referat dina final edit oke
TRANSCRIPT

1
I. PENDAHULUAN
Kehamilan manusia berlangsung selama durasi rata-rata 280 hari (dihitung dari
periode menstruasi terakhir), dengan durasi normal antara 259 dan 294 hari.
Kelahiran sebelum usia kehamilan ini dianggap kelahiran prematur dan jika terus
berlanjut melewati periode ini disebut dengan post-term. Dalam praktik modern,
kelahiran post-term langka terjadi karena persalinan seringkali diinduksi pada atau
sebelum 14 hari terakhir taksiran persalinan. Sebaliknya, kelahiran prematur
adalah penyebab terbesar morbiditas dan mortalitas neonatal, yang angkanya
semakin meningkat di semua negara maju dengan prevalensi saat ini 12% di
Amerika Serikat dan 8% di Inggris. Namun, meskipun telah banyak kemajuan
dalam pengetahuan medis, faktor – faktor yang bertanggung jawab atas inisiasi
persalinan manusia baik dalam persalinan aterm dan prematur tidak dipahami
dengan baik, sehingga membatasi pilihan terapi.1
Para ilmuwan awalnya telah mempelajari regulasi hormonal dalam kelahiran
domba. Pada domba kortisol janin memicu partus dengan menginduksi enzim
plasenta yang memungkinkan konversi progesteron (P4) konversi ke estradiol
(E2). Peningkatan E2/P4 merangsang reseptor oksitosin miometrium,
memproduksi prostaglandin, dan memicu sintesis dan pelepasan oksitosin di
hipofisis, dan pada akhirnya menyebabkan persalinan. Sementara itu pada manusia
masih menjadi teka-teki apakah yang diaktifkan adalah jalur yang sama dengan
domba. Penelitian mengenai mekanisme ini telah dimulai seiring dengan
penemuan oksitosin pada awal 1900-an, prostaglandin, faktor pertumbuhan
(growth factor), sitokin, oksida nitrat, endothelins, dan, yang terbaru, cortisol-
releasing factor plasenta telah dianggap sebagai mekanisme yang mungkin
terlibat.2
Pada awal kehamilan, pertumbuhan uterus disebabkan oleh hiperplasia sel otot
polos, dengan hipertrofi (mungkin disebabkan oleh peregangan) menyebabkan
kehamilan bisa terus berlanjut. Asal usul miosit baru tidak diketahui, tetapi
mungkin timbul dari populasi sel induk miometrium. Dengan semakin

2
berkembangnya kehamilan, miometrium dipertahankan dalam keadaan ”tenang”,
memungkinkan janin untuk tetap bertahan di dalam uterus. Serviks uterus tetap
tertutup, menjaga janin dalam rahim meskipun berat badan janin terus meningkat
ditambah dengan plasenta dan amnion. Menjelang akhir kehamilan, serviks mulai
mengalami perubahan, mengalami pembukaan sebagai akibat kontraksi uterus.
Perubahan serviks ini dikenal sebagai pematangan serviks, dan perubahan
struktural ini dapat diidentifikasi pada pemeriksaan beberapa hari sebelum onset
persalinan.1,2,3
Tanda-tanda persalinan yang paling jelas adalah timbulnya kontraksi reguler
miometrium, yang dapat disertai dengan pecahnya ketuban. Bukti ini
menunjukkan bahwa proses ini lebih banyak dipicu reaksi inflamasi. Beberapa
kelompok peneliti mulai tertarik dengan hipotesis ini dan menyelidiki apakah agen
anti-inflamasi yang dapat digunakan sebagai terapi persalinan prematur. Baru-baru
ini, prokineticins telah terbukti menginduksi mediator pro-inflamasi dalam
jaringan kehamilan manusia.1
Tujuan penulisan referat ini adalah untuk menelaah peranan prokineticin
dalam menginflamasi miometrium dan menginisiasi persalinan.
II. TEORI INISIASI PERSALINAN
A. Faktor Endokrin
Mekanisme persalinan manusia tidak sepenuhnya dipahami. Mekanisme
partus yang telah dipahami dengan baik adalah mekanisme pada domba, dan
penerapan mekanisme ini pada manusia cukup memuaskan. Prinsip dasar
persalinan adalah adanya aktivasi adrenal janin yang menyebabkan
peningkatan produksi kortisol, lalu kortisol akan menginduksi peningkatan
aktivitas 17-m-hidroksilase plasenta dan C-17 lyase, dan pergeseran plasenta
dari memperoduksi progesteron ke estrogen. Akibatnya, tingkat estrogen
dalam plasma janin dan ibu meningkat dan progesteron menurun. Perubaan
mendasar lingkungan hormon steroid dihubungkan dengan peningkatan

3
produksi prostaglandin oleh kotiledon ibu (plasenta), miometrium dan selaput
janin, 'dan juga dengan semakin meningkatnya aktivitas oksitosin.4
Sementara estrogen merangsang produksi prostaglandin intrauterin,
penurunan progesteron meningkatkan sintesis prostaglandin. Pada peneleitian
yang dilkuakn oleh Schwartz, dkkdengan mengamati perubahan hormonal
pada domba selama masa kehamilan didapatkan formasi penurunan
progesteron dan peningkatan estrogen pada domba berhubungan dengan
pembentukan miosit, sehingga meningkatkan kepekaan miometrium.
Perubahan yang disebabkan oleh steroid ini menyebabkan persalinan pada
domba.2,4
Pada wanita hamil, sebelum persalinan terjadi peningkatan glukokortikoid
janin dan estrogen plasenta, namun, tidak ada perubahan tingkat progesteron
dalam plasma perifer ibu yang terukur. Berbeda dengan plasenta domba, tidak
mungkin untuk memicu 17-a-hidroksilase atau Cl 7,20 lyase dalam plasenta
manusia. Progesteron dapat dibentuk oleh plasenta manusia baik dari
lipoprotein low-density ibu atau dari pregnenolon-sulfat janin, tetapi tidak
dikonversi langsung menjadi estrogen dalam plasenta. Sebaliknya, estrogen
plasenta terbentuk dari prekursor CL9 yang disediakan baik oleh adrenal janin
atau ibu. Akibatnya, penurunan progesterone plasenta yang memicu
persalinan di domba betina hamil tidak terjadi pada wanita. 4
Meskipun ada perbedaan biokimia dasar sebelum persalinan pada
manusia dan domba, peningkatan produksi prostaglandin intrauterine terjadi
pada kedua spesies selama persalinan. Bahkan, peningkatan produksi
prostaglandin intrauterine adalah sebuah temuan yang konsisten dalam setiap
penelitian inisiasi kelahiran mamalia. Pada kehamilan manusia, prostaglandin
dapat menginduksi persalinan setiap saat selama kehamilan, inhibitor sintesis
prostaglandin dapat memperpanjang kehamilan, dan prostaglandin ampuh
berperan sebagai agen kontraktil miometrium agen kontraktil, sehingga
memiliki peran kunci inisiasi persalinan.2,3,4

