artikel-sudarko
TRANSCRIPT

5/10/2018 artikel-sudarko - slidepdf.com
http://slidepdf.com/reader/full/artikel-sudarko 1/14
MODIFIKASI ASETILKOLINESTERASE
DENGAN MUTASI KOMBINASI SECARA IN SILICO
UNTUK BIOSENSOR ORGANOFOSFAT
Sudarko, Devit Suwardiyanto, dan A.A. Istri Ratnadewi2)
Jurusan Kimia, FMIPA, Universitas Jember
ABSTRAK
Pengawasan lingkungan dari pestisida yang mempengaruhi kesehatan
manusia dan ekosistem, telah menjadi pusat perhatian karena penggunaan pestisida di dunia. Deteksi pestisida di lingkungan dapat dilakukan dengan biosensor yang menggunakan asetilkolinesterase. Dalam penelitian ini telahdilakukan modifikasi asetilkolinesterase Torpedo californica secara in silico(simulasi komputer). Modifikasi ini bertujuan untuk meningkatkan sensitifitas dankespesifikan terhadap organofosfat. Modifikasi asetilkolinesterase dilakukansecara in silico dengan program Modeller 6.2 dan untuk mengetahui efek modifikasi terhadap sensitifitas dan kespesifikannya digunakan programAutoDock 3.0. Strategi yang digunakan untuk memperoleh mutan yaitu dengankombinasi mutan tunggal yang diharapkan dapat meningkatkan sensitifitas enzimterhadap organofosfat. Dalam penelitian ini tidak dapat memperoleh
asetilkolinesterase yang lebih sensitif dan spesifik terhadap organofosfat.Kesulitan prediksi efek mutasi terhadap inhibisi organofosfat terjadi karenaadanya dua model pengikatan organofosfat yang dipengaruhi oleh konsentrasi.
Namun dengan strategi ini diperoleh mutan yang lebih sensitif dan spesifik terhadap karbamat. Hasil terbaik untuk peningkatan sensitifitas dan kespesifikankarbamat terjadi pada mutan F330A dengan peningkatan k i sebesar 0,42 untuk karbaril dan 0,31 untuk karbofuran.
Kata kunci: asetilkolinesterase, biosensor, insektisida, pemodelan homologi,docking .
1

5/10/2018 artikel-sudarko - slidepdf.com
http://slidepdf.com/reader/full/artikel-sudarko 2/14
MODIFICATION ACETYLCHOLINESTERASE WITH
COMBINATION MUTATION BY IN SILICO
FOR BIOSENSOR ORGANOPHOSPHATE
Sudarko, Devit Suwardiyanto, dan A.A. Istri Ratnadewi
Department of Chemistry, Faculty of Mathematics and Natural SciencesUniversity of Jember
ABSTRACT
To detect traces of insecticides in environment using biosensors, weengineered Torpedo c. acetylcholinesterase (AChE) by in silico to increase itssensitivity and spesifity to organophosphate. The mutant made by homology
program using MODELER 6.2 modules of TRITON 3.0. To show the effect of modification, we use AUTODOCK 3.0 program. Mutant ACHe was obtainsthrough the combination of several mutations that increase sensitivity toorganophosphate. Two types of insecticides that used in this research, isorganophosphate and carbamate. Carbamate will be use as comparator for lookingspecification AChE to organophosphate. In this research, AChE that moresensitive to organophosphate cannot be obtains. The difficulty of predicting the
effect mutation, because there are two binding models that influence byconcentration. Nevertheless, the use of these strategies allowed us to obtain AChEwhich more sensitive to carbamate. The best result that increase sensitivity andspecifity to carbamat are mutant F330A. Mutant F330A increase the inhibitionconstant about 0.42 for carbaryl and 0.31 for carbofuran.
Key words: Acetylcholinesterase, biosensor, insecticide, homology modeling,docking.
2

5/10/2018 artikel-sudarko - slidepdf.com
http://slidepdf.com/reader/full/artikel-sudarko 3/14
PENDAHULUAN
Pengawasan lingkungan dari pestisida yang mempengaruhi manusia dan
ekosistem, telah menjadi pusat perhatian. Organofosfat dan karbamat merupakan
pestisida yang banyak digunakan dan memiliki kemampuan untuk menggantikan
organoklorin seperti DDT, aldrin, lindane, dan lain-lain. Pestisida ini memiliki
persistansi lingkungan yang rendah dibanding organoklorin, tetapi memiliki
tingkat keracunan yang lebih tinggi (Kok et al., 2002).
