aktivitas antibakteri ekstrak zingiber zerumbet asal … · senyawa-senyawa aktif terhadap enzim...
TRANSCRIPT

AKTIVITAS ANTIBAKTERI EKSTRAK Zingiber zerumbet
ASAL PULAU TIMOR dan SIMULASI DOCKING
INHIBISI SENYAWA-SENYAWA AKTIF
TERHADAP ENZIM MurA
ORIGENES BOY KAPITAN
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2016


PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Aktivitas Antibakteri
Ekstrak Zingiber zerumbet Asal Pulau Timor dan Simulasi Docking Inhibisi
Senyawa-Senyawa Aktif Terhadap Enzim MurA adalah benar karya saya dengan
arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada
perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya
yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam
teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut
Pertanian Bogor.
Bogor, Januari 2016
Origenes Boy Kapitan NIM G851130111


RINGKASAN
ORIGENES BOY KAPITAN. Aktivitas Antibakteri Ekstrak Zingiber zerumbet
Asal Pulau Timor dan Simulasi Docking Inhibisi Senyawa-Senyawa Aktif
Terhadap Enzim MurA. Dibimbing oleh LAKSMI AMBARSARI dan SYAMSUL
FALAH.
Zingiber zerumbet merupakan tanaman herbal yang secara tradisional
digunakan sebagai obat borok oleh penduduk di Pulau Timor. Tanaman ini tumbuh
secara liar. Dari studi literatur diketahui bahwa tanaman ini memiliki aktivitas
antimikroba. Secara empiris penggunaan rimpang tanaman oleh masyarakat di
Timor dalam mengobati luka borok menjadi menarik untuk dikaji. Kondisi wilayah
Pulau Timor yang kering dengan iklim semi arid menimbulkan dugaan adanya
aktivitas senyawa metabolit sekunder yang tinggi dalam menyembuhkan luka
borok. Luka borok lebih banyak disebabkan oleh kesalahan penangan luka dan juga
serangan bakteri. Tujuan dilakukannya penelitian ini adalah untuk menguji aktivitas
antibakteri dan mencari nilai KHTM dari ekstrak tanaman, mengetahui senyawa-
senyawa aktif yang terkandung pada ekstrak dengan aktivitas antibakteri terbaik,
dan mempelajari interaksi senyawa-senyawa tersebut dengan enzim MurA melalui
in silico molecular docking.
Senyawa-senyawa aktif dari Z. zerumbet diekstraksi menggunakan pelarut n-
heksan, etil asetat, etanol, dan air. Selanjutnya diuji aktivitas antibakteri masing-
masing ekstrak menggunakan metode difusi sumur terhadap bakteri Eschericia coli,
Pseudomonas aeruginosa, Staphylococcus aureus, dan Bacillus subtilis. Ekstrak
dengan aktivitas terbaik digunakan untuk mencari nilai konsentrasi hambat tumbuh
minumun (KHTM) terhadap bakteri. Ekstrak tersebut selanjutnya diuji secara
kualitatif kandungan metabolit sekundernya menggunakan metode Harborne yang
meliputi uji fenolik hidrokuinon, flavonoid, alkaloid, saponin, tanin dan
triterpenoid-steroid, serta dianalisis kandungan senyawa-senyawa aktif
menggunakan LCMS. Senyawa-senyawa tersebut selanjutnya digunakan sebagai
ligan uji dalam mempelajari penghambatan enzim MurA secara simulasi docking.
Hasil penelitian menunjukkan bahwa keempat ekstrak memiliki aktivitas
antibakteri dan ekstrak etanol memiliki aktivitas antibakteri terbaik. Nilai KHTM
untuk S. aureus, B. subtilis, E. coli dan P. aeruginosa berturut-turut adalah sebesar
50, 100, 150, 250 mg mL-1. Hasil uji fitokimia mengindikasikan ekstrak kasar etanol
rimpang tanaman mengandung senyawa-senyawa terpenoid, flavonoid, tanin dan
fenolik. Hasil analisis LCMS memperlihatkan bahwa ekstrak kasar etanol
mengandung senyawa zerumbon dan gingerglikolipid B. Energi afinitas (ΔGbinding)
hasil penambatan molekular untuk ligan alami (substrat), ligan obat (fosfomisin),
ligan zerumbon, dan ligan gingerglikolipid B, berturut-turut adalah senilai -10.1, -
4.7, -8.3, dan -8,4 kkal mol-1. Hasil simulasi docking menunjukkan bahwa terjadi
mekanisme reaksi kompetitif antara ligan uji zerumbon dengan substrat uridine-
diphosphate-N-asetylglucosamine pada enzim karena ligan uji menempati tempat
yang sama dengan substrat sehingga menghambat terbentuknya peptidoglikan
penyusun dinding sel bakteri.
Kata kunci : Gingerglikolipid B, Jahe Liar, Molecular Docking, Peptidoglikan,
Zerumbon

SUMMARY
ORIGENES BOY KAPITAN. Antibacterial Activity of Zingiber zerumbet Extracts
From Timor Island and Inhibition Docking Simulation of Active Compounds
Against MurA Enzyme. Supervised by LAKSMI AMBARSARI and SYAMSUL
FALAH.
Zingiber zerumbet is a medicinal plant which is traditionally used by
Timorese people to treat ulcerative lesions in Timor island. This plant grows wildly.
According to the literature, it is known that these plants have an antimicrobial
activity. Empirically, the use of rhizomes of plants by people in Timor to treat
ulcerative lesions is an interesting topic to study. The antibacterial activity of this
plant can be attributed to the presence of specific secondary metabolites which is
due to the dry and semi arid climate of Timor island region. Ulcers can be caused
by a fault treatment of injury and also bacterial attack. The objective of this study
was to determine the antibacterial activity and the minimun inhibitory concentration
(MIC) value of Z. zerumbet extract and to determine the active compounds
contained in the rhizome extract which had the highest antibacterial activity using
LC-MS and to study the interaction between the test compounds and MurA enzyme
through molecular docking simulations.
Initially, the active compounds of Z. zerumbet were extracted using a n-
hexane, ethyl acetate, ethanol and aqueous solvent. Antibacterial activity test was
done using the diffusion method wells of the four crude extracts of plants Z.
zerumbet against bacteria Escherichia coli, Pseudomonas aeruginosa,
Staphylococcus aureus, and Bacillus subtilis. Extract with the highest activity was
then used to find the MIC value against the bacteria. The extracts were also tested
to determine their secondary metabolite content using Harborne methods which
include test phenolic hydroquinone, flavonoids, alkaloids, saponins, tannins and
triterpenoids-steroid, and analyzed the content of the active compounds using
LCMS. The active compounds were used as ligands in studying the MurA enzyme
inhibition test in simulated docking.
It was found that ethanol extract had the highest antibacterial activity
compared to other extracts. The minimum inhibitory concentration (MIC) values of
ethanol extract against S. aureus, B. subtilis, E. coli, and P. aeruginosa were
respectively determined to be 50, 100, 150, 250 mg mL-1. The result of LC-MS
analysis showed that the most active antibacterial compound in the ethanol extract
were zerumbone and gingerglycolipid B. Energy affinity (ΔGbinding) result tethering
molecule to the natural ligand (substrate), a drug ligand (fosfomycin), zerumbon
ligand, and the ligand gingerglikolipid B, respectively is worth -10.1, -4.7, -8.3, and
-8.4 kcal mol -1. Further docking simulations indicated that there was a competitive
reaction mechanism between test ligand with an uridine-diphosphate-N-
asetylglucosamine substrate in the enzyme as a test ligand occupied the same place
as the substrate therefore inhibiting the formation of peptidoglycan, a major
constituent of bacterial cell wall.
Keywords: Gingerglycolipid B, Molecular docking, Peptidoglycan, Wild ginger,
Zerumbone

© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis ini dalam bentuk apa pun tanpa izin IPB


ORIGENES BOY KAPITAN
Tesis
sebagai salah satu syarat untuk memperoleh gelar
Magister Sains
pada
Program Studi Biokimia
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2016
AKTIVITAS ANTIBAKTERI EKSTRAK Zingiber zerumbet
ASAL PULAU TIMOR dan SIMULASI DOCKING
INHIBISI SENYAWA-SENYAWA AKTIF
TERHADAP ENZIM MurA

Penguji Luar Komisi pada Ujian Tesis : Dr I Made Artika, MAppSc


PRAKATA
Puji dan syukur penulis panjatkan kehadirat Tuhan Yang Maha Esa atas
segala rahmat dan karunia-Nya, sehingga penulis dapat menyelesaikan tesis dengan
baik yang berjudul Aktivitas Antibakteri Ekstrak Zingiber zerumbet Asal Pulau
Timor dan Simulasi Docking Inhibisi Senyawa-Senyawa Aktif Terhadap Enzim
MurA dengan total penelitian selama 7 bulan sejak bulan Desember 2014 sampai
Juli 2015 di Laboratorium Biokimia dan Laboratorium Bakteriologi, Divisi
Mikrobiologi Medik FKH IPB.
Penulis juga mengucapkan terima kasih kepada Dr. Dra. Laksmi Ambarsari,
MS selaku pembimbing pertama dan Dr. Syamsul Falah, S. Hut, M. Si sebagai
pembimbing kedua yang dengan sabar memberikan arahan, bimbingan, perhatian,
motivasi dan masukkan selama penelitian serta dalam penyusunan tesis.
Tidak lupa juga penulis mengucapkan terima kasih kepada keluarga (Bapa
David, Mama Martha, Ka Nadus dan Yati, Ka Meri, dan Ka Olivianus), sahabat
(Frans, Tirta, Ridho, Rini, Irma, Vida, Mb. Asih, Ka Fidelis, dan Ka Ocin), teman-
teman Laboratorium Biokimia dan Laboratorium Bakteriologi, teman-teman SPs
IPB program studi Biokimia 2013, dan teman-teman Gamanusratim untuk
dukungan yang tidak henti-hentinya kepada penulis.
Penyusunan tesis ini tentunya tidak terlepas dari kekurangan. Oleh karena itu,
penulis mengharapkan saran dan kritik untuk menyempurnakan penyusunan tesis
ini. Semoga hasil penelitian ini dapat bermanfaat bagi kemajuan ilmu pengetahuan.
Bogor, Januari 2016
Origenes Boy Kapitan

vii
DAFTAR ISI Hal
DAFTAR ISI vii
DAFTAR GAMBAR viii DAFTAR TABEL viii DAFTAR LAMPIRAN viii 1 PENDAHULUAN 1
Latar Belakang 1 Rumusan Masalah 2
Tujuan Penelitian 2 Manfaat Penelitian 2
2 METODE PENELITIAN 3 Bahan 3 Alat 3 Prosedur Penelitian 3
3 HASIL 6
Identifikasi Tumbuhan 6
Rendemen Ekstrak 6 Aktivitas Antibakteri Ekstrak N-Heksan, Etil Asetat, Etanol, dan Air 7 Konsentrasi Hambat Tumbuh Minimun Ekstrak Etanol 8
Hasil Uji Fitokimia 8 Analisis Kandungan Senyawa Ekstrak Etanol 9
Karakteristik Enzim MurA 9 Kestabilan Struktur Reseptor 11
Potensi Ligan Menurut Aturan Lipinski 11 Energi Afinitas (ΔGbinding) Penambatan Molekular 12 Interaksi Penambatan Molekul Ligan pada Enzim MurA 13
4 PEMBAHASAN 14 Identifikasi Tumbuhan dan Rendemen Ekstrak 14
Aktivitas Antibakteri Ekstrak N-Heksan, Etil Asetat, Etanol, dan Air dan
KHTM Ekstrak Etanol 15 Analisis Kandungan Senyawa Ekstrak Etanol 17 Simulasi Molecular Docking Ligan Senyawa Aktif Z. zerumbet Terhadap
Reseptor MurA 18 5 SIMPULAN dan SARAN 21
Simpulan 21 Saran 21
DAFTAR PUSTAKA 22 LAMPIRAN 25 RIWAYAT HIDUP 35

viii
DAFTAR GAMBAR
DAFTAR TABEL
DAFTAR LAMPIRAN
1 Tanaman Z. zerumbet: A. Rimpang; B. Batang; C. Daun; D. Bunga 6 2 Zona bening yang terbentuk pada ekstrak etanol tanaman Z. zerumbet
ketika diuji terhadap bakteri S. aureus 7 3 Karakteristik enzim Mur A (a) Struktur 3D kompleks ternary enzim
(b) Plot Ramachandran enzim 10 4 Struktur sekunder reseptor Mur A 10 5 Afinitas energi (ΔG) ligan alami, ligan fosfomisin (obat),
ligan zerumbon, dan ligan gingerglikolipid B terhadap enzim MurA 12
1 Persentasi rendemen ekstrak 7
2 Hasil uji aktivitas antibakteri 8 3 Hasil uji KHTM ekstak etanol Z. zerumbet 8 4 Hasil uji fitokimia ekstrak etanol Z. zerumbet 9 5 Struktur 2D ligan (substrat atau UDP-N-GluAc, fosfomisin, zerumbon,
dan gingerglikolipid B) 11
6 Hasil filter Lipinski antara ligan alami, ligan obat, dan ligan uji 12 7 Hasil interaksi penambatan molekuler antara ligan dengan
enzim MurA 13
1 Diagram alir penelitian 26
2 Hasil identifikasi tanaman 27
3 Ekstrak kasar n-heksan, etil asetat, etanol dan aquades tanaman Z. zerumbet 28 4 Profil dan kromatogram hasil analisis LCMS ekstrak etanol Z. zerumbet 28 5 Hasil analisis kromatogram LCMS m/z 219.12
menggunakan database HMDB 30
6 Hasil analisis kromatogram LCMS m/z 679.26
menggunakan database HMDB 30
7 Energi afinitas hasil penambatan ligan alami terhadap MurA 31
8 Energi afinitas hasil penambatan ligan obat fosfomisin terhadap MurA 31
9 Energi afinitas hasil penambatan ligan uji zerumbon terhadap MurA 32
10 Energi afinitas hasil penambatan ligan uji gingerglikolipid B
terhadap MurA 32
11 Visualisasi 3D kompleks substrat, fosfomisin
dan zerumbon pada enzim MurA 32
12 Visualisasi 3D kompleks substrat, fosfomisin
dan gingerglikolipid B pada enzim MurA 33
13 Visualisasi 2D interaksi antara substrat, fosfomisin, zerumbon,
dan gingerglikolipid B dengan residu enzim MurA 33
14 Visualisasi 2D kesamaan interaksi antara fosfomisin
dan zerumbon dengan residu substrat enzim MurA 34

