pemilihan jenis tanaman restorasi berdasarkan beberapa ... · klorofil sehingga laju...
TRANSCRIPT

5
TINJAUAN PUSTAKA
Fotosintesis
Proses utama fotosintesis terjadi di kloroplas. Pada tumbuhan C3 sebagian
besar kloroplas terdapat dalam sel mesofil daun. Proses reaksi fotosintesis pada
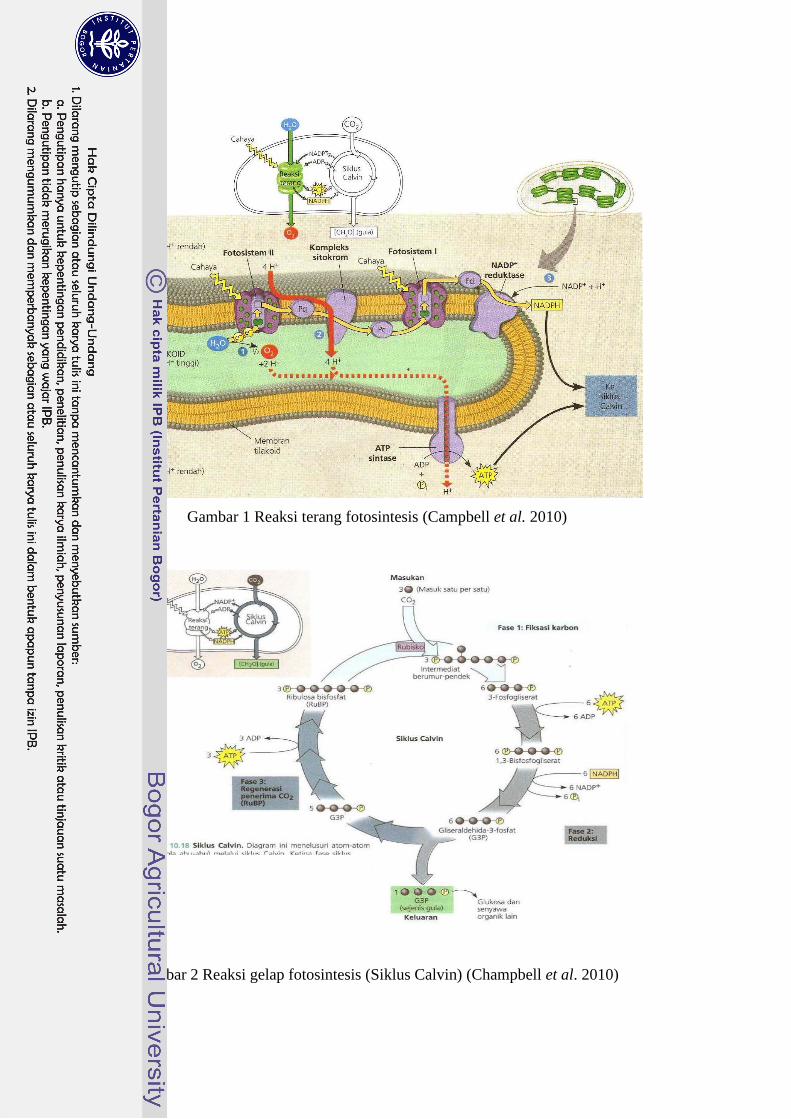
tumbuhan tingkat tinggi berlangsung dua tahap, yaitu: 1) reaksi terang (Gambar1),
dan 2) reaksi gelap (Gambar 2). Tahapan reaksi fotosintesis adalah sebagai berikut
(Taiz & Zeiger 2003; Lambers et al. 2008; Campbell et al. 2010).
1. Penyerapan foton oleh pigmen, terutama klorofil, bekerja sama dengan dua
fotosistem. Pigmen tersimpan dalam membran bagian dalam tilakoid dan
menyerap sebagian besar energi dari radiasi aktif fotosintesis (PAR =
Photosynthetically Active Radiation) pada panjang gelombang 400-700 nm.
Pigmen ini mentransfer energi eksitasi ke pusat reaksi pada fotosistem di mana
proses selanjutnya dimulai.
2. Elektron yang dihasilkan dari pemecahan molekul air dengan produksi oksigen
berkesinambungan ditranspor melalui rantai transpor elektron yang tertanam
dalam membran tilakoid. NADPH dan ATP yang dihasilkan dari proses ini
digunakan untuk proses reaksi gelap.
Kedua reaksi ini bergantung pada energi cahaya, oleh karena itu disebut reaksi
terang fotosintesis. Reaksi terang mengubah energi surya menjadi energi kimia
dalam ATP dan NADPH.
3. NADPH dan ATP digunakan dalam siklus reduksi karbon fotosintesis (siklus
Calvin). Dalam siklus ini CO2, yang masuk melalui stomata, diasimilasi
membentuk senyawa berkarbon tiga (triosa fosfat). Proses ini dapat
berlangsung tanpa cahaya, oleh karena itu disebut reaksi gelap. Proses ini
berlangsung di dalam stroma.
6
Gambar 1 Reaksi terang fotosintesis (Campbell et al. 2010)
Gambar 2 Reaksi gelap fotosintesis (Siklus Calvin) (Champbell et al. 2010)

7
Pada fotosintesis, tumbuhan memanfaatkan energi matahari untuk
mengoksidasi air untuk melepaskan O2, dan mereduksi CO2 untuk membentuk
senyawa karbon yang lebih besar, terutama gula (Champbell et al. 2010).
Faktor abiotik seperti cahaya, suhu, konsentrasi CO2, uap air, keberadaan
nutrisi memiliki pengaruh utama pada fotosintesis bersih, yang selanjutnya
mempengaruhi pertumbuhan dan produktivitas. Kapasitas fotosintesis tidak hanya
dipengaruhi oleh lingkungan abiotik tetapi juga dipengaruhi oleh umur dan posisi
daun pada kanopi. Jenis atau genus yang berbeda memiliki laju fotosintesis yang
berbeda. Daun Populus memiliki laju fotosintesis maksimum sebelum daun
berukuran maksimum, sedangkan daun Quercus mencapai laju fotosintesis
maksimum beberapa minggu setelah daun berukuran maksimum (Ceulemans &
Saugier 1991).
Laju fotosintesis daun tanaman dipengaruhi oleh beberapa faktor, yaitu:
a. Perbedaan jenis tumbuhan
Tumbuhan berdasarkan metabolisme fotosintesisnya dapat dibagi menjadi 3
golongan besar, yaitu jenis C-3, C-4, dan CAM (Crassulacean Acid Metabolism).
Tanaman kehutanan umumnya termasuk C-3. Tumbuhan C-4 secara umum
mempunyai laju fotosintesis tertinggi, sementara tumbuhan CAM memiliki laju
fotosintesis terendah (Lakitan 2010). Contoh tanaman C-4 seperti jagung, tebu,
dan sorgum. Nanas dan kaktus termasuk ke dalam tanaman CAM. Ketiga jenis
tumbuhan tersebut juga memiliki anatomi yang berbeda. Tumbuhan C-4 sel
seludangnya berkembang baik dan memiliki klorofil (Taiz & Zeiger 2003;
Campbell et al. 2010). Perbedaan dalam golongan tumbuhan yang sama juga
terjadi. Tanaman kacang tanah dan bunga matahari merupakan tanaman yang
yang senang cahaya matahari (sun-adapted), tingkat kejenuhan terhadap
cahayanya rendah. Tanaman C-4 dapat beradaptasi pada intensitas cahaya dan
suhu tinggi, dan jenis tanaman ini lebih efisien dalam memanfaatkan air dalam
kondisi tersebut. Tanaman C-3 cenderung mencapai puncak laju fotosintesis pada
intensitas cahaya dan suhu moderat, dan akan terganggu oleh suhu tinggi dan
intensitas cahaya penuh (Odum 1996).

