perbanyakan vegetatif kelapa_sawit_unggul_secara_kultur_jaringan-andal_yakinudin-g84090018-ipb-2012
TRANSCRIPT

i
PERBANYAKAN VEGETATIF KELAPA SAWIT UNGGUL
SECARA KULTUR JARINGAN
Laporan Praktik Lapangan
di Lab Clonal Oil Palm Production Unit
PT Tunggal Yunus Estate, Asian Agri
Desa Pangkalan Kerinci, Kec. Kerinci, Kab. Pelalawan, Prov. Riau
ANDAL YAKINUDIN
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
BOGOR
2012

ii
PERBANYAKAN VEGETATIF KELAPA SAWIT UNGGUL
SECARA KULTUR JARINGAN
ANDAL YAKINUDIN
Laporan Praktik Lapang
Sebagai Salah Satu Syarat Memperoleh Gelar
Sarjana Sains pada
Departemen Biokimia
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
BOGOR
2012

iii
Judul : Perbanyakan Vegetatif Kelapa Sawit Unggul Secara Kultur Jaringan
Nama : Andal Yakinudin
NIM : G84090018
Disetujui,
Komisi Pembimbing
Drs. Djarot Sasongko, M.S Ir. Ida S. Febriantine, M.Si
Pembimbing Utama Pembimbing Lapang
Diketahui,
Ketua Departemen Biokimia
Dr. I Made Artika, M. App. Sc
NIP 19630117 198903 1 001
Tanggal Lulus:

iv
PRAKATA
Puji dan syukur penulis panjatkan kehadirat Allah SWT atas rahmat dan
karunia-Nya sehingga penulis dapat menyelesaikan laporaan kegiatan praktik
lapang yang berjudul “Kultur Jaringan: Perbanyakan Vegetatif Kelapa Sawit Sawit
Unggul di PT Tunggal Yunus Estate, Asian Agri”.
Ucapan terima kasih penulis sampaikan kepada Ir. Ida S. Febriantine,
M.Si. sebagai pembimbing lapang sekaligus Manager COPPU, Drs. Djarot
Sasongko, M.S. selaku pembimbing utama, dan Pak Ang Boon Beng sebagai
General Manager PT Tunggal Yunus Estate. Ucapan terima kasih juga penulis
sampaikan kepada Pak Gogoh, Pak Ari, Pak Erwin, Pak Beni, dan Bu Rike, selaku
pengawas lapang selama penulis melaksanakan praktik lapang di COPPU. Terima
kasih penulis ucapkan juga kepada Pak Abdur Rahman, Pak Rizki, Pak Sucipto,
Pak Izharul, Pak Ridwan, Pak Kukuh, dan Pak Harkingto, yang telah banyak
membantu dan memberikan pengarahan kepada penulis selama menimba ilmu di
OPRS Topaz.
Tidak lupa juga penulis mengucapkan terima kasih kepada Bu Vika
sebagai Manager Tanoto Foundation, Pak Indra Setiawan, dan seluruh pegawai
lainnya yang menjadi teman penulis selama melaksanakan praktik lapang.
Terimakasih juga penulis ucapkan kepada seluruh pegawai AAALI yang telah
membantu akomodasi penulis selama praktik lapang. Penulis juga mengucapkan
terimakasih kepada Waliyuddin, Munjiati, dan Ester, sebagai teman seperjuangan
selama praktik lapang. Ucapan terimakasih yang terdalam penulis sampaikan
kepada kedua orang tua yang tidak pernah lelah mendoakan dan memberikan
semangat selama penulis melaksanakan praktik lapang.
Akhir kata, penulis berharap laporan ini dapat bermanfaat bagi pihak-
pihak yang membutuhkan demi kemajuan bangsa Indonesia.
Pelalawan, Agustus 2012
Andal Yakinudin

v
DAFTAR ISI
Halaman
PRAKATA………………………………………………………………………...iv
DAFTAR ISI……………………………………………………………………… v
DAFTAR TABEL…………………………………………………………………vi
DAFTAR GAMBAR…………………………………………………………….. vi
DAFTAR LAMPIRAN…………………………………………………………... vi
PENDAHULUAN…………………………………………………………………1
KEADAAN UMUM PT TUNGGAL YUNUS ESTATE………………………….4
Sejarah dan Perkembangan Perusahaan………………………………………... 4
Lokasi dan Tata Letak Perusahaan……………………………………………... 6
Struktur Organisasi Perusahaan………………………………………………… 6
Ketenagakerjaan………………………………………………………………... 7
Jenis Produk……………………………………………………………………..8
TINJAUAN PUSTAKA…………………………………………………………. 11
Kelapa Sawit…………………………………………………………………... 11
Kultur Jaringan………………………………………………………………... 13
Metabolit Sekunder Fenolik…………………………………………………... 18
BAHAN DAN METODE……………………………………………………….. 22
Alat dan Bahan………………………………………………………………... 22
Metode Penelitian……………………………………………………………... 22
HASIL DAN PEMBAHASAN………………………………………………….. 25
Hasil Inokulasi Eksplan dan Kalus……………………………………………. 25
Hasil Transfer EC Rendah dan EC Tinggi……………………………………..29
Hasil Transfer SD-RI………………………………………………………….. 34
Aklimatisasi Ramet…………………………………………………………… 35
Kontaminasi……………………………………………………………………36
SIMPULAN DAN SARAN……………………………………………………... 39
Simpulan………………………………………………………………………. 39
Saran…………………………………………………………………………... 39
DAFTAR PUSTAKA……………………………………………………………. 40

vi
DAFTAR TABEL
Halaman
“Balance sheet” dari ion-ion pada media-media……………………………….. 15
Beberapa fungsi fenilpropanoid………………………………………………… 18
DAFTAR GAMBAR
Logo Asian Agri.................................................................................................... 4
Garis besar produk metabolisme senyawa fenilpropanoid.................................... 19
Reaksi deaminasi fenilalanina oleh PAL.............................................................. 20
Sintesis asam hidroksisinamat dari asam trans-sinamat………………………... 21
Hasil inokulasi umbut kelapa sawit……………………………………………... 26
Kultur kalus pada kultur jaringan kelapa sawit..................................................... 27
Kultur EC rendah pada kultur jaringan kelapa sawit............................................ 30
Kultur EC tinggi pada kultur jaringan kelapa sawit…………………………….. 31
Kultur SD pada kultur jaringan kelapa sawit........................................................ 34
Kultur RI pada kultur jaringan kelapa sawit……………………………………. 34
Ramet hasil kultur jaringan kelapa sawit……………………………………….. 36
Kultur yang terkontaminasi……………………………………………………... 37
DAFTAR LAMPIRAN
Diagram alir kegiatan praktik lapangan……………………………………….... 44
Struktur Organisasi RGE Group............................................................................ 45
Tahapan kultur jaringan kelapa sawit di COPPU……………………………….. 46
Gambar berbagai tahapan kultur jaringan kelapa.................................................. 47
Komposisi media kultur jaringan tanaman............................................................ 49

PENDAHULUAN
Program Sarjana Biokimia merupakan salah satu program pendidikan yang
diselenggarakan oleh Departemen Biokimia, Fakultas Matematika dan Ilmu
Pengetahuan Alam, Institut Pertanian Bogor. Program ini memiliki tujuan untuk
menghasilkan lulusan tingkat sarjana yang memahami kaidah ilmiah, landasan
keilmuan yang kuat, mampu berpikir secara logis dan sistematis, terampil, serta
kreatif dalam menemukan solusi bagi masalah kehidupan yang berkaitan dengan
biokimia untuk kesejahteraan umat manusia.
Praktik lapang berguna untuk melatih sekaligus menyelaraskan ilmu yang
telah diperoleh di perkuliahan dengan dunia kerja agar bisa menjadi bekal saat
memasuki dunia kerja nantinya. Melalui kegiatan praktik lapangan (PL),
mahasiswa dituntut untuk dapat mengembangkan aspek kognitif, afektif dan
psikomotorik melalui penerapan ilmu yang diiringi dengan latihan kerja pada
instansi atau industri terkait, serta pengamatan metode-metode yang diterapkan di
lapangan sesuai dengan bidang keahlian. Selain itu, mahasiswa juga diharapkan
dapat melatih sikap kooperatif dan interaktif, agar mampu menjadi tenaga yang
terampil dan professional dalam dunia kerja.
Indonesia merupakan negara yang sangat potensial untuk pengembangan
usaha perkebunan kelapa sawit. Saat ini, kelapa sawit sudah secara luas ditanam
di Pulau Sumatra, Kalimantan, Jawa, Sulawesi, Papua, dan pulau-pulau lainnya.
Potensi lahan untuk penanaman baru kebun kelapa sawit masih cukup luas.
Lahan-lahan tersebut terutama berada di Kalimantan dan Papua. Perkebunan
kelapa sawit membawa dampak yang besar terhadap kemajuan perekonomian
masyarakat Indonesia, terutama pada daerah perkebunan dan pengolahan hasil
sawit. Oleh karena sumbangan yang besar dalam perekonomian Indonesia,
ekspansi dan intensifikasi lahan sawit di Indonesia terus dilakukan hingga saat ini.
Bibit unggul kelapa sawit dan teknologi-teknologi pendukung diperlukan
untuk mendukung ekspansi dan intensifikasi kebun kelapa sawit. Bibit unggul
diperlukan dalam jumlah besar bisa diperoleh dari berbagai macam teknik. Teknik
pemuliaan tanaman dapat menghasilkan bibit unggul dengan cara menyeleksi
indukan kelapa sawit unggul dan pembuahan dilakukan pada bibit sawit unggul
pula. Hal ini diharapkan bisa menghasilkan anakan kelapa sawit yang unggul

2
seperti induknya. Cara perbanyakan kelapa sawit dengan perkawinan ini masih
memiliki kelemahan, yaitu fenotip anak yang dihasilkan tidak bisa dijamin persis
keunggulannya dan menghasilkan banyak variasi fenotip dalam tiap perkawinan.
Hal ini karena pencampuran gen akan membuat banyak sekali variasi pada anakan
dan variasi-variasi yang terjadi belum tentu merupakan sifat yang diinginkan.
Asian Agri melalui PT Tunggal Yunus Estate melakukan perbanyakan
kelapa sawit unggul menggunakan teknik vegetatif, yaitu melalui kultur jaringan,
untuk mengatasi kelemahan reproduksi generatif kelapa sawit dalam penyediaan
benih unggul. Indukan kelapa sawit dengan sifat-sifat yang luar biasa unggul
diperbanyak dan ditanam kembali di lapangan untuk melihat hasilnya. Selain
digunakan untuk perbanyakan kelapa sawit untuk ditanam di lahan produksi, PT
Tunggal Yunus Estate melalui COPPU juga melakukan perbanyakan pohon-pohon
induk yang digunakan dalam produksi benih unggul. Beberapa pohon-pohon
induk Dura dan Pisifera yang digunakan memiliki sifat-sifat yang sangat unggul
tetapi jumlahnya terbatas. Untuk mengatasi hal ini, kultur jaringan merupakan
salah satu solusinya.
Perbanyakan kelapa sawit unggul dengan teknik kultur jaringan ini
diharapkan mampu memenuhi kebutuhan bibit dalam usaha Asian Agri untuk
meningkatkan produktivitas kebun. Hal ini perlu dilakukan karena areal untuk
membuka perkebunan baru semakin sulit untuk didapat. Kultur jaringan juga
digunakan untuk perbanyakan pohon-pohon induk unggul sebagai sumber benih
Tenera. Selain itu, teknik kultur jaringan ini bisa menyediakan perbanyakan
pohon-pohon induk untuk penyediaan bibit unggul kelapa sawit secara generatif.
Hasil penanaman klon kelapa sawit unggul ini diharapkan juga bisa meningkatkan
produktivitas kebun karena klon tersebut merupakan perbanyakan dari kelapa
sawit yang sudah terbukti sangat ungul di lapangan.
Salah satu tantangan dalam kultur jaringan kelapa sawit adalah proses
kultur jaringan yang efisien (Kushairi et al 2010). Saat ini, PT Tunggal Yunus
Estate melalui COPPU sedang mengembangkan proses tersebut. Salah satu
kendala dalam kultur jaringan di COPPU, walaupun dalam jumlah kasus terbatas,
adalah masalah pencoklatan (browning) pada saat proses subkultur. Pencoklatan
dapat menyebabkan kematian jaringan saat proses subkultur dan peningkatan

3
kejadian abnormalitas pada tanaman yang dihasilkan saat ditumbuhkan di
lapangan. Hal ini sangat mempengaruhi tingkat multiplikasi tanaman dan kualitas
tanaman yang dihasilkan, sehingga berpengaruh langsung pada efisiensi proses
kultur jaringan tersebut.
Kegiatan praktik lapang dilaksanakan selama bulan Juli dan Agustus 2012
di Clonal Oil Palm Production Unit (COPPU), PT Tunggal Yunus Estate, Asian
Agri. Lokasi laboratorium berada di Kebun Buatan, Eko 2, Desa Pangkalan
Kerinci, Kecamatan Kerinci, Kabupaten Pelalawan, Provinsi Riau. Beberapa
bagian kegiatan praktik lapang juga dilaksanakan di OPRS Topaz, Desa
Petapahan, Kecamatan Tapung, Kabupaten Kampar, Riau. Kegiatan di OPRS
Topaz dilaksanakan selama satu minggu.
Pelaksanaan praktik lapang di COPPU bertujuan untuk melakukan praktik
transfer embryoid culture tinggi (EC tinggi), transfer shoot development-root
induction (SD-RI), transfer root induction (RI), preparasi media, cek kontaminasi
subkultur, skoring (new embryoid (Ne) & EC tinggi), seleksi & input ramet, dan
seleksi bad culture. Adapun bagian kegiatan praktik lapang yang dilakukan di
OPRS Topaz adalah penanganan ramet (hardening, pre nursery, main nursery,
maintenance & selection), overview Seed Production Unit (SPU), sampling bunch
di lapangan, oil extraction in Lab Bunch Analysis, pisifera & pollen processing,
bagging & pollination, dan overview breeding & selection untuk pemilihan ortet.
Kegiatan praktik lapang ini dilakukan dengan metode observasi dan praktik
terhadap alur kerja (metode) kultur jaringan kelapa sawit di COPPU. Tujuan
khusus dari praktik lapang ini adalah mempelajari dan memahami proses-proses
kultur jaringan kelapa sawit, mulai dari pengambilan eksplan di lapangan hingga
menghasilkan benih kelapa sawit yang siap tanam.