1
1 PENDAHULUAN
Latar Belakang
Tanaman Zingiberaceae mempunyai sifat aromatik dan efek farmakologis
yang dipengaruhi oleh senyawa-senyawa metabolit sekunder pada rimpangnya
(Voravuthikuncai et al. 2006; Chen et al. 2008). Salah satu anggota Zingiberaceae
yang menjadi daya tarik bagi para peneliti dunia karena beragamnya potensi
pengobatan yang dikandung adalah Zingiber zerumbet (L.) Smith (Singh et al.
2012). Tanaman ini tersebar di wilayah Asia Selatan, Asia Tenggara, Pasifik dan
Oceania (Kress et al. 2002; Kress 2014). Di Indonesia, tanaman ini dikenal dengan
sebutan lempuyang dan tersebar di wilayah Sumatra, Jawa dan Kalimantan.
Tanaman ini tumbuh dan diduga tersebar secara merata di Pulau Timor serta hidup
secara liar. Secara empirik, masyarakat di Pulau Timor hanya memanfaatkan
tanaman ini dalam penyembuhan luka borok secara topikal, sementara itu di
beberapa daerah yang lain tanaman ini dimanfaatkan baik dalam kondisi ekstrak
segar maupun ekstrak rebusan sebagai pembersih darah, obat disentri, nyeri perut,
batu ginjal, sakit kuning, antelmintik, luka, obat borok, bisul, penurun bengkak, dan
gatal-gatal, serta pereda nyeri (Sinaga et al. 2011).
Voravuthikuncai et al. 2006 melaporkan bahwa Z. zerumbet yang berasal dari
Thailand diketahui efektif dalam menghambat bakteri S. aureus dengan nilai
konsentrasi hambat tumbuh minimum (KHTM) sebesar 0,79 mg mL-1. Sari et al.
(2013) melaporkan bahwa ekstrak segar Z. zerumbet asal Sumatra Barat
mempunyai aktivitas antimikrob dengan daya hambat berturut-turut sebesar 9,13
mm; 9,2 mm; dan 9,6 mm ketika diujikan pada mikrob uji S. aureus, E. coli, dan C.
albicans. Uji antimikrob ekstrak kasar etanol terhadap 30 jenis patogen dan 3 jenis
jamur menunjukkan bahwa tanaman Z. zerumbet asal wilayah Bangladesh
mempunyai kemampuan yang sangat potensial sebagai antimikrob dengan
kemampuan hambat sebesar 6-10 mm (Kader et al. 2011). Kemampuan aktivitas
antibakteri tanaman disebabkan oleh kandungan metabolit sekundernya yang
dipengaruhi oleh kondisi lingkungan tumbuh tanaman. Pulau Timor beriklim semi arid yang dicirikan oleh bulan-bulan kering yang
lebih lama dari bulan-bulan basahnya, sehingga tanahnya dipengaruhi oleh kondisi
kering, bersuhu tinggi dan berkelembaban rendah. Keadaan daerah yang beriklim
semiarid mengakibatkan kondisi tanamannya berada dalam keadaan cekaman
kekeringan dan suhu tinggi dengan intensitas penyinaran matahari lebih panjang.
Kondisi ini memungkinkan tanaman Z. zerumbet untuk mensintesis dan
mengakumulasikan senyawa aktif tertentu yang berbeda dengan daerah lain sebagai
respon terhadap cekaman kekeringan dan panas. Senyawa-senyawa tersebut diduga
memiliki aktivitas antibakteri, terlihat dari penggunaannya secara empiris dalam
menyembuhkan luka borok.
Kelangsungan hidup bakteri bergantung pada aktivitas enzim UDP-N-
acetylglucosamine enolpyruvyl transferase, EC 2.5.1.7 (MurA) (Skarzynski et al.
1996; Eschenburg et al. 2005). MurA mengkatalisis tahapan pertama biosintesis
peptidoglikan dinding sel bakteri dengan mengkondensasikan fosfoenolpiruvat
(PEP) dan UDP-N-asetilglukosamin (UDP-Glc-NAc) menjadi UDP-GlcNAc-
enolpiruvat. Tahapan ini tidak terjadi pada mamalia sehingga MurA menjadi target
dalam pengembangan obat antibakteri (Eschenburg et al. 2005). Obat komersial

2
yang bekerja menghambat inhibitor kerja enzim MurA adalah Fosfomisin
(Skarzynki et al. 1996). Fosfomisin diketahui memiliki kekurangan karena semakin
meningkatkan tingkat resistensi bakteri terhadap obat ini (Eschenburg et al. 2005),
sehingga perlu untuk dicari obat inhibitor kerja enzim yang baru.
Computer aided drug design (CADD) adalah metode pengembangan obat-
obatan berbasis ilmu komputasi (Scheneider dan Baringhaus 2008; Meng et al.
2011). Salah satu CADD yang digunakan adalah simulasi molecular docking.
Tujuan simulasi molecular docking adalah untuk memahami dan memprediksi
rekognisi molekuler (Scheneider dan Baringhaus 2008; Yanuar 2012). Molecular
docking telah sering digunakan dalam mempelajari interaksi obat dan reseptor
dengan memberikan prediksi orientasi ikatan atau kecocokan geometris yang paling
mungkin dan prediksi afinitas ikatan atau kecocokan energi dari kandidat ligan pada
protein target (Scheneider dan Baringhaus 2008; Meng et al. 2011; Yanuar 2012).
Simulasi docking juga dapat digunakan untuk memperoleh mekanisme kerja suatu
senyawa kimia atau makromolekul dalam skala molekuler sehingga dimungkinkan
untuk mendesain obat berbasis struktur ligan (Vijesh et al. 2013).
Rumusan Masalah
Secara empiris penggunaan rimpang Zingiber zerumbet oleh masyarakat di
Timor dalam mengobati luka borok menjadi menarik untuk dikaji. Kondisi wilayah
Pulau Timor yang kering dengan iklim semi arid membuat peneliti menduga adanya
aktivitas senyawa-senyawa kimia aktif (metabolit sekunder) yang tinggi dalam
menyembuhkan luka borok. Luka borok lebih banyak disebabkan oleh kesalahan
penangan luka dan juga serangan bakteri. Sebelum mengisolasi senyawa yang
berperan dalam penyembuhan luka borok, dilakukan uji aktivitas antibakteri
terlebih dahulu dan dilanjutkan dengan mempelajari mekanisme penghambatan
kerja enzim Mur A oleh senyawa-senyawa kimia aktif dari Z. zerumbet secara in
silico.
Tujuan Penelitian
Tujuan dilakukannya penelitian ini adalah menguji aktivitas antibakteri dan
mencari nilai konsentrasi hambat tumbuh minumun dari ekstrak n-heksan, etil
asetat, etanol, dan air tanaman Z. zerumbet terhadap bakteri Eschericia coli,
Pseudomonas aeruginosa, Staphylococcus aureus, dan Bacillus subtilis. Selain itu,
penelitian ini juga dilakukan untuk mengetahui senyawa aktif yang terkandung
pada ekstrak Z. zerumbet, dan mempelajari interaksi senyawa tersebut dengan
enzim MurA melalui in silico molecular docking.
Manfaat Penelitian
Manfaat yang dapat diperoleh dari penelitian ini adalah dapat berkontribusi
pada bidang farmasi mengenai informasi akan kandungan senyawa-senyawa aktif
Z. zerumbet dan potensi hambatnya terhadap bakteri penginfeksi luka, sehingga
dapat dikembangkan sebagai salah satu kandidat obat antibakteri baru.

3
2 METODE PENELITIAN
Waktu dan Tempat Penelitian
Penelitian dilakukan pada bulan Desember 2014 sampai Juli 2015 di
Laboratorium Biokimia, Departemen Biokimia IPB dan Laboratorium
Bakteriologi, Divisi Mikrobiologi Medik FKH IPB.
Bahan
Bahan yang digunakan dalam penelitian ini adalah rimpang Zingiber
zerumbet segar yang dikoleksi pada bulan Juni dari wilayah Amarasi, Kabupaten
Kupang, Nusa Tenggara Timur. Untuk bakteri uji digunakan bakteri Eschericia
coli, Pseudomonas aeruginosa, Staphylococcus aureus dan Bacillus subtilis. Bahan
pendukung yang digunakan antara lain aquadest, alkohol 70%, fosfomyacin, n-
heksana, etil asetat, etanol 96%, dimetilsulfoksida (DMSO), media Potato Dextrose
Agar (PDA), media Nutrient Agar (NA), Nutrient Broth (NB), dan media Mueller
Hinton Agar (MHA), file berekstensi Fasta, PDB, PDBQT, struktur kimia enzim
MurA, struktur kimia zerumbon, struktur kimia gingerglikolipid B, struktur kimia
fosfomisin, ADT 1.5.6, VMD 1.9.2, Marvin Sketch, Discovery Studio V3.5,
LigPlot+ 4.5.3, dan Autodock Vina.
Alat
Alat yang digunakan adalah timbangan digital (ACIS), blender, magnetic
stirrer, rotavapor, cawan petri, tabung reaksi, Erlenmeyer, pipet, gelas ukur, batang
pengaduk, jarum ose, vortex mixer XH-D, autoklaf (N-Biotech. Inc), inkubator
(Firlabo), shaker (N-Biotech. Inc), LCMS (Qmicro QAA 642), jangka sorong
(Varnier), pipet mikroliter (Gilsong), laminar air flow (ESCO), pembakar Bunsen,
perforator, Laptop (Intel core i3, 2GB RAM ddr2, 250 seagate HDD, dan VGA ati
radeon x1200).
Prosedur Penelitian
Preparasi dan Ekstraksi Sampel
Sampel diidentifikasi oleh Bidang Botani, Pusat Penelitian Biologi, LIPI-
Bogor. Ekstraksi rimpang Z. zerumbet menggunakan metode maserasi. Sampel
dihaluskan dengan digiling menggunakan blender, selanjutnya diayak
menggunakan ayakan berukuran 40 mesh. Sebanyak 50 gram simplisia diekstraksi
mengunakan pelarut n-heksana, etil asetat, dan etanol masing-masing sebanyak 150
mL selama 24 jam pada suhu ruang, sedangkan untuk pelarut akuades dilakukan
pemanasan dengan suhu = 80oC selama 2 jam. Filtrat hasil maserasi disaring dan
dipekatkan dengan rotavapor vakum pada suhu 60oC.
Uji Aktivitas Antibakteri (Kader et al. 2011)
Uji aktivitas antibakteri dilakukan dengan metode difusi agar (sumuran).
Bakteri-bakteri patogen Gram negatif (E. coli dan P. Aeruginosa) dan Gram positif
(S. aureus dan B. subtilis) diperoleh dari koleksi Laboratorium Bakteriologi
Fakultas Kedokteran Hewan IPB. Inokulan bakteri ditumbuhkan pada media

4
Nutrien Agar (NA) (OxoidTM). Biakan bakteri kemudian diencerkan dengan NaCl
0,85% menggunakan metode McFarland 0.5 (setara dengan 108 CFU mL-1).
Sebanyak 0.1 mL masing-masing suspensi bakteri yang telah diencerkan kemudian
dicampurkan ke dalam 20 mL media Mueller-Hinton Agar (MHA) (OxoidTM)
suhu ± 45oC. Setelah memadat, media dilubangi dengan cork borrer berdiameter
5.3 mm. Sebanyak 50 μL dari masing-masing ekstrak dimasukkan ke dalam
sumuran. Uji aktivitas antibakteri masing-masing ekstrak dilakukan pada
konsentrasi 500 mg mL-1. Fosfomisin 0.4 mg mL-1 digunakan sebagai kontrol
positif dan akuades steril sebagai kontrol negatif. Selanjutnya diinkubasi pada suhu
37oC selama 24 jam. Zona bening yang terlihat di sekeliling lubang menjadi
petunjuk adanya aktivitas antibakteri dan merupakan zona hambat dari ekstrak
tanaman Z. zerumbet. Zona bening tersebut kemudian diukur menggunakan jangka
sorong.
Ekstrak yang menunjukkan aktivitas terbaik kemudian diukur nilai
konsentrasi hambat tumbuh minimum (KHTM). Pengujian KHTM dilakukan
menggunakan metode difusi agar dengan prosedur sama seperti yang telah
dijelaskan sebelumnya. Penentuan KHTM dilakukan dengan membuat konsentrasi
yang bervariasi yaitu 5, 10, 25, 50, 100, 150, dan 250 mg mL-1.
Penapisan Kandungan Fitokimia (Harborne 1996)
Ekstrak dengan aktivitas antibakteri terbaik selanjutnya diuji fitokimia untuk
mengetahui secara kualitatif kandungan senyawa bioaktifnya. Uji ini meliputi uji
fenolik hidrokuinon, flavonoid, alkaloid, saponin, tanin dan triterpenoid-steroid.
Semua uji fitokimia dilakukan menurut Harbone (1996). Senyawa fenolik
hidrokuinon diuji menggunakan pereaksi FeCl3. Terbentuknya warna hijau atau
hijau biru menunjukkan adanya senyawa fenol dalam ekstrak. Senyawa
triterpenoid-steroid diuji menggunakan reagent Lieberman Burchard. Reaksi
campuran akan menghasilkan warna merah untuk adanya triterpenoid, warna hijau
atau biru untuk adanya steroid, sedangkan warna ungu menunjukkan adanya
senyawa triterpenoid dan steroid. Senyawa alkaloid diuji menggunakan reagent
Mayer, Dragendrof, dan Wagner secara berturut-turut yang akan memberikan
endapan berwarna putih, merah, dan coklat bila positif mengandung senyawa
tersebut. Senyawa golongan saponin diuji dengan memanaskan ekstrak dalam
akuades, kemudian dikocok. Adanya saponin diindikasikan dengan terbentuknya
busa. Tanin diuji dengan memanaskan ekstrak dalam akuades kemudian campuran
disaring. Filtrat direaksikan dengan FeCl 1% (w/v). Keberadaan tanin diindikasikan
oleh terbentuknya warna hijau gelap atau biru gelap. Senyawa flavonoid diuji
dengan memanaskan ekstrak dalam akuades dan disaring. Filtrat kemudian
direaksikan dengan H2SO4 10% (w/v). Adanya flavonoid ditandai oleh
terbentuknya warna merah, kuning, atau jingga pada lapisan amil alkohol.
Analisis Kandungan Senyawa Ekstrak teraktif
Analisis kandungan senyawa yang terkandung pada ekstrak teraktif dilakukan
menggunakan Liquid chromatography mass spectra (LCMS) di Laboratorium
Kesehatan Daerah DKI Jakarta. Spesifikasi instrumen yang digunakan adalah
Qmicro QAA 642, dengan ion mode adalah ES+. Pelarut yang digunakan adalah
metanol dan air dengan perbandingan 10:90 (v/v) dengan laju alir 0.20 mL mnt-1,
dan temperatur kolom sebesar 40oC.

5
Analisis kromatogram massa molekul ion puncak senyawa-senyawa terkandung
dilakukan menggunakan database dari The Human Metabolome Database (HMDB).
Senyawa-senyawa kimia hasil analisis kemudian digunakan sebagai ligan uji dalam
uji simulasi pendaratan molekul terhadap enzim MurA.
Penambatan Molekuler Senyawa Aktif Terhadap Enzim MurA Secara In-
Silico (Kumar et al. 2012)
Tahap awal untuk melakukan simulasi docking adalah menentukan reseptor
dan ligan yang akan diuji. Reseptor yang digunakan adalah enzim MurA. Protein
target atau reseptor diperoleh dengan mendownload file pdb enzim dari pdb bank
melalui situs www.rscb.org/pdb. Proses optimasi geometri reseptor dilakukan
dengan perangkat lunak AutoDock Tools (ADT) 1.5.6. Tahap awal yang dilakukan
adalah penghilangan molekul air (H2O) disekitar protein, hetero atom dan ligan
alami. Selanjutnya adalah penambahan muatan Gasteiger dan Hidrogen. File
disimpan dalam format PDBQT. Kestabilan struktur dapat dilihat dengan VMD
1.9.2 melalui diagram plot Ramachandran.
Struktur ligan diperoleh dengan menggambar struktur dua dimensi (2D) dan
tiga dimensi (3D) menggunakan piranti lunak Marvin Sketch. Ligan yang akan
digunakan dirancang menggunakan perangkat lunak Marvin sketch dengan format
penyimpanan PDB. Proses optimasi dilakukan menggunakan ADT 1.5.6. Tahap
awal adalah dilakukan merged nonpolar hidrogen, memberikan muatan Gasteiger
dan semua file disimpan dalam format PDBQT. Selanjutnya dilakukan filter
menurut aturan Lipinski untuk semua senyawa ligan menggunakan online access
http://www.scfbio-iitd.res.in/ software/ utility/ lipinskifilters.jsp (Lipinski 2009).
Selanjutnya dilakukan penambatan molekuler yang dimulai dengan proses Grid dan
validasi parameter penambatan dilakukan dengan ADT 1.5.6, dan dilanjutkan
dengan penambatan molekular yang dilakukan menggunakan AutoDock Vina
(Scripps Research Institute, USA) dan di asumsikan semua rotatable bond (ikatan
siklik) dari ligan dapat berotasi (fleksibel) dan makromolekulnya adalah tetap
(rigid). Parameter berikut adalah parameter yang telah divalidasi. Ukuran grid box
yang dipilih adalah 80 x 80 x 80 Å dengan spacing centered 0.375 Å pada site sisi
aktif, center_x = 36.021, center_y = 19.91, center_z = 44.388 dan luas box
melingkupi keseluruhan struktur MurA. Exaustiveness di atur pada angka 250.
Letakkan folder vina di C:\Vina kemudian isikan file CONF.TXT dengan parameter
diatas sesuai dengan angka center dan size nya. Eksekusi perintah penambatan
menggunakan command pada jendela CMD, panggil Vina.exe lalu ketik perintah
“C:\vina --config conf.txt --log log.txt”, lalu tekan enter dan tunggu proses sampai
selesai lalu keluarkan num-mode 20. Ligan dalam kompleks MurA yang memiliki
energi bebas Gibbs (∆Gbinding) terkecil dari daftar disimpan dalam format PDBQT
(ADT 1.5.6) dan dikonversi ke format PDB dengan DSV 3.5. Hasil penambatan di
analisis energi bebas Gibbs (∆G), ikatan hidrogen, RMSD, residu yang berikatan,
gaya Van der waals dan gugus fungsi ligan menggunakan Ligplot+ 4.5.3 dengan
format .pdb untuk visualisasi 2D, DSV 3.5 untuk analisis gaya Van der waals dan
mode ikatan dengan DNA serta VMD digunakan untuk visualisasi struktur 3D.