8
b. Umur daun
Daun muda umumnya mempunyai kemampuan fotosintesis yang masih
rendah. Kemampuan fotosintesis akan meningkat dengan bertambahnya umur dan
luasan daun. Setelah ukuran daun mencapai maksimum, maka daun akan menjadi
tua dan berubah warna menjadi kuning karena klorofil mulai rusak. Rusaknya
klorofil akan menurunkan kemampuan fotosintesis daun (Salisbury & Ross 1995;
Lakitan 2010).
c. Letak daun
Daun yang terletak di bagian dalam tajuk kurang mendapat cahaya matahari.
Laju fotosintesis daun yang terletak di bagian dalam tajuk akan lebih rendah
dibanding dengan daun yang terletak di tepi luar tajuk (Salisbury & Ross 1995;
Lakitan 2010). Susunan daun dalam tajuk juga mempengaruhi efektifitas
penyerapan cahaya matahari. Jika letak daun mendatar dan sebagian besar cahaya
datang dari atas, maka daun bagian atas akan terpajan pada cahaya matahari
penuh, sehingga fotosintesis pada daun tersebut akan terlalu jenuh, dan banyak
cahaya yang diserap menjadi tidak berguna (Salisbury & Ross 1995).
d. Fase pertumbuhan
Tumbuhan yang sedang tumbuh, sedang berbunga, dan berbuah, memiliki
laju fotosintesis yang tinggi dan laju translokasi fotosintat yang juga tinggi.
Tumbuhan yang sedang dalam fase istirahat memiliki laju fotosintesis yang
rendah (Lakitan 2010).
e. Intensitas cahaya matahari
Setiap jenis tumbuhan membutuhkan intensitas cahaya matahari yang
berbeda-beda. Ada jenis tumbuhan yang pertumbuhannya baik pada cahaya
matahari penuh (sun plant), ada juga tumbuhan yang pertumbuhannya baik pada
kondisi ternaungi (shade plant). Bahwa cahaya sering membatasi fotosintesis
terlihat dengan menurunnya laju penambatan CO2 ketika tumbuhan terkena
bayangan awan sebentar. Daun naungan umumnya mempunyai klorofil lebih
banyak, khususnya klorofil b, terutama karena tiap kloroplas mempunyai lebih
banyak grana dibandingkan pada daun matahari (Salisbury & Ross 1995; Lambers
et al. 2008; Lakitan 2010).

9
f. Konsentrasi gas CO2
Gas CO2 merupakan bahan yang dibutuhkan untuk fotosintesis. Jika
konsentrasi gas meningkat, maka hasil fotosintesis akan meningkat pula. Akan
tetapi secara umum konsentrasi gas yang melebihi 1000-2000 ppm akan
berpengaruh buruk pada fotosintesis (Salisbury & Ross 1995; Lambers et al.
2008; Lakitan 2010).
g. Suhu udara
Rentang suhu yang memungkinkan tumbuhan berfotosintesis sangat luas.
Konifer dapat berfotosintesis sangat lambat pada suhu -6 oC atau lebih rendah.
Pengaruh suhu terhadap fotosintesis bergantung pada spesies, keadaan lingkungan
tempat tumbuh, dan keadaan lingkungan saat pengukuran. Secara umum, suhu
optimum untuk fotosintesis sama dengan suhu siang hari di tempat tumbuhan
tersebut biasa hidup. Enzim sensitif terhadap suhu. Proses reduksi karbondioksida
pada karbohidrat memiliki banyak reaksi enzim. Salah satu enzim yang terdapat
dalam daun dengan konsentrasi tinggi yaitu ribulosa bifosfat karboksilase
(Rubisco) (Salisbury & Ross 1995; Lakitan 2010).
h. Ketersediaan air
Air merupakan bahan baku fotosintesis selain CO2. Kekurangan air dapat
menghambat laju fotosintesis karena pengaruhnya terhadap turgiditas sel penjaga
stomata. Jika tumbuhan kekurangan air, maka turgiditas sel penjaga akan
menurun, sehingga stomata menutup. Menutupnya stomata akan menghambat
serapan CO2 (Hopkins & Hüner 2004; Lakitan 2010).
i. Unsur hara
Unsur hara secara tidak langsung mempengaruhi proses fotosintesis. Jika
unsur esensial tidak terpenuhi, laju fotosintesis akan berkurang. Kapasitas
fotosintesis umumnya sensitif dengan kandungan nitrogen. Nitrogen sangat
berperan dalam proses fotosintesis, yakni sebagai bagian dari molekul klorofil,
pembawa redoks pada rantai transpor elektron, dan berperan pada semua reaksi
enzimatis dalam metabolisme fotosintesis. Nitrogen juga menentukan
produktivitas primer (Hopkins & Hüner 2004). Tumbuhan menanggapi kurangnya
pasokan unsur esensial dengan menunjukkan gejala kekahatan yang khas. Pada
tanaman yang kekurangan Mg, karena Mg adalah bagian esensial molekul

