sejarah penemuan dna
DESCRIPTION
biokimiaTRANSCRIPT

BIOKIMIA II Ririn Vidiastuti
SEJARAH PENEMUAN DNA
Sejarah Penemuan DNA (Deoxyribonucleic Acid)... Molekul Deoxyribonucleic
Acid atau DNA pertama ditemukan oleh seorang ahli ilmu kimia berkebangsaan
Jerman bernama Friedrich Miescher pada tahun 1869. Miescher menyelidiki susunan
kimia dari nukleus sel. Ia mengetahui bahwa nukleus sel tidak terdiri dari karbohidrat,
protein maupun lemak, melainkan terdiri dari zat yang mempunyai pengandungan
fosfor sangat tinggi. Oleh karena zat itu terdapat di dalam nukleus sel, maka zat itu
disebutnya nuklein. Nama ini kemudian diubah menjadi asam nukleat, karena asam
ikut menyusunnya.
Penelitian berikutnya dilakukan oleh Fisher pada tahun 1880. Dari hasil risetnya
ditemukan adanya zat-zat pirimidin dan purin di dalam asam nukleat. Temuan ini
dikembangkan lagi oleh Albreent Kossel yang menghasilkan temuan dua pirimidin
yaitu sitosin dan timin dan dua purin yaitu adenin dan guanin di dalam asam nukleat,
sehingga atas penemuannya ia mendapatkan hadiah nobel pada tahun 1910.
Pada tahun 1920-an, dengan pewarna ungu DNA yang khas, yang
dikembangkan oleh ahli kimia Jerman, Robert Feulgen, DNA ditemukan terletak secara
ekslusif pada kromosom. Karena itu, DNA merupakan lokasi yang diharapkan bagi
suatu bahan genetik. Pada tahun yang sama Phoebus Levine dari Institut Rockefeller
(seorang ahli biokimia kelahiran Rusia) mengungkapkan bahwa gula DNA adalah
deoksiribosa (karena itu namanya asam deoksiribonukleat).
Avery Machlead dan Mc Arthy (1944) memberi penegasan terhadap penemuan
terdahulu bahwa DNA mempunyai hubungan langsung dengan keturunan.
Selanjutnya penelitian Chargaff di tahun 1955, melalui hidrolisis DNA membuktikan
bahwa pada berbagai macam makhluk ternyata banyaknya adenin selalu kira-kira
sama dengan banyaknya timin (A=T), demikian pula dengan sitosin dan guanin
(S=G). Dengan perkataan lain, aturan Ghargaff menyatakan bahwa perbandingan A/T
dan S/G selalu mendekati satu.
Penelitian selanjutnya dilakukan oleh ahli biologi molekuler, James Dewey
Watson dan Francis H.C. Crick pada tahun 1953. Hasil penelitian tersebut
memperlihatkan bahwa DNA tidak berdiri sendiri sebagai suatu rantai tunggal
melainkan sebagai dua rantai yang saling berpilin, dengan basa pada rantai yang satu
melekat pada basa rantai yang lain. Dengan lain perkataan, DNA adalah suatu heliks
ganda. Teori model ini dikukuhkan dan disempurnakan oleh M.A.F. Wilkins pada tahun
1961. Oleh karena penemuan ini mereka bertiga mendapat hadiah nobel pada tahun
1962 dalam kedokteran dan fisiologi.

BIOKIMIA II Ririn Vidiastuti

BIOKIMIA II Ririn Vidiastuti
ASAM NUKLEAT
Asam nukleat adalah biopolymer yang berbobot molekul tinggi dengan unit
monomernya mononukleotida. Asam nukleat terdapat pada semua sel hidup dan
bertugas untuk menyimpan dan mentransfer genetic, kemudian menerjemahkan
informasi ini secara tepat untuk mensintesis protein yang khas bagi masing-masing
sel. Asam nukleat (bahasa Inggris: nucleic acid) adalah makromolekul biokimia yang
kompleks, berbobot molekul tinggi, dan tersusun atas rantai nukleotida yang
mengandung informasi genetik. Asam nukleat yang paling umum adalah Asam
deoksiribonukleat (DNA) and Asam ribonukleat (RNA). Asam nukleat ditemukan pada
semua sel hidup serta pada virus. Asam nukleat dinamai demikian karena keberadaan
umumnya di dalam inti (nukleus) sel.
Pada tahun 1879, Albrecht Kossel menemukan asam nukleat yang tersusun
oleh suatu gugus gula, gugus fosfat, dan gugus basa. Asam nukleat berbentuk rantai
linier yang merupakan gabungan monomer nukleotida sebagai unit pembangunnya.
Molekul ini menyimpan informasi pertumbuhan sel dan reproduksi.
Monomer nukleotida sebagai struktur primer asam nukleat diperoleh dari hasil
hidrolisis asam nukleat. Proses hidrolisis lebih lanjut dari monomer nukleotida akan
dihasilkan asam fosfat dan nukleosida. Proses hidrolisis ini dilakukan dalam suasana
basa. Jika hidrolisis dilanjutkan kembali terhadap senyawa nukleosida dalam larutan
asam berair akan dihasilkan molekul gula dan basa nitrogen dengan bentuk
heterosiklik.
STRUKTUR, MACAM, DAN FUNGSI ASAM NUKLEAT
Makrobiomolekul ini mempunyai susunan yang sangat unik, yaitu berupa
polimer yang tersusun atas monomer yang disebut nukleotida. Tiap nukleotida terdiri
atas nukleosida dan asam fosfat. Nukleosida terdiri atas gula pentose (ribose atau
deoksiribosa) dan basa nitrogen heterosiklik, yaitu turunan purina (adenine dan
guanine) dan turunan pirimidina (sitosin, urasil, dan timin).
(Sumardjo,2006)
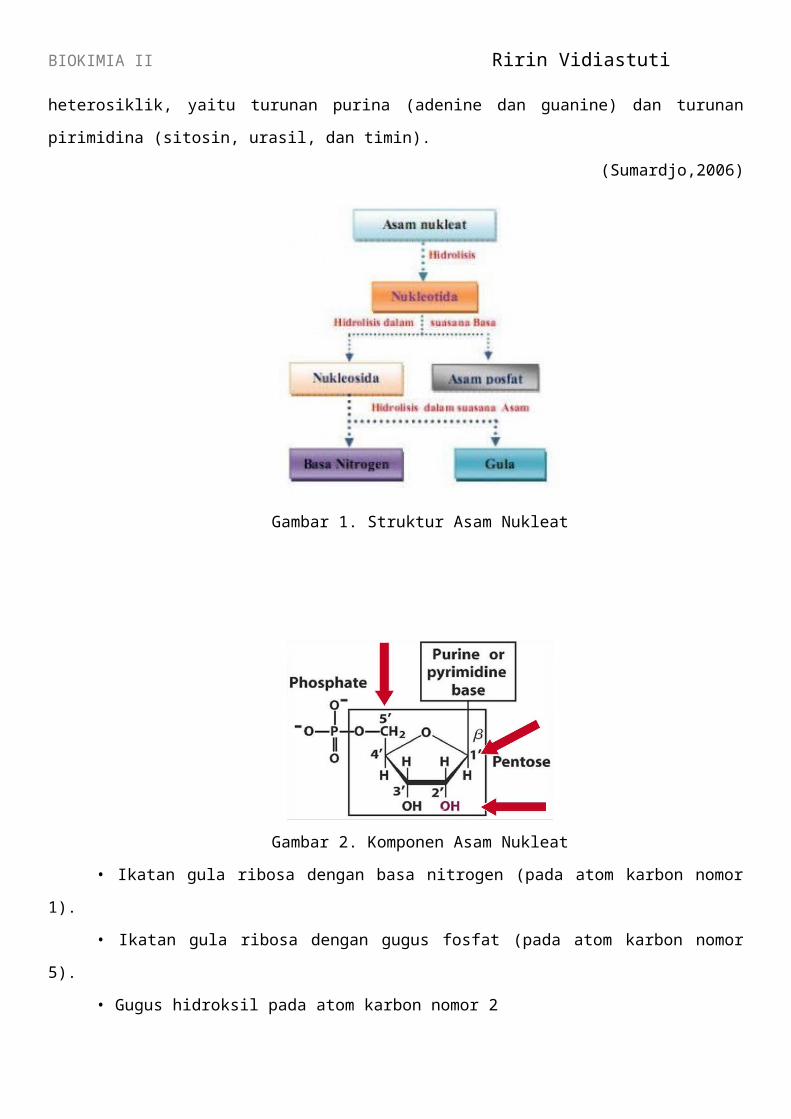
BIOKIMIA II Ririn Vidiastuti
Gambar 1. Struktur Asam Nukleat
Gambar 2. Komponen Asam Nukleat
• Ikatan gula ribosa dengan basa nitrogen (pada atom karbon nomor 1).
• Ikatan gula ribosa dengan gugus fosfat (pada atom karbon nomor 5).
• Gugus hidroksil pada atom karbon nomor 2
Di antara ketiga komponen monomer asam nukleat tersebut di atas, hanya basa
N-lah yang memungkinkan terjadinya variasi. Pada kenyataannya memang urutan
(sekuens) basa N pada suatu molekul asam nukleat merupakan penentu bagi
spesifisitasnya. Dengan perkataan lain, identifikasi asam nukleat dilakukan
berdasarkan atas urutan basa N-nya sehingga secara skema kita bisa
menggambarkan suatu molekul asam nukleat hanya dengan menuliskan urutan
basanya saja.

