uji mikronuklei dengan pengeblokan … lusiyanti dkk 2011.pdf · dosis dan sifat biologik yaitu...
TRANSCRIPT

Seminar Nasional Keselamatan Kesehatan dan Lingkungan VII
Jakarta, 6-7 Juli 2011
PTKMR-BATAN, BAPETEN, KEMENKES-RI dan Pusarpedal-KLH 57
UJI MIKRONUKLEI DENGAN PENGEBLOKAN SITOKENESIS
PADA LIMFOSIT DAN APLIKASINYA SEBAGAI
BIODOSIMETRI RADIASI
Yanti Lusiyanti dan Zubaidah Alatas
Pusat Teknologi Keselamatan dan Metrologi Radiasi - BATAN
ABSTRAK
UJI MIKRONUKLEI DENGAN PENGEBLOKAN SITOKENESIS PADA LIMFOSIT DAN
APLIKASINYA SEBAGAI BIODOSIMETRI RADIASI. Mikronuklei adalah salah satu indikasi
kerusakan struktur pada kromosom akibat radiasi, yang dapat diamati dari sel dengan dua inti (binukleat),
dengan cara memblok proses pembelahan pada tahap sitokinesis menggunakan sitokalasin B yang dikenal
dengan Cytokinesis Block (CB). Bila metoda uji mikronuklei akan dijadikan sebagai biodosimetri radiasi,
maka perlu diketahui kurva respon dosis pada kisaran luas dari berbagai kualitas radiasi. Kurva respon
mikronuklei yang diinduksi oleh radiasi LET rendah, antara lain sinar-X, sinar γ dan partikel β
diperlihatkan dengan model persamaan linier quadratik, sedangkan radiasi LET tinggi hanya menghasilkan
persamaan linier. Variasi antar individu dalam pembentukkan mikronukleus sampai kini masih
dipertanyakan, karena data yang diperoleh dari beberapa penelitian belum menunjukkan hasil yang konstan
terutama untuk frekuensi mikronukleus latar. Untuk menerapkan uji mikronukleus sebagai biodosimetri
radiasi, diperlukan kalibrasi kurva mikronukleus pada individu pra pajanan terutama untuk dosis rendah. Kata kunci : mikronuklei, pengeblokan sitokinensis, radiasi, biodosimetri
ABSTRACT
MICRONUCLEI ASSAY USING CYTOKINESIS BLOCK IN LYMPHOCYTES AND ITS
APPLICATION AS RADIATION BIODOSIMETRY.Micronuclei is one of structural damage indicators
caused by ionizing radiation that observed on cells with binucleate by blocking cell proliferation at
cytokinesis stage using cytochalasin B, known as cytokinesis Block (CB). If micronuclei assay is used as a
biological dosimeter, it is essensial to have dose-respon curve on a wide range of radiation qualities. The
dose respons curve of micronuclei for low LET radiation i,e X-rays, γ rays and β particle represented in the
linear quadratiq model and for high LET i.e neutron and ά particle represented in linear model. Individual
variation in micronuclei is unclear because the uncertainties are predominately for the base line
miccronucleus. One of the mayor limitation in measuring exposure to low doses may be the interindividual
variation in radiosensitivity, and the practical implementation of the micronucleus assay for radiation
biodosimetry may be limited by a need to perform individual pre-exposure calibrations.
Keywords : micronuclei, cytokinetic blocking, radiation, biodosimetry

Seminar Nasional Keselamatan Kesehatan dan Lingkungan VII
Jakarta, 6-7 Juli 2011
PTKMR-BATAN, BAPETEN, KEMENKES-RI dan Pusarpedal-KLH 58
I. PENDAHULUAN
Pengukuran biodosimetri berkaitan
dengan besarnya dosis serap radiasi yang
diterima dan berkontribusi serta terhadap
risiko kesehatan yang diakibatkan oleh
paparan radiasi. Prinsip biodosimetri adalah
menentukan dosis serap radiasi dengan
mengukur perubahan yang terjadi pada
materi biologi akibat paparan radiasi pada
tubuh manusia. Salah satu yang paling utama
dari biodosimetri adalah dalam kasus
kecelakaan radiasi yang terjadi pada individu
yang tidak menggunakan dosimeter fisik.
Kadangkala metode dosimetri fisik harus
dilengkapi atau didukung oleh uji biologik,
sebagai contoh terjadinya pajanan sebagian
tubuh (parsial) dengan dosimetri fisik diluar
area radiasi.
Cek silang dosis yang diukur secara
fisik diperlukan pada kondisi tertentu. Akan
tetapi, jika dosis ditentukan secara biologik,
variabilitas biologik akan mempengaruhinya,
karena diyakini untuk individu yang
radiosensitif akan memiliki efek yang lebih
besar pada materi biologiknya daripada
ukuran rata-rata. Biodosimetri didasarkan
pada marker biologi atau biomarker yang
diinduksi oleh paparan radiasi dan dapat
diterapkan untuk mengestimasi dosis ketika
dosimetri fisik tidak tersedia, atau untuk
melengkapi dosimetri fisik. Biomarker
radiasi yang telah secara luas dikenal dan
diaplikasikan sebagai dosimetri biologi atau
biodosimetri radiasi adalah aberasi
kromosom menggunakan metode sitogenetik
untuk mengetahui frekuensi kromosom
bentuk disentrik, asentrik fragmen dan cincin
(ring). Analisis aberasi kromosom disentrik
telah dibuktikan sebagai gold standar (baku
standar) yang sensitif untuk mengetahui
kerusakan sel akibat radiasi dan telah
dijadikan sebagai biodosimetri radiasi [1].
Metode biodosimetri yang cepat sangat
diperlukan untuk mengkaji efek radiasi tunda
(jangka panjang). Selain teknik analisis
aberasi kromosom pada limfosit, teknik uji
lain yang juga dapat digunakan dalam
biodosimetri adalah uji mikronukleus (MN).
Pengujian MN dari limfosit perifer pertama
kali diperkenalkan oleh Countryman and
Heddle [2]. Pada metode awal hanya
dilakukan pada sel yang telah selesai
membelah secara in-vitro. Suatu pendekatan
telah dikembangkan dengan menggunakan
inhibitor cytokenesis yaitu cytochalasin-B.