4
Berikut ini adalah ulasan mengenai peran sumbu hipotalamus-
pituitary-adrenal diikuti oleh peran zat-zat kunci dalam mekanisme persalinan,
yaitu progesteron, estrogen, oksitosin, dan prostaglandin.
1. Aksis Hipotalamus-Hipofisis-Adrenal
Pematangan dan aktivasi sumbu hypothalamicpituitary- adrenal pada
janin domba telah banyak diulas. Secara sederhana, mekanisme adalah :
peningkatan sekresi corticotropin- releasing hormone (CRH)
berhubungan dengan peningkatan produksi hormon adrenocorticotropin
(ACTH) yang menyebabkan pematangan dan peningkatan cortisol janin.
Namun mekanisme ini kurang jelas dalam persalinan manusia. Janin
anencephaly tanpa oligohidramnion tidak memperpanjang usia
kehamilan. Pengukuran CRH janin, ACTH dan kortisol tidak dapat
diperoleh dalam kehamilan manusia, tetapi data yang tersedia (darah tali
pusat, cairan ketuban) tidak membuktikan korelasi yang sama dengan
mekanisme pada domba. Namun hal yang penting untuk dicatat adalah
bahwa plasenta dan selaput janin dapat menghasilkan CRH dan ACTH. 3,4
2. Progesteron
Progesteron diperlukan untuk pemeliharaan uterus selama kehamilan
di sebagian besar mamalia. Dalam spesies non-primata, persalinan
didahului dengan peningkatan rasio estrogen. Sebelumnya telah
dinyatakan bahwa diperlukan penghilangan blok progesteron untuk
memicu aktivitas uterus yang diperlukan untuk memicu persalinan
manusia. Namun, dalam manusia dan monyet rhesus, konsentrasi plasma
progesterone dan estrogen perifer tidak berubah secara signifikan tepat
sebelum persalinan aterm dan preterm. Sebuah studi baru-baru ini,
mengukur estriol dan progesterone dalam saliva wanita sebelum, selama,
dan setelah persalinan spontan, penelitian ini juga tidak menemukan

5
perubahan konsentrasi steroid yang signifikan segera sebelum atau selama
persalinan. Di sisi lain antagonis RU486, menginduksi aborsi pada wanita
selama trimester pertama dan persalinan aterm. Selain itu, ditemukan
peningkatan estradiol dalam uterus wanita hamil. Pengamatan ini
menunjukkan bahwa perubahan dalam rasio estrogen progesteron
mungkin memainkan peran dalam mengontrol aktivitas uterus. Sebuah
studi di monyet rhesus di akhir kehamilan, telah menunjukkan bahwa
meskipun pemberian RU486 akan menginduksi aktivitas rahim, efeknya
tidak akan menginduksi hingga menyebabkan kelahiran. Selain itu, dari
catatan kehamilan pada wanita dengan hypobetalipoproteinemia di
antaranya ada memiliki penurunan tingkat sirkulasi progesteron.3
Telah dikemukakan bahwa modulasi biosintesis progesteron dan
estrogen dalam membran janin manusia dapat terjadi pada jangka waktu
panjang sehingga daerah penurunan progesteron dan peningkatan
estradiol tidak akan terdeteksi oleh pengukuran konsentrasi di sirkulasi
perifer ibu. Amnion, chorion dan desidua pada manusia adalah jaringan
steroidogenik, estrogen dan progesteron terutama diproduksi dari
precursor sulfat. 3
Mitchell & Challis menemukan bahwa aktivitas sulfohydrolase steroid
di chorion meningkat setelah persalinan spontan. Enzim 17P, 20cL-
hidroksisteroid dehidrogenase (17P, 20a-HSD) juga hadir dalam
membran janin, desidua dan plasenta. Enzim ini mengkatalisis konversi
estron agar membentuk estradiol yang secara biologis lebih aktif,
demikian pula dengan progesteron menjadi progestin tidak aktif 20-a
dihydroprogesterone.3
Ada beberapa bukti bahwa aktivitas 17P, 20a-HSD di plasenta
meningkat dalam jangka waktu panjang. Pengaturan pasokan substrat dan
dan progesteron lokal dalam membran janin. Perubahan estrogen dan

6
progesteron di awal persalinan akan berpengaruh pada produksi
prostaglandin lokal.4
3. Estrogen
Estrogen diperkirakan memainkan peran kunci dalam mekanisme
persalinan di banyak spesies hewan. Pada wanita, seperti yang
disebutkan sebelumnya, pemberian estrogen akan menginduksi aktivitas
uterus. Estrogen diketahui menginduksi reseptor oksitosin, sekresi
oksitosin, dan peningkatan produksi prostaglandin. Namun demikian
tidak ada perubahan dalam tingkat sirkulasi estrogen pada persalinan
wanita. Awal persalinan pada wanita yang diberikan RU486 atau inhibitor
3P-hidroksisteroid dehidrogenase (3p-HSD) tidak menyebabkan
perubahan aktivitas estrogen. Mayoritas wanita dengan defisiensi
sulphatase steroid akan memulai persalinan aterm. Dalam situasi tersebut,
tingkat sirkulasi estrogen unconjugated (yang rendah) tidak meningkat
pada awal persalinan.4
4. Oksitosin
Secara historis oksitosin dianggap bertanggung jawab sebagai faktor
primer onset persalinan. Oksitosin adalah salah agen kontraktil yang
paling ampuh dan spesifik. Oksitosin eksogen diberikan untuk
menginduksi kontraksi persalinan dan efektif dalam mendorong
persalinan di semua spesies mamalia yang dipelajari. Namun, peran
oxytocin endogen dalam inisiasi persalinan manusia tetap meragukan
karena kadarnya dalam plasma ibu tidak meningkat hingga tahap kedua
persalinan. Tingkat oksitosin yang relatif tinggi ditemukan dalam darah
tali pusat (arteri > vena) setelah yang persalinan normal, hal ini
menunjukkan bahwa oksitosin janin mungkin memainkan peran dalam

7
inisiasi persalinan. Tingginya kadar oxytocinase di cairan ketuban dan
plasma ibu dapat membantah perdebatan mengenai hal ini.3,4
Ketidakmampuan untuk menunjukkan peningkatan kadar oksitosin
sebelum persalinan telah menyebabkan keyakinan bahwa oksitosin tidak
penting untuk inisiasi persalinan melainkan berfungsi untuk
mengintensifkan persalinan. Namun, beberapa telah mendalilkan bahwa
meningkatkan sensitivitas miometrium ke tingkat basal oksitosin saat
aterm adalah kunci onset persalinan. Dukungan untuk hipotesis ini berasal
dari pengamatan bahwa jumlah reseptor oksitosin meningkat dalam
miometrium dan desidua wanita selama persalinan. Hingga saat ini belum
diketahui apa yang menginduksi peningkatan jumlah reseptor oksitosin,
tetapi telah diketahui bahwa oksitosin dikendalikan oleh lingkungan
hormon steroid.4
Selain memicu kontraktilitas uterus ada bukti bahwa oksitosin juga
akan menstimulasi biosintesis prostaglandin di beberapa jaringan.
Oksitosin merangsang produksi prostaglandin dalam desidua manusia dan
selaput janin, dan efek ini secara signifikan lebih besar dalam jaringan
dari wanita bersalin. Data ini menunjukkan oksitosin mungkin penting
dalam merangsang biosintesis prostaglandin intrauterine saat aterm.3,4
Baru-baru ini, telah ditunjukkan bahwa jaringan intrauterin dapat
menghasilkan oksitosin. Selain itu, telah ada bukti bahwa ada ekspresi
gen oksitosin 3-4 pada saat persalinan. Hal ini menyediakan mekanisme
dimana oksitosin bisa menghindari konsentrasi oxytocinase tinggi pada
wanita.2
5. Prostaglandin
Prostaglandin dianggap sebagai kunci awal persalinan. Bukti untuk
mendukung pandangan ini telah banyak diulas. Partus disertai dengan
meningkatnya konsentrasi prostaglandin dalam darah, urin, dan cairan