10
klorofil, maka klorofil tak terbentuk tanpa Mg atau terbentuk dalam jumlah sedikit
bila konsentrasi Mg rendah (Salisbury & Ross 1995).
j. Kesehatan daun
Daun yang terserang penyakit tidak dapat melakukan fotosintesis dengan
baik. Penyakit pada daun akibat patogen dapat menyebabkan klorosis dan
nekrosis. Daun yang mengalami klorosis dan nekrosis akan mengalami degradasi
klorofil sehingga laju fotosintesisnya terhambat (Hopkins & Hüner 2004)
k. Polutan udara
Beberapa polutan dapat mempengaruhi fotosintesis, seperti gas SOx, NOx,
ozon,logam berat, dan hujan asam. Zat-zat tersebut dapat mengganggu proses
pembentukan atau mempengaruhi umur kloroplas, serta mengganggu proses
biokimia yang terjadi di daun (Hopkins & Hüner 2004). Lamanya pemaparan
polutan pada tumbuhan akan mengakibatkan terakumulasinya polutan tersebut.
Hal ini antara lain dapat terjadi pada sistem membran kloroplas, tempat proses
awal fotosintesis, terjadi perubahan pada strukturnya (Treshow & Anderson
1991).
Klorofil dan Karakteristiknya
Klorofil terdapat dalam kloroplas dalam jumlah banyak, yaitu pada
membran tilakoid, sering terikat longgar pada protein, tetapi mudah diekstraksi ke
dalam pelarut lipid seperti aseton dan eter (Harborne 1987).
Tumbuhan tingkat tinggi mengandung klorofil a dan klorofil b. Klorofil a
berwarna hijau tua dengan rumus kimia C55H72O5N4Mg, sedang klorofil b
berwarna hijau muda dengan rumus kimia C55H70O6N4Mg. Rumus bangunnya
berupa cincin yang terdiri atas 4 pirol (tetrapirol) dengan Mg sebagai inti. Struktur
klorofil a mempunyai gugus metil (CH3), sedangkan klorofil b mempunyai gugus
aldehida (CHO) (Harborne 1987; Champbell et al. 2010), seperti terlihat pada
Gambar 3.
Molekul klorofil terdiri dari dua bagian yaitu kepala porfirin dan rantai
hidrokarbon yang panjang. Porfirin adalah tetrapirol siklik, yang terdiri dari empat
nitrogen yang mengikat cincin pirol yang dihubungkan dengan empat rantai
metana (Champbell et al. 2010).

11
Faktor-faktor yang berpengaruh terhadap pembentukan klorofil yaitu:
1) Faktor genetik, 2) Cahaya. Pada beberapa kecambah tanaman Angiospermae,
klorofil dapat terbentuk tanpa cahaya. Tanaman lain yang ditumbuhkan di tempat
gelap tidak berhasil membentuk klorofil, atau mengalami klorosis. Terlalu banyak
cahaya berpengaruh buruk kepada klorofil. Larutan klorofil yang dihadapkan
kepada sinar kuat tampak berkurang hijaunya. Daun-daun yang terus-menerus
terpapar sinar matahari langsung, warnanya menjadi hijau kekuning-kuningan. 3)
Oksigen. Kecambah yang ditumbuhkan di tempat gelap tidak dapat membentuk
klorofil jika tidak diberikan oksigen kepadanya. 4) Karbohidrat dalam bentuk gula
sangat menolong dalam pembentukan klorofil, terutama pada daun yang tumbuh
di tempat gelap. 5) Nitrogen dan magnesium merupakan bahan pembentuk
klorofil, dan merupakan suatu keharusan. Kekurangan salah satu zat tersebut akan
mengakibatkan klorosis. 6) Air merupakan faktor keharusan. Kekurangan air
mengakibatkan desintegrasi klorofil, seperti terjadi pada rumput dan pohon-pohon
di musim kering. 7) Temperatur yang paling baik untuk pembentukan klorofil
adalah antara 26o-30
oC (Dwidjoseputro 1986).
Gambar 3 Struktur molekul klorofil (Chambell et al. 2010)

12
Molekul klorofil berperan sebagai antena untuk menangkap cahaya dan
mentransfer energi eksitasinya ke pusat reaksi fotosistem I. Sejumlah besar
klorofil terdapat dalam LHC (Light Harvesting Complex). Rasio klorofil a dan
klorofil b sekitar 1.12 untuk LHC. Klorofil b bekerja sama penuh dengan LHC
dan sedikit dengan kompleks PSI dan PSII. Daun terlihat berwarna hijau dalam
cahaya putih karena klorofil lebih efisien berfungsi sebagai penyerap cahaya pada
panjang gelombang biru dan merah daripada pada spektrum hijau, yang praktis
dipantulkan seluruhnya (Lambers et al. 2008) sehingga daun nampak berwarna
hijau. Cahaya (dalam hal ini satuan partikel cahaya terkecil yaitu foton) yang
ditangkap oleh klorofil akan mengeksitasi klorofil ke tingkat energi yang lebih
tinggi. Klorofil yang menyerap cahaya biru akan tereksitasi ke tingkat energi yang
lebih tinggi dari pada klorofil yang menyerap cahaya merah. Pada tingkat energi
yang lebih tinggi, klorofil dalam kondisi tidak stabil, sehingga akan melepaskan
energinya dalam bentuk panas ke lingkungan dan akan kembali ke tingkat eksitasi
yang lebih rendah, di mana tingkat eksitasi ini akan stabil selama maksimum
beberapa nanodetik (10-9
detik). Oleh karena itu, proses pemanenan energi harus
terjadi dalam waktu yang sangat singkat. Energi inilah yang digunakan untuk
proses fotosintesis (Taiz & Zeiger 2003).
Pada tingkat tereksitasi terendah ada beberapa kemungkinan penyaluran
energi terjadi. Kemungkinan pertama, energi akan dilepas perlahan secara radiatif
sebagai foton pada panjang gelombang yang lebih panjang melalui fluorosen.
Kemungkinan kedua, energi akan segera dilepaskan kembali dalam bentuk panas.
Kemungkinan ini umumnya yang cukup besar terjadi pada tumbuhan di alam.
Kemungkinan ketiga, klorofil mungkin akan menyalurkan energinya kepada
molekul lain seperti molekul oksigen. Kemungkinan keempat, energi akan
dimanfaatkan untuk reaksi fotokimia dari fotosintesis untuk membentuk ATP dan
NADPH (Taiz & Zeiger 2003).
Keempat bentuk penyaluran energi ini merupakan kemungkinan umum dari
mekanisme ebergi dalam sistem cahaya fotosintesis. Pada kondisi lingkungan
yang baik bagi tanaman, maka penyaluran dalam bentuk reaksi fotokimia relatif
besar, sehingga proses fotosintesis akan berjalan dengan laju yang tinggi, sejalan
dengan tingginya laju transpor elektron dalam reaksi terang fotosintesis. Namun