6
3 HASIL
Identifikasi Tumbuhan
Berdasarkan hasil identifikasi, identitas sampel tumbuhan adalah Zingiber
zerumbet (L.) Sm., famili Zingiberaceae. Morfologi tanaman ini ditampilkan pada
Gambar 1. Z. zerumbet merupakan tanaman semak berumur tahunan (terna
parennial). Tanaman ini tumbuh di daerah dataran rendah sampai ketinggian 1200
m di atas permukaan laut. Berdasarkan klasifikasi botaninya, tanaman ini termasuk
ke dalam:
Divisi : Spermatophyta
Subdivisi : Angiospermae
Kelas : Monocotyledonae
Bangsa : Zingiberales
Suku : Zingiberaceae
Genus : Zingiber
Spesies : Zingiber zerumbet
A B C D
Gambar 1 Tanaman Z. zerumbet: A. Rimpang; B. Batang; C. Daun; D. Bunga
Rendemen Ekstrak
Ekstrak adalah sedian yang diperoleh dengan mengekstraksi zat aktif dari
simplisia menggunakan pelarut yang sesuai dan selanjutnya semua pelarut diuapkan
sehingga tersisa massanya. Teknik ekstraksi yang digunakan dalam penelitian ini
adalah maserasi yaitu simplisia bahan direndam dalam larutan pengekstrak yang
sesuai selama 24 jam pada suhu ruang dan pada suhu 80oC. Penarikan senyawa
dapat terjadi karena memanfaatkan sifat bahan sehingga senyawa larut dalam
kepolaran larutan pengekstrak yang sesuai. Ini dikenal dengan prinsip like disolved
like.
Rendemen ekstrak merupakan perbandingan antara bobot ekstrak yang
dihasilkan dengan bobot sampel awal yang diekstrak. Persentasi rendemen ekstrak
pada Tabel 1, memperlihatkan kemampuan pelarut air lebih besar dalam
mengekstraksi komponen aktif tanaman Z. zerumbet, diikuti oleh pelarut etanol, etil
asetat dan N-heksana. Rendemen ekstrak air yang lebih besar kemungkinan

7
dipengaruhi oleh polaritasnya. Polaritas relatif pelarut dapat dilihat dari nilai
konstanta dielektrik masing-masing pelarut.
Tabel 1 Persentasi rendemen ekstrak
Pelarut Bobot sampel (g) Bobot Ekstrak (g) Rendemen (%b/b)
n-Heksana 200 6.30 3.15
Etil Asetat 200 7.60 3.80
Etanol 100 6.20 6.20
Air 150 10.4 6.90
Aktivitas Antibakteri Ekstrak N-Heksan, Etil Asetat, Etanol, dan Air
Pengujian aktivitas antibakteri merupakan teknik untuk mengukur berapa
besarnya potensi atau konsentrasi suatu senyawa dapat memberikan efek toksik
bagi bakteri. Ekstrak Z. zerumbet dinyatakan mempunyai aktivitas antibakteri jika
terbentuk zona bening di sekeliling sumuran berisi ekstrak yang ditumbuhkan pada
media yang telah diinokulasi oleh bakteri (Gambar 2).
Keterangan: A: Konsentrasi ekstrak uji sebesar 250 mg mL-1; B. Konsentrasi ekstrak sebesar 150
mg mL-1; C. Konsentrasi ekstrak sebesar 100 mg mL-1; D. Konsentrasi ekstrak sebesar
50 mg mL-1; E. Konsentrasi ekstrak sebesar 25 mg mL-1; F. Konsentrasi ekstrak sebesar
10 mg mL-1; G. Konsentrasi ekstrak sebesar 5 mg mL-1; H. Kontrol positif; I. Kontrol
negatif
Gambar 2 Zona bening yang terbentuk pada ekstrak etanol tanaman Z. zerumbet
ketika diuji terhadap bakteri S. aureus
Diameter hambat merupakan ukuran kekuatan hambatan substansi
antibakteri. Lebarnya diameter zona bening yang terbentuk ditentukan oleh
konsentrasi senyawa yang menjadi dasar pengujian kuantitatif dan
mengindikasikan bahwa senyawa tersebut bisa bebas berdifusi ke seluruh medium.
Hasil uji aktivitas antibakteri ekstrak Z. zerumbet dapat dilihat pada Tabel 2.
Hasil uji menunjukkan bahwa ekstrak etanol memiliki diameter hambat yang
lebih besar dibanding ekstrak n-Heksan, etil asetat, dan air. Ekstrak etanol terlihat
lebih menghambat pada bakteri Gram positif (B. subtilis dan S. aureus) dibanding
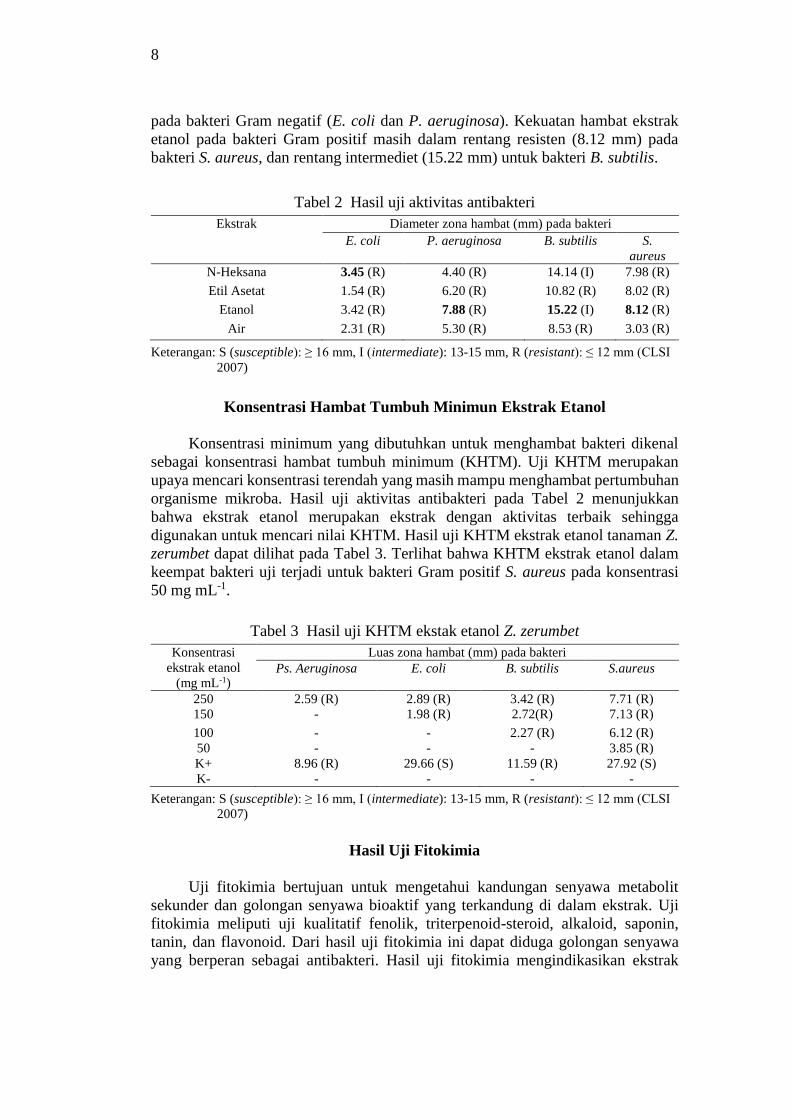
8
pada bakteri Gram negatif (E. coli dan P. aeruginosa). Kekuatan hambat ekstrak
etanol pada bakteri Gram positif masih dalam rentang resisten (8.12 mm) pada
bakteri S. aureus, dan rentang intermediet (15.22 mm) untuk bakteri B. subtilis.
Tabel 2 Hasil uji aktivitas antibakteri
Ekstrak Diameter zona hambat (mm) pada bakteri
E. coli P. aeruginosa B. subtilis S.
aureus
N-Heksana 3.45 (R) 4.40 (R) 14.14 (I) 7.98 (R)
Etil Asetat 1.54 (R) 6.20 (R) 10.82 (R) 8.02 (R)
Etanol 3.42 (R) 7.88 (R) 15.22 (I) 8.12 (R)
Air 2.31 (R) 5.30 (R) 8.53 (R) 3.03 (R)
Keterangan: S (susceptible): ≥ 16 mm, I (intermediate): 13-15 mm, R (resistant): ≤ 12 mm (CLSI
2007)
Konsentrasi Hambat Tumbuh Minimun Ekstrak Etanol
Konsentrasi minimum yang dibutuhkan untuk menghambat bakteri dikenal
sebagai konsentrasi hambat tumbuh minimum (KHTM). Uji KHTM merupakan
upaya mencari konsentrasi terendah yang masih mampu menghambat pertumbuhan
organisme mikroba. Hasil uji aktivitas antibakteri pada Tabel 2 menunjukkan
bahwa ekstrak etanol merupakan ekstrak dengan aktivitas terbaik sehingga
digunakan untuk mencari nilai KHTM. Hasil uji KHTM ekstrak etanol tanaman Z.
zerumbet dapat dilihat pada Tabel 3. Terlihat bahwa KHTM ekstrak etanol dalam
keempat bakteri uji terjadi untuk bakteri Gram positif S. aureus pada konsentrasi
50 mg mL-1.
Tabel 3 Hasil uji KHTM ekstak etanol Z. zerumbet
Konsentrasi
ekstrak etanol
(mg mL-1)
Luas zona hambat (mm) pada bakteri
Ps. Aeruginosa E. coli B. subtilis S.aureus
250 2.59 (R) 2.89 (R) 3.42 (R) 7.71 (R)
150 - 1.98 (R) 2.72(R) 7.13 (R)
100 - - 2.27 (R) 6.12 (R)
50 - - - 3.85 (R)
K+ 8.96 (R) 29.66 (S) 11.59 (R) 27.92 (S)
K- - - - -
Keterangan: S (susceptible): ≥ 16 mm, I (intermediate): 13-15 mm, R (resistant): ≤ 12 mm (CLSI
2007)
Hasil Uji Fitokimia
Uji fitokimia bertujuan untuk mengetahui kandungan senyawa metabolit
sekunder dan golongan senyawa bioaktif yang terkandung di dalam ekstrak. Uji
fitokimia meliputi uji kualitatif fenolik, triterpenoid-steroid, alkaloid, saponin,
tanin, dan flavonoid. Dari hasil uji fitokimia ini dapat diduga golongan senyawa
yang berperan sebagai antibakteri. Hasil uji fitokimia mengindikasikan ekstrak
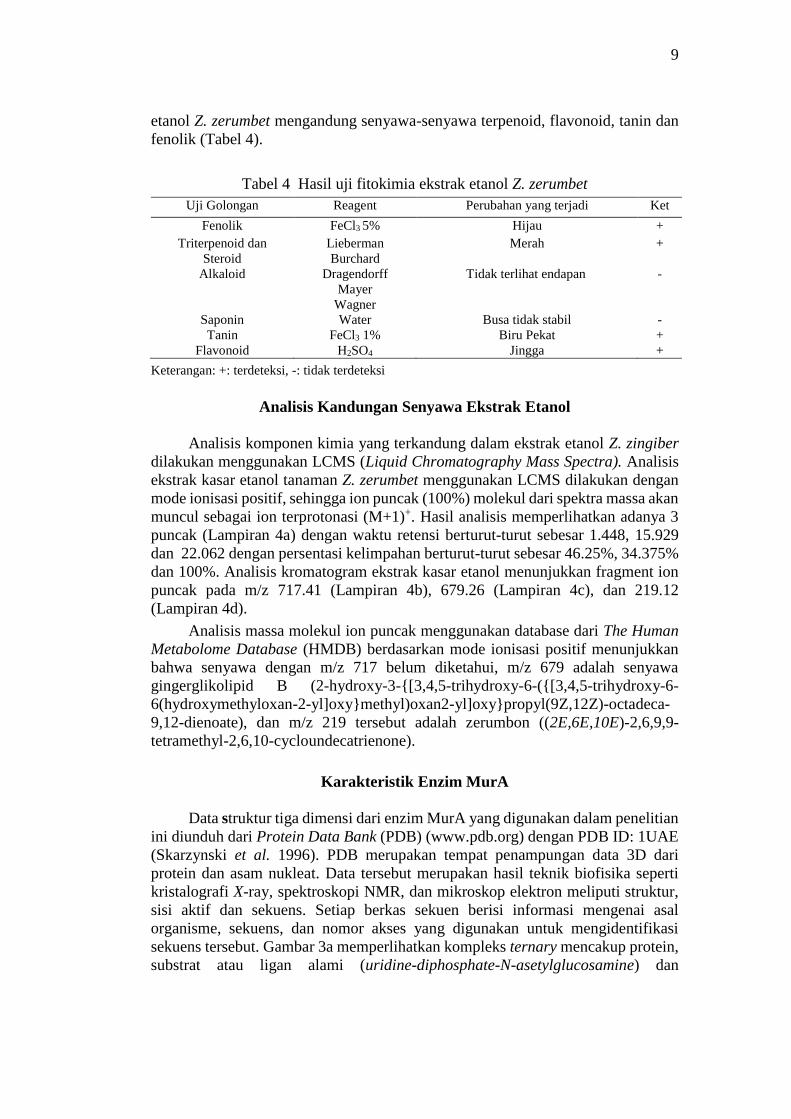
9
etanol Z. zerumbet mengandung senyawa-senyawa terpenoid, flavonoid, tanin dan
fenolik (Tabel 4).
Tabel 4 Hasil uji fitokimia ekstrak etanol Z. zerumbet
Uji Golongan Reagent Perubahan yang terjadi Ket
Fenolik FeCl3 5% Hijau +
Triterpenoid dan
Steroid
Lieberman
Burchard
Merah +
Alkaloid Dragendorff
Mayer
Wagner
Tidak terlihat endapan -
Saponin Water Busa tidak stabil -
Tanin FeCl3 1% Biru Pekat +
Flavonoid H2SO4 Jingga +
Keterangan: +: terdeteksi, -: tidak terdeteksi
Analisis Kandungan Senyawa Ekstrak Etanol
Analisis komponen kimia yang terkandung dalam ekstrak etanol Z. zingiber
dilakukan menggunakan LCMS (Liquid Chromatography Mass Spectra). Analisis
ekstrak kasar etanol tanaman Z. zerumbet menggunakan LCMS dilakukan dengan
mode ionisasi positif, sehingga ion puncak (100%) molekul dari spektra massa akan
muncul sebagai ion terprotonasi (M+1)+. Hasil analisis memperlihatkan adanya 3
puncak (Lampiran 4a) dengan waktu retensi berturut-turut sebesar 1.448, 15.929
dan 22.062 dengan persentasi kelimpahan berturut-turut sebesar 46.25%, 34.375%
dan 100%. Analisis kromatogram ekstrak kasar etanol menunjukkan fragment ion
puncak pada m/z 717.41 (Lampiran 4b), 679.26 (Lampiran 4c), dan 219.12
(Lampiran 4d).
Analisis massa molekul ion puncak menggunakan database dari The Human
Metabolome Database (HMDB) berdasarkan mode ionisasi positif menunjukkan
bahwa senyawa dengan m/z 717 belum diketahui, m/z 679 adalah senyawa
gingerglikolipid B (2-hydroxy-3-{[3,4,5-trihydroxy-6-({[3,4,5-trihydroxy-6-
6(hydroxymethyloxan-2-yl]oxy}methyl)oxan2-yl]oxy}propyl(9Z,12Z)-octadeca-
9,12-dienoate), dan m/z 219 tersebut adalah zerumbon ((2E,6E,10E)-2,6,9,9-
tetramethyl-2,6,10-cycloundecatrienone).
Karakteristik Enzim MurA
Data struktur tiga dimensi dari enzim MurA yang digunakan dalam penelitian
ini diunduh dari Protein Data Bank (PDB) (www.pdb.org) dengan PDB ID: 1UAE
(Skarzynski et al. 1996). PDB merupakan tempat penampungan data 3D dari
protein dan asam nukleat. Data tersebut merupakan hasil teknik biofisika seperti
kristalografi X-ray, spektroskopi NMR, dan mikroskop elektron meliputi struktur,
sisi aktif dan sekuens. Setiap berkas sekuen berisi informasi mengenai asal
organisme, sekuens, dan nomor akses yang digunakan untuk mengidentifikasi
sekuens tersebut. Gambar 3a memperlihatkan kompleks ternary mencakup protein,
substrat atau ligan alami (uridine-diphosphate-N-asetylglucosamine) dan