BIOKIMIA II Ririn Vidiastuti
NUKLEOTIDA DAN NUKLEOSIDA
Gambar 3. Nukleotida dan Nukleosida
Penomoran posisi atom C pada cincin gula dilakukan menggunakan tanda aksen
(1’, 2’, dan seterusnya), sekedar untuk membedakannya dengan penomoran posisi
pada cincin basa. Posisi 1’ pada gula akan berikatan dengan posisi 9 (N-9) pada basa
purin atau posisi 1 (N-1) pada basa pirimidin melalui ikatan glikosidik atau glikosilik
(Gambar 2.2). Kompleks gula-basa ini dinamakan nukleosida.
Di atas telah disinggung bahwa asam nukleat tersusun dari monomer-monomer
berupa nukleotida, yang masing-masing terdiri atas sebuah gugus fosfat, sebuah gula
pentosa, dan sebuah basa N. Dengan demikian, setiap nukleotida pada asam nukleat
dapat dilihat sebagai nukleosida monofosfat. Namun, pengertian nukleotida secara
umum sebenarnya adalah nukleosida dengan sebuah atau lebih gugus fosfat. Sebagai
contoh, molekul ATP (adenosin trifosfat) adalah nukleotida yang merupakan
nukleosida dengan tiga gugus fosfat.
Jika gula pentosanya adalah ribosa seperti halnya pada RNA, maka
nukleosidanya dapat berupa adenosin, guanosin, sitidin, dan uridin. Begitu pula,
nukleotidanya akan ada empat macam, yaitu adenosin monofosfat, guanosin
monofosfat, sitidin monofosfat, dan uridin monofosfat. Sementara itu, jika gula
pentosanya adalah deoksiribosa seperti halnya pada DNA, maka (2’-
deoksiribo)nukleosidanya terdiri atas deoksiadenosin, deoksiguanosin, deoksisitidin,
dan deoksitimidin.
Ikatan fosfodiester
Selain ikatan glikosidik yang menghubungkan gula pentosa dengan basa N,
pada asam nukleat terdapat pula ikatan kovalen melalui gugus fosfat yang
menghubungkan antara gugus hidroksil (OH) pada posisi 5’ gula pentosa dan gugus

BIOKIMIA II Ririn Vidiastuti
hidroksil pada posisi 3’ gula pentosa nukleotida berikutnya. Ikatan ini dinamakan
ikatan fosfodiester karena secara kimia gugus fosfat berada dalam bentuk diester.
Oleh karena ikatan fosfodiester menghubungkan gula pada suatu nukleotida
dengan gula pada nukleotida berikutnya, maka ikatan ini sekaligus menghubungkan
kedua nukleotida yang berurutan tersebut. Dengan demikian, akan terbentuk suatu
rantai polinukleotida yang masing-masing nukleotidanya satu sama lain dihubungkan
oleh ikatan fosfodiester.
Kecuali yang berbentuk sirkuler, seperti halnya pada kromosom dan plasmid
bakteri, rantai polinukleotida memiliki dua ujung. Salah satu ujungnya berupa gugus
fosfat yang terikat pada posisi 5’ gula uanine. Oleh karena itu, ujung ini dinamakan
ujung P atau ujung 5’. Ujung yang lainnya berupa gugus hidroksil yang terikat pada
posisi 3’ gula uanine sehingga ujung ini dinamakan ujung OH atau ujung 3’. Adanya
ujung-ujung tersebut menjadikan rantai polinukleotida linier mempunyai arah
tertentu.
Pada pH netral adanya gugus fosfat akan menyebabkan asam nukleat
bermuatan uanine. Inilah uanine pemberian nama ’asam’ kepada molekul
polinukleotida meskipun di dalamnya juga terdapat banyak basa N. Kenyataannya,
asam nukleat memang merupakan anion asam kuat atau merupakan polimer yang
sangat bermuatan uanine.
Sekuens asam nukleat
Telah dikatakan di atas bahwa urutan basa N akan menentukan spesifisitas
suatu molekul asam nukleat sehingga biasanya kita menggambarkan suatu molekul
asam nukleat cukup dengan menuliskan urutan basa (sekuens)-nya saja. Selanjutnya,
dalam penulisan sekuens asam nukleat ada kebiasaan untuk menempatkan ujung 5’
di sebelah kiri atau ujung 3’ di sebelah kanan. Sebagai contoh, suatu sekuens DNA
dapat dituliskan 5’-ATGACCTGAAAC-3’ atau suatu sekuens RNA dituliskan 5’-
GGUCUGAAUG-3’.
Jadi, spesifisitas suatu asam nukleat selain ditentukan oleh sekuens basanya,
juga harus dilihat dari arah pembacaannya. Dua asam nukleat yang memiliki sekuens
sama tidak berarti keduanya sama jika pembacaan sekuens tersebut dilakukan dari
arah yang berlawanan (yang satu 5’→ 3’, sedangkan yang lain 3’→ 5’).

BIOKIMIA II Ririn Vidiastuti
Gambar 4. Basa Nitrogen
( Susanto, 2012)
Ada dua jenis asam nuklet:
1. DNA (deoxyribonucleid acid)
Asam ini adalah polimer yang terdiri atas molekul-molekul deoksiribonukleotida
yang terikat satu sama lain sehingga membentuk rantai polinukleotida yang panjang.
Molekul DNA yang panjang ini terbentuk oleh ikatan antara atom C nomor 3 dengan
atom C nomor 5 pada molekul deoksiribosa dengan perantaraan gugus fosfat. Secara
kimia DNA mengandung karakteri/sifat sebagai berikut:
a. Memiliki gugus gula deoksiribosa.
b. Basa nitrogennya uanine (G), sitosin ©, timin (T) dan uanine (A).
c. Memiliki rantai heliks ganda anti parallel
d. Kandungan basa nitrogen antara kedua rantai sama banyak dan berpasangan
spesifik satu dengan lain. Guanin selalu berpasangan dengan sitosin ( G –C), dan
uanine berpasangan dengan timin (A – T), sehingga jumlah uanine selalu sama
dengan jumlah sitosin. Demikian pula uanine dan timin.
2. RNA (ribonucleid acid)
Asam ribonukleat adalah suatu polimer yang terdiri atas molekul-molekul
ribonukleotida. Seperti DNA asam ribonukleat terbentuk oleh adanya ikatan antara
atom C nomor 3 dengan atom C nomor 5 pada molekul uanin dengan perantaraan
gugus fosfat. Rumus strukturnya sama dengan gambar 10.2 tetapi gulanya adalah

BIOKIMIA II Ririn Vidiastuti
uanin ( atom C nomor 2 mengikat gugus OH) RNA memiliki sifat spesifik yang berbeda
dengan sifat kimia DNA, yakni dalam hal:
a. Gula pentosanya adalah ribose
b. RNA memiliki ribonukleotida uanine(G), sitosin ©, uanine (A) dan Urasil (U)
pengganti Timin pada DNA.
c. Untai fosfodiesternya adalah untai tunggal yang bisa melipat membentuk
jepit rambut seperti untai ganda.Beda dengan DNA bentuk molekulnya heliks ganda.
d. Prosentasi kandungan bas tidak harus sama, pasangan uanine tidak harus
sama dengan urasil, dan sitosin tidak harus sama dengan uanine.
Ada tiga jenis RNA yaitu tRNA (transfer RNA), mRNA (messenger RNA) dan rRNA
(ribosomal RNA). Ketiga macam RNA ini mempunyai fungsi yang berbeda-beda, tetapi
ketiganya secara bersama-sama mempunyai peranan penting dalam sintesis protein.
( Mustofa,2012)
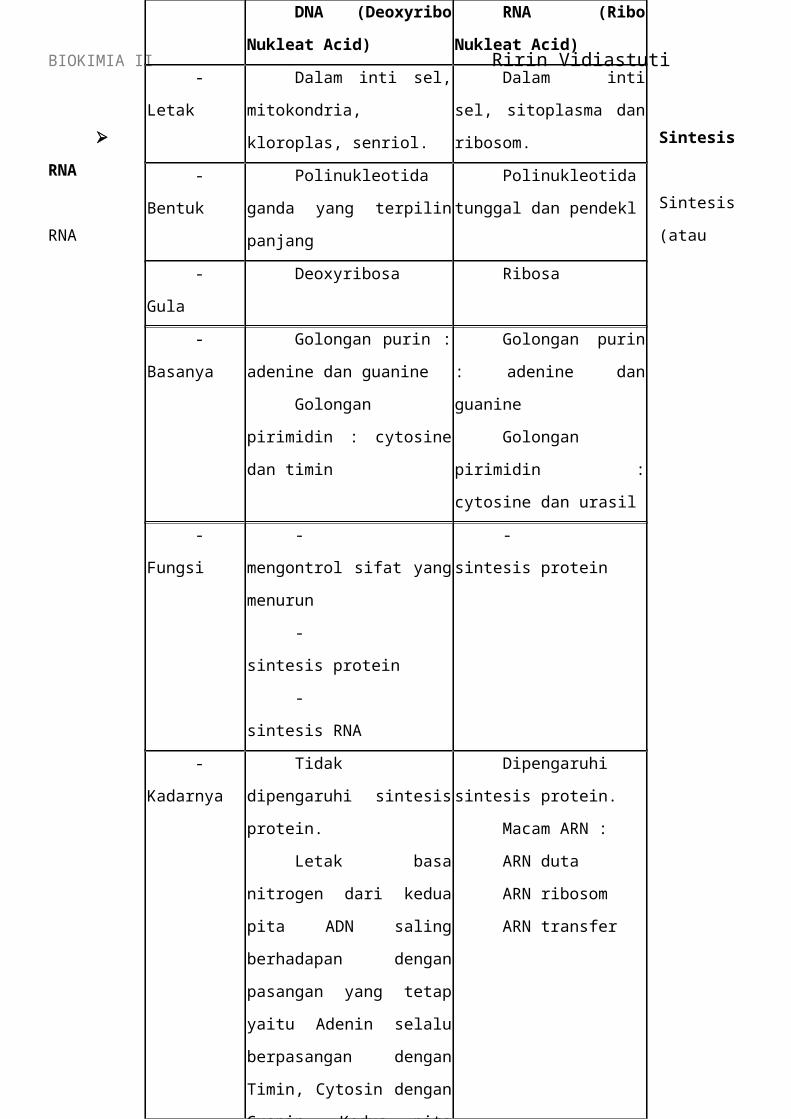
DNA RNA
Gula :
deoksiribosa
Basa : AGCT
Untai Ganda
Prokariot :
Sitoplasma
Eukariot : Inti
Penyimpan
informasi
Gula : Ribosa
Basa : AGCU
Untai Tunggal
Prokariot :
sitoplasma
Eukariot : inti
dan sitoplasma
Hasil transkripsi
Beberapa fungsi penting asam nukleat adalah menyimpan, menstransmisi, dan
mentranslasi informasi genetik; metabolisme antara (intermediary metabolism) dan
reaksi-reaksi informasi energi; koenzim pembawa energi; koenzim pemindah asam
asetat, zat gula, senyawa amino dan biomolekul lainnya; koenzim reaksi oksidasi
reduksi.
( Mustofa, 2012)
2.2 Sifat – sifat asam nukleat
Sifat-sifat Fisika-Kimia Asam Nukleat