13
dalam keadaan lingkungan yang kurang menguntungkan, seperti cekaman
kekeringan, keasaman, dan suhu rendah/tinggi, penyaluran energi ke arah
fotokimia akan mengalami hambatan (Taiz & Zeiger 2003).
Prinsip dasar penyerapan cahaya, sering disebut Hukum Stark Einstein,
adalah bahwa setiap molekul hanya dapat menyerap satu foton setiap kali, dan
foton ini menyebabkan eksitasi satu elektron saja. Energi dalam tiap foton
berbanding terbalik dengan panjang gelombang. Cahaya biru dan ungu dengan
gelombang yang lebih pendek memiliki lebih banyak foton energetik dibanding
cahaya merah atau jingga dengan gelombang yang lebih panjang. Satu mol (6.02 x
1023
) foton sering disebut sebagai satu einstein (Salisbury & Ross 1995; Campbell
et al. 2010; Lakitan 2010).
Tumbuhan sebagai Penyerap CO2
Agus dan van Noordwijk (2007) melaporkan bahwa pembakaran hutan
alami pada lahan gambut menyebabkan pelepasan CO2 sebanyak 734 ton ha-1
yang berasal dari C yang tersimpan di vegetasi sebasar 200 ton ha-1
. Tetapi jumlah
tersebut mungkin masih lebih rendah dari jumlah CO2 yang diemisikan
sebenarnya, karena selama pembakaran hutan lapisan atas gambut juga terbakar
dan melepaskan CO2. Seandainya gambut yang terbakar setebal 10 cm, maka akan
terjadi penambahan emisi CO2 sebesar 220 ton ha-1
karena tanah gambut
mengandung C sekitar 6 ton ha-1
cm-1
.
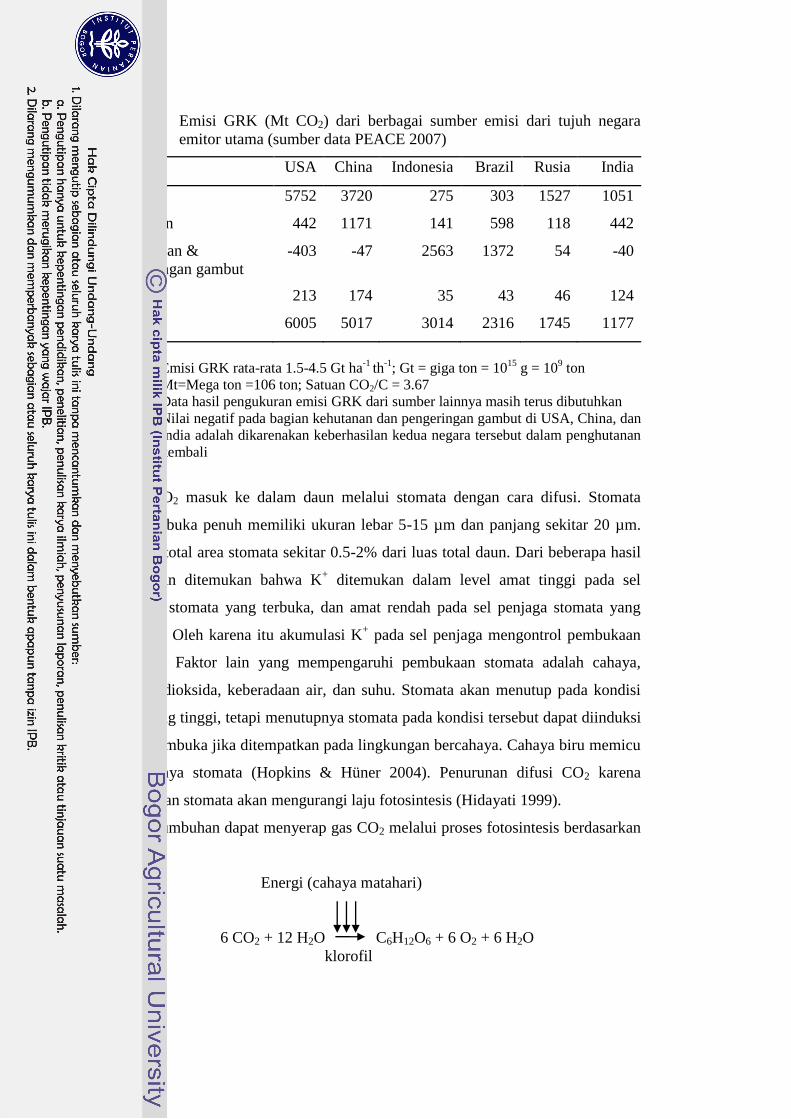
Pada tahun 2007 (PEACE 2007) Indonesia dianggap sebagai salah satu
negara penghasil emisi GRK terbesar di dunia. Emisi GRK terutama berasal dari
kegiatan alih guna lahan hutan dan pengeringan lahan gambut menjadi lahan
pertanian (Tabel 1).
Negara emitor GRK terbesar sebenarnya adalah USA dan China, jumlah
GRK yang diemisikan dua kali lipat lebih besar dari emisi asal Indonesia.
Bedanya, emisi GRK dari kedua negara industri tersebut berasal dari penggunaan
bahan bakar fosil dan industri (Hairiah 2008).
14
Tabel 1 Emisi GRK (Mt CO2) dari berbagai sumber emisi dari tujuh negara
emitor utama (sumber data PEACE 2007)
Emisi USA China Indonesia Brazil Rusia India
Energi 5752 3720 275 303 1527 1051
Pertanian 442 1171 141 598 118 442
Kehutanan &
pengeringan gambut
-403 -47 2563 1372 54 -40
Limbah 213 174 35 43 46 124
Total 6005 5017 3014 2316 1745 1177
Catatan:
1. Emisi GRK rata-rata 1.5-4.5 Gt ha-1
th-1
; Gt = giga ton = 1015
g = 109 ton
Mt=Mega ton =106 ton; Satuan CO2/C = 3.67
2. Data hasil pengukuran emisi GRK dari sumber lainnya masih terus dibutuhkan
3. Nilai negatif pada bagian kehutanan dan pengeringan gambut di USA, China, dan
India adalah dikarenakan keberhasilan kedua negara tersebut dalam penghutanan
kembali
CO2 masuk ke dalam daun melalui stomata dengan cara difusi. Stomata
yang terbuka penuh memiliki ukuran lebar 5-15 µm dan panjang sekitar 20 µm.
Jumlah total area stomata sekitar 0.5-2% dari luas total daun. Dari beberapa hasil
penelitian ditemukan bahwa K+ ditemukan dalam level amat tinggi pada sel
penjaga stomata yang terbuka, dan amat rendah pada sel penjaga stomata yang
tertutup. Oleh karena itu akumulasi K+ pada sel penjaga mengontrol pembukaan
stomata. Faktor lain yang mempengaruhi pembukaan stomata adalah cahaya,
karbon dioksida, keberadaan air, dan suhu. Stomata akan menutup pada kondisi
CO2 yang tinggi, tetapi menutupnya stomata pada kondisi tersebut dapat diinduksi
agar membuka jika ditempatkan pada lingkungan bercahaya. Cahaya biru memicu
terbukanya stomata (Hopkins & Hüner 2004). Penurunan difusi CO2 karena
penutupan stomata akan mengurangi laju fotosintesis (Hidayati 1999).
Tumbuhan dapat menyerap gas CO2 melalui proses fotosintesis berdasarkan
rumus:
Energi (cahaya matahari)
6 CO2 + 12 H2O C6H12O6 + 6 O2 + 6 H2O
klorofil