Biosensor berdasar inhibisi pada aktifitas beberapa enzim telah digunakan
untuk pengukuran polutan dalam lingkungan (Kuswandi and Mascini, 2005). Hal
ini, karena sistem sensor tersebut telah mampu memberikan cara analisis suatu
polutan secara cepat, mudah dan handal pada jumlah renik. Dalam pengembangan
biosensor, penggunaan enzim sangat bermanfaat dan menjanjikan (Kuswandi and
Narayanaswamy , 1998; Wolfbies and Li, 1993).
Dari penelitian yang telah dilakukan Kovarik et al (2003) dapat diketahui
bahwa subtitusi residu F338A dan Y337A (penomoran berdasarkan
asetilkolinesterase tikus), menghasilkan peningkatan konstanta inhibisi
organofosfat sebesar dua kali lipat. Mutasi yang sama dapat dilakukan pada
asetilkolinesterase Torpedo c., yaitu pada F331 dan Y334 (Barril et al., 2001).
Modifikasi asetilkolinesterase Drosophila m. untuk deteksi insektisida
yang dilakukan oleh Boublik et al (2002), menunjukkan bahwa mutasi F330L,
Y370A, dan F371I dapat meningkatkan sensitifitas enzim terhadap organofosfat.
Subtitusi residu F371I menghasilkan sensitifitas yang lebih tinggi dibanding
subtitusi residu F371A. Mutasi yang sama dapat dilakukan pada
asetilkolinesterase Torpedo c., yaitu pada residu F290, Y330, dan F331.
Salah satu strategi untuk meningkatkan kinerja asetilkolinesterase untuk
biosensor organofosfat adalah dengan meningkatkan kespesifikan dan sensitifitas
enzim terhadap organofosfat. Pada penelitian ini dilakukan modifikasi sisi aktif
asetilkolinesterase secara in silico (simulasi komputer), selanjutnya untuk
mengetahui kespesifikan dan sensitifitas enzim hasil modifikasi tersebut
3

5/10/2018 artikel-sudarko - slidepdf.com
http://slidepdf.com/reader/full/artikel-sudarko 4/14
dilakukan uji interaksi antara enzim hasil modifikasi dengan substrat maupun
beberapa jenis inhibitor (heptenofos, diklorfos, karbofuran, karbaril).
METODE PENELITIAN
Penelitian ini dilaksanakan pada bulan Mei 2005 sampai Juni 2005 di
Jurusan kimia, Fakultas Matematika dan Ilmu Pengetahuan Alam, Universitas
Jember. Alat yang digunakan dalam penelitian ini antara lain; komputer Intel
Xeon 2,4 MHz dual prosessor dengan memori 512 MB dan system operasi Linux
Debian Sarge Testing yang dilengkapi dengan program aplikasi AutoDockTools,
Autodock 3.05 dan TRITON 3.0; serta komputer Intel Pentium 4 2,26 MHz
dengan memori 128 MB dan sistem operasi Microsoft Windows XP Pro, Linux
Debian Sarge Testing yang dilengkapi dengan program aplikasi AutoDockTools,
AutoDock 3.05, Pymol dan ChemOffice 2000.
Preparasi Ligand dan Protein
Ligand dibuat menggunakan ChemOffice 2000. Pertama, struktur 2D
ligand digambar dengan ChemDraw Ultra (modul ChemOffice 200). Kemudian,
struktur 2D diekspor ke struktur 3D dengan Chem3D. struktur 3D diminimalisasi
energinya dengan teori PM3 dalam MOPAC 7 (modul ChemOffice 200). Struktur
mutan dan wild type Torpedo c. dibuat dengan mutasi virtual dari pemodelan
homologi.
Pemodelan Homologi
Struktur X-ray tunggal digunakan sebagai template untuk pemodelan
homologi. Struktur X-ray diperoleh dari protein data bank (E105). Data struktur
dimanipulasi menggunakan MODELLER 6.2 yang terdapat pada TRITON 3.0.
