4 hasil dan pembahasan - repository.ipb.ac.id · dengan cover glass. cover glass ditekan dengan...
TRANSCRIPT

12
letakkan di gelas arloji yang telah ditetesi aseto orcein 2% kemudian ditutup dengan cover glass. Cover glass ditekan dengan tangan agar tidak bergeser dan dipukul-pukul menggunakan ujung penghapus pensil agar sel-sel terpisah. Bersihkan sisa aseto orcein 2% yang menempel diluar cover glass menggunakan kertas saring kemudian dipanaskan sedikit menggunakan api bunsen. Cover glass ditekan halus dan dihangatkan lagi sedikit. Sekeliling cover glass diberikan cat kuku bening dan dikeringkan agar tidak bergeser. Preparat diamati menggunakan mikroskop pada perbesaran 10 x 100. Pengamatan dilakukan pada lima sel dari setiap preparat
4 HASIL DAN PEMBAHASAN
Kondisi Umum
Planlet (tunas pucuk dan tunas samping) dan embrio somatik jeruk Siam Simadu yang digunakan pada penelitian ini berasal dari planlet dan embrio somatik yang tersedia di Laboratorium Biologi Sel dan Jaringan, BB-Biogen. Planlet dan embrio somatik tersebut berasal dari nuselus yang diisolasi dari buah muda jeruk Siam Simadu yang telah berumur 30-90 hari setelah anthesis. Tunas pucuk, tunas samping, dan embrio somatik jeruk Siam Simadu yang telah diberikan perlakuan kolkisin disimpan pada ruang kultur dengan penyinaran menggunakan lampu 20 watt selama 16 jam/hari dengan intensitas cahaya rata-rata 1500-2000 lux, dengan suhu 16-20 oC. Laboratorium tempat dilakukannya penelitian sangat menjaga kestabilan suhu ruang kultur supaya tetap terjaga pada kisaran 16 - 20oC (Gambar 5). Kestabilan suhu ruang kultur tersebut dibantu dengan kondisi Air Conditioner (AC) yang tetap dihidupkan selama 24 jam.
Gambar 5 Kondisi ruang kultur dan ruang tanam
Tingkat kontaminasi yang terjadi cukup tinggi. Kontaminasi merupakan gangguan utama yang terjadi pada eksplan yang ditanam secara in vitro. Kontaminasi pada media mengganggu pertumbuhan eksplan bahkan dapat menyebabkan kematian. Kontaminasi disebabkan oleh cendawan dan bakteri yang tumbuh di permukaan media atau disekeliling

13
eksplan. Keberadaan cendawan ditandai dengan munculnya cendawan berwarna putih, hijau, dan hitam di permukaan media. Kemungkinan terjadinya kontaminasi disebabkan oleh larutan kolkisin yang kurang steril, wadah yang digunakan untuk menyimpan larutan kolkisin kurang steril, kertas saring yang digunakan kurang steril, eksplan belum benar-benar kering ketika ditiriskan saat perlakuan sehingga media menjadi berair dan meyebabkan kontaminasi.
Kendala yang terjadi secara umum saat penelitian adalah kesulitan mengukur pH larutan kolkisin hingga 5.6. Kendala yang terjadi saat induksi tetraploid tunas pucuk adalah kesulitan memotong daun terkecil tunas pucuk karena apabila tidak hati-hati meristem apikal akan ikut terpotong (Gambar 6 A). Pemotongan daun ini bertujuan agar kolkisin mudah masuk ke dalam sel-sel meristem apikal dan mengurangi terjadinya kimera. Kendala yang terjadi saat induksi tetraploid tunas samping adalah kesulitan memotong tunas samping apabila tidak hati-hati meristem akan ikut terpotong karena kebanyakan planlet jeruk Siam Simadu yang tersedia memiliki buku (node) yang pendek (Gambar 6 B). Kendala yang terjadi saat induksi tetraploid embrio somatik adalah globular yang memiliki ukuran beragam sehingga sulit memilih globular dengan ukuran yang sama agar keragaman yang dihasilkan dari percobaan ini sedikit (Gambar 6 C).
Gambar 6 (A) Daun kecil tunas pucuk, (B) buku (node) jeruk Siam Simadu yang pendek, (C) globular yang memiliki ukuran beragam
Gambar 7 (A) Tunas samping yang membentuk kalus hijau bertekstur kompak, (B) tunas samping yang hanya membesar, (C) tunas samping yang membentuk kalus putih bertekstur remah
14
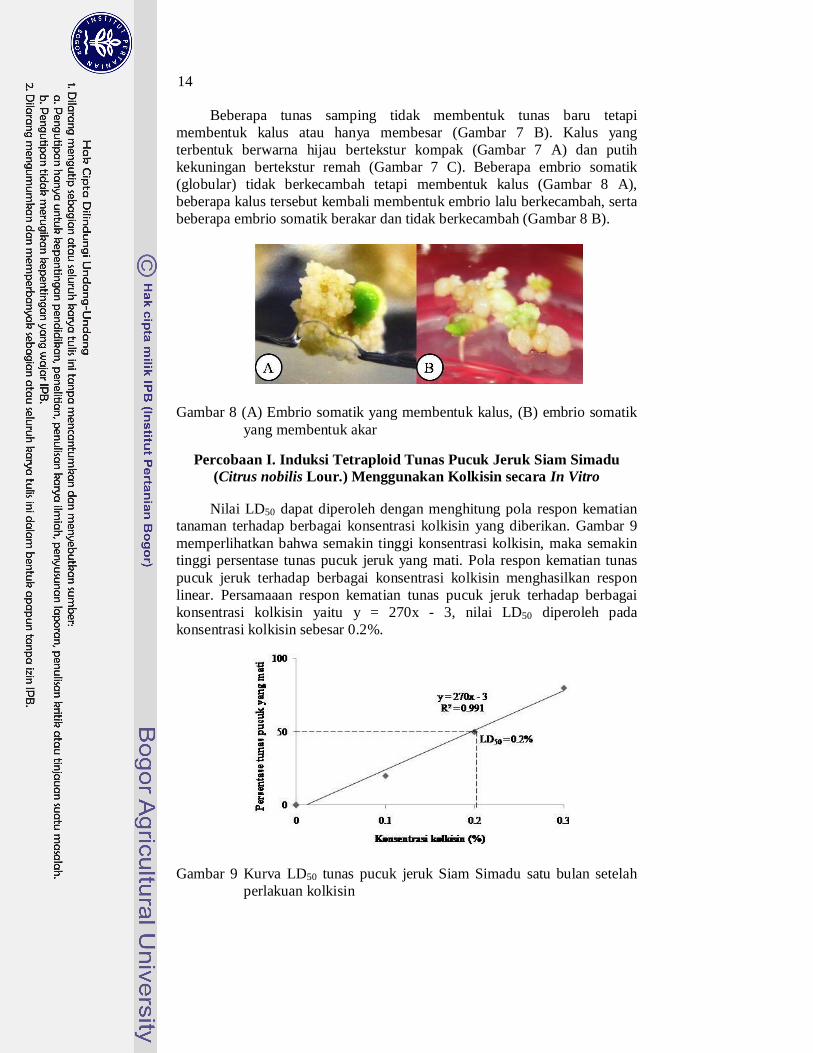
Beberapa tunas samping tidak membentuk tunas baru tetapi membentuk kalus atau hanya membesar (Gambar 7 B). Kalus yang terbentuk berwarna hijau bertekstur kompak (Gambar 7 A) dan putih kekuningan bertekstur remah (Gambar 7 C). Beberapa embrio somatik (globular) tidak berkecambah tetapi membentuk kalus (Gambar 8 A), beberapa kalus tersebut kembali membentuk embrio lalu berkecambah, serta beberapa embrio somatik berakar dan tidak berkecambah (Gambar 8 B).
Gambar 8 (A) Embrio somatik yang membentuk kalus, (B) embrio somatik
yang membentuk akar
Percobaan I. Induksi Tetraploid Tunas Pucuk Jeruk Siam Simadu (Citrus nobilis Lour.) Menggunakan Kolkisin secara In Vitro
Nilai LD50 dapat diperoleh dengan menghitung pola respon kematian tanaman terhadap berbagai konsentrasi kolkisin yang diberikan. Gambar 9 memperlihatkan bahwa semakin tinggi konsentrasi kolkisin, maka semakin tinggi persentase tunas pucuk jeruk yang mati. Pola respon kematian tunas pucuk jeruk terhadap berbagai konsentrasi kolkisin menghasilkan respon linear. Persamaaan respon kematian tunas pucuk jeruk terhadap berbagai konsentrasi kolkisin yaitu y = 270x - 3, nilai LD50 diperoleh pada konsentrasi kolkisin sebesar 0.2%.