BIOKIMIA II Ririn Vidiastuti
Di bawah ini akan dibicarakan sekilas beberapa sifat fisika-kimia asam nukleat.
Sifat-sifat tersebut adalah stabilitas asam nukleat, pengaruh asam, pengaruh alkali,
denaturasi kimia, viskositas, dan kerapatan apung.
a. Stabilitas asam nukleat
Ketika kita melihat struktur tangga berpilin molekul DNA atau pun struktur
sekunder RNA, sepintas akan nampak bahwa struktur tersebut menjadi stabil akibat
adanya ikatan hidrogen di antara basa-basa yang berpasangan. Padahal, sebenarnya
tidaklah demikian. Ikatan hidrogen di antara pasangan-pasangan basa hanya akan
sama kuatnya dengan ikatan hidrogen antara basa dan molekul air apabila DNA
berada dalam bentuk rantai tunggal. Jadi, ikatan hidrogen jelas tidak berpengaruh
terhadap stabilitas struktur asam nukleat, tetapi sekedar menentukan spesifitas
perpasangan basa.
Penentu stabilitas struktur asam nukleat terletak pada interaksi penempatan
(stacking interactions) antara pasangan-pasangan basa. Permukaan basa yang
bersifat hidrofobik menyebabkan molekul-molekul air dikeluarkan dari sela-sela
perpasangan basa sehingga perpasangan tersebut menjadi kuat.
b. Pengaruh asam
Di dalam asam pekat dan suhu tinggi, misalnya HClO4 dengan suhu lebih dari
100ºC, asam nukleat akan mengalami hidrolisis sempurna menjadi komponen-
komponennya. Namun, di dalam asam mineral yang lebih encer, hanya ikatan
glikosidik antara gula dan basa purin saja yang putus sehingga asam nukleat
dikatakan bersifat apurinik.
c. Pengaruh alkali
Pengaruh alkali terhadap asam nukleat mengakibatkan terjadinya perubahan
status tautomerik basa. Sebagai contoh, peningkatan pH akan menyebabkan
perubahan struktur guanin dari bentuk keto menjadi bentuk enolat karena molekul
tersebut kehilangan sebuah proton. Selanjutnya, perubahan ini akan menyebabkan
terputusnya sejumlah ikatan hidrogen sehingga pada akhirnya rantai ganda DNA
mengalami denaturasi. Hal yang sama terjadi pula pada RNA. Bahkan pada pH netral
sekalipun, RNA jauh lebih rentan terhadap hidrolisis bila dibadingkan dengan DNA
karena adanya gugus OH pada atom C nomor 2 di dalam gula ribosanya.

BIOKIMIA II Ririn Vidiastuti
d. Denaturasi kimia
Sejumlah bahan kimia diketahui dapat menyebabkan denaturasi asam nukleat
pada pH netral. Contoh yang paling dikenal adalah urea (CO(NH2)2) dan formamid
(COHNH2). Pada konsentrasi yang relatif tinggi, senyawa-senyawa tersebut dapat
merusak ikatan hidrogen. Artinya, stabilitas struktur sekunder asam nukleat menjadi
berkurang dan rantai ganda mengalami denaturasi.
e. Viskositas
DNA kromosom dikatakan mempunyai nisbah aksial yang sangat tinggi karena
diameternya hanya sekitar 2 nm, tetapi panjangnya dapat mencapai beberapa
sentimeter. Dengan demikian, DNA tersebut berbentuk tipis memanjang. Selain itu,
DNA merupakan molekul yang relatif kaku sehingga larutan DNA akan mempunyai
viskositas yang tinggi. Karena sifatnya itulah molekul DNA menjadi sangat rentan
terhadap fragmentasi fisik. Hal ini menimbulkan masalah tersendiri ketika kita hendak
melakukan isolasi DNA yang utuh.
f. Kerapatan apung
Analisis dan pemurnian DNA dapat dilakukan sesuai dengan kerapatan apung
(bouyant density)-nya. Di dalam larutan yang mengandung garam pekat dengan
berat molekul tinggi, misalnya sesium klorid (CsCl) 8M, DNA mempunyai kerapatan
yang sama dengan larutan tersebut, yakni sekitar 1,7 g/cm3. Jika larutan ini
disentrifugasi dengan kecepatan yang sangat tinggi, maka garam CsCl yang pekat
akan bermigrasi ke dasar tabung dengan membentuk gradien kerapatan. Begitu
juga, sampel DNA akan bermigrasi menuju posisi gradien yang sesuai dengan
kerapatannya. Teknik ini dikenal sebagai sentrifugasi seimbang dalam tingkat
kerapatan (equilibrium density gradient centrifugation) atau sentrifugasi
isopiknik.
Oleh karena dengan teknik sentrifugasi tersebut pelet RNA akan berada di dasar
tabung dan protein akan mengapung, maka DNA dapat dimurnikan baik dari RNA
maupun dari protein. Selain itu, teknik tersebut juga berguna untuk keperluan analisis
DNA karena kerapatan apung DNA (ρ) merupakan fungsi linier bagi kandungan GC-
nya. Dalam hal ini, ρ = 1,66 + 0,098% (G + C).
Sifat-sifat Spektroskopik-Termal Asam Nukleat

BIOKIMIA II Ririn Vidiastuti
Sifat spektroskopik-termal asam nukleat meliputi kemampuan absorpsi sinar
UV, hipokromisitas, penghitungan konsentrasi asam nukleat, penentuan kemurnian
DNA, serta denaturasi termal dan renaturasi asam nukleat. Masing-masing akan
dibicarakan sekilas berikut ini.
a. Absorpsi UV
Asam nukleat dapat mengabsorpsi sinar UV karena adanya basa nitrogen yang
bersifat aromatik; fosfat dan gula tidak memberikan kontribusi dalam absorpsi UV.
Panjang gelombang untuk absorpsi maksimum baik oleh DNA maupun RNA adalah
260 nm atau dikatakan λmaks = 260 nm. Nilai ini jelas sangat berbeda dengan nilai
untuk protein yang mempunyai λmaks = 280 nm. Sifat-sifat absorpsi asam nukleat
dapat digunakan untuk deteksi, kuantifikasi, dan perkiraan kemurniannya.
b. Hipokromisitas
Meskipun λmaks untuk DNA dan RNA konstan, ternyata ada perbedaan nilai yang
bergantung kepada lingkungan di sekitar basa berada. Dalam hal ini, absorbansi pada
λ 260 nm (A260) memperlihatkan variasi di antara basa-basa pada kondisi yang
berbeda. Nilai tertinggi terlihat pada nukleotida yang diisolasi, nilai sedang diperoleh
pada molekul DNA rantai tunggal (ssDNA) atau RNA, dan nilai terendah dijumpai pada
DNA rantai ganda (dsDNA). Efek ini disebabkan oleh pengikatan basa di dalam
lingkungan hidrofobik. Istilah klasik untuk menyatakan perbedaan nilai absorbansi
tersebut adalah hipokromisitas. Molekul dsDNA dikatakan relatif hipokromik (kurang
berwarna) bila dibandingkan dengan ssDNA. Sebaliknya, ssDNA dikatakan hiperkromik
terhadap dsDNA.
c. Penghitungan konsentrasi asam nukleat
Konsentrasi DNA dihitung atas dasar nilai A260-nya. Molekul dsDNA dengan
konsentrasi 1mg/ml mempunyai A260 sebesar 20, sedangkan konsentrasi yang sama
untuk molekul ssDNA atau RNA mempunyai A260 lebih kurang sebesar 25. Nilai A260
untuk ssDNA dan RNA hanya merupakan perkiraan karena kandungan basa purin dan
pirimidin pada kedua molekul tersebut tidak selalu sama, dan nilai A260 purin tidak
sama dengan nilai A260 pirimidin. Pada dsDNA, yang selalu mempunyai kandungan
purin dan pirimidin sama, nilai A260 -nya sudah pasti.

BIOKIMIA II Ririn Vidiastuti
d. Kemurnian asam nukleat
Tingkat kemurnian asam nukleat dapat diestimasi melalui penentuan nisbah
A260 terhadap A280. Molekul dsDNA murni mempunyai nisbah A260 /A280 sebesar 1,8.
Sementara itu, RNA murni mempunyai nisbah A260 /A280 sekitar 2,0. Protein, dengan
λmaks = 280 nm, tentu saja mempunyai nisbah A260 /A280 kurang dari 1,0. Oleh karena
itu, suatu sampel DNA yang memperlihatkan nilai A260 /A280 lebih dari 1,8 dikatakan
terkontaminasi oleh RNA. Sebaliknya, suatu sampel DNA yang memperlihatkan nilai
A260 /A280 kurang dari 1,8 dikatakan terkontaminasi oleh protein.
e. Denaturasi termal dan renaturasi
Di atas telah disinggung bahwa beberapa senyawa kimia tertentu dapat
menyebabkan terjadinya denaturasi asam nukleat. Ternyata, panas juga dapat
menyebabkan denaturasi asam nukleat. Proses denaturasi ini dapat diikuti melalui
pengamatan nilai absorbansi yang meningkat karena molekul rantai ganda (pada
dsDNA dan sebagian daerah pada RNA) akan berubah menjadi molekul rantai tunggal.
Denaturasi termal pada DNA dan RNA ternyata sangat berbeda. Pada RNA
denaturasi berlangsung perlahan dan bersifat acak karena bagian rantai ganda yang
pendek akan terdenaturasi lebih dahulu daripada bagian rantai ganda yang panjang.
Tidaklah demikian halnya pada DNA. Denaturasi terjadi sangat cepat dan bersifat
koperatif karena denaturasi pada kedua ujung molekul dan pada daerah kaya AT akan
mendestabilisasi daerah-daerah di sekitarnya.
Suhu ketika molekul asam nukleat mulai mengalami denaturasi dinamakan titik
leleh atau melting temperature (Tm). Nilai Tm merupakan fungsi kandungan GC
sampel DNA, dan berkisar dari 80 ºC hingga 100ºC untuk molekul-molekul DNA yang
panjang.
DNA yang mengalami denaturasi termal dapat dipulihkan (direnaturasi) dengan
cara didinginkan. Laju pendinginan berpengaruh terhadap hasil renaturasi yang
diperoleh. Pendinginan yang berlangsung cepat hanya memungkinkan renaturasi
pada beberapa bagian/daerah tertentu. Sebaliknya, pendinginan yang dilakukan
perlahan-lahan dapat mengembalikan seluruh molekul DNA ke bentuk rantai ganda
seperti semula. Renaturasi yang terjadi antara daerah komplementer dari dua rantai
asam nukleat yang berbeda dinamakan hibridisasi.
f. Superkoiling DNA