10
fosfomisin atau ligan obat. Fosfomisin sebagai obat komersial yang menghambat
kerja enzim ini dengan menjadi analog substrat yang bereaksi dengan residu Cys115.
(A) (B)
Gambar 3 Karakteristik enzim MurA: (A) Struktur 3D kompleks ternary enzim
(Skarzynski et al. 1996); (B) Plot Ramachandran enzim
Enzim Mur A terdiri dari 418 residu dengan komposisi helixnya sebesar
34% yang terdiri dari 18 helix dengan 143 residu asam amino. Sedangkan beta
sheetnya terdiri dari 34 strand dengan 113 residu. Struktur sekunder enzim Mur A
dapat dilihat pada gambar 4.
Gambar 4 Struktur sekunder reseptor MurA:
= Beta hairpin
= Turn
= Alfa heliks
= 3/10-heliks

11
Kestabilan Struktur Reseptor
Kestabilan struktur enzim MurA dilihat dari plot Ramachandran (Gambar
3b). Plot Ramachandran digunakan untuk mengetahui kualitas struktur enzim.
Setiap asam amino penyusun protein akan memiliki satu sudut phi (Ф) dan psi (ψ),
sehingga setiap residu asam tergambarkan sebagai satu plot (koordinat). Visualisasi
plot Ramachandran diunduh pada Protein Data Bank (PDB). Kualitas struktur
protein dapat diketahui dengan melihat sudut dihedral yang dilarang (dissalowed
region). Bila residu non glisin yang berada pada dissalowed region lebih dari 15%
maka dapat dikatakan bahwa protein tersebut memiliki kualitas struktur yang sangat
tidak stabil (kurang baik). Enzim MurA bersifat stabil terlihat dari data asam amino
yang berada pada daerah yang paling disukai (favorable) sebesar 97.8% (407/406)
dan daerah yang diizinkan (allowed) sebesar 100.0% (416/416) serta daerah yang
tidak diijinkan (disallowed) kurang dari 1%.
Potensi Ligan Menurut Aturan Lipinski
Struktur ligan yang digunakan dalam penelitian ini ditunjukkan pada Tabel
5. Sementara itu potensi ligan untuk dapat masuk ke dalam membran sel dan diserap
diuji menggunakan software Lipinski dengan parameternya adalah berat molekul,
jumlah gugus donor ikatan hidrogen, jumlah gugus penerima ikatan hidrogen, nilai
log P dan refrakfititas molar (Lipinski et al. 2001). Pengujian dilakukan
menggunakan tools online pada situs http://scfbio-
iitd.res.in/software/utility/lipinski filters.jsp. Hasil filter Lipinski antara ligan alami,
ligan fosfomisin (obat), ligan gingerglikolipid B (senyawa uji), dan ligan zerumbon
(senyawa uji) dapat dilihat pada Tabel 6.
Tabel 5 Struktur 2D ligan (substrat atau UDP-N-GluAc, fosfomisin, zerumbon,
dan gingerglikolipid B)
Structur 2D Nama IUPAC Nama
umum
[({[2R,3S,4R,5R)-5-(2,4-dioxo-
1,2,3,4-tetrahydropyrimidin-1-yl)-
3,4-dihydroxyoxolan-2-
yl]methoxy}(hydroxyphosphoryl)o
xy]({[2R,3R,4R,5S,6R)-3-
acetamido-4,5-dihydroxy-6-
(hydroxymethyl)oxan-2-
yl]oxy}phosphinic acid
Uridine-
diphosphate
-N-acetyl-
glucosamine
[(1R)-1-hydroxypropyl]phosphonic
acid
Fosfomisin
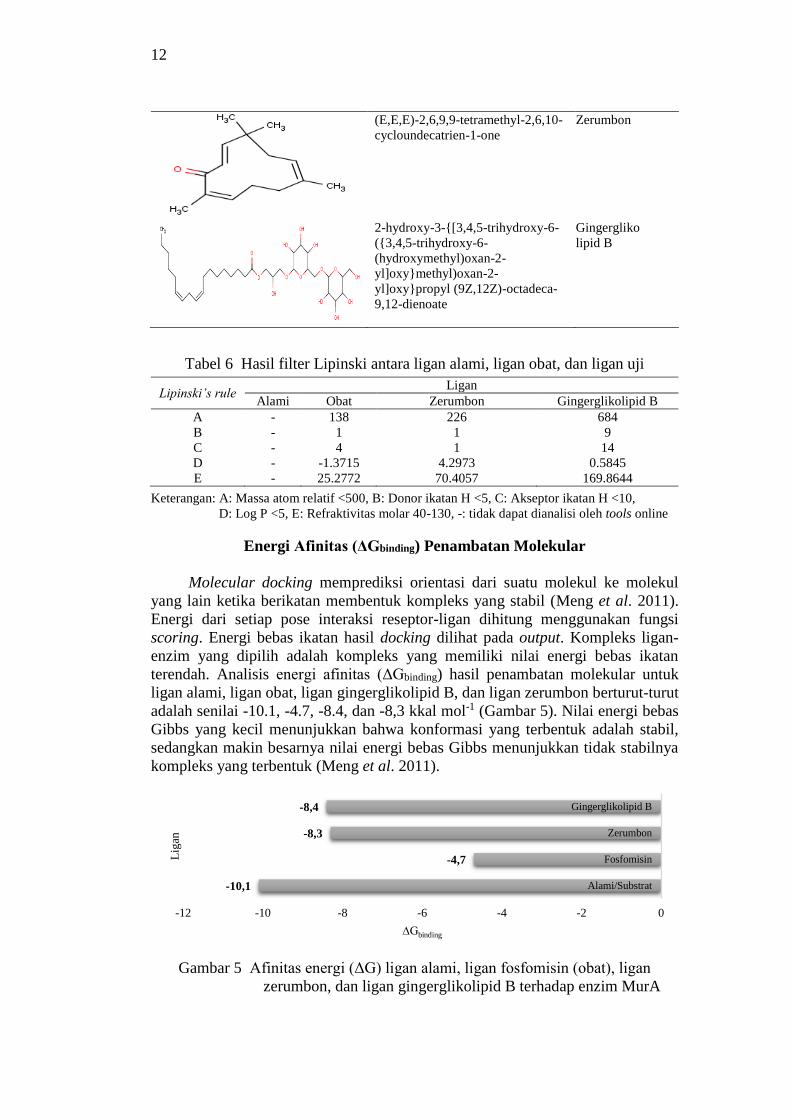
12
(E,E,E)-2,6,9,9-tetramethyl-2,6,10-
cycloundecatrien-1-one
Zerumbon
2-hydroxy-3-{[3,4,5-trihydroxy-6-
({3,4,5-trihydroxy-6-
(hydroxymethyl)oxan-2-
yl]oxy}methyl)oxan-2-
yl]oxy}propyl (9Z,12Z)-octadeca-
9,12-dienoate
Gingergliko
lipid B
Tabel 6 Hasil filter Lipinski antara ligan alami, ligan obat, dan ligan uji
Lipinski’s rule Ligan
Alami Obat Zerumbon Gingerglikolipid B
A - 138 226 684
B - 1 1 9
C - 4 1 14
D - -1.3715 4.2973 0.5845
E - 25.2772 70.4057 169.8644
Keterangan: A: Massa atom relatif <500, B: Donor ikatan H <5, C: Akseptor ikatan H <10,
D: Log P <5, E: Refraktivitas molar 40-130, -: tidak dapat dianalisi oleh tools online
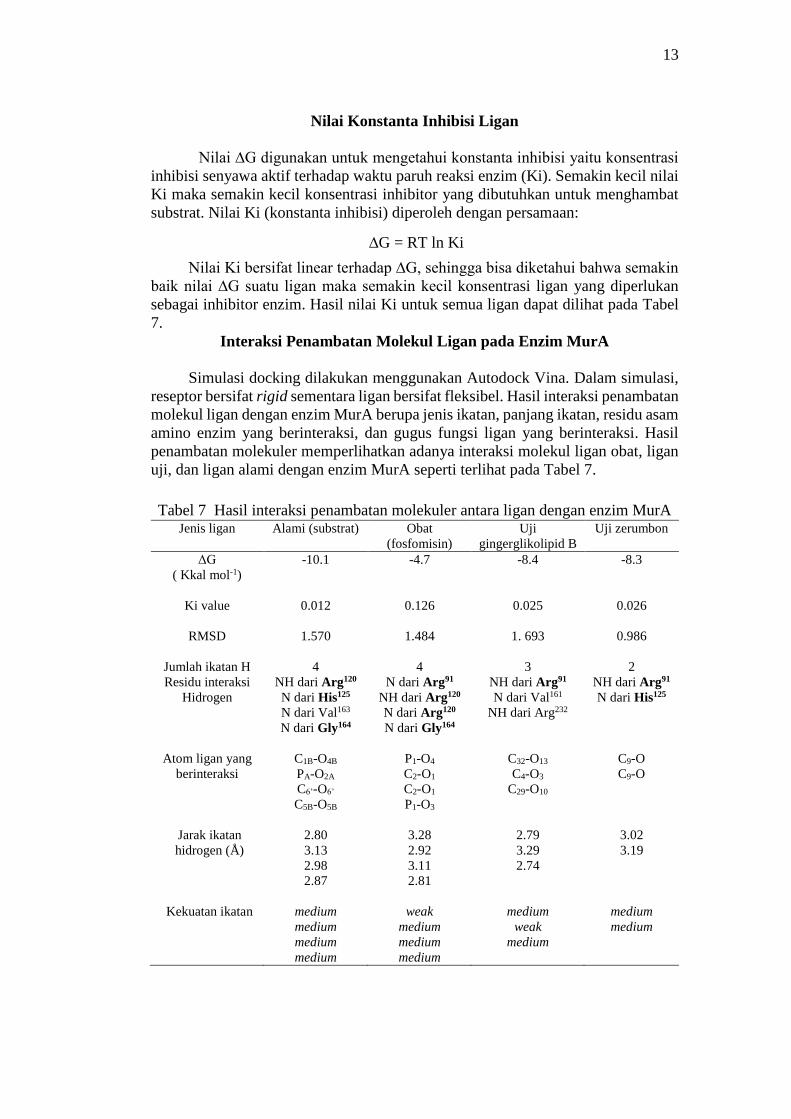
Energi Afinitas (ΔGbinding) Penambatan Molekular
Molecular docking memprediksi orientasi dari suatu molekul ke molekul
yang lain ketika berikatan membentuk kompleks yang stabil (Meng et al. 2011).
Energi dari setiap pose interaksi reseptor-ligan dihitung menggunakan fungsi
scoring. Energi bebas ikatan hasil docking dilihat pada output. Kompleks ligan-
enzim yang dipilih adalah kompleks yang memiliki nilai energi bebas ikatan
terendah. Analisis energi afinitas (ΔGbinding) hasil penambatan molekular untuk
ligan alami, ligan obat, ligan gingerglikolipid B, dan ligan zerumbon berturut-turut
adalah senilai -10.1, -4.7, -8.4, dan -8,3 kkal mol-1 (Gambar 5). Nilai energi bebas
Gibbs yang kecil menunjukkan bahwa konformasi yang terbentuk adalah stabil,
sedangkan makin besarnya nilai energi bebas Gibbs menunjukkan tidak stabilnya
kompleks yang terbentuk (Meng et al. 2011).
Gambar 5 Afinitas energi (ΔG) ligan alami, ligan fosfomisin (obat), ligan
zerumbon, dan ligan gingerglikolipid B terhadap enzim MurA
-10,1
-4,7
-8,3
-8,4
-12 -10 -8 -6 -4 -2 0
Alami/Substrat
Fosfomisin
Zerumbon
Gingerglikolipid B
∆Gbinding
Lig
an
13
Nilai Konstanta Inhibisi Ligan
Nilai ∆G digunakan untuk mengetahui konstanta inhibisi yaitu konsentrasi
inhibisi senyawa aktif terhadap waktu paruh reaksi enzim (Ki). Semakin kecil nilai
Ki maka semakin kecil konsentrasi inhibitor yang dibutuhkan untuk menghambat
substrat. Nilai Ki (konstanta inhibisi) diperoleh dengan persamaan:
Nilai Ki bersifat linear terhadap ∆G, sehingga bisa diketahui bahwa semakin
baik nilai ∆G suatu ligan maka semakin kecil konsentrasi ligan yang diperlukan
sebagai inhibitor enzim. Hasil nilai Ki untuk semua ligan dapat dilihat pada Tabel
7.
Interaksi Penambatan Molekul Ligan pada Enzim MurA
Simulasi docking dilakukan menggunakan Autodock Vina. Dalam simulasi,
reseptor bersifat rigid sementara ligan bersifat fleksibel. Hasil interaksi penambatan
molekul ligan dengan enzim MurA berupa jenis ikatan, panjang ikatan, residu asam
amino enzim yang berinteraksi, dan gugus fungsi ligan yang berinteraksi. Hasil
penambatan molekuler memperlihatkan adanya interaksi molekul ligan obat, ligan
uji, dan ligan alami dengan enzim MurA seperti terlihat pada Tabel 7.
Tabel 7 Hasil interaksi penambatan molekuler antara ligan dengan enzim MurA
Jenis ligan Alami (substrat) Obat
(fosfomisin)
Uji
gingerglikolipid B
Uji zerumbon
ΔG
( Kkal mol-1)
-10.1 -4.7 -8.4 -8.3
Ki value
0.012 0.126 0.025 0.026
RMSD
1.570 1.484 1. 693 0.986
Jumlah ikatan H 4 4 3 2
Residu interaksi
Hidrogen
NH dari Arg120
N dari His125
N dari Val163
N dari Gly164
N dari Arg91
NH dari Arg120
N dari Arg120
N dari Gly164
NH dari Arg91
N dari Val161
NH dari Arg232
NH dari Arg91
N dari His125
Atom ligan yang
berinteraksi
C1B-O4B
PA-O2A
C6’-O6’
C5B-O5B
P1-O4
C2-O1
C2-O1
P1-O3
C32-O13
C4-O3
C29-O10
C9-O
C9-O
Jarak ikatan
hidrogen (Å)
2.80
3.13
2.98
2.87
3.28
2.92
3.11
2.81
2.79
3.29
2.74
3.02
3.19
Kekuatan ikatan medium
medium
medium
medium
weak
medium
medium
medium
medium
weak
medium
medium
medium
∆G = RT ln Ki

14
4 PEMBAHASAN
Identifikasi Tumbuhan dan Rendemen Ekstrak
Z. zerumbet memiliki ukuran rimpang yang besar berwarna kuning gading,
dengan rasa yang cukup pahit, agak pedas, dan bau yang khas dari senyawa
aromatik (spice taste) (Gambar 1A). Tinggi tanaman berkisar 1.500-1.800 cm.
Memiliki batang semu dan tumbuh tegak serta berwarna kemerahan (Gambar 1B).
Daunnya berbentuk obovate (bulat telur terbalik) dengan pangkal daun membulat
dan ujung daunnya agak meruncing (subacuminate). Daun bertrikom halus
(subglabrous) serta memiliki tangkai (petiole) bermembran, ligula bermembran,
dan pelepah (sheath) (Gambar 1C). Bunga tanaman tumbuh dari pangkal rimpang
berbentuk seperti bonggol (cone), dan bunganya berwarna kuning pucat (Gambar
1D). Braktea merupakan kantong tempat munculnya bunga dimana satu bunga
dalam satu braktea. Braktea berwarna hijau sewaktu muda dan berubah warna
menjadi merah. Perubahan warna menjadi merah terjadi setelah pembuahan (Larsen
et al. 1999).
Ekstraksi dilakukan dengan teknik meserasi yang mana pengambilan
komponen atau zat aktif dilakukan dengan cara merendam bahan dalam cairan
ekstrak yang sesuai selama beberapa waktu dan suhu tertentu. Maserasi
dimaksudkan untuk dapat mengekstrak keseluruhan senyawa yang larut dalam
pengekstrak yang digunakan. Komponen aktif tersebut akan larut dengan prinsip
like disolved like. Penggunaan pelarut dengan tingkat kepolaran yang berbeda
bertujuan untuk mengekstrak komponen aktif yang belum sepenuhnya diketahui
dalam rimpang Z. zerumbet. Selama proses meserasi dilakukan pengadukan dan
penggantian cairan penyari setiap hari. Endapan yang diperoleh kemudian
dipisahkan dan filtratnya dipekatkan (Lampiran 3). Persentasi rendemen ektrak
pada Tabel 1, memperlihatkan kemampuan pelarut air lebih besar dalam
mengekstraksi komponen aktif tanaman Z. zerumbet, diikuti oleh pelarut etanol, etil
Residu interaksi
hidrofobik
Lys22, Asn23,
Leu24, Leu26,
Trp95, Arg91,
Ala92, Pro121,
Ser162, Val317,
Phe328
Ser162, Val163,
Phe32
Asn23, Val157,
Lys160, Val163,
Glu183, Asn184,
Glu188, Asp211,
Pro298, Thr304,
Asp305, Val327,
Phe328
Trp95, Ser162,
Val163,
Gly164, Val327,
Phe328
Kesamaan
interaksi
asam amino
terhadap Substrat
Lys22, Asn23,
Leu26, Arg91,
Ala92, Trp95,
Arg120, Pro121,
Leu124, His125,
Val161, Ser162,
Val163, Gly164,
Val327, Phe328
Arg91, Arg120,
Ser162, Val163,
Gly164, Phe328
Asn23, Arg91,
Val157, Lys160,
Val161, Val163,
Glu183, Asn184,
Glu188, Asp211,
Arg232, Pro298,
Thr304, Asp305,
Val327, Phe328
Arg91, Trp95,
His125, Ser162,
Val163, Gly164,
Val327, Phe328
Persentase
kesamaan
interaksi asam
amino terhadap
substrat (%)
16 (100)
6 (37,50)
6 (37,50)
8 (50)