8
ketuban. Produksi prostaglandin dalam jaringan intrauterin (amnion dan
desidua) meningkat saat persalinan. Pemberian prostaglandin akan
menginduksi persalinan atau aborsi pada setiap tahap kehamilan.
Prostaglandin juga berperan dalam pematangan serviks. Pemberian
inhibitor sintesis prostaglandin (yaitu non-steroid anti-inflammatory
drugs) akan menunda persalinan dan memperpanjang interval dalam
induksi aborsi. Studi terbaru menunjukkan bahwa prostaglandin berperan
dalam persiapan persalinan serta mekanisme yang sebenarnya secara
langsung mengakibatkan kontraksi rahim.3,4
B. Faktor Inflamatori
Konsep persalinan tradisional dan bukti pada hewan, menunjukkan bahwa
persalinan adalah peristiwa hormonal, terutama didorong oleh perubahan
progesteron dan/atau sintesis kortisol. Model seperti itu dapat diterapkan pada
partus manusia tetapi seringkali tidak cocok. Selama 10 tahun terakhir, beberapa
peneliti telah mengusulkan bahwa peristiwa-peristiwa utama persalinan manusia
adalah proses inflamasi, dengan perubahan hormonal yang bersifat sekunder atau
pelengkap.5
Bukan pengamatan baru bahwa persalinan pada manusia adalah sebuah proses
inflamasi, namun dalam beberapa tahun terakhir telah banyak perkembangan baru
yang menarik telah membantu memperkuat dan mengambangkan konsep ini. Pada
tingkat yang paling dasar, konsep ini didukung oleh adanya leukosit, (sebagian
besar neutrofil dan makrofag, dan juga sel T) yang menginvasi miometrium,
serviks, dan membran fetal sesaat atau segera setelah awal persalinan, yang dipicu
oleh adanya peningkatan ekspresi chemokine dan molekul adhesi sel dalam
jaringan. Secara paralel, leukosit yang beredar dalam darah perifer meningkatkan
ekspresi adhesi sel molekul ligan seperti CD11a dan CD11b, memfasilitasi
emigrasi leukosit ke dalam miometrium dan serviks.6

9
Peningkatan ekspresi sitokin dalam miometrium dan serviks dalam persalinan
muncul di bagian yang diserang leukosit dengan bukti terbaru menunjukkan
bahwa kapasitas peningkatan leukosit selama persalinan untuk mengekspresikan
sitokin dimulai dalam sirkulasi perifer. Pro-inflamasi sitokin memiliki banyak efek
pada jaringan reproduksi, termasuk efek stimulasi kontraktilitas miometrium
melalui sejumlah mekanisme yang berbeda, yang akan dibahas di bawah ini.
Sitokin juga terlibat dalam renovasi jaringan Dan menarik lebih banyak leukosit
dalam mekanisme umpan balik positif, yang meaugmentasi proses persalinan.
Aktivitas faktor nuclear kappaB (NF-KB), sebuah faktor transkripsi yang
mengatur gen yang terlibat dalam berbagai proses seluler termasuk peradangan,
juga meningkat pada persalinan. Peningkatan NF-KB paling jelas dalam
hubungannya dengan persalinan terjadi di membran janin, meskipun NF-KB juga
muncul dalam miometrium. NF-KB dapat menyediakan mekanisme untuk memicu
proses inflamasi dalam persalinan, karena NF-KB sendiri diinduksi oleh stimuli
pro-inflamasi.1,5
Persalinan berhubungan dengan invasi leukosit, aktivasi NF-KB dan produksi
pro-inflamasi sitokin dalam beberapa jaringan reproduksi yang relevan. Data ini
sangat mendukung hipotesis bahwa persalinan merupakan proses inflamasi.
Dampak fungsional dari kejadian pro-inflamasi dijelaskan di bawah ini.6
1. Miometrium
Upregulasi pro-inflamasi sitokin dalam miometrium menjelang persalinan
merangsang dan memperkuat kontraksi uterus. Tribe dkk telah
menunjukkan bahwa interleukin (IL)-1β menginduksi masuknya kalsium
basal dan masuknya simpanan kalsium di sel otot polos miometrium,
sehingga secara langsung meningkatkan kontraktilitas. Selain itu, IL-1β
dan tumor nekrosis faktor (TNF)–α merangsang pelepasan asam
arakidonat, dan ekspresi siklooksigenase (COX)-2 sehingga meningkatkan

10
produksi prostaglandin di kultur sel miometrium, dengan aktivitas NF-KB
yang lebih besar.1,6
Prostaglandin telah lama dikenal sebagai stimulator dari kontraksi
miometrim sehingga ada potensi hubungan kausal yang jelas antara proses
inflamasi yang diamati pada miometrium dan proses klinis persalinan.6
2. Serviks
Pematangan serviks ditandai dengan kerusakan kolagen dan renovasi
elemen jaringan ikat pada serviks. Invasi leukosit dan peningkatan
konsentrasi protein IL-8, IL-6 dan granulocyte-CSF (G-CSF) diamati
dalam jaringan ikat serviks aterm manusia dan selanjutnya renovasi serviks
sangat penting untuk memfasilitasi pembukaan selama persalinan. Secara
fungsional, IL-8 dapat merangsang pelepasan degradatif enzim dari
neutrofil. Enzim ini termasuk protease serin dan matriks metalloproteinase
((MMP) -8 dan (MMP-9)) yang kemudian berpartisipasi dalam degradasi
kolagen dan glikosaminoglikan, komponen matriks ekstraseluler sehingga
menyebabkan pematangan serviks. Produksi MMP-1, MMP-3 dan MMP-9
dari fibroblas dan sel-sel otot polos mungkin juga ditingkatkan oleh IL-1β
dalam leher rahim sementara mendownregulasi ekspresi inhibitor jaringan
metalloproteinase (TIMP)-2, inhibitor endogen MMP-2, dengan peristiwa
yang lebih diperkuat oleh produksi NF-KB. NF-KB sendiri dapat
diaktifkan melalui toll-like receptor (TLR)-4 oleh protein matriks
ekstraseluler seperti fragmen fibronektin janin, yang rusak selama renovasi
serviks, sehingga memperkuat proses peradangan yang sedang
berlangsung. Dengan demikian, di akhir kehamilan dan selama persalinan,
serviks tetap kokoh dan tertutup meskipun tekanan meningkat selama
kehamilan, lalu di bawah pengaruh sejumlah mediator inflamasi, serviks
melembut dan melebar.1,6

11
3. Membran Janin
Pola pro-inflamasi serupa dengan yang diamati dalam miometrium terlihat
dalam membran janin. Produksi IL-8, TNF-α, IL-6 dan IL-1β semua
meningkat pada membran janin dan cairan ketuban selama persalinan.
Ditemukan juga peningkatan kadar MMP-9, dan penurunan tingkat TIMPs.
Secara fungsional, rasio MMP/TIMP yang meningkat melemahkan
membran janin, memfasilitasi pecahnya ketuban yang sering menjadi
prekursor persalinan.6
Pemeriksaan selaput janin dari spesimen pra-persalinan yang diperoleh
pada operasi caesar telah menunjukkan bahwa zona kelemahan ada di
wilayah yang melapisi serviks. Fitur dari zona ini termasuk penurunan
ketebalan dibandingkan dengan sisa selaput janin, peningkatan MMP-9 dan
penurunan TIMP-3 dan juga peningkatan pembelahan poli (ADP-ribose)
polimerase-1 (PARP1). PARP1 adalah enzim yang terlibat dalam
perbaikan DNA dan apoptosis dan dengan demikian peningkatannya
menunjukkan peningkatan apoptosis. Tampaknya menjelang akhir
kehamilan selaput janin dipersiapkan untuk persalinan melaluo proses
renovasi untuk membentuk daerah lemah yang akan menjadi lokasi ruptur.
Apa yang memulai dan mengembangkan proses ini tidak diketahui, tetapi
pemahaman yang lebih besar mengenai peristiwa molekuler yang
menyebabkan pecahnya membran mengarah pada pengembangan strategi
terapi baru untuk menghentikan persalinan onset prematur.6
Membran janin memiliki peran penting, karena mereka adalah gerbang
antara janin dan ibu, dan dapat mengirimkan sinyal pra-persalinan dari bayi
ke miometrium ibu dan serviks. Protein surfaktan paru yang berasal dan
fosfolipid mungkin menjadi kunci untuk hal ini: dengan semakin
matangnya janin, semakin banyak jumlah yang diproduksi maka kadarnya
akan meningkat dalam cairan ketuban. Protein surfaktan ini juga dapat
berkontribusi pada respon inflamasi dalam membran janin dan kemudian