BIOKIMIA II Ririn Vidiastuti
Banyak molekul dsDNA berada dalam bentuk sirkuler tertutup atau closed-
circular (CC), misalnya DNA plasmid dan kromosom bakteri serta DNA berbagai
virus. Artinya, kedua rantai membentuk lingkaran dan satu sama lain dihubungkan
sesuai dengan banyaknya putaran heliks (Lk) di dalam molekul DNA tersebut.
Sejumlah sifat muncul dari kondisi sirkuler DNA. Cara yang baik untuk
membayangkannya adalah menganggap struktur tangga berpilin DNA seperti gelang
karet dengan suatu garis yang ditarik di sepanjang gelang tersebut. Jika kita
membayangkan suatu pilinan pada gelang, maka deformasi yang terbentuk akan
terkunci ke dalam sistem pilinan tersebut. Deformasi inilah yang disebut sebagai
superkoiling.
g. Interkalator
Geometri suatu molekul yang mengalami superkoiling dapat berubah akibat
beberapa faktor yang mempengaruhi pilinan internalnya. Sebagai contoh,
peningkatan suhu dapat menurunkan jumlah pilinan, atau sebaliknya, peningkatan
kekuatan ionik dapat menambah jumlah pilinan. Salah satu faktor yang penting
adalah keberadaan interkalator seperti etidium bromid (EtBr). Molekul ini
merupakan senyawa aromatik polisiklik bermuatan positif yang menyisip di antara
pasangan-pasangan basa. Dengan adanya EtBr molekul DNA dapat divisualisasikan
menggunakan paparan sinar UV.
(Susanto, 2012)
2.3 Proses sintesis DNA dan RNA
REPLIKASI DNA (Asam deoksiribonukleat)
Replikasi DNA sangat penting agar DNA kita tidak termutasi, dimana DNA
menyimpan cetak biru bagi segala aktivitas sel. Pada proses replikasi terdapat pos-
pos pengecekan, sehingga jika ada kode yang salah tercetak, dapat langsung dikenali
dan langsung diperbaiki. Ada beberapa teori yang mencoba menjelaskan bagaimana
proses replikasi DNA ini terjadi, yaitu konservatif, semikonservatif, dan dispersif.
Tiga kemungkinan terjadinya replikasi DNA
1. Model konservatif, yaitu dua rantai DNA lama tetap tidak berubah, berfungsi
sebagai cetakan untuk dua rantai DNA baru. Replikasi ini mempertahankan molekul
dari DNA lama dan membuat molekul DNA baru.

BIOKIMIA II Ririn Vidiastuti
2. Model semikonservatif, yaitu dua rantai DNA lama terpisah dan rantai baru
disintesis dengan prinsip komplementasi pada masing-masing rantai DNA lama.
Akhirnya dihasilkan dua rantai DNA baru yang masing-masing mengandung satu
rantai cetakan molekul DNA lama dan satu rantai baru hasil sintesis.
3. Model dispersif, yaitu beberapa bagian dari kedua rantai DNA lama digunakan
sebagai cetakan untuk sintesis rantai DNA baru. Oleh karena itu, hasil akhirnya
diperoleh rantai DNA lama dan baru yang tersebar pada rantai DNA lama dan baru.
Replikasi ini menghasilkan dua molekul DNA lama dan DNA baru yang saling
berselang-seling pada setiap untai.
Setelah berhasil membuat model struktur DNA, Watson dan Crick memprediksi
bahwa DNA bereplikasi dengan cara semikonservatif. Kemudian pada tahun 1958,
Matthew Meselson dan Franklin Stahl melakukan percobaan untuk menguji ketiga
alternatif hipotesis replikasi DNA tersebut dengan menggunakan DNA bakteri
Eschericia coli. Hasilnya ternyata mendukung model replikasi semikonservatif yang
telah diprediksi oleh Watson dan Crick.
Berikut ini merupakan animasi proses replikasi DNA :
Gambar 5. Animasi replikasi DNA
Proses replikasi memerlukan protein atau enzim pembantu; salah satu
yang terpenting dikenal dengan nama DNA polimerase, yang merupakan enzim
pembantu pembentukan rantai DNA baru yang merupakan suatu polimer.
Proses replikasi diawali dengan pembukaan untaian ganda DNA pada titik-titik
tertentu di sepanjang rantai DNA. Proses pembukaan rantai DNA ini dibantu oleh
enzim helikase yang dapat mengenali titik-titik tersebut, dan enzim girase yang
mampu membuka pilinan rantai DNA. Jika dianalogikan, dua rantai DNA heliks ganda
yang terpisah berbentuk seperti ritsleting terbuka. Setelah cukup ruang terbentuk

BIOKIMIA II Ririn Vidiastuti
akibat pembukaan untaian ganda ini, DNA polimerase masuk dan mengikat diri pada
kedua rantai DNA yang sudah terbuka secara lokal tersebut. Proses pembukaan rantai
ganda tersebut berlangsung disertai dengan pergeseran DNA polimerase mengikuti
arah membukanya rantai ganda. Setiap rantai DNA yang “lama” akan berfungsi
sebagai cetakan yang menentukan urutan nukleotida di sepanjang rantai DNA
komplementer baru yang bersesuaian dengan cara mendeteksi basa komplemennya.
Monomer DNA yang berupa nukleotida ditambahkan di kedua sisi rantai yang
membuka setiap kali DNA polimerase bergeser. Setelah mendapatkan pasangan yang
sesuai, nukleotida yang baru tersebut disambung satu sama lain untuk membentuk
tulang punggung gula-fosfat rantai DNA yang baru. Jadi, setiap molekul DNA terdiri
atas satu rantai DNA “lama” dan satu rantai DNA “baru”. Setelah seluruh rantai
benar-benar terpisah, maka,terdapat dua molekul DNA yang sama persis dengan satu
molekul DNA induk.
Enzim DNA polimerase memiliki fungsi lain, yaitu mengoreksi DNA yang
baru terbentuk, membetulkan setiap kesalahan replikasi, dan memperbaiki DNA yang
rusak. Adanya fungsi tersebut menjadikan rangkaian nukleotida DNA sangat stabil
dan mutasi jarang terjadi.Yang berikut ini merupakan awal dari proses replikasi DNA
yang diambil dengan menggunakan mikroskop. Enzim DNA polimerase memisahkan
dua rantau DNA heliks ganda :
Proses replikasi DNA ini merupakan proses yang rumit namun teliti. Proses
sintesis rantai. Proses replikasi memerlukan protein atau enzim pembantu; salah satu
yang terpenting dikenal dengan nama DNA polimerase, yang merupakan enzim
pembantu pembentukan rantai DNA baru yang merupakan suatu polimer. Proses
replikasi diawali dengan pembukaan untaian ganda DNA pada titik-titik tertentu di
sepanjang rantai DNA. Proses pembukaan rantai DNA ini dibantu oleh enzim helikase
yang dapat mengenali titik-titik tersebut, dan enzim girase yang mampu membuka
pilinan rantai DNA. Setelah cukup ruang terbentuk akibat pembukaan untaian ganda
ini, DNA polimerase masuk dan mengikat diri pada kedua rantai DNA yang sudah
terbuka secara lokal tersebut. Proses pembukaan rantai ganda tersebut berlangsung
disertai dengan pergeseran DNA polimerase mengikuti arah membukanya rantai
ganda. Monomer DNA ditambahkan di kedua sisi rantai yang membuka setiap kali
DNA polimerase bergeser. Hal ini berlanjut sampai seluruh rantai telah benar-benar
terpisah. Proses replikasi DNA ini merupakan proses yang rumit namun teliti.
Proses sintesis rantai DNA baru memiliki suatu mekanisme yang
mencegah terjadinya kesalahan pemasukan monomer yang dapat berakibat fatal.

BIOKIMIA II Ririn Vidiastuti
Karena mekanisme inilah kemungkinan terjadinya kesalahan sintesis amatlah kecil.
Jadi, replikasi DNA adalah proses penggandaan molekul DNA untai ganda. Pada sel,
replikasi DNA terjadi sebelum pembelahan sel. Prokariota terus-menerus melakukan
replikasi DNA. Pada eukariota, waktu terjadinya replikasi DNA sangatlah diatur, yaitu
pada fase S siklus sel, sebelum mitosis atau meiosis I. Penggandaan tersebut
memanfaatkan enzim DNA polimerase yang membantu pembentukan ikatan antara
nukleotida-nukleotida penyusun polimer DNA. Proses replikasi DNA dapat pula
dilakukan in vitro dalam proses yang disebut reaksi berantai polimerase (PCR).
Mekanisme replikasi DNA
Ada tiga cara teoretis replikasi DNA yang pernah diusulkan, yaitu konservatif,
semikonservatif, dan dispersif. Pada replikasi konservatif seluruh tangga berpilin DNA
awal tetap dipertahankan dan akan mengarahkan pembentukan tangga berpilin baru.
Pada replikasi semikonservatif tangga berpilin mengalami pembukaan terlebih dahulu
sehingga kedua untai polinukleotida akan saling terpisah. Namun, masing-masing
untai ini tetap dipertahankan dan akan bertindak sebagai cetakan (template) bagi
pembentukan untai polinukleotida baru. Sementara itu, pada replikasi dispersif kedua
untai polinukleotida mengalami fragmentasi di sejumlah tempat. Kemudian, fragmen-
fragmen polinukleotida yang terbentuk akan menjadi cetakan bagi fragmen
nukleotida baru sehingga fragmen lama dan baru akan dijumpai berselang-seling di
dalam tangga berpilin yang baru. antara ketiga cara replikasi DNA yang diusulkan
tersebut, hanya cara semikonservatif yang dapat dibuktikan kebenarannya melalui
percobaan yang dikenal dengan nama sentrifugasi seimbang dalam tingkat kerapatan
atau equilibrium density-gradient centrifugation.Percobaan ini dilaporkan hasilnya
padatahun1958 oleh M.S. Meselson dan F.W. Stahl.
Garpu replikasi
Garpu replikasi atau cabang replikasi (replication fork) ialah struktur
yangterbentuk ketika DNA bereplikasi. Garpu replikasi ini dibentuk akibat enzim
helikase yang memutus ikatan-ikatan hidrogen yang menyatukan kedua untaian DNA,
membuat terbukanya untaian ganda tersebut menjadi dua cabang yang masing-
masing terdiri dari sebuah untaian tunggal DNA. Masing-masing cabang tersebut
menjadi "cetakan" untuk pembentukan dua untaian DNA baru berdasarkan urutan
nukleotida komplementernya. DNA polimerase membentuk untaian DNA baru dengan
memperpanjang oligonukleotida yang dibentuk oleh enzim primase dan disebut
primer. DNA polimerase membentuk untaian DNA baru dengan menambahkan
nukleotida—dalam hal ini, deoksiribonukleotida—ke ujung 3'-hidroksil bebas