Feneh dan Morley [3] telah menerapkan
Cytocalasin–B yang mampu memperlihatkan
bahwa sel yang telah membelah dapat
diakumulasi dan dikenali sebagai sel
binukleat (dua inti) dan MN dapat secara
spesifik dan efisien terlihat di dekat sel
binucleat sementara sel mononuclear yang
tidak membelah tidak mampu mengekspresi-
kan adanya MN secara in vitro.
MN telah digunakan sejak tahun 1937
sebagai indikator pajanan genotoksik

Seminar Nasional Keselamatan Kesehatan dan Lingkungan VII
Jakarta, 6-7 Juli 2011
PTKMR-BATAN, BAPETEN, KEMENKES-RI dan Pusarpedal-KLH 59
berdasarkan pada studi radiasi oleh Brenneke
dan Mather. Sejak saat itu banyak studi lain
dilakukan pada sel tumbuhan, hewan dan
manusia, baik secara in vitro maupun in vivo.
Penelitian frekuensi MN juga mendukung
asumsi bahwa MN ini merupakan produk
awal proses karsinogenik pada manusia
Bila induksi MN akan digunakan
untuk biodosimetri radiasi, maka sangat perlu
membuat suatu kurva data respon-dosis,
untuk kisaran yang luas dari berbagai kualitas
radiasi, terutama yang banyak digunakan
dalam penelitian lingkungan, dan sangat
sesuai untuk proteksi radiasi. Di samping itu
sangat penting untuk memperoleh konsistensi
terutama antara laboratorium acuan dengan
laboratorium lainnya. Faktor yang berperan
terhadap pembentukkan MN antara lain
adalah faktor fisik seperti LET, dosis dan laju
dosis dan sifat biologik yaitu variasi antar
individu, yang dipengaruhi oleh umur, jenis
kelamin dan kinetika sel. Dalam makalah ini
dibahas mengenai uji MN dengan metode
cytokinesis block (CB) dan kemungkinan
implementasinya bila digunakan sebagai
biodosimetri radiasi.
II. MIKRONUKLEI
Paparan radiasi dapat menginduksi
terjadinya pembentukan fragmen kromosom
Asentrik (kromosom tanpa centromer dan
malsegregasi kromosom utuh (whlole
kromosom). Fragmen kromosom asentrik dan
whole kromosom yang tidak mampu
berinteraksi dengan benang spindel lag
behind anaphase., sehingga akhirnya fragmen
tersebut tidak ikut ke dalam nuklei anak
utama. Potongan fragmen kromosom atau
kromosom tersebut membentuk nuckleus
kecil yang terpisah dan dinamakan
mikronuklei, dengan demikian mikronuklei
merupakan materi nukleus (DNA)) terlihat
sebagai lingkaran kecil dalam sitoplasma di
luar nukleus, dengan struktur dan intensitas
warna serupa dengan nukleus Gambar 1.
Mikronuklei terbentuk dari fragmen asentrik
yang gagal bergabung dengan sel anak
selama proses pembelahan sel. Dapat juga
terbentuk dari sebuah kromosom yang
tertinggal, atau tidak terbawa dalam proses
mitosis, atau terjadi akibat konfigurasi
kromosom yang kompleks, pada waktu
proses anafase. Namun demikian terdapat
bukti kuat yang menunjukkan bahwa radiasi
dapat menginduksi pembentukkan
mikronukleus adalah terutama berasal dari
fragmen asentrik [1,2].
Gambar 1. Sel binukleat yang
memiliki 1 mikronuklei [1]

Seminar Nasional Keselamatan Kesehatan dan Lingkungan VII
Jakarta, 6-7 Juli 2011
PTKMR-BATAN, BAPETEN, KEMENKES-RI dan Pusarpedal-KLH 60
Kriteria mikronuklei antara lain ;
diameter kurang dari seperlima diameter
nukleus (10µm), terletak dalam sitoplasma
dan di luar nukleus, tidak ada kontak dengan
nukleus [1]. Keterangan lain menyatakan
bahwa ukuran mikronukleus yang diinduksi
radiasi cenderung berukuran antara 6-12 µm.
Kira-kira 80% mikronukleus yang diinduksi
oleh sinar gamma mengandung DNA sekitar
6% atau kurang dari nukleus interfase,
menandakan asal mikronukleus berasal dari
fragmen asentrik. Pada individu normal atau
frekuensi mikronuklei untuk 1000 sel
binukleate adalah 3- 30 [1,4].
Beberapa keunggulan dari uji MN
adalah sbb. :
1. Dapat dikombinasi dengan deteksi mutasi
kromosom dan genom sekaligus.
2. Dapat membedakan antara klastogen dan
aneugen.
3. Ada kemungkinan mendeteksi apoptosis
atau nekrosis secara bersamaan.
4. Dapat digunakan untuk banyak jenis sel,
cepat, murah, dan sederhana.
5. Ada kemungkinan otomatisasi dan unggul
secara statistik.
6. Dapat membedakan antara sel yang
sedang membelah dan tidak membelah.
7. Mampu mendeteksi jembatan disentrik
(dicentric bridges) sebagai jembatan
nukleoplasmik dan pengkajian proliferasi
sel (persentase sel binukleat).
III. TEKNIK PENGEBLOKAN
SITOKINESIS
Mikronuklei terbentuk akibat
kerusakan struktur dari kromosom yang
terjadi pada fase G0-G1 dari siklus sel,
sehingga mikronukleus muncul setelah sel
mengalami pembelahan inti.
Pengujian teknik scoring mikronuklei dalam
sebuah sel, yang telah diblok pada limfosit
tepi, digunakan sebagai metoda alternatif
realistis untuk mengetahui secara kuantitatif
adanya kerusakan kromosom akibat radiasi,
selain penghitungan kromosom disentrik
[4,5]. Metoda CB yaitu menggunakan
penambahan zat Sitokalasin-B terhadap
kultur limfosit, berfungsi untuk memblok
proses sitokinesis sehingga sel berada pada
tingkat pembelahan sel binukleat (sel dengan
dua inti), dan mikronklei yang terbentuk akan
teramati pada sel binukleat tersebut [3,6].
Teknik pengeblokan yang
dimaksudkan untuk memperoleh mikronuklei
ini telah diperkenalkan lebih dari 30 tahun
lalu, tepatnya tahun 1975. Proses atau
mekanisme pembentukan mikronuklei
dijelaskan dalam Gambar 2.

Seminar Nasional Keselamatan Kesehatan dan Lingkungan VII
Jakarta, 6-7 Juli 2011
PTKMR-BATAN, BAPETEN, KEMENKES-RI dan Pusarpedal-KLH 61
Gambar 2. Proses pembentukan MN yang diinduksi dengan sitochalasin-B [1].