KEADAAN UMUM PT TUNGGAL YUNUS ESTATE
Sejarah dan Perkembangan Perusahaan
PT Tunggal Yunus Estate merupakan pengelola dari Oil Palm Research
Station di kebun Topaz (OPRS Topaz). Selain OPRS Topaz, Asian Agri juga
memiliki pusat riset di Bahilang (R&D Bahilang). OPRS Topaz menginduk pada
R&D Bahilang. OPRS Topaz mulai dirintis sejak tahun 1992 dengan seleksi dan
persilangan pohon induk di Kosta Rika. Selanjutnya, pohon induk yang dihasilkan
ditanam di Kebun Topaz pada tahun 1996.
Sebagai produsen benih kelapa sawit, OPRS Topaz bertujuan untuk
menghasilkan bahan tanaman kelapa sawit dengan dengan produktifitas minyak
yang tinggi dan mempunyai keunggulan sekunder yang diperoleh melalui aktifitas
pemuliaan yang sistematis dan berkelanjutan. Untuk mendukung riset dalam
pemuliaan dan produksi bahan tanaman kelapa sawit ini, PT Tunggal Yunus Estate
mendirikan Clonal Oil Palm Production Unit (COPPU) di Kebun Buatan Eko 2,
di Pangkalan Kerinci.
COPPU adalah laboratorium riset dan produksi skala besar bibit sawit
unggul yang didapat dengan teknik kultur jaringan. Lab kultur jaringan ini
berlokasi di Kebun Buatan Eko 2. Laboratorium ini mulai dioperasikan tahun
2005 dengan nama awal Tissue Culture Laboratory (TC Lab) yang tergabung
dalam Royal Golden Eagle Technology Centre (RGE-TC). RGE-TC merupakan
semua tempat riset untuk RGE Group. RGE-TC merupakan kolaborasi antara
RAPP Fiber-Mill-AA. Ada empat lab di RGE-TC; Soil Analitycal Lab, Pest &
Disease Lab, Pulp & Paper Lab, Tissue Culture Lab. Logo Asian Agri disajikan
pada Gambar 1.
Gambar 1 Logo Asian Agri.

5
Pada tanggal 23 Maret 2012, gedung laboratorium dipindahkan ke lokasi
yang sekarang dan namanya berubah menjadi nama yang digunakan hingga
sekarang. Unit laboratorium ini dikepalai oleh seorang manager dan dan dibantu
oleh lima orang asisten. Para asisten dan manager mengatur karyawan lainnya di
COPPU yang bertugas di bagian preparasi media, sub kultur, gudang,
administrasi-personalia, dan bagian pemrosesan data.
COPPU merupakan salah satu divisi dari PT Tunggal Yunus Estate (PT
TYE) yang bertugas dalam riset kelapa sawit unggul dan perbanyakannya dengan
teknik kultur jaringan. Kantor pusat PT TYE berada di kebun Topaz, Desa
Petapahan, Kecamatan Tapung, Kabupaten Kampar, Riau. PT TYE sendiri
mempunyai 4 divisi dan 1 unit pelaksana marketing. Keempat divisi tersebut
adalah Seed Production Unit (SPU), Nursery and Extention, Breeding, dan
COPPU. SPU, Nursery and Extention, dan Breeding berada di kebun Topaz dan
hanya COPPU yang berlokasi di Kebun Buatan, Kerinci.
PT TYE merupakan bagian dari grup perusahaan Asian Agri dan bergerak
dibidang produksi bibit kelapa sawit. Produksi benih kelapa sawit yang dijual oleh
PT TYE ke masyarakat maupun perusahaan lain masih berupa kecambah dan bibit
kelapa sawit dari hasil persilangan varietas Dura dan Pisifera unggul. Saat ini,
produksi klon kelapa sawit di COPPU masih terbatas. Hal ini bukan karena
kapasitas laboratorium yang terbatas, namun karena masih dalam tahap percobaan
untuk melakukan uji potensi produksi serta insiden abnormalitas dalam tanaman
klon tersebut. Setelah dipastikan bahwa klon tersebut memiliki tingkat produksi
TBS dan minyak yang tinggi, tidak ada abnormalitas, serta tidak ada perbedaan
dengan induknya, maka akan dilakukan kloning ulang atau recloning. Proses
recloning tentu akan menjadi mudah dan dapat dijalankan dengan skala produksi
yang lebih besar dengan hasil yang lebih baik.
PT TYE melalui COPPU saat ini terus melakukan riset kultur jaringan
kelapa sawit untuk mendukung produksi benih unggul kelapa sawit skala besar.
Riset kultur jaringan ini juga dibarengi dengan program pemuliaan kelapa sawit
dan didukung oleh para pakar yang telah berpengalaman di bidang penelitian
kelapa sawit baik dari dalam maupun luar negeri. Perpaduan riset dalam teknik
kultur jaringan dan pemuliaan tanaman akan menghasilkan keselarasan dalam

6
mencapai tujuan PT TYE, yaitu menghasilkan bahan tanaman kelapa sawit
dengan produktivitas minyak yang tinggi dan mempunyai keunggulan sekunder
yang diperoleh melalui aktifitas pemuliaan yang sistematis dan berkelanjutan.
Lokasi dan Tata Letak Perusahaan
Kantor pusat Asian Agri di Indonesia terletak di Uniplaza Building lantai 6
Menara Timur, Jl. Letjend MT Haryono No. A-1 Medan 20231, Sumatra Utara.
PT Tunggal Yunus Estate memiliki kantor pusat di OPRS Topaz, Desa Petapahan,
Kecamatan Tapung, Kabupaten Kampar, Provinsi Riau. PT Tunggal Yunus Estate
memiliki perkebunan kelapa sawit seluas 4000 ha dan digunakan untuk produksi
dan penelitian benih unggul kelapa sawit. OPRS Topaz memliki jarak sekitar 80
Km dari Kota Pekanbaru dan dapat ditempuh dengan mobil dalam waktu sekitar 2
jam. PT Tunggal Yunus Estate juga memiliki Marketing Office di Jl. Jend.
Sudirman No. 68, Pekanbaru, Riau.
Struktur Organisasi Perusahaan
PT Tunggal Yunus Estate termasuk dalam kelompok usaha Asian Agri.
Asian Agri sendiri merupakan bagian dari Royal Golden Eagle Group (RGE
Group). RGE Group memiliki lima bidang kelompok usaha. Kelima kelompok
usaha tersebut dibagi bardasarkan bidang perusahaannya; Pacific Oil &
Gas/PO&G (bidang minyak bumi dan gas), Sateri (bidang produksi rayon),
PechTec (bidang konstruksi), Asian Agri/AA (bidang perkebunan), dan Riau
Andalan Pulp and Paper/RAPP (bidang kehutanan, produksi bubur kertas dan
kertas Paper One). Asian Agri memliki 12 perusahaan yang bergerak di bidang
usaha tani kelapa sawit. PT Tunggal Yunus Estate (PT TYE) sendiri secara
spesifik bergerak dalam usaha penyediaan bibit unggul kelapa sawit.
PT TYE dipimpin oleh seorang general manager bernama Bapak Ang
Boon Beng. PT TYE sendiri terdiri dari empat divisi dan satu unit pelaksana kerja.
Divisi tersebut adalah SPU, Nursery & Extention, Breeding, dan COPPU. Satu
unit kerja di PT TYE adalah Marketing. Tiap divisi/unit kerja dipimpin oleh
seorang manager; SPU oleh Bapak Tan Kim Seng, Nursery & Extention oleh
Bapak Juliarto Barus, Breeding oleh Bapak Kukuh Setiawan, COPPU oleh Ibu Ida
Febriantine, dan Marketing oleh Bapak Ronny Susilo.

7
Ibu Ida Febriantine sendiri dalam memimpin COPPU dibantu oleh lima
orang yang bertugas sebagai supervisor. COPPU memiliki empat bagian dalam
melaksanakan fungsinya dan masing masing bagian ditangani secara terpisah oleh
para supervisor tersebut. Keempat bagian tersebut dan supervisor yang
menanganinya adalah: Media Preparation oleh Pak Beni Oktora, Subkultur oleh
Pak Gogoh Sulaksono dan Ibu Rike Novikasari, Eksplan dan Kalus oleh Pak
Nazhri Jambak, dan bagian Ramet serta Data Processing oleh Pak Erwin
Simanjuntak. Bagian pendukung yang berperan sangat penting pada kelancaran
pekerjaan adalah Administration, Personalia dan Common Service. Bagian
pendukung ini dikepalai oleh seorang KTU, Pak Indra Setiawan.
Ketenagakerjaan
Terdapat sistem tingkatan karyawan di Asian Agri. Para staff (General
Manager, Manager, dan Asisten) masuk dalam kelas eksekutif, para pemimpin
divisi atau unit kerja masuk dalam tingkatan senior manager atau manager. Para
pemimpin divisi atau unit kerja dibantu oleh para asisten di setiap unit kerja
masing-masing. Tingkatan asisten dimulai dari asisten satu hingga asisten empat.
Asisten empat disebut asisten kepala (head assistant). Mulai dari asisten hingga
tingkat yang lebih tinggi, para karyawan tersebut disebut sebagai staff. Adapun
para pekerja lainnya yang tingkatannya dibawah asisten, statusnya disebut non
staff.
Tingkatan pekerja non staff dibagi dalam tiga bagian, dimulai dari yang
paling atas yaitu SKU-B, SKU-H, dan Pekerja Harian Lepas (PHL). Dalam
tingkat SKU-B pun terdapat tingkatan lagi mulai dari tingkat A1, hingga G8.
Urutan tingkat SKU-B berurut mulai abjad A hingga G dan dari angka 1 hingga 8
(A1, A2, A3, …, A8, B1, B2, … dan seterusnya hingga G8). Para PHL merupakan
tingkatan pekerja yang paling bawah dan memiliki kuantitas terbesar. Para
karyawan non staff bisa diangkat ke jenjang yang lebih tinggi oleh HR Personalia
Region Office dengan persetujuan Regional Head berdasarkan masa kerja,
produktivitas, kedisiplinan, dan hal-hal lain yang dianggap penting. Evaluasi dan
pengajuan promosi disampaikan oleh asisten kepada manager.

8
Jenis Produk
Asian Agri melalui PT Tunggal Yunus Estate saat ini memproduksi benih
kelapa sawit persilangan varietas dura dan pisifera unggul (D×P) menggunakan
tetua dura dan pisifera terseleksi dari Kosta Rika. Tetua dura yang digunakan
berjumlah 228 keturunan inbred lines Dura Deli (D×D) yang berasal dari lembaga
riset ternama seperti MARDI Serdang (Malaysia), OPRS Banting (Malaysia),
OPRS Dami (Papua New Guinea), Stasiun Riset Chemara (Malaysia), Socfin
Johor Labis (Malaysia) dan San Alejo (Honduras). Tetua pisifera terseleksi
sejumlah 50 keturunan yang berasal dari AVROS (H & C, Malaysia), AVROS
Dami, Ekona, Ghana, Nigeria, La Me dan Yangabi. Pohon-pohon induk tersebut
ditanam di OPRS Topaz dan produksi serta pengolahan lanjutan benih kelapa
sawit juga dilakukan disana.
Berdasarkan hasil pengujian dan seleksi yang seksama, telah terpilih tetua
dura dan pisifera untuk memproduksi varietas unggul kelapa sawit D×P Topaz
yaitu Topaz 1 (hasil persilangan Dura Deli dengan Pisifera Nigeria), Topaz 2
(hasil persilangan Dura Deli dengan Pisifera Ghana), Topaz 3 (hasil persilangan
Dura Deli dengan Pisifera Ekona), Topaz 4 (hasil persilangan Dura Deli dengan
Pisifera Yangambi).
Varietas unggul kelapa sawit D×P Topaz telah memperoleh Ijin Pelepasan
Varietas sesuai Surat Keputusan Menteri Pertanian RI Nomor 57, 58,59, dan
60/KPTS/SR.120/I/2004 masing-masing tertanggal 16 Januari 2004. OPRS Topaz
telah memperoleh sertifikasi ISO 14001:2004 (SGS) dan ISO 9001:2000 (TÜV
NORD)
Varietas Topaz telah diuji menggunakan 440 progeni D×P yang merupakan
persilangan antara 223 Dura Deli dengan 50 pisifera dan ditanam pada 3 lokasi
dengan jenis tanah yang berbeda; tanah organik/aluvial, gambut dangkal, dan
gambut dalam. Luas total areal percobaan lebih dari 600 hektar di Sumatra Utara
dan Riau. Keunggulan D×P Topaz yaitu potensi hasil minyak yang tinggi,
produksi yang tinggi mulai panen pertama, rendemen minyak yang tinggi,
pertumbuhan meninggi yang lambat, dan mampu beradaptasi dengan baik pada
tanah gambut.

9
Kontrol pengumpulan tepung sari dari tetua pisifera dan penyerbukan pada
bunga betina dilakukan dengan ketelitian yang sangat tinggi. Proses selanjutnya
adalah menjaga kemurnian benih yang dihasilkan saat pemrosesan benih lebih
lanjut. Standard seleksi yang tinggi dan kontrol kualitas yang ketat memberikan
jaminan bahan tanaman yang dihasilkan berkualitas tinggi.
Saat ini, produksi ramet (benih dari kultur jaringan) kelapa sawit yang
dihasilkan di COPPU masih sekitar 28.000 ramet per tahun. Ramet ini belum
diproduksi untuk tujuan komersial karena masih dalam tahap penelitian serta
penggunaannya terbatas untuk kepentingan perusahaan saja. Jika hasil kultur
jaringan telah terbukti unggul dan tidak ada abnormalitas, baru tingkat produksi
secara komersial akan dilakukan.