BIOKIMIA II Ririn Vidiastuti
nukleotida rantai DNA yang sedang tumbuh. Dengan kata lain, rantai DNA baru
disintesis dari arah 5'→3', sedangkan DNA polimerase bergerak pada DNA "induk"
dengan arah 3'→5'. Namun demikian, salah satu untaian DNA induk pada garpu
replikasi berorientasi 3'→5', sementara untaian lainnya berorientasi 5'→3', dan
helikase bergerak membuka untaian rangkap DNA dengan arah 5'→3'. Oleh karena
itu, replikasi harus berlangsung pada kedua arah berlawanan tersebut.
Pembentukan leading strand
Pada replikasi DNA, untaian pengawal (leading strand) ialah untaian DNA yang
disintesis dengan arah 5'→3' secara berkesinambungan. Pada untaian ini, DNA
polimerase mampu membentuk DNA menggunakan ujung 3'-OH bebas dari sebuah
primer RNA dan sintesis DNA berlangsung secara berkesinambungan, searah dengan
arah pergerakan garpu replikasi.
Pembentukan lagging strand
Lagging strand ialah untaian DNA yang terletak pada sisi yang berseberangan
dengan leading strand pada garpu replikasi. Untaian ini disintesis dalam segmen-
segmen yang disebut fragmen Okazaki. Pada untaian ini, primase membentuk primer
RNA. DNA polimerase dengan demikian dapat menggunakan gugus OH 3' bebas pada
primer RNA tersebut untuk mensintesis DNA dengan arah 5'→3'. Fragmen primer RNA
tersebut lalu disingkirkan (misalnya dengan RNase H dan DNA Polimerase I) dan
deoksiribonukleotida baru ditambahkan untuk mengisi celah yang tadinya ditempati
oleh RNA. DNA ligase lalu menyambungkan fragmen-fragmen Okazaki tersebut
sehingga sintesis lagging strand menjadi lengkap.
Dinamika pada garpu replikasi
Bukti-bukti yang ditemukan belakangan ini menunjukkan bahwa enzim dan
protein yang terlibat dalam replikasi DNA tetap berada pada garpu replikasi
sementara DNA membentuk gelung untuk mempertahankan pembentukan DNA ke
dua arah. Hal ini merupakan akibat dari interaksi antara DNA polimerase, sliding
clamp, dan clamp loader.
Sliding clamp pada semua jenis makhluk hidup memiliki struktur serupa dan
mampu berinteraksi dengan berbagai DNA polimerase prosesif maupun non-prosesif
yang ditemukan di sel. Selain itu, sliding clamp berfungsi sebagai suatu faktor
prosesivitas. Ujung-C sliding clamp membentuk gelungan yang mampu berinteraksi
dengan protein-protein lain yang terlibat dalam replikasi DNA (seperti DNA polimerase
dan clamp loader). Bagian dalam sliding clamp memungkinkan DNA bergerak

BIOKIMIA II Ririn Vidiastuti
melaluinya. Sliding clamp tidak membentuk interaksi spesifik dengan DNA. Terdapat
lubang 35A besar di tengah clamp ini. Lubang tersebut berukuran sesuai untuk dilalui
DNA dan air menempati tempat sisanya sehingga clamp dapat bergeser pada
sepanjang DNA. Begitu polimerase mencapai ujung templat atau mendeteksi DNA
berutas ganda (lihat di bawah), sliding clamp mengalami perubahan konformasi yang
melepaskan DNA polimerase.Clamp loader merupakan protein bersubunit banyak
yang mampu menempel pada sliding clamp dan DNA polimerase. Dengan hidrolisis
ATP, clamp loader terlepas dari sliding clamp sehingga DNA polimerase menempel
pada sliding clamp. Sliding clamp hanya dapat berikatan pada polimerase selama
terjadinya sintesis utas tunggal DNA. Jika DNA rantai tunggal sudah habis, polimerase
mampu berikatan dengan subunit pada clamp loader dan bergerak ke posisi baru
pada lagging strand. Pada leading strand, DNA polimerase III bergabung dengan
clamp loader dan berikatan dengan sliding clamp.
Replikasi diprokariota dan eukariota
Replikasi DNA prokariota\Replikasi DNA kromosom prokariota, khususnya
bakteri, sangat berkaitan dengan siklus pertumbuhannya. Daerah ori pada E. coli,
misalnya, berisi empat buah tempat pengikatan protein inisiator DnaA, yang masing-
masing panjangnya 9 pb. Sintesis protein DnaA ini sejalan dengan laju pertumbuhan
bakteri sehingga inisiasi replikasi juga sejalan dengan laju pertumbuhan bakteri. Pada
laju pertumbuhan sel yang sangat tinggi; DNA kromosom prokariota dapat mengalami
reinisiasi replikasi pada dua ori yang baru terbentuk sebelum putaran replikasi yang
pertama berakhir. Akibatnya, sel-sel hasil pembelahan akan menerima kromosom
yang sebagian telah bereplikasi.
Protein DnaA membentuk struktur kompleks yang terdiri atas 30 hingga 40
buah molekul, yang masing-masing akan terikat pada molekul ATP. Daerah ori akan
mengelilingi kompleks DnaA-ATP tersebut. Proses ini memerlukan kondisi superkoiling
negatif DNA (pilinan kedua untai DNA berbalik arah sehingga terbuka). Superkoiling
negatif akan menyebabkan pembukaan tiga sekuens repetitif sepanjang 13 pb yang
kaya dengan AT sehingga memungkinkan terjadinya pengikatan protein DnaB, yang
merupakan enzim helikase, yaitu enzim yang akan menggunakan energi ATP hasil
hidrolisis untuk bergerak di sepanjang kedua untai DNA dan memisahkannya.
Untai DNA tunggal hasil pemisahan oleh helikase selanjutnya diselubungi oleh
protein pengikat untai tunggal atau single-stranded binding protein (Ssb) untuk

BIOKIMIA II Ririn Vidiastuti
melindungi DNA untai tunggal dari kerusakan fisik dan mencegah renaturasi. Enzim
DNA primase kemudian akan menempel pada DNA dan menyintesis RNA primer yang
pendek untuk memulai atau menginisiasi sintesis pada untai pengarah. Agar replikasi
dapat terus berjalan menjauhi ori, diperlukan enzim helikase selain DnaB. Hal ini
karena pembukaan heliks akan diikuti oleh pembentukan putaran baru berupa
superkoiling positif. Superkoiling negatif yang terjadi secara alami ternyata tidak
cukup untuk mengimbanginya sehingga diperlukan enzim lain, yaitu topoisomerase
tipe II yang disebut dengan DNA girase. Enzim DNA girase ini merupakan target
serangan antibiotik sehingga pemberian antibiotik dapat mencegah berlanjutnya
replikasi DNA bakteri.
Seperti telah dijelaskan di atas, replikasi DNA terjadi baik pada untai pengarah
maupun pada untai tertinggal. Pada untai tertinggal suatu kompleks yang disebut
primosom akan menyintesis sejumlah RNA primer dengan interval 1.000 hingga 2.000
basa. Primosom terdiri atas helikase DnaB dan DNA primase. Primer baik pada untai
pengarah maupun pada untai tertinggal akan mengalami elongasi dengan bantuan
holoenzim DNA polimerase III. Kompleks multisubunit ini merupakan dimer, separuh
akan bekerja pada untai pengarah dan separuh lainnya bekerja pada untai tertinggal.
Dengan demikian, sintesis pada kedua untai akan berjalan dengan kecepatan yang
sama. Masing-masing bagian dimer pada kedua untai tersebut terdiri atas subunit a,
yang mempunyai fungsi polimerase sesungguhnya, dan subunit e, yang mempunyai
fungsi penyuntingan berupa eksonuklease 3’– 5’. Selain itu, terdapatsubunit b yang
menempelkan polimerase pada DNA. Begitu primer pada untai tertinggal dielongasi
oleh DNA polimerase III, mereka akan segera dibuang dan celah yang ditimbulkan
oleh hilangnya primer tersebut diisi oleh DNA polimerase I, yang mempunyai aktivitas
polimerase 5’ – 3’, eksonuklease 5’ – 3’, dan eksonuklease penyuntingan 3’ – 5’.
Eksonuklease 5’ - 3’ membuang primer, sedangkan polimerase akan mengisi celah
yang ditimbulkan. Akhirnya, fragmen-fragmen Okazaki akan dipersatukan oleh enzim
DNA ligase. Secara in vivo, dimer holoenzim DNA polimerase III dan primosom diyakini
membentuk kompleks berukuran besar yang disebut dengan replisom. Dengan
adanya replisom sintesis DNA akan berlangsung dengan kecepatan 900 pb tiap detik.
Kedua garpu replikasi akan bertemu kira-kira pada posisi 180°C dari ori. Di
sekitar daerah ini terdapat sejumlah terminator yang akan menghentikan gerakan
garpu replikasi. Terminator tersebut antara lain berupa produk gen tus, suatu inhibitor
bagi helikase DnaB. Ketika replikasi selesai, kedua lingkaran hasil replikasi masih
menyatu. Pemisahan dilakukan oleh enzim topoisomerase IV. Masing-masing