Pengujian mikronuklei telah digunakan
sejak tahun 1937 sebagai indikator pajanan
genotoksik berdasarkan pada studi radiasi
oleh Brenneke dan Mather. Sejak saat itu
banyak studi lain dilakukan pada sel
tumbuhan, hewan dan manusia, baik secara
in vitro maupun in vivo. Penelitian frekuensi
MN juga mendukung asumsi bahwa MN ini
merupakan produk awal proses karsinogenik
pada manusia.
Penggunaan teknik mikronukleus
dalam penghitungan lebih mudah cepat dan
sel yang dapat diamati dalam jumlah banyak,
terutama apabila menggunakan sistem image
komputer otomatis. Sehingga teknik
mikronukleus memungkinkan digunakan
sebagai sebagai prosedur rutin, dan dapat
diamati pada dosis rendah antara 0,05-1 Gy,
yang merupakan rentang batas dosis terendah
untuk mendeteksi adanya aberasi kromosom
[5,7,8]. Keunggulan lain dari uji ini adalah
waktu prosesnya dimana mikronuklei dapat
dihitung dengan cepat dan sangat sesuai
untuk deteksi awal pada sejumlah besar
korban kecelakaan radiasi. Pengujian MN
dengan Metode CB ini juga telah dilakukan
terhadap Para pekerja radiasi di BATAN,
dengan hasil yang ditemukan frekuensi MN
masih berada dalam kisaran frekuensi latar
untuk mikronuklei [9].
Beberapa kriteria dari sel BNC yang
layak untuk dihitung frekuensi
mikronukleinya harus memenuhi ketentuan
sebagai berikut.
- Sel harus dalam bentuk binukleat (terdiri
dari dua nukleus).
- Kedua inti dalam binukleat sel harus
dalam kondisi bersentuhan dengan
membran inti dan berada dalam satu
lingkaran sitoplasma yang sama.
- Kedua inti dalam sel binukleat harus
memiliki ukuran, penyerapan warna dan
intensitas pewarnaan yang sama.
SITOCHALASIN B

Seminar Nasional Keselamatan Kesehatan dan Lingkungan VII
Jakarta, 6-7 Juli 2011
PTKMR-BATAN, BAPETEN, KEMENKES-RI dan Pusarpedal-KLH 62
- Kedua inti dalam sel binukleat mungkin
tidak dihubungkan atau mungkin
menempel satu atau lebih oleh jembatan
nukleoplasma dan ukurannya tidak kurang
lebih 1/4 diameter dari inti.
- Kedua inti utama dalam sel binukleat
mungkin bersentuhan satu sama lain
namun idealnya harus tidak overlap satu
sama lain. Sel dengan kondisis nuklei
yang overlaping dapat dihitung hanya
apabila lingkaran inti dari inti yang lain
dapat dibedakan.
- Lingkaran sitoplasma atau membran
dari selbinukleat harus berinteraksi dan
secara jelas dapat dibedakan dari
lingkaran sitoplasmic dari sel sekitarnya.
Akan tetapi uji ini memiliki
keterbatasan terutama oleh adanya frekuensi
background yang lebih besar dan lebih
bervariasi dibandingkan dengan disentrik. Uji
MN juga tidak mampu mendeteksi semua
aberasi kromosom struktural (hanya
asentrik), membutuhkan pembelahan sel
untuk ekspresi MN, ada kemungkinan
interferensi oleh sitochalasin-B seperti
spindle poison, ada kemungkinan interferensi
dengan penghambat lain dari sitokinesis, dan
sitotoksisitas sitochalasin-B itu sendiri
bervariasi antar-jenis sel atau bahkan antar-
sub jenis dari jenis sel yang sama [1].
IV. INDUKSI MIKRONUKLEI OLEH
RADIASI
Teknik pengujian MN dengan
metode CB menggunakan sitochalasin-B
untuk menginduksi mikronuklei juga menjadi
andalan banyak peneliti dalam menentukan
dosis radiasi. Mikronuklei dalam sel dua inti
terbentuk selama transisi metafase-anafase
ketika seluruh kromosom hilang (kejadian
aneugenik) atau fragment kromosom asentrik
setelah terjadi patahan kromosom (kejadian
clastogenik) yang tidak bergabung ke dalam
inti sel anak. Karena lebih sederhana, lebih
cepat, tidak mahal serta bentuk mikronuklei
yang sederhana, mudah dikenali dan ada
potensi untuk otomatisasi dengan sitometri
maka teknik pengeblokan sitokinesis ini juga
diandalkan oleh para peneliti]. Meninjau
awal mula MN muncul, telah diketahui
dengan baik bahwa sebagian besar MN
akibat radiasi terutama berasal dari fragmen
kromosom asentrik yang merupakan hasil
patahan kromosom. Sejumlah kecil MN
akibat radiasi dapat berasal dari kromosom
utuh yang gagal (lag) setelah anaphase
disebabkan karena beberapa kelainan pada
tingkat spindle atau protein kinetochore.
Gambaran mikronuklei yang diinduksi oleh
radiasi gamma 1 Gy terlihat pada Gambar 3.

Seminar Nasional Keselamatan Kesehatan dan Lingkungan VII
Jakarta, 6-7 Juli 2011
PTKMR-BATAN, BAPETEN, KEMENKES-RI dan Pusarpedal-KLH 63
Gambar 3. Contoh hasil uji mikronuklei yang diinduksi oleh radiasi sinar gamma dosis
1 Gy. Satu (kiri) dan tiga (kanan) mikronuklei yang berada di samping BNC di dalam
sitoplasma [10].
Untuk mengetahui apakah MN yang
diinduksi adalah akibat paparan iradiasi atau
senyawa kimia, para peneliti
mengembangkan teknik deteksi MN
menggunakan teknik flourescence in situ
hybridization (FISH) menggunakan pan
centromeric probe dan hasilnya diperlihatkan
dalam Gambar 4. Pengaruh radiasi dalam
menginduksi pembentukan mikronukleus
dipengaruhi oleh Linier Energi Transfer
(LET), laju dosis dan besarnya dosis radiasi.