11
TINJAUAN PUSTAKA
Kelapa Sawit
Kelapa sawit masuk ke dalam Divisi Tracheophyta, Subdivisi Pteropsida,
Kelas Angiospermae, Subkelas Monocotyledonae, Ordo Cocoidae, Famili Palmae,
dan Genus Elaeis. Terdapat tiga spesies pada kelapa sawit, yaitu Elaeis guineensis
Jacq., Elaeis oleifera (H.B.K) Cortes, Elaeis odora (Fauzi et al 2008).
E. guineensis diusahakan secara komersial di Afrika, Amerika Selatan,
Asia Tenggara, Pasifik Selatan, serta beberapa daerah lain dengan skala yang
lebih kecil. Tanaman kelapa sawit berasal dari Afrika dan Amerika Selatan
(Brazil). Di Brazil, tanaman ini dapat ditemukan tumbuh liar atau setengah
disepanjang tepi sungai. Kelapa sawit yang termasuk dalam Subfamili Cocoideae
merupakan tanaman asli Amerika Selatan, termasuk spesies E. oleifera dan E.
odora (Pahan 2006).
Kelapa sawit yang dibudidayakan terdiri dari dua jenis yaitu E.
guineensis dan E. oleifera. Jenis pertama yang terluas dibudidayakan orang.
Kedua spesies kelapa sawit ini memiliki keunggulan masing-masing. E.
guineensis memiliki produksi yang sangat tinggi dan E. oleifera memiliki
kandungan minyak tak jenuh tinggi serta tegakan tanaman yang rendah. Banyak
orang menyilangkan kedua spesies ini untuk mendapatkan spesies yang tinggi
produksinya dan mudah dipanen. E. oleifera sekarang mulai dibudidayakan pula
untuk menambah keanekaragaman sumber daya genetik (Pahan 2006).
Pemulia tanaman seringkali melihat tipe kelapa sawit berdasarkan
ketebalan cangkang, yang terdiri dari Dura, Pisifera, dan Tenera. Dura
merupakan sawit yang buahnya memiliki cangkang tebal sehingga dianggap
memperpendek umur mesin pengolah namun biasanya tandan buahnya besar-
besar dan kandungan minyak per tandannya berkisar 18%. Pisifera buahnya tidak
memiliki cangkang, sehingga tidak memiliki inti (kernel) yang menghasilkan
minyak ekonomis dan bunga betinanya steril sehingga sangat jarang
menghasilkan buah. Tenera adalah persilangan antara induk Dura dan jantan
Pisifera. Jenis ini dianggap bibit unggul sebab melengkapi kekurangan masing-
masing induk dengan sifat cangkang buah tipis namun bunga betinanya tetap
fertile (Fauzi et al 2008).

12
Tanaman kelapa sawit berakar serabut. Perakarannya sangat kuat karena
tumbuh ke bawah dan ke samping membentuk akar primer, sekunder, tersier, dan
kuarter (Fauzi et al 2008). Akar primer tumbuh ke bawah di dalam tanah sampai
batas permukaan air tanah. Akar sekunder, tertier, dan kuarter tumbuh sejajar
dengan permukaan air tanah bahkan akar tertier dan kuarter menuju ke lapisan
atas atau ke tempat yang banyak mengandung zat hara (Lubis 2008).
Batang kelapa sawit tumbuh tegak lurus (phototropi) dibungkus pelepah
daun (frond base) (Lubis 2008). Batang berbentuk silindris berdiameter 0,5 m
pada tanaman dewasa. Bagian bawah umumnya lebih besar disebut bongkol
batang atau bowl. Pada tanaman yang masih muda batangnya tidak terlihat karena
tertutup oleh pelepah daun. Daun kelapa sawit mirip kelapa yaitu membentuk
susunan daun majemuk, bersirip genap, dan bertulang sejajar. Daun-daun
membentuk satu pelepah yang panjangnya mencapai 7.5 - 9 m. Jumlah anak daun
di setiap pelepah berkisar antara 250 - 400 helai (Fauzi et al 2008).
Tanaman kelapa sawit di lapangan mulai berbunga pada umur 12–14
bulan, tetapi baru ekonomis untuk dipanen pada umur 2.5 tahun (Lubis 2008).
Dari setiap ketiak pelepah daun akan keluar satu tandan bunga jantan atau betina.
Diferensiasi seks terjadi 17–25 bulan sebelum antesis dan setelah antesis
membutuhkan waktu 5–6 bulan baru matang panen. Secara visual tandan bunga
jantan atau betina baru dapat diketahui setelah muncul dari ketiak pelepah daun
yaitu 7–8 bulan sebelum matang (Fauzi et al 2008).
Bunga betina setelah dibuahi akan berkembang pada spikelet. Karena
kondisi terjepit maka buah yang terletak di bagian dalam akan lebih kecil dan
kurang sempurna bentuknya dibandingkan dengan yang terletak di bagian luar
(Lubis 2008). Kematangan buah dibedakan atas matang morfologis dan matang
fisiologis. Matang morfologis adalah kematangan buah yang telah sempurna
bentuknya serta kandungan minyak optimal. Matang fisiologis adalah kematangan
buah yang sudah lebih lanjut yaitu telah siap untuk tumbuh dan berkembang
biasanya satu bulan sesudah matang morfologis (Pahan 2006).
Beberapa hal yang harus diperhatikan dalam memilih seleksi induk untuk
kultur jaringan yang baik adalah oil yield (>10 ton/ha), pohon harus sehat dan
tidak tinggi, pelepahnya rapi, serta tidak ada penyakit, dan batangnya tidak terlalu

13
besar. Bunch index, yaitu perhitungan rasio antara produk tandan dengan jumlah
hasil fotosintesis, menjadi tidak kalah pentingnya. Bunch index yang ideal adalah
>6 yang berarti 60% hasil fotosintesis digunakan untuk produksi tandan buah
segar (TBS) kelapa sawit. Kelapa sawit dengan bunch index yang tinggi lebih
efisien dalam produksi tandan dibanding kelapa sawit yang bunch index-nya
rendah. Dengan nilai indeks yang tinggi, biasanya bentuk pohon dan batangnya
akan lebih kecil, pelepahnya akan lebih pendek karena hasil fotosintesis lebih
banyak digunakan untuk produksi TBS (Lubis 2008).
Kultur Jaringan
Salah satu hal terpenting dalam siklus hidup organisme adalah fertilisasi
yang melibatkan penggabungan dari dua gamet yang berlawanan (berbeda tipe)
dan membentuk zigot. Zigot ini merupakan asal dari seluruh bagian multiselular
dan multiorgan dari organisme tingkat tinggi. Secara teoritis, semua sel di tubuh
tanaman seharusnya menerima material genetik yang sama dengan zigot. Adanya
fakta ini mengharuskan adanya beberapa faktor lain yang berdampak pada
karakter genetik sel yang memberikan variasi ekspresi genetik yang besar dari sel-
sel yang identik secara genetik. Proses yang terlibat untuk menghasilkan variasi-
variasi ini disebut diferensiasi (Bhojwani dan Razdan 1983).
Kultur jaringan tanaman merupakan teknik menumbuh-kembangkan
bagian tanaman, baik berupa sel, jaringan,atau organ dalam kondisi aseptik secara
in vitro. Teknik ini dicirikan oleh kondisi kultur yang aseptik, penggunaan media
kultur buatan dengan kandungan nutrisi lengkap dan ZPT (zat pengatur tumbuh),
serta kondisi ruang kultur yang suhu dan pencahayaannya terkontrol (Yusnita
2008). Berdasarkan bagian tanaman yang dikulturkan, secara lebih spesifik
terdapat beberapa tipe kultur, yaitu kultur kalus, kultur suspensi sel, kultur akar,
kultur pucuk tunas, kultur embrio, kultur ovul, kultur anter, dan kultur kuncup
bunga. Namun, semua jenis kultur tersebut sering disebut dalam istilah umum,
yaitu kultur jaringan (Yusnita 2008).
Praktik kultur jaringan tanaman bermula dari pembuktian sifat totipotensi
(total genetic potensial) sel, yaitu bahwa setiap sel tanaman yang hidup dilengkapi
dengan informasi genetik dan perangkat fisiologis yang lengkap untuk tumbuh
dan dan berkembang menjadi tanaman utuh, jika kondisinya sesuai. Teori ini

14
dikemukakan oleh Schwan dan Scheiden pada tahun 1838. Gotlieb Haberlant, ahli
botani Jerman, dianggap sebagai pelopor dalam sejarah perkembangan kultur
jaringan tanaman. Dalam publikasi tahun 1902, Haberlant mengemukakan bahwa
sel tumbuhan yang diisolasi dan dikondisikan dalam lingkungan yang sesuai akan
tumbuh dan berkembang menjadi tanaman yang lengkap. Namun, percobaan
untuk membuktikan idenya itu menemui kegagalan. Hal ini diduga karena
keterbatasan pengetahuan tentang hormon dan nutrisi tanaman pada waktu itu
(Yusnita 2008).
Totipotensi sel berhasil dibuktikan pada pertengahan sampai akhir tahun
1930-an. Setiap sel tumbuhan atau bagian kecil tanaman dapat tumbuh dan
berkembang menjadi individu tanaman baru yang lengkap. Penemuan ZPT dan
upaya pengembangan formulasi media berperan penting dalam menentukan
keberhasilan teknik kutur jaringan atau kultur in vitro secara umum (Bhojwani
dan Razdan 1983).
Kebutuhan nutrisional untuk pertumbuhan optimal dari jaringan dalam
tabung dapat bervariasi berdasarkan spesiesnya. Bahkan jaringan dari bagian
tanaman yang berbeda mungkin mempunyai kebutuhan yang berbeda untuk
pertumbuhan yang memuaskan (Murashige dan Skoog 1962). Sebagai contoh,
tidak ada medium tunggal yang bisa disarankan agar seluruhnya cocok untuk
semua tipe jaringan dan organ tanaman.
Beberapa media kultur jaringan paling awal, seperti medium kultur akar
dari White dan medium kultur kalus dari Gautheret, dikembangkan dari larutan
nutrien yang sebelumnya digunakan untuk kultur tanaman utuh. White
mengembangkan mediumnya dari Uspenski dan medium Uspenki untuk alga, dan
medium Gautheret berdasarkan larutan garam Knop. Semua formulasi media
selanjutnya berdasarkan pada media White dan Gautheret (Bhojwani dan Razdan
1983).
Beberapa jaringan tanaman seperti wortel, blackberry, dan sebagian besar
jaringan tumor, dapat tumbuh pada media yang sederhana yang hanya
mengandung garam anorganik dan gula sederhana. Untuk sebagaian besar
jaringan tanaman lainnya, penting untuk menambahkan pada mediumnya dengan
vitamin, asam amino, dan zat tumbuh dengan kombinasi kualitatif dan kuantitatif

15
yang berbeda-beda (Bhojwani dan Razdan 1983). Seringkali, campuran nutritif
kompleks ditambahkan pada media kultur jaringan tanaman. Medium yang hanya
mengandung bahan yang terdefinisi secara kimia disebut sebagai medium sintetik
(Bhojwani dan Razdan 1983). Bahkan dalam medium sintetik pun, penguraian
dari beberapa bahan (sukrosa, vitamin, dll) selama proses autoklaf dan interaksi
antara berbagai bahan selama preparasi media akan mengubah komposisi akhir
dari medium tersebut (Yusnita 2008).
Kandungan media pada kultur jaringan menurut Bhojwani dan Razdan
(1983) dibagi menjadi lima bagian; nutrien anorganik, nutrien organik, hormon
pertumbuhan, bahan pemadat (solidifying agent), dan tingkat keasaman
(konsentrasi ion H+ pada media, dinyatakan dengan pH).
Elemen mineral sangat penting dalam kehidupan tanaman. Mineral
berperan menjadi bagian dari pigmen tanaman, bagian dari dinding sel, bagian
dari stuktur biomolekul, dan lain-lain. Selain C, H, dan O, ada 12 elemen lain
yang dikenal sangat esensial untuk pertumbuhan tanaman. Elemen-elemen
tersebut adalah nitrogen, fosfor, sulfur, kalsium, kalium, magnesium, besi,
mangan, tembaga, seng, boron, dan molybdenum (Yusnita 2008). Enam elemen
pertama dari elemen-elemen tersebut, secara komparatif dibutuhkan dalam jumlah
besar, sehingga disebut elemen makro. Enam elemen lainnya secara komparatif
dibutuhkan dalam jumlah sedikit, sehingga disebut elemen-elemen mikro.
Berdasarkan rekomendasi dari dari Internasional Association for Plant
Physiology, elemen-elemen yang dibutuh oleh tanaman pada konsentrasi lebih
besar dari 0,5 mmol-1 mmol termasuk dalam makroelemen dan elemen-elemen
yang dibutuhkan pada konsentrasi kurang dari 0,5 mmol-1 mmol termasuk dalam
mikroelemen (De Fossard 1976). Kelima belas elemen yang penting untuk
pertumbuhan tanaman utuh juga terbukti penting untuk kultur jaringan.
Salah satu tipe ion mungkin bisa didapatkan lebih dari satu jenis garam
mineral. Perbandingan dua jenis media kultur jaringan yang berbeda dapat terlihat
dari konsentrasi total ion-ion yang berbeda dalam suatu medium. “Balance sheet”
dari ion-ion pada ketujuh media yang diberikan di Tabel 1.
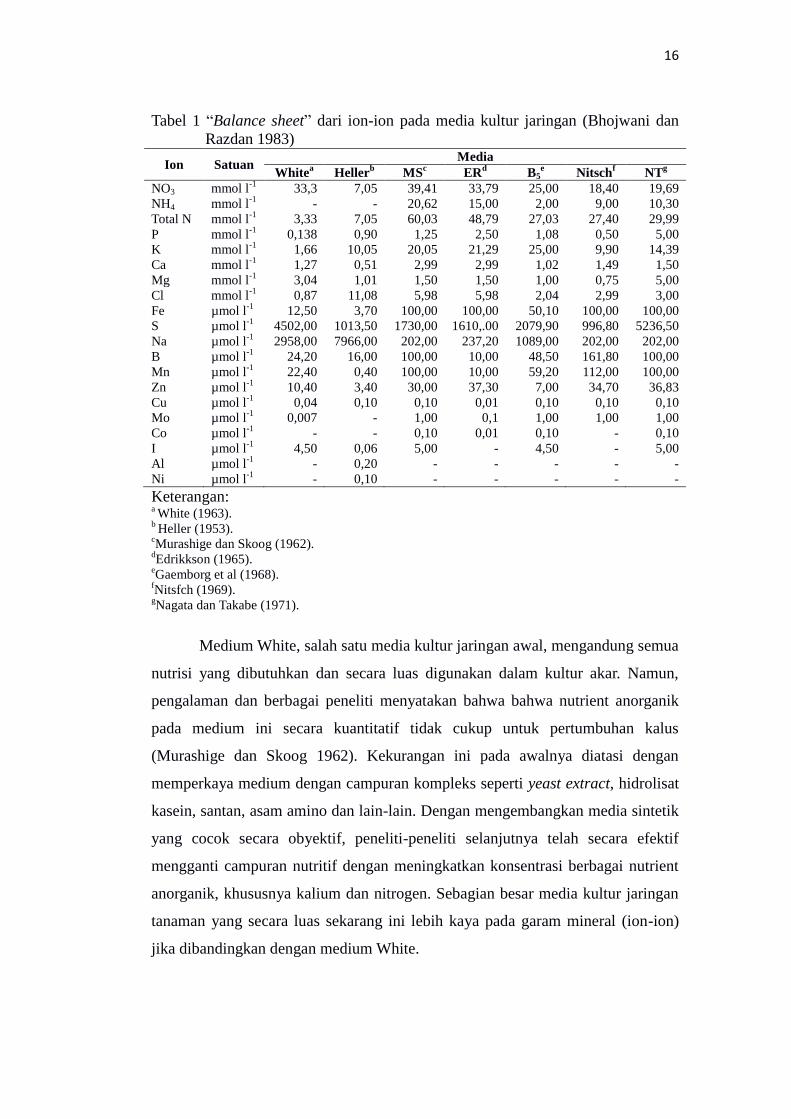
16
Tabel 1 “Balance sheet” dari ion-ion pada media kultur jaringan (Bhojwani dan
Razdan 1983)
Ion Satuan Media
Whitea
Hellerb
MSc
ERd
B5e
Nitschf
NTg
NO3 mmol l-1
33,3 7,05 39,41 33,79 25,00 18,40 19,69
NH4 mmol l-1
- - 20,62 15,00 2,00 9,00 10,30
Total N mmol l-1
3,33 7,05 60,03 48,79 27,03 27,40 29,99
P mmol l-1
0,138 0,90 1,25 2,50 1,08 0,50 5,00
K mmol l-1
1,66 10,05 20,05 21,29 25,00 9,90 14,39
Ca mmol l-1
1,27 0,51 2,99 2,99 1,02 1,49 1,50
Mg mmol l-1
3,04 1,01 1,50 1,50 1,00 0,75 5,00
Cl mmol l-1
0,87 11,08 5,98 5,98 2,04 2,99 3,00
Fe µmol l-1
12,50 3,70 100,00 100,00 50,10 100,00 100,00
S µmol l-1
4502,00 1013,50 1730,00 1610,.00 2079,90 996,80 5236,50
Na µmol l-1
2958,00 7966,00 202,00 237,20 1089,00 202,00 202,00
B µmol l-1
24,20 16,00 100,00 10,00 48,50 161,80 100,00
Mn µmol l-1
22,40 0,40 100,00 10,00 59,20 112,00 100,00
Zn µmol l-1
10,40 3,40 30,00 37,30 7,00 34,70 36,83
Cu µmol l-1
0,04 0,10 0,10 0,01 0,10 0,10 0,10
Mo µmol l-1
0,007 - 1,00 0,1 1,00 1,00 1,00
Co µmol l-1
- - 0,10 0,01 0,10 - 0,10
I µmol l-1
4,50 0,06 5,00 - 4,50 - 5,00
Al µmol l-1
- 0,20 - - - - -
Ni µmol l-1
- 0,10 - - - - -
Keterangan: a White (1963).
b Heller (1953).
cMurashige dan Skoog (1962).
dEdrikkson (1965).
eGaemborg et al (1968).
fNitsfch (1969).
gNagata dan Takabe (1971).
Medium White, salah satu media kultur jaringan awal, mengandung semua
nutrisi yang dibutuhkan dan secara luas digunakan dalam kultur akar. Namun,
pengalaman dan berbagai peneliti menyatakan bahwa bahwa nutrient anorganik
pada medium ini secara kuantitatif tidak cukup untuk pertumbuhan kalus
(Murashige dan Skoog 1962). Kekurangan ini pada awalnya diatasi dengan
memperkaya medium dengan campuran kompleks seperti yeast extract, hidrolisat
kasein, santan, asam amino dan lain-lain. Dengan mengembangkan media sintetik
yang cocok secara obyektif, peneliti-peneliti selanjutnya telah secara efektif
mengganti campuran nutritif dengan meningkatkan konsentrasi berbagai nutrient
anorganik, khususnya kalium dan nitrogen. Sebagian besar media kultur jaringan
tanaman yang secara luas sekarang ini lebih kaya pada garam mineral (ion-ion)
jika dibandingkan dengan medium White.