15
asetat dan N-heksana. Rendemen ekstrak air yang lebih besar kemungkinan
dipengaruhi oleh polaritasnya. Polaritas relatif pelarut dapat dilihat dari nilai
konstanta dielektrik masing-masing pelarut. Polaritas pelarut air adalah sebesar
0,90 yang mana lebih besar dari pelarut etanol (0.68), etil asetat (0.38) dan heksana
(0.00) (Yasni 2013). Hal ini menunjukkan rendemen ekstrak makin meningkat
seiring meningkatnya kepolaran pelarut. Tingginya kemampuan pelarut air dalam
mengekstraksi komponen aktif berkaitan dengan polaritas (ε) pelarut yang berarti
komponen senyawa yang terkandung dalam rimpang Z. zerumbet sebagian besar
merupakan senyawa polar. Sifat kepolaran senyawa dilihat dari gugus polarnya
seperti gugus OH, COOH, dan lain-lain. Hal lain yang memperbesar kemampuan
pelarut air dalam menarik komponen aktif adalah proses ekstraksi yang
menggunakan pemanasan pada suhu 80oC yang mana proses pemanasan akan
memperbesar kelarutan.
Aktivitas Antibakteri Ekstrak N-Heksan, Etil Asetat, Etanol, dan Air dan
KHTM Ekstrak Etanol
Pengujian aktivitas antibakteri ekstrak masing-masing dilakukan pada
konsentrasi 500 mg mL-1 untuk mengetahui ada atau tidaknya kemampuan ekstrak
tanaman menghambat pertumbuhan bakteri patogen. Kontrol positif yang
digunakan adalah fosfomisin, sedangkan kontrol negatif yang digunakan adalah
aquadest. Ekstrak Z. zerumbet dinyatakan mempunyai aktivitas antibakteri jika
terbentuk zona bening di sekeliling sumuran berisi ekstrak yang ditumbuhkan pada
media yang telah diinokulasi oleh bakteri. Aktivitas antibakteri ekstrak Z. zerumbet
dapat dilihat pada Tabel 2. Dari data di atas, terlihat bahwa ekstrak etanol
memberikan batas daerah hambat yang terefektif dengan diameter terbesar adalah
15.22 mm pada bakteri B. subtilis, dan diameter terkecil adalah 3.42 mm pada
bakteri E. coli. Kader et al. (2011) melaporkan bahwa ekstrak kasar etanol Z.
zerumbet dengan konsentrasi 400 µg disk-1 mampu menghambat 5 jenis bakteri
Gram positif dan 8 jenis bakteri Gram negatif dengan luas zona hambat berkisar 6-
10 mm. Sifat antibakteri ini berkaitan dengan senyawa-senyawa yang terkandung
yakni zederon (suatu sesquiterpen), fenolik, saponin, dan terpenoid (Hashemi et al.
2008; Kader et al. 2010). Penghambatan aktivitas bakteri oleh senyawa aktif
tanaman dapat disebabkan oleh beberapa faktor, yakni gangguan pada senyawa
penyusun dinding sel, peningkatan permeabilitas membran sel yang menyebabkan
kehilangan senyawa penyusun sel, menginaktivasi enzim metabolik, dan destruksi
fungsi material genetik (Yasni 2013). Pengaruh senyawa antibakteri terhadap sel
bakteri dapat menyebabkan kerusakan sel yang berlanjut pada kematian. Kerusakan
bakteri merupakan hasil interaksi senyawa antibakteri dengan bagian tertentu pada
sel. Terjadinya proses tersebut karena perlekatan senyawa antibakteri pada
permukaan sel maupun akibat berdifusinya senyawa tersebut ke dalam sel. Interaksi
tersebut mengakibatkan sejumlah perubahan atau kerusakan pada sel bakteri yang
mempengaruhi fungsi metabolisme sel.
Data penelitian menunjukkan bahwa ekstrak etanol Z. zerumbet lebih
menghambat bakteri Gram positif (S. aureus dan B. Subtilis) dibanding bakteri
Gram negatif (E. coli dan P. aeruginosa). Hal ini kemungkinan dipengaruhi oleh
perbedaan komposisi dan struktur pada dinding sel bakteri Gram positif dan Gram
negatif. Struktur dinding sel bakteri Gram positif lebih sederhana, yakni berlapis

16
tunggal (90% peptidoglikan) dengan ketebalan bervariasi antara 20-40 nm (Kim et
al. 2015) dengan kandungan lipid yang rendah (1-4%) sehingga memudahkan
bahan bioaktif masuk ke dalam sel. Sementara itu, struktur dinding sel bakteri
negatif lebih kompleks, yaitu berlapis tiga terdiri dari lapisan luar lipoprotein,
lapisan tengah lipopolisakarida yang berperan menghalangi masuknya bahan
bioaktif antibakteri (Griffin 2000), dan lapisan dalam berupa peptidoglikan yang
berkisar 5-10 % (6.25±0.53 nm untuk E. coli dan 2.41±0.54 nm untuk Ps.
aeruginosa) dengan kandungan lipid tinggi (11-12%) (Vollmer dan Seligman
2009).
Data pada Tabel 3 menunjukkan nilai KHTM terendah terjadi pada bakteri S.
aureus. S. aureus merupakan bakteri Gram positif, yang memiliki 40 lapisan
peptidoglikan yang merupakan 50% dari bahan penyusun dinding sel. Perhusip
(2006) mengungkapkan bahwa setiap zat yang menghambat salah satu langkah
dalam biosintesis peptidoglikan akan menakibatkan dinding sel bakteri yang
tumbuh menjadi rapuh sehingga sel akan mengalami lisis. Rusaknya dinding sel
atau terhambatnya sintesis oleh senyawa antibakteri mengakibatkan terbentuknya
sel-sel yang peka terhadap tekanan osmosis, terutama pada bakteri Gram positif
yang mana kekuatan dinding selnya berasal dari peptidoglikan. Tekanan osmosis
dalam sel bakteri menyebabkan terjadinya lisis, yang merupakan dasar efek
bakterisidal pada bakteri (Perhusip 2006).
Dari data hasil uji KHTM (Tabel 3), memperlihatkan ekstrak etanol Z.
zerumbet mempunyai aktivitas yang sedang. Ini berkaitan dengan kondisi ekstrak
yang masih belum murni. Banyaknya senyawa yang terkandung dalam ekstrak
kasar etanol ini mengakibatkan adanya sifat saling meniadakan (antagonis) antar
senyawa dalam ekstrak sehingga mengurangi aktivitas antibakteri ekstrak etanol Z.
zerumbet. Pemisahan atau fraksinasi senyawa dalam ekstrak lebih lanjut berperan
penting dalam meningkatkan kemampuan hambat terhadap bakteri. Kader et al.
2011 melaporkan bahwa ekstrak etanol yang telah difraksinasi memiliki
kemampuan hambat terendah sebesar 128-256 µg disk-1 saat diujikan pada bakteri
E. coli, P. aeruginosa, S. typhi, B. cereus, S. lutea, dan V. Parahemolyticus.
Uji Fitokimia Ekstrak Etanol
Uji fitokimia menggambarkan akan golongan senyawa yang terkandung
dalam ekstrak. Hasil uji fitokimia mengindikasikan rimpang tanaman Z. zerumbet
mengandung senyawa-senyawa terpenoid, flavonoid, tanin dan fenolik (Tabel 3).
Flavanoid dan tanin merupakan bagian dari senyawa fenolik (Harbone 1996).
Alkaloid diketahui tidak terkandung dalam ekstrak etanol Z. zerumbet, hal ini
diduga berkaitan dengan keberadaan alkaloid dalam jaringan tumbuhan kurang dari
1 % sehingga dapat menyebabkan uji skrining alkaloid memberikan hasil negatif
(Harborne 1996). Nag et al. (2013) melaporkan bahwa Z. zerumbet mengandung
sesquiterpenoid, flavonoid, senyawa-senyawa aromatik, vanillin, kaempferol dan
senyawa turunannya, serta polifenol.
Senyawa-senyawa fenolik, flavanoid, tanin, dan terpenoid pada tanaman
zingiberaceae diketahui memiliki aktivitas antibakteri (Nursal et al. 2006; Lawalata
2012). Senyawa terpen merupakan senyawa antibakteri utama dalam rempah-
rempah (Yasni 2013). Terpenoid diketahui dapat bersifat sebagai antibakteri
(Cowan 1999). Mekanisme terpenoid sebagai antibakteri adalah bereaksi dengan

17
fraksi lipid membran plasma bakteri yang mengakibatkan perubahan permeabilitas
membran yang jika diakumulasikan terus-menerus dapat mengakibatkan lisisnya
material intraseluler akibat terbentuknya rongga pada lipid bilayer (Griffin 2000).
Senyawa tanin dan flavonoid merupakan senyawa polifenol yang bersifat sebagai
antibakteri (Cowan 1999). Senyawa flavanoid dalam aktivitas kerjanya akan
membentuk ikatan kompleks dengan dinding sel bakteri sehingga menurunkan
permeabilitas dinding sel (Nagappan et al. 2011) dan merusak membran sel bakteri
akibat sifat lipofiliknya (de Fatima et al. 2006). Demikian halnya tanin, tanin diduga
berikatan dengan dinding sel bakteri sehingga akan menginaktifkan kemampuan
menempel bakteri dan menghambat pertumbuhan bakteri (Cowan 1999).
Analisis Kandungan Senyawa Ekstrak Etanol
Analisis komponen kimia yang terkandung dalam ekstrak etanol Z. zerumbet
dilakukan menggunakan LCMS (Liquid Chromatography Mass Spectra). Hasil
analisis ekstrak etanol Zingiber zerumbet menggunakan KCSM (kromatografi cair
spektra massa) memperlihatkan adanya 3 puncak dengan waktu retensi berturut-
turut sebesar 1.448, 15.929 dan 22.062 dengan persentasi kelimpahan berturut-
turut sebesar 46.25%, 34.375% dan 100%.
Analisis ekstrak etanol Z. zerumbet menggunakan KCMS dilakukan dengan
mode ionisasi positif, sehingga ion puncak (100%) molekul dari spektra massa akan
muncul sebagai ion terprotonasi (M+1)+. Analisis massa molekul ion puncak
menggunakan database dari The Human Metabolome Database (HMDB) berdasarkan
mode ionisasi positif menunjukkan bahwa senyawa dengan m/z 219 tersebut adalah
zerumbon ((2E,6E,10E)-2,6,9,9-tetramethyl-2,6,10-cycloundecatrienone). Abdul et al.
(2008) mengemukakan bahwa massa ion fragmentasi terbesar (218 m/z) sebanding
dengan bobot molekul senyawa. Zerumbon merupakan konstituen terbesar pada
rimpang tanaman dengan kelimpahan berkisar antara 12.6-73.1 % (Yob et al. 2011;
Singh et al. 2012). Zerumbon mempunyai rumus kimia C15H22O dengan berat molekul
sebesar 218.3346 (Tabel 5). Zerumbon adalah senyawa monosiklik sesquiterpen dan memiliki kekhasan
dalam strukturnya dimana mempunyai suatu konjugasi silang keton pada 11
anggota cincinnya (Kitayama et al. 2001). Zerumbon memiliki tiga ikatan rangkap
di mana ikatan rangkap pada posisi C6 terisolasi sedangkan dua ikatan pada posisi
C2 dan C10 terkonjugasi dalam sistem dienon (Kitayama et al. 2011). Ikatan
rangkap pada C10 terkonjugasi silang dengan gugus karbonil pada struktur cincin
beranggota 11 (Kitayama et al. 2011). Gugus keton pada atom C-8 mengakibatkan
zerumbon bersifat polar sehingga bisa terekstrak pada pelarut polar dan semi polar.
Sifat polaritas senyawa merupakan sifat kimia yang sangat penting. Kepolaran ini
berkaitan kelarutannya dalam air dan sifat hidrofilik sehingga zat antibakteri ini
dapat larut dalam fase air dimana mikroba umumnya tumbuh dalam fase air. Abdul
et al. 2008 melaporkan bahwa zerumbon memiliki aktivitas antibakteri pada S.
choleraesuis tetapi tidak memiliki kemampuan menghambat pada pada bakteri
resisten methicillin dari S. aureus, P. Aeruginosa, S. choleraesuis dan B. subtilis.
(Kumar et al. 2013) melaporkan bahwa konsentrasi MIC zerumbon dalam
menghambat bakteri B. cereus, S. aureus, E. coli, dan Y. Enterocolitica berturut-
turut sebesar 100, 125, 75, dan 250 ppm. Senyawa sesquiterpena diketahui memiliki
aktivitas antibakteri lebih baik pada bakteri Gram positif dari pada bakteri Gram

18
negatif. Ini diduga berkaitan dengan dimilikinya membran luar pada bakteri Gram
negatif yang mengelilingi dinding sel, dimana membatasi difusi senyawa hidrofilik
melalui lipopolysaccha.
Senyawa dengan massa molekul ion puncak m/z 679 juga menjadi penyusun
ekstrak etanol tanaman. Analisis menggunakan database HMDB memberikan hasil
bahwa senyawa tersebut adalah Gingerglikolipid B (2-hydroxy-3-{[3,4,5-
trihydroxy-6-({[3,4,5-trihydroxy-6-6(hydroxymethyloxan-2-
yl]oxy}methyl)oxan2-yl]oxy}propyl(9Z,12Z)-octadeca-9,12-dienoate). Senyawa
ini memiliki rumus kimia C33H58O14 dengan berat molekul sebesar 678.8052 (Tabel
5). Gingerglikolipid B umumnya ditemukan sebagai senyawa penyusun pada
tanaman Zingiber officinale dan diketahui bersifat sebagai anti borok (Yoshikawa
et al. 1994). Gingerglikolipid B merupakan suatu monoasildigalaktosilgliserol.
Beragamnya senyawa-senyawa aktif yang terkandung pada tanaman Z.
zerumbet dipengaruhi oleh kondisi lingkungan tanaman tersebut tumbuh. Pulau
Timor merupakan wilayah berkarakteristik semi arid, dimana mengalami musim
kering yang panjang dan musim penghujan yang pendek. Keadaan daerah yang
beriklim semiarid mengakibatkan kondisi tanamannya berada dalam keadaan cekaman
kekeringan dan suhu tinggi dengan intensitas penyinaran matahari lebih panjang
(Selmar 2008). Ini mengakibatkan tanaman yang tumbuh mengalami kondisi
cekaman kekeringan. Kondisi stres abiotik ini mengakibatkan tanaman
mengakumulasikan secara berlebih senyawa-senyawa metabolit tertentu yang
berbeda dengan daerah lain sebagai toleransi akan kondisi tersebut (Sopandie
2014).
Simulasi Molecular Docking Ligan Senyawa Aktif Z. zerumbet Terhadap
Reseptor MurA
Kelangsungan hidup bakteri bergantung pada aktivitas enzim UDP-N-
acetylglucosamine enolpyruvyl transferase, EC 2.5.1.7 (MurA) (Skarzynski et al.
1996; Eschenburg et al. 2005). MurA mengkatalisis tahapan pertama biosintesis
peptidoglikan dengan mengkondensasikan fosfoenolpiruvat (PEP) dan UDP-N-
asetilglukosamin (UDP-Glc-NAc) menjadi UDP-GlcNAc-enolpiruvat.
Peptidoglikan adalah heteropolimer glikan yang dihubungsilangkan dengan asam
amino. Peptidoglikan merupakan komponen spesifik penyusun terbesar pada
dinding sel bakteri, yang terletak di luar membran sitoplasma (Vollmer et al. 2008).
Peptidoglikan sangat esensial bagi kehidupan bakteri karena berperan dalam
menahan tekanan turgor (Vollmer et al. 2008; de Pedro dan Cava 2015) dan
memberi bentuk atau kekakuan sel (Margolin 2009) sehingga melindungi bakteri
dari kerusakan akibat tekanan osmosis.
Terhambatnya aktivitas enzim MurA mengakibatkan ikut terhambatnya
biosintesis dinding sel bakteri. Rusaknya dinding sel atau terhambatnya sintesis
oleh senyawa antibakteri mengakibatkan terbentuknya sel-sel yang peka terhadap
tekanan osmosis, terutama pada bakteri Gram positif yang mana kekuatan dinding
selnya berasal dari peptidoglikan. Tekanan osmosis dalam sel bakteri menyebabkan
terjadinya lisis, yang mengakibatkan kematian bakteri. Molecular docking adalah metode yang digunakan untuk memprediksi oreintasi
antara satu molekul dengan molekul lainnya ketika terjadi suatu gaya antara satu
dengan yang lain untuk membentuk suatu ikatan yang stabil (Scheneider dan