Nilai LET adalah jumlah energi yang
terdeposit sepanjang jejak lintasan radiasi
yang dilaluinya, maka nilai LET
berhubungan erat dengan efektifitas suatu
jenis radiasi pengion, yang menyebabkan
kerusakan pada materi biologi yang
dilintasinya. Semakin besar LET, semakin
besar daya rusak radiasi tersebut pada materi
biologis yang dikenal dengan istilah Relatif
Biological Effectiveness (RBE) [9,10].
Gambar 4. Teknik deteksi MN menggunakan flourescence in situ hybridization (FISH)
menggunakan pan centromeric probe. MN hasil induksi kimia (kiri) dan radiasi
(kanan).

Seminar Nasional Keselamatan Kesehatan dan Lingkungan VII
Jakarta, 6-7 Juli 2011
PTKMR-BATAN, BAPETEN, KEMENKES-RI dan Pusarpedal-KLH 64
Perbedaan utama antara partikel alfa
sebagai radiasi LET tinggi, dengan radiasi
gamma/sinar-X sebagai radiasi LET rendah,
yaitu dalam hal distribusi energi pada
populasi sel atau jaringan yang terpajan.
Ionisasi akan terjadi pada setiap interval 100
nm atau lebih sepanjang lintasan radiasi
gamma/-X akan menembus suatu jaringan
sedalam beberapa cm, sebelum melepaskan
semua energinya. Sehingga terjadi distribusi
energi yang merata dalam jaringan, dengan
demikian dosis radiasi yang diterima oleh sel
dalam jaringan adalah sama dengan tingkatan
pajanan yang sangat rendah. Sedangkan
radiasi alfa, terjadi ionisasi setiap 0,2-0,5
nm, sehingga deposisi energi yang besar
terjadi pada satu lokasi tertentu. Umumnya
partikel alfa melintas hanya sejauh 50 μm
sebelum semua energinya habis dilepaskan
[1,11].
Pada radiasi dengan LET tinggi,
induksi mikronukleus tampak jelas, sesuai
dengan yang digambarkan yaitu untuk
partikel α dan neutron, jauh lebih efektif
dalam menginduksi pembentukan
mikronukleus dibanding sinar-X, sinar
gamma maupun partikel β. Hal tersebut
terlihat pada penelitian yang dilaporkan
bahwa induksi mikronukleus oleh neutron
pada energi 5,5 MeV dengan nilai LET 20
keV μm-1
, nilai ini hampir mendekati untuk
partikel α yaitu 20-23 keV μm-1
. Keduanya
memperlihatkan persamaan linier 0,374 ±
0,012 untuk neutron dan 0,336 ± 0,039 untuk
partikel alfa serta tidak berbeda secara
bermakna [12,8].
Sinar-X dan sinar gamma merupakan
radiasi dengan LET rendah, mempunyai
kemampuan menginduksi dengan kerusakan
yang tidak sama. Pada saat radiasi
gelombang elektromagnetik (X dan γ)
berinteraksi dengan sebuah atom, maka
keduanya akan melepaskan elektron
sekunder. Radiasi gamma dari Co-60 dengan
energi 1,1 MV mempunyai nilai LET lebih
rendah yaitu 0,2 kev/ μm dari Sinar-X 250
kV yaitu 2 kev/ μm, dengan demikian
efektifitas sinar gamma untuk merusak
materi biologi menjadi lebih rendah, sekitar
10% . Hal yang sama terjadi pada sinar-X
dengan energi 25 MV mempunyai nilai LET
0,2 kev/ μm [8,13].
Induksi mikronukleus pada LET
rendah diketahui bahwa Sinar-X cenderung
sedikit lebih efektif, dibanding sinar gamma
walaupun antara kedua kurva tampak tidak
menunjukkan perbedaan yang nyata. Hasil
penelitian lain yang dilakukan pada beberapa
kualitas radiasi berbeda, diketahui bahwa
pada dosis yang tinggi yaitu 10 Gy untuk
sinar-X dan 1 Gy untuk neutron, dapat
mempengaruhi kapasitas proliferasi pada sel
limposit yang telah distimulasi. Telah
diketahui bahwa partikel α dan neutron jauh
lebih efektif dalam menginduksi
pembentukan mikronukleus dibanding sinar-
X, sinar gamma maupun partikel β [8].
Seminar Nasional Keselamatan Kesehatan dan Lingkungan VII
Jakarta, 6-7 Juli 2011
PTKMR-BATAN, BAPETEN, KEMENKES-RI dan Pusarpedal-KLH 65
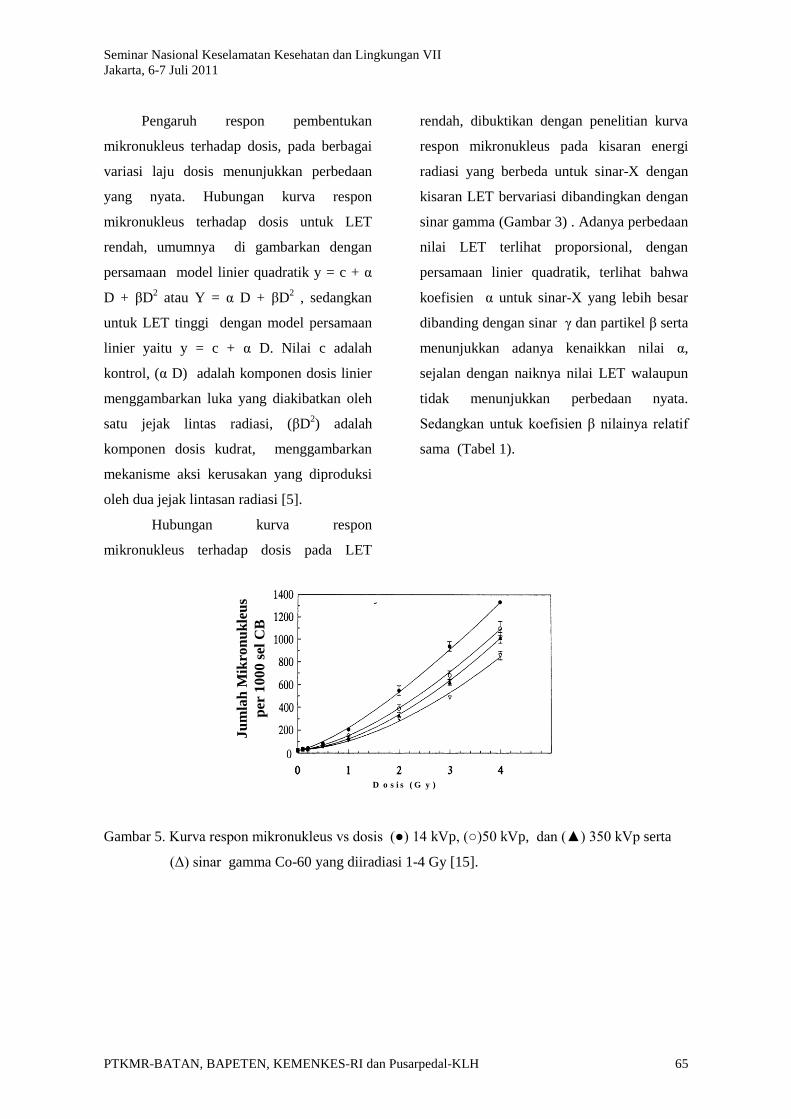
Pengaruh respon pembentukan
mikronukleus terhadap dosis, pada berbagai
variasi laju dosis menunjukkan perbedaan
yang nyata. Hubungan kurva respon
mikronukleus terhadap dosis untuk LET
rendah, umumnya di gambarkan dengan
persamaan model linier quadratik y = c + α
D + βD2 atau Y = α D + βD
2 , sedangkan
untuk LET tinggi dengan model persamaan
linier yaitu y = c + α D. Nilai c adalah
kontrol, (α D) adalah komponen dosis linier
menggambarkan luka yang diakibatkan oleh
satu jejak lintas radiasi, (βD2) adalah
komponen dosis kudrat, menggambarkan
mekanisme aksi kerusakan yang diproduksi
oleh dua jejak lintasan radiasi [5].