17
Menurut Heller (1965), gejala defisiensi dari beberapa elemen yang
ditunjukkan oleh jaringan kalus saat kekurangan nitrogen adalah beberapa
jaringan (Virginia creeper) menunjukkan kemunculan antosianin yang hebat;
bentuk tidak terjadi. Saat terjadi defisiensi nitrogen, kalium, dan fosfor terjadi
hipertrofi sel dan pengurangan jaringan kambium. Defisiensi sulfur
mengakibatkan jaringan mengalami klorosis. Kekurangan besi mengakibatkan
penghentian pembelahan sel. Saat defisiensi boron pada jaringan, sel mengalami
penghambatan pembelahan dan pemanjangan. Kekurangan mangan dan
molybdenum dapat mempengaruhi pemanjangan sel.
Sebagian besar kultur sel tanaman mampu membentuk semua vitamin
esensial, namun dalam jumlah yang kurang optimal. Untuk mencapai
pertumbuhan terbaik dari jaringan, seringkali penting untuk menambahkan
medium dengan vitamin dan asam amino. Thiamin (vitamin B1) secara umum
terbukti sebagai sebuah bahan yang penting. Vitamin lain, terutaman piridoksin
(vitamin B6), asam nikotinat (vitamin B3), kalsium pantotenat (vitamin B5), dan
inositol juga dikenal meningkatkan pertumbuhan bagian tanaman yang dikultur.
Berbagai media standard menunjukkan perbedaan yang luas pada komposisi
berbagai vitamin dan asam amino (Yusnita 2008).
Beberapa campuran nutritif kompleks seperti hidrolisat kasein (CH),
santan kelapa (CM), jus tomat (JC), corn milk, ekstrak malt (ME), dan yeast
extract (YE), juga telah digunakan untuk meningkatkan pertumbuhan beberapa
jenis kalus dan organ-organ. Penggunaan ekstrak alami sebisa mungkin dihindari
karena berpengaruh pada reproduksibilitas hasil kultur. Hal ini terjadi karena
kualitas dan kuantitas bahan perangsang pertumbuhan dalam ekstrak ini seringkali
bervariasi tergantung umur jaringannya dan variasi organisme donornya
(Bhojwani dan Razdan 1983).
Jaringan yang awalnya berwarna hijau secara perlahan akan kehilangan
pigmen hijaunya dalam kultur dan tergantung pada sumber karbon luar. Bahkan
jaringan yang mendapatkan pigmen hijau melalui perubahan yang tiba-tiba atau
dibawah kondisi khusus selama periode kultur, tidak menjadi bersifat autotrof
untuk karbon. Tegakan yang hijau dan berorgan lengkap dalam kultur juga

18
menunjukkan pertumbuhan dan proliferasi yang lebih baik dengan penambahan
sumber karbon yang cocok pada medium kultur (Yusnita 2008).
Sumber karbon yang paling sering digunakan adalah sukrosa, dengan
konsentrasi antara 2% hingga 5%. Glukosa dan sukrosa juga dikenal dapat
mendukukung pertumbuhan yang baik pada beberapa jaringan. Secara umum,
potongan akar tanaman dikotil paling baik tumbuh dengan sukrosa dan potongan
akar tanaman monokotil paling baik tumbuh dengan dekstrosa (glukosa).
Beberapa bentuk karbon dalam kultur jaringan tanaman yang dapat digunakan
adalah maltosa, galaktosa, mannosa, dan laktosa (Gautheret 1959). Beberapa
kultur tanaman bahkan bisa memetabolisme pati sebagai sumber karbon mereka.
Selain penambahan nutrient, secara umum juga penting untuk
menambahkan satu atau lebih bahan pengatur tumbuh, seperti auksin, sitokinin,
dan giberelin, untuk mendukung pertumbuhan jaringan dan organ yang baik.
Namun, kebutuhan bahan-bahan ini bervariasi tergantung pada jaringannya, dan
dipercaya bahwa hal ini juga tergantung dari kandungan endogen bahan-bahan
tersebut dalam jaringan (Bhojwani dan Razdan 1983).
Metabolit Sekunder Fenolik
Tanaman mengandung keragaman yang besar dalam senyawa turunan
fenolik. Contohnya adalah fenol sederhana, flavonoid, stilben, tanin, lignan, dan
lignin. Bersama dengan asam karboksilat rantai panjang, senyawa fenolik juga
merupakan komponen dari suberin dan kutin. Senyawa yang lebih bervariasi ini
memiliki fungsi yang penting sebagai antibiotik, pestisida alami, senyawa sinyal
untuk pembentukan simbiosis dengan Rhizobia, menarik polinator, agen protektif
terhadap sinar UV, bahan penyalut untuk membuat dinding sel impermeabel
terhadap udara dan air, dan bahan stuktural untuk memberikan stabilitas tanaman.
Semua senyawa ini diturunkan dari fenilalanina, dan dalam beberapa tanaman,
juga dari tirosina (Saxena 2001).
Fenilalanina dan tirosina dibentuk oleh jalur shikimate. Karena senyawa
fenolik diturunkan dari dua asam amino yang mengandung sebuah cincin fenil
dengan sebuah cincin samping C3, senyawa-senyawa ini secara kolektif disebut
fenilpropanoid (Heldt 2005). Garis besar produk metabolisme senyawa
fenilpropanoid disajikan pada gambar 2.

19
Gambar 2 Garis besar produk metabolisme senyawa fenilpropanoid (Heldt 2005).
Tabel 2 Beberapa fungsi fenilpropanoid (Heldt 2005) Fenilpropanoid Fungsi
Kumarin Antibiotik, racun bagi tanaman pengganggu
Lignan Antibiotik, racun bagi tanaman pengganggu
Lignin Bahan penyusun dinding sel
Suberin dan Kutin Pembentuk lapisan impermiabel
Stilben Antibiotik, khususnya fungisida
Flavonoid Antibiotik, sinyal untuk interaksi dengan simbion, pigmen pada bunga, senyawa pelindung terhadap cahaya berlebih
Tanin Bahan penyamak, fungisida, perlindungan terhadap herbivore
Senyawa-senyawa turunan asam benzoat, termasuk asam salisilat,
dibentuk oleh pemecahan dari fragmen C2 dari fenilpropana. Asam salisilat dapat
mempengaruhi tumbuhan. Telah diamati bahwa tanaman tembakau yang diberi
perlakuan aspirin (asetil salisilat) atau asam salisilat dapat meningkatkan
ketahanan terhadap patogen, seperti Tobacco mosaic virus (TMV) (Saxena 2001).
Beberapa fungsi fenilpropanoid juga disajikan pada tabel 2.
Banyak tumbuhan menunjukkan peningkatan kandungan asam salisilat
setelah diinfeksi oleh virus atau jamur. Peningkatan kandungan asam salisilat juga

20
terjadi setelah tanaman terpapar sinar UV atau tekanan ozon. Asam salisilat
menunjukkan fungsi sebagai senyawa sinyal yang penting yang bekerja dengan
mengaktifkan rangkaian sinyal untuk ekspresi enzim yang menjadi bagian dari
reaksi pertahanan terhadap virus, bakteri, dan jamur (Heldt 2005).
Fenilalanina amonia liase (PAL) mengkatalisis reaksi awal dari
metabolisme fenilpropanoid. Enzim ini mengkatalisis reaksi deaminasi dari
fenilalanina dan pembentukan sebuah ikatan ganda antar karbon. Pada beberapa
tumbuhan rerumputan, tirosina diubah menjadi asam 4-hidroksisinamat dengan
cara yang analog oleh tirosina ammonia liase. PAL adalah enzim yang paling
intensif dipelajari pada metabolisme sekunder tanaman. Enzim ini merupakan
tetramer dengan berat subunit 77 hingga 83 kDa. Pembentukan fitoaleksin
fenilpropanoid setelah infeksi jamur melibatkan induksi yang sangat cepat dari
PAL. PAL dihambat oleh produknya sendiri. Analog fenilalanina, asam
aminoksifenilpropanoid, juga merupakan inhibitor PAL yang sangat potenisal
(Heldt 2005). Reaksi oleh PAL dan struktur inhibitornya diberikan pada gambar 3.
Gambar 3 Reaksi deaminasi fenilalanina oleh PAL (Heldt 2005).
keterangan : a. pembentukan asam trans-sinamat.
b. asam aminoksifenilpropanoat, analog struktural
fenilalanina, menghambat PAL.
Enzim-enzim monooksigenase terlibat dalam sintesis fenol. Proses
pembentukan gugus hidroksil pada cincin fenil asam sinamat (hidroksilasi)
melalui sebuah reaksi terkatalisis monooksigenase menggunakan sitokrom P450
(cyt-P450) sebagai situs pengikatan O2 berdasarkan reaksi:
NADPH + H+ R-CH3 + O2 NADP
+ + RCH2-OH + H2O
a b

21
Pada reaksi ini, elektron ditransfer dari NADPH melalui FAD ke cyt-P450,
dan dari sana ke O2. Dari molekul O2, hanya satu atom O yang tergabung dalam
gugus hidroksil yang terbentuk. Atom O yang tersisa direduksi menghasilkan
H2O. Karena alasan ini, reaksi ini disebut monooksigenase. Selain O2, cyt-P450
juga bisa mengikat CO. karena hal ini, monooksigenase P450 dihambat oleh CO
(Heldt 2005). Reaksi sintesis asam hidroksisinamat dari asam trans-sinamat
dijelaskan pada Gambar 4.
Gambar 4 Sintesis asam hidroksisinamat dari asam trans-sinamat (Heldt 2005).
Monooksigenase P450 secara luas terdistribusi pada kingdom tumbuhan
dan hewan. Mayoritas protein ini mungkin terlibat dalam pembentukan gugus
hidroksil untuk sintesis hormon tanaman dan metabolit sekunder. Selain itu,
kelompok enzim ini juga berperan penting dalam proses detoksifikasi (Heldt
2005).

