ecoli
DESCRIPTION
BakteriTRANSCRIPT

5
II. TINJAUAN PUSTAKA
2.1. Karakteristik Morfologi Escherichia coli
Bakteri E. coli merupakan spesies dengan habitat alami dalam saluran
pencernaan manusia maupun hewan. E. coli pertama kali diisolasi oleh Theodor
Escherich dari tinja seorang anak kecil pada tahun 1885. Bakteri ini berbentuk
batang, berukuran 0,4-0,7 x 1,0-3,0 µm, termasuk gram negatif, dapat hidup
soliter maupun berkelompok, umumnya motil, tidak membentuk spora, serta
fakultatif anaerob (Gambar 1) (Carter & Wise 2004).
Gambar 1. Morfologi E. coli
(Sumber: Kunkel 2009)
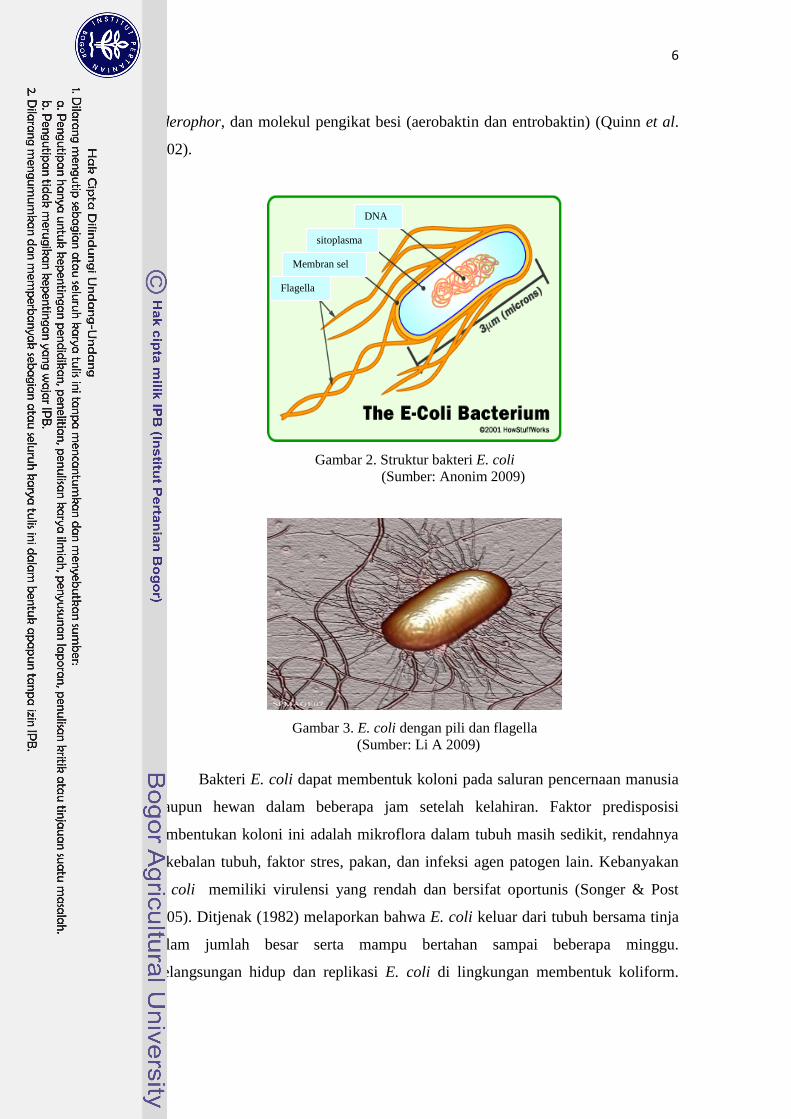
Struktur sel E. coli dikelilingi oleh membran sel, terdiri dari sitoplasma
yang mengandung nukleoprotein (Gambar 2). Membran sel E. coli ditutupi oleh
dinding sel berlapis kapsul. Flagela dan pili E. coli menjulur dari permukaan sel
(Gambar 3) (Tizard 2004). Tiga struktur antigen utama permukaan yang
digunakan untuk membedakan serotipe golongan E. coli adalah dinding sel,
kapsul dan flagela. Dinding sel E. coli berupa lipopolisakarida yang bersifat
pirogen dan menghasilkan endotoksin serta diklasifikasikan sebagai antigen O.
Kapsul E. coli berupa polisakarida yang dapat melindungi membran luar dari
fagositik dan sistem komplemen, diklasifikasikan sebagai antigen K. Flagela
E. coli terdiri dari protein yang bersifat antigenik dan dikenal sebagai antigen H.
Faktor virulensi E. coli juga disebabkan oleh enterotoksin, hemolisin, kolisin,
6
siderophor, dan molekul pengikat besi (aerobaktin dan entrobaktin) (Quinn et al.