12
akan mempengaruhi serviks dan miometrium mengingat bahwa mereka
merangsang aktivitas COX-2 melalui TLR-4 pada tikus dan prostaglandin
E2 berikutnya (PGE2) produksi dalam amnion.6
Peningkatan produksi PGE2 amnion dapat menyebar melalui korion dan
desidua untuk merangsang produksi prostaglandin di sel miometrium dan
karenanya menyebabkan kontraktilitas. Janin dan plasenta juga dapat
bersama-sama memicu pecahnya ketuban karena prostaglandin, TNF-β dan
corticotropin releasing hormon (CRH) (yang juga meningkat dalam cairan
amnion aterm) merangsang produksi MMP-9, yang melemahkan membran
dan dengan demikian memfasilitasi pecahnya ketuban.6
Jadi jika onset persalinan manusia memiliki fitur perubahan inflamasi
dan endokrin, mana yang lebih dominan? Ada sedikit bukti menarik yang
menunjukkan bahwa pada manusia perubahan hormonal merupakan sinyal
akhir kehamilan dan memulai proses persalinan. Tidak seperti spesies
hewan, manusia tampaknya tidak tergantung pada penurunan progesteron,
maupun peningkatan estrogen untuk memulai persalinan. Sebaliknya,
tampaknya banyak proses yang kompleks dan saling terkait yang tergabung
untuk mengakhiri fase “tenang” kehamilan dan memulai persalinan.
Mitchell dan Taggart telah menyarankan model akumulasi modular sistem
fisiologis dimana beberapa proses fisiologis saling berhubungan dan
berkembang secara paralel sampai massa kritis tercapai dan menghasilkan
persalinan. Model mereka juga menjelaskan bagaimana persalinan
prematur dan aktivasi dari beberapa 'modul' yang tidak tepat bisa
mengakibatkan persalinan prematur sehingga konsep seperti itu bisa
menjelaskan terjadinya persalinan aterm dan prematur. Penelitian lain
mengusulkan bahwa hubungan antara proses tersebut didominasi oleh
sistem inflamasi, peranan hormon dalam partus manusia telah diragukan,
sistem kekebalan tubuh memiliki peranan yang lebih penting.1,6

13
Progesteron telah lama digambarkan sebagai steroid anti-inflamasi dan
ini didukung oleh banyak data yang menunjukkan interaksinya dengan
sistem kekebalan tubuh. Progesteron sendiri menghambat produksi IL-6
dalam arteri fetoplasenta, dan progestogen seperti medroxyprogesterone
acetate menghambat berbagai sitokin pro-inflamasi dan chemokin di
miometrium manusia. Yang menarik adalah interaksi progesteron dengan
NF- KB, faktor transkripsi yang memiliki fungsi kunci dalam partus
manusia. NF-KB meningkat dalam membran janin selama persalinan,
tetapi secara negatif ditekan oleh aktivasi reseptor progesteron. Sebaliknya,
NF-KB sendiri merepresi aktivitas reseptor progesteron, penelitian dari
kelompok Carole Mendelson menunjukkan bahwa aktivasi NF-KB di
miometrium manusia meregulasi isoform inhibitor PR. Penghilangan
imunosupresif mempengaruhi dengan withdrawal progesteron fungsional.
Fakta ini diinisiasi oleh peradangan dan aktivasi NF-KB.1,6
Estrogen dapat menginduksi baik pro-inflamasi atau efek anti-inflamasi,
tergantung pada rangsangan kekebalan tubuh, jenis sel yang terlibat, organ
target, konsentrasi estrogen dan reseptor. Sehubungan dengan ekspresi
sitokin, estrogen dalam konsentrasi tinggi yang terlihat pada kehamilan
dapat menghambat jalur pro-inflamasi yang diaktifkan oleh IL-1β, IL-6 ,
IL-8 dan TNF-α serta menghambat aktivitas natural killer cells.
Sebaliknya, sekresi anti-inflamasi interleukin IL-4, IL-10 dan perubahan
faktor pertumbuhan (TGF)-β dirangsang oleh estrogen tinggi.6
Meskipun efek anti-inflamasi jelas dari estrogen, estradiol in vivo dapat
meningkatkan produksi pro-inflamasi sitokin oleh sel T CD4+
kemungkinan melalui efek langsung sirkulasi leukosit yang
mengekspresikan ER-α. Dengan demikian, meskipun estrogen memiliki
efek stimulasi pada miometrium, interaksi yang sangat sistem kekebalan
tubuh yang kompleks mungkin berkontribusi pada ketenangan yang
dipertahankan selama kehamilan.5

14
CRH dan sumbu HPA sangat erat kaitannya dengan sistem kekebalan
tubuh dan memiliki efek kompleks pada reproduksi sistem wanita manusia.
CRH telah terbukti mengatur produksi sitokin, dan akibatnya inflamasi
sitokin mempengaruhi produksi CRH. Baik TNF-α dan IL-1β merangsang
produksi CRH, dan sebaliknya CRH menghambat produksi IL-1 dan IL-6
oleh sel mononuklear darah perifer. CRH dapat mengurangi sitotoksisitas
natural killer cells dan juga menghambat proliferasi sel. Peningkatan
produksi kortisol janin, juga terkait dengan pematangan paru janin dan
produksi protein surfaktan dan fosfolipid yang hadir dalam cairan ketuban.
Senyawa-senyawa tersebut adalah pro-inflamasi dan diperkirakan bahwa
mereka mungkin menstimulasi produksi prostaglandin dan menginisiasi
peradangan pada membran janin.1,6
Efek inflamasi sitokin pada OTR agak bertentangan dengan yang
dijelaskan dalam literatur. Schmid dkk melaporkan downregulation mRNA
OTR oleh IL-1β dalam miometrium, dan Rauk et al. juga melaporkan
downregulation OTR mRNA setelah pengobatan dengan IL-1β dan setelah
pengobatan dengan IL-6 di SD sel otot halus manusia. Namun, Terzidou et
al. menunjukkan upregulation transien mRNA OTR dengan pengobatan
miosit primer dengan IL-1β.
Promotor OTR berisi situs binding transkripsi putatif CCAAT/enhancer
binding protein (C/EBP) dan NF-KB, yang merupakan faktor transkripsi
yang diaktifkan oleh sitokin seperti IL-1β dan IL-6. IL-1β dan IL-6
meningkatkan sekresi oksitosin dalam sel otot polos uterus dan IL-1β telah
diamati menambah kontraktilitas miometrium yang dirangsang oksitosin.
Dengan demikian, ada kemungkinan bahwa hasil dari infiltrasi sel
inflamasi dan peningkatan sitokin pro-inflamasi adalah upaya untuk
mengaktifkan jaringan miometrium (setidaknya sebagian melalui
peningkatan ekspresi OTR) dan untuk mempromosikan kontraktilitas.1,6