22
BAHAN DAN METODE
Alat dan Bahan
Alat-alat yang digunakan dalam proses kultur jaringan di Laboratorium
COPPU adalah Laminar Air Flow (LAF), pinset, scalpel, sterile blade, pembakar
bunsen, tatakan, kertas koran, korek api, cutter, gunting, petridish, specimen tube
(ST), tray dari besi dan platik, test tube (TT), botol selai ukuran sedang dan besar,
trolley, rak, autoklaf (merk Hirayama, Astell, dan ASP), pH meter, panci, neraca
analitik, labu erlenmeyer 100 ml dan 500 ml, kompor gas, magnetic stirer,
spatula, alat penjetek, dan pipet Ependorf.
Bahan-bahan yang digunakan adalah umbut (calon daun), air RO, media
MS modifikasi, sukrosa, gelrite, glukosa, natrium hipoklorit, hormon auksin,
alkohol, dan wrapping plastik.
Metode Penelitian
Perbanyakan vegetatif kelapa sawit unggul di COPPU menggunakan
metode kultur jaringan dari Bhojwani dan Razdan (1983). Tetapi, metode yang
dipakai saat ini telah mengalami berbagai pengembangan dari hasil penelitian dan
saran dari konsultan. Komposisi bahan kimia dan zat pengatur tumbuh (ZPT)
yang digunakan tidak diizinkan untuk dipublikasikan oleh manajemen COPPU.
Preparasi Media
Formulasi medium yang digunakan mengacu pada medium Murashige dan
Skoog (1962) dengan beberapa modifikasi. Media yang digunakan terdiri atas dua
jenis, yaitu medium padat dan medium cair. Perbedaan formulasi medium padat
dan cair terletak pada penggunaan bahan pemadat. Medium padat menggunakan
agar sebagai bahan pemadat, sedangkan medium cair menggunakan gelrite
sebagai bahan peningkat konsistensi media. Setelah semua larutan stok
diformulasi dan ditambahkan ZPT serta sukrosa, penetapan pH media baru
dilakukan. Media ditetapkan pH-nya pada 5,70+0,01. Setelah penetapan pH,
penambahan baru dilakukan. Pemasakkan media dilakukan dalam panci dengan
pemanasan kompor hingga mendidih. Selama pemasakkan media, media harus
terus diaduk menggunakan alat pengaduk elektrik. Setelah mendidih, media
dituang dalam vessel sesuai dengan kebutuhan. Media padat yang telah dituang

23
dalam vessel kemudian disterilisasi menggunakan autoklaf pada suhu 114 oC
selama 2 jam (modifikasi Rasool et al 2011).
Inokulasi Eksplan
Eksplan yang digunakan di COPPU berasal dari umbut. Umbut yang
diambil sebagai eksplan berasal dari daun minus, yaitu daun yang belum keluar
dari tempat munculnya daun. Kemudian, eksplan disterilisasi menggunakan
NaHClO dengan konsentrasi tertentu. Setelah sterilisasi, eksplan dibilas dengan
akuades steril beberapa kali dan disimpan dalam larutan glukosa 2% selama
menunggu ekplan ditransfer. Umbut yang digunakan sebagai eksplan dipotong
melintang menggunakan scalple steril dengan ketebalan sekitar 2 mm. setelah itu,
eksplan diinokulasikan pada media padat di cawan petri. Satu cawan petri
maksimal hanya bisa diisi oleh 3 potong eksplan (modifikasi Singh et al 2011).
Inokulasi Kalus
Setelah kalus muncul dari eksplan, maka eksplan tersebut dipindahkan ke
tahap callus development. Eksplan yang sudah memunculkan kalus dipindahkan
pada media MS padat modifikasi di cawan petri. Pemindahan dilakukan secara
aseptik mengunakan alat bantu pinset (Mineo 1990).
Transfer EC Rendah
New embryoid (Ne) yang dihasilkan dari tahap callus development
dipindahkan ke tahap embryoid culture rendah dengan kode EC1. Ne
dipindahkan secara aseptik pada medium MS padat modifikasi dalam sampel tube
(ST). Proses transfer dibantu dengan pinset steril. Embryoid yang muncul
dipotong (diurai) dengan scalple steril berdasarkan retakannya agar kultur
berkembang dengan baik. Transfer penyegaran media subkultur dilakukan tiap 2
bulan (Chehmalee dan Te-chato 2008).
Transfer EC Tinggi
Embryoid dan shoot yang berkembang pada tahapan EC tinggi dipisahkan
(dipotong) berdasarkan alur/retakannya menggunakan scalple steril. Embryoid
dipindahkan secara aseptik pada medium MS padat modifikasi pada botol sedang
dengan alat bantu pinset steril. Ne yang muncul (jika ada) dipindahkan ke ST

24
berisi MS modifikai padat. Seleksi dilakukan pada tunas yang telah memiliki
panjang 3 cm atau lebih untuk selanjutnya dimasukkan dalam tahapan SD. Buang
embrioid yang mengalami pencoklatan dan hyperhydricity, akar napas
(pneumorf), tunas abnormal, dan jaringan yang mati. Transfer dilakukan hanya
pada embryoid yang bagus dan tunas yang normal (Chehmalee dan Te-chato
2008).
Transfer SD-RI
Shoot yang sudah memiliki panjang 7 cm atau lebih pada tahap SD atau
EC tinggi dipindahkan ke medium penginduksi akar pada botol sedang. Akar dan
seludang daun yang muncul dipotong dan dibuang menggunakan scaple dan
pinset steril. Daerah bekas pemotongan akar dibuat berbentuk lancip agar menjadi
tempat tumbuhnya akar hasil induksi di tahapan RI (Chehmalee dan Te-chato
2008).
Aklimatisasi Ramet
Ramet hasil kultur jaringan kelapa sawit ditempatkan dalam net house. Di
dalam net house, ramet dikeluarkan dari TT dan dicuci untuk membersihkan sisa
media, kemudian dibilas dengan fungisida (Tiplo®) dan ditanam dalam polibag
kecil. Ramet kemudian ditempatkan dalam sungkup plastik. Pada pagi dan sore
hari, ramet disiram untuk memenuhi kebutuhan air dan menjaga kelembapan agar
tetap tinggi. Selama proses aklimatisasi, sungkup plastik dibuka secara bertahap
mulai dari satu sisi, kedua sisi, hingga sungkup plastik dilepas semua (modifikasi
Mineo 1990).
Cek Kontaminasi
Semua kultur yang disimpan dalam dalam growth room diambil untuk
diperikasa terhadap kontaminasi setiap 2 minggu sekali. Kultur diamati secara
langsung di dalam growth room dan jika ditemukan kultur yang terkontaminasi,
kultur tersebut dipisahkan. Kultur yang terkontaminasi selanjutnya didata dan
dimusnahkan. Kultur yang terkontaminasi dimusnahkan dengan autoklaf hingga
semua kontaminan di dalamnya mati (Yusnita 2008)

25
HASIL DAN PEMBAHASAN
Hasil Inokulasi Eksplan dan Kalus
Kultur jaringan adalah suatu metode untuk mengisolasi bagian dari
tanaman seperti sekelompok sel atau jaringan yang ditumbuhkan dengan kondisi
aseptik, sehingga bagian tanaman dapat memperbanyak diri tumbuh menjadi
tanaman lengkap kembali (Hameed et al 2006). Teknik kutur jaringan
memanfaatkan prinsip perbanyakan tumbuhan secara vegetatif. Berbeda dari
teknik perbanyakan tumbuhan secara konvensional, teknik kultur jaringan
dilakukan dalam kondisi aseptik di dalam botol kultur dengan medium dan
kondisi tertentu. Karena hal ini, teknik ini sering disebut kultur in vitro (Gunawan
1987).
Eksplan adalah material tanaman awal yang digunakan untuk perbanyakan
vegetatif tanaman menggunakan teknik kultur jaringan. Material tanaman yang
sering dikulturkan diantaranya kambium vaskular, parenkim penyimpan, pericycle
dari akar, kotiledon, mesofil daun, dan jaringan provaskular. Faktanya, semua
tanaman multiselular adalah sumber eksplan yang potensial untuk inisiasi kalus
(Dodds dan Lorin 1985).
Pada kelapa sawit, sumber eksplan yang telah diteliti adalah umbut, akar,
dan bunga. Umbut merupakan daun muda yang yang masih belum keluar. Kultur
jaringan di COPPU menggunakan umbut sebagai sumber eksplannya. Kelebihan
umbut sebagai sumber eksplan adalah kontamin bawaan relatif lebih sedikit, dan
merupakan jaringan yang relatif muda sehingga masih mudah membelah
(Zulkarnain 2009). Kultur jaringan adalah teknik perbanyakan tanaman dengan
menggunakan bagian dari tanaman tersebut (organ, protoplasma, sel) yang
dikerjakan dalam kondisi steril. Kondisi steril merupakan syarat umum dalam
teknik kultur jaringan (Bhojwani dan Razdan 1983). Berbeda dengan umbut,
penggunaan bunga sebagai sumber eksplan masih dalam tahap pengembangan dan
belum ada yang melaporkan keberhasilan perbanyakan kelapa sawit dengan
eksplan bunga. Akar tidak diutamakan digunakan sebagai sumber eksplan karena
asal pohon induk sangat sulit ditelusur dengan pasti (karena letaknya di bawah
tanah), dan kandungan kontaminan bawaan yang tinggi (Zulkarnain 2009).

26
Gambar 5 Hasil inokulasi umbut kelapa sawit sebagai eksplan kultur jaringan.
keterangan: a. kultur eksplan sebelum inkubasi,
b. kultur eksplan sesudah inkubasi.
Tahapan awal dari kultur jaringan adalah inokulasi bagian tanaman
(eksplan) yang akan dikultur. Sebelum diinokulasi, umbut harus disterilisasi
terlebih dahulu untuk menghilangkan kemungkinan terikutnya kontaminan
(Hutami 2008). Tujuan dari tahapan inokulasi adalah menstimulir munculnya
kalus.
Eksplan yang digunakan di COPPU berasal dari umbut. Sebelum
ditransfer ke media tumbuh, bagian dari umbut tersebut harus diseleksi dulu agar
didapatkan bagian umbut yang layak untuk dikulturkan. Umbut yang diambil
sebagai eksplan berasal dari daun minus, yaitu daun yang belum keluar dari
tempat munculnya daun. Daun minus diambil karena lokasinya berada di bagian
dalam sehingga kontaminanya sedikit. Saat pertama kali diinokulasi, eksplan ini
masuk dalam tahapan Ep1 (kultur eksplan pertama) dan akan diinkubasi selama 6
bulan dalam dark room hingga mengeluarkan kalus (Gambar 5). Pada awal
inokulasi, eksplan masih terlihat segar dan berwarna cerah. Warna eksplan akan
semakin coklat, bahkan menghitam, selama masa inkubasi. Setiap 2 minggu
sekali, dilakukan pengecekan kultur untuk melihat adanya kontaminasi dan
melihat perkembangan eksplan. Jika eksplan sudah memunculkan kalus, maka
kalus beserta eksplan tersebut dipindahkan ke media dalam petri lain dan masuk
ke tahap kultur kalus (Gambar 6).

27
Gambar 6 Kultur kalus pada kultur jaringan kelapa sawit.
keterangan: a. kalus, b. eksplan, c. media, d. newembryoid.
Setelah kalus muncul pada kultur eksplan, maka kalus tersebut akan
masuk dalam tahap callus development (kultur kalus). Dalam tahap ini, kalus
dikulturkan lebih lanjut dan akan melalui tahapan somatic embryogenesis
sehingga menghasilkan embrio (Chehmalee dan Te-chato 2008). Kalus merupakan
masa sel yang belum terdiferensiasi dengan bentuk yang tidak beraturan, dan
berasal dari hasil dediferensiasi jaringan induk (Dodds dan Lorin 1985). Secara
histologi, kalus berasal dari pembelahan berkali-kali sel-sel parenkim di sekitar
berkas pengangkut kecuali xilem. Kalus sering muncul pada ujung potongan akar
atau daun sebagai hasil pelukaan. Dengan menggunakan teknik kultur jaringan,
pembentukan kalus bisa diinduksi di sejumlah jaringan dan organ tanaman yang
biasanya tidak membentuk kalus dalam respon pelukaan. Kalus dapat diinduksi
dengan menambahkan ZPT yang sesuai pada media kultur, misalnya auksin dan
sitokinin yang disesuaikan. Jika konsentrasi auksin lebih besar daripada sitokinin,
maka kalus akan terbentuk. Sedangkan jika konsentrasi sitokinin yang lebih besar
dibandingkan dengan auksin, maka yang terbentuk bukanlah kalus, melainkan
tunas (Gunawan 1987). Pada kultur jaringan, kalus akan mengalami somatik
embryogenesis sehingga menghasilkan embrio yang bisa berkembang menjadi
tanaman utuh (Yusnita 2008).
Pengamatan yang dilakukan pada kultur kalus yang sudah diinkubasi pada
dark room mendapati adanya perkembangan embrio somatik dari hasil somatik
embryogenesis (Gambar 6). Embrio somatik ini disebut sebagai new embryoid
(Ne). Kalus hasil kultur jaringan di COPPU terlihat putih dan berair. Chemale dan
Te-chato (2008) berhasil menginisiasi pembentukan kalus pada tanaman kelapa
sawit dengan eksplan embrio dan mendapatkan kalus berbentuk nodular callus

28
(NC), embriogenic callus (EC), dan root like callus (RLC). Pengamatan pada
proses kultur jaringan di COPPU menunjukkan ketiga jenis kalus muncul tersebut
muncul pada kultur eksplan dan kultur kalus.
Somatik embryogenesis in vitro dari tanaman umumnya melibatkan dua
tahap, yaitu induksi dediferensiasi dan rediferensiasi (Jun-Yan et al 2000). Induksi
dediferensiasi bertujuan untuk menghasilkan kalus. Induksi dediferensiasi dari sel
yang sudah terdiferensiasi pada eksplan biasanya terjadi dengan penambahan ZPT
pada medium kultur. ZPT (hormon tanaman) pada medium kultur membuat sel
yang dikultur mulai kembali membelah menghasilkan kalus. Rediferensiasi
bertujuan untuk menghasilkan embrio somatik dari kalus yang bersifat
embriogenik. Hanya kalus yang bersifat embriogenik yang bisa berubah menjadi
embrio somatik. Embrio somatik ini lalu bisa menyempurnakan
perkembangannya dan berubah menjadi plantlet pada medium bebas hormon.
Jadi, embrio somatik bisa membentuk tanaman tanpa stimulasi dari hormon
tanaman. Pengamatan pada somatik embryogenesis dari beberapa tanaman
(contohnya wortel) membuktikan bahwa embrio somatik berasal dari sel tunggal
dalam kalus embriogenik (Jun-Yan et al 2000).
Somatik embryogenesis dibagi menjadi menjadi somatik embryogenesis
langsung dan somatik embryogenesis tidak langsung. Somatik embryogenesis
langsung tidak melibatkan fase kalus dalam pembentukan embrio somatik, tetapi
embrio muncul langsung dari eksplan. Somatik embryogenesis tidak langsung
melibatkan fase kalus untuk menghasilkan embrio somatik. Pada somatik
embryogenesis tidak langsung, embrio somatik muncul dari diferensiasi kalus.
Kelapa sawit adalah tanaman monokotil, dan tidak seperti kebanyakan
spesies tanaman lain, perbanyakan vegetatif hanya bisa dilakukan melalui kultur
jaringan (Kushairi et al 2010). Kultur jaringan kelapa sawit memberikan jalan
untuk konservasi pohon induk unggul, baik melalui penanaman klon di lapangan,
maupun penyimpanan kultur kalus embriogenik dan embrio somatik secara
cryopreservation (Jun-Yan et al 2000).
Eksplan yang tidak memunculkan kalus selama 6 bulan inkubasi harus
ditransfer ke medium yang baru. Setelah ditransfer, eksplan ini akan masuk dalam
tahap Ep2 (kultur eksplan kedua) dan akan ditunggu lagi selama 6 bulan untuk