MODELLER mengimplementasikan pendekatan pemodelan menggunakan
perbandingan struktur protein (Sali et al., 1993). Pemodelan dimulai dengan
penataan sequence yang akan dimodelkan (target) dengan struktur protein tiga
dimensi yang telah diketahui (cetakan). Setelah itu dilakukan penghitungan
batasan pada sequence target yang dihitung dari penataannya dengan struktur tiga
dimensi template. Batasan tersebut diperoleh dari analisis statistik hubungan
pasangan-pasangan protein homolog. Analisis ini berdasar pada database penataan
4

5/10/2018 artikel-sudarko - slidepdf.com
http://slidepdf.com/reader/full/artikel-sudarko 5/14
105 famili yang termasuk 416 protein yang telah diketahui strukrur tiga
dimensinya (Sali et al., 1994). Dengan penelusuran database, akan diperoleh tabel
mengenai berbagai korelasi, seperti korelasi antara jarak ekivalen Cα - Cα, atau
sudut dihedral dari dua protein yang berhubungan. Selanjutnya, batasan hubungan
dan term energi CHARM memperkirakan stereokimia yang cocok, dan
mengombinasikannya ke dalam fungsi objektif. Terakhir, model diperoleh dengan
mengoptimasi fungsi objektif dalam ruang kartesian (Sali et al., 2004).
Docking
AutoDock merupakan program yang dikembangkan untuk memprediksi
interaksi ligand dengan target biomakromolekul. Dalam AutoDock, ligand pada
awalnya diacak diluar protein, kemudian melakukan translasi, orientasi, dan
konformasi sampai sisi ideal ditemukan (Morris et al., 1998). Program
AUTOTORS menentukan ikatan yang mungkin berotasi pada molekul ligand.
Titik grid yang digunakan dalam penelitian ini sebesar 80x80 dengan spasi grid
3,75 Å yang dipusatkan pada sisi aktif serin. Energi potensial setiap titik grid
dihitung menggunakan program AUTOGRID. Kemudian program AutoDock
menghitung energi interaksi antara konformasi ligand fleksibel dan setiap titik grid melalui kombinasi algoritma pencarian. Tahap translasi yang digunakan
sebesar 0,2 Å dan tahap rotasi 5°. Posisi awal dan sudut dihedral dipilih secara
acak dan terakhir simulasi 150 docking dijalankan untuk memperoleh struktur
kompleks dari ligand yang secara statistik diterima.
AutoDock 3.0 dapat menggunakan algoritma genetik (AG) Lamarckian
sebagai fungsi pencarian untuk simulasi docking . Konsep AG berdasar pada
evolusi biologis. Dalam simulasi docking, penataan ligand didefinisikan dengansebuah set variabel keadaan yang mendefinisikan translasi, orientasi, dan
konformasi ligand. Dalam AG, setiap variabel sesuai dengan genotip dan
koordinat atom sesuai dengan fenotip. Dari gen tersebut, fenotip ligand dapat
dihitung energinya berdasar fungsi energi bebas.
AutoDock menggunakan AG sebagai metode pencarian global dan sebuah
pencarian lokal (PL) adaptive. Pencarian lokal diturunkan dari sebuah algoritma
oleh Solis dan Wets, dimana langkah-langkahnya ditentukan, dan konformasi baru
5

5/10/2018 artikel-sudarko - slidepdf.com
http://slidepdf.com/reader/full/artikel-sudarko 6/14
yang diperoleh dievaluasi dengan fungsi energi. Perubahan energi antara dua
konformasi dihitung seperti pada algoritma Monte Carlo. Jika perubahannya lebih
disukai, maka diterima dan sebalikknya jika tidak, maka algoritma akan
mengambil langkah yang berlawanan (Morris et al, 2001).
HASIL DAN PEMBAHASAN
Proses Mutasi in Silico
Mutasi kombinasi mutan tunggal pada asetilkolinesterase, yang meliputi
F288L, F290L, F330A, F331I, dan Y334A dilakukan dengan program
MODELLER. Proses tersebut membutuhkan waktu kurang lebih 40 menit untuk
tiap mutan. Kombinasi kelima macam mutan tunggal tersebut menghasilkan tiga
puluh dua macam mutan termasuk asetilkolinesterase asli (wild type) hasil
optimasi MODELLER.
Efek Mutasi Asetilkolinesterase pada Substrat
Asetilkolin sebagai substrat didocking ke semua mutan dan dibandingkan
hasilnya dengan asetilkolinesterase wild type. Proses perhitungan docking
membutuhkan waktu rata-rata delapan jam tiap perhitungan. Dari hasil docking
diketahui bahwa mutan tunggal dan kombinasinya meningkatkan k 1 substrat (data
hasil docking pada semua mutan tidak ditampilkan pada artikel ini).