BIOKIMIA II Ririn Vidiastuti
lingkaran hasil replikasi kemudian disegregasikanke dalam kedua sel hasil
pembelahan. Replikasi DNA eukariota
Pada eukariota, replikasi DNA hanya terjadi pada fase S di dalam interfase.
Untuk memasuki fase S diperlukan regulasi oleh sistem protein kompleks yang
disebut siklin dan kinase tergantung siklin atau cyclin-dependent protein kinases
(CDKs), yang berturut-turut akan diaktivasi oleh sinyal pertumbuhan yang mencapai
permukaan sel. Beberapa CDKs akan melakukan fosforilasi dan mengaktifkan protein-
protein yang diperlukan untuk inisiasi pada masing-masing ori.Berhubung dengan
kompleksitas struktur kromatin, garpu replikasi pada eukariota bergerak hanya
dengan kecepatan 50 pb tiap detik. Sebelum melakukan penyalinan, DNA harus
dilepaskan dari nukleosom pada garpu replikasi sehingga gerakan garpu replikasi
akan diperlambat menjadi sekitar 50 pb tiap detik. Dengan kecepatan seperti ini
diperlukan waktu sekitar 30 hari untuk menyalin molekul DNA kromosom pada
kebanyakan mamalia.
Sederetan sekuens tandem yang terdiri atas 20 hingga 50 replikon mengalami
inisiasi secara serempak pada waktu tertentu selama fase S. Deretan yang mengalami
inisasi paling awal adalah eukromatin, sedangkan deretan yang agak lambat adalah
heterokromatin. Daerah sentromer dan telomer dari DNA bereplikasi paling lambat.
Pola semacam ini mencerminkan aksesibilitas struktur kromatin yang berbeda-beda
terhadap faktor inisiasi.
Seperti halnya pada prokariota, satu atau beberapa DNA helikase dan Ssb yang
disebut dengan protein replikasi A atau replication protein A (RP-A) diperlukan untuk
memisahkan kedua untai DNA. Selanjutnya, tiga DNA polimerase yang berbeda
terlibat dalam elongasi. Untai pengarah dan masing-masing fragmen untai tertinggal
diinisiasi oleh RNA primer dengan bantuan aktivitas primase yang merupakan bagian
integral enzim DNA polimerase a. Enzim ini akan meneruskan elongasi replikasi tetapi
kemudian segera digantikan oleh DNA polimerase d pada untai pengarah dan DNA
polimerase e pada untai tertinggal. Baik DNA polimerase d maupun e mempunyai
fungsi penyuntingan. Kemampuan DNA polimerase d untuk menyintesis DNA yang
panjang disebabkan oleh adanya antigen perbanyakan nuklear sel atau proliferating
cell nuclear antigen (PCNA), yang fungsinya setara dengan subunit b holoenzim DNA
polimerase III pada E. coli. Selain terjadi penggandaan DNA, kandungan histon di
dalam sel juga mengalami penggandaan selama fase S.
Mesin replikasi yang terdiri atas semua enzim dan DNA yang berkaitan dengan
garpu replikasi akan diimobilisasi di dalam matriks nuklear. Mesin-mesin tersebut

BIOKIMIA II Ririn Vidiastuti
dapat divisualisasikan menggunakan mikroskop dengan melabeli DNA yang sedang
bereplikasi. Pelabelan dilakukan menggunakan analog timidin, yaitu
bromodeoksiuridin (BUdR), dan visualisasi DNA yang dilabeli tersebut dilakukan
dengan imunofloresensi menggunakan antibodi yang mengenali BUdR.
Ujung kromosom linier tidak dapat direplikasi sepenuhnya karena tidak ada DNA
yang dapat menggantikan RNA primer yang dibuang dari ujung 5’ untai tertinggal.
Dengan demikian, informasi genetik dapat hilang dari DNA. Untuk mengatasi hal ini,
ujung kromosom eukariota (telomer) mengandung beratus-ratus sekuens repetitif
sederhana yang tidak berisi informasi genetik dengan ujung 3’ melampaui ujung 5’.
Enzim telomerase mengandung molekul RNA pendek, yang sebagian sekuensnya
komplementer dengan sekuens repetitif tersebut. RNA ini akan bertindak sebagai
cetakan (templat) bagi penambahan sekuens repetitif pada ujung 3’.
Hal yang menarik adalah bahwa aktivitas telomerase mengalami penekanan di
dalam sel-sel somatis pada organisme multiseluler, yang lambat laun akan
menyebabkan pemendekan kromosom pada tiap generasi sel. Ketika pemendekan
mencapai DNA yang membawa informasi genetik, sel-sel akan menjadi layu dan mati.
Fenomena ini diduga sangat penting di dalam proses penuaan sel. Selain itu,
kemampuan penggandaan yang tidak terkendali pada kebanyakan sel kanker juga
berkaitan dengan reaktivasi enzim telomerase.
Gambar 6. DNA
Replikasi DNA. Mula-mula, heliks ganda DNA (merah) dibuka menjadi dua untai
tunggal oleh enzim helikase (9) dengan bantuan topoisomerase (11) yang
mengurangi tegangan untai DNA. Untaian DNA tunggal dilekati oleh protein-protein
pengikat untaian tunggal (10) untuk mencegahnya membentuk heliks ganda kembali.
Primase (6) membentuk oligonukleotida RNA yang disebut primer (5) dan molekul

BIOKIMIA II Ririn Vidiastuti
DNA polimerase (3 & 8) melekat pada seuntai tunggal DNA dan bergerak sepanjang
untai tersebut memperpanjang primer, membentuk untaian tunggal DNA baru yang
disebut leading strand (2) dan lagging strand (1). DNA polimerase yang membentuk
lagging strand harus mensintesis segmen-segmen polinukleotida diskontinu (disebut
fragmen Okazaki (7)). Enzim DNA ligase (4) kemudian menyambungkan potongan-
potongan lagging strand tersebut.
(Anonymous a,2012)
Jenis RNA
RNA dapat dibedakan menjadi dua kelompok utama, yaitu RNA genetik dan RNA
non-genetik.
1. RNA genetik
RNA genetik memiliki fungsi yang sama dengan DNA, yaitu sebagai pembawa
keterangan genetik. RNA genetik hanya ditemukan pada makhluk hidup tertentu yang
tidak memiliki DNA, misalnya virus. Dalam hal ini fungsi RNA menjadi sama dengan
DNA, baik sebagai materi genetik maupun dalam mengatur aktivitas sel.
2. RNA non-genetik
RNA non-genetik tidak berperan sebagai pembawa keterangan genetik sehingga
RNA jenis ini hanya dimiliki oleh makhluk hidup yang juga memiliki DNA. Berdasarkan
letak dan fungsinya, RNA non-genetik dibedakan menjadi mRNA, tRNA, dan rRNA.
1) mRNA (messenger RNA) atau ARNd (ARN duta)
mRNA merupakan RNA yang urutan basanya komplementer (berpasangan)
dengan salah satu urutan basa rantai DNA. RNA jenis ini merupakan polinukleotida
berbentuk pita tunggal linier dan disintesis oleh DNA di dalam nukleus. Panjang
pendeknya mRNA berhubungan dengan panjang pendeknya rantai polipeptida yang
akan disusun. Urutan asam amino yang menyusun rantai polipeptida itu sesuai
dengan urutan kodon yang terdapat di dalam molekul mRNA yang bersangkutan.
mRNA bertindak sebagai pola cetakan pembentuk polipeptida. Adapun fungsi utama
mRNA adalah membawa kode-kode genetik dari DNA di inti sel menuju ke ribosom di
sitoplasma. mRNA ini dibentuk bila diperlukan dan jika tugasnya selesai, maka akan
dihancurkan dalam plasma.
2) tRNA (transfer RNA) atau ARNt (ARN transfer)
RNA jenis ini dibentuk di dalam nukleus, tetapi menempatkan diri di dalam
sitoplasma. tRNA merupakan RNA terpendek dan bertindak sebagai penerjemah
kodon dari mRNA. Fungsi lain tRNA adalah mengikat asam-asam amino di dalam

BIOKIMIA II Ririn Vidiastuti
sitoplasma yang akan disusun menjadi protein dan mengangkutnya ke ribosom.
Bagian tRNA yang berhubungan dengan kodon dinamakan antikodon.
3) rRNA (ribosomal RNA) atau ARNr (ARN ribosomal)
RNA ini disebut ribosomal RNA karena terdapat di ribosom meskipun dibuat di
dalam nukleus. RNA ini berupa pita tunggal, tidak bercabang, dan fleksibel. Lebih dari
80% RNA merupakan rRNA. Fungsi dari RNA ribosom adalah sebagai mesin perakit
dalam sintesis protein yang bergerak ke satu arah sepanjang mRNA. Di dalam
ribosom, molekul rRNA ini mencapai 30-46%.
Dengan adanya penjelasan materi di atas, terdapat perbedaan antara DNA dan
RNA.
Tabel . Perbedaan DNA dan RNA

BIOKIMIA II Ririn Vidiastuti
Sintesis
RNA
Sintesis
RNA (atau
transkripsi) dapat didefinisikan sebagai proses mentransfer informasi dari urutan
DNA (Deoxyribo
Nukleat Acid)
RNA (Ribo
Nukleat Acid)
-
Letak
Dalam inti sel,
mitokondria, kloroplas,
senriol.
Dalam inti sel,
sitoplasma dan
ribosom.
-
Bentuk
Polinukleotida
ganda yang terpilin
panjang
Polinukleotida
tunggal dan pendekl
-
Gula
Deoxyribosa Ribosa
-
Basanya
Golongan purin :
adenine dan guanine
Golongan
pirimidin : cytosine dan
timin
Golongan purin :
adenine dan guanine
Golongan
pirimidin : cytosine dan
urasil
-
Fungsi
- mengontrol
sifat yang menurun
- sintesis
protein
- sintesis RNA
- sintesis
protein
-
Kadarnya
Tidak dipengaruhi
sintesis protein.
Letak basa
nitrogen dari kedua pita
ADN saling berhadapan
dengan pasangan yang
tetap yaitu Adenin selalu
berpasangan dengan
Timin, Cytosin dengan
Guanin. Kedua pita itu
diikatkan oleh ikatan
hidrogen.
Dipengaruhi
sintesis protein.
Macam ARN :
ARN duta
ARN ribosom
ARN transfer