2002).
Gambar 2. Struktur bakteri E. coli
(Sumber: Anonim 2009)
Gambar 3. E. coli dengan pili dan flagella
(Sumber: Li A 2009)
Bakteri E. coli dapat membentuk koloni pada saluran pencernaan manusia
maupun hewan dalam beberapa jam setelah kelahiran. Faktor predisposisi
pembentukan koloni ini adalah mikroflora dalam tubuh masih sedikit, rendahnya
kekebalan tubuh, faktor stres, pakan, dan infeksi agen patogen lain. Kebanyakan
E. coli memiliki virulensi yang rendah dan bersifat oportunis (Songer & Post
2005). Ditjenak (1982) melaporkan bahwa E. coli keluar dari tubuh bersama tinja
dalam jumlah besar serta mampu bertahan sampai beberapa minggu.
Kelangsungan hidup dan replikasi E. coli di lingkungan membentuk koliform.
sitoplasma
Membran sel
Flagella
DNA

7
E. coli tidak tahan terhadap keadaan kering atau desinfektan biasa. Bakteri ini
akan mati pada suhu 600
C selama 30 menit.
2.1.1. Klasifikasi
Klasifikasi E. coli menurut Songer dan Post (2005) adalah sebagai berikut:
Kingdom : Bacteria
Filum : Proteobacteria
Kelas : Gamma Proteobacteria
Ordo : Enterobacteriales
Famili : Enterobacteriaceae
Genus : Escherichia
Spesies : Escherichia coli
Berdasarkan perbedaan serotipe dan virulensi, strain E. coli patogen yang
menyebabkan penyakit pada saluran pencernaan dibedakan menjadi enam
golongan, yaitu enterotoksigenik (ETEC), enteroinvasif (EIEC), enteropatogenik
(EPEC), enterohemorhagik (EHEC), enteroagregatif (EAEC), dan
nekrotoksigenik (NTEC) (Sommer et al. 1994).
Golongan ETEC merupakan penyebab diare enterotoksigenik pada
mamalia, seperti anak sapi, anak babi, dan anak domba. Gejala klinis yang terjadi
antara lain diare, dehidrasi, asidosis, bahkan kematian (Hanif et al. 2003). Faktor
virulensi yang digunakan untuk identifikasi ETEC adalah enterotoksin dan
antigen pili (fimbriae). Enterotoksin ETEC berupa toksin labil panas (heat-labile
toxins/ LT) dan toksin stabil panas (heat-stabile toxins/ ST). ETEC dapat
menghasilkan satu atau dua enterotoksin tergantung pada plasmid (massa DNA
ekstra kromosom). Makhluk hidup yang terinfeksi bakteri mengandung kedua
plasmid biasanya mengalami diare yang lebih berat dan lebih lama. Enterotoksin
akan diabsorbsi oleh sel epitel yeyunum dan ileum serta dapat merusak motilitas
usus sehingga memfasilitasi keberadaan ETEC di dalam lumen usus (Salyers &
Whitt 1994).
ETEC yang mempunyai antigen perlekatan K99 merupakan penyebab
utama diare neonatal dan kematian anak sapi (Supar et al. 1998). ETEC K99

8
dapat terdeteksi pada hari kedua sampai hari kelima dari ulas rektal anak sapi
yang menderita diare dan tidak ditemukan lagi pada anak sapi yang diare setelah
lebih dari lima hari (Supar 1986). Adapun faktor yang mempengaruhi infeksi
ETEC pada inang, yaitu umur, pH lambung, dan kehadiran antibodi spesifik
terhadap permukaan antigen ETEC (Supar 2001).
2.1.2. Patogenesa Infeksi ETEC
Mekanisme infeksi ETEC di dalam tubuh, yaitu ETEC menempel pada sel
enterosit melalui pili (fimbriae). ETEC kemudian berproliferasi dan berkolonisasi
pada mukosa usus sehingga terjadi peningkatan jumlah ETEC di dalam saluran
pencernaan dan muncul lesio. Diare terjadi karena dinding usus mengalami
kerusakan dan menghalangi reabsorbsi cairan (Biowey & Weaver 2003).