Kombinasi mutan tunggal yang dilakukan dalam penelitian ini
menyebabkan bertambah besarnya ruang gorge asetilkolinesterase. Bertambahnya
ukuran ruang menyebabkan energi internal ligand berkurang dan substrat lebih
mudah ke sisi aktif, sehingga k 1 substrat meningkat. Pada beberapa mutan,
perbesaran ruang gorge menyebabkan perubahan orientasi substrat. Dari hasil
docking , beberapa mutan memilki probabilitas pengikatan substrat pada sisi
periperal.
Ikatan asetilkolin seperti pada gambar 1, distabilkan oleh kantung oksi
anion yang terdiri dari residu G118, G119, dan A201. Gugus asetil asetilkolin
terikat pada kantung oksi anion melalui ikatan hidrogen antara gugus karbonil
asetilkolin dengan gugus NH peptida. Gugus ekor kuarterner trimetilamonium
terikat pada sisi anionik yang terdiri dari residu W84, Y130 dan E199. W84 dan
6

5/10/2018 artikel-sudarko - slidepdf.com
http://slidepdf.com/reader/full/artikel-sudarko 7/14
Y130 berikatan dengan bagian kation melalui interaksi kation-π dan interaksi
hidrofobik, sedangkan E199 melalui interaksi elektrostatik.
Gambar 1. Model Pengikatan Substrat pada Sisi Aktif Serin
Dari data probabilitas orientasi substrat dapat dikelompokkan substrat
yang memiliki jumlah probabilitas orientasi substrat ke sisi aktif serin dengan
nilai lebih besar dari 50% disajikan pada gambar 2.
0.00
2.00
4.00
6.00
8.00
10.00
12.00
14.00
16.00
18.00
20.00
W i l d
T y p e F 2
8 8 L
F 3 3 0
A F 3
3 1 I
F 2 8 8 L / F
2 9 0 L
F 2 8 8 L / F
3 3 0 A
F 2 8 8 L / Y
3 3 4 A
F 2 9 0 L / F
3 3 0 A
F 2 9 0 L / F
3 3 1 I
F 2 9 0 L / Y
3 3 4 A
F 3 3 0
A / Y 3
3 4 A
F 3 3 1 I / Y
3 3 4 A
F 2 8 8 L / F
3 3 0 A
/ Y 3 3