BIOKIMIA II Ririn Vidiastuti
nukleotida DNA ke urutan RNA.Dalam organisme multi-selular (eukariota), mereka
telah berevolusi jauh lebih kompleks mekanisme pengaturan transkripsi dari
organisme uniseluler (prokariota).Sebuah enzim besar yang dikenal sebagai
transkripsi RNA polimerase mengkatalisis. Dengan peran yang berbeda dan sifat,
berbagai transkripsi RNA polimerase mengkatalisis tipe RNA yang berbeda. Meskipun
ada perbedaan besar dalam ukuran dan jumlah subunit polipeptida, struktur
keseluruhan dari polimerase RNA di prokariota dan eukariota cukup mirip,
mengungkap asal evolusi yang sama.
Berikut ini adalah garis besar dari RNA polimerase pada eukariota:
RNA polimerase I terletak di nucleolus dan mentranskripsi ribosomal RNA
(rRNA). Ribosomal RNA, bersama dengan berbagai bentuk protein ribosom. Satu rRNA
tertentu adalah katalis untuk pembentukan ikatan peptida. Berbagai jenis rRNA
berbagai ukuran dari 120-4700 basis amina. Eukariotik dan prokariotik rRNA yang
jelas berbeda, namun kedua spesies berumur panjang dan stabil.
RNA polimerase II lokal dengan inti, dan mentranskripsi messenger RNA (mRNA)
dan RNA nuklir paling kecil. Messenger RNA adalah pembawa informasi genetik pada
struktur primer protein dari DNA, bersama dengan fitur khusus yang memungkinkan
untuk menempel ribosom dan fungsi dalam sintesis protein. Ukurannya tergantung
pada ukuran protein untuk yang kode. Ini cenderung relatif pendek-hidup, dan umur
bervariasi dari spesies secara molekuler spesies molekul tergantung pada peran
biologis protein.
RNA polimerase III mentranskripsi RNA transfer (tRNA) dan RNA kecil lainnya.
tRNA adalah molekul kecil (65-110 nukleotida) dirancang untuk membawa asam
amino diaktifkan untuk tempat sintesis protein, ribosom. Ini adalah berumur panjang
dan stabil.
Transkripsi berlangsung dalam tiga (3) tahap:
1. Inisiasi,
2. Perpanjangan, dan
3. Penghentian.
Berikut ini menjelaskan transkripsi eukariotik:
Pada tahap inisiasi, RNA polimerase DNA pencarian untuk situs inisiasi, juga
disebut situs promotor, atau promotor. Pada tahap ini, DNA untai ganda ("tertutup").
Ini RNA polimerase / luka-struktur DNA yang disebut sebagai kompleks ditutup.

BIOKIMIA II Ririn Vidiastuti
polimerase RNA kemudian unwinds bentangan pendek DNA heliks ganda untuk
menghasilkan beruntai tunggal ("terbuka") template DNA dari yang dibutuhkan
petunjuk. Ini RNA polimerase / diterminasi-struktur DNA disebut kompleks terbuka.
Pada tahap perpanjangan, RNA polimerase memilih trifosfat ribonucleoside
benar dan mengkatalisis pembentukan ikatan fosfodiester. Proses ini diulangi
sebanyak yang enzim unidirectionally bergerak sepanjang template DNA. transkrip A
disintesis dari awal sampai akhir oleh molekul RNA polimerase tunggal.
Pada tahap terminasi, polimerase RNA mendeteksi sinyal terminasi yang
menentukan di mana transkrip berakhir.Kimia sintesis RNA adalah identik untuk
semua bentuk RNA, termasuk messenger RNA, transfer RNA, dan RNA ribosomal.
Langkah-langkah dasar hanya dijelaskan juga berlaku untuk semua bentuk. proses
sintetik mereka berbeda terutama dalam peraturan, pengolahan posttranscriptional,
dan polimerase khusus yang berpartisipasi.
RNA sintesis dapat dirangsang dengan pemberian RNA eksogen. Ditunjukkan
dalam beberapa percobaan, al Grabowska et. menentukan tingkat penggabungan 5p
uridin radioaktif-trifosfat (UTP) ke dalam RNA disintesis (dikembangkan dalam sistem
sel-bebas dengan kromatin dari hati tikus dan DNA polimerase RNA bergantung dari E
coli). Mereka menemukan transkripsi yang meningkat lima kali lipat setelah
penambahan hati tikus RNA [6]. Kanehisa et al. and Dobrzelewski et al. memperoleh
hasil yang sama dalam sistem dari DNA ayam hidup atau betis kromatin timus dan
polimerase RNA dari E coli.
(Dendhi, 2012)
2.4 Proses transkripsi dan translokasi
Transkripsi
Transkripsi merupakan pembentukan/sintesis RNA dari salah satu rantai DNA,
sehingga terjadi proses pemindahan informasi genetik dari DNA ke RNA. Fungsi ini
disebut fungsi heterokatalis DNA karena DNA mampu mensintesis senyawa lain
yaitu RNA. Sebuah rantai DNA digunakan untuk mencetak rantai tunggal mRNA
dengan bantuan enzim polimerase. Enzim tersebut menempel pada kodon
permulaan, umumnya adalah kodon untuk asam amino metionin. Pertama-tama,
ikatan hidrogen di bagian DNA yang disalin terbuka. Akibatnya, dua utas DNA
berpisah. Salah satu polinukleotida berfungsi sebagai pencetak atau sense, yang lain
sebagai gen atau antisense. Misalnya pencetak memiliki urutan basa G-A-G-A-C-T,
dan yang berfungsi sebagai gen memiliki urutan basa komplemen C-T-C-T-G-A.
Karena pencetaknya G-A-G-A-C-T, maka RNA hasil cetakannya C-U-C-U-G-A. Jadi, RNA

BIOKIMIA II Ririn Vidiastuti
C-U-C-U-G-A merupakan hasil kopian dari DNA C-T-C-T-G-A (gen), dan merupakan
komplemen dari pencetak.
Transkripsi DNA akan menghasilkan mRNA (messenger RNA). Pada organisme
eukariot, mRNA yang dihasilkan itu tidak langsung dapat berfungsi dalam sintesis
polipeptida, sebab masih mengandung segmen-segmen yang tidak berfungsi yang
disebut intron. Sedangkan segmen-segmen yang berfungsi untuk sintesis protein
disebut ekson. Di dalam nukleus terjadi pematangan/pemasakan mRNA yaitu dengan
jalan melepaskan segmen-segmen intron dan merangkaikan segmen-segmen ekson.
Gabungan segmen-segmen ekson membentuk satu rantai/utas mRNA yang
mengandung sejumlah kodon untuk penyusunan polipeptida. Rantai mRNA ini dikenal
sebagai sistron.
Gambar 7. Proses pematangan mRNA dengan membuang bagian
intron
Proses transkripsi ini terjadi di dalam inti sel (nukleus). DNA tetap berada di
dalam nukleus, sedangkan hasil transkripsinya dikeluarkan dari nukleus menuju
sitoplasma dan melekat pada ribosom. Ini dimaksudkan agar gen asli tetap
terlindung, sementara hasil kopinya ditugaskan untuk melaksanakan pesan-pesan
yang dikandungnya. Jika RNA rusak, akan segera diganti dengan hasil kopian yang
baru
1. Inisiasi (permulaan)
Daerah DNA di mana RNA polimerase melekat dan mengawali transkripsi
disebut sebagai promoter. Suatu promoter menentukan di mana transkripsi dimulai,
juga menentukan yang mana dari kedua untai heliks DNA yang digunakan sebagai
cetakan.