ETEC memproduksi enterotoksin heat labile toxin (LT) atau heat stable
toxin (ST) (Sommer et al. 1994). Menurut Ganong (2002), toksin akan berikatan
dengan reseptor dan masuk ke dalam sel. Toksin stabil bekerja mengaktivasi
guanilat siklase sehingga menyebabkan akumulasi cairan dan elektrolit di dalam
lumen usus serta memblokade absorbsi. Toksin labil akan mengikat ribose
adenosin difosfat (ADP) sehingga menghambat kegiatan GTPase (pemecah
protein G). Akibatnya, protein G ini meningkat dan merangsang adenilil siklase
sel epitel yang berkepanjangan sehingga menyebabkan peningkatan jumlah adenil
monofosfat (AMP). Peningkatan AMP akan menyebabkan peningkatan sekresi
sel-sel kelenjar di dalam usus, yaitu merangsang seksresi Cl- (hipersekresi) dengan
membuka saluran klorida pada sel kripta dan menghambat absorbsi Na+
dari
lumen ke dalam sel epitel usus. Peningkatan kadar elektrolit dan air di dalam
lumen usus menyebabkan diare.
Diare merupakan gejala gangguan pencernaan yang ditandai dengan
pengeluaran feses dalam jumlah melebihi normal, konsistensi cair, dan frekuensi
pengeluaran yang melebihi normal. Feses dikeluarkan oleh penderita tanpa
kesulitan karena terjadi peningkatan peristaltik usus (Ganong 2002). Frekuensi
diare pada anak sapi berhubungan dengan keadaan imunodefisiensi neonatus.
Imunodefisiensi pada anak sapi disebabkan oleh kegagalan transfer kekebalan
pasif pada neonatus akibat tidak diberi kolostrum atau diberi susu berkualitas

9
rendah, belum optimalnya kemampuan absorbsi dari epitel usus, populasi terlalu
padat, sanitasi buruk, stres akibat perubahan pakan, higiene pakan, panas, dan
perubahan lingkungan (Khan & Khan 1996), serta kurangnya respon imun dan
mikroflora intestinal. Anak sapi yang diare terus-menerus akan memperlihatkan
gejala klinis berupa lemas, lesu, tidak mau menyusu, daerah di sekitar perineal
kotor oleh feses, mukosa mulut kering, pucat, kebiruan, turgor kulit jelek, dan
dapat menimbulkan kematian (Setiawan et al. 1983).
Cairan yang diseksresikan oleh kelenjar mukosa usus mengandung banyak
NaHCO3 sehingga ion Na+ dan HCO3
- akan ditarik dari darah, akibatnya derajat
asam (pH) darah menurun dan terjadi asidosis. Asidosis yang ditimbulkan oleh
keadaan ini akan menyebabkan kolapsnya sistem peredaran darah yang segera
diikuti shock dan kematian (Subronto 1985).
2.2. Sapi Friesian Holstein
Sapi merupakan hewan ternak sebagai sumber daging, susu, tenaga kerja,
dan kebutuhan lainnya (misalnya upacara agama). Sapi menghasilkan sekitar 50%
kebutuhan daging, 95% kebutuhan susu, dan 85% kebutuhan kulit di dunia (Ismail
2008).
Sapi Friesian Holstein (FH) berasal dari Belanda Utara dan Friesland
Barat. Bangsa sapi ini dikembangkan dari sapi liar Bos taurus. Sapi FH murni
mempunyai ciri-ciri berwarna belang hitam putih, ada juga berwarna merah putih
dengan batas warna yang jelas. Sapi FH merupakan sapi perah dengan produksi
susu tertinggi dibandingkan dengan bangsa-bangsa sapi perah lainnya dengan
kadar lemak susu rendah (3-7%). Dewasa kelamin sapi ini lambat, umur pertama
kali dikawinkan berkisar antara 15-18 bulan. Anak sapi FH neonatus memiliki
berat badan berkisar antara 35-45 kg (Sudono 1999). Berat badan sapi FH betina
dan jantan dewasa masing-masing sekitar 625 kg dan 900 kg. Umumnya terdapat
warna putih berbentuk segitiga pada dahi, kaki bagian bawah dan rambut di
bagian ekor berwarna putih, bertanduk pendek serta mengarah ke depan (Gambar
4). Sapi FH bersifat jinak dan tenang sehingga mudah untuk ditangani (Anonim
2008).

10
Klasifikasi sapi perah menurut Tyler dan Ensminger (2006) adalah sebagai
berikut:
Kingdom : Animalia
Filum : Chordata
Subfilum : Vertebrata
Kelas : Mamalia
Ordo : Artiodactyla
Subordo : Ruminansia
Famili : Bovidae
Genus : Bos
Spesies : Bos taurus
Sapi FH mempunyai adaptasi lingkungan yang baik pada dataran tinggi
(sekurang-kurangnya 700 m di atas permukaan laut), pada temperatur berkisar
antara 16-24 0C dan curah hujan sekitar 2000 mm/ tahun. Produksi susu sapi FH
tidak berselisih jauh dibandingkan dengan negara asalnya bila suhu lingkungan
sejuk yaitu pada suhu 18,3 0C dengan kelembaban udara 55%. Produksi susu rata-
rata sapi FH dapat mencapai 6360 kg/ tahun (Sutardi 1983). Sapi perah akan
mengalami cekaman panas yang berakibat pada menurunnya produktivitas jika
berada di lokasi yang memiliki suhu tinggi dan kelembaban udara yang tidak
mendukung (Anonima 2009).