4 A
F 2 8 8 L / F
3 3 1 I / Y 3 3
4 A
F 2 9 0 L / F
3 3 0 A
/ Y 3 3
4 A
F 2 9 0 L / F
3 3 1 I / Y 3
3 4 A
Mutan
k i
( 1 0 - 5 )
Gambar 2. Grafik K 1 Substrat dengan Probabilitas Lebih Besar dari50%
Efek Mutasi Asetilkolinesterase pada Inhibisi Organofosfat
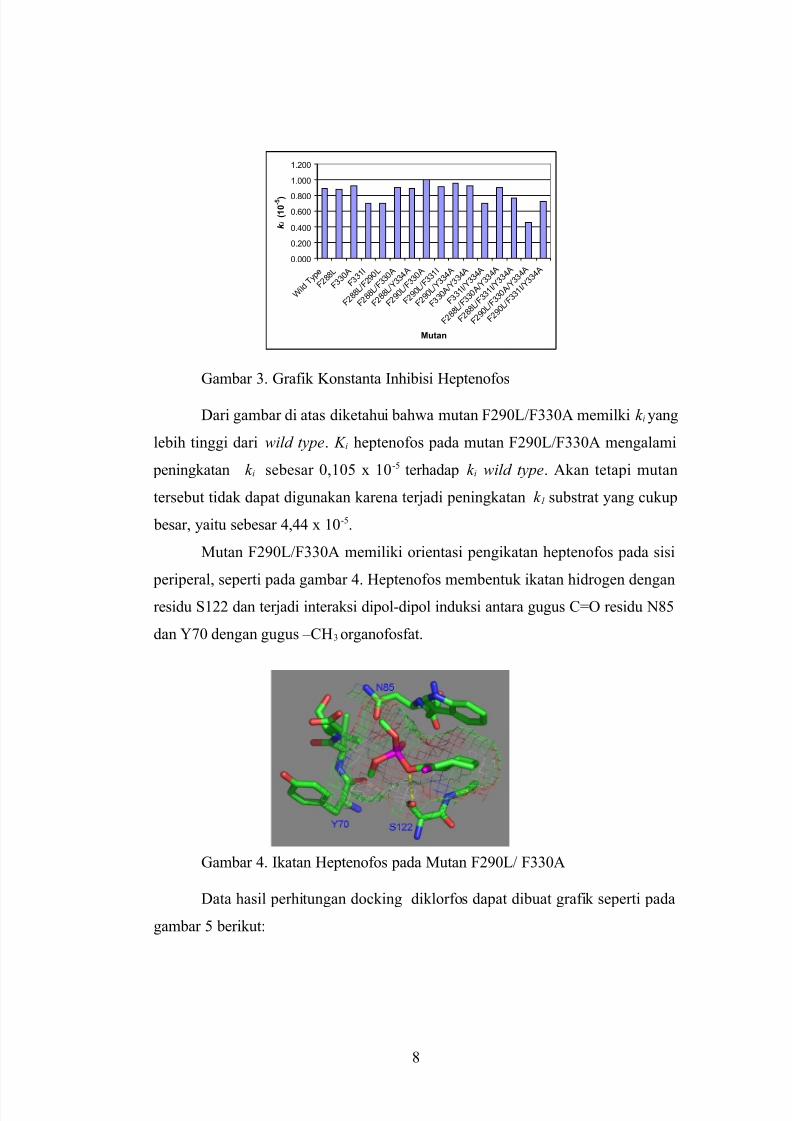
Data hasil perhitungan docking heptenofos dapat dibuat grafik seperti
pada gambar 3 (grafik hanya menampilkan mutan yang orientasi substratnya ke
sisi aktif serin).
7
5/10/2018 artikel-sudarko - slidepdf.com
http://slidepdf.com/reader/full/artikel-sudarko 8/14
0.000
0.200
0.400
0.600
0.800
1.000
1.200
W i l d
T y p e F 2
8 8 L
F 3 3 0
A F 3
3 1 I
F 2 8 8 L / F
2 9 0 L
F 2 8 8 L / F
3 3 0 A
F 2 8 8 L / Y
3 3 4 A
F 2 9 0 L / F
3 3 0 A
F 2 9 0 L / F
3 3 1 I
F 2 9 0 L / Y
3 3 4 A
F 3 3 0 A / Y 3
3 4 A
F 3 3 1 I / Y
3 3 4 A
F 2 8 8 L / F
3 3 0 A
/ Y 3 3
4 A
F 2 8 8 L / F
3 3 1 I / Y 3
3 4 A
F 2 9 0 L / F
3 3 0 A
/ Y 3 3
4 A
F 2 9 0 L / F
3 3 1 I / Y 3
3 4 A
Mutan
k i (
1 0 - 5 )
Gambar 3. Grafik Konstanta Inhibisi Heptenofos
Dari gambar di atas diketahui bahwa mutan F290L/F330A memilki k i yang
lebih tinggi dari wild type. K i heptenofos pada mutan F290L/F330A mengalami
peningkatan k i sebesar 0,105 x 10-5 terhadap k i wild type. Akan tetapi mutan
tersebut tidak dapat digunakan karena terjadi peningkatan k 1 substrat yang cukup
besar, yaitu sebesar 4,44 x 10-5.
Mutan F290L/F330A memiliki orientasi pengikatan heptenofos pada sisi
periperal, seperti pada gambar 4. Heptenofos membentuk ikatan hidrogen denganresidu S122 dan terjadi interaksi dipol-dipol induksi antara gugus C=O residu N85
dan Y70 dengan gugus –CH3 organofosfat.