19
Baringhaus 2008). Teknik ini memprediksikan apakah suatu molekul dapat berikatan
dengan reseptor, protein, DNA dan ligan docking dengan teknik penempatan pada area
tertentu. Tujuan simulasi molecular docking adalah untuk memahami dan
memprediksi rekognisi molekuler dengan mencari kestabilan ikatan antara ligan
dan reseptor pada keadaan energi minimum (Scheneider dan Baringhaus 2008;
Yanuar 2012). Simulasi docking dapat dipergunakan untuk memperoleh mekanisme
kerja suatu senyawa kimia atau makromolekul seperti protein maupun peptida, dalam
skala molekuler sehingga dimungkinkan untuk mendesain obat berbasis struktur. Data struktur tiga dimensi dari enzim MurA ligase yang digunakan dalam
penelitian ini diunduh dari Protein Data Bank (PDB) (www.pdb.org) dengan PDB
ID: 1UAE. Data tersebut merupakan hasil teknik biofisika seperti kristalografi X-
ray maupun spektroskopi NMR meliputi struktur, sisi aktif dan sekuens. Gambar
3A memperlihatkan kompleks ternary mencakup protein, substrat (uridine-
diphosphate-N-asetylglucosamine) dan ligan (fosfomisin). Fosfomisin bereaksi
dengan residu Cys115.
File berformat .pdb memiliki data-data yang dibutuhkan untuk keperluan
perancangan obat. Biasanya terdapat data dari posisi atom, koordinat molekul,
rantai-rantai asam amino dari suatu makromolekul. Terdapat juga data jumlah dari
atom protein, asam nukleat atom, heterogen atom, dan solvent atom. Bagian
terpenting yang terkandung adalah susunan dari rantai asam amino yang
membentuk rantai peptida dari makromolekul tersebut.
Kestabilan struktur enzim MurA ligase dilihat dari plot Ramachandran
(Gambar 3A). Visualisasi plot Ramachandran diunduh pada PDB. Diagram
Ramachandran memiliki empat kuadran, yakni most favoured regions pada kuadran
I, additional allowed regions pada kuadran II, generously allowed regions pada
kuadran III, dan disallowed regions pada kuadran IV. Phi (Φ) menyatakan sumbu
x, sedangkan psi (ψ) menyatakan sumbu y dari asam amino pada struktur protein
(Bosco & Brasseur 2005). MurA ligase bersifat stabil terlihat dari data asam amino
yang berada pada daerah yang paling disukai (favorable) sebesar 97.8% (407/406)
dan daerah yang diizinkan (allowed) sebesar 100.0% (416/416) serta daerah yang
tidak diijinkan (disallowed) kurang dari 1%. Semakin besar residu asam amino
yang berada pada most favored region dan semakin rendah persentase residu pada
disallowed region maka kualitas struktur semakin bagus dan stabil (Bosco &
Brasseur 2005).
Potensi ligan untuk dapat masuk ke dalam membran sel dan diserap diuji
menggunakan software Lipinski dengan parameternya adalah berat molekul,
jumlah gugus donor ikatan hidrogen, jumlah gugus penerima ikatan hidrogen, nilai
log P dan refrakfititas molar. Hasil filter Lipinski pada Tabel 6 menunjukkan bahwa
ligan fosfomisin dan gingerglikolipid B tidak memenuhi aturan Lipinski, sementara
ligan zerumbon memenuhi aturan Lipinski. Ligan fosfomisin memiliki nilai
refraktivitas molar dibawah 40, sehingga dinilai tidak optimal diserap dalam tubuh.
Sementara itu, ligan gingerglikolipid B memiliki massa atom relatif dan nilai
refraktivitas molar yang lebih besar dari yang disyaratkan sehingga disarankan
untuk tidak menjadi kandidat obat pengganti fosfomisin karena tidak akan optimal
diserap dalam tubuh.
Molecular docking memprediksi orientasi dari suatu molekul ke molekul
yang lain ketika berikatan membentuk kompleks yang stabil (Funkhouser 2007).
Analisis energi afinitas (ΔGbinding) hasil penambatan molekular untuk ligan alami,

20
ligan obat, ligan zerumbon, dan ligan gingerglikolipid B berturut-turut adalah
senilai -10.1, -4.7, -8.3, dab -8,4 kkal mol-1. Score energi bebas yang diperoleh
menggambarkan interaksi dan aktivitas biologis dari ligan terhadap reseptor. Nilai
energi bebas Gibbs yang kecil menunjukkan bahwa konformasi yang terbentuk
adalah stabil, sedangkan makin besarnya nilai energi bebas Gibbs menunjukkan
tidak stabilnya kompleks yang terbentuk (Meng et al. 2011; Yanuar 2012).
Interaksi antara reseptor dengan ligan dikendalikan oleh pengaturan interaksi
molekuler yang kompleks pada daerah binding pocket (Scheneider dan Baringhaus
2008). Interaksi yang spesifik di daerah binding pocket dikenal sebagai direct
docking. Pendekatan direct docking dilakukan dengan penentuan grid box pada
kantung ikatan ligan dari protein. Grid box digunakan untuk mengarahkan senyawa
ligan agar berinteraksi atau terikat ke daerah katalitik reseptor (Scheneider dan
Baringhaus 2008; Meng et al. 2011). Hasil interaksi penambatan molekul ligan
dengan MurA ligase berupa jenis ikatan, panjang ikatan, residu asam amino enzim
yang berinteraksi, dan gugus fungsi ligan yang berinteraksi. Hasil penambatan
molekuler memperlihatkan adanya interaksi molekul ligan obat dengan enzim
MurA ligase seperti terlihat pada Tabel 7 dan lampiran 11, 12, 13, serta 14.
Interaksi ligan obat dan ligan uji berbeda dalam menempati kantung sisi aktif enzim
tempat terikatnya substrat atau ligan alami. Ligan uji zerumbon memiliki kesamaan
interaksi dengan 8 residu asam amino substrat (50%) pada sisi aktif kompleks
enzim. Kesamaan residu interaksi antara ligan zerumbon dengan substrat MurA
menunjukkan bahwa ligan zerumbon menempati sisi aktif yang sama dengan
substrat pada enzim sehingga menjadi inhibitor kompetitif pada enzim MurA.
Interaksi ligan uji zerumbon dengan residu asam amino dalam kantung sisi
aktif enzim adalah sama dengan interaksi ligan obat dan ligan alami. Kesamaan
residu asam amino yang terikat pada zerumbon dan ligan obat yakni residu Arg91,
sementara dengan ligan alami yakni residu His125. Ligan alami mengikat residu
asam amino yang sama dengan tempat terikat zerumbon dan ligan obat pada asam-
asam amino Arg120 dan His125. Hasil ini menunjukkan bahwa terjadi mekanisme
reaksi kompetitif antara ligan uji zerumbon dengan substrat (ligan alami) pada
enzim karena menempati tempat yang sama dengan substrat. Ligan obat mengikat
residu asam amino Arg120 sehingga substrat terhalang untuk mengikat residu asam
amino pada tempat yang sama. Ini menunjukkan bahwa ligan obat selain berperan
sebagai analog untuk substrat dengan cara alkilasi balik gugus tiol asam amino
Cys115, juga terjadi reaksi juga terjadi reaksi kompetitif antara ligan obat dengan
substrat.
Ligan gingerglikolipid B memiliki kesaman tempat pengikatan dengan ligan
obat pada sisi aktif enzim, yang ditandai dengan kesamaan mengikat residu asam
amino Arg91. Diduga ligan ini juga menempati tempat yang sama dengan ligan
alami dari enzim MurA, yang mana terlihat juga dari kesamaan residu interaksi
hidrofobik, yakni Asn23, Arg91, dan Phe328.
Dilihat dari nilai energi affinitas, ∆Gbinding Gingerglikolipid B lebih baik dari
∆Gbinding zerumbon, namun perlu dipertimbangkan lagi untuk digunakan sebagai
obat pengganti fosfomisin karena tidak memenuhi aturan Lipinski. Aturan Lipinski
mengatur terdistribusinya obat dalam tubuh. Oleh karena itu, dapat diasumsikan
bahwa zerumbon lebih mampu berdistribusi di dalam tubuh, sehingga lebih
dipertimbangkan untuk menjadi kandidat obat pengganti obat fosfomisin. Senyawa
zerumbon lebih dipilih untuk dipertimbangkan sebagai kandidat obat penghambat

21
sintesis dinding sel bakteri karena dilihat dari hasil interaksi antara zerumbon
dengan MurA, dimana zerumbon menempati tempat atau berikatan dengan residu-
residu asam amino yang sama dengan substrat maupun obat fosfomisin.
Hasil simulasi molecular docking yang menunjukkan bahwa senyawa
zerumbon dapat bertindak sebagai inhibitor kompetitif terhadap substrat enzim
MurA. Ini mengakibatkan zerumbon dapat menghambat kerja enzim MurA
sehingga mengganggu biosintesis peptidoglikan. Hasil simulasi molecular docking
berkorelasi positif dengan hasil uji in vitro antibakteri. Ekstrak etanol Z. zerumbet
sangat menghambat bakteri Gram positif dibanding pada bakteri Gram negatif
(Tabel 2). Zerumbon merupakan komponen utama pada ekstrak etanol Z. zerumbet.
Salah satu pembeda antara bakteri Gram positif dan Gram negatif adalah kandungan
peptidoglikan pada dinding sel bakteri. Peptidoglikan merupakan penyusun
terbesar (90%) dinding sel bakteri Gram positif dan hanya 5-10% pada bakteri
Gram negatif. Terhambatnya sintesis peptidoglikan oleh zerumbon mengakibatkan
terbentuknya sel-sel yang peka terhadap tekanan osmosis, terutama pada bakteri
Gram positif yang mana kekuatan dinding selnya berasal dari peptidoglikan
sehingga menyebabkan terjadinya lisis yang mengakibatkan kematian bakteri.
5 SIMPULAN dan SARAN
Simpulan
Rimpang Z. zerumbet asal Pulau Timor yang diekstrak dengan pelarut air,
etanol, etil asetat, dan n-heksana memiliki aktivitas antibakteri. Ekstrak etanol
memiliki aktivitas antibakteri yang lebih besar daripada ekstrak lainnya. Nilai
KHTM ekstrak etanol terhadap S. aureus, B. subtilis, E. coli, P. aeruginosa secara
berurutan adalah 50, 100, 150, 250 mg mL-1. Hasil analisis LC-MS ekstrak etanol
menunjukkan bahwa senyawa yang terkandung dalam ekstrak adalah zerumbon dan
gingerglikolipid B. Simulasi docking menunjukkan bahwa terjadi mekanisme
reaksi kompetitif antara ligan uji zerumbon dengan substrat uridine-diphosphate-
N-asetylglucosamine pada enzim karena ligan uji menempati tempat yang sama
dengan substrat sehingga menghambat terbentuknya peptidoglikan penyusun
dinding sel bakteri.
Saran
Belum diketahuinya salah satu komponen metabolit sekunder ekstrak etanol
hasil LCMS pada m/z 717.41 merupakan hal yang menarik untuk dikaji lebih lanjut
dan juga perlu penelitian lebih lanjut mengenai aktivitas antibakteri menggunakan
ekstrak segar tanaman Z. zerumbet asal Pulau Timor.

22
DAFTAR PUSTAKA
Abdul A B, Abdelwahab S I, Al-Zubairi A S, Elhassan M M, Murali S M. 2008.
Anticancer and antimicrobial activities of Zerumbone from the rhizomes of
Zingiber zerumbet. Int, J. Pharmacol., 4 (4): 301-304
Bosco KH, Brasseur R. 2005. The Ramachandran plots of glycine and pre-proline.
BMC Struc Bio. 5:1-14.
Brown ED, Vivas EI, Walsh CT, Kolter R. 1995. MurA (MurZ), the Enzyme That
Catalyzes the First Committed Step in Peptidoglycan Biosynthesis, Is
Essential in Escherichia coli. J. Bacteriol. 177(14): 4194–4197.
Chen IN, Chan CC, Ng CC, Wang CY, Shyu YT, Chang TL. 2008. Antioxidant and
Antimicrobial activity of Zingiberaceae Plants in Taiwan. Plant Foods Hum
Nutr. 63: 15-20
Clinical and Laboratory Standards Institute (CLSI). 2007. Performance standards
for antimicrobial susceptibility testing; seventeenth informational
supplement. M100-S17. Vol. 27 No. 1
Cowan M M. 1999. Plant products as antimicrobial agents. Clin. Microbiol. Rev.
Vol 12, No. 4
de Pedro MA, Cava F. 2015. Structural constraints and dynamics of bacterial cell
wall architecture. Front Microbiol., 6: 1-10. doi:
10.3389/fmicb.2015.00449.
de Fatima A, Modo LV, Conegero LS, Pilli RA, Ferreira CV, Kohn LK, de
Carvalho JE. 2006. Lactones and their derivatives biological activities,,
mechanism of action and potential leads for drugs design. J. Med. Chem.
(13): 3371-3384
Eschenburg S, Priestman MA, Abdul-Latif FA, Delachaume C, Fassy F,
Schonbrunn E. 2005. A Novel Inhibitor That Suspends the Induced Fit
Mechanism of UDP-N-acetylglucosamine Enolpyruvyl Transferase
(MurA). J Biol Chem. 280 (14): 14070–14075. doi:
10.1074/jbc.M414412200.
Funkhouser T. 2007. Lecture: Protein-ligand docking methods. Pricenton
University
Griffin S. 2000. Aspect of antimicrobial activity of terpenoids and the relationship
to their molecular structure [Disertation]. New South Wales (AU):
University of Western Sidney
Harborne J B. 1996. Metode Fitokimia: penuntun cara modern menganalisis
tumbuhan, diterjemahkan oleh Kosasih Padmawinata dan Iwang Soediro.
Bandung (ID): Penerbit ITB
Hashemi SR, Zulkifli I, Hair Bejo M, Farida A, Somchit MN. 2008. Acute toxicity
study and phytochemical screening of selected herbal aqueous extract in
broiler chikens. Int. J. Pharmacol. 4(5): 352-360.
Kader M G, Habib M R, Nikkon F, Yeasmin T, Rashid M A, Rahman M M,
Gibbons S. 2010. Zedorone from the rhizomes of Zingiber zerumbet and its
antistaphylococcal activity. BLACPMA, 9 (1), 63-68
Kader M G, Nikkon F, Rashid M A, Yeasmin T. 2011. Antimicrobial activities of
the rhizome extract of Zingiber zerumbet Linn. Asian Pac J Trop Biomed.
1(5):409-412