Gambar 4. Sapi perah jenis Friesian Holstein (Sumber : Anonim 2008)

11
2.2.1. Sistem Pencernaan Sapi Neonatus
Saat dilahirkan, ruminansia pada umumnya memiliki lambung depan yang
kecil dan belum berfungsi. Lambung depan yang terdiri dari rumen, retikulum,
dan omasum, hanya menempati 30% dari keseluruhan lambung (Triakoso 2008).
Perkembangan lambung pada ruminansia muda dibagi ke dalam empat tahap,
yaitu tahap baru lahir (0-24 jam), tahap pre-ruminan (1 hari-3 minggu), tahap
transisi (3-8 minggu), serta tahap sebelum dan sesudah penyapihan (8 minggu-
dewasa) (Leek 1993). Tahap pre-ruminan, pakan cair akan masuk melalui
oesophageal groove, yang dapat menutup. Penutupan oesophageal groove
merupakan refleks yang diaktifkan oleh adanya gerakan menyusu dari anak
neonatus. Makanan dari esofagus langsung masuk ke dalam abomasum tanpa
melalui lambung depan (Imran 2010).
Abomasum secara fisik dan biokimiawi mampu mencerna bahan pakan
utama anak sapi yaitu susu. Abomasum mensekresi renin pada masa pre-ruminan.
Renin adalah enzim proteolitik dan berfungsi memecah susu menjadi kasein dan
whey. Whey masuk ke dalam duodenum dalam waktu lima menit setelah minum
susu, sedangkan kasein akan tetap berada di dalam abomasum. Kasein didegradasi
secara bertahap oleh renin atau pepsin serta asam klorida. Pencernaan protein ini
akan berlangsung selama 24 jam. Pencernaan dilanjutkan di dalam usus oleh
enzim-enzim seperti tripsin, kimotripsin dan karbopeptidase yang disekresikan
oleh pankreas serta peptidase lain yang disekresi oleh usus. Asam amino yang
terbentuk diabsorbsi di dalam usus halus, terutama pada yeyunum (Ruckebusch et
al. 1983).
Perubahan renin menjadi pepsin di dalam abomasum dipengaruhi faktor
umur. Semakin dewasa anak sapi, maka pepsin yang terbentuk semakin banyak.
Aktivitas pepsin masih rendah pada anak sapi yang berumur kurang dari tiga
minggu. Peningkatan jumlah pepsin terjadi jika anak sapi mulai mengkonsumsi
pakan selain susu dan pepsin bekerja optimal pada cairan abomasum dengan pH 2
(Triakoso 2008). Pakan yang dikonsumsi juga akan menggertak perkembangan
populasi mikroba dan fungsi ruminoretikulum. Sistem pencernaan anak sapi sudah
berfungsi penuh pada umur lebih dari delapan minggu (Sudono 1999).

12
2.3. Kolostrum dan Susu Sapi
Kolostrum merupakan sekresi yang dihasilkan kelenjar ambing mamalia
pada tahap akhir kebuntingan sampai beberapa hari setelah melahirkan (Tizard
2004). Kolostrum mulai diproduksi sekitar 3-6 minggu sebelum melahirkan
(Lazzaro 2000), berwarna kuning, konsistensi kental, dengan komposisi zat nutrisi
tinggi (Sutardi 1983). Menurut Parakkasi (1998), komposisi nutrisi dan sifat fisik
kolostrum dipengaruhi bangsa, paritas (jumlah kelahiran), ransum prepartum, dan
lamanya masa kering kandang.
Transfer imunoglobulin dari sirkulasi darah induk ruminansia menuju
kelenjar ambing dimulai pada beberapa minggu menjelang induk melahirkan dan
berhenti segera menjelang induk melahirkan (Esfandiari 2005). Antibodi di dalam
kolostrum melintasi epitel kelenjar ambing secara transitosis dan memasuki
sirkulasi anak melalui usus halus dan memberi imunitas pasif terhadap infeksi
(Arthington et al. 2000).