15
Dari persamaan tersebut menunjukkan bahwa fotosintesis merupakan reaksi
antara CO2 dan air untuk memproduksi glukosa, karbohidrat berkarbon 6 atau
disebut heksosa (Hopkins & Hüner 2004). Fotosintesis tidak dapat berjalan tanpa
energi dari sinar matahari dan molekul klorofil sebagai penangkap energinya.
Hutan mempunyai potensi yang besar dalam pengurangan kadar CO2
melalui konservasi dan manajemen hutan. Hutan sekunder muda dalam
pertumbuhannya memerlukan CO2 sehingga banyak dihasilkan biomassa
tumbuhan melalui proses fotosintesis hutan yang sedang tumbuh (Heriyanto &
Siregar 2007).
Fotosintesis dan Pertumbuhan
Pertumbuhan tanaman bergantung kepada kemampuan tanaman tersebut
untuk menyerap karbon di atmosfer dan mengolahnya menjadi senyawa organik
dengan memanfaatkan energi cahaya melalui proses fotosintesis. Kurang lebih
40% dari berat kering tanaman mengandung karbon, yang dihasilkan dari
fotosintesis. Proses ini vital untuk pertumbuhan dan berlangsung pada setiap
tanaman serta merupakan bagian utama dari siklus pertumbuhan (Lambers et al.
2008).
Pertumbuhan merupakan akumulasi bahan organik. Kandungan karbon,
komponen struktural utama tanaman berbeda untuk jenis tanaman yang berbeda.
Contohnya, kandungan karbon pada pinus sebesar 50% dari berat kering. Model
pertumbuhan tegakan pohon bergantung pada keseimbangan karbon, laju
fotosintesis dan respirasi. Pertumbuhan bersih merupakan selisih antara
pertumbuhan dan senesen. Proporsi respirasi dari produksi fotosintesis total pada
hutan Pinus sylvestris sekitar 40-50% dan mencapai 90% pada hutan Pinus
radiata (Raghavendra 1991).
Pohon dengan laju pertumbuhan cepat memiliki laju asimilasi CO2 relatif
lebih tinggi pada wilayah beriklim tropis (Hidayati et al. 2009). Diduga laju
asimilasi CO2 dapat dijadikan indikator untuk mengevaluasi karakteristik
pertumbuhan. Kajian mengenai fotosintesis pohon jauh lebih sedikit dipelajari
dibandingkan dengan fotosintesis tanaman budidaya karena beberapa alasan yaitu:
ukuran pohon dewasa mempersulit proses pengambilan sampling, pohon di hutan

16
alami memiliki tingkat keragaman yang tinggi, kesulitan untuk pengukuran laju
fotosintesis seluruh jenis tegakan pohon di hutan, minimnya model pertumbuhan
pohon ditinjau dari sudut pandang proses fisiologi (Raghavendra 1991).
Deskripsi Tanaman Restorasi TNGGP
Beleketebe (Sloanea sigun (Blume) K. Schumann)
Beleketebe atau ki somang adalah nama daerah S. sigun. Tumbuh di Jawa
khususnya Jawa Barat pada ketinggian 600 – 2100 m dpl. Daerah penyebaran
selain Indonesia adalah Birma, Kamboja, India, Malaysia, Myanmar, dan
Thailand. Jenis ini tumbuh tersebar atau kadang-kadang berkelompok di hutan
hujan primer yang lembab, terutama pada tanah subur. Pohon ini berbunga kecil,
kuning muda sampai putih. Bunga-bunga tersebut tumbuh berkelompok atau
tunggal pada ranting baru. Buahnya berukuran sedang, bulat, dan berbulu panjang
seperti rambutan, berwarna hijau muda (Lembaga Biologi Nasional Indonesia –
LIPI 1977).
Jenis ini termasuk keluarga Elaeocarpaceae. Pohonnya dapat mencapai 30 m
dengan diameter 120 cm. Diameter bebas cabang tidak berdaun mencapai 40 cm.
Batangnya lurus, tidak berbanir, dengan kulit yang licin dan berwarna abu-abu
tua. Tajuknya lebat sekali, bulat, dan daunnya gugur selama ± dua hari yang
diikuti oleh pertumbuhan daun muda, kemudian berbunga di ujung-ujung ranting.
Daunnya tunggal, tersusun dalam bentuk spiral dan kadang-kadang agak
berhadapan, berbentuk bulat telur sampai jorong, ujungnya melancip, tidak
berbulu, permukaan atasnya mengkilap bertangkai panjang (Lembaga Biologi
Nasional – LIPI 1977). Tangkai daun berukuran 2-2.5 cm, ramping, bengkak di
ujung. Helai daun berbentuk elips berukuran 12-14 cm × 5-6 cm, tipis, vena
lateral berjumlah 5 atau 6 per sisi, tepi daun bergigi. Bunga soliter, sepal
berjumlah 4, kelopak 4, berwarna kekuningan. Benang sari berukuran 5-7 mm.
Ovarium bulat telur (Backer & van Den Brink 1965). Termasuk pohon lapis
pertama atau kanopi (van Steenis 2006).