Gambar 4. Ikatan Heptenofos pada Mutan F290L/ F330A
Data hasil perhitungan docking diklorfos dapat dibuat grafik seperti pada
gambar 5 berikut:
8

5/10/2018 artikel-sudarko - slidepdf.com
http://slidepdf.com/reader/full/artikel-sudarko 9/14
0
5
10
15
20
2530
35
40
45
W i l d
T y p e F 2
8 8 L
F 3 3 0
A F 3
3 1 I
F 2 8 8 L / F 2
9 0 L
F 2 8 8 L / F
3 3 0 A
F 2 8 8 L / Y
3 3 4 A
F 2 9 0 L / F
3 3 0 A
F 2 9 0 L / F
3 3 1 I
F 2 9 0 L / Y
3 3 4 A
F 3 3 0
A / Y 3
3 4 A
F 3 3 1 I / Y
3 3 4 A
F 2 8 8 L / F
3 3 0 A
/ Y 3 3
4 A
F 2 8 8 L / F
3 3 1 I / Y 3
3 4 A
F 2 9 0 L / F
3 3 0 A
/ Y 3 3
4 A
F 2 9 0 L / F 3
3 1 I / Y
3 3 4 A
Mutan
k i (
1 0 - 5 )
Gambar 5. Grafik Konstanta Inhibisi Diklorfos
Dari gambar 5 diketahui bahwa mutan F290L/F330A/Y334A memiliki k i
yang lebih tinggi dari wild type. Konstanta inhibisi diklorfos mutan
F290L/F330A/Y334A mengalami peningkatan sebesar 4,2 x 10-5 terhadap k i wild
type. Akan tetapi pada mutan tersebut juga terjadi peningkatan k 1 substrat yang
cukup besar, yaitu 65,75 x 10-5. Model pengikatan diklorfos pada mutan
F290L/F330A/Y334A seperti pada gambar 6.
Gambar 6. Ikatan diklorfos pada mutan F290L/F330A/Y334A
Dari hasil docking diperoleh probabilitas terbesar pengikatan organofosfat
terjadi pada sisi periperal dan hanya beberapa run (ulangan) yang menghasilkan
orientasi organofosfat ke sisi aktif. Pada asetilkolinesterase wild type yang
diinhibisi diklorfos, diperoleh tiga belas run dari 150 run yang menunjukkan
orientasi ligan ke sisi aktif. Sedangkan yang mengunakan inhibitor heptenofos
diperoleh dua belas run dari 150 run. Probabilitas pada mutan lainnya juga
menunjukkan kecenderungan pengikatan ke sisi periperal. Hal tersebut
9

5/10/2018 artikel-sudarko - slidepdf.com
http://slidepdf.com/reader/full/artikel-sudarko 10/14
menunjukkan bahwa organofosfat juga dapat langsung memfosforilasi sisi
katalitik tanpa berikatan dengan sisi periperal terlebih dahulu walaupun dengan
probabilitas yang kecil.
Secara keseluruhan, pemodelan inhibisi organofosfat pada
asetilkolinesterase dengan AutoDock tidak dapat memprediksi pengaruh mutasi.
Hal tersebut disebabkan inhibisi organofosfat memiliki lebih dari satu tahap
inhibisi dan dipengaruhi oleh konsentrasi. AutoDock memiliki keterbatasan tidak
dapat memprediksi model inhibisi yang dipengaruhi oleh konsentrasi. AutoDock
hanya dapat memprediksi probabilitas terbesar keadaan akhir model pengikatan
suatu ligand pada makromolekul.
Ada dua macam model inhibisi organofosfat, yaitu fosforilasi irreversibel
pada sisi aktif dan interaksi reversibel pada sisi periperal (Friboult et al , 1990).
Pengikatan organofosfat pada asetilkolinesterase juga dipengaruhi oleh
konsentrasi organofosfat. Pada konsentrasi rendah organofosfat menyerang sisi
periperal. Ketika sisi periperal telah jenuh, organofosfat akan menyerang sisi aktif
serin (Kardos and Sultatos, 2000).
Model kinetika inhibisi organofosfat pada sisi periperal asetilkolinesterase
dapat diilustrasikan pada gambar 7. Konstanta kecepatan k +1 dan k -1 menjelaskan
ikatan reversibel AB ke sisi periperal, dan dua titik sebelah kiri asetilkolinesterase
menunjukkan ikatan reversibel AB ke sisi periperal. k i’ menunjukkan perubahan
k i sebagai hasil adanya ikatan pada sisi periperal. Sedangkan k 3 menunjukkan
reaktifasi enzim (Kardos and Sultatos, 2000).
AB
ABAB
B
AB + AChE AChE-A AChE
+ +
AB + AB AChE AChE-A AChE
k i k 3
k -1k +1
k i' k 2
+
Gambar 7. Kinetika Inhibisi Organofosfat pada Asetilkolinesterase(Kardos and Sultatos, 2000).