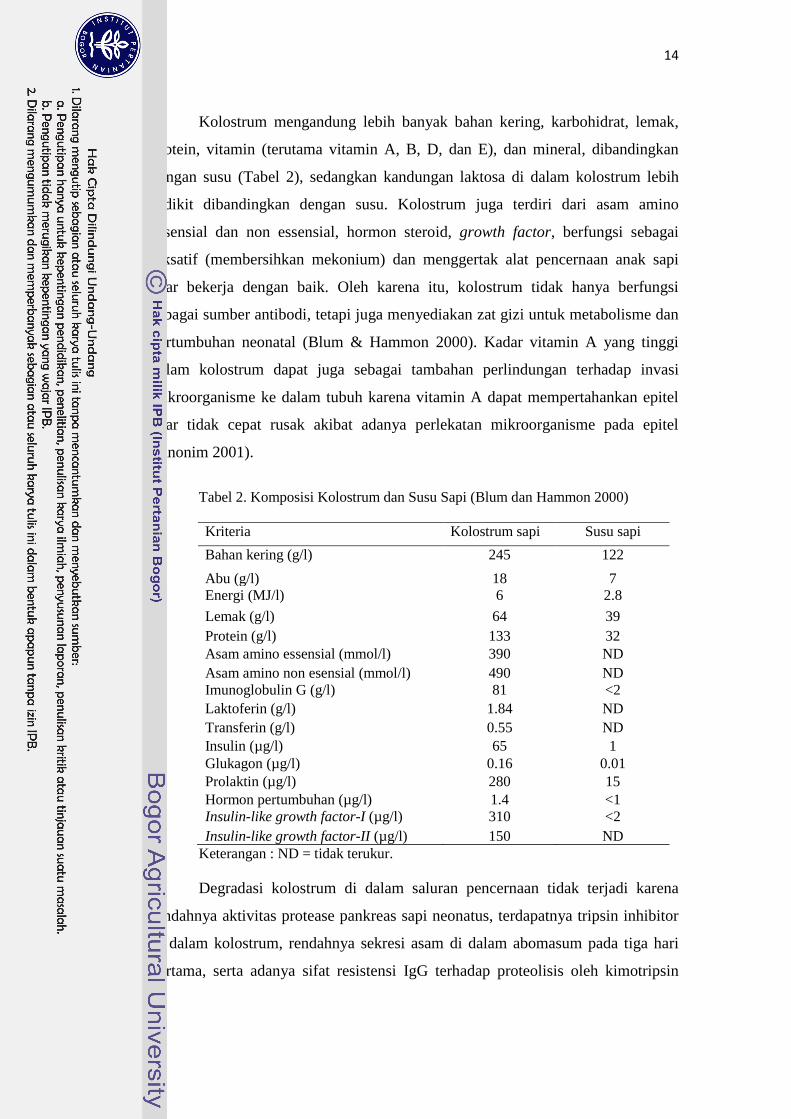
Imunoglobulin utama yang terkandung di dalam kolostrum sapi meliputi
IgG (90%), IgM (7%), dan IgA (5%) (Stott et al. 1979). Fraksi globulin kolostrum
disintesa oleh sel-sel plasma di dalam kelenjar ambing dari asam-asam amino
bebas di dalam darah (Toelihere 1979). Imunoglobulin utama di dalam kolostrum
hewan domestik pada umumnya adalah IgG, yaitu 65-90% dari total antibodi,
sedangkan kandungan IgA dan imunoglobulin lainnya hanya sedikit. Namun
setelah kolostrum berubah menjadi susu terjadi perubahan konsentrasi
imunoglobulin, tergantung spesies. IgG merupakan imunoglobulin paling
dominan pada ruminansia baik di dalam kolostrum maupun susu, sedangkan IgA
merupakan imunoglobulin yang paling dominan di dalam susu non-ruminansia
(Tabel 1) (Tizard 2004).
13
Tabel 1. Kandungan imunoglobulin dalam kolostrum dan susu pada hewan
domestik (Tizard 2004)
IgG merupakan antibodi utama yang berperan di dalam pengaturan respon
kekebalan sekunder, fiksasi komplemen, bertindak sebagai opsonin oleh
makrofag, dan imunoglobulin utama yang berperan dalam transfer kekebalan pasif
untuk anak neonatus. IgA berperan dalam melindungi selaput lendir, menetralisir
toksin atau virus dan mencegah perlekatan toksin atau virus pada permukaan sel
sasaran. IgA juga dapat meningkatkan efek bakteriolitik dengan cara
mengaktifkan komplemen. IgM berperan dalam perlindungan primer melawan
septikemia, fiksasi komplemen, dan proses agglutinasi (Roitt et al. 1998).
Pernyataan ini didukung oleh Subronto (1985) bahwa IgG dan IgM kolostrum
diperlukan untuk melindungi hewan neonatus dari penyakit sistemik, sedangkan
IgA mempunyai fungsi lokal dalam saluran percernaan.