29
menghasilkan kalus. Cek kontaminasi sekaligus pengamatan kemunculan kalus
dilakukan setiap 2 minggu sekali dalam tahapan Ep1 maupun Ep2. Jika setelah 6
bulan eksplan tidak muncul kalus di Ep2, maka kultur eksplan tersebut akan
dibuang (discard). Jika telah muncul kalus, eksplan tersebut dipindahkan ke
tahapan kultur kalus (C) dan diberi nomor line (urutan kemunculan kalus dalam
satu nomor palm yang diperbanyak. Transfer ke medium baru setelah 6 bulan
kultur eksplan perlu dilakukan karena inkubasi pada tenggat waktu tersebut akan
dibarengi dengan pengurangan nutrisi esensial dan pengeringan perlahan pada
medium akibat penguapan air (Dodds dan Lorin 1985).
Tahapan kultur kalus mulai dari C1 hingga C6. Perpindahan tahapan
dilakukan setiap 2 bulan sekali. Cek kemunculan Ne dilakukan setiap 2 minggu
pada kultur kalus. Tahap kultur kalus bertujuan untuk menginduksi munculnya Ne
dari kalus sebagai sumber embrio somatik untuk perbanyakan vegetatif kelapa
sawit. Jika muncul Ne, Ne tersebut dipindahkan untuk masuk ke dalam kultur EC
rendah. Skoring terhadap Ne sebelum proses transfer dilakukan untuk menentukan
kualitas Ne tersebut.
Hasil Transfer EC Rendah dan EC Tinggi
Tahapan selanjutnya yang akan dilakukan pada embrio yang sudah
terbentuk pada kultur kalus adalah multiplikasi dan polyembryoid. Multiplikasi
adalah kemunculan embryoid dari kalus yang muncul dari eksplan. Sedangkan
tahapan lebih lanjut pada multiplikasi adalah polyembryoid. Tahapan multiplikasi
embryoid ada 18 tahapan subkultur, dibedakan EC rendah (1-4) dan EC tinggi (5-
18). Pada tahapan polyembryoid, embryoid mulai berdiferensiasi menjadi organ,
yaitu tunas dan akar (Bhojwani dan Razdan 1983). Pembagian EC rendah dan EC
tinggi hanya semata-mata dilakukan karena ada perbedaaan penekanan. Angka 1
hingga 18 merupakan jumlah penyegaran medium yang telah dilakukan pada
kultur tersebut.
EC rendah merupakan fondasi dari kultur embryoid yang ada di
laboratorium sehingga lebih diutamakan dalam hal pencegahan kontaminasi dan
ketepatan waktu subkultur (Yusnita 2008). EC rendah didominasi oleh embrio
berbentuk new embryoid (embrio yang terbentuk langsung dari kalus). Klasifikasi

30
Gambar 7 Kultur EC rendah pada kultur jaringan kelapa sawit.
keterangan: a. NodAgg, b. NodFr, c. Embryoid.
Ne berdasarkan bentuk dan terdiri dari nodular friable (NodFr) dan nodular
aggregate (NodAgg) (Chehmalee dan Te-chato 2008). EC tinggi didominasi
embryoid dengan banyak variasi (chunky, finger like structure, wavy, dan molar
like stucture) dan tunas (shoot). Dalam perjalanan poliembryoid di EC tinggi,
luaran yang diharapkan adalah tegakan (shoot) (Yusnita 2008).
Pada kultur EC rendah (Gambar 7), teramati adan Ne berbentuk NodAgg
dan NodFr yang berkembang dari Ne subkultur sebelumnya. Ne yang dikultur
juga berkembang menjadi embryoid berberbentuk chungky. NodAgg terbentuk
karena perkembangan embrio somatik yang cenderung mengumpul dalam satu
agregat kompak. NodFr terbentuk karena perkembangan masing-masing embrio
somatik menyebar dalam suatu daerah sehingga terlihat pecah (Chehmalee dan
Te-chato 2008).
Ne yang muncul pada EC rendah dan EC tinggi harus dipisahkan dan
dikultur pada vessel yang berbeda (ST) saat proses subkultur. Hal yang perlu
diperhatikan dalam transfer EC adalah waktu transfer. Waktu transfer EC
diusahakan secepat mungkin karena jika waktu transfer lebih dari 10 menit, kultur
akan mengalami pencoklatan dan akan menurunkan tingkat keberhasilan kultur
dan kualitas kultur selanjutnya.

31
Gambar 8 Kultur EC tinggi pada kultur jaringan kelapa sawit.
keterangan: a. embryoid, b. NodFr, c. jaringan yang hyperhydricity, d.
d.ipencoklatan jaringan.
Pengamatan pada EC tinggi mendapati adanya jaringan yang mengalami
hyperhydricity dan pencoklatan (browning) (Gambar 8). Kemunculan jaringan
hyperhydricity, pencoklatan, dan kematian jaringan eksplan pada kultur jaringan
di COPPU merupakan salah satu masalah yang dihadapi oleh manajemen
laboratorium. Pencoklatan sangat umum terjadi pada spesies tanaman berkayu,
terutama bila eksplan diambil dari pohon dewasa. Penghambatan pertumbuhan
biasanya sangat kuat pada beberapa spesies yang umumnya mengandung senyawa
tanin atau hidroksifenol dengan konsentrasi tinggi (Hutami 2008). Pencoklatan
pada jaringan muda lebih sedikit dibandingkan dengan jaringan yang tua (George
dan Sherrington 1984). Karena hal ini, kultur jaringan di COPPU menggunakan
jaringan meristem yang lebih muda seperti bagian umbut pada kelapa sawit.
Kejadian pencoklatan sangat menurunkan tingkat proliferasi dan pertumbuhan
dari kultur. Kultur yang mengalami pencoklatan memiliki pertumbuhan yang
lambat dan perkembangan yang tidak bagus. Jika pencoklatan terjadi pada
tingkatan yang sangat parah, kultur tersebut diseleksi dengan kualifikasi bad
culture karena kultur jenis ini bisa meningkatkan tingkat abnormalitas tanaman
kelapa sawit yang dihasilkan.
Pencoklatan jaringan terjadi karena aktivitas enzim oksidase yang
mengandung tembaga seperti katekol oksidase (EC 1.10.3.1) dan monofenol
monooksigenase (EC 1.14.18.1) yang dilepaskan atau disintesis dan tersedia pada

32
kondisi oksidatif ketika jaringan dilukai. Katekol oksidase dan monofenol
monooksigenase memilki nama lain polifenol oksidase (PPO) (daftar enzim
oksidoreduktase di NC-IUBMB). Substrat untuk enzim ini ada bermacam-macam
pada jaringan yang berbeda, yang umum adalah tirosin dan katekol. Berikut
adalah reaksi yang dikatalisis oleh dua enzim tersebut.
Polifenol Oksidase : 2 katekol + O2 2 1,2-benzokuinon + 2 H2O
Monofenol Monooksigenase : L-tirosina + L-dopa + O2 L-dopa + dopaquinon
+ H2O
Enzim dan substrat dalam keadaan normal akan tertahan dalam ruang
berbeda di dalam sel dan akan keluar bersama-sama pada saat sel dilukai atau
hampir mati. Fenol mempunyai fungsi alami penting dalam mengatur oksidasi
IAA. Beberapa monofenol seperti asam sinaptat dan asam ferulat pada konsentrasi
yang rendah akan menghambat oksidasi enzimatik dari IAA. Hal ini
menghasilkan perpanjangan dan pembelahan sel, serta peningkatan pertumbuhan
dan perkembangan tanaman.
Toksisitas fenol kemungkinan disebabkan oleh ikatan reversibel antara
hidrogen dan protein. Penghambatan pertumbuhan yang tidak dapat diperbaiki
terjadi ketika fenol teroksidasi menjadi senyawa aktif quinon yang tinggi yang
kemudian memutar, memolimerase dan/atau mengoksidasi protein menjadi
senyawa melanat yang makin meningkat (Hutami 2008). Ozyigit et al (2007)
melaporkan bahwa terbentuknya senyawa fenol dipengaruhi oleh struktur
kimianya, spesies tanaman, proses biologi (organogenesis atau somatik
embriogenesis), dan tahap perkembangannya. Oksidasi fenol yang berubah
menjadi quinon dan senyawa lain (polimerisasinya) yang sangat beracun
menyebabkan pencoklatan medium dan kematian eksplan.
Penanggulangan pencoklatan pada jaringan, khususnya eksplan yang baru
diisolasi, dan pada media tumbuh yang digunakan, seringkali dilakukan dengan
menghilangkan senyawa fenol pada eksplan, modifikasi potensial redoks,
penghambatan aktivasi enzim fenol oksidase, dan penurunan aktivitas fenolase
serta ketersediaan substrat (George dan Sherrington 1984 dalam Hutami 2008).
Kemunculan jaringan hyperhydricity pada kultur jaringan di COPPU
merupakan salah satu masalah yang dihadapi oleh manajemen. Jaringan

33
hyperhydricity merupakan kultur abnormal dan akan dibuang pada saat proses
transfer. Hyperhydricity (terminologi awal disebut vitrifikasi) adalah kelainan
fisiologi yang terjadi pada tanaman herba dan berkayu selama propagasi vegetatif
in vitro. Ciri jaringan yang mengalami hyperhydricity jaringan yang menebal dan
transparan, daun menebal, keriting dan atau bergelombang, sering berukuran
sangat besar, dan mudah pecah. Beberapa peneliti mendefinisikan sebagai
malformasi hingga kekurangan klorofil, dan kelebihan kandungan air pada sel.
Tipe kalus berair yang kehilangan elemen xilem juga digolongkan sebagai
hyperhydricity (Gaspar et al 1987).
Petumbuhan yang rendah dari jaringan yang mengalami hyperhydricity
disebabkan laju multiplikasi yang rendah. Kultur seperti ini akan cepat kehilangan
semua kapasitas untuk propagasi sehingga cenderung untuk dibuang (discard).
Proses hyperhydricity terkadang dapat kembali menjadi normal. Jaringan yang
mengalami hyperhydricity jika ditempatkan pada medium yang tidak
menyebabkan vitrifikasi seringkali menunjukkan perkembangan anatomi menjadi
tanaman normal (Gaspar et al 1987).
Terdapat beberapa perbedaaan fisiologis dan biokimiawi pada jaringan
hyperhydricity. Jaringan yang mengalami hyperhydricity memiliki kandungan air
yang lebih tinggi dibanding jaringan normal. Kekurangan proses lignifikasi juga
terjadi pada dinding sel kalus yang mengalami hyperhydricity. Hal ini
menyebabkan dinding sel mengandung lebih sedikit lignin dan selulosa dibanding
kalus normal. PAL dan peroksidase asam, enzim yang terlibat dalam proses
lignifikasi, memiliki aktivitas lebih kecil pada jaringan hyperhydricity. Jaringan
hyperhydricity juga memilki kandungan klorofil serta aktivitas fotosintesis yang
lebih kecil dibanding jaringan normal (Gaspar et al 1987).
Salah satu tahapan dalam kultur jaringan kelapa sawit adalah shoot
development (SD). Tahapan ini bertujuan memperpanjang panjang tegakan yang
didapat dari proses transfer EC tinggi hingga mencapai tinggi minimal 7 cm.
Perlakuan hormon-hormon tertentu pada medium yang digunakan pada tahap SD
membuat tegakan cenderung bertambah panjang. Shoot yang masuk ke tahap SD
harus memilki panjang 3 cm atau lebih.

34
Gambar 9 Kultur SD pada kultur jaringan kelapa sawit.
Pengamatan pada kultur SD yang ada di COPPU menunjukkan
pertambahan tinggi tegakan selama inkubasi di light room (Gambar 9). Inkubasi
di light room diperlukan untuk merangsang pembentukan kloroplas dan
mengarahkan shoot agar tumbuh tegak. Kultur SD menggunakan botol besar agar
vessel ini mampu menampung pertumbuhan kultur yang semakin panjang.
Pada SD, kemungkinan akan berkembang juga akar pada shoot. Namun,
perkembangannya tidak sebaik yang perpanjangan shoot. Akar ini cenderung tidak
berfungsi baik dan mudah lepas jika dibiarkan sampai penanaman di lapangan.
Maka, saat transfer ke RI, akar ini akan dibuang.
Hasil Transfer SD-RI
Saat panjang tegakan sudah mencapai 7 cm atau lebih, maka tegakan
tersebut harus dipindahkan pada tahapan selanjutnya, yaitu tahapan root induction
(RI). Tahapan RI bertujuan merangsang pertumbuhan akar pada tunas kelapa
sawit karena perlakuan hormon-hormon perangsang pertumbuhan akar. Hasil
transfer SD-RI disajikan pada Gambar 10.
Gambar 10 Hasil transfer SD-RI pada kultur jaringan kelapa sawit.