17
Jamuju (Dacrycarpus imbricatus (Blume) de Laub.)
D. imbricatus yang memiliki nama padanan Podocarpus imbricatus Blume,
termasuk ke dalam famili Podocarpaceae yang merupakan kelompok tanaman
berdaun jarum atau konifer. Jamuju, nama umum, juga dikenal dengan nama
lokal: ampinur bunga (Karo), medang cemara (Melayu), ambun (Minangkabau),
ki jamuju, ki mades, kiputri, kicemara (Sunda), aru, taji, tekit, camara ukung,
cemara rante, cemara waris (Jawa), cemara binek (Madura), kaju (Sumbawa), kaju
angin, camba-camba (Banten) (Heyne 1987), embun (Sumatera, Sulawesi),
cemba-cemba (Sulawesi), igem (Filipina), sha-mo-pin (Burma), podo churcher
atap (Malaysia), thong nang (Vietnam). Sebaran tumbuh meliputi selatan China,
Indochina, Burma, Thailand, Malaysia, hingga Vanuatu dan Fiji. Di Indonesia
jenis ini tersebar di Sumatera Utara, Sumatera Barat, Aceh, dan Jawa Barat.
Merupakan tanaman dataran tinggi, yaitu pada ketinggian 750 – 2500 dpl. Tinggi
pohon jamuju rata-rata 50 m, termasuk pohon mencuat (van Steenis 2006) dengan
diameter mencapai 100-200 cm. Termasuk tanaman moderat-lambat, walaupun
pertumbuhannya lebih cepat di daerah yang disinari matahari penuh. Jamuju
berdaun majemuk berbentuk lancip membentuk apiculus yang halus. Daun
dewasa jamuju menyebar, lurus, seperti skala, rebah pada 1 sampai 4 sisi,
berukuran 1.0-1.8 mm x 0.4-1.0 mm. Daun involucralnya menyebar dan
membuka pada reseptakel pada saat muda. Jamuju termasuk tanaman yang selalu
hijau, tidak menggugurkan daunnya pada musim kemarau, memiliki kanopi yang
lebat berbentuk piramid hingga oval (Backer & van Den Brink 1963).
Jamuju memiliki pepagan keras, permukaan kasar berlentisel disana-sini,
pada pohon tua mengelupas dalam bentuk lempengan tebal kecil-kecil memanjang
vertikal menggelendong. Hidup di hutan campuran basah atau di hutan cemara.
Penyebaran jenis D. imbricatus di Taman Nasional Gede Pangrango
mengelompok, karena jenis ini banyak ditemukan pada ketinggian 1400 sampai
2045 m dpl. D. imbricatus di Taman Nasional Gede Pangrango tumbuh pada
ketinggian tempat 1400 sampai 2045 m dpl dan kelerengan 3 sampai 40 % dengan
suhu 15 sampai 25 oC dengan kelembaban 73 sampai 100%. Jenis tanah yang
cocok untuk jenis D. imbricatus adalah jenis tanah regosol dan andosol dengan

18
kemasaman tanah yang termasuk kategori asam sampai sangat asam (Bramasto
2008).
Ki Sireum (Syzygium lineatum (Bl.) Merr. & Perry)
Pohon ini masuk ke dalam keluarga Myrtacea. S. lineatum memiliki nama
lokal kayu udang Sumatera), ki sireum (Sunda, Jawa), nagasari rangkang (Jawa),
phung kha, kelat puteh, daeng sok phlueak, khwat (Thailand), lagi-lagi
(Philipina), kelat lapis (Malaysia). Nama sinonim jenis ini di antaranya adalah S.
longiflorum.
Habitusnya pohon. Cabangnya berwarna coklat kehitaman ketika kering,
berbentuk silinder tetapi biasanya sedikit runcing di kedua ujungnya, membulat
pada bagian melintang, permukaannya halus; cabang tua putih keabu-abuan.
Tangkai daun berukuran 1-1.2 cm; daun elips berukuran 6-8 x 2.5-3.5 cm, kasar,
abaksial (bagian bawah daun) sedikit berwarna saat kering, adaksial (bagian atas
daun) berwarna coklat tua dan mengkilap saat kering. Abaksial memiliki kelenjar
banyak, adaksial berkelenjar kecil, tulang daun sekunder padat, terpisah dengan
jarak 1 mm, dengan sudut 75o dari tulang daun bagian tengah. Tulang daun
intramarginal kurang dari 1 mm dari batas pinggir, bagian dasar lebar dan tumpul.
Puncak daun melancip dengan titik puncak 1-1.5 cm. Berbunga banyak dengan
ukuran 8-10 cm, bercabang tiga, kuncup bunga berukuran 6-7 mm. Kelopak
bunga berbentuk lobus semiorbital pendek. Kelopak bunga terpisah berukuran 3
mm. Benang sari berukuran 5-7 mm (Backer & van Den Brink 1965).
Lame (Alstonia scholaris (L) R. Br.)
Lame merupakan anggota famili Apocynaceae, memiliki nama daerah kayu
gabus, kayu skola, pulai, lame, pule (Indonesia); chattin (Bengal); lettok (Burma);
birrba, black board tree, dita bark, milk wood, milkwood pine, milky pine, white
cheesewood (Inggris); dalipoen, dita (Filipina); chatian (India); pulai, pule, rite
(Indonesia); pule (Jawa); pulai, pulai linlin (Malaysia); chataun, chhatiwan
(Nepal); sattaban, teenpet, teenpethasaban (Thailand); chatiyan wood, pulai,
shaitan wood, white cheese wood (nama dagang); caay mof cua, caay suwxa
(Vietnam). Habitus A. scholaris adalah pohon besar, dengan tinggi sekitar 40 m.
Lame memiliki lateks susu yang mengalir cepat ketika dipotong. Batang berwarna