Komponen kekebalan di dalam kolostrum mempunyai dua fungsi.
Pertama, antibodi kolostrum diabsorpsi ke dalam sirkulasi untuk mencegah invasi
mikroorganisme. Kedua, antibodi kolostrum tidak diabsorpsi karena usus halus
tidak lagi permeabel karena sel-sel di dalam usus sudah mature, akibatnya
antibodi tetap berada di dalam lumen usus dan berperan sebagai imunitas pasif
lokal (Scott et al. 2004). Frandson (1992) melaporkan bahwa kolostrum harus
segera diberikan setelah anak lahir karena permeabilitas usus paling tinggi segera
setelah lahir dan menurun dengan cepat terutama setelah 24 jam. Lebih dari waktu
tersebut usus tidak lagi permeabel menyerap protein antibodi kolostrum.
Imunoglobulin (mg/dL)
Spesies Fluida IgA IgM IgG
Kuda Kolostrum 500-1500 100-350 1500-5000
Susu 50-100 05-10 20-50
Sapi Kolostrum 100-700 300-1300 3400-8000
Susu 10-50 10-20 50-750
Domba Kolostrum 100-700 400-1200 4000-6000
Susu 5-12 0-7 60-100
Babi Kolostrum 950-1050 250-320 3000-7000
Anjing Kolostrum 500-2200 14-57 120-300
Susu 110-620 10-54 1-3
14
Kolostrum mengandung lebih banyak bahan kering, karbohidrat, lemak,
protein, vitamin (terutama vitamin A, B, D, dan E), dan mineral, dibandingkan
dengan susu (Tabel 2), sedangkan kandungan laktosa di dalam kolostrum lebih
sedikit dibandingkan dengan susu. Kolostrum juga terdiri dari asam amino
essensial dan non essensial, hormon steroid, growth factor, berfungsi sebagai
laksatif (membersihkan mekonium) dan menggertak alat pencernaan anak sapi
agar bekerja dengan baik. Oleh karena itu, kolostrum tidak hanya berfungsi
sebagai sumber antibodi, tetapi juga menyediakan zat gizi untuk metabolisme dan
pertumbuhan neonatal (Blum & Hammon 2000). Kadar vitamin A yang tinggi
dalam kolostrum dapat juga sebagai tambahan perlindungan terhadap invasi
mikroorganisme ke dalam tubuh karena vitamin A dapat mempertahankan epitel
agar tidak cepat rusak akibat adanya perlekatan mikroorganisme pada epitel
(Anonim 2001).
Tabel 2. Komposisi Kolostrum dan Susu Sapi (Blum dan Hammon 2000)
Kriteria Kolostrum sapi Susu sapi
Bahan kering (g/l) 245 122
Abu (g/l) 18 7
Energi (MJ/l) 6 2.8
Lemak (g/l) 64 39
Protein (g/l) 133 32
Asam amino essensial (mmol/l) 390 ND
Asam amino non esensial (mmol/l) 490 ND
Imunoglobulin G (g/l) 81 <2
Laktoferin (g/l) 1.84 ND
Transferin (g/l) 0.55 ND
Insulin (µg/l) 65 1
Glukagon (µg/l) 0.16 0.01
Prolaktin (µg/l) 280 15
Hormon pertumbuhan (µg/l) 1.4 <1
Insulin-like growth factor-I (µg/l) 310 <2
Insulin-like growth factor-II (µg/l) 150 ND
Keterangan : ND = tidak terukur.
Degradasi kolostrum di dalam saluran pencernaan tidak terjadi karena
rendahnya aktivitas protease pankreas sapi neonatus, terdapatnya tripsin inhibitor
di dalam kolostrum, rendahnya sekresi asam di dalam abomasum pada tiga hari
pertama, serta adanya sifat resistensi IgG terhadap proteolisis oleh kimotripsin

15
(Stott et al. 1979). Tizard (2004) melaporkan bahwa pada hewan neonatus,
kegiatan proteolitik di dalam saluran pencernaan rendah, karena itu protein
kolostrum tidak dipecah dan tidak digunakan sebagai sumber makanan, melainkan
utuh sampai di usus halus terutama yeyunum. Protein diserap di usus halus oleh
sel epitel usus melalui proses pinositosis. Melalui sel ini protein masuk ke dalam
saluran limfe dan kapiler usus, kemudian mencapai sirkulasi darah. Usus pada
ruminansia bersifat permeabel tapi tidak selektif. Semua imunoglobulin diabsorpsi
walaupun IgA berangsur-angsur diseksresi kembali ke dalam lumen usus. Setelah
absorbsi berhenti, antibodi yang diperoleh secara pasif ini akan segera menurun
konsentrasinya melalui proses katabolisme normal. Tingkat penurunan antibodi
tergantung pada kelas imunoglobulin dan konsentrasi awal.