Hubungan kurva respon
mikronukleus terhadap dosis pada LET
rendah, dibuktikan dengan penelitian kurva
respon mikronukleus pada kisaran energi
radiasi yang berbeda untuk sinar-X dengan
kisaran LET bervariasi dibandingkan dengan
sinar gamma (Gambar 3) . Adanya perbedaan
nilai LET terlihat proporsional, dengan
persamaan linier quadratik, terlihat bahwa
koefisien α untuk sinar-X yang lebih besar
dibanding dengan sinar γ dan partikel β serta
menunjukkan adanya kenaikkan nilai α,
sejalan dengan naiknya nilai LET walaupun
tidak menunjukkan perbedaan nyata.
Sedangkan untuk koefisien β nilainya relatif
sama (Tabel 1).
D o s i s ( G y )
Gambar 5. Kurva respon mikronukleus vs dosis (●) 14 kVp, (○)50 kVp, dan (▲) 350 kVp serta
(Δ) sinar gamma Co-60 yang diiradiasi 1-4 Gy [15].
Ju
mla
h M
ikro
nu
kle
us
per
1000 s
el C
B
Seminar Nasional Keselamatan Kesehatan dan Lingkungan VII
Jakarta, 6-7 Juli 2011
PTKMR-BATAN, BAPETEN, KEMENKES-RI dan Pusarpedal-KLH 66
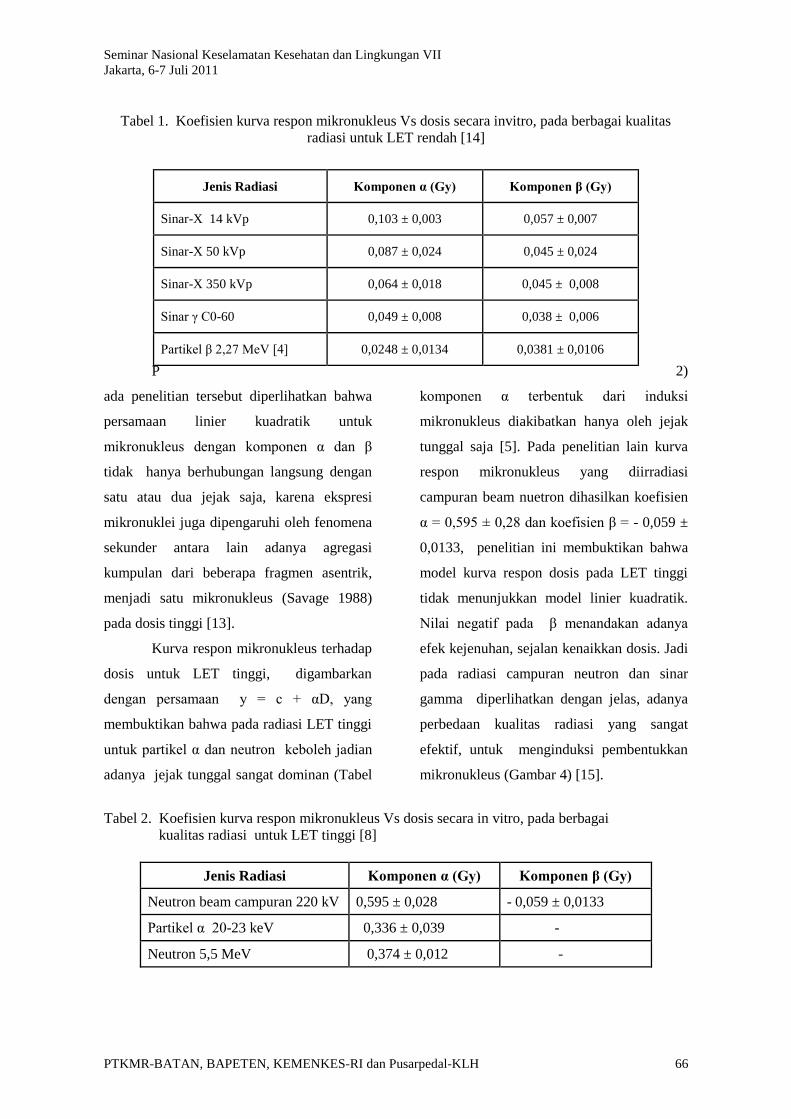
Tabel 1. Koefisien kurva respon mikronukleus Vs dosis secara invitro, pada berbagai kualitas
radiasi untuk LET rendah [14]
P
ada penelitian tersebut diperlihatkan bahwa
persamaan linier kuadratik untuk
mikronukleus dengan komponen α dan β
tidak hanya berhubungan langsung dengan
satu atau dua jejak saja, karena ekspresi
mikronuklei juga dipengaruhi oleh fenomena
sekunder antara lain adanya agregasi
kumpulan dari beberapa fragmen asentrik,
menjadi satu mikronukleus (Savage 1988)
pada dosis tinggi [13].