19
abu-abu gabus agak ke abu-abu-putih kulit. Daun berkarang di aksila atas dengan
jumlah 4-8 daun. Tangkai daun tangkai berukuran 1-1.5 cm, elips atau lanset,
gundul atau berbulu jarang, meruncing ke arah dasar, dengan ukuran daun 11.5-23
x 4-7.5 cm. Permukaan atas hijau tua, bagian bawah hijau-putih dengan 25-40
pasang urat lateral pada setiap sisi pelepah berjarak 2-6 mm terpisah. Ujung daun
bulat atau meruncing ke arah dasar. Bunga majemuk, malai terminal, hingga 120
cm; berukuran 7-10 mm berwarna putih, krem atau hijau; tabung berbulu; lobus
jarang atau padat puber, 1,5-4 mm, margin kiri tumpang tindih; sangat wangi.
Buah terjumbai, dengan dua lobus, folikelnya pecah, coklat atau hijau, kering atau
berkayu, berbentuk gelendong lonjong, berukuran 15-32 cm, 4-6 mm, biji
cokelat, berukuran 4-5 mm x 0.9 -1.2 mm, dengan seberkas rambut 7-13 mm pada
setiap akhir.
Lame ditemukan pada ketinggian 0-900 m, dengan suhu tahunan rata-rata
12-32 oC dan curah hujan tahunan rata-rata 1200-1400 mm. Jenis ini baik hidup
pada jenis tanah bermacam-macam, termasuk aluvial. Lateksnya bisa dibuat
permen karet berkualitas baik. Kayunya direkomendasikan sebagai spesies kayu
bakar untuk daerah Patana, Sri Lanka. Kulit kayu menghasilkan serat, dan kayu
dianggap sebagai cocok untuk produksi pulp dan kertas. Bunga A. scholaris
menghasilkan minyak esensial. Kedokteran aborigin Australia menggunakan kulit
kayu untuk pengobatan sakit perut dan demam, lateks untuk neuralgia dan sakit
gigi. Di India, kulit kayu digunakan untuk mengobati keluhan usus
(worldagroforestrycentre.org; Backer & van Den Brink 1965).
Manglid (Manglietia glauca Bl.)
Manglid masuk ke dalam keluarga Magnoliaceae. Nama botaninya adalah
Manglietia glauca, dengan nama sinonim Magnolia blumei
(worldagroforestrycentre.org; Backer & van Den Brink 1963). Jenis ini memiliki
nama daerah: manglid (Sunda); baros, cempaka bulus (Jawa); antuang, madang
limpaung, sitibai (Minangkabau); cempaka. Manglid berupa pohon, dengan
tinggi mencapai 25 – 40 m dengan bebas cabang 25 m dan diameter mencapai 150
cm, tersebar di ketinggian 1000 – 1500 m dpl. Hidupnya berkelompok dan di
tempat yang lembab. Tajuk membulat, lebat, percabangannya berbentuk garpu

20
yang dimulai jauh dari atas tanah (Heyne 1987). Daun tunggal bentuk elips
memanjang atau elips melebar, kebanyakan bulat telur memanjang, ukuran 13-18
cm, panjang kadang sampai 25 cm. Ujung dan pangkal daun runcing, tangkai
daun panjang. Tidak berbulu, abaksial daun berwarna abu-abu kebiruan, bagian
adaksial hijau muda agak mengkilap, tersusun spiral.
Bunga terminal, soliter, besar, tangkai panjang 2.5 - 4 cm, berwarna kuning
muda, harum, kelopak 9-13 tersusun dalam 3 lingkaran, benang sari banyak dan
tersusun spiral, tangkai benang sari panjang atau pendek. Ovary ada 4 atau lebih
pada masing-masing karpel. Penyerbukan dibantu oleh lebah madu dan berbunga
sepanjang tahun. Buah majemuk, berbentuk kerucut (kegelvormig) panjang 6-8
cm, pada permukaan berwarna hijau dengan titik-titik putih, kemudian menjadi
coklat hitam. Biji 2-6 banyaknya, kadang sampai 12, berwarna merah
(http://www.dephut.go.id).
Puspa (Schima wallichii (DC.) Korth.)
S. wallichii termasuk famili Theaceae, tinggi pohonnya dapat mencapai 40
m, termasuk pohon lapis kedua/kanopi (van Steenis 2006), dengan panjang batang
bebas cabang sampai 25 m dan diameter sampai 250 cm. Tidak berbanir, kulit luar
berwarna merah muda, merah tua sampai hitam, beralur dangkal dan mengelupas.
Kulit hidup tebalnya sampai 15 mm berwarna merah dan di dalamnya terdapat
miang gatal. Tajuknya bulat sampai lonjong, lebat, hijau tua, mengkilat, dengan
daun muda berwarna merah jambu. Daunnya tunggal, tebal, bagian abaksial hijau
kebiru-biruan, berbentuk jorong. Bunganya putih, tunggal, terdapat pada ketiak
daun dan berkelompok pada ujung ranting. Buahnya agak bulat, berwarna putih
perak seperti sutra saat muda, coklat dan pecah bila sudah tua (Sastrapradja et al.
1977).
Puspa tumbuh pada tanah kering dan tidak memiliki keadaan tekstur dan
kesuburan tanah, sehingga baik untuk reboisasi padang alang-alang, belukar dan
tanah kritis. Jenis ini memerlukan iklim basah sampai agak kering, hidup pada
dataran rendah sampai di dataran pegunungan dengan ketinggian 1000 m dpl
(Martawijaya et al. 1989). Puspa dapat tumbuh pada kisaran iklim, habitat dan
tanah yang luas. Kebutuhan akan cahaya tergolong sedang, sering berkelompok

21
dan terdapat pada dataran rendah sampai hutan dataran tinggi, akan tetapi
terutama terdapat pada hutan yang terganggu dan hutan sekunder. Adapun
persebaran alami Puspa yaitu Brunei, India, China, Indonesia, Malaysia,
Myanmar, Nepal, Papua, Philipina, Thailand dan Vietnam (Agroforestry Data
Base 2007).
Rasamala (Altingia excelsa Noronha)
Rasamala dikelompokkan ke dalam famili Hamamelidaceae, memiliki nama
daerah rasamala, mala, tulasan, atau mandung. Jenis ini menyebar mulai dari
Himalaya menuju wilayah lembab di Myanmar hingga Semenanjung Malaysia, ke
Sumatera dan Jawa. Di Jawa, jenis ini hanya tumbuh di wilayah barat dengan
ketinggian 500-1500 m dpl, di hutan bukit dan pegunungan lembab. Di Sumatera,
A. excelsa tersebar di Bukit Barisan. Tumbuh alami terutama pada tapak lembab
dengan curah hujan lebih 100 mm per bulan dan tanah vulkanik. Jenis ini
digunakan untuk penanaman terutama di Jawa Barat dan Jawa Tengah. Ditanam
pada jarak rapat, karena pohon muda cenderung bercabang jika mendapat banyak
sinar matahari.
Pohon selalu hijau, tinggi dapat mencapai 40-60 m dengan tinggi bebas
cabang 20-35 m, diameter hingga 80-150 cm. Kulit kayu halus, abu-abu, dan
kayunya merah. Pohon yang masih muda bertajuk rapat dan berbentuk piramid,
kemudian berangsur menjadi bulat setelah tua. Letak daun bergiliran, bentuknya
lonjong, panjangnya 6 - 12 cm, dan lebarnya 2.5-5.5 cm, dengan tepi daun
bergerigi halus. Bunga berkelamin satu. Bunga jantan dan betina terpisah pada
pohon yang sama. Malai betina terdiri dari 14-18 bunga, berkumpul menyerupai
kepala. A. excelsa tumbuh alami terutama di bukit campuran yang lembab dan
hutan pegunungan. Jenis ini sering ditemukan hidup berkelompok dengan spesies
Podocarpus, Quercus dan Castanopsis. Ditemukan pada ketinggian 500-1500 m,
dengan rata-rata curah hujan tahunan lebih 100 mm. Rasamala baik hidup di
tanah vulkanik atau kadang-kadang pada tanah di atasnya terdapat batuan sedimen
(http://www.worldagroforestry.org).