15
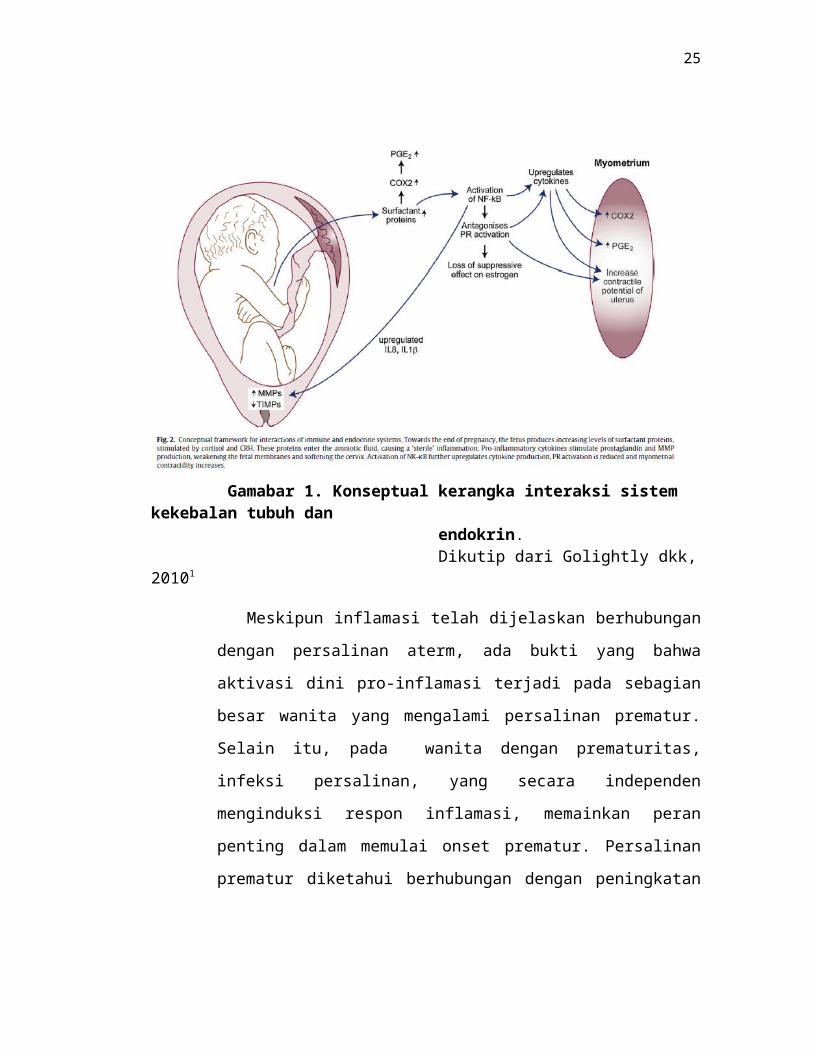
Gamabar 1. Konseptual kerangka interaksi sistem kekebalan tubuh dan endokrin. Dikutip dari Golightly dkk, 20101
Meskipun inflamasi telah dijelaskan berhubungan dengan persalinan
aterm, ada bukti yang bahwa aktivasi dini pro-inflamasi terjadi pada
sebagian besar wanita yang mengalami persalinan prematur. Selain itu,
pada wanita dengan prematuritas, infeksi persalinan, yang secara
independen menginduksi respon inflamasi, memainkan peran penting
dalam memulai onset prematur. Persalinan prematur diketahui
berhubungan dengan peningkatan kadar IL6 dan sitokin pro-inflamasi
lainnya dalam cairan ketuban, yang seringkali muncul akibat infeksi.
Lainnya telah menunjukkan peningkatan kadar pro-inflamasi sitokin dalam
cairan serviks wanita baik dalam persalinan prematur aktif atau mereka
yang dalam ancaman prematur.1,6
Mengingat peran inflamasi pada awal persalinan, beberapa kelompok
telah menyelidiki agen anti-inflamasi untuk mengatasi persalinan prematur.
Antagonis IL1b menghambat IL1b yang menginduksi persalinan prematur

16
pada tikus. Data menunjukkan bahwa IL10 dan prostaglandin J2
menghambat lipopolisakarida (LPS) yang menginduksi persalinan
prematur pada tikus, keduanya tampaknya menghambat kontraksi rahim.
Baru-baru ini, prokineticins telah ditunjukkan memiliki peran dalam
menginduksi pro-inflamasi mediator dalam jaringan kehamilan manusia.1
III. PROKINETICIN
Pada tahun 1999 sebuah protein kecil kaya residu sistein diisolasi dari sekresi
kulit katak Bombina variegata dan diberi nama Bv8, untuk menunjukkan asal-
usulnya (B. variegata) dan yang berat molekul (kDa 8). Bv8 juga ditemukan di
guinea pig, ileum tikus, usus tikus, dan induksi hiperalgesia pada tikus. Karena
banyaknya urutan peptida bioaktif yang disekresikan oleh kelenjar olocryne kulit
amfibi yang dilestarikan sepanjang evolusi untuk mempertahankan selular dasar
atau proses perkembangan, tingginya tingkat identitas antara amfibi, ikan dan
reptil, peptida menyerupai Bv8 mungkin juga ditemukan pada mamalia. Dalam
beberapa tahun setelah penemuan Bv8, kloning cDNA mengkonfirmasikan
adanya Bv8 hortologues di tikus, sapi,dan monyet.7
Protein ini di mamalia diberi nama prokineticin1 (PK1 atau EGVEGF,
endocrine gland-derived vascular endothelial growth factor) dan prokineticin2
(PK2 atau mBv8) karena kemampuan mereka untuk mengkontraksikan ileum
marmot. Mereka dikosifikasikan oleh dua gen berbeda, PROK1 dan PROK2.
Bentuk kedua dari PK2 telah diidentifikasi dan diberi nama PK2b karena
menyisipkan 21 dasar asam amino dalam urutan nya.7
Tiga kelompok independen kemudian mengidentifikasi dua reseptor G-
protein-coupled yang berhubungan erat untuk Bv8/PKs, prokineticin reseptor 1
(PKR1) dan reseptor prokineticin 2 (PKR2). Penelitian intensif selama beberapa
tahun terakhir telah menunjukkan bahwa aktivitas biologis protein
Bv8/Prokineticin mencakup angiogenesis dan keterlibatan dalam reproduksi dan
kanker, survival neuronal dan neurogenesis, hormon hipotalamus. Sekresi,

17
kontrol ritme sirkadian dan modulasi kompleks perilaku, seperti makan dan
minum. Tingginya tingkat ekspresi Bv8/PK2 manusia dalam tulang sumsum,
organ limfoid dan leukosit menunjukkan keterlibatan peptida ini di
hematopoiesis dan inflamasi dan proses imunomodulator. Selain itu, penurunan
ambang nyeri dramatis yang dihasilkan oleh Bv8 bertindak pada PKRs dalam
neuron sensorik menunjukkan bahwa Bv8/prokineticins dan reseptor mereka
dapat bertindak sebagai mediator inflamasi dan nyeri neuropatik.7,8
1. Struktur Protein
Prokinecticins (PKs) adalah dua protein yang baru diidentifikasi dan berperan
dalam kondisi fisiologis dan patologis. Istilah PK1 dan PK2 dikemukakan
oleh Li et al. untuk mencerminkan fungsi mereka dalam mendorong kontraksi
spesifik dan kuat pada otot polos saluran pencernaan. Selanjutnya, LeCouter
et al. menjelaskan faktor pertumbuhan yang memicu respon mitogenik yang
kuat dan direproduksi dalam kelenjar endokrin yang diturunkan sel endotel.
Efek yang sama yang disebabkan oleh protein ini dan oleh faktor
pertumbuhan endotel vaskular (VEGF) maka protein ini diberi nama kelenjar
endokrin VEGF (EG-VEGF). Meskipun ada beberapa kesamaan dalam fungsi
dan mekanisme kontrol VEGF dan EG-VEGF, dua faktor ini secara struktural
tidak berhubungan. Urutan asam amino untuk PK1 dan EG-VEGF identik,
dan, untuk tujuan ulasan ini, istilah 'PK1' yang akan digunakan di seluruh
pembahasan.7,8,9
PK1 adalah ortholog manusia yang diisolasi dari protein tidak beracun dari
bisa ular mamba hitam (Dendroaspis polylepis) dan diberi nama protein
venom A (VPRA) atau mamba intestinal toxin 1 (MIT1), karena
kemampuannya untuk mengkontraksikan ileum marmot. PK1 adalah 80%
homolog dengan VPRA/MIT1 dan merupakan protein yang matang dari 86
asam amino, dengan sinyal peptida dari 19 asam amino. Gen yang mengkode