Gambar 9 Kurva LD50 tunas pucuk jeruk Siam Simadu satu bulan setelah perlakuan kolkisin
15
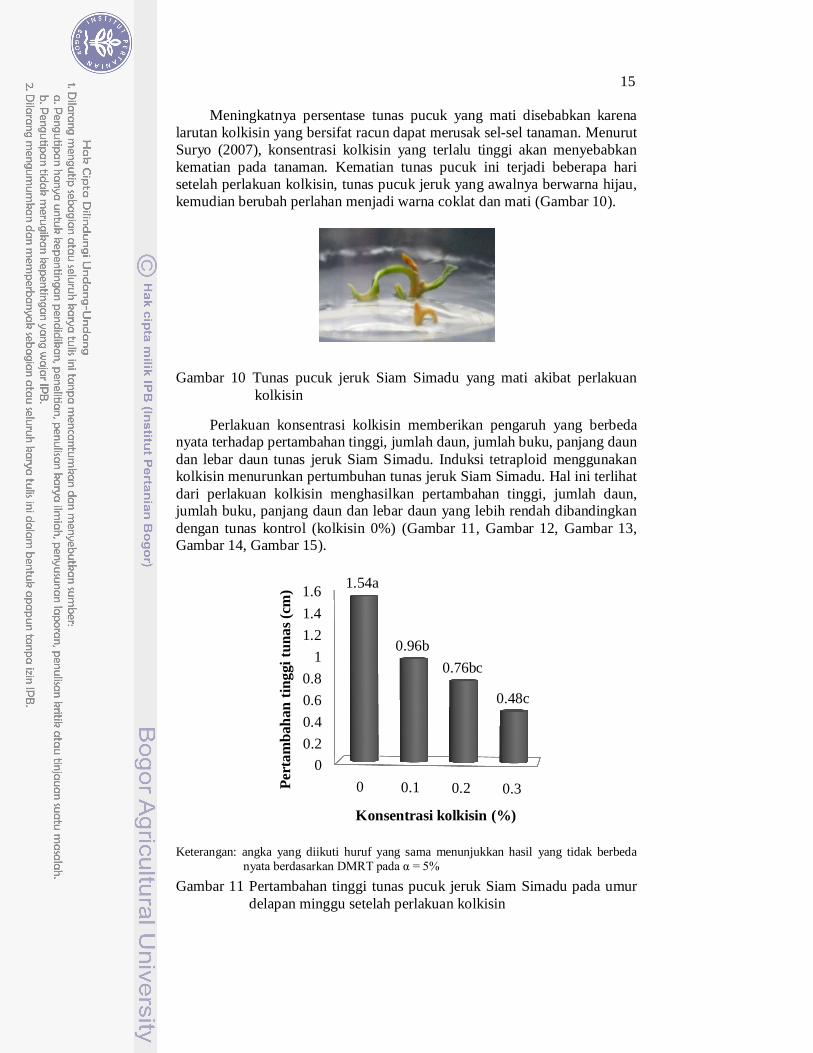
Meningkatnya persentase tunas pucuk yang mati disebabkan karena larutan kolkisin yang bersifat racun dapat merusak sel-sel tanaman. Menurut Suryo (2007), konsentrasi kolkisin yang terlalu tinggi akan menyebabkan kematian pada tanaman. Kematian tunas pucuk ini terjadi beberapa hari setelah perlakuan kolkisin, tunas pucuk jeruk yang awalnya berwarna hijau, kemudian berubah perlahan menjadi warna coklat dan mati (Gambar 10).
Gambar 10 Tunas pucuk jeruk Siam Simadu yang mati akibat perlakuan kolkisin
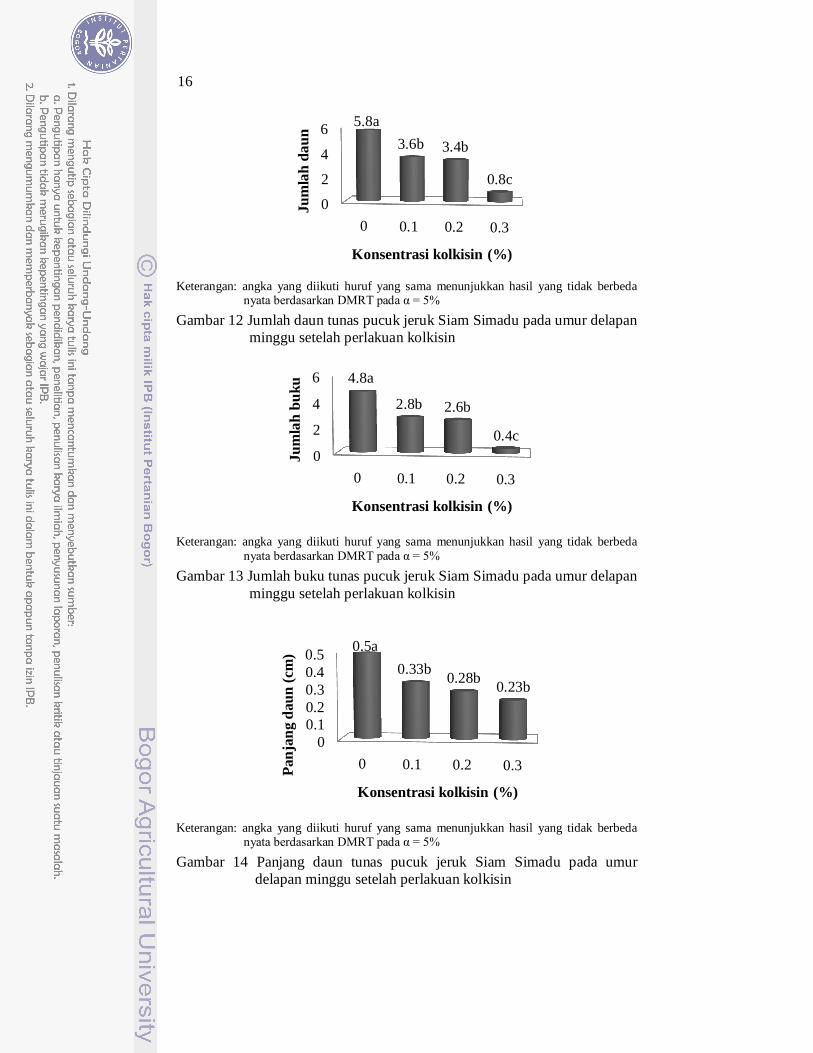
Perlakuan konsentrasi kolkisin memberikan pengaruh yang berbeda nyata terhadap pertambahan tinggi, jumlah daun, jumlah buku, panjang daun dan lebar daun tunas jeruk Siam Simadu. Induksi tetraploid menggunakan kolkisin menurunkan pertumbuhan tunas jeruk Siam Simadu. Hal ini terlihat dari perlakuan kolkisin menghasilkan pertambahan tinggi, jumlah daun, jumlah buku, panjang daun dan lebar daun yang lebih rendah dibandingkan dengan tunas kontrol (kolkisin 0%) (Gambar 11, Gambar 12, Gambar 13, Gambar 14, Gambar 15).