Dengan mendocking ulang kompleks ligand yang telah terikat pada sisi
periperal asestilkolinesterase akan diperoleh kompleks organofosfat yang terikat
10

5/10/2018 artikel-sudarko - slidepdf.com
http://slidepdf.com/reader/full/artikel-sudarko 11/14
di sisi periperal dan sisi aktif sekaligus. Organofosfat kedua memiliki probabilitas
100% masuk ke sisi aktif asetilkolinesterase dengan nilai k i sebesar 1,31 x 10-5
untuk heptenofos dan 4,14 x 10-5 untuk diklorfos. Hal tersebut sesuai dengan
asumsi Kardos and Sultatos (2000) bahwa organofosfat dapat membentuk
kompleks AB--AChE-A.
Efek Mutasi Asetilkolinesterase pada Inhibisi Karbamat
Dari hasil docking karbaril dan karbofuran pada semua asetilkolinesterase
mutan dan wild type dapat dibuat grafik seperti pada gambar 8. Untuk
memperoleh mutan yang spesifik untuk karbamat, perlu dibandingkan konstanta
inhibisi karbaril dengan karbofuran. Mutan yang memiliki peningkatan konstanta
inhibisi yang hampir sama bersifat spesifik untuk karbamat.
0
0.5
1
1.5
2
2.5
3
W i l d
T y p e
F 2 8 8 L
F 3 3 0
A F 3
3 1 I
F 2 8 8 L / F
2 9 0 L
F 2 8 8 L / F
3 3 0 A
F 2 8 8 L / Y
3 3 4 A
F 2 9 0 L / F
3 3 0 A
F 2 9 0 L / F
3 3 1 I
F 2 9 0 L / Y
3 3 4 A
F 3 3 0
A / Y 3
3 4 A
F 3 3 1 I / Y
3 3 4 A
F 2 8 8 L / F
3 3 0 A
/ Y 3 3
4 A
F 2 8 8 L / F
3 3 1 I / Y 3
3 4 A
F 2 9 0 L / F
3 3 0 A
/ Y 3 3
4 A
F 2 9 0 L / F
3 3 1 I / Y 3
3 4 A
Mutan
k i (
1 0 - 5 )
Karbaril
Karbofuran
Gambar 8. Grafik Perbandingan Konstanta Inhibisi Karbarildengan Karbofuran
Jika dibandingkan dengan k 1 substrat maka mutan yang paling bagus
adalah F330A dan F331I. Pada mutan F331I k 1 karbaril meningkat dari 1,42x10-5
ke 1,78 x10-5 dan k 1 karbofuran meningkat dari 1,02x10-5 ke 1,45x10-5. Model
pengikatan karbaril pada mutan F331I ditunjukkan pada gambar 9. Atom oksigen
karbonil karbaril berikatan hidrogen dengan residu S200 dan gugus benzil
mengarah pada sisi anionik. Inhibisi karbofuran mengalami perubahan orientasi
ligan. Orientasi ligan berubah dari sisi esterase ke sisi periperal, seperti
ditunjukkan pada gambar 10. Perubahan tersebut disebabkan perbesaran ruang
11

5/10/2018 artikel-sudarko - slidepdf.com
http://slidepdf.com/reader/full/artikel-sudarko 12/14
kantung asil dan perubahan kepolaran residu 331. Subtitusi F331I menyebabkan
berkurangnya kepolaran residu tersebut.
Gambar 9. Inhibisi Karbaril pada Mutan F331I
Gambar 10. Inhibisi Karbofuran pada Mutan F331I
Subtitusi residu F330A menghasilkan k 1 yang hampir sama dengan mutan
F331I. Peningkatan k 1 ini sesuai dengan eksperimen secara in vitro yang dilakukan
oleh Boublik et al (2002) pada Drosophila m. Konstanta inhibisi karbaril
meningkat dari 1,42x10-5 ke 1,84 x10-5 dan konstanta inhibisi karbofuran
meningkat dari 1,02x10-5 ke 1,33x10-5. Mutan F330A bersifat lebih spesifik untuk
karbamat dibandingkan dengan mutan F331I. Seperti pada gambar 11 dan 12,
orientasi karbamat tetap pada sisi aktif serin dan tidak menyebabkan perubahan
yang besar pada aktifitas asetilkolinesterase terhadap substrat. Subtitusi F330A
tidak mempengaruhi ruang kantung asil. Gugus CH3 karbofuran berinteraksi
dengan gugus A330 melalui interaksi hidrofobik. Gugus benzil karbaril dan
karbofuran mengarah pada sisi anionik.