Kurva respon mikronukleus terhadap
dosis untuk LET tinggi, digambarkan
dengan persamaan y = c + αD, yang
membuktikan bahwa pada radiasi LET tinggi
untuk partikel α dan neutron keboleh jadian
adanya jejak tunggal sangat dominan (Tabel
2)
komponen α terbentuk dari induksi
mikronukleus diakibatkan hanya oleh jejak
tunggal saja [5]. Pada penelitian lain kurva
respon mikronukleus yang diirradiasi
campuran beam nuetron dihasilkan koefisien
α = 0,595 ± 0,28 dan koefisien β = - 0,059 ±
0,0133, penelitian ini membuktikan bahwa
model kurva respon dosis pada LET tinggi
tidak menunjukkan model linier kuadratik.
Nilai negatif pada β menandakan adanya
efek kejenuhan, sejalan kenaikkan dosis. Jadi
pada radiasi campuran neutron dan sinar
gamma diperlihatkan dengan jelas, adanya
perbedaan kualitas radiasi yang sangat
efektif, untuk menginduksi pembentukkan
mikronukleus (Gambar 4) [15].
Tabel 2. Koefisien kurva respon mikronukleus Vs dosis secara in vitro, pada berbagai
kualitas radiasi untuk LET tinggi [8]
Jenis Radiasi Komponen α (Gy) Komponen β (Gy)
Neutron beam campuran 220 kV 0,595 ± 0,028 - 0,059 ± 0,0133
Partikel α 20-23 keV 0,336 ± 0,039 -
Neutron 5,5 MeV 0,374 ± 0,012 -
Jenis Radiasi Komponen α (Gy) Komponen β (Gy)
Sinar-X 14 kVp 0,103 ± 0,003 0,057 ± 0,007
Sinar-X 50 kVp 0,087 ± 0,024 0,045 ± 0,024
Sinar-X 350 kVp 0,064 ± 0,018 0,045 ± 0,008
Sinar γ C0-60 0,049 ± 0,008 0,038 ± 0,006
Partikel β 2,27 MeV [4] 0,0248 ± 0,0134 0,0381 ± 0,0106
Seminar Nasional Keselamatan Kesehatan dan Lingkungan VII
Jakarta, 6-7 Juli 2011
PTKMR-BATAN, BAPETEN, KEMENKES-RI dan Pusarpedal-KLH 67
Dosis (Gy)
Ju
mla
h M
N p
er 1
00
0 s
el C
B
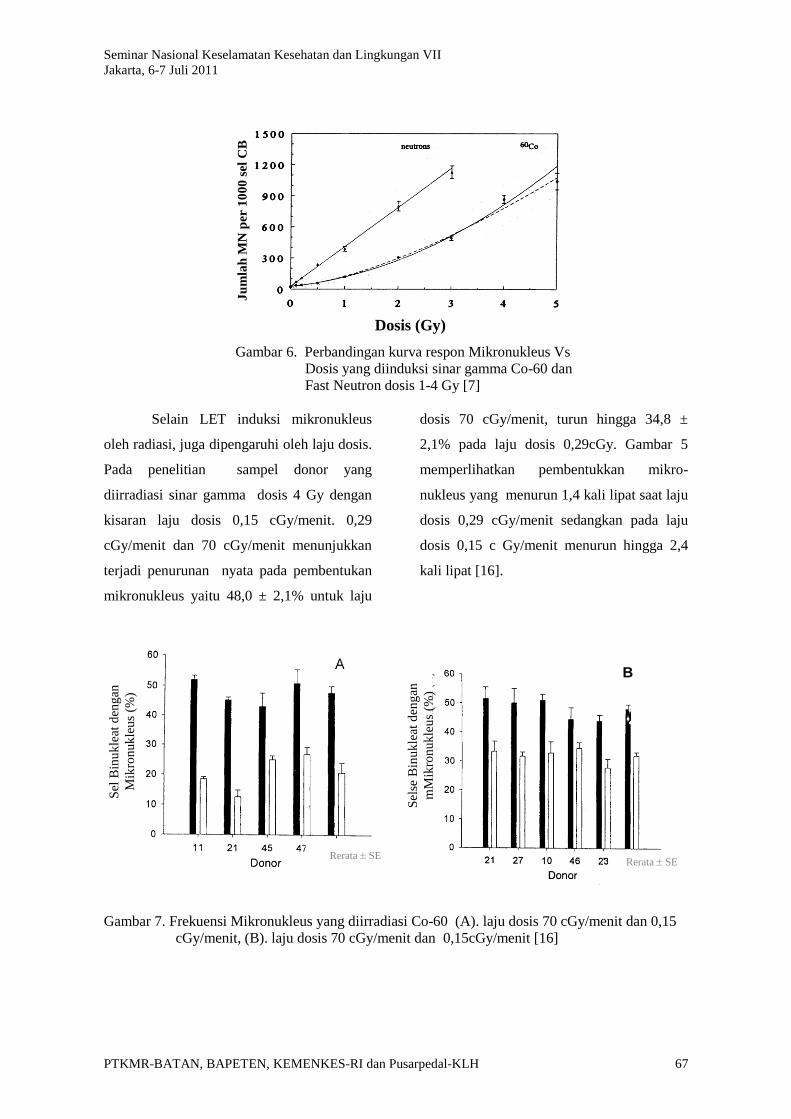
Gambar 6. Perbandingan kurva respon Mikronukleus Vs
Dosis yang diinduksi sinar gamma Co-60 dan
Fast Neutron dosis 1-4 Gy [7]
Selain LET induksi mikronukleus
oleh radiasi, juga dipengaruhi oleh laju dosis.
Pada penelitian sampel donor yang
diirradiasi sinar gamma dosis 4 Gy dengan
kisaran laju dosis 0,15 cGy/menit. 0,29
cGy/menit dan 70 cGy/menit menunjukkan
terjadi penurunan nyata pada pembentukan
mikronukleus yaitu 48,0 ± 2,1% untuk laju
dosis 70 cGy/menit, turun hingga 34,8 ±
2,1% pada laju dosis 0,29cGy. Gambar 5
memperlihatkan pembentukkan mikro-
nukleus yang menurun 1,4 kali lipat saat laju
dosis 0,29 cGy/menit sedangkan pada laju
dosis 0,15 c Gy/menit menurun hingga 2,4
kali lipat [16].