23
Kim SJ, Chang J, Singh M. 2015. Peptidoglycan architecture of Gram-positive
bacteria by solid-state NMR. Biochim Biophys Acta. 1848: 350-362. doi:
10.1016/j.bbamem.2014.05.031.
Kitayama T, Yamamoto K, Utsumi R, Takatani M, Hill R K, Kawai Y, Sawada S,
Okamoto T. 2001. Chemistry of Zerumbone. 2. Regulation of ring bond
cleavage and uniqui antibacterial activities of zerumbone derivatives.
Biosci. Biotechnol. Biochem., 65 (10), 2193-2199
Krees WJ, Prince LM, Wiliam KJ. 2002. Phylogeny and a new classification of the
gingers (zingiberaceae): evidance from molecular data. Am. J. Botany 89
(11): 1682-1696.
Kress WJ, 2014. Zingiberales Research Website. Washington DC, USA:
Smithsonian Institution.
http://botany.si.edu/zingiberales/families/familypage.cfm?myfamily=Zingi
beraceae
Kumar DR, Lakshmi PS, Saravani N, Marimuthu S. 2012. In silico Molecular
Docking on Porcine pancreatic phospholipase A2 against plant extract of
phenolic inhibitor. IJRBB. 2 (3): 8-16.
Larsen K, Ibrahim H, Khaw SH, Saw LG. 1999. Pollination and seed dispersal, p.
19-20. In: K. M. Wong (Ed.). Ginger of Penninsular Malysia and Singapura.
Natural History Publication (Borneo). Kinabalu
Lawalata V N. 2012. Rekayasa proses ekstraksi kulit buah Langsat (Lansium
domesticum var. langsat) sebagai bahan antibakteri dan antioksidan.
[disertasi].Bogor (ID): Program pascasarjana, Institut Pertanian Bogor
Lipinski CA, Lombardo F, Segawa T, Ko D. 2001. Experimental and computational
approaches to estimate solubility and permeability in drug discovey and
development setting. Adv Drug Deliv Rev . 46: 3-26.
Kumar DR, Lakshmi PS, Saravani N, Marimuthu S. 2012. In silico Molecular
Docking on Porcine pancreatic phospholipase A2 against plant extract of
phenolic inhibitor. IJRBB. 2 (3): 8-16.
Margolin W. 2009. Sculpting the bacterial cell. Curr Biol. 19(17): 812-822. doi:
10.1016/j.cub.2009.06.033.
Meng XY, Zhang HX, Mezei M, Cui M. 2011. Molecular docking: a powerful
approach for structure-based drug discovery. Curr Comput Aided Drugs
Des. 7(2):146-157
Nag A, Bandyopadhyay M, Mukherjee A. 2013. Antioxidant activities and
cytotoxicity of Zingiber zerumbet (L.) Smith rhizome. J Pharmacogn
Phytochem. 2 (3): 102-108
Naggapan T, Ramasamy P, Wahid MEA, Segaran TC, Vairappan CS. 2011.
Biological activity of carbazole alkaloids and essential oil of Murraya
koenigii againts antibiotic resistant microbes and cancer cell lines. J.
Molecule. (16): 9651-9664
Nursal W, Sri, Wilda S. 2006. Bioaktifitas Ekstrak Jahe (Zingiber officinale Roxb.)
Dalam Menghambat Pertumbuhan Koloni Bakteri Escherichia coli dan
Bacillus subtilis. Jurnal Biogenesis 2(2): 64-66
Parhusip AJN. 2006. Kajian mekanisme antibakteri ekstrak andaliman
(Zanthoxylum acanthopodium DC) terhadap bakteri patogen pangan
[disertasi]. Bogor (ID): Sekolah Pascarjana Institut Pertanian Bogor.

24
Sari KIP, Periadnadi, Nasir N. 2013. Uji antimikroba ekstrak segar jahe-jahean
(Zingiberaceae) terhadap S. aureus, E. coli dan C. albicans. J. Bio. UA. 2
(1)-Maret 2013
Selmar D. 2008. Potensial of salt and drought stress to increase pharmaceutical
significant secondary coumpounds in plants. Agriculture and forestry
research. 58:139-144
Sinaga E, Suprihatin, Wiryanti I. 2011. Perbandingan daya sitotoksik ekstrak
rimpang 3 jenis tumbuhan Zingiberaceae terhadap sel kanker MCF-7.
Jurnal Farmasi Indonesia Vol. 5 No. 3. Januari 2011: 125-133
Singh CB, Nongalleima Kh, Brojendrosingh S, Ningobam S, Lokendrajit N, Singh
LW. 2012 Biological and chemical properties of Zingiber zerumbet Smith:
a review. Phytochem Rev. 11:113-125
Skarzynski T, Mistry A, Wonacott A, Hutchinson SE, Kelly VA, Duncan K. 1996.
Structure of UDP-N-acetylglucosamine enolpyruvyl transferase, an enzyme
essential for the synthesis of bacterial peptidoglycan, complexed with
substrate UDP-N-acetylglucosamine and the drug fosfomycin. Structure. 4:
1465-1474.
Sopandie D. 2014. Fisiologi Adaptasi Tanaman Terhadap Cekaman Abiotik pada
Agroekositem Tropika. Bogor (ID): IPB Press
Staniek A, Bouwmeester, Fraser PD, Kayser O, Martens S, Tissier A, Van der Krol
S, Wessjohann L, Warzecha H. 2013. Natural-products-modifying metabolite
pathways in plants. Biotechnol. J. 8: 1159-1171
Vijesh AM, Isloor AM, Sandeep T, Arulmoli T, Fun H-K. 2013. Molecular docking
studies of some new imidazole derivatives for antimicrobial properties.
Arabian J Chem. 6(2): 197-204. doi: 10.1016/j.arabjc.2011.10.007.
Vishwanatha H N, Niraguna B P, Gowrishankar B S, Shridhar S B. 2012.
Antimicrobial activity of zerumbone from zingiber zerumbet against
staphylococcus epidermidis and Aspergillus spp. IJABPT, Vol 3, issue 4,
Oct-Dec 2012
Vollmer W, Blanot D, de Pedro MA. 2008. Peptidoglycan structure and
architecture. FEMS Microbiol Rev. 32(2): 149-167. doi: 10.1111/j.1574-
6976.2007.00094.x.
Voravuthikuncai SP, Limsuwan S, Supapol O, Subhadhirasakul S. 2006.
Antibacterial activity of extracts from family Zingiberaceae against
foodborne pathogens. J Food Safety. 26: 325-334.
Yanuar A. 2012. Penambatan Molekular: Praktek dan Aplikasi pada Virtual
Screening. Jakarta (ID): Fakultas Farmasi Universitas Indonesia
Yasni S. 2013. Teknologi Pengolahan dan Pemanfaatan Produk Ekstraktif
Rempah. Bogor: IPB Press.
Yob N J, Jofrry S. M, Affandi M M R, Teh L K, Salleh M Z, Zakaria Z A. Zingiber
zerumbet (L.) Smith: A Review of Its Ethnomedicinal, Chemical, and
Pharmacological Uses. Doi: 10.1155/2011/543216
Yoshikawa M, Yamaguchi S, Kunimi K, Matsuda H, Okuno Y, Yamahara J,
Murakami N. 1994. Stomachic principles in Ginger III. An anti-ulcer
principle, 6-Gingesulfonic acid, and three monoacyldigalactosylglycerols,
Gingerglycolipids A, B, and C, from Zingiberis Rhizoma Originating in
Taiwan. Chem Pharm Bull. 42 (6) 1226-1230

25
LAMPIRAN
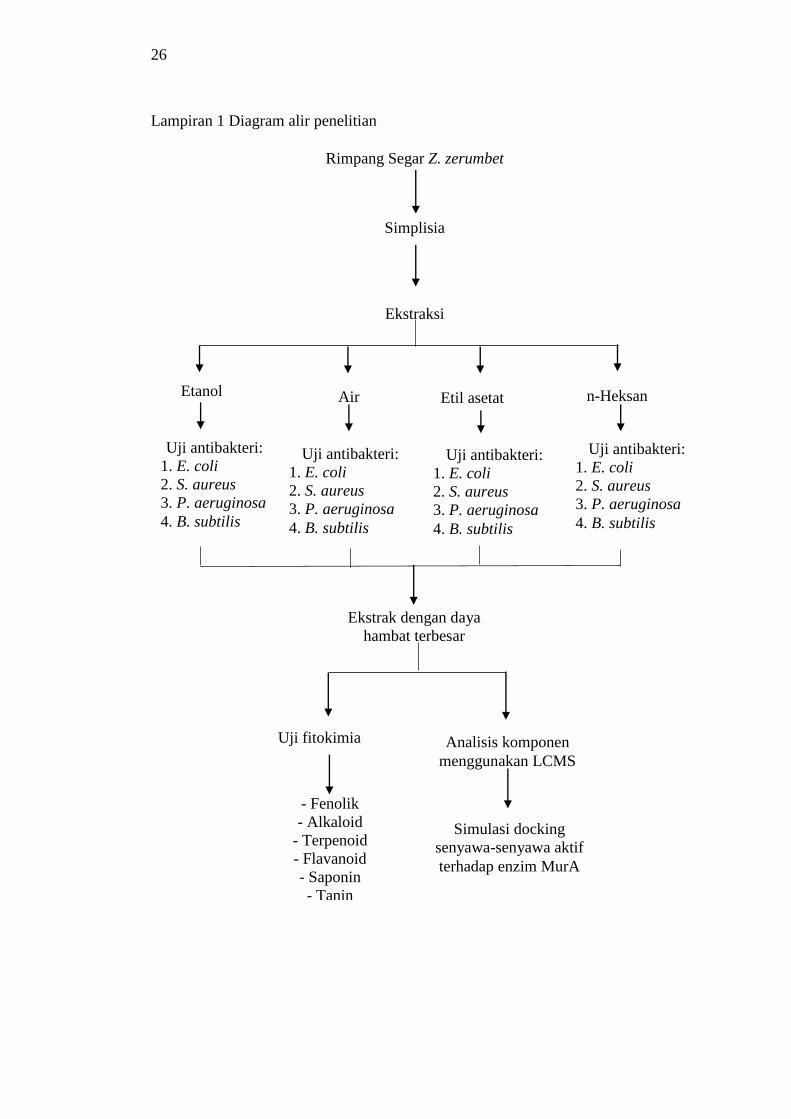
26
Lampiran 1 Diagram alir penelitian
Ekstraksi
Etanol Air n-Heksan
Uji fitokimia
Ekstrak dengan daya
hambat terbesar
Simplisia
Rimpang Segar Z. zerumbet
Analisis komponen
menggunakan LCMS
- Fenolik
- Alkaloid
- Terpenoid
- Flavanoid
- Saponin
- Tanin
Simulasi docking
senyawa-senyawa aktif
terhadap enzim MurA
Uji antibakteri:
1. E. coli
2. S. aureus
3. P. aeruginosa
4. B. subtilis
Uji antibakteri:
1. E. coli
2. S. aureus
3. P. aeruginosa
4. B. subtilis
Uji antibakteri:
1. E. coli
2. S. aureus
3. P. aeruginosa
4. B. subtilis
Etil asetat
Uji antibakteri:
1. E. coli
2. S. aureus
3. P. aeruginosa
4. B. subtilis