Kolostrum penting untuk anak sapi karena adanya imunitas pasif dari
induk ke anak melalui pemindahan imunoglobulin asal kolostrum yang diserap
di dalam usus halus anak sapi. Kegagalan transfer imunoglobulin menyebabkan
anak sapi akan memiliki kadar imunoglobulin yang rendah atau tidak cukup di
dalam serumnya sehingga beresiko besar terhadap koliseptikemi, pneumoni, dan
infeksi lainnya (Biowey & Weaver 2003). Supar et al. (1998) melaporkan bahwa
anak sapi yang lahir dari induk sapi yang divaksin tetapi tidak mendapat
kolostrum, dan kemudian ditantang dengan E. coli K99, meskipun tidak mati akan
menderita diare dan bobot badan relatif tidak bertambah sampai umur 42 hari.
2.3.1. Kekebalan Humoral Neonatus
Tipe plasenta ruminansia adalah sindesmokhorial. Tipe plasenta tersebut
memiliki struktur jaringan pemisah antara sirkulasi darah induk dan fetus, yang
terdiri dari beberapa lapis yang tersusun atas endotel kapiler induk, jaringan
uterus, epitel uterus, epitel khorion, jaringan ikat fetus, dan endotel kapiler fetus.
Transfer imunoglobulin (Ig) transplasenta pada tipe plasenta ini benar-benar
terhambat akibatnya tidak memungkinkan terjadinya transfer Ig di dalam uterus
(Tizard 2004).
Anak sapi lahir dalam keadaan hipogammaglobulinemia, oleh karena itu
sapi neonatus sangat tergantung pada antibodi yang diterima melalui kolostrum
sampai sistem imun anak sapi aktif (Khan & Khan 1996). Sapi neonatus yang

16
belum menyusu mengandung sedikit sekali konsentrasi antibodi di dalam
serumnya, sedangkan sapi neonatus yang sudah menyusu mengandung
konsentrasi antibodi yang tinggi dalam serumnya (Supar et al. 1998).
Rendahnya konsentrasi imunoglobulin berhubungan dengan tingginya
tingkat morbiditas dan mortalitas. Sebanyak 53,6% resiko terjadinya mortalitas
disebabkan oleh konsentrasi IgG di dalam serum <10 g/l akibat kurangnya
transfer IgG dari kolostrum. Konsentrasi IgG normal di dalam serum agar anak
sapi sehat adalah sekitar 17,5-18 g/l (Stott et al. 1979). Menurut Loucks et al.
(1985), frekuensi kejadian hipo dan agammaglobulinemia diakibatkan oleh
kegagalan ingesti kolostrum, rendahnya konsentrasi IgG di dalam kolostrum,
keterlambatan pemberian kolostrum, dan kegagalan absorbsi kolostrum.
Kekebalan pasif yang rendah pada anak sapi akan menurunkan laju pertumbuhan
bobot badan, menurunkan produksi susu pada laktasi pertama, serta meningkatkan
angka kesakitan pada enam bulan pertama setelah lahir (Sudono 1999).
Antibodi adalah molekul glikoprotein yang bersirkulasi dalam darah,
berperan di dalam pencegahan dan pengobatan suatu penyakit karena dapat
bereaksi dengan antigen yang merangsang pembentukannya. Antibodi memiliki
kemampuan berikatan secara khusus dengan antigen serta mempercepat
penghancuran dan penyingkiran antigen. Molekul ini disintesa oleh sel plasma (sel
B) sebagai respon kekebalan terhadap suatu antigen dan bersifat spesifik terhadap
antigen tersebut (Tizard 2004).
Antibodi di dalam serum memiliki konsentrasi lebih tinggi dibandingkan
dengan cairan tubuh lainnya (susu, sekresi nasal, saliva, air mata, urin, sekresi
vagina, dan cairan broncho-alveolar), namun lebih rendah dibandingkan dengan
konsentrasi antibodi di dalam kolostrum. Konsentrasi tertinggi imunoglobulin
dalam serum umumnya dicapai antara 12-24 jam setelah lahir. Imunoglobulin
dalam serum merupakan kompleks antibodi yang ditujukan terhadap bermacam-
macam determinan antigen (Norman et al. 1981).