35
Tahapan RI dalam kultur jaringan kelapa sawit dibagi menjadi 2 bagian
berdasarkan jenis medianya, yaitu bagian yang menggunakan media cair dan
media padat. Media padat disebut media RI dan media cair disebut media RG.
Subkultur pada tahapan RI meliputi R1, R2, dan R3. Tahapan pertama dilakukan
selama 2 bulan hingga ada akar induksi yang muncul pada tunas. Jika akar induksi
ini muncul dan telah memenuhi syarat ramet, maka bisa langsung dikeluakan dari
proses subkultur. Jika telah muncul akar induksi, tetapi belum memenuhi syarat
ramet, maka tunas tersebut dipindahkan ke tahapan R2 menggunakan medium RG.
Bila dalam tahap R1 tidak muncul akar induksi, tahapan selanjutnya adalah R2
dengan masih menggunakan medium R1. Tahapan R3 hanya ada untuk medium
RG dan tunas yang dikultur berasal dari tahapan R2. Proses transfer tunas dari
medium RI ke RG disebut transfer RI-RG.
Aklimatisasi Ramet
Luaran dari proses kultur jaringan di lab adalah ramet. Ramet atau plantlet
adalah tanaman hasil dari kultur jaringan yang sudah memiliki organ utama yang
lengkap. Organ utama yang dimaksud adalah akar, batang, dan daun. Sebelum
ditanaman di lapangan, ramet harus menjalani proses yang disebut aklimatisasi.
Ramet hasil kultur jaringan kelapa sawit di COPPU ditampilkan pada Gambar 11.
Proses aklimatisasi (hardening) dilakukan selama sebulan. Selama proses
ini, ramet di tempatkan dalam net house agar terlindung dari intensitas cahaya
berlebih. Di dalam net house, ramet dikeluarkan dari TT dan dicuci untuk
membersihkan sisa media, kemudian dibilas dengan fungisida (Tiplo®) dan
ditanam dalam polibag kecil. Sisa media harus dibersihkan agar akar ramet bisa
mengambil nutrisi dari tanah dengan mudah.
Ramet tersebut ditempatkan dalam sungkup plastik untuk menjaga
kelembapan tetap tinggi. Pada pagi dan sore hari, ramet disiram untuk memenuhi
kebutuhan air dan menjaga kelembapan agar tetap tinggi. Selama proses
hardening, sungkup plastik dibuka secara bertahap mulai dari satu sisi, kedua sisi,
hingga sungkup plastik dilepas semua. Hal ini penting agar ramet memiliki waktu
untuk beradaptasi. Waktu yang dibutuhkan untuk aklimatisasi ramet kelapa sawit
adalah 1 bulan.

36
Gambar 11 Ramet hasil kultur jaringan kelapa sawit.
Ramet hasil kultur jaringan yang diregenerasikan dalam lingkungan
dengan kelembaban tinggi dan bersifat heterotrof, harus menjadi autotrof bila
dipindahkan ke lapangan. Proses pemindahan merupakan langkah akhir dari
prosedur mikropropagasi dan diistilahkan sebagai tahap aklimatisasi. Aklimatisasi
adalah suatu upaya mengkondisikan plantlet atau tunas minor hasil perbanyakan
melalui in vitro ke lingkungan in vivo yang septik (Yusnita 2008).
Masa aklimatisasi merupakan masa yang kritis karena pucuk dan plantlet
yang diregenerasikan dari kultur in vitro menunjukkan beberapa sifat yang
menguntungkan, seperti lapisan lilin (kutikula) tidak berkembang dengan baik,
kurangannya lignifikasi batang, jaringan pembuluh dari akar ke pucuk kurang
berkembang, dan stomata seringkali tidak berfungsi. Keadaan ini menyebabkan
pucuk-pucuk in vitro sangat peka terhadap transpirasi, serangan cendawan dan
bakteri, cahaya dengan intensitas tinggi, dan suhu tinggi. Oleh karena itu,
aklimatisasi pucuk-pucuk in vitro memerlukan penanganan khusus, bahkan
diperlukan modifikasi terhadap kondisi lingkungan terutama dalam kaitannya
dengan suhu, kelembapan, dan intensitas cahaya (Zulkarnain 2009).
Kontaminasi
Kontaminasi menjadi salah satu masalah dalam kultur jaringan.
Kontaminasi bisa menggagalkan kultur yang sedang ditumbuhkan dengan
merusak media, memperlambat pertumbuhan kultur, bahkan mematikan kultur.
Kontaminasi besar yang kerap terjadi adalah jamur dan bakteri (Gambar 8b dan
8c). Sumber kontaminasi lainnya yang kerap ditemui adalah sejenis laba-laba

37
penghisap yang disebut mites (Gambar 8a). Mites hampir tidak terlihat oleh mata
telanjang dan harus dilihat dengan kaca pembesar agar bisa terlihat lebih jelas.
Kontaminasi oleh jamur dan bakteri dapat mudah dikenali karena terlihat jelas
oleh mata telanjang. Penampakan kultur yang terkontaminasi bakteri terlihat pada
permukaan media dan retakan-retakan di dalam media. Kultur yang
terkontaminasi bakteri akan terlihat memiliki koloni bakteri di permukaan yang
berpenampakan keruh. Retakan-retakan pada media yang terkontaminasi bakteri
juga akan jelas terlihat keruh akibat kolonisasi bakteri.
Untuk kontaminasi jamur, penampakan kultur akan menunjukkan koloni
hifa jamur yang terlihat sangat jelas. Koloni hifa jamur ini memilki variasi warna
mulai dari putih, coklat, bahkan bisa berwarna hitam. Jamur sangat cepat
menyebar karena berkembang dengan spora. Kontaminasi oleh mites bisa
menyebabkan kultur tersebut mengalami kelayuan, bahkan kematian. Mites
menghisap makanan dari kultur sehingga kultur mengalami kerusakan yang
signifikan. Selain itu, mites juga membawa kontaminsi jamur pada kultur karena
selama menginvasi kultur, mites bisa membawa spora jamur yang menempel
padanya (Zulkarnain 2009).
Untuk mengendalikan tingkat kontaminasi pada kultur, COPPU
memberlakukan aturan-aturan di lab yang membuat kontamin mempunyai
peluang minimal untuk masuk dalam kultur. Aturan-aturan tersebut meliputi
teknik pengambilan umbut, sterilisasi laminar air flow dan peralatan yang
digunakan, pembersihan setiap ruangan secara teratur, cek tingkat kontaminasi di
ruangan (laminar air flow, growth room dan ruang transfer), dan POS di
laboratorium (Zulkarnain 2009).
Gambar 12 Kultur yang terkontaminasi.
Keterangan: a. Mites, b. Bakteri, c. Jamur.

38
Teknik pengambilan umbut dari ortet harus hati-hati agar kontamin yang
terbawa minimal. Saat pengiriman ke lab, kultur harus dibungkus dahulu dan
disimpan dalam ruangan dingin untuk menghambat perkembangan kontaminan.
LAF dan alat-alat yang digunakan untuk transfer kultur harus disterilisasi setiap
hari agar tidak membawa kontaminan pada kultur. Pembersihan ruang transfer,
ruang subkultur, ruang preparasi media, dan ruang-ruang lainnya dalam
lingkungan lab harus dibersihkan setiap hari. Untuk mengecek tingkat
kontaminasi dari ruangan, secara teratur tiap bulan dilakukan cek kontaminasi
ruangan dengan menaruh media terbuka dalam ruangan yang dicek. Kontaminasi
ruangan selain di LAF dijaga seminimal mungkin. POS yang diberlakukan di
COPPU juga dibuat untuk meminimalkan kontaminasi pada kultur.
Cek kontaminasi pada kultur dilakukan setiap dua minggu agar
kontaminasi tidak menyebar dalam growth room. Cek kontaminasi akan
menyeleksi kultur-kultur yang terkontaminasi dan memusnahkannya. Jumlah
kultur yang dimusnahkan akan mempengaruhi tingkat keberhasilan kultur
jaringan.

SIMPULAN DAN SARAN
Simpulan
Kultur jaringan berhasil digunakan untuk memperbanyak bibit kelapa
sawit unggul di PT Tunggal Yunus Estate. Pencoklatan, kontaminasi dan bad
culture, didapatkan berpengaruh terhadap kualitas dan multiplikasi tanaman.
Kegiatan praktik lapang di PT Tunggal Yunus Estate menjadi salah satu
perwujudan penerapan ilmu biokimia dalam dunia kerja nyata. Praktik lapang di
PT Tunggal Yunus Estate ini juga telah memberikan mahasiswa tambahan
pengetahuan dan keterampilan dalam teknik kultur jaringan skala besar.
Saran
Perlu beberapa penelitian lanjutan untuk mencari cara penanggulangan
pencoklatan, bad culture, dan kontaminasi yang lebih baik dan efisien secara
ekonomi.

40
DAFTAR PUSTAKA
Andersone U, Levinsh G. 2002. Changes of morphogenic competence in mature
Pinus sylvestris L. buds in vitro. Annals of Botany. 90:293-298.
Antolovich et al. 2000. Sample preparation of phenolic compound in fruits.
Analyst. 125:989-1009.
Bhojwani SS, Razdan MK. 1983. Plant Tissue Culture: Theory and Practice.
Amsterdam: Elsevier Science Publisher.
Chehmalee S, Te-chato S. 2008. Induction of somatic embryogenesis and plantlet
regeneration from cultured zygotic embryo of oil palm. J Agri Tech. 4(2):
137-146.
Creasy LL. 1968. The increase in phenylalanine ammonia-lyase activity in
strawberry leaf disks and its correlation with flavonoid synthesis.
Phytochem 7:441-446.
Dodds JH, Roberts LW. 1985. Experiment in Plant Tissue Culture, Second
Edition. Cambridge University Press: Cambridge.
Fauzi Y et al. 2008. Kelapa Sawit: Budi Daya Pemanfaatan Hasil dan Limbah
Analisis Usaha dan Pemasaran. Jakarta: Penebar Swadaya.
Gaspar TH, Kevers C, Debergh P, Maene L, Paques M, Boxus PH. 1987.
Vitrivication: morphological, physiological, and ecological aspects.
Dalam: JM Bonga dan DJ Durzan (Editor). Cell and Tissue Culture in
Forestry, Volume 1: General Principle and Biotechnology, Kluwer
Academic Publisher Group: Dordrecht. Hal: 152-166.
George EF. and Sherrington PD. 1984. Plant Propagation by Tissue Culture. Hand
Book and Directory of Comercial Laboratories. London: Eastern Press.
Gunawan LW. 1987. Teknik Kultur Jaringan. Bogor: Laboratorium Kultur
Jaringan PAU Bioteknologi IPB.
Hameed N, Shabbir A, Ali A, Bajwa R. 2006. In vitro micropropagation of disease
free rose (Rosa indica L.). Mycopath. 4: 35-38.
Heldt HW. 2005. Plant Biochemistry, 3rd
Edition. San Diego: Elsevier Academic
Press.
Huang LC, Lee YL, Huang BL, Kuo CI, Shaw JF. 2002. High polyphenol oxidase
activity and low titratable acidity in browning bamboo tissue culture. In
Vitro Cell Dev Biol Plant. 38(4): 358-365.
Hutami S. 2006. Penggunaan arang aktif dalam kultur in vitro. Berita Biologi
8(1): 83-89.
Hutami S. 2008. Masalah pencoklatan pada kultur jaringan. J Agro Biogen. 4(2):
83-88.

41
Jun-Yan Z, Fu-Xing G, Razdan MK. 2000. Somatic embryogenesis and
germplasm conservation of plant. Dalam: MK Razdan dan EC Cocking
(Editor). Conservation of Plant Genetic Resource In Vitro, Volume 2:
Aplication and Limitation, Science Publishers Inc.: Enfield. Hal: 167-192.
Kushairi A, AH Tarmizi, I Zamzuri, Ong-Abdullah M, Samsul KR, Ooi SE, N
Rajanaidu. 2010. Makalah pada International Seminar on Advance in Oil
Palm Tissue Culture, International Society for Oil Palm Breeders (ISOPB),
di Yogyakarta, Indonesia. 29 Mei 2010.
Loomis, WD. and Battail J. 1966. Plant phenolic compounds and the isolation of
plant enzym. Phytochem. 5: 423-438.
Lubis AU. 2008. Kelapa Sawit (Elaeis guineensis Jacq) di Indonesia, Edisi 2.
Medan: Pusat Penelitian Kelapa Sawit Marihat.
Mineo L. 2010. Plant tissue culture techniques. Dalam Tested Studies for
Laboratory Teaching. Vol 11. C A Goldman. Editor. Prosiding dari
Workshop/Konferensi dari Asscociation for Biology Laboratory Education
(ABLE).
NC-IUBMB. The Enzime List: Class 1–Oxidoreductases [terhubung berkala]
http://www.enzyme-database.org/ ( 21 Sept 2012).
Newton, RJ, Tang W, Harris LC, and Outhavong V. 2004. Antioxidants enhance in
vitro plant regeneration by inhibiting the accumulation of peroxidase in
Virginia pine (Pinus virginiana Mill.). Plant Cell Rep. 22(12):871-877.
Nhut D.T., N.T.M. Hanh, P.Q. Tuan, L.T.M. Nguyet, N.T.H. Tram, N.C. Chinh,
N.H. Nguyen, and D.N. Vinh. 2006. Liquid culture as a positive condition
to induce and enhance quality and quantity of somatic embryogenesis of
Lilium longiflorum. Sci Hort. 110(1):93-97.
Ozyigit II, MV Kahraman, and O Ercan. 2007. Relation between explant age, total
phenols and regeneration response in tissue cultured cotton (Gossypium
hirsutum L.). African J Biotechnol. 6(1):003-008.
Pahan I. 2006. Panduan Lengkap Kelapa Sawit: Manajemen Bisnis dari Hulu
hingga Hilir. Jakarta: Penebar Swadaya.
Rasool SN, Jahererunnisa S, Jayaveera KN, Suresh KC. 2011. In vitro callus
induction and in vivo antioxidant activity of Passiflora foetida L. leaves.
Intl J App Res Nat Prod. 4(1): 1-10.
Rodriguez R. 1982. Callus initiation and root formation from in vitro culture of
walnut cotyledons. Sci Hort. 17(20):195-196.
Saxena PK, Editor. 2001. Development of Plant-Based Medicines: Concervation,
Eficacy and Safety. Amsterdam: Kluwer Academic Publisher.

42
Singh N, Yadav K, Kumari S, Renu. 2011. Metabolic change during
differentiation in callus culture of Stevia Rebaudiana (Bertoni). J Phytol.
3(3): 63-67.
Tabiyeh, D.T., F. Bernard, and H. Shacker. 2006. Investigation of glutathione,
salicylic acid and GA3 effects on browning in Pistacia vera shoot tips
culture. ISHS Acta Hort. 726.
Murashige T, Skoog F. 1962. A revised medium for rapid growth and bioassays
with tobacco tissue cultures. Physiol Plant. 15: 473-497.
Vaughn K, Duke SO. 1984. Function of polyphenol oxidase in higer plants.
Physiol Plant. 60(1): 1275-1283
Yusnita. 2008. Kultur Jaringan: Cara Memperbanyak Tanaman secara Efisien.
Jakarta: Agromedia Pustaka.
Zulkarnain. 2009. Kultur Jaringan Tanaman: Solusi Perbanyakan Tanaman Budi
Daya. Bumi Aksara: Jakarta.