12

5/10/2018 artikel-sudarko - slidepdf.com
http://slidepdf.com/reader/full/artikel-sudarko 13/14
Gambar 11. Inhibisi Karbaril pada Mutan F330A
Gambar 12. Inhibisi Karbofuran pada Mutan F330A
KESIMPULAN DAN SARAN
Pengaruh modifikasi asetilkolinesterase terhadap inhibisi organofosfat
tidak dapat diprediksi menggunakan AutoDock 3.0 karena organofosfat memiliki
dua model pengikatan, yaitu pengikatan ligand pada sisi periperal dan sisi aktif
serin. Oleh sebab itu perlu dilakukan penelitian lebih lanjut untuk mempelajari
mekanisme inhibisi organofosfat.
Mutan yang spesifik untuk karbamat yaitu mutan F330A. Mutan F330A
memiliki konstanta inhibisi satu setengah kali lebih besar dibanding dengan
asetilkolinesterase wild type. Dari hasil penelitian ini perlu dilakukan eksperimen
lebih lanjut untuk memperoleh asetilkolinesterase yang lebih spesifik dan sensitif
untuk karbamat, yaiutu dengan subtitusi residu F330A.
Referensi
Barril, X., Kalko, S. G., Orozco, M. and Luque, F. J., 2002. Rational Design of Reversible Acetylcholinesterase Inhibitors. Mini Review in Medicinal
Chemistry. 2:27-36.
13

5/10/2018 artikel-sudarko - slidepdf.com
http://slidepdf.com/reader/full/artikel-sudarko 14/14
Boublik, Y., Aguet, P. S., Lougarre, A., Arnaud, M., Villatte, F., Mondacca, S. E.,and Fournier, D. 2002. Acetylcholinesterase engineering for detection of
insecticide residues. Protein Eng. 15: 43-50.Friboult, A., Rieger, F., Goudou, D., Amitai, G., and Taylor, P. 2000. Interaction
of an organophosphate with peripheral site on acetylcholinesterase. Biochemistry, 29: 914-920.
Kardos, S.A., and Sultatos, L.G. 2000. Interactions of the Organophosphates
Paraoxon and Methyl Paraoxon with Mouse Brain Asetilkolinesterase.Toxicological Sciences, 58: 118-126.
Kok, F.N, Bozoglu, F., and Hasirci, V. 2002. Constustion of an
Acethylcholinesterase-Choline oxidase biosensor for aldicarb detection.
Biosensors and Bioelectronics, 17: 531-539.Kovarik, Z., Radic, Z. Berman, H.A., Simeon-rudolf, V., Reiner, E., and Taylor,
P. 2003. Acetylcholinesterase Active Center and Gorge Conformation
Analised by Combinatorial mutations and Enantiomeric Phosphonates. Biochem.J. 373: 33-40.
Kuswandi, B. and Narayanaswamy. R. 1998. A Simple Optical Flow Injection Ammonia sensors. Anal.Lett , 31: 395-410.
Kuswandi, B and Mascini, M. 2005. Enzyme Inhibition Based Biosensor for Environmental Monitoring . Current Enzyme Inhibition, 1: 11-20. BenthamScience Publisher Ltd.
Morris, G. M., Goodsell, D. S., Halliday, R. S., Huey, R., Hart, W. E., Belew, R.K., and Olson, A. J., 1998. Automated Docking Using a Lamarkian
Genetic Algorithm and an Empirical Binding Free Energy Function. J.
Comp. Chem. 19: 1639-1662.
Morris, G. M., Goodsell, D. S., Huey, R., Hart, W. E., Halliday, S., Belew, R., andOlson, A. J. 2001. User’s Guide : AutoDock Version 3.0.5.
Sali, A. and Overington, J. P. 1994. Derivation of rules for comparative protein
modeling from a databse of protein structure aligments. Protein Science,3: 1582-1596.
Sali, Andrej. Webb, B., Madhusudhan, M.S., Shen, M.Y., Renom, M.A.M.,Eswar, N., Alber, F., Oliva, B., Fiser, A., Sanchez, R., Yerkovich, B.,Badretdinov, A., Melo, F., Overington, J.P., and Feyfant, E. 2004 Manual
: MODELLER (A Program for Protein Structure Modeling), Release 7v7.Wolfbies, O.S and Li, H.1993. Flourescence Optical Urea Biosensor with an
Ammonium Optrode as Transducer. Biosensors&Bioelectronics , 8: 161-166
14