) (Coleoptera: Curculionidae) with entomopathogenic fungi Beauveria bassiana using a modified sex pheromone trap in the field. Appl Entomol and Zool 34: 501-505.
Yasuda, K. 2000. Integrated pest management of Wesr Indian sweet potato weevil Euscepes postfasciatus (Fairmaire) and sweet potato weevil Cylas formicarius (Fabricius) in Oki-nawa, Japan. Food Fertilizer Technology Center (FFTC). http://www.ffc.agner. org/library/abstract/ eb493b.html [12 Agu 2011].