18
prekursor PK1 terletak pada kromosom 1p21 manusia dan dikodekan oleh tiga
ekson.8
PK2 adalah paralog manusia PK1 dan ortholog dari protein yang diisolasi
dari sekresi kulit katak Bombina variegata, yang dikenal sebagai Bv8. Sebuah
varian PK2 telah dijelaskan dalam manusia, tikus dan testis banteng, memiliki
insert arginin dan lisin kaya residu 21 asam amino. Hal ini menyebabkan
varian yang disebut 'Bv8-dasar' (Bv8-b). Prekursor PK2 terletak pada
kromosom manusia 3p21.1 dan dikodekan oleh empat ekson, dengan ekson
ketiga menjadi subjek splicing alternatif.8,9
PK1 dan PK2 berbagi 44% identitas asam amino dan juga berbagi struktur
umum motif protein. Mereka memiliki urutan N-terminal (AVITGA), yang
penting untuk aktivitas dari protein. Mutasi untuk urutan ini, dengan
memasukkan sebuah metionin sebelum N-terminal alanin, substitusi dari
alanin N-terminal dengan metionin atau penghapusan dari dua asam amino
pertama, akan menghasilkan antagonis PK reseptor (PKR). Fitur lain dari PKS
adalah adanya sepuluh sistein kekal, yang diperkirakan akan membentuk lima
ikatan disulfida. Ikatan-ikatan disulfida ini diperkirakan untuk membentuk
lipatan dalam molekul PK1 mirip dengan yang dibentuk dalam colipase
protein terkait, dan Dickkopfs, yang penting untuk aktivitas protein
rekombinan, mutasi sistein atau penggantian sistein menghasilkan protein
yang memiliki aktivitas yang sama dengan PKRs.7,8
PKS adalah ligan untuk dua G-protein-coupled receptors (GPCRs) yang
saling terkait erat, disebut PKR1 dan PKR2, yang berbagi identitas asam
amino 85% dan menunjukkan perbedaan terbesar dalam urutan N-terminal
mereka. Urutan mereka hampir identik dalam domain transmembran,
menunjukkan bahwa mekanisme aktivasi mereka identik dan molekul analog
kecil tidak akan bisa dibedakan, seperti halnya PK1 dan PK2. Afinitas dari
faktor-faktor reseptor mereka mirip, dengan PK2 yang menunjukkan afinitas
yang cukup tinggi untuk kedua reseptor.7

19
2. Reseptor
Semua prokineticins mengerahkan fungsi biologis mereka melalui aktivasi
dari dua protein G- reseptor, prokineticin reseptor-1 dan -2 (PKR1 dan PK-
R2) yang dikode oleh gen yang terletak di kromosom manusia daerah 2q14
dan 20p13. Reseptor ini berbagi 85% identitas asam amino dan berbeda
terutama dalam urutan N-terminal mereka. Reseptor berpasangan Gq, Gi dan
Gs untuk memediasi mobilisasi kalsium intraseluler, fosforilasi p44/p42
mitogen yang diaktifkan protein kinase, serin/treonin kinase Akt dan cAMP.
Aktivasi reseptor telah menunjukkan proses proliferasi, anti-apoptosis,
diferensiasi dan migrasi/mobilisasi sel target di berbagai sistem. Pola ekspresi
prokineticins dan reseptor mereka telah dilaporkan dalam berbagai jaringan,
yang memberikan isyarat untuk fungsi biologis spesifik dalam jaringan. Selain
itu, Pola ekspresi G-protein diferensial dan beberapa G-protein coupling dari
reseptor lebih meningkatkan kompleksitas fungsional dari sistem. Ini
memungkinkan sel-sel untuk melakukan fungsi fisiologis yang berbeda dalam
menanggapi rangsangan ligan yang sama.7,9
Ekspresi profil dinamis Prok-1 juga telah dilaporkan dalam uterus,
ovarium dan plasenta dalam menanggapi perubahan hormonal seluruh siklus
menstruasi dan selama kehamilan. Mengenai ekspresi spesifik jaringan pada
sistem, diketahui bahwa estrogen, progesteron chorionic, gonadotropin dan
hypoxia-inducible factor (HIF-1α) memediasi ekspresi dinamis dari Prok-1 di
saluran reproduksi. Aktivasi jalur mitogen-activated protein kinase (MAPK),
mungkin melalui faktor transkripsi c-jun/fos, yang sangat penting untuk
ekspresi PK-R1 di pial neural sel enterik.8
Belum banyak yang diketahui mengenai reseptor PKs. Karakterisasi
promotor prokineticins dan reseptor mereka karena akan sangat memudahkan
identifikasi jaringan-spesifik regulator untuk sistem. Hal ini juga dicatat
bahwa ekspresi profil prokineticins dan reseptor mereka dalam tikus sedikit

20
berbeda dari yang di manusia, menunjukkan bahwa inaktivasi dari orthologue
tikus dapat mengakibatkan fenotipe yang mungkin secara substansial berbeda
dari manusia.8
3. Aktivitas Biologis
a). Gastrointestinal
Prok-1 dan Prok-2 memiliki fungsi yang sama dan terlibat dalam berbagai
kegiatan di berbagai jaringan. Nama PKs diambil karena kemampuannya
untuk mengkontraksi usus tikus. PK-R1 dihasilkan oleh pleksus
myenterik dari bagian proksimal usus tikus di mana Prok-1 melemaskan
proksimal usus melalui mekanisme nitrat dimediasi oksida sedangkan
Prok-2 tidak mempengaruhi kontraksi otot tikus. Perbedaan menunjukkan
kekhususan jaringan dan jenis sistem. Sejalan dengan peran regulasi PKs
dalam pleksus myenteric, Prok-1 dan PK-R1 baru-baru ini telah
terdeteksi dalam saraf mukosa/mesenkim dan puncak sel enterik (NCC)
dari usus embrio tikus.
Prok-1 memodulasi baik proliferasi dan diferensiasi NCC enterik
selama perkembangan sistem saraf enterik (ENS), yang pada akhirnya
berkontribusi terhadap pembentukan myenteric dan submukosa pleksus
enterik. Secara khusus, Prok-1 juga menginduksi ekspresi
neurotransmiter, seperti nitrat neuronal oksida sintase (nNOS) dan
vasoaktif usus peptida (VIP), Prok-1 juga dapat memediasi formasi dari
inhibitori musculomotor neuron.7,8,9
b). Jaringan steroidogenik dan saluran reproduksi
Prok-1 juga disebut endocrine gland-derived vascular endothelial growth
factor (EG VEGF) karena bentuknya yang unik dan memiliki efek
angiogenik selektif dalam kelenjar endokrin. Prok-1 ditunjukkan untuk
mempromosikan angiogenesis dalam ovarium dan testis dan