22
Saninten (Castanopsis argentea (Bl.) A.DC.)
Saninten masuk ke dalam famili Fagaceae. Ia memiliki nama daerah kandik
kurus, paning-paning, rasak, selasik (Sumatera), berangan, saninten, sarangan, dan
wrakas (Jawa). Daerah penyebaran meliputi Sumatera Barat, Jawa, Papua,
Myanmar, dan Malaysia (Heyne 1987). Habitus pohon bertajuk lebar dengan
ketinggian 15-35 m, diameter 0.5-1 m, dengan panjang batang bebas cabang
sampai 25 m, tidak berbanir (http://www.d-forin.com). Daun pohon saninten
berbentuk lancip memanjang (lanset) dengan ukuran panjang 7-12 cm, lebar 2-3.5
cm, bagian terlebar di tengah, permukaan daun licin berlilin, dan bagian
bawahnya berwarna abu-abu keperakan ditutupi bulu-bulu menyerupai bintang
atau sisik yang lebat. Jika dibanding dengan daun jenis lain, C. Argentea lebih
terlihat keperakan (Backer & van Den Brink 1965). Tumbuhan ini berdaun
tunggal dengan kedudukan berseling dan tersusun seperti spiral dan daun
penumpu mudah luruh. Ahli botani van Steenis (1972) menyatakan bahwa daun
penumpu (stipula) ditutupi bulu yang lebat, panjang daun berkisar antara 10-15
mm dan lebar 2-3 cm. Salah satu ciri khas organ vegetatifnya, yaitu bila daun
dilipat maka akan terlihat garis lilin berwarna putih memanjang pada bagian daun
di sebelah atas. Kulit batang pohon berwarna hitam, kasar, dan pecah-pecah
dengan permukaan batang tidak rata, terdapat alur-alur memanjang pada batang
yang tak lain adalah garis empulur yang menonjol keluar. Hal ini merupakan salah
satu ciri khas organ vegetatif famili Fagaceae. Kayu terasnya berwarna coklat
kelabu sampai merah muda, kayu gubal/bagian tengah berwarna putih, kuning
muda, dan kadang-kadang kemerah-merahan dengan ketebalan 5-6 cm (Prawira
1990).
Pohon saninten berbunga pada Agustus-Oktober dan berbuah pada
November-Februari. Bunga jantan tersusun dalam untaian berbentuk bulir
sepanjang 15-25 cm, bunga betina tumbuh menyendiri dengan panjang 5-15 cm,
diameter 2-4 mm, dan bunga berwarna kuning keputihan. Buahnya bertangkai
seperti buah rambutan, berkelompok di mana kulit buah ditutupi oleh duri yang
tumbuh berkelompok, ramping, tajam, dan berkayu. Buah berbentuk bulat telur
dengan duri mencuat pada empat sisi yang berisi tiga biji berbentuk tipis dan

23
cekung. Biji biasanya dimanfaatkan sebagai bahan makanan dengan cara direbus
atau dibakar (van Steenis 1972).
Keadaan Umum Lokasi Penelitian
Area restorasi terletak di Resort Bodogol Taman Nasional Gunung Gede
Pangrango (TNGGP) (06o46’.241” LS; 106
o50’.447”BT) pada ketinggian 620-
709 m dpl, berdekatan dengan Pusat Pendidikan Konservasi Alam (PPKA)
Bodogol (Gambar 4).
Gambar 4 Lokasi restorasi Resort Bodogol TNGGP (tanda panah menunjukkan
lokasi) (Balai TNGGP)
Bagian dari wilayah ini merupakan lahan pertanian aktif dan sisanya
merupakan lahan pertanian yang telah ditinggalkan. Lahan pertanian di wilayah
ini merupakan milik TNGGP. Menurut sejarahnya, lahan ini dulunya milik Perum
Perhutani Unit III, perusahaan publik yang bertugas mengelola produksi kayu,
dari tahun 1978-2003. Tiga spesies kayu utama yang ditanam monokultur di
wilayah ini adalah Altingia excelsa, Agathis damara, dan Pinus merkusii. Setelah
penebangan pada tahun 2003, wilayah yang terbuka dimanfaatkan oleh petani
24
lokal sampai sekarang, dan wilayah ini menjadi bagian dari TNGGP. Para petani
menanam tanaman tahunan seperti jagung, singkong, cabai, kacang panjang, dan
lain-lainnya. Sementara, lahan yang ditinggalkan didominasi oleh tanaman liar.
Karakteristik lahan dapat dilihat pada Tabel 2.
Tahun 2009 UPT Balai Konservasi Tumbuhan Kebun Raya Cibodas
menanam sekitar 2400 benih dari 8 jenis tanaman yang secara alami dapat
ditemukan di hutan TNGGP. Penanaman dilaksanakan di lahan seluas 4 ha secara
acak. Kedelapan spesies pohon tersebut adalah Altingia excelsa, Alstonia
scholaris, Castanopsis argentea, Dacrycarpus imbricatus, Manglietia glauca,
Syzygium lineatum, Schima wallichii, dan Sloanea sigun. Sampel tanaman
sebanyak 240 pohon (30 pohon untuk setiap jenis) diberi label dan dimonitor
pertumbuhannya (Rahman et al. 2011).
Tabel 2 Karakteristik lahan restorasi
Parameter Wilayah
Restorasi
Hutan Alami
TNGGP
pH tanah (0-10 cm) 5.27 5.10
(10-20 cm) 4.75 4.76
Bahan organik (%) (0-10 cm) 6.73 9.86
(10-20 cm) 6.48 10.2
Bulk density (g/cm3) (0-10 cm) 0.66 0.56
(10-20 cm) 0.67 0.72
Pasir (%) 9.11
Debu (%) 49.69
Liat (%) 41.20
Kemiringan lahan (o) 48-60
Suhu udara (oC)
Intensitas cahaya rata-rata (lux)
Jam 09.00-11.00
23.3-33.0
32 000