Berdasarkan ukuran molekul, waktu paruh di dalam plasma, kandungan
karbohidrat, dan aktivitas biologi, antibodi dikelompokkan menjadi IgG, IgM,
IgA, IgE, dan IgD. Antibodi yang paling berlimpah di dalam sirkulasi darah

17
adalah imunoglobulin gamma (IgG). Antibodi umumnya hanya berikatan khusus
dengan antigen yang merangsang pembentukannya (Kuby 2004).
Antibodi yang secara umum meningkat setelah paparan antigen adalah
IgM dan IgG. IgM adalah kelas imunoglobulin yang terdapat dalam konsentrasi
tertinggi kedua setelah IgG di dalam serum kebanyakan hewan. IgM lebih efisien
dibandingkan dengan IgG pada aktivasi komplemen, opsonisasi, netralisasi virus,
dan aglutinasi walaupun diproduksi dalam jumlah relatif kecil. IgM akan
terbentuk sebagai respon paling awal dan selanjutnya konsentrasi akan menurun
dengan cepat. Sementara itu IgG akan terus-menerus meningkat hingga level
maksimum dalam periode yang relatif lama. IgG adalah imunoglobulin yang
terdapat dalam konsentrasi tertinggi dalam serum darah karena lebih mudah
berdifusi ke dalam cairan ekstravaskular dibandingkan dengan imunoglobulin lain
sehingga berperan utama dalam mekanisme pertahanan yang diperantai antibodi
(Roitt 1991).
Respon kekebalan humoral utama terhadap infeksi E. coli bekerja secara
langsung melawan bakteri atau produknya, seperti faktor kolonisasi dan toksin.
Antibodi terhadap antigen kapsul (K) dapat membantu menetralkan sifat
antifagosit dari kapsul (mengopsonisasi organisme) dan menyebabkan
penghancuran bakteri oleh sel fagositik. Antibodi juga akan berikatan dengan
reseptor sel usus sehingga ETEC tidak dapat melekat pada dinding usus dan tidak
patogen (Carter & Wise 2004). Salyers dan Whitt (1994) melaporkan bahwa
antibodi dalam usus halus akan melapisi permukaan usus halus sehingga
menghambat terjadinya perlekatan antara reseptor pada vili enterosit dengan
ETEC, akibatnya bakteri tidak dapat melekat pada dinding usus sehingga
kolonisasi E. coli pada vili dan produksi enterotoksin dapat dihindari.
2.4. Enzyme-Linked Immunosorbent Assay (ELISA)
Enzyme-Linked Immunosorbent Assay (ELISA) merupakan salah satu jenis
uji pengikatan primer. Uji ini dapat digunakan untuk mengidentifikasi dan
mengukur antibodi atau antigen. Prinsip dasar ELISA adalah mengukur langsung
interaksi antara antigen dengan antibodi. Adanya antibodi menunjukkan adanya
paparan antigen ke dalam tubuh inang yang diperiksa (Tizard 2004). Menurut

18
Burgess (1995), teknik ELISA dapat bekerja dengan konsentrasi bahan yang
cukup kecil dengan tingkat sensitifitas yang tinggi. Terdapat dua macam teknik
ELISA yang merupakan metode dasar ELISA, yaitu ELISA langsung dan ELISA
tidak langsung. ELISA langsung maupun tidak langsung digunakan untuk
mendeteksi antigen dan antibodi dengan syarat salah satunya diketahui. Indikator
enzim untuk reaksi imunologi merupakan ciri utama teknik ELISA.
ELISA tidak langsung merupakan konfigurasi paling sederhana yang dapat
digunakan untuk mengukur titer antibodi. ELISA tidak langsung digunakan
sebagai uji serologik karena cepat, sederhana dan relatif murah (Parede & Ginting
1996). Ikatan antigen dan antibodi pada ELISA tidak langsung ada dua macam,
yaitu ikatan antigen-antibodi primer dan ikatan antibodi primer-antibodi sekunder.
Ikatan antigen-antibodi primer bersifat spesifik, terjadi antara epitop antigen
dengan paratop pada rantai Fab antibodi. Antibodi primer tidak berlabel dapat
diperoleh dari serum atau cairan tubuh lain. Ikatan antibodi primer-antibodi
sekunder bersifat tidak spesifik, artinya ikatan antara antibodi dan anti-antibodi
dapat terjadi pada semua macam antibodi. Antibodi sekunder (sering disebut juga
dengan konjugat) terikat pada enzim (berlabel enzim). Enzim ini dapat
menguraikan substrat yang ditambahkan sehingga terjadi perubahan warna
larutan. Kekuatan warna ini tergantung dari banyaknya substrat yang terurai.
Banyaknya substrat yang terurai tergantung dari banyaknya enzim dalam larutan.
Kekuatan ini menunjukkan jumlah ikatan antigen-antibodi primer (Burgess 1995).