21
menginduksi proliferasi, migrasi dan fenestration sel endotel yang berasal
dari kelenjar adrenal. Namun demikian, tidak seperti faktor pertumbuhan
endotel vaskular (VEGF), Prok-1 adalah mitogen angiogenik yang sangat
spesifik dan hanya mengatur kelenjar endokrin endotelium pembuluh
darah, tetapi tidak sel endotel yang berasal dari aorta vena, umbilkus atau
kornea. Selain sel endotel, Prok-1 juga diekspresikan dalam sel-sel
Leydig testis manusia dan diyakini untuk mempromosikan angiogenesis
interstisial untuk mendukung aktivitas endokrin testis. Ekspresi Prok-2, di
sisi lain, dibatasi hanya pada spermatosit primer. Ekspresi komplementer
Prok-1 dan VEGF telah dilaporkan dalam sel granulosa, yang
mengelilingi sel stroma (sel teka) serta korpus luteum dalam ovarium. Uji
in vitro menunjukkan bahwa Prok-1 mempromosikan proliferasi dan
kelangsungan hidup sel endotel dari korpus luteum sapi. Hal ini juga
secara tidak langsung menginduksi angiogenesis dengan merangsang
ekspresi VEGF. Dalam uterus, ekspresi puncak dari Prok-1 ditemukan
dalam epitel kelenjar midluteal endometrium peri-implantasi,
menunjukkan bahwa mungkin Prok-1 memfasilitasi implantasi dengan
meningkatkan permeabilitas mikrovaskuler endometrium. Pada plasenta,
ekspresi yang kuat dari Prok-1 oleh sinsitiotrofoblas, menunjukkan bahwa
ia juga bertindak sebagai faktor pertumbuhan plasenta untuk diferensiasi
trofoblas.1,7,8,9
c). Sistem saraf sentral dan peripheral
Prok-2, tetapi tidak Prok-1, adalah tombol pengatur beragam proses
biologis dalam sistem saraf pusat. Hal ini banyak diekspresikan dalam
otak tikus dan bertindak sebagai faktor neurotropik endogen untuk
mendukung survival saraf. Selain itu, Prok-2 dan PK-R2 juga banyak
ditemukan di inti suprachiasmatic dan mengendalikan ritme sirkadian
perilaku. Dalam olfactory bulb, Prok-2 menginduksi zona migrasi yang

22
diturunkan subventricular neuronal dan mengatur morfogenesis olfactory
bulb. Dalam sistem saraf perifer, PK-Rs dinyatakan dalam ganglion akar
dorsal (DRG) dan Prok-2 ditemukan untuk terlibat dalam nociception
tersebut. Dalam kultur in vitro pada DRG neuron tikus menunjukkan
bahwa Prok-2 secara signifikan menurunkan ambang nociceptor dari
rangsangan fisik dan kimia. Penelitian selanjutnya menunjukkan bahwa
Prok-2 penyebab hiperalgesia dengan menginduksi reseptor potensial
vanilloid 1 transien (TRPV1) di neuron DRG, pada gilirannya,
mengurangi ambang nociceptive untuk rangsangan termal dan mekanik.
Secara konsisten, Pk-r1-null tikus juga menunjukkan nociception akut
gangguan dan nyeri inflamasi serta sensasi panas.7,8
d). Sumsum tulang, perifer dan kardiomiosit
Peran prokineticins lainnya adalah haematopoiesis dan regulasi dari
respon kekebalan. Prokineticins dapat mempromosikan survival dan
diferensiasi
granulocytic dan moncytic, juga merangsang mobilisasi sel hematopoietik
dan memodulasi respon kekebalan. Peran sinyal Prok-2/PK-R1 tidak
dibatasi hanya dalam mempromosikan pertumbuhan endotel kapiler,
namun juga kelangsungan hidup kardiomiosit. Prok-2, melalui aktivasi
PK-R1, menginduksi pembentukan pembuluh darah jantung dalam kultur
independen VEGF sel endotel. Selain itu, Prok-2 juga mengaktifkan Akt
untuk melindungi kardiomiosit terhadap stres oksidatif dan
menyelamatkan miokardium dari miokard infark dalam model tikus.7,8
IV. PROKINETICIN DAN INISIASI PERSALINAN
A. Prokineticins sebagai mediator inflamasi
Prokineticins diekspresikan oleh sel-sel kekebalan tubuh, termasuk sel B dan T
dan umumnya terdeteksi di lokasi respon inflamasi . Yang penting, reseptor

23
untuk prokineticins umumnya terdeteksi pada permukaan sel-sel kekebalan
tubuh seperti makrofag dan neutrofil, hal ini menunjukkan bahwa prokineticins
mungkin memainkan peran penting dalam regulasi autokrin respon inflamasi
dan kekebalan tubuh.10
Ada bukti yang berkembang bahwa kadar prokineticins yang meningkat di
lokasi peradangan dan terlibat dalam mediasi inflamasi nyeri. Dalam model
hewan PROK1 telah ditunjukkan mengikat tulang sel sumsum mononuklear
dan meningkatkan pembentukan populasi sel adeheren. Sel-sel ini menunjukkan
fenotipe makrofag seperti PROK1 yang menginduksi diferensiasi progenitor sel
makrofag. Efek PROK1 ini juga telah dikonfirmasi pada manusia di mana
stimulasi sumsum tulang CD34 + dengan PROK1 secara dramatis mempercepat
status pematangan dan mengakibatkan pengembangan populasi dari monosit /
makrofag. Selanjutnya, monosit yang dirangsang PROK1 mengekspresikan
tingkat TNF yang lebih tinggi dengan menurunkan secara simultan anti-
inflamasi IL10, hal ini menunjukkan bahwa PROK1 tidak hanya merangsang
diferensiasi monosit tetapi juga mengubah fungsi mereka dengan meningkatkan
pro-inflamasi.1,10
Efek pro-inflamasi PROK1 juga ditunjukkan oleh kemampuannya untuk
menginduksi ekspresi chemokin CCl4, CXCL1 dan CXCL8 pada monosit
manusia. Hal ini menunjukkan bahwa PROK1 mungkin merupakan mekanisme
perekrutan leukosit yang signifikan dan menginvasi jaringan yang diamati
dalam peradangan. PROK2 dan ortholog amfibi Bv8 telah terbukti penting
dalam memediasi haematopoiesis dan respon imun bawaan. 10,11
Efek pro-inflamasi PROK2 telah dibuktikan pada makrofag tikus dimana
PROK2 meningkatkan kemotaksis dan mempromosikan produksi IL1b dan
TNF dengan penurunan paralel sekresi anti-inflamasi IL10. Secara signifikan,
semua efek dimediasi melalui PROKR1 dan efek ini tidak ditemukan dalam
prokr12/2 tikus. Peran kunci dari PROKR1 dalam memediasi respon inflamasi
ditunjukkan dengan respon dalam produksi sitokin di splenocytes tikus. Studi

24
ini menunjukkan bahwa PROK2 via PROKR1 secara signifikan meng-
upregulasi pro-inflamasi IL1b berdampingan dengan downregulasi dari anti-
inflamasi IL4 dan IL10. Selain itu, neutrofil yang dirangsang dengan PROK2
memperoleh peningkatan potensi migrasi dan mobilisasi sel dari sumsum tulang
ke situs tumor pankreas tikus tergantung pada aktivasi PROK2.12
Secara kolektif penelitian ini menunjukkan bahwa prokineticins
mempromosikan peradangan dan ekspresi mereka dipengaruhi oleh faktor pro-
inflamasi, dan ini merupakan mekanisme penting dalam respon imun bawaan.
Hal ini menunjukkan bahwa mungkin prokineticins berfungsi sebagai faktor
penting yang terlibat dalam gangguan inflamasi. Saat ini telah muncul berbagai
laporan mengenai disregulasi prokineticin pada beberapa penyakit inflamasi,
misalnya PROK1 telah terbukti meningkat dalam jaringan endometriotik
ektopik, dan secara signifikan meningkat pada pasien pre-eklampsia. PROK2
adalah mediator neutrofil tergantung angiogenesis penting pada tahap awal
perkembangan neoplastik, dan PROK1 dan PROKR1 diinduksi dengan cepat
dalam model murine dengan kolitis.10,11
B. Peran prokinetisin dalam inisiasi persalinan
Ekspresi prokineticin telah dilaporkan di unit uteroplasenta aterm, ini
menunjukkan adanya fungsi prokineticins dalam induksi persalinan.
Peningkatan ekspresi PROK2 di miometrium dan serviks telah ditunjukkan
dengan analisis microarray. Dalam plasenta, ekspresi PROK1 dan PROKR1
terdeteksi di plasenta trimester ketiga. Dalam studi ini, PROKR1 ditemukan
dalam pembuluh darah dan makrofag plasenta, dan menunjukkan bahwa
PROK1 berpotensi mengatur kekebalan tubuh dan fungsi sel pembuluh darah
dan menanggapi inflamasi dalam jaringan kehamilan. Dalam jaringan plasenta,
PROK1 telah ditunjukkan meningkatkan ekspresi mediator inflamasi seperti
COX2 dan IL8. COX2 diturunkan prostaglandin, bersama dengan chemokin
seperti IL8, bertindak untuk mengaktifkan sel-sel kekebalan tubuh
25
permeabilitas, vaskular dan infiltrasi sel inflamasi selama persalinan.
Prostaglandin juga terlibat dalam pematangan serviks dan kontraksi uterus,
sehingga tingkat COX2yang lebih tinggi merupakan penanda penting dari
persalinan yang sedang berlangsung. 1,10,12
Oleh karena itu, PROK1 dan PROKR1 merupakan jalur inisiasi untuk
respon inflamasi dalam plasenta trimester ketiga mengarah ke terjadinya partus.
Pro-inflamasi yang serupa dengan yang diamati dalam plasenta juga dilaporkan
dalam miometrium dan serviks selama persalinan dimana terdeteksi
peningkatan kadar prokineticins.1,13
Gambar 2. Mekanisme kerja prokinetisins dan reseptor nya dalam persalinan
sebagai respon terhadap infeksi bakteri
Dikutip dari Denison dkk, 20101
Jalur inflamasi Prokineticin dapat bertindak untuk meningkatkan chemotaxis
sel imun polimorfonuklear seperti neutrofil melalui induksi ekspresi chemokin
seperti IL8 dan CXCL1. Neutrofil juga mengekspresikan reseptor prokineticins
yang ketika dirangsang meningkatkan potensi migrasi mereka. Prokineticins