Gambar 7. Frekuensi Mikronukleus yang diirradiasi Co-60 (A). laju dosis 70 cGy/menit dan 0,15
cGy/menit, (B). laju dosis 70 cGy/menit dan 0,15cGy/menit [16]
Sel
Bin
uk
leat
den
gan
Mik
ron
uk
leu
s (%
)
Rerata SE
Sel
se B
inu
kle
at d
eng
an
mM
ikro
nuk
leu
s (%
)
Rerata SE
B

Seminar Nasional Keselamatan Kesehatan dan Lingkungan VII
Jakarta, 6-7 Juli 2011
PTKMR-BATAN, BAPETEN, KEMENKES-RI dan Pusarpedal-KLH 68
Induksi mikronukleus dipengaruhi
oleh laju dosis pada penelitian lain
dilaporkan bahwa pembentukkan mikro-
nukleus yang diirradiasi dengan laju dosis 0,7
cGy/menit sampai 2,6 cGy/menit, responnya
kurang efektif dibanding dengan laju dosis 40
Gy/jam. Demikian pula pada penelitian
Boreham diperlihatkan pola yang sama yaitu
pada laju dosis 0,29 cGy/menit dan 0,15
cGy/menit menunjukkan efek yang lebih
rendah, dibanding dengan laju dosis tinggi
yaitu 70 cGy/menit [16,17].
Penurunan laju dosis dari
0,29cGy/menit hingga 0,15 cGy/menit
mungkin telah mampu menginduksi
penundaan siklus sel sedangkan pada laju
dosis tinggi tidak dipengaruhi. Kemungkinan
penundaan siklus sel yamg diinduksi pada
laju dosis 0,15 cGy/menit tidak diperlukan
untuk pembentukkan mikronukleus pada laju
dosis 0,29 cGy/menit diperlihatkan
penundaan siklus sel tidak merupakan bagian
utama untuk kemampuan menaikkan proses
perbaikan patahan kromosom pada waktu
terjadi penurunan laju dosis. Diduga
penundaan siklus sel mungkin hanya sebagai
efek samping pada laju dosis yang sangat
rendah [16,17].
V. MIKRONUKLEI SEBAGAI
DOSIMETRI BIOLOGI
Beberapa karakteristik yang menjadi
syarat sebagai dosimetri biologi, antara lain
adalah adanya variasi mikronukleus antar
individu. Penelitian oleh Lee TK dkk
mengisyaratkan bahwa mikronuklei dalam
limfosit darah perifer dapat dijadikan sebagai
biodosimeter untuk pajanan akut dan
mungkin juga kronik setelah radiasi in
vivo[18]. Pembedaan antara paparan radiasi
pengion dengan non radiasi juga dapat
diketahui dengan uji mikronuklei
menggunakan pelabelan kromosom. Namun
terdapat variasi yang cukup besar antar-
laboratorium atau antar-individu, terutama
untuk dosis dengan LET rendah, sehingga
masing- masing laboratorium harus
merekontruksi kurva dosis-respon sendiri.
Jumlahnya yang menurun dengan waktu dan
sensitivitasnya rendah menyebabkan uji ini
perlu dipertimbangkan sebagai dosimetri
biologi. Dengan kejadian mikronuklei
spontan antara 3-30 per 1000 BNC, uji ini
juga menjadi kurang diminati, selain dapat
meninggi dengan bertambahnya umur untuk
kejadian MN spontan [1,18]. Peneliti lain
beranggapan bahwa variasi individu tersebut
dipertimbangkan /dianggap sangat kecil dan
dianggap sebagai refleksi dosis fisik yang
diterima setiap individu.
Variasi individu dalam pembentukan
mikronukleus yang di induksi oleh radiasi,
tidak menunjukkan perbedaan yang
bermakna diantara donor karena
kemungkinan pada penelitian tersebut ada
koinsidensi pada seleksi donor dan hasil
penelitian lain dilaporkan bahwa ada variasi
individu yang positif dalam pembentukan
mikronukleus yang dihubungkan dengan
umur yang dipengaruhi oleh jenis kelamin.

Seminar Nasional Keselamatan Kesehatan dan Lingkungan VII
Jakarta, 6-7 Juli 2011
PTKMR-BATAN, BAPETEN, KEMENKES-RI dan Pusarpedal-KLH 69
[3,17]. Data yang diperoleh dari hasil
penelitian terhadap ke 8 variasi donor,
dengan kisaran umur 23 sampai 55 tahun,
secara jelas terdapat keterkaitan antara
frekuensi latar mikronukleus dengan dosis.
Radiasi yang menginduksi frekuensi
mikronukleus menunjukkan tidak ada
keterkaitan umur yang bermakna dengan
frekuensi mikronukleus yang terbentuk,
namun demikian variasi yang besar terjadi
antar individu [8].
Dibanding dengan metoda sitogenetik
klasik kajian dosis dengan metoda CB untuk
mikronukleus dalam limfosit tepi ,relatif
lebih sederhana. Dapat dihitung lebih cepat
dan variasi antar scorer kurang berpengaruh.
Salah satu kekurangannya adalah dalam hal
data dasar /latar dari frekuensi mikronukleus,
baik dari segi jumlah, maupun variasi
individu. dibanding dengan aberasi
kromosom. Dengan menghitung 4000-5000
sel pada dosis terendah 0,05 Gy sudah dapat
mendeteksi adanya pembentukkan
mikronukleus. Studi lain telah dilaporkan
menggunakan sistem scoring otomatis
melalui komputer, dengan menghitung sel
binukleat dalam 103, semua keraguan
/ketidakpastian dalam pengkajian dosis
individual adalah 0,25 Gy, sementara dosis
kurang dari 0,3 Gy tidak dapat dideteksi
dengan jelas. Keraguan tersebut terutama
karena variasi mikronukleus antar individu.
Pengembangan untuk kisaran dosis rendah
diperlukan pengetahuan data dasar frekuensi
mikronukleus individu, sebelum irradiasi,
yaitu dengan cara mengukur respon dosis
untuk setiap individu [6].