27
Lampiran 2 Hasil identifikasi tanaman

28
Lampiran 3 Ekstrak kasar n-heksan, etil asetat, etanol, dan aquades tanaman Z.
zerumbet
Lampiran 4 Profil dan kromatogram hasil analisis LCMS ekstrak etanol Z.
zerumbet
a
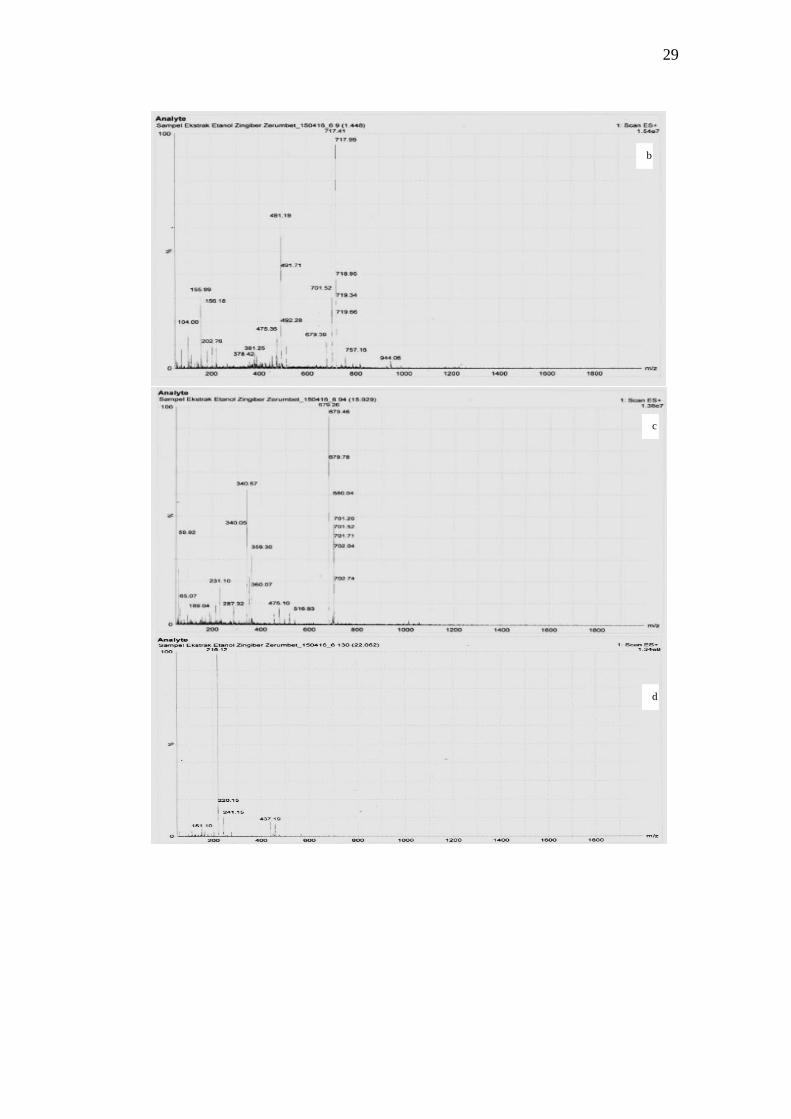
29
c
b
d
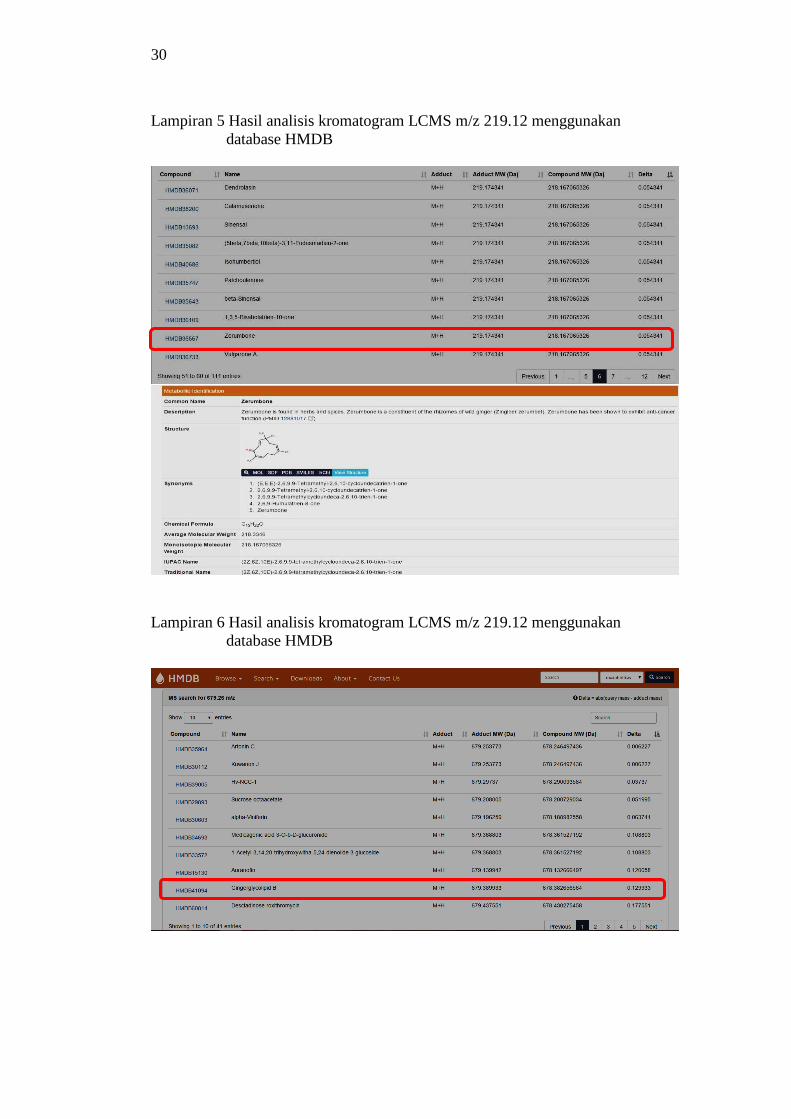
30
Lampiran 5 Hasil analisis kromatogram LCMS m/z 219.12 menggunakan
database HMDB
Lampiran 6 Hasil analisis kromatogram LCMS m/z 219.12 menggunakan
database HMDB
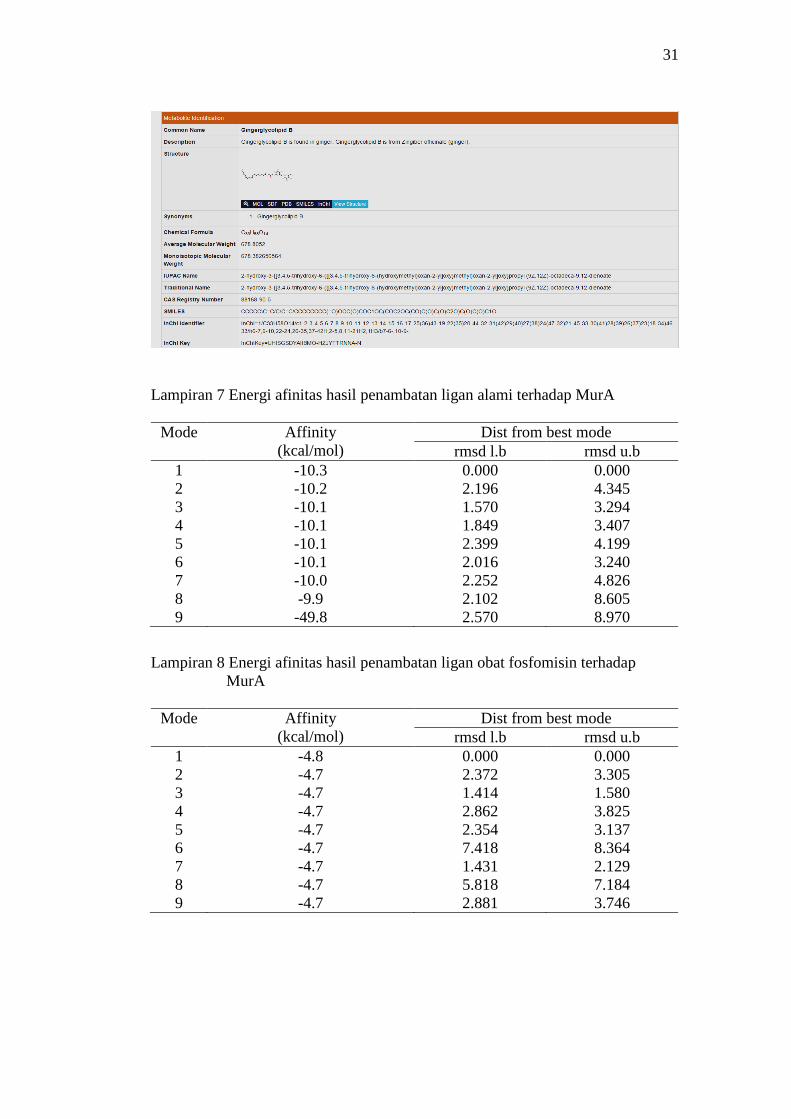
31
Lampiran 7 Energi afinitas hasil penambatan ligan alami terhadap MurA
Mode Affinity
(kcal/mol)
Dist from best mode
rmsd l.b rmsd u.b
1 -10.3 0.000 0.000
2 -10.2 2.196 4.345
3 -10.1 1.570 3.294
4 -10.1 1.849 3.407
5 -10.1 2.399 4.199
6 -10.1 2.016 3.240
7 -10.0 2.252 4.826
8 -9.9 2.102 8.605
9 -49.8 2.570 8.970
Lampiran 8 Energi afinitas hasil penambatan ligan obat fosfomisin terhadap
MurA
Mode Affinity
(kcal/mol)
Dist from best mode
rmsd l.b rmsd u.b
1 -4.8 0.000 0.000
2 -4.7 2.372 3.305
3 -4.7 1.414 1.580
4 -4.7 2.862 3.825
5 -4.7 2.354 3.137
6 -4.7 7.418 8.364
7 -4.7 1.431 2.129
8 -4.7 5.818 7.184
9 -4.7 2.881 3.746
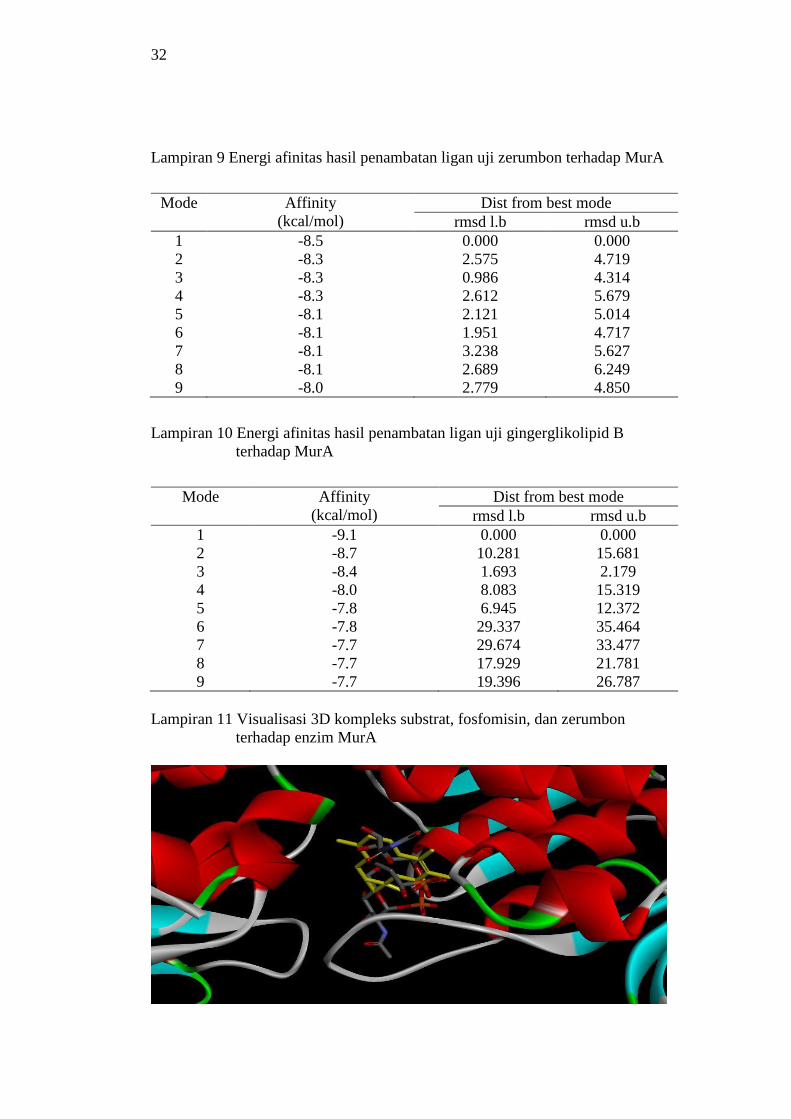
32
Lampiran 9 Energi afinitas hasil penambatan ligan uji zerumbon terhadap MurA
Mode Affinity
(kcal/mol)
Dist from best mode
rmsd l.b rmsd u.b
1 -8.5 0.000 0.000
2 -8.3 2.575 4.719
3 -8.3 0.986 4.314
4 -8.3 2.612 5.679
5 -8.1 2.121 5.014
6 -8.1 1.951 4.717
7 -8.1 3.238 5.627
8 -8.1 2.689 6.249
9 -8.0 2.779 4.850
Lampiran 10 Energi afinitas hasil penambatan ligan uji gingerglikolipid B
terhadap MurA
Mode Affinity
(kcal/mol)
Dist from best mode
rmsd l.b rmsd u.b
1 -9.1 0.000 0.000
2 -8.7 10.281 15.681
3 -8.4 1.693 2.179
4 -8.0 8.083 15.319
5 -7.8 6.945 12.372
6 -7.8 29.337 35.464
7 -7.7 29.674 33.477
8 -7.7 17.929 21.781
9 -7.7 19.396 26.787
Lampiran 11 Visualisasi 3D kompleks substrat, fosfomisin, dan zerumbon
terhadap enzim MurA
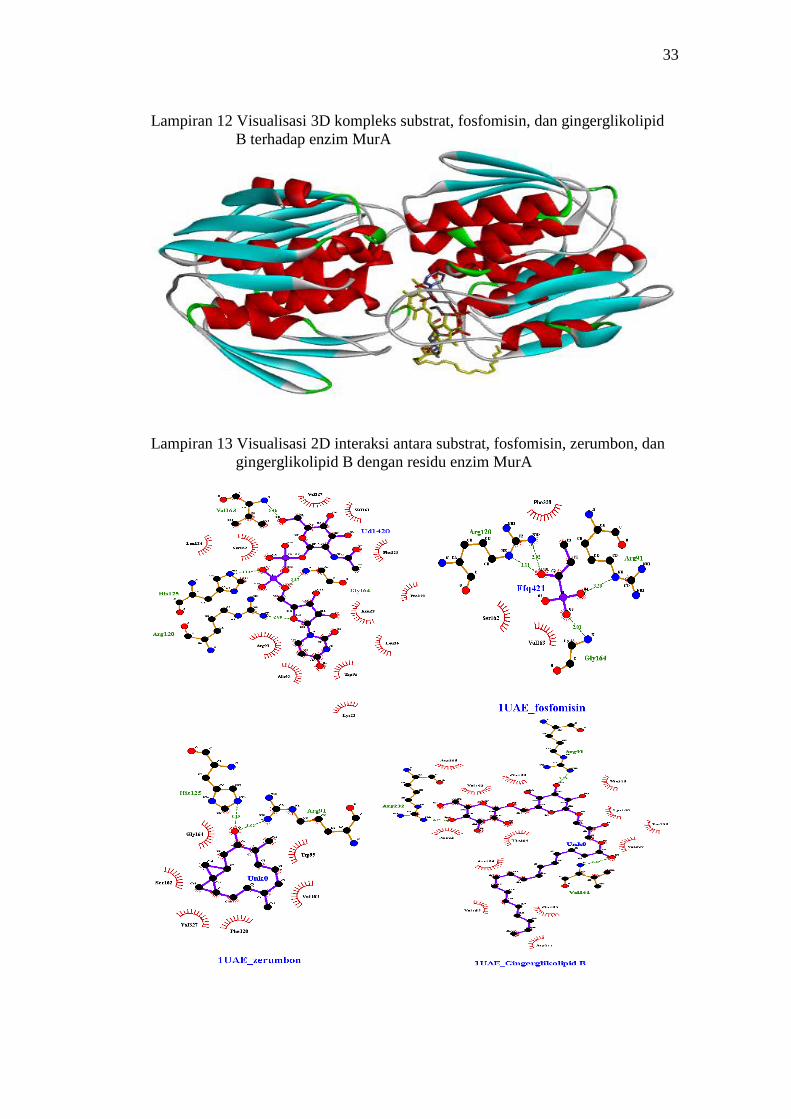
33
Lampiran 12 Visualisasi 3D kompleks substrat, fosfomisin, dan gingerglikolipid
B terhadap enzim MurA
Lampiran 13 Visualisasi 2D interaksi antara substrat, fosfomisin, zerumbon, dan
gingerglikolipid B dengan residu enzim MurA
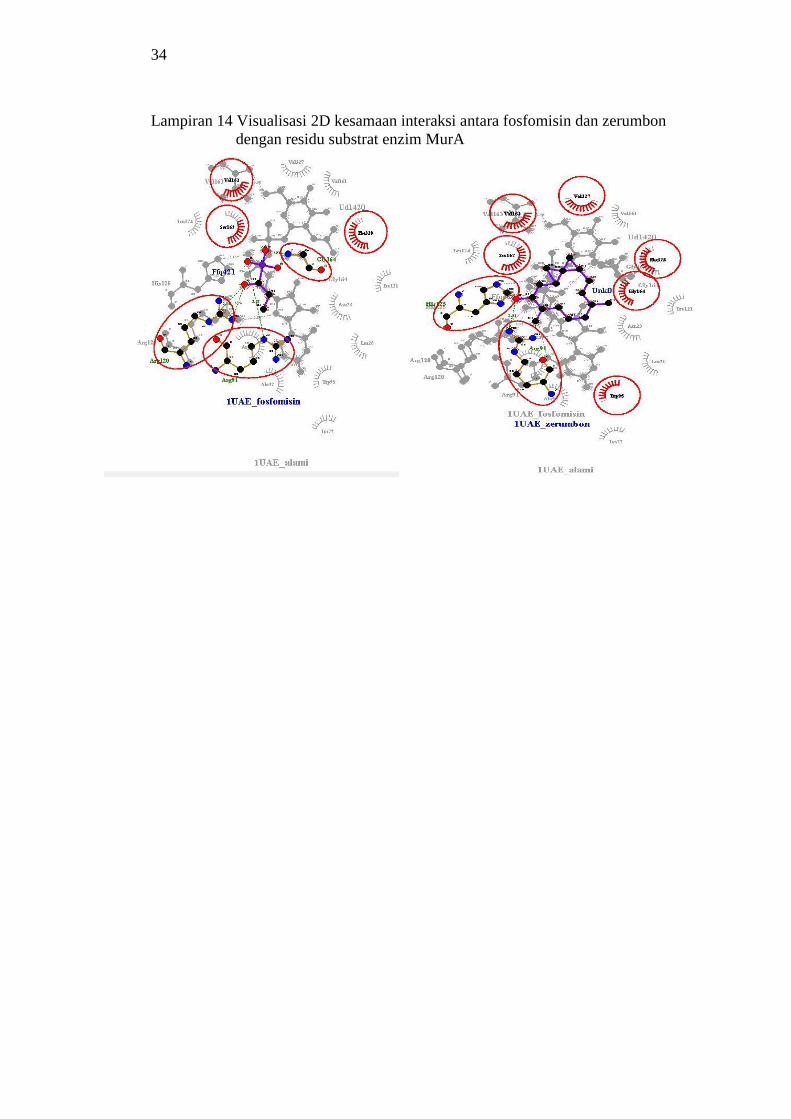
34
Lampiran 14 Visualisasi 2D kesamaan interaksi antara fosfomisin dan zerumbon
dengan residu substrat enzim MurA

35
RIWAYAT HIDUP
Penulis dilahirkan di Soba-Amarasi, 2 Oktober 1988 dari ayah David Jibrael
Kapitan dan Ibu Martha Kadrida Kapitan-Ottemusu. Penulis merupakan putra
keempat dari empat bersaudara. Penulis menyelesaikan pendidikan menengah atas
di SMAN 2 Kupang tahun 2006. Penulis meneruskan pendidikan di Jurusan Kimia,
Fakultas Sains dan Teknik, Universitas Nusa Cendana tahun 2007 dan penulis
mendapatkan gelar sarjana (S1) pada tahun 2012. Tahun 2013 penulis melanjutkan
studinya di Departemen Biokimia, Fakultas Matematika dan Ilmu Pengetahuan
Alam, Institut Pertanian Bogor dengan beasiswa BPPDN (Beasiswa Pendidikan
Pascasarjana Dalam Negeri) DIKTI 2013.
Selama mengikuti perkuliahan penulis sering mengikuti pelatihan dan
seminar yang diadakan baik di dalam kampus maupun di luar kampus. Selan itu,
penulis juga aktif dalam kegiatan organisasi kemahasiswaan Undana sebagai
Pengurus Himpunan Mahasiswa Jurusan (2008/2009, 2009/2010), Pengurus Badan
Eksekutif Mahasiswa Fakultas (2010/2011, 2011/2012), organisasi ekstra kampus
pada Gerakan Mahasiswa Kristen Indonesia (GMKI) cabang Kupang (2008/2011).
Penulis juga pernah menjadi asisten mata kuliah praktikum Kimia Dasar
(2011/2013), Kimia Analitik I dan II (2010/2013) untuk mahasiswa Kimia, dan
Biokimia untuk mahasiswa Politeknik Kesehatan Kupang (2010/2013) dan
Fakultas Kedokteran Hewan Undana (2011/2013) di Laboratorium Kimia Undana.
Penulis juga aktif sebagai tenaga lapangan dalam pemantauan kualitas air dan udara
pada Laboratorium Kimia Undana. Selama mengikuti perkuliahan di Pascasarjana
IPB, penulis pernah mengikuti kegiatan yang diadakan oleh Forum Mahasiswa
Pascasarjana (Forum WACANA) IPB.
Penulis menulis artikel penelitian yang berjudul Uji fitokimia dan
identifikasi senyawa-senyawa metabolit sekunder ekstrak etil asetat, etanol, dan air
tanaman Zingiber zerumbet asal Pulau Timor yang dipublikasikan di jurnal Current
Biochemistry. Selain itu penulis juga mempublikasikan tulisan yang berjudul
Inhibition docking simulation of Zerumbone, Gingerglycolipid B, and Curzerenone
compound of Zingiber zerumbet from Timor island against MurA enzyme di
Journal of Applied Chemical Science pada Volume 3 No. 1: 9-19.