26
juga berkontribusi dalam menstimulasi sitokin seperti IL1b oleh neutrofil, yang
diakui sebagai mediator penting dari inflamasi dalam proses persalinan. Selain
itu, peningkatan ekspresi MMPs dalam miometrium selama persalinan dan
prokineticins mungkin memainkan peran penting dalam induksi ekspresi MMP
dalam miometrium melalui fosforilasi reseptor faktor pertumbuhan epidermal
yang merupakan mekanisme intraseluler yang dikenal untuk mengatur ekspresi
MMP.1,13
Prokineticins awalnya dikenal karena memiliki efek kuat terhadap
kontraktilitas otot polos seperti pada saluran pencernaan. Efek kontraktil
dimediasi melalui induksi masuknya kalsium dalam sel otot polos. Oleh karena
itu, prokineticins juga dapat berkontribusi ke awal persalinan dengan
menginduksi kontraktilitas otot halus miometrium. Prokineticins dapat
menginduksi kontraktilitas miometrium langsung melalui PROKR1 seperti
yang ditemukan dalam miometrium kehamilan aterm.1 Prostaglandin juga
diakui sebagai regulator kunci dari kontraktilitas miometrium persalinan.
Prokineticins juga dapat berkontribusi untuk memicu kontraktilitas miometrium
secara tidak langsung melalui induksi mediator inflamasi seperti ekspresi
COX2 dan pelepasan prostaglandin.1,10,13
V. RINGKAAN
Konsep persalinan tradisional dan bukti pada hewan, menunjukkan bahwa
persalinan adalah peristiwa hormonal, namun beberapa tahun terakhir, beberapa
peneliti telah mengusulkan bahwa peristiwa-peristiwa utama persalinan manusia
adalah proses inflamasi, dengan perubahan hormonal yang bersifat sekunder atau
pelengkap.
Ekspresi prokineticin telah dilaporkan di unit uteroplasenta aterm, ini
menunjukkan adanya fungsi prokineticins dalam induksi persalinan Peningkatan
ekspresi PROK2 di miometrium dan serviks telah ditunjukkan dengan analisis
microarray. Dalam plasenta, ekspresi PROK1 dan PROKR1 terdeteksi di plasenta

27
trimester ketiga. PROKR1 ditemukan dalam pembuluh darah dan makrofag
plasenta, dan menunjukkan bahwa PROK1 berpotensi mengatur kekebalan tubuh
dan fungsi sel pembuluh darah dan menanggapi inflamasi dalam jaringan
kehamilan. Dalam jaringan plasenta, PROK1 telah ditunjukkan meningkatkan
ekspresi mediator inflamasi seperti COX2 dan IL8. COX2 diturunkan
prostaglandin, bersama dengan chemokin seperti IL8, bertindak untuk
mengaktifkan sel-sel kekebalan tubuh permeabilitas, vaskular dan infiltrasi sel
inflamasi selama persalinan. Prostaglandin juga terlibat dalam pematangan serviks
dan kontraksi uterus, sehingga tingkat COX2yang lebih tinggi merupakan penanda
penting dari persalinan yang sedang berlangsung.
Jalur inflamasi Prokineticin dapat bertindak untuk meningkatkan chemotaxis
sel imun polimorfonuklear seperti neutrofil melalui induksi ekspresi chemokin
seperti IL8 dan CXCL1. Neutrofil juga mengekspresikan reseptor prokineticins
yang ketika dirangsang meningkatkan potensi migrasi mereka. Prokineticins juga
berkontribusi dalam menstimulasi sitokin seperti IL1b oleh neutrofil, yang diakui
sebagai mediator penting dari inflamasi dalam proses persalinan. Selain itu,
peningkatan ekspresi MMPs dalam miometrium selama persalinan dan
prokineticins mungkin memainkan peran penting dalam induksi ekspresi MMP
dalam miometrium melalui fosforilasi reseptor faktor pertumbuhan epidermal yang
merupakan mekanisme intraseluler yang dikenal untuk mengatur ekspresi MMP.
VI. RUJUKAN
1. Catalano, Lannagan, Gorowiec, Denison, Norman, and Jabbour1.Prokineticins: novel mediators of inflammatory and contractile pathways at parturition? Molecular Human Reproduction, 2010; 16 : 311–319,
2. Schwartz, LB. Understanding human parturition. The Lancet, 1997; 350:1792-3
3. Mitchell, MD. The initiation of parturition. Current Obstetrics and Gynecology, 1994; 4: 74-78

28
4. Lockwood, CJ. The initiation of parturition at term. Obstet Gynecol Clin N Am. 2004; 31: 935– 947
5. Pearson, RD. Immunogenicity, parturition and the prostaglandins. Medical Hypotheses, 1979; 5: 1297-1303
6. Golightlya, Jabbourb, Normana. Endocrine immune interactions in human parturition. Molecular and Cellular Endocrinology, 2011; 335: 52–59
7. Negri, Lattanzi, Giannini, Melchiorri. Bv8/Prokineticin proteins and their receptors. Life Sciences, 2007;8: 1103–1116
8. Ngan, Tam. Prokineticin-signaling pathway. The International Journal of Biochemistry & Cell Biology, 2008;40: 1679–1684
9. Ralbovsky et al. Triazinediones as prokineticin 1 receptor antagonists. Part 1: SAR, synthesis and biological evaluation. Bioorganic & Medicinal Chemistry Letters, 2009; 19: 2661–2663
10. Hoffmann, Feige, Alfaidy. Placental Expression of EG-VEGF and its Receptors PKR1 (Prokineticin Receptor-1) and PKR2 Throughout Mouse Gestation. Placenta, 2007; 28: 1049-58
11. Balasubramanian, et al. The puzzles of the prokineticin 2 pathway in human reproduction. Molecular and Cellular Endocrinology, 2011; 346: 44–50
12. Evans et al. Prokineticin 1 Signaling and Gene Regulation in Early Human Pregnancy Endocrinology, 2008; 149: 2877–2887.
13. Fiona et al. Prokineticin-1: A Novel Mediator of the Inflammatory Response in Third-Trimester Human Placenta. Endocrinology; 149: 3470–3477