BIOKIMIA II Ririn Vidiastuti
2. Elongasi (pemanjangan)
Saat RNA bergerak di sepanjang DNA, RNA membuka untaian heliks ganda DNA
dengan bantuan enzim polimerase, sehingga terbentuklah molekul RNA yang akan
lepas dari cetakan DNA-nya.
3. Terminasi (pengakhiran)
Transkripsi berlangsung sampai RNA polimerase mentranskripsi urutan DNA
yang disebut terminator. Terminator yang ditranskripsi merupakan suatu urutan
RNA yang berfungsi sebagai kodon terminasi (kode stop) yang sesungguhnya. Pada
sel prokariotik, transkripsi biasanya berhenti tepat pada akhir kodon terminasi, yaitu
ketika polimerase mencapai titik terminasi sambil melepas RNA dan DNA. Sebaliknya,
pada sel eukariotik polimerase terus melewati sinyal terminasi, suatu urutan AAUAAA
di dalam mRNA. Pada titik yang jauh kira-kira 10 hingga 35 nukleotida, mRNA ini
dipotong hingga terlepas dari enzim tersebut.
(Anonymous b, 2012)
Translokasi
Translokasi merupakan mutasi kromosom dimana sebagian dari salah satu
kromosom ditransfer ke kromosom lain. Translokasi adalah perpindahan bahan
terlarut yang dapat terjadi di seluruh bagian tumbuhan. Translokasi ini membahas
yang terjadi pada Floem. Ada dua macam translokasi yaitu translokasi resiprok dan
translokasi Robertsonian.
Gambar 8. Translokasi
Mekanisme dan Pola Translokasi
Sejak lama para ahli fisiologi tumbuhan bermaksud mengukur langsung
translokasi dalam system pengangkutan dengan cara mengikuti pergerakan bahan
bertanda. Mula – mula menggunakan zat warna : fluoresein bergerak dengan mudah
dalam sel floem dan masih digunakan sebagai perunut yang efektif. Virus dan
herbisida juga pernah digunakan. Penggunakan fosfor, belerang, klorin, kalsium,
stronsium, rubidium, kalium, hydrogen dalam kajian ini, namun hingga saat ini nuklida
radioaktif yang paling penting.
BIOKIMIA II Ririn Vidiastuti
Perunut radioaktif bisa dilacak perjalannya dengan pelacak radiasi yang
disentuhkan pada batang atau bagian lain dari tumbuhan. Metode lainnya adalah
autoradiografi. Tumbuhan diletakkan bersinggungan dengan sehelai film sinar – X
selama beberapa hari hingga bulan. Kemudian,film tersebut dikembangkan dan
ditemui letak radioaktivitasnya pada tanaman tersebut.
Model E. Munch di Jerman pada tahun 1926 adalah model pengangkutan floem
yang dianut sampai sekarang. Konsepnya yaitu model aliran – tekanan. Menggunakan
dua osmometer. Osmometer yang dilakukan di laboratorium direndam dalam larutan.
Osmometer pertama berisi larutan yang lebih pekat daripada larutan sekitar,
osmometer kedua berisi larutan kurang pekat dari osmometer pertama dan harus
lebih pekat dari medium sekelilingnya.
Osmometer pertama dialokasikan dengan daun (sebagai sumber); sedangkan
osmometer kedua dialokasikan dengan organ-organ penerima (sebagai limbung,
misal buah, jaringan meristem, dan akar). Perbedaan antara model osmometer
dengan pengangkutan floem yang sesungguhnya terletak pada sumber dan
lingbungnya. Pada daun, bahan terlarut yang telah terangkut segera ditambahkan
kembali dari hasil fotosintesis (phloem loading); dan bahan terlarut yang telah sampai
ke limbung akan dikeluarkan dari pembuluh floem (phloem unloading). Dimanfaatkan
untuk pertumbuhan atau ditimbun di organ penampung, misalnya dalam bentuk pati
atau lemak. Larutan perendam pada osmometer setara dengan bagian apoplas
tanaman, yakni dinding sel dan pembuluh xylem.
Material Translokasi
Fungsi floem adalah sebagai jaringan translokasi bahan organik yang terutama
berisi karbohidrat. Crafts dan Lorenz (1994) mendapatkan persentase nitrogen (dalam
bentuk protein) sebesar 45%. Sebenarnya gula yang menjadi linarut terbesar yang
ditranslokasikan dalam cairan floem. Diantara gula ini, sukrosa yang paling banyak
jumlahnya. Gula lain seperti gula rafinosa : glukosa, rafinosa, stakiosa, dan fruktosa
juga ada pada gula alcohol: manitol, sorbitol, galaktitol, serta mio-inositol.
Tingkat Pergerakan
Diestimasi dengan cara menghitung penambahan berat organ tersebut selama
kurun waktu tertentu untuk mengetahui laju pengangkutan melalui pembuluh floem
ke suatu organ. Kemudian diukur luas penampang melintang dari pembuluh floem.
Berdasarkan data tersebut dapat dihitung laju transfer massa (mass transfer rate).
Laju perpindahan masa merupakan jumlah bahan yang melintasi suatu irisan
melintang tabung taapis per satuan waktu.

BIOKIMIA II Ririn Vidiastuti
Dikemukakan oleh Alden S. Crafts dan O.Lorenz (dari University of California di Davis)
berasumsi bahwa bahan kering yang diangkut melalui floem mempunyai gravitas
spesifik (atau kepadatan) sebesar 1,5 g.cm-3.Nilai ini jika dibagi dengan laju transfer
massa akan diperoleh velositas sebesar 110 mm.jam-1. Tentu saja, bahan yang
diangkut dalam pembuluh floem tidak dalam bentuk kering, tetapi terlarut didalam
air. Dengan demikian velositas sesungguhnya adalah lebih cepat. Potensi osmotik
larutan floem yang umum terukur adalah antara -2 Mpa sampai -3 Mpa, yang setara
dengan 20% sampai 30% larutan sukrosa (Sukrosa merupakan bahan terlarut yang
dominan pada larutan floem. Berdasarkan nilai ini, maka volaritas pengangkutan pada
pembuluh floem adalah antara 363 sampai 550 mm.jam-1.
Dengan teknik yang lebih maju, pengukuran velositas dapat dilakukan dengan
isotop11C dalam bentuk CO2 yang diberikan pada daun. Isotop ini akan terkandung
dalam fotosintat yang akan diangkut melalui pembuluh floem. Pada 2 atau lebih posisi
pada batang ditempatkan pendeteksi radiasi dengan jarak yang telah ditetapkan.
Phloem Loading dan Unloading
Proses peningkatan konsentrasi gula pada sel-sel floem yang berada dekat
dengan sel-sel fotosintetik pada daun disebut proses pengisian floem (phloem
loading). Berdasarkan pengukuran pada berbagai spesies, terlihat bahwa potensi
osmotik sel-sel mesofil (sekitar -0,8 MPa sampai -1,8 MPa) lebih tinggi dibanding pada
pembuluh floem (antara -2,0 MPa sampai -3,0 MPa). Karena bahan terlarut (sukrosa)
pada pembuluh floem lebih tinggi dibanding pada sel-sel mesofil.
Serapan sukrosa oleh sel peneman floem ini yang dikarenakan oleh sel
peneman ini lebih besar dan lebih aktif dibandingkan sel-sel lain pada jaringan floem
dan juga adanya penumbuhan ke dalam (ingrowth) yang menyebabkan luas
permukaan membran sel ini menjadi 3 kali lebih luas. Menyebabkan potensi osmotic
sitoplasma sel ini menjadi turun (lebih negatif) dan ini akan merangsang air untuk
masuksecara osmosis kedalam sel ini dari sel-sel mesofil disekitarnya. Sebagai
akibatnya tekanan internal pada sel peneman akan meningkat dan mengakibatkan
sukrosa bergerak masuk ke pembuluh floem secara simplastik melalui
plasmodesmata. Masuknya larutan yang mengandung sukrosa ke pembuluh floem
dari sel-sel peneman ini yang mengakibatkan tekanan internal pada pembuluh floem
pada daun lebih tinggi, yang kemudian menjadi faktor pendorong dari aliran larutan
floem, berarti pengangkutan senyawa-senyawa yang terlarut didalamnya.
Proses pengisian embrane bersifat selektif. Jenis material yang di translokasi
seperti gula rafinosa : glukosa, rafinosa, dan stakiosa juga ada pada gula alcohol:

BIOKIMIA II Ririn Vidiastuti
manitol, sorbitol, galaktitol, serta mio-inositol. Fruktosa jarang diangkut kedalam
pembuluh floem. Demikian juga dengan asam amino dan mineral.sifat selektif ini
memperkuat argumentasi bahwa senyawa – senyawa yang akan dimuat kedalam
pembuluh floem diserap dari apoplas oleh sel – sel peneman floem. Sifat selektif ini
berkaitan dengan peranan senyawa pembawa pada embrane, yang menyangkut pada
senyawa – senyawa tertentu.
Kompetisi antara organ atau jaringan limbung ditentukan oleh laju pengeluaran bahan
dari pembuluh floem (phloem unloading). Limbung yang dapat memanfaatkan hasil
terlarut (sukrosa) dari pembuluh floem dan akan berpeluang besar untuk memperoleh
lebih banyak lagi bahan terlarut dari organ sumber. Hal ini disebabkan sukrosa
diserap sel – sel organ limbung dari pembuluh floem, maka potensi air sel – sel
limbung tersebut turun. Mengakibatkan air akan bergerak keluar dari pembuluh floem
dan tekanan internal pembuluh floem pada organ atau jaringan limbung akan turun.
Hal ini akan lebih memacu laju pengangkutan dari sumber ke limbung karena
perbedaan tekanan internal yang lebih besar antara kedua ujung pembuluh floem
tersebut.
( Lakitan, 1993)

BIOKIMIA II Ririn Vidiastuti
BAB III
PENUTUP
3.1 Kesimpulan
Bagaimana proses sintesis DNA dan RNA?
Bagaimana proses transkripsi dan translokasi?
Asam nukleat tersusun atas nukleotida, yang mana nukleotida tersusun
atas nukleosida dan fosfat. Nukleosida tersusun atas gula pentose dan basa nitrogen
(purin dan pirimidin).
Asam nukleat memiliki sifat kimia, yaitu stabilitas asam nukleat,
pengaruh asam, pengaruh alkali, denaturasi kimia, viskositas, dan kerapatan apung,
sedangkan sifat spektroskopik-termal asam nukleat meliputi kemampuan absorpsi
sinar UV, hipokromisitas, penghitungan konsentrasi asam nukleat, penentuan
kemurnian DNA, serta denaturasi termal dan renaturasi asam nukleat.
Proses sintesis DNA berupa replikasi DNA yang terdiri atas 3 macam, yaitu
model konservatif, model semikonservatif, dan model dispersif. Sedangkan sintesis
RNA terdiri atas RNA polimerase I terletak di nucleolus dan mentranskripsi ribosomal
RNA (rRNA), lalu, RNA polimerase II lokal dengan inti, dan mentranskripsi messenger
RNA (mRNA) dan RNA nuklir paling kecil. RNA polimerase III mentranskripsi RNA
transfer (tRNA) dan RNA kecil lainnya.
Proses Transkripsi terdiri atas insiasi, elongasi, dan terminasi.

BIOKIMIA II Ririn Vidiastuti
DAFTAR PUSTAKA
Anonymous a. 2012. Replikasi di prokariota dan eukariota.
http://www.google.com
Di akses 20 Mei 2012
Anonymous b. 2012. Transkripsi. http://desybio.wordpress.com/tag/1-
transkripsi/
Di akses 20 Mei 2012
Dendhi, Deny. 2012. Sintesis DNA.
http://denydendhi.blogspot.com/2011/03/replikasi-dna.html Di akses 20 Mei
2012
Lakitan, Benyamin.1993. Dasar – Dasar Fisiologi Tumbuhan. PT RajaGrafindo
Persada : Jakarta
Mustofa, Kharis. 2012. Komponen Asam Nukleat.
http://kharism.blog.unsoed.ac.id/
Di Akses 20 Mei 2012
Sumardjo, Damin. 2006. Pengantar Kimia : Buku Pnduan Kuliah Mahasiswa
Kedokteran dan Program Strata I Fakultas Bioeksata. Jakarta : Penerbit Buku
Kedokteran EGC
Susanto, Agus H. 2012. BAB II Asam Nukleat.
http://biomol.wordpress.com/bahan-ajar/asam-nukleat/ Di akses 20 Mei 2012