VI. PENUTUP
Dari hasil kajian diatas, studi
mengenai pembentukan mikronukleui akibat
radiasi dengan menggunakan tehnik CB,
pada limfosit tepi merupakan metoda
radiobiologi yang relevan, bila digunakan
sebagai bidosimetri radiasi. Hubungan respon
mikronukleus dengan dosis untuk LET
rendah digambarkan dengan persamaan
linier qudratik, sedangkan untuk LET tingi
hanya persamannya linier. Induksi
mikronulei juga dipengaruhi oleh laju dosis,
pada laju dosis rendah frekuensinya semakin
turun. Salah satu kekurangannya adalah
dalam hal data dasar /latar dari frekuensi
mikronukleus, baik dari segi jumlah, maupun
variasi individu. Sehingga untuk menerapkan
uji mikronukleus sebagai dosimetri biologi,
diperlukan kalibrasi kurva mikronukleus
pada individu pra pajanan terutama untuk
dosis rendah.
DAFTAR PUSTAKA
1. INTERNATIONAL ATOMIC ENERGY
AGENCY, Biological Dosimetry
Chromosomal Aberration Analysis for
Dose Assessments. Technical Reports
Series No. 260, IAEA, Viena, 25-31,
2001.
2. COUNTRYMAN,P.I., HEDDLE,
J.A.,The production of Micronuclei from
Chromosome Aberration in Irradiated
Culture of Human Lymfocytes, Mutation
Research, 41, 321-331, 1979.

Seminar Nasional Keselamatan Kesehatan dan Lingkungan VII
Jakarta, 6-7 Juli 2011
PTKMR-BATAN, BAPETEN, KEMENKES-RI dan Pusarpedal-KLH 70
3. FENECH, M and MARLEY, A.A.
Measurement Of Micronuklei in
Limphocytes. Mutation Research, 147,
29-36 (1985).
4. HEDDLE., J.B and CARRANO, The
DNA Content of Micronuclei Induced in
Bone Marrow by Gamma Irradiation: in
Evident that micronuclei arise from
acentric Chromosomal Fragments.
Mutation Research 44, 63-70, 1977.
5. ALMASSY, Z., KREPINSKY. A.B.,
BIANCO. et. al. The Present State and
Perspectives of Micronucleus Essay in
Radiation Protection. In A Review. Appl
Radiat Isotop .38, 241-249, 1987.
6. KOKSAL, G., DALCI, D.O., and
PALA, F.S., Micronuclei in Human
Lymphocytes: The Co-60 Gamma Rays
Dose Response Mutation Research 161,
193-195, 1996.
7. VRAL, A., VERHAEGEN, H.
THIERENS, H., et.al. The InVitro
Cytokinesis-block Micronucleus Assay:
Detailed Description of an Improved
Slide Preparation Tehnique for The
Automated Detection of Micronuclei in
Human Lymphocytes. Mutagenesis 9,
439-443, 1994.
8. MILL, A.J., WELLS, J., HALL, S.C., et.
al, A. Micronucleus Induction in
Human Lymphocytes: Comparative
Effects of X Rays, Alpha Particles,
Beta Particles and Neutrons and
Implications for Biological Dosimetry,
Radiation Research, 145, 575-585, 1996.
9. PURNAMI, S., LUSIYANTI, Y.,
SYAIFUDIN, M., RHAMADANI, D.,
NURHAYATI, S., TETRIANA, D.,
Radiation Induced Micronuclei in
Lymphocyte Cell of Radiation Workers.
Prociding The International Conference
On Basic Science, Brawijaya University
Malang Indonesia, February 17-18, 2011.
10. SYAIFUDIN, M., dan KANG, C.,
Induksi Aberasi kromosom dan
mikronuklei dalam limfosit manusia
akibat radiasi gamma dan keandalannya
sebagai dosimeter biologi.Prosiding
Seminar Nasional Fisika Universitas
Andalas Padang, 5 September 2007,
ISBN 978-979-25-1951-B.
11. HALL, E.J. Radiobiology for
Radiologist, Fourth Edition. J.B.
Lippincot Company, Philadelphia,
Baltimore New York, London, 161-165,
1993.
12. LITTLE, J.B. What are the Risks of Low
Level Exposure to α Radiation from
Radon. Proc.Natl.Acad. Sci USA. 94,
5996-5997, 1997.
13. VRAL, A., FERHAGEN, F.,
THIERENS, H., et. al. Micronuclei
Induced by Fast Neutron Versus Co-60
Gamma Rays in Human peripheral Blood
Lymphocytes, Int.J. Radiat. Biol 65, no
3, 321-328, 1994.
14. VERHAEGEN. F and VRAL A.
Sensitivity of Micronucleus Induction in
Human Lymphovytes to Low LET
Radiation Qualities : in RBE and
Correlation of RBE and LET. Radiation
Research 139, 208-213, 1994.
15. HUBER, R., SCHRAUBE, H.,
NAHRSTEDT, U., et. al. Dose Response
Relationship of Micronuclei in Human
Lymphocytes Induced by Fission
Neutrons and by Low LET Radiations,
Mutation Research, 306, 135-141, 1994.
16. BOREHAM, D.R., DOLLING, J.A,
MAVES, S.R, et. al. Dose Rate Effect
for Apoptosis and Mikronucleus
Formation in Gamma Irradiated Human
Lymphocytes, Radiation Research 153,
579-586, 2000.
17. VRAL, A., THIERENS, H., BAEYENS,
A. et. al. Study of Dose Rate and Split
Dose Effects on The in vitro
Micronucleus yield in Human
Lymphocytes exposed to X-Rays. Int. J.
Radiat Biol 61. 777-784, 1992.
18. LEE, T-K., ALLISON, R. R.,O'BRIEN,
K. F., NAVES, J. L.KARLSSON, U. L.
and WILEY, A.L., Jr. Persistence of
micronuclei in lymphocytes of cancer
patients after radiotherapy. Radiation
Research,157, 678–684, 2002.

Seminar Nasional Keselamatan Kesehatan dan Lingkungan VII
Jakarta, 6-7 Juli 2011
PTKMR-BATAN, BAPETEN, KEMENKES-RI dan Pusarpedal-KLH 71
TANYA JAWAB
Penanya : Maria Lina
Pertanyaan :
- Dari ke empat uji yang telah dijelaskan,
uji manakah yang paling sensitif dan
spesifik untuk mengetahui kerusakan
kromosom ?
- Apa kelebihan Uji mikronukleus untuk
menganalisis kerusakan kromosom
akibat radiasi?
Jawaban
- Pengujian aberasi kromosom disentrik
adalah pengujian yang paling sensitive
untuk mengetahui kerusakan kromosom
akibat radiasi.
- Untuk uji mikronukleus dari segi
pengamatan lebih cepat dan mudah serta
tidak membutuhkan skill khusus.