43
LAMPIRAN
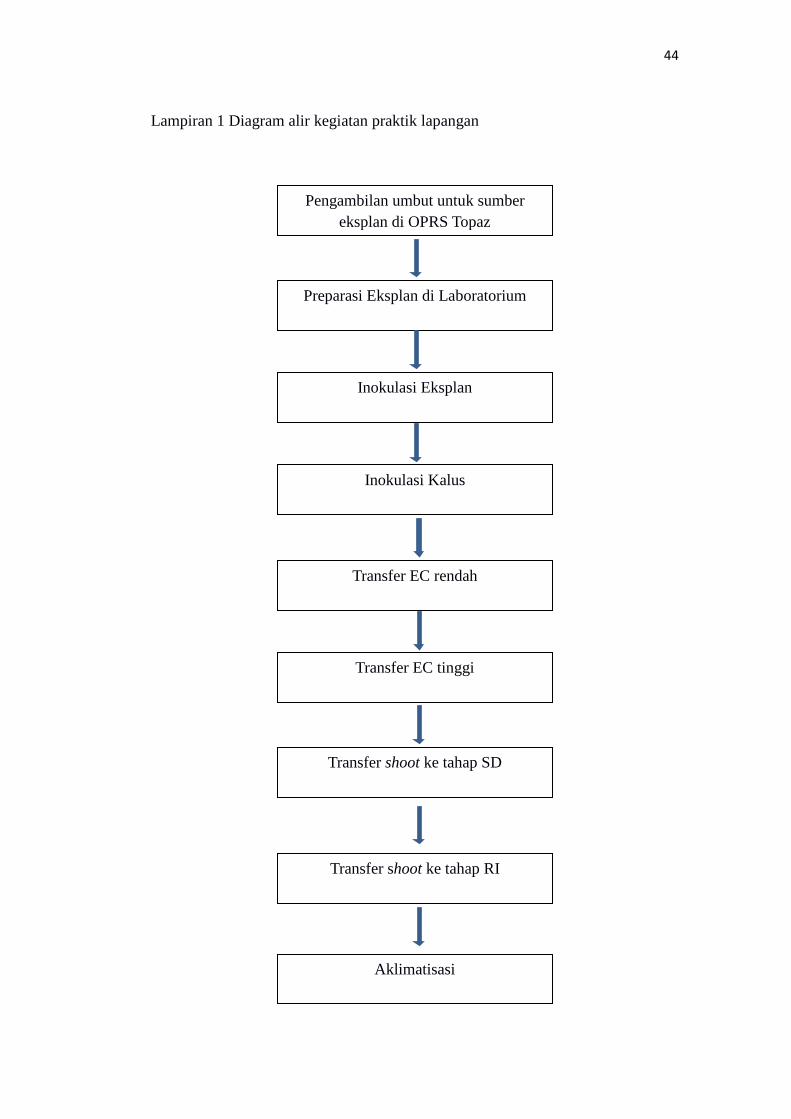
44
Lampiran 1 Diagram alir kegiatan praktik lapangan
Pengambilan umbut untuk sumber
eksplan di OPRS Topaz
Preparasi Eksplan di Laboratorium
Inokulasi Eksplan
Transfer EC rendah
Inokulasi Kalus
Transfer EC tinggi
Transfer shoot ke tahap SD
Aklimatisasi
Transfer shoot ke tahap RI

45
Lampiran 2 Struktur Organisasi RGE Group
RGE Group
PO&G Sateri RAPP
PT Tunggal Yunus Esatate
Asian Agri PechTech
Breeding Seed Production Unit
Marketing
Nursery & Extention COPPU
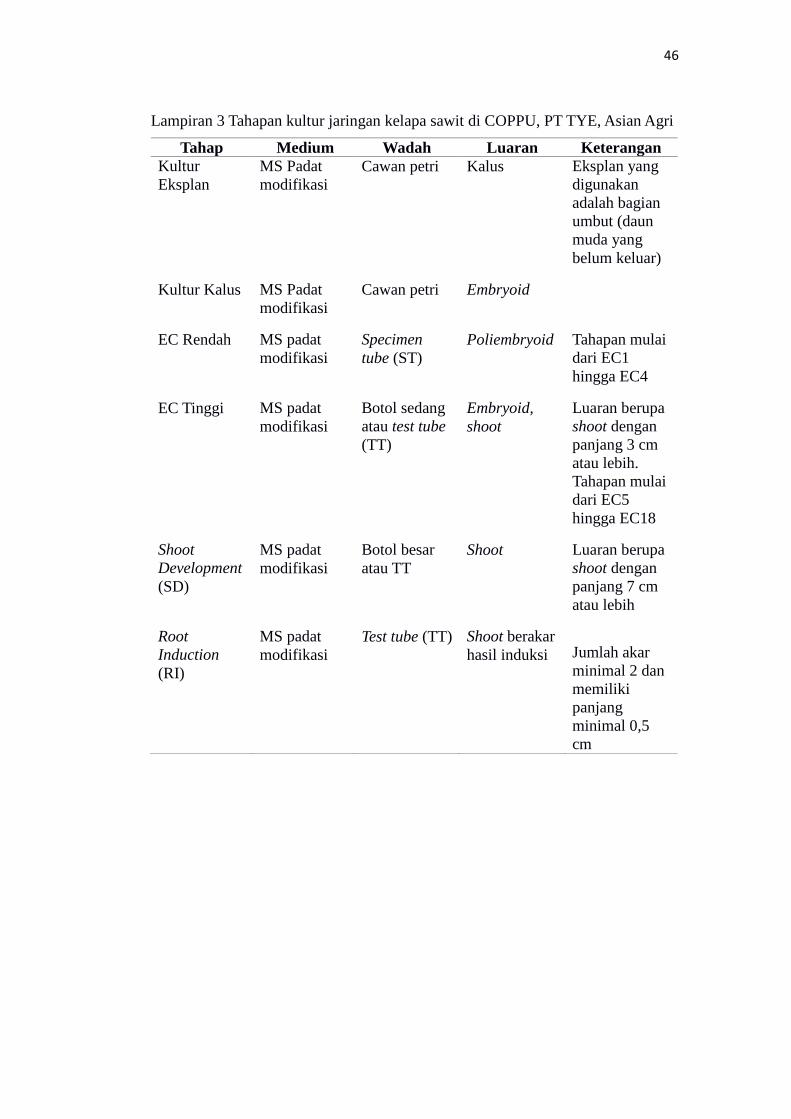
46
Lampiran 3 Tahapan kultur jaringan kelapa sawit di COPPU, PT TYE, Asian Agri
Tahap Medium Wadah Luaran Keterangan
Kultur
Eksplan
MS Padat
modifikasi
Cawan petri Kalus Eksplan yang
digunakan
adalah bagian
umbut (daun
muda yang
belum keluar)
Kultur Kalus MS Padat
modifikasi
Cawan petri Embryoid
EC Rendah MS padat
modifikasi
Specimen
tube (ST)
Poliembryoid Tahapan mulai
dari EC1
hingga EC4
EC Tinggi MS padat
modifikasi
Botol sedang
atau test tube
(TT)
Embryoid,
shoot
Luaran berupa
shoot dengan
panjang 3 cm
atau lebih.
Tahapan mulai
dari EC5
hingga EC18
Shoot
Development
(SD)
MS padat
modifikasi
Botol besar
atau TT
Shoot Luaran berupa
shoot dengan
panjang 7 cm
atau lebih
Root
Induction
(RI)
MS padat
modifikasi
Test tube (TT)
Shoot berakar
hasil induksi
Jumlah akar
minimal 2 dan
memiliki
panjang
minimal 0,5
cm

47
Lampiran 4 Hasil Kultur Kelapa Sawit dari Berbagai Proses Kultur Jaringan di
Laboratorium COPPU.
(1) (2) (3)
(4) (5) (6)
Keterangan :
(1) = Kultur kelapa sawit dalam tahap EC tinggi.
(2) = Kultur kelapa sawit dalam tahap EC rendah.
(3) = New Embroid.
(4) = Kultur kelapa sawit dalam tahap SD.
(5) = Kultur kelapa sawit tahap SD.
(6) = Kultur kelapa sawit tahap RI, terlihat sudah keluar akar.

48
Lampiran 4 lanjutan
(7) (8) (9)
Keterangan :
(7) = Kultur kelapa sawit yang terkena serangan jamur.
(8) = Kultur kelapa sawit yang terkena serangan bakteri.
(9) = Kultur kelapa sawit yang terkena serangan mites.
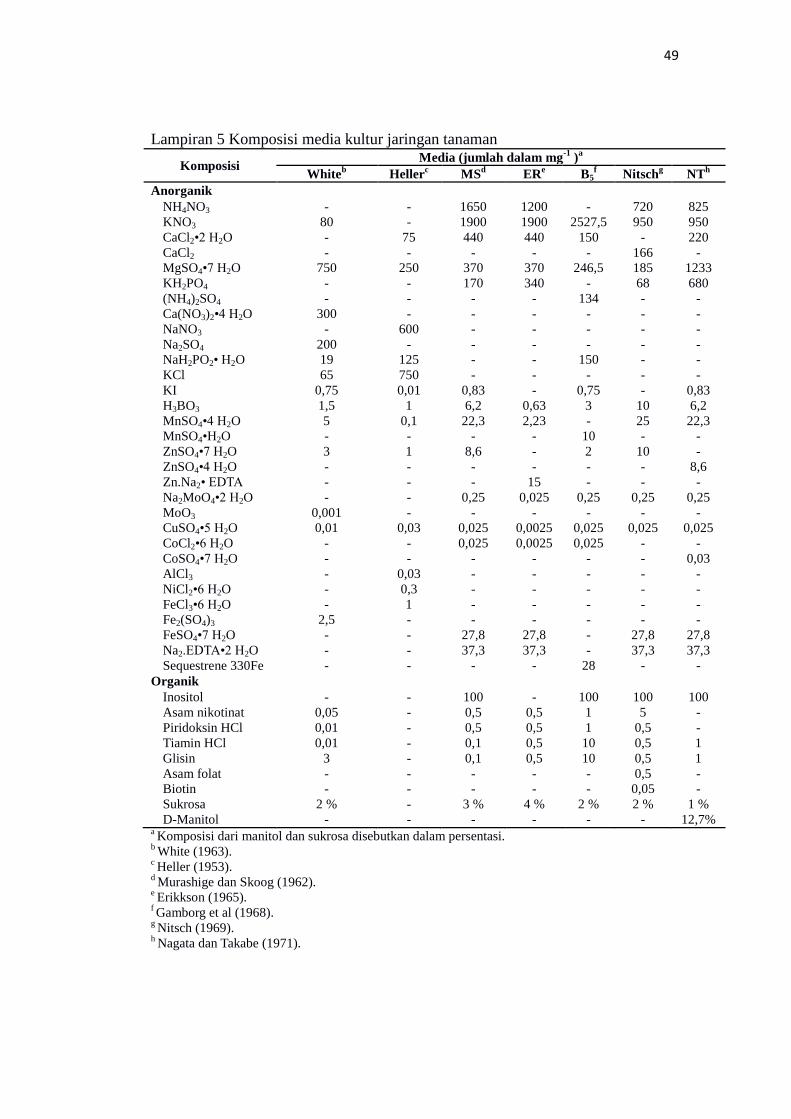
49
Lampiran 5 Komposisi media kultur jaringan tanaman
Komposisi Media (jumlah dalam mg
-1 )
a
Whiteb
Hellerc
MSd
ERe
B5f
Nitschg
NTh
Anorganik NH4NO3 - - 1650 1200 - 720 825
KNO3 80 - 1900 1900 2527,5 950 950
CaCl2•2 H2O - 75 440 440 150 - 220
CaCl2 - - - - - 166 -
MgSO4•7 H2O 750 250 370 370 246,5 185 1233
KH2PO4 - - 170 340 - 68 680
(NH4)2SO4 - - - - 134 - -
Ca(NO3)2•4 H2O 300 - - - - - -
NaNO3 - 600 - - - - -
Na2SO4 200 - - - - - -
NaH2PO2• H2O 19 125 - - 150 - -
KCl 65 750 - - - - -
KI 0,75 0,01 0,83 - 0,75 - 0,83
H3BO3 1,5 1 6,2 0,63 3 10 6,2
MnSO4•4 H2O 5 0,1 22,3 2,23 - 25 22,3
MnSO4•H2O - - - - 10 - -
ZnSO4•7 H2O 3 1 8,6 - 2 10 -
ZnSO4•4 H2O - - - - - - 8,6
Zn.Na2• EDTA - - - 15 - - -
Na2MoO4•2 H2O - - 0,25 0,025 0,25 0,25 0,25
MoO3 0,001 - - - - - -
CuSO4•5 H2O 0,01 0,03 0,025 0,0025 0,025 0,025 0,025
CoCl2•6 H2O - - 0,025 0,0025 0,025 - -
CoSO4•7 H2O - - - - - - 0,03
AlCl3 - 0,03 - - - - -
NiCl2•6 H2O - 0,3 - - - - -
FeCl3•6 H2O - 1 - - - - -
Fe2(SO4)3 2,5 - - - - - -
FeSO4•7 H2O - - 27,8 27,8 - 27,8 27,8
Na2.EDTA•2 H2O - - 37,3 37,3 - 37,3 37,3
Sequestrene 330Fe - - - - 28 - -
Organik Inositol - - 100 - 100 100 100
Asam nikotinat 0,05 - 0,5 0,5 1 5 -
Piridoksin HCl 0,01 - 0,5 0,5 1 0,5 -
Tiamin HCl 0,01 - 0,1 0,5 10 0,5 1
Glisin 3 - 0,1 0,5 10 0,5 1
Asam folat - - - - - 0,5 -
Biotin - - - - - 0,05 -
Sukrosa 2 % - 3 % 4 % 2 % 2 % 1 %
D-Manitol - - - - - - 12,7% a Komposisi dari manitol dan sukrosa disebutkan dalam persentasi.
b White (1963).
c Heller (1953).
d Murashige dan Skoog (1962).
e Erikkson (1965).
f Gamborg et al (1968).
g Nitsch (1969).
h Nagata dan Takabe (1971).