Keterangan: angka yang diikuti huruf yang sama menunjukkan hasil yang tidak berbeda
nyata berdasarkan DMRT pada α = 5%
Gambar 11 Pertambahan tinggi tunas pucuk jeruk Siam Simadu pada umur delapan minggu setelah perlakuan kolkisin
00.20.40.60.8
11.21.41.6
0 0.1 0.2 0.3
1.54a
0.96b0.76bc
0.48c
Pert
amba
han
tingg
i tun
as (c
m)
Konsentrasi kolkisin (%)
16
Keterangan: angka yang diikuti huruf yang sama menunjukkan hasil yang tidak berbeda
nyata berdasarkan DMRT pada α = 5%
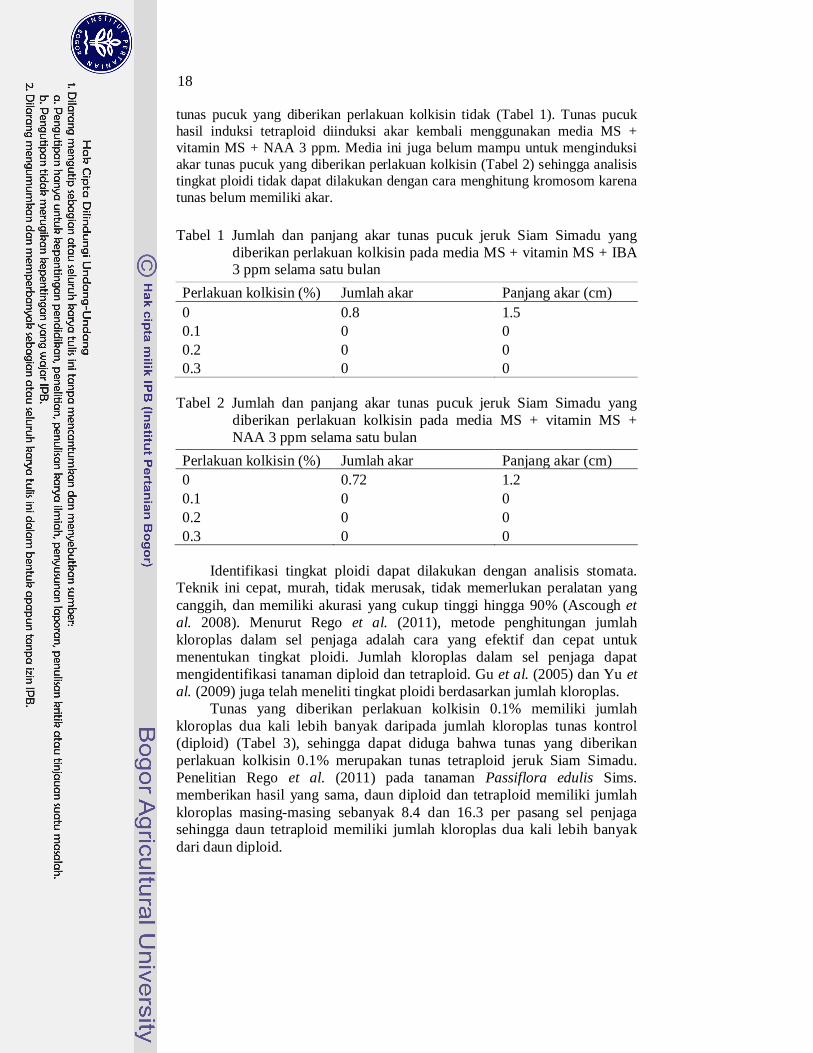
Gambar 12 Jumlah daun tunas pucuk jeruk Siam Simadu pada umur delapan minggu setelah perlakuan kolkisin
Keterangan: angka yang diikuti huruf yang sama menunjukkan hasil yang tidak berbeda
nyata berdasarkan DMRT pada α = 5%
Gambar 13 Jumlah buku tunas pucuk jeruk Siam Simadu pada umur delapan minggu setelah perlakuan kolkisin
Keterangan: angka yang diikuti huruf yang sama menunjukkan hasil yang tidak berbeda
nyata berdasarkan DMRT pada α = 5%
Gambar 14 Panjang daun tunas pucuk jeruk Siam Simadu pada umur delapan minggu setelah perlakuan kolkisin
0
2
4
6
0 0.1 0.2 0.3
5.8a3.6b 3.4b
0.8c
Jum
lah
daun
Konsentrasi kolkisin (%)
0
2
4
6
0 0.1 0.2 0.3
4.8a
2.8b 2.6b
0.4c
Jum
lah
buku
Konsentrasi kolkisin (%)
00.10.20.30.40.5
0 0.1 0.2 0.3
0.5a0.33b 0.28b 0.23b
Panj
ang
daun
(cm
)
Konsentrasi kolkisin (%)

17
Keterangan: angka yang diikuti huruf yang sama menunjukkan hasil yang tidak berbeda
nyata berdasarkan DMRT pada α = 5% Gambar 15 Lebar daun tunas pucuk jeruk Siam Simadu pada umur delapan
minggu setelah perlakuan kolkisin
Semakin tinggi konsentrasi kolkisin yang diberikan maka pertambahan tinggi, jumlah daun, jumlah buku, panjang daun dan lebar daun tunas juga semakin sedikit sehingga ukuran tanaman menjadi lebih kecil dibandingkan dengan tanaman kontrol (Gambar 16 A, B, C, D). Hasil yang diperoleh dari penelitian ini memiliki kesamaan dengan penelitian-penelitian sebelumnya yaitu pada beberapa tanaman rentan terhadap perlakuan kolkisin yang menyebabkan penurunan pertumbuhan pada tanaman Gossypium arboretum L (Rauf et al. 2006), Plantanus acerifolia (Liu et al. 2007), dragonhead (Omidbaigi et al. 2010a), dan Pelargonium x hortorum (Jadrna et al. 2010).
Terhambatnya pertumbuhan tunas diduga karena proses pembelahan sel yang abnormal akibat pengaruh kolkisin. Damayanti dan Mariska (2003) menyebutkan bahwa pemberian kolkisin dapat mengakibatkan penundaan pertumbuhan akibat jaringan yang rusak dan memerlukan waktu lama untuk tumbuh. Menurut Suryo (2007) pembelahan sel menjadi lambat disebabkan jumlah kromosom yang mengganda.
Salah satu ciri fisik tunas poliploid yang umum adalah meningkatnya ukuran sel sehingga mengakibatkan daun menjadi lebih tebal (Griffith et al. 1999). Data Tabel 3 menunjukkan bahwa daun tunas pucuk yang diberikan perlakuan kolkisin lebih tebal dibandingkan dengan daun tunas pucuk kontrol. Semakin tinggi konsentrasi kolkisin, semakin tebal juga daun yang dihasilkan (Gambar 16 E, F, G, H). Hasil penelitian Omidbaigi et al. (2010) pada Ocimum basilicum L. menunjukkan bahwa tanaman tetraploid memiliki daun yang lebih tebal dibandingkan dengan tanaman diploid.
Induksi akar dilakukan dengan zat pengatur tumbuh (ZPT) golongan auksin. Auksin merupakan golongan zat pengatur tumbuh yang dapat mengatur terjadinya pertumbuhan akar (Liu et al. 1998). Gaspar et al. (1996) menyatakan bahwa auksin sangat diperlukan dalam organogenesis termasuk dalam pembentukan akar.
Berdasarkan hasil penelitian Merigo (2011), media induksi akar terbaik tunas jeruk keprok Batu 55 yaitu media MS + IBA 3 ppm. Oleh karena itu, induksi akar pada tunas pucuk hasil induksi tetraploid dilakukan pada media MS + vitamin MS + IBA 3 ppm. Penambahan auksin IBA 3 ppm dapat menginduksi pertumbuhan akar tunas pucuk kontrol (kolkisin 0%) sedangkan
0
0.1
0.2
0.3
0 0.1 0.2 0.3
0.3a 0.25ab 0.23b 0.2b
Leba
r dau
n (c
m)
Konsentrasi kolkisin (%)
18
tunas pucuk yang diberikan perlakuan kolkisin tidak (Tabel 1). Tunas pucuk hasil induksi tetraploid diinduksi akar kembali menggunakan media MS + vitamin MS + NAA 3 ppm. Media ini juga belum mampu untuk menginduksi akar tunas pucuk yang diberikan perlakuan kolkisin (Tabel 2) sehingga analisis tingkat ploidi tidak dapat dilakukan dengan cara menghitung kromosom karena tunas belum memiliki akar. Tabel 1 Jumlah dan panjang akar tunas pucuk jeruk Siam Simadu yang
diberikan perlakuan kolkisin pada media MS + vitamin MS + IBA 3 ppm selama satu bulan
Perlakuan kolkisin (%) Jumlah akar Panjang akar (cm) 0 0.8 1.5 0.1 0 0 0.2 0 0 0.3 0 0
Tabel 2 Jumlah dan panjang akar tunas pucuk jeruk Siam Simadu yang
diberikan perlakuan kolkisin pada media MS + vitamin MS + NAA 3 ppm selama satu bulan
Perlakuan kolkisin (%) Jumlah akar Panjang akar (cm) 0 0.72 1.2 0.1 0 0 0.2 0 0 0.3 0 0
Identifikasi tingkat ploidi dapat dilakukan dengan analisis stomata.
Teknik ini cepat, murah, tidak merusak, tidak memerlukan peralatan yang canggih, dan memiliki akurasi yang cukup tinggi hingga 90% (Ascough et al. 2008). Menurut Rego et al. (2011), metode penghitungan jumlah kloroplas dalam sel penjaga adalah cara yang efektif dan cepat untuk menentukan tingkat ploidi. Jumlah kloroplas dalam sel penjaga dapat mengidentifikasi tanaman diploid dan tetraploid. Gu et al. (2005) dan Yu et al. (2009) juga telah meneliti tingkat ploidi berdasarkan jumlah kloroplas.
Tunas yang diberikan perlakuan kolkisin 0.1% memiliki jumlah kloroplas dua kali lebih banyak daripada jumlah kloroplas tunas kontrol (diploid) (Tabel 3), sehingga dapat diduga bahwa tunas yang diberikan perlakuan kolkisin 0.1% merupakan tunas tetraploid jeruk Siam Simadu. Penelitian Rego et al. (2011) pada tanaman Passiflora edulis Sims. memberikan hasil yang sama, daun diploid dan tetraploid memiliki jumlah kloroplas masing-masing sebanyak 8.4 dan 16.3 per pasang sel penjaga sehingga daun tetraploid memiliki jumlah kloroplas dua kali lebih banyak dari daun diploid.

19
Tabel 3 Pengaruh perlakuan konsentrasi kolkisin terhadap tebal daun dan jumlah kloroplas per pasang sel penjaga
Konsentrasi kolkisin (%) Rata-rata tebal daun (µm) Jumlah kloropas/pasang sel
penjaga 0 146.01 ± 6.26 8.67 ± 0.58 0.1 185.01 ± 4.82 18.25 ± 1.71 0.2 197.04 ± 29.00 24.67 ± 3.21 0.3 205.24 ± 0.00* 32.00 ± 1.41 Keterangan : * hanya ada satu tanaman
Gambar 16 Tunas dengan perlakuan kolkisin (A) 0%, (B) 0.1%, (C) 0.2%,
(D) 0.3%, tebal daun dengan perlakuan kolkisin (E) 0%, (F) 0.1%, (G) 0.2%, (H) 0.3%, stomata dengan perlakuan kolkisin (I) 0%, (J) 0.1%, (K) 0.2%, (L) 0.3%
Lozykowska (2003) menyatakan bahwa ukuran stomata berhubungan dengan jumlah kloroplas pada sel penjaga. Peningkatan jumlah kloroplas pada sel penjaga mengakibatkan ukuran stomata menjadi lebih besar (Gambar 16 I, J, K, L). Tunas yang diberikan perlakuan kolkisin memiliki ukuran panjang dan lebar stomata yang lebih besar dibandingkan dengan tunas kontrol karena tunas yang diberikan perlakuan kolkisin memiliki jumlah kloroplas yang lebih banyak (Tabel 4). Tunas perlakuan kolkisin 0.1% memiliki ukuran panjang dan lebar stomata yang lebih besar daripada

20
tunas kontrol. Hasil penelitian Tang et al. (2011), tanaman diploid memiliki ukuran stomata yang lebih kecil dibandingkan dengan tanaman tetraploid turunannya.
Ukuran stomata yang semakin besar mengakibatkan kerapatan stomata menjadi semakin rendah (Tabel 4). Tunas yang diberikan perlakuan kolkisin memiliki kerapatan stomata lebih rendah dibandingkan dengan tunas kontrol karena tunas yang diberikan perlakuan kolkisin memiliki ukuran stomata yang lebih besar. Tunas perlakuan kolkisin 0.1% memiliki kerapatan stomata yang lebih rendah daripada tunas kontrol. Hasil yang sama juga diperoleh dari penelitian Rego et al. (2011) pada tanaman Passiflora edulis Sims., daun tetraploid memiliki kerapatan stomata yang lebih rendah dibandingkan dengan daun diploid.
Tabel 4 Pengaruh perlakuan konsentrasi kolkisin terhadap ukuran panjang
dan lebar stomata serta kerapatan stomata Konsentrasi kolkisin (%)
Panjang stomata (µm)
Lebar stomata (µm)
Kerapatan stomata (jumlah stomata mm-2)
0 18.32 ± 1.24 14.96 ± 2.84 67.52 ± 7.64 0.1 23.66 ± 2.63 21.44 ± 3.78 59.87 ± 8.70 0.2 22.34 ± 0.85 18.19 ± 1.09 54.35 ± 15.57 0.3 19.59 ± 2.52 17.31 ± 1.09 47.56 ± 51.55
Gambar 17 Daun pertama, daun ketiga, dan daun kelima tunas jeruk Siam
Simadu perlakuan kolkisin 0.1% yang dianalisis jumlah kloroplas
Tunas pucuk perlakuan kolkisin 0.1% dianalisis jumlah kloroplas pada daun pertama, ketiga, dan kelima (Gambar 17) karena diduga sebagai tunas tetraploid jeruk Siam Simadu. Daun pertama, ketiga, dan kelima Tunas 1 dan Tunas 3 memiliki jumlah kloroplas yang tidak berbeda nyata (Tabel 5) sehingga mutan yang dihasilkan cukup solid. Tunas 2 memiliki jumlah kloroplas yang sama pada daun pertama dan ketiga, tetapi memiliki jumlah kloroplas yang berbeda nyata pada daun kelima sehingga mutan yang dihasilkan tidak solid atau terjadi kimera (Tabel 5).

21
Tabel 5 Jumlah kloroplas daun pertama, ketiga, dan kelima tunas perlakuan kolkisin 0.1%
Daun Jumlah kloroplas Tunas 1 Tunas 2 Tunas 3
Pertama 15.50 ± 1.00 16.40 ± 1.14 16.00 ± 0.82 Ketiga 16.33 ± 1.53 17.33 ± 0.58 15.75 ± 0.96 Kelima 16.67 ± 0.58 24.67 ± 2.08 17.33 ± 1.15
Kimera dapat terjadi karena tunas pucuk terdiri atas banyak sel.
Bagian sel yang terluar akan mengalami penggandaan kloroplas lebih awal karena sel-sel langsung terkena kolkisin. Sel-sel meristematik dan bakal daun yang lebih muda pada meristem apikal, yang terlindung oleh bakal daun yang lebih tua (Gambar 18) akan mengganda setelah larutan kolkisin berhasil masuk ke bagian tersebut sehingga mengakibatkan penggandaan kloroplas yang berbeda dan terjadi kimera. Induksi tetraploid menggunakan tunas pucuk dapat menyebabkan persentase kimera (miksoploid) yang tinggi (Thao et al. 2003; Gu et al. 2005). Kimera juga ditemukan pada penelitian Aleza et al. (2009) dalam menghasilkan tanaman jeruk tetraploid.
Gambar 18 Struktur tunas pucuk (sel-sel meristematik, bakal daun yang lebih muda, dan bakal daun yang lebih tua) (UIC 2013)
Percobaan II. Induksi Tetraploid Tunas Samping Jeruk Siam Simadu
Menggunakan Kolkisin secara In Vitro
Pola respon kematian tunas samping jeruk terhadap berbagai konsentrasi kolkisin menghasilkan respon linear. Persamaaan respon kematian tunas samping jeruk terhadap berbagai konsentrasi kolkisin yaitu y = 279.9x + 5.447, nilai LD50 diperoleh pada konsentrasi kolkisin sebesar 0.16%. Gambar 19 memperlihatkan bahwa semakin tinggi konsentrasi kolkisin, maka semakin tinggi persentase tunas samping jeruk yang mati. Kematian tunas samping ini terjadi beberapa hari setelah
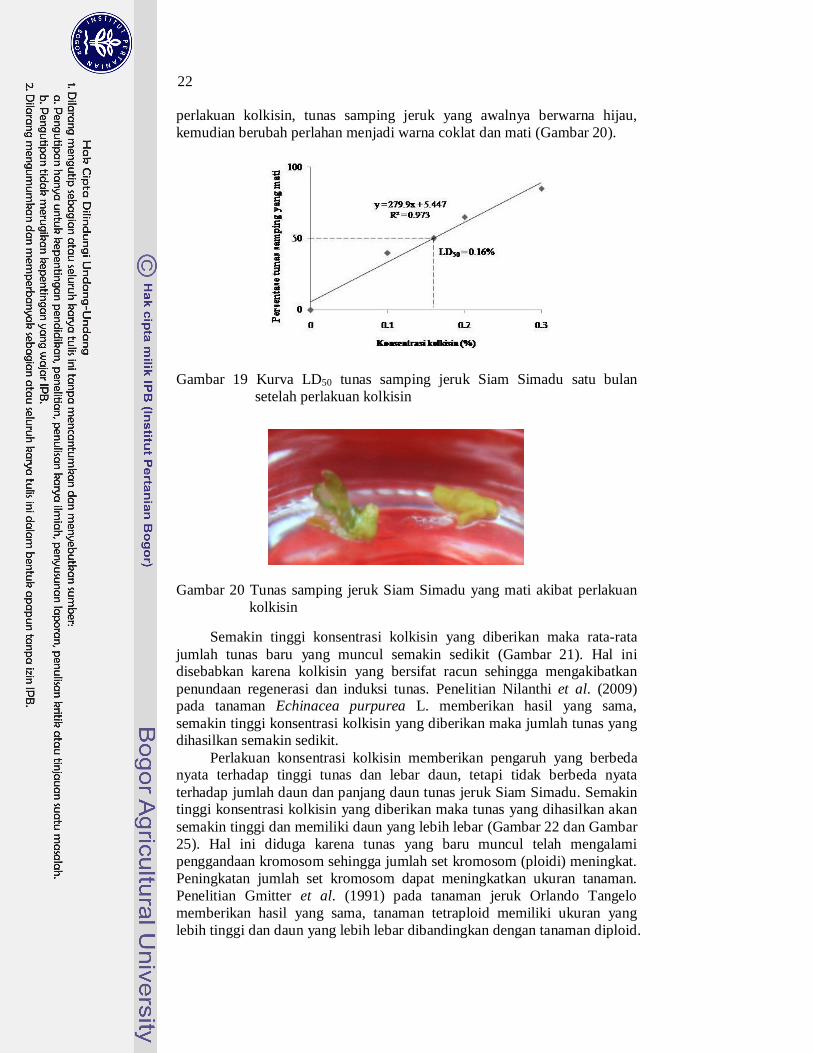
22
perlakuan kolkisin, tunas samping jeruk yang awalnya berwarna hijau, kemudian berubah perlahan menjadi warna coklat dan mati (Gambar 20).
Gambar 19 Kurva LD50 tunas samping jeruk Siam Simadu satu bulan setelah perlakuan kolkisin
Gambar 20 Tunas samping jeruk Siam Simadu yang mati akibat perlakuan kolkisin
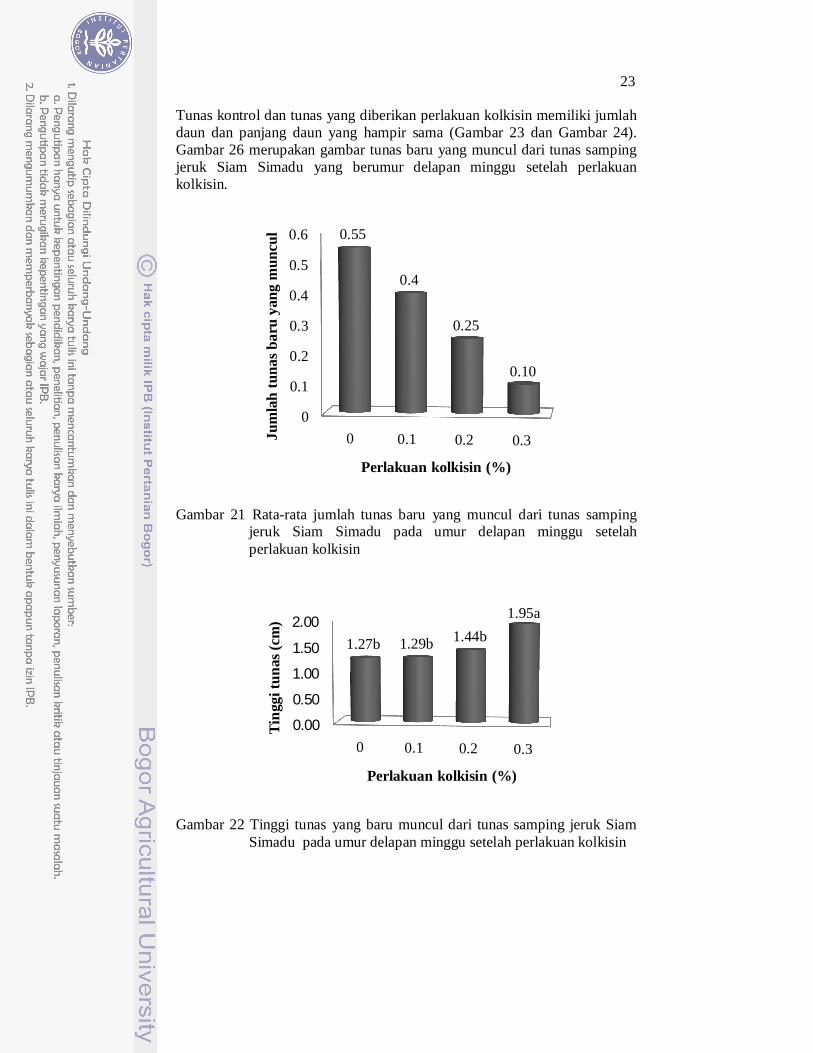
Semakin tinggi konsentrasi kolkisin yang diberikan maka rata-rata jumlah tunas baru yang muncul semakin sedikit (Gambar 21). Hal ini disebabkan karena kolkisin yang bersifat racun sehingga mengakibatkan penundaan regenerasi dan induksi tunas. Penelitian Nilanthi et al. (2009) pada tanaman Echinacea purpurea L. memberikan hasil yang sama, semakin tinggi konsentrasi kolkisin yang diberikan maka jumlah tunas yang dihasilkan semakin sedikit.
Perlakuan konsentrasi kolkisin memberikan pengaruh yang berbeda nyata terhadap tinggi tunas dan lebar daun, tetapi tidak berbeda nyata terhadap jumlah daun dan panjang daun tunas jeruk Siam Simadu. Semakin tinggi konsentrasi kolkisin yang diberikan maka tunas yang dihasilkan akan semakin tinggi dan memiliki daun yang lebih lebar (Gambar 22 dan Gambar 25). Hal ini diduga karena tunas yang baru muncul telah mengalami penggandaan kromosom sehingga jumlah set kromosom (ploidi) meningkat. Peningkatan jumlah set kromosom dapat meningkatkan ukuran tanaman. Penelitian Gmitter et al. (1991) pada tanaman jeruk Orlando Tangelo memberikan hasil yang sama, tanaman tetraploid memiliki ukuran yang lebih tinggi dan daun yang lebih lebar dibandingkan dengan tanaman diploid.
23
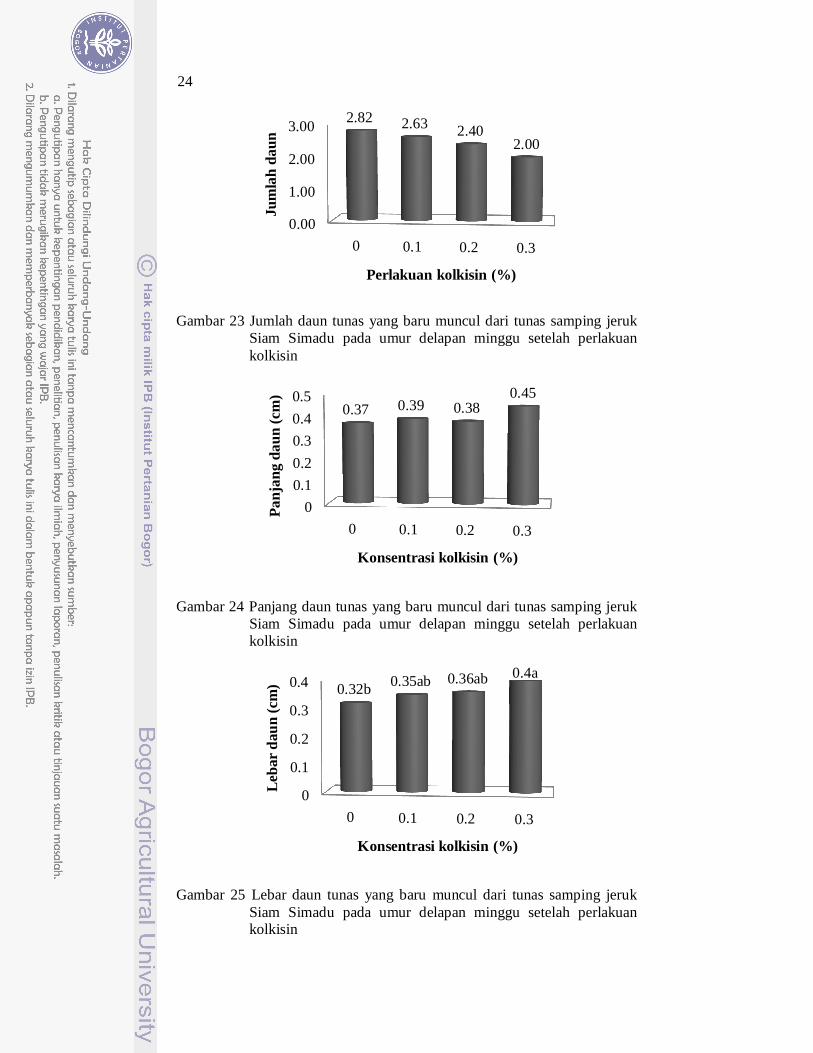
Tunas kontrol dan tunas yang diberikan perlakuan kolkisin memiliki jumlah daun dan panjang daun yang hampir sama (Gambar 23 dan Gambar 24). Gambar 26 merupakan gambar tunas baru yang muncul dari tunas samping jeruk Siam Simadu yang berumur delapan minggu setelah perlakuan kolkisin.
Gambar 21 Rata-rata jumlah tunas baru yang muncul dari tunas samping jeruk Siam Simadu pada umur delapan minggu setelah perlakuan kolkisin
Gambar 22 Tinggi tunas yang baru muncul dari tunas samping jeruk Siam Simadu pada umur delapan minggu setelah perlakuan kolkisin
0
0.1
0.2
0.3
0.4
0.5
0.6
0 0.1 0.2 0.3
0.55
0.4
0.25
0.10
Jum
lah
tuna
s bar
u ya
ng m
uncu
l
Perlakuan kolkisin (%)
0.00
0.50
1.00
1.50
2.00
0 0.1 0.2 0.3
1.27b 1.29b 1.44b1.95a
Ting
gi tu
nas (
cm)
Perlakuan kolkisin (%)
24
Gambar 23 Jumlah daun tunas yang baru muncul dari tunas samping jeruk Siam Simadu pada umur delapan minggu setelah perlakuan kolkisin
Gambar 24 Panjang daun tunas yang baru muncul dari tunas samping jeruk Siam Simadu pada umur delapan minggu setelah perlakuan kolkisin
Gambar 25 Lebar daun tunas yang baru muncul dari tunas samping jeruk Siam Simadu pada umur delapan minggu setelah perlakuan kolkisin
0.00
1.00
2.00
3.00
0 0.1 0.2 0.3
2.82 2.63 2.402.00
Jum
lah
daun
Perlakuan kolkisin (%)
00.10.20.30.40.5
0 0.1 0.2 0.3
0.37 0.39 0.380.45
Panj
ang
daun
(cm
)
Konsentrasi kolkisin (%)
0
0.1
0.2
0.3
0.4
0 0.1 0.2 0.3
0.32b 0.35ab 0.36ab 0.4a
Leba
r dau
n (c
m)
Konsentrasi kolkisin (%)

25
Gambar 26 Tunas baru yang muncul dari tunas samping jeruk Siam Simadu yang berumur delapan minggu setelah perlakuan kolkisin (A) 0%, (B) 0.1%, (C) 0.2%, dan (D) 0.3%
Data Tabel 8 menunjukkan bahwa daun tunas yang diberikan perlakuan kolkisin lebih tebal dibandingkan dengan daun tunas kontrol. Semakin tinggi konsentrasi kolkisin, semakin tebal juga daun yang dihasilkan (Gambar 27). Hasil penelitian Chen et al. (2010) pada Anthurium andraeanum menunjukkan bahwa tanaman tetraploid memiliki daun yang lebih tebal dibandingkan dengan tanaman diploid.
Gambar 27 Tebal daun tunas yang baru muncul dari tunas samping jeruk
Siam Simadu yang berumur delapan minggu setelah perlakuan kolkisin (A) 0%), (B) 0.1%, (C) 0.2%, dan (D) 0.3%
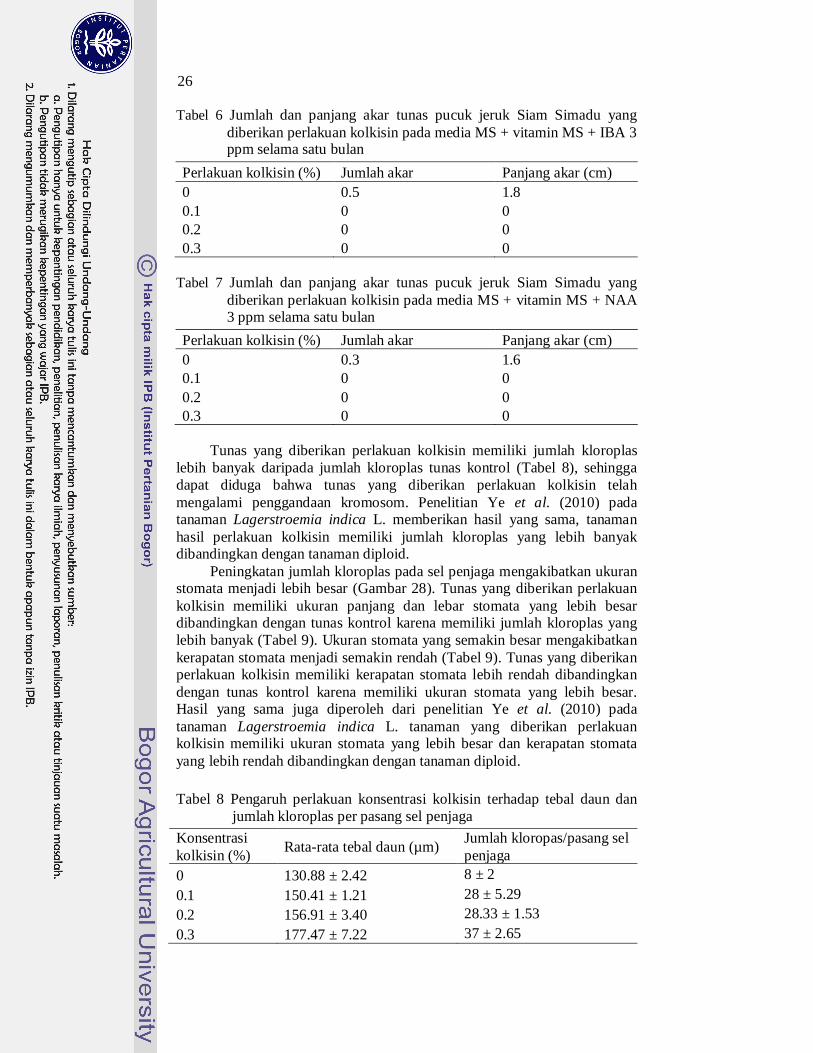
Induksi akar dilakukan dengan zat pengatur tumbuh (ZPT) golongan auksin. Penambahan auksin IBA 3 ppm dapat menginduksi pertumbuhan akar tunas pucuk kontrol (kolkisin 0%) sedangkan tunas pucuk yang diberikan perlakuan kolkisin tidak (Tabel 6). Tunas pucuk hasil induksi tetraploid diinduksi akar kembali menggunakan media MS + vitamin MS + NAA 3 ppm. Media ini juga belum mampu untuk menginduksi akar tunas pucuk yang diberikan perlakuan kolkisin (Tabel 7) sehingga analisis tingkat ploidi tidak dapat dilakukan dengan cara menghitung kromosom karena tunas belum memiliki akar.
26
Tabel 6 Jumlah dan panjang akar tunas pucuk jeruk Siam Simadu yang diberikan perlakuan kolkisin pada media MS + vitamin MS + IBA 3 ppm selama satu bulan
Perlakuan kolkisin (%) Jumlah akar Panjang akar (cm) 0 0.5 1.8 0.1 0 0 0.2 0 0 0.3 0 0
Tabel 7 Jumlah dan panjang akar tunas pucuk jeruk Siam Simadu yang
diberikan perlakuan kolkisin pada media MS + vitamin MS + NAA 3 ppm selama satu bulan
Perlakuan kolkisin (%) Jumlah akar Panjang akar (cm) 0 0.3 1.6 0.1 0 0 0.2 0 0 0.3 0 0
Tunas yang diberikan perlakuan kolkisin memiliki jumlah kloroplas
lebih banyak daripada jumlah kloroplas tunas kontrol (Tabel 8), sehingga dapat diduga bahwa tunas yang diberikan perlakuan kolkisin telah mengalami penggandaan kromosom. Penelitian Ye et al. (2010) pada tanaman Lagerstroemia indica L. memberikan hasil yang sama, tanaman hasil perlakuan kolkisin memiliki jumlah kloroplas yang lebih banyak dibandingkan dengan tanaman diploid.
Peningkatan jumlah kloroplas pada sel penjaga mengakibatkan ukuran stomata menjadi lebih besar (Gambar 28). Tunas yang diberikan perlakuan kolkisin memiliki ukuran panjang dan lebar stomata yang lebih besar dibandingkan dengan tunas kontrol karena memiliki jumlah kloroplas yang lebih banyak (Tabel 9). Ukuran stomata yang semakin besar mengakibatkan kerapatan stomata menjadi semakin rendah (Tabel 9). Tunas yang diberikan perlakuan kolkisin memiliki kerapatan stomata lebih rendah dibandingkan dengan tunas kontrol karena memiliki ukuran stomata yang lebih besar. Hasil yang sama juga diperoleh dari penelitian Ye et al. (2010) pada tanaman Lagerstroemia indica L. tanaman yang diberikan perlakuan kolkisin memiliki ukuran stomata yang lebih besar dan kerapatan stomata yang lebih rendah dibandingkan dengan tanaman diploid.
Tabel 8 Pengaruh perlakuan konsentrasi kolkisin terhadap tebal daun dan
jumlah kloroplas per pasang sel penjaga Konsentrasi kolkisin (%) Rata-rata tebal daun (µm) Jumlah kloropas/pasang sel
penjaga 0 130.88 ± 2.42 8 ± 2 0.1 150.41 ± 1.21 28 ± 5.29 0.2 156.91 ± 3.40 28.33 ± 1.53 0.3 177.47 ± 7.22 37 ± 2.65

27
Tabel 9 Pengaruh perlakuan konsentrasi kolkisin terhadap ukuran panjang dan lebar stomata serta kerapatan stomata
Konsentrasi kolkisin (%)
Panjang stomata (µm)
Lebar stomata (µm)
Kerapatan stomata (jumlah stomata mm-2)
0 16.81 ± 0.38 14.44 ± 0.40 68.12 ± 2.06 0.1 29.51 ± 7.74 30.80 ± 5.11 45.21 ± 5.49 0.2 34.48 ± 1.47 35.42 ± 0.55 43.26 ± 2.19 0.3 38.32 ± 0.34 38.42 ± 0.23 40.77 ± 1.07
Gambar 28 Stomata daun tunas yang baru muncul dari tunas samping jeruk Siam Simadu yang berumur delapan minggu setelah perlakuan kolkisin (A) 0%), (B) 0.1%, (C) 0.2%, dan (D) 0.3%
Percobaan III. Induksi Tetraploid Embrio Somatik Jeruk Siam Simadu
Menggunakan Kolkisin secara In Vitro
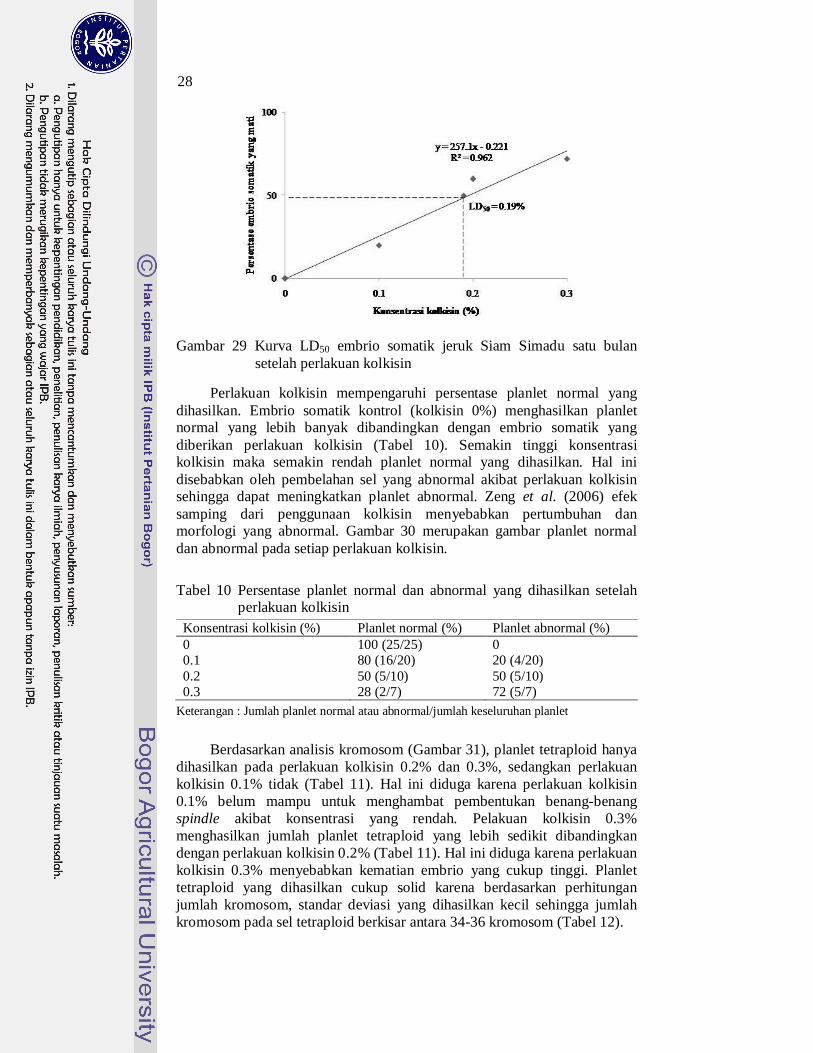
Pola respon kematian embrio somatik jeruk terhadap berbagai konsentrasi kolkisin menghasilkan respon linear. Persamaaan respon kematian embrio somatik jeruk terhadap berbagai konsentrasi kolkisin yaitu y = 257.1x - 0.221, nilai LD50 diperoleh pada konsentrasi kolkisin sebesar 0.19%. Gambar 29 memperlihatkan bahwa semakin tinggi konsentrasi kolkisin, maka semakin tinggi persentase embrio somatik jeruk yang mati.
28
Gambar 29 Kurva LD50 embrio somatik jeruk Siam Simadu satu bulan setelah perlakuan kolkisin
Perlakuan kolkisin mempengaruhi persentase planlet normal yang dihasilkan. Embrio somatik kontrol (kolkisin 0%) menghasilkan planlet normal yang lebih banyak dibandingkan dengan embrio somatik yang diberikan perlakuan kolkisin (Tabel 10). Semakin tinggi konsentrasi kolkisin maka semakin rendah planlet normal yang dihasilkan. Hal ini disebabkan oleh pembelahan sel yang abnormal akibat perlakuan kolkisin sehingga dapat meningkatkan planlet abnormal. Zeng et al. (2006) efek samping dari penggunaan kolkisin menyebabkan pertumbuhan dan morfologi yang abnormal. Gambar 30 merupakan gambar planlet normal dan abnormal pada setiap perlakuan kolkisin. Tabel 10 Persentase planlet normal dan abnormal yang dihasilkan setelah
perlakuan kolkisin Konsentrasi kolkisin (%) Planlet normal (%) Planlet abnormal (%) 0 100 (25/25) 0 0.1 80 (16/20) 20 (4/20) 0.2 50 (5/10) 50 (5/10) 0.3 28 (2/7) 72 (5/7)
Keterangan : Jumlah planlet normal atau abnormal/jumlah keseluruhan planlet
Berdasarkan analisis kromosom (Gambar 31), planlet tetraploid hanya dihasilkan pada perlakuan kolkisin 0.2% dan 0.3%, sedangkan perlakuan kolkisin 0.1% tidak (Tabel 11). Hal ini diduga karena perlakuan kolkisin 0.1% belum mampu untuk menghambat pembentukan benang-benang spindle akibat konsentrasi yang rendah. Pelakuan kolkisin 0.3% menghasilkan jumlah planlet tetraploid yang lebih sedikit dibandingkan dengan perlakuan kolkisin 0.2% (Tabel 11). Hal ini diduga karena perlakuan kolkisin 0.3% menyebabkan kematian embrio yang cukup tinggi. Planlet tetraploid yang dihasilkan cukup solid karena berdasarkan perhitungan jumlah kromosom, standar deviasi yang dihasilkan kecil sehingga jumlah kromosom pada sel tetraploid berkisar antara 34-36 kromosom (Tabel 12).
29
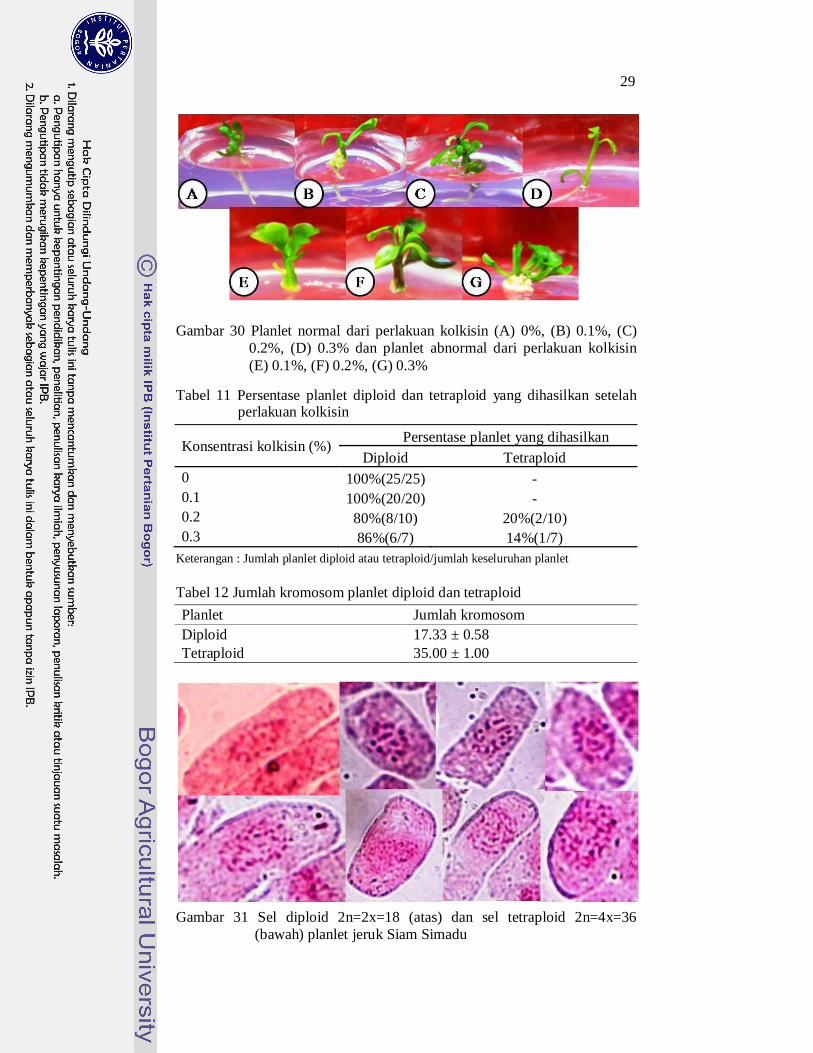
Gambar 30 Planlet normal dari perlakuan kolkisin (A) 0%, (B) 0.1%, (C)
0.2%, (D) 0.3% dan planlet abnormal dari perlakuan kolkisin (E) 0.1%, (F) 0.2%, (G) 0.3%
Tabel 11 Persentase planlet diploid dan tetraploid yang dihasilkan setelah perlakuan kolkisin
Konsentrasi kolkisin (%) Persentase planlet yang dihasilkan Diploid Tetraploid
0 100%(25/25) - 0.1 100%(20/20) - 0.2 80%(8/10) 20%(2/10) 0.3 86%(6/7) 14%(1/7)
Keterangan : Jumlah planlet diploid atau tetraploid/jumlah keseluruhan planlet Tabel 12 Jumlah kromosom planlet diploid dan tetraploid Planlet Jumlah kromosom Diploid 17.33 ± 0.58 Tetraploid 35.00 ± 1.00
Gambar 31 Sel diploid 2n=2x=18 (atas) dan sel tetraploid 2n=4x=36
(bawah) planlet jeruk Siam Simadu

30
Perbandingan antara Induksi Tetraploid Tunas Pucuk, Tunas Samping, dan Embrio Somatik Jeruk Siam Simadu Menggunakan Kolkisin
secara In Vitro
Induksi tetraploid jeruk Siam Simadu lebih baik menggunakan eksplan embrio somatik fase globular daripada tunas pucuk dan tunas samping karena embrio somatik fase globular memiliki jumlah sel yang lebih sedikit dibandingkan dengan tunas pucuk dan tunas samping sehingga munculnya kimera dapat berkurang. Embrio somatik dapat secara langsung menghasilkan akar dibandingkan dengan tunas pucuk dan tunas samping yang harus melalui tahap induksi akar terlebih dahulu karena embrio somatik berbentuk bipolar yaitu mempunyai calon meristem yaitu akar dan meristem tunas (Husni et al. 2010). Induksi tetraploid tunas pucuk dan tunas samping sampai saat ini belum dapat menginduksi akar, walaupun telah ditanam pada media yang ditambahkan auksin sehingga tidak dapat dilakukan analisis kromosom. Induksi tetraploid embrio somatik memerlukan waktu perendaman yang lebih cepat dibandingkan dengan tunas pucuk dan tunas samping karena embrio somatik memiliki ukuran yang lebih kecil dibandingkan dengan tunas pucuk dan tunas samping.
Pembahasan Umum
Induksi tetraploid menggunakan kolkisin dapat menurunkan pertumbuhan tunas jeruk Siam Simadu. Semakin tinggi konsentrasi kolkisin yang diberikan maka pertambahan tinggi, jumlah daun, jumlah buku, panjang daun dan lebar daun tunas juga semakin sedikit sehingga ukuran tanaman menjadi lebih kecil dibandingkan dengan tanaman kontrol. Hasil yang diperoleh dari penelitian ini memiliki kesamaan dengan penelitian-penelitian sebelumnya yaitu pada beberapa tanaman rentan terhadap perlakuan kolkisin yang menyebabkan penurunan pertumbuhan pada tanaman Gossypium arboretum L (Rauf et al. 2006), Plantanus acerifolia (Liu et al. 2007), dragonhead (Omidbaigi et al. 2010a), dan Pelargonium x hortorum (Jadrna et al. 2010).
Terhambatnya pertumbuhan tunas diduga karena proses pembelahan sel yang abnormal akibat pengaruh kolkisin. Damayanti dan Mariska (2003) menyebutkan pemberian kolkisin dapat mengakibatkan penundaan pertumbuhan akibat jaringan yang rusak dan memerlukan waktu lama untuk tumbuh. Menurut Suryo (2007) pembelahan sel menjadi lambat disebabkan jumlah kromosom yang mengganda.
Salah satu ciri fisik tunas poliploid yang umum adalah meningkatnya ukuran sel sehingga mengakibatkan daun menjadi lebih tebal (Griffith et al. 1999). Daun tunas pucuk yang diberikan perlakuan kolkisin lebih tebal dibandingkan dengan daun tunas pucuk kontrol. Semakin tinggi konsentrasi kolkisin, semakin tebal juga daun yang dihasilkan. Hasil penelitian Omidbaigi et al. (2010) pada Ocimum basilicum L. menunjukkan bahwa tanaman tetraploid memiliki daun yang lebih tebal dibandingkan dengan tanaman diploid.

31
Identifikasi tingkat ploidi dapat dilakukan dengan analisis stomata. Teknik ini cepat, murah, tidak merusak, tidak memerlukan peralatan yang canggih, dan memiliki akurasi yang cukup tinggi hingga 90% (Ascough et al. 2008). Menurut Rego et al. (2011), metode penghitungan jumlah kloroplas dalam sel penjaga adalah cara yang efektif dan cepat untuk menentukan tingkat ploidi. Jumlah kloroplas dalam sel penjaga dapat mengidentifikasi tanaman diploid dan tetraploid. Gu et al. (2005) dan Yu et al. (2009) juga telah meneliti tingkat ploidi berdasarkan jumlah kloroplas.
Tunas yang diberikan perlakuan kolkisin 0.1% memiliki jumlah kloroplas dua kali lebih banyak daripada jumlah kloroplas tunas kontrol (diploid) sehingga dapat diduga bahwa tunas yang diberikan perlakuan kolkisin 0.1% merupakan tunas tetraploid jeruk Siam Simadu. Penelitian Rego et al. (2011) pada tanaman Passiflora edulis Sims. memberikan hasil yang sama, daun tetraploid memiliki jumlah kloroplas dua kali lebih banyak dari daun diploid.
Lozykowska (2003) menyatakan bahwa ukuran stomata berhubungan dengan jumlah kloroplas pada sel penjaga. Peningkatan jumlah kloroplas pada sel penjaga mengakibatkan ukuran stomata menjadi lebih besar. Tunas yang diberikan perlakuan kolkisin memiliki ukuran panjang dan lebar stomata yang lebih besar dibandingkan dengan tunas kontrol karena tunas yang diberikan perlakuan kolkisin memiliki jumlah kloroplas yang lebih banyak. Tunas perlakuan kolkisin 0.1% memiliki ukuran panjang dan lebar stomata yang lebih besar daripada tunas kontrol. Hasil penelitian Tang et al. (2011), tanaman diploid memiliki ukuran stomata yang lebih kecil dibandingkan dengan tanaman tetraploid turunannya.
Ukuran stomata yang semakin besar mengakibatkan kerapatan stomata menjadi semakin rendah. Tunas yang diberikan perlakuan kolkisin memiliki kerapatan stomata lebih rendah dibandingkan dengan tunas kontrol karena tunas yang diberikan perlakuan kolkisin memiliki ukuran stomata yang lebih besar. Tunas perlakuan kolkisin 0.1% memiliki kerapatan stomata yang lebih rendah daripada tunas kontrol. Hasil yang sama juga diperoleh dari penelitian Rego et al. (2011) pada tanaman Passiflora edulis Sims., daun tetraploid memiliki kerapatan stomata yang lebih rendah dibandingkan dengan daun diploid.
Induksi tetraploid menggunakan kolkisin dapat menurunkan rata-rata jumlah tunas baru yang muncul dari tunas samping jeruk Siam Simadu. Hal ini disebabkan karena kolkisin yang bersifat racun sehingga mengakibatkan penundaan regenerasi dan induksi tunas. Penelitian Nilanthi et al. (2009) pada tanaman Echinacea purpurea L. memberikan hasil yang sama, semakin tinggi konsentrasi kolkisin yang diberikan maka jumlah tunas yang dihasilkan semakin sedikit.
Semakin tinggi konsentrasi kolkisin yang diberikan maka tunas baru yang muncul dari tunas samping akan semakin tinggi dan memiliki daun yang lebih lebar. Hal ini diduga karena tunas yang baru muncul telah mengalami penggandaan kromosom sehingga jumlah set kromosom (ploidi) meningkat. Peningkatan jumlah set kromosom dapat meningkatkan ukuran tanaman. Penelitian Gmitter et al. (1991) pada tanaman jeruk Orlando Tangelo memberikan hasil yang sama, tanaman tetraploid memiliki ukuran

32
yang lebih tinggi dan daun yang lebih lebar dibandingkan dengan tanaman diploid.
Daun dari tunas yang diberikan perlakuan kolkisin lebih tebal dibandingkan dengan daun tunas kontrol. Semakin tinggi konsentrasi kolkisin, semakin tebal juga daun yang dihasilkan. Hasil penelitian Chen et al. (2010) pada Anthurium andraeanum menunjukkan bahwa tanaman tetraploid memiliki daun yang lebih tebal dibandingkan dengan tanaman diploid.
Tunas yang diberikan perlakuan kolkisin memiliki jumlah kloroplas lebih banyak daripada jumlah kloroplas tunas kontrol sehingga dapat diduga bahwa tunas yang diberikan perlakuan kolkisin telah mengalami penggandaan kromosom. Penelitian Ye et al. (2010) pada tanaman Lagerstroemia indica L. memberikan hasil yang sama, tanaman hasil perlakuan kolkisin memiliki jumlah kloroplas yang lebih banyak dibandingkan dengan tanaman diploid.
Tunas yang diberikan perlakuan kolkisin memiliki ukuran panjang dan lebar stomata yang lebih besar dan kerapatan stomata yang lebih rendah dibandingkan dengan tunas kontrol. Hasil yang sama juga diperoleh dari penelitian Ye et al. (2010) pada tanaman Lagerstroemia indica L. tanaman yang diberikan perlakuan kolkisin memiliki ukuran stomata yang lebih besar dan kerapatan stomata yang lebih rendah dibandingkan dengan tanaman diploid.
Induksi akar pada tunas hasil induksi tetraploid dilakukan dengan zat pengatur tumbuh (ZPT) golongan auksin yaitu IBA dan NAA. Penambahan IBA 3 ppm dapat menginduksi pertumbuhan akar tunas kontrol (kolkisin 0%) sedangkan tunas yang diberikan perlakuan kolkisin tidak. Tunas hasil induksi tetraploid diinduksi akar kembali menggunakan media yang ditambahkan NAA 3 ppm. Penambahan NAA 3 ppm belum mampu untuk menginduksi akar tunas yang diberikan perlakuan kolkisin sehingga analisis tingkat ploidi tidak dapat dilakukan dengan cara menghitung kromosom karena tunas belum memiliki akar.
Perlakuan kolkisin mempengaruhi persentase planlet normal yang dihasilkan dari embrio somatik. Embrio somatik kontrol (kolkisin 0%) menghasilkan planlet normal yang lebih banyak dibandingkan dengan embrio somatik yang diberikan perlakuan kolkisin. Semakin tinggi konsentrasi kolkisin maka semakin rendah planlet normal yang dihasilkan. Hal ini disebabkan oleh pembelahan sel yang abnormal akibat perlakuan kolkisin sehingga dapat meningkatkan planlet abnormal. Zeng et al. (2006) efek samping dari penggunaan kolkisin menyebabkan pertumbuhan dan morfologi yang abnormal.
Berdasarkan analisis kromosom, planlet tetraploid hanya dihasilkan pada perlakuan kolkisin 0.2% dan 0.3%, sedangkan perlakuan kolkisin 0.1% tidak. Hal ini diduga karena perlakuan kolkisin 0.1% belum mampu untuk menghambat pembentukan benang-benang spindle akibat konsentrasi yang rendah. Pelakuan kolkisin 0.3% menghasilkan jumlah planlet tetraploid yang lebih sedikit dibandingkan dengan perlakuan kolkisin 0.2%. Hal ini diduga karena perlakuan kolkisin 0.3% menyebabkan kematian embrio yang